Embed Size (px)
Citation preview

Lựa chọn các điều kiện lên men xốp tối ưu và
nghiên cứu các đặc tính xylanase từ các chủng
vi khuẩn ưu nhiệt
Nguyễn Thị Nhiên
Trường Đại học Khoa học Tự nhiên
Luận văn Thạc sĩ ngành: Vi sinh vật; Mã số: 60 42 40
Người hướng dẫn: TS. Đào Thị Lương
Năm bảo vệ: 2011
Abstract: Khảo sát và tuyển chọn những chủng vi khuẩn và xạ khuẩn bền nhiệt có
hoạt độ enzyme xylanase cao. Nghiên cứu đặc điểm sinh học và phân loại các chủng
vi khuẩn và xạ khuẩn nghiên cứu. Lựa chọn các điều kiện lên men xốp tối ưu cho các
chủng vi khuẩn và xạ khuẩn nghiên cứu. Nghiên cứu đặc tính và tinh sạch enzyme của
các chủng nghiên cứu.
Keywords: Vi sinh vật học; Vi khuẩn; Lên men; Sinh học
Content
MỞ ĐẦU
1. Lý do lựa chọn đề tài
Trong nhiều năm gần đây, nhu cầu sử dụng enzyme có nguồn gốc từ vi sinh vật ngày
càng tăng, đặc biệt là enzyme xylanase. Xylanase được sử dụng trong nhiều ngành sản xuất
trên toàn thế giới [26, 15]. Một trong những ứng dụng quan trọng của xylanase là được dùng
để bổ sung vào thức ăn chăn nuôi. Sự có mặt của xylanase trong thức ăn chăn nuôi có tác
dụng thúc đẩy quá trình tiêu hóa, cải thiện hệ vi sinh vật đường ruột của động vật [11, 6].
Ngoài ra, xylanase còn được ứng dụng trong nhiều lĩnh vực khác như: phân hủy rác thải; tẩy
trắng trong công nghiệp sản xuất giấy, sản xuất nhiên liệu sinh học,… [33, 30].
Xét ở quy mô công nghiệp, sản xuất enzyme là một lĩnh vực đang ngày càng phát triển
của công nghệ sinh học nói chung và công nghệ thực phẩm nói riêng. Nhu cầu sử dụng
enzyme có nguồn gốc từ vi sinh vật ngày càng tăng cao, đặc biệt là enzyme xylanase được sử
dụng trong nhiều ngành sản xuất trên toàn thế giới [14].
Trong sinh giới, động vật và thực vật không có khả năng tự sinh xylanase, do đó vai trò
thủy phân xylan phụ thuộc chủ yếu vào nguồn vi sinh vật, đặc biệt quan trọng là các nhóm vi
khuẩn, xạ khuẩn và nấm. Trước đây, việc nghiên cứu xylanase chủ yếu được tiến hành trên
đối tượng nấm sợi. Tuy nhiên, gần đây cùng với sự phát mẽ của khoa học công nghệ, ngày

2
càng nhiều chủng vi sinh vật có khả năng sinh xylanase được phát hiện và nghiên cứu sâu.
Đồng thời, các lĩnh vực liên quan đến enzyme xylanase và ứng dụng của loại enzyme này ở vi
khuẩn và xạ khuẩn được các nhà khoa học ngày càng quan tâm hơn.
Lên men xốp được đánh giá là con đường tốt nhất để sản xuất enzyme và các sản
phẩm bền nhiệt khác. Lên men xốp ngày càng được sử dụng rộng rãi trong sản xuất cũng như
trong nghiên cứu vì những ứng dụng mang tính thực tiễn và kinh tế của nó. Lên men xốp
được dùng trong công nghiệp chế biến và sản xuất thực phẩm chủ yếu là lĩnh vực sản xuất các
loại hương liệu, sản xuất enzyme (a-amylase, fructosyl transferase, lipase, pectinase,
xylanase), các acid hữu cơ (acid lactic, acid citric) và các loại kẹo cao su [8].
Trước đây, trong sản xuất enzyme người ta thường sử dụng kỹ thuật lên men dịch thể.
Tuy nhiên, khi tiến hành so sánh hiệu quả của các loại hình lên men, người ta thấy hàm lượng
enzyme vi sinh vật có thể sinh ra trong lên men xốp cao hơn nhiều lần so với trong lên men
dịch thể khi sử dụng cùng một chủng vi sinh vật trong cùng điều kiện lên men [14].
Để góp phần bổ sung các số liệu cần thiết về việc ứng dụng kỹ thuật lên men xốp
trong công nghiệp sản xuất enzyme chúng tôi thực hiện đề tài: “Lựa chọn các điều kiện lên
men xốp tối ưu và nghiên cứu đặc tính xylanase từ các chủng vi khuẩn ưa nhiệt”.
2. Nhiệm vụ và mục đích nghiên cứu
- Khảo sát và tuyển chọn những chủng vi khuẩn và xạ khuẩn bền nhiệt có hoạt độ
enzyme xylanase cao.
- Nghiên cứu đặc điểm sinh học và phân loại các chủng vi khuẩn và xạ khuẩn nghiên
cứu.
- Lựa chọn các điều kiện lên men xốp tối ưu cho các chủng vi khuẩn và xạ khuẩn
nghiên cứu.
- Nghiên cứu đặc tính và tinh sạch enzyme của các chủng nghiên cứu.
3. Những đóng góp mới của đề tài
Đê tai chúng tôi thực hiện có kê thưa cac công trình nghiên cứu đã công bố gần đây về
mặt phương phap nghiên cưu , tuy nhiên kêt qua cua no hoan toan mơi và độc lập . Do đo đê tai
sẽ góp phần đóng góp thêm số liệu vào cơ sở dữ liệu về quá trình sản xuất enzyme xylanase
bằng kỹ thuật lên men xốp.
Chƣơng 1. TỔNG QUAN TÀI LIỆU
1.1. XYLAN
1.1.1. Xylan
Xylan là thành phần chính của hemicellulose thực vật, là polysaccharide phổ biến thứ
hai trong tự nhiên chỉ sau cellulose và được tìm thấy trong thành tế bào thực vật [27]. Xylan
còn tham gia vào cấu trúc và giúp lignocellulose bền hơn.
Thực vật trên cạn có xylan là chuỗi D-xylosyl được nối với nhau bằng liên kết β-1,4,
tảo biển tổng hợp xylan với một cấu trúc hóa học khác đó là các monomer D-xylose nối với
nhau bằng liên kết β-1,3. Ở một vài lớp Chlorophycaea và Rhodophycaea không có sự hiện
diện của cellulose, xylan tạo nên vật liệu hình sợi trong suốt. Thông thường, xylan chiếm
3
khoảng 15-30% trọng lượng chất khô trong cây hạt kín và khoảng 7-15% trọng lượng chất
khô của cây hạt trần. Đặc biệt trong cây lá rộng, xylan chiếm tới 35% tổng trọng lượng chất
khô của thực vật [21].
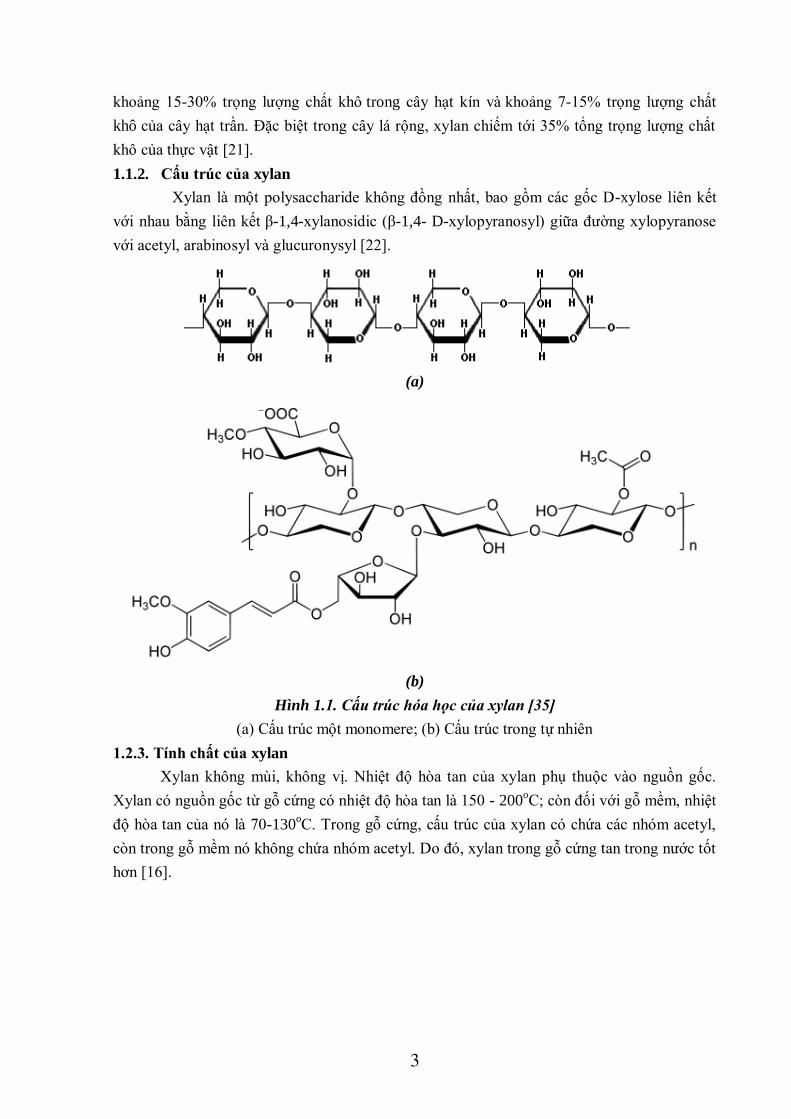
1.1.2. Cấu trúc của xylan
Xylan là một polysaccharide không đồng nhất, bao gồm các gốc D-xylose liên kết
với nhau bằng liên kết β-1,4-xylanosidic (β-1,4- D-xylopyranosyl) giữa đường xylopyranose
với acetyl, arabinosyl và glucuronysyl [22].
(a)
(b)
Hình 1.1. Cấu trúc hóa học của xylan [35]
(a) Cấu trúc một monomere; (b) Cấu trúc trong tự nhiên
1.2.3. Tính chất của xylan
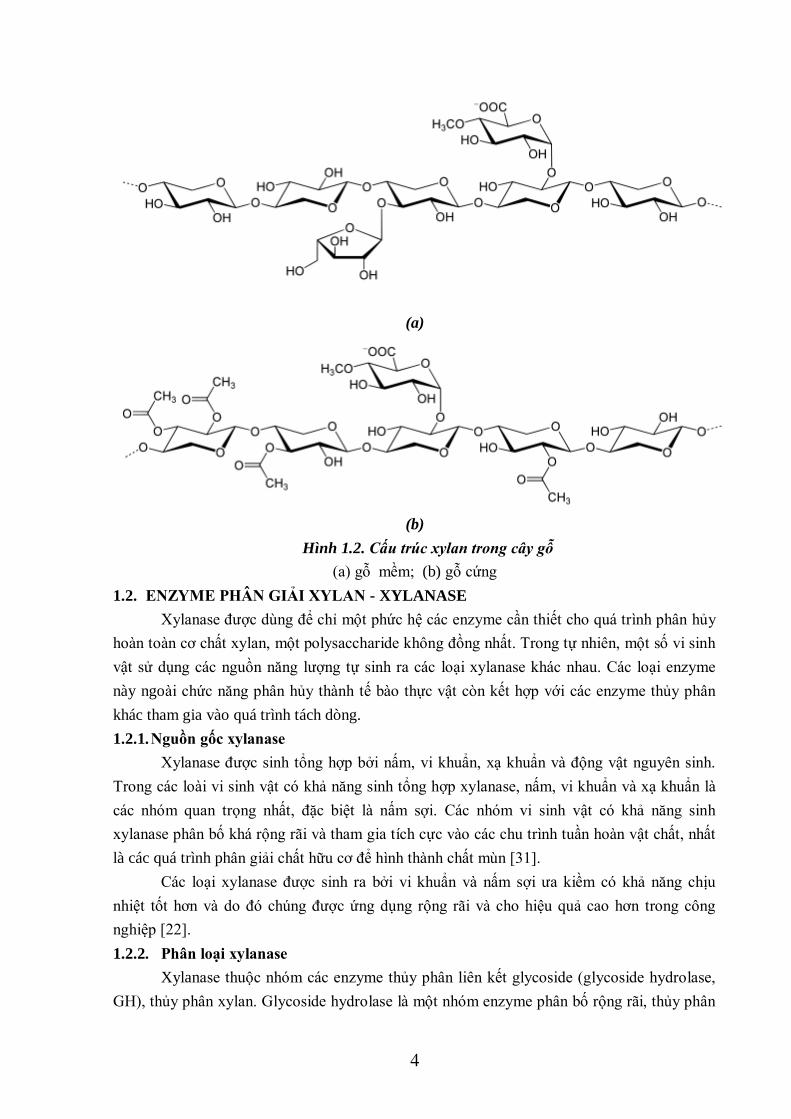
Xylan không mùi, không vị. Nhiệt độ hòa tan của xylan phụ thuộc vào nguồn gốc.
Xylan có nguồn gốc từ gỗ cứng có nhiệt độ hòa tan là 150 - 200oC; còn đối với gỗ mềm, nhiệt
độ hòa tan của nó là 70-130oC. Trong gỗ cứng, cấu trúc của xylan có chứa các nhóm acetyl,
còn trong gỗ mềm nó không chứa nhóm acetyl. Do đó, xylan trong gỗ cứng tan trong nước tốt
hơn [16].
4
(a)
(b)
Hình 1.2. Cấu trúc xylan trong cây gỗ
(a) gỗ mềm; (b) gỗ cứng
1.2. ENZYME PHÂN GIẢI XYLAN - XYLANASE
Xylanase được dùng để chỉ một phức hệ các enzyme cần thiết cho quá trình phân hủy
hoàn toàn cơ chất xylan, một polysaccharide không đồng nhất. Trong tự nhiên, một số vi sinh
vật sử dụng các nguồn năng lượng tự sinh ra các loại xylanase khác nhau. Các loại enzyme
này ngoài chức năng phân hủy thành tế bào thực vật còn kết hợp với các enzyme thủy phân
khác tham gia vào quá trình tách dòng.
1.2.1. Nguồn gốc xylanase
Xylanase được sinh tổng hợp bởi nấm, vi khuẩn, xạ khuẩn và động vật nguyên sinh.
Trong các loài vi sinh vật có khả năng sinh tổng hợp xylanase, nấm, vi khuẩn và xạ khuẩn là
các nhóm quan trọng nhất, đặc biệt là nấm sợi. Các nhóm vi sinh vật có khả năng sinh
xylanase phân bố khá rộng rãi và tham gia tích cực vào các chu trình tuần hoàn vật chất, nhất
là các quá trình phân giải chất hữu cơ để hình thành chất mùn [31].
Các loại xylanase được sinh ra bởi vi khuẩn và nấm sợi ưa kiềm có khả năng chịu
nhiệt tốt hơn và do đó chúng được ứng dụng rộng rãi và cho hiệu quả cao hơn trong công
nghiệp [22].
1.2.2. Phân loại xylanase
Xylanase thuộc nhóm các enzyme thủy phân liên kết glycoside (glycoside hydrolase,
GH), thủy phân xylan. Glycoside hydrolase là một nhóm enzyme phân bố rộng rãi, thủy phân

5
các liên kết glycoside trong chuỗi polysaccharide hoặc oligosaccharide. Do cấu trúc phức tạp
của các carbonhydrate trong tự nhiên, nên các enzyme đặc hiệu cơ chất khác nhau cũng tồn tại
số lượng lớn song song với nó. GH từ các nguồn khác nhau được phân loại thành các họ khác
nhau dựa vào trình tự axit amin tương đồng. Nguyên tắc cơ bản là từ trình tự axit amin bậc
một có thể thấy được mối liên kết trực tiếp giữa cấu trúc bậc này và sự cuộn gấp của các
enzyme đối với các thành viên trong một hệ nhất định [7].
Dựa vào sự tương đồng trình tự acid amin và các kết quả phân tích về cấu trúc không
gian, xylanase chủ yếu được xếp vào họ 10 và 11 của hệ enzyme thủy phân glycoside (Collin
& cộng sự, 2005; Jeya & cộng sự, 2009; Sibtain & cộng sự, 2009). Xylanase cả hai họ 10 và
11 đều thủy phân xylan bằng cơ chế thay thế hai lần liên quan đến hai acid glutamic (Sinnot,
1990). Hai phân tử acid này nằm tại trung tâm hoạt động của enzyme và quyết định một phần
đến sự hình thành cấu trúc của xylanase. Một phân tử acid hoạt động như một chất xúc tác
bằng cách thêm proton vào cơ chất, trong khi phân tử acid thứ hai có ái lực mạnh với nhân, có
vai trò mở đầu cho phản ứng phân cắt và trong sự hình thành enzyme α-glycosyl (Collin &
cộng sự, 2005) [12].
Các thành viên họ xylanase 10 có cấu trúc α/β gấp cuộn 8 lần và trọng lượng phân tử
xấp xỉ 48 kDa, pI thấp, và thủy phân các liên kết glycoside với sự giữ được hình thể nguyên
vẹn, trong khi đó, họ xylanase 11 có cấu trúc mạch β gập hình bánh sandwich gần giống với
cấu trúc bàn tay phải và có khối lượng phân tử nhỏ hơn, vào khoảng 20 kDa, pI cao và thủy
phân các liên kết glycoside với cơ chế xúc tác đổi chỗ hai lần (Hakylinen & cộng sự, 2003;
Zhou & cộng sự, 2009).
Xylanase thuộc hệ GH10 thuộc nhóm acid xylanase. Xylanase thuộc hệ GH11 thuộc
nhóm alkaline xylanase [36].
1.2.3. Cấu trúc
Cấu trúc bậc ba của phân tử xylanase được xác định bằng cách sử dụng mô hình
Insight II Molecular. Theo đó, cấu trúc xylanase bao gồm một miền đơn có chứa 2 vùng cấu
trúc β và một miền có chuỗi xoắn α, các cấu trúc này chỉ có ở xylanase hệ GH11. Cấu trúc
chung của phân tử xylanase được ví như ―bàn tay phải‖ với các ngón tay ở phía dưới và ngón
cái nằm phía
trên. Trung tâm
hoạt động là
ngón tay cái,
lòng bàn tay và
ngón tay.
6
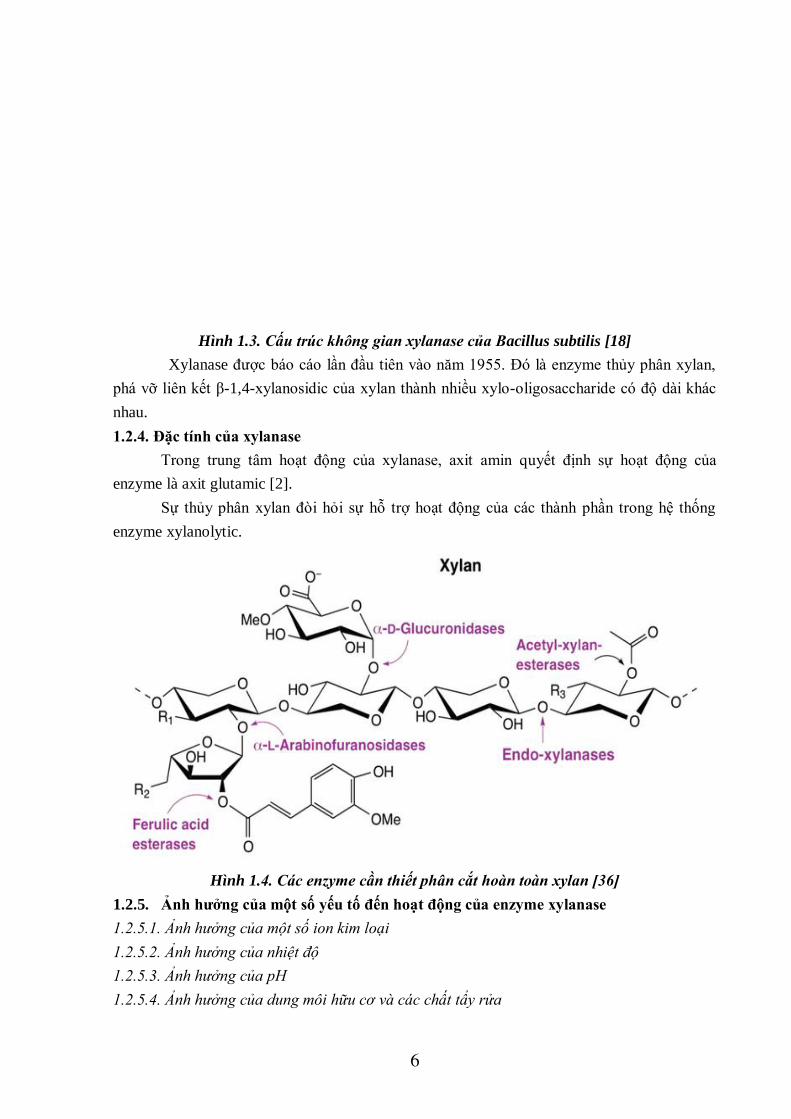
Hình 1.3. Cấu trúc không gian xylanase của Bacillus subtilis [18]
Xylanase được báo cáo lần đầu tiên vào năm 1955. Đó là enzyme thủy phân xylan,
phá vỡ liên kết β-1,4-xylanosidic của xylan thành nhiều xylo-oligosaccharide có độ dài khác
nhau.
1.2.4. Đặc tính của xylanase
Trong trung tâm hoạt động của xylanase, axit amin quyết định sự hoạt động của
enzyme là axit glutamic [2].
Sự thủy phân xylan đòi hỏi sự hỗ trợ hoạt động của các thành phần trong hệ thống
enzyme xylanolytic.
Hình 1.4. Các enzyme cần thiết phân cắt hoàn toàn xylan [36]
1.2.5. Ảnh hƣởng của một số yếu tố đến hoạt động của enzyme xylanase
1.2.5.1. 1.2.5.1. Ảnh hưởng của một số ion kim loại
1.2.5.2. Ảnh hưởng của nhiệt độ
1.2.5.3. Ảnh hưởng của pH
1.2.5.4. Ảnh hưởng của dung môi hữu cơ và các chất tẩy rửa

7
1.2.6. Ứng dụng của xylanase
1.2.6.1. Ứng dụng của xylanase trong công nghiệp thực phẩm
1.2.6.2. Trong công nghiệp sản xuất giấy và bột giấy
1.2.6.3. Ứng dụng của xylanase trong sản xuất nguyên liệu sinh học
1.2.6.4. Ứng dụng trong công nghiệp vải sợi
1.2.6.5. Ứng dụng trong nông nghiệp
1.2.6.6. Ứng dụng của xylanase trong xử lý môi trường
1.2.7. Vi sinh vật sinh xylanase
Như đã trình bày ở mục 1.2.2, xylanase được sinh tổng hợp bởi nấm, vi khuẩn, xạ
khuẩn và động vật nguyên sinh. Xylanase chủ yếu thuộc hệ 10 và 11 của hệ enzyme thủy
phân glycoside. Xylanase thường được tổng hợp bởi một số vi sinh vật như Streptomyces,
Bacillus, Trichoderma, Aspergillus, Thermobacillus …[36].
Trong các loài vi sinh vật có khả năng sinh tổng hợp xylanase, chi Bacillus và
Streptomyces đặc biệt được quan tâm nghiên cứu. Vi khuẩn thuộc chi Bacillus phân bố rộng
rãi trong tự nhiên, đa dạng về sinh thái. Các loài thuộc chi Bacillus đã, đang và ngày càng trở
thành những vi sinh vật quan trọng hàng đầu về mặt ứng dụng. Các ứng dụng của chúng bao
trùm hàng loạt lĩnh vực, từ sản xuất thực phẩm thủ công truyền thống đến công nghệ lên men
hiện đại, đến sinh học phân tử, y-dược học chữa các bệnh hiểm nghèo, mỹ phẩm, xử lý môi
trường ô nhiễm, thu hồi bạc kim loại từ các phế liệu. Chính vì lẽ đó nên đã có ngày càng
nhiều các nghiên cứu sâu về chi Bacillus này cũng như mở rộng ứng dụng của chúng đối với
đời sống con nguời, trong đó có rất nhiều nghiên cứu công bố liên quan đến khả năng sinh
xylanase của các chủng thuộc chi này [3].
Chi Streptomyces được Wakman và Henrici đặt tên năm 1943 [32]. Đây là chi có số
lượng loài được mô tả lớn nhất. Streptomyces có tiêm năng rât lơn trong viêc sinh tông hơp
nhiêu hơp chât có hoạt độ sinh hoc, phân giai nhiêu hơp chât phưc tap , kích thích sự phát triển
của cây trồng , có vai tro quan trong trong cac chu trình vật chất ngoài tự nhiên . Đặc biệt ,
Streptomyces có khả năng sinh tổng hợp nhiều loại enzyme thủy phân có hoạt tính cao , trong
đó phải kể đến xylanase [13,25,29,5].
1.3. LÊN MEN XỐP
1.3.1. Khái niệm lên men xốp
Thuật ngữ lên mên xốp được dùng cho các quá trình mà trong đó vi sinh vật sinh
trưởng bằng cách sử dụng các vật liệu không tan trong nước, trong quá trình lên men này độ
ẩm không vượt quá hàm lượng bão hòa của cơ chất xốp. Nước là yếu tố thiết yếu cho sự sinh
trưởng của vi sinh vật trong lên men xốp, mặc dù chỉ cần một lớp nước mỏng và thông thường
nước sẽ được hấp thụ vào bên trong cơ chất.
1.3.2. Ƣu điểm của kỹ thuật lên men xốp
Trước đây, ở các nước phương Tây, lên men xốp vẫn ít được chú ý so với lên men
dịch thể. Sự khác nhau cơ bản nhất giữa các quá trình lên men này là ở một số yếu tố như độ
ẩm tối ưu, sự hình thành gradient nhiệt độ, gradient dinh dưỡng, các sản phẩm lên men, cơ

8
chế hình thành bào tử cũng như khả năng sinh enzyme và cơ chế hình thành các chất trao đổi
thứ cấp trong quá trình lên men như các chất kháng sinh, acit citric và các chất tạo mùi.
Các cơ chất được sử dụng trong lên men xốp được thu thập từ các phế phẩm công
nông nghiệp (ví dụ, trấu lúa mì hoặc lúa mì, bã mía, bã cà phê, bã nho, cùi dừa,…) hoặc các
nguyên liệu trơ (các chất nhựa trong trao đổi ion) cũng được sử dụng. Cơ chất sử dụng gần
như không phải xử lý trước khi dùng, trường hợp phải tiền xử lý thì chỉ cần rửa qua bằng
nước.
Các ưu điểm chính của kỹ thuật lên men xốp:
- Môi trường nuôi cấy đơn giản. Một số cơ chất có thể được sử dụng trực tiếp làm môi
trường xốp hoặc được cho thêm một số dưỡng chất trước khi sử dụng.
- Sản phẩm enzym thu được ở dạng đậm đặc.
- Độ ẩm tối ưu thấp và phần lớn giống cấy được sử dụng trong lên men xốp giảm được
tối đa nguy cơ nhiễm độc vi sinh vật.
- Lượng chất thải sinh ra trong lên men xốp ít hơn so với lên men dịch thể
- Enzyme tạo ra ít nhạy cảm với các chất kìm hãm hoặc ức chế trao đổi chất.
Mặc dù có nhiều ưu điểm vượt trội nhưng kỹ thuật lên men xốp cũng vẫn tồn tại một
số ít nhược điểm như sau:
- Vi sinh vật được dùng trong nghiên cứu bị giới hạn khả năng sinh trưởng vì độ ẩm
của môi trường hạn chế.
- Việc xác định các thông số như độ ẩm, pH, hàm lượng oxy và CO2 tự do là một vấn
đề khó khăn vì thiếu dụng cụ tiến hành.
- Phạm vi ứng dụng kỹ thuật lên men xốp còn chưa được nghiên cứu rộng rãi.
1.4. TINH SẠCH ENZYME XYLANASE
Tinh sạch enzyme là một chuỗi các quá trình liên tiếp nhằm tách riêng một loại
enzyme từ hỗn hợp ban đầu. Tinh sạch enzyme là quá trình thiết yếu để xác định được các
đặc tính về cấu trúc - chức năng, động lực học, cơ chế xúc tác và sự tương tác giữa các loại
enzyme quan trọng. Nguyên liệu khởi đầu được sử dụng cho quá trình tinh sạch thường là một
loại mô sinh học hoặc dịch nuôi cấy vi sinh vật. Quá trình tinh sạch enzyme cho phép tách
chiết một loại enzyme từ một hỗn hợp ban đầu có chứa loại enzyme đó, tách riêng các phần
có bản chất là protein và phần không có bản chất là protein trong hỗn hợp. Quá trình tách
chiết này là một quá trình khá phức tạp. Các bước tách chiết này dựa vào kích thước phân tử,
đặc tính lý hóa, khả năng bám dính và hoạt độ sinh học của từng loại enzyme.
Mục đích của việc tinh sạch enzyme nhằm tối ưu hóa hoạt độ xúc tác của enzyme, tinh
sạch tối đa được enzyme và nghiên cứu các đặc tính của chúng. Tinh sạch enzyme còn được
ứng dụng rộng rãi trong lĩnh vực y tế và công nghiệp.
Muốn tinh sạch được enzyme mà vẫn giữ nguyên được đặc tính của nó thì chúng ta có
thể sử dụng nhiều phương pháp khác nhau, dựa vào nhiệt độ, nồng độ proton (pH), hoặc các
tác nhân hóa học (các muối trung tính (NH4)2SO4, Na2SO4, …).

9
Để tinh sạch được enzyme, người ta thường sử dụng kết hợp đồng thời nhiều kỹ thuật
khác nhau: thu hồi enzyme bằng muối ammonisunfat, sắc ký trao đổi ion, điện di SDS-PAGE
và điện di trên gel hoạt tính.
1.4.1. Tủa enzyme bằng muối ammonisunfat
1.4.2. Tủa enzyme bằng dung môi hữu cơ
1.4.3. Sắc ký
Sắc ký là quá trình tách dựa trên sự phân bố liên tục các cấu tử chất phân tích trên 2 pha: Một
pha đứng yên có khả năng hấp thụ chất phân tích gọi là pha tĩnh, một pha di chuyển qua pha
tĩnh gọi là pha động; do các cấu tử chất phân tích có ái lực khác nhau với pha tĩnh, chúng di
chuyển với tốc độ khác nhau. Các chất có ái lực lớn với pha tĩnh sẽ chuyển động chậm hơn
qua hệ thống sắc ký so với các chất tương tác yếu hơn với pha này. Nhờ đặc điểm này mà
người ta có thể tách phân đoạn các chất nhờ quá trình sắc ký.
1.4.3.1. Sắc ký lọc gel
1.4.3.2. Sắc ký trao đổi ion
1.4.3.3. Sắc ký ái lực
1.4.3.4. Sắc ký tương tác kỵ nước
1.4.3.5. Các kỹ thuật sắc ký lỏng hiệu năng cao - HPLC
1.4.4. Điện di
Điện di protein (Protein Electrophorus) là dùng dòng điện 1 chiều để dịch chuyển các
tiểu phân tử protein trên môi trường gel dựa trên lực Lorenz.
Những phân tử khác nhau sẽ dịch chuyển với tốc độ khác nhau mà ta có thể quan sát được
trên điện di đồ. Tốc độ di chuyển này được quyết định bởi 3 yếu tố: điện tích, kích thước và
khối lượng phân tử.
1.4.4.1. Điện di trên gel agarose
1.4.4.2. Điện di trên gel polyacrylamide
CHƢƠNG 2 - NGUYÊN LIỆU VÀ PHƢƠNG PHÁP NGHIÊN CỨU
2.1 CHỦNG VI SINH VẬT, MÔI TRƢỜNG VÀ THIẾT BỊ NGHIÊN CỨU
2.1.1. Chủng vi sinh vật
- Sử dụng 26 chủng vi sinh vật đươc lưu giữ tại Viện Vi sinh vật và Công nghệ Sinh
học - Đại học Quốc gia Hà Nội.
2.1.2. Môi trƣờng nghiên cứu
2.1.2.1. Môi trường nhân giống (g/l)
- Xạ khuẩn: sử dụng môi trường YG (cao nấm men -10, Glucose-10, Nước - 1, pH-7).
Khử trùng 121oC trong 15 phút.
- Vi khuẩn: sử dụng môi trường canh thang (Peptone - 5, cao thịt - 3, NaCl 5, Nước -
1, pH 7.0). Khử trùng 121oC trong 15 phút.
2.1.2.2. Môi trường nuôi dịch thể

10
a. Môi trường nuôi vi khuẩn: gồm 5 môi trường ký hiệu từ V1- V5. Khử trùng ở 1210C, 15
phút.
b. Môi trường nuôi xa khuân : gôm 5 môi trương ky hiêu tư X 1 tơi X5. Khử trùng ở 1210C,
15 phút.
2.1.2.3. Môi trường kiểm tra hoạt độ enzyme
Bản thạch chứa cơ chất (g/l): Xylan - 1, agar - 16, đệm Tris-HCl 50 mM, pH 7.0, khử
trùng 121oC trong 15 phút.
2.1.2.4. Môi trường nuôi xốp
Nguyên liêu cho môi trương xôp gôm co : bôt ngô, cám gạo , gạo lức, malt, đâu tương
(các cơ chất này có thể dễ dàng mua được trên thi trương).
2.1.3. Thiết bị nghiên cứu
Các thiết bị và máy móc của Bảo tàng giống chuẩn vi sinh vật, Viện Vi sinh vật và Công nghệ
Sinh học -Đại học Quốc gia Hà Nội:
- Máy đọc trình tự ADN-3100 Avant Genetic Analyzer (Mỹ).
- Máy khuếch đại gen- Gene Amp PCR System 9700 (Mỹ).
- Máy ly tâm lạnh- Centrifuge C30P Satorius Group (Đức).
- Kính hiển vi BX 51-Olympus (Nhật).
- Máy soi gel Bio-Rat Gen doc (Mỹ).
- Máy cô mẫu chân không- AES 1010 Speed Vac System-Thermo Savant(Mỹ).
- Máy đo quang phổ- UV 1600 (Thụy Điển).
- Máy lắc ổn nhiệt (B. Braun Biotech International -Mỹ).
- Máy đông khô (Dura-Dry, FTS Systems, Inc. -Mỹ)…
- Máy rung (Eyela Cute Mixer-1000 -Nhật).
- Máy ly tâm (A15-B.Braun-Đức).
- Máy cô quay chân không (RV05-ST, KIKA Labortechnik -Đức).
- Máy sắc ký lỏng cao áp HPLC (Agilent Technologies -Mỹ).
- Bể ổn nhiệt (Ecoline RE 104 -Nhật).
- Thiết bị điện di nằm cho ADN (Bio Rad -Mỹ).
- Bộ chạy điện di đứng cho proteing (Biorad - Mỹ).
- Cột sắc ký trao đổi ion,…
2.2. PHƢƠNG PHÁP NGHIÊN CỨU
2.2.1. Phƣơng pháp xác định hoạt tính xylanase
2.2.1.1. Phương pháp định tính (khuếch tán trên thạch)
Vi khuẩn và xạ khuẩn được nuôi cấy lắc 200 vòng/phút với các điều kiện nhiệt độ,
môi trường và pH thích hợp. Sau các khoảng thời gian cần thiết, lấy dịch li tâm với tốc độ
12000 vòng/phút trong 10 phút. Thu dịch enzyme nhỏ vào bản thạch chứa cơ chất đã khoan
lỗ. Đặt trong tủ ấm (500C). Sau 24 giờ, dùng thuốc thử Công gô đỏ nhuộm bản thạch, sau đó
rửa bằng dung dịch NaCl 1M và đọc kết quả. Hoạt tính xylanase được xác định bằng kích
thước vòng phân giải (VPG) (D-d, mm), trong đó, D là kích thước vòng phân giải, d là kích
thước lỗ đục.
11
2.2.1.2. Phương pháp định lượng
Xác định hoạt độ xylanasebằng phương pháp DNS theo Miller (1999). Hoạt độ
xylanase được xác định theo phương pháp của Bailey, 1992.
Phương pháp DNS
a. Chuẩn bị hóa chất
b. Cách tiến hành
c. Dựng đường chuẩn
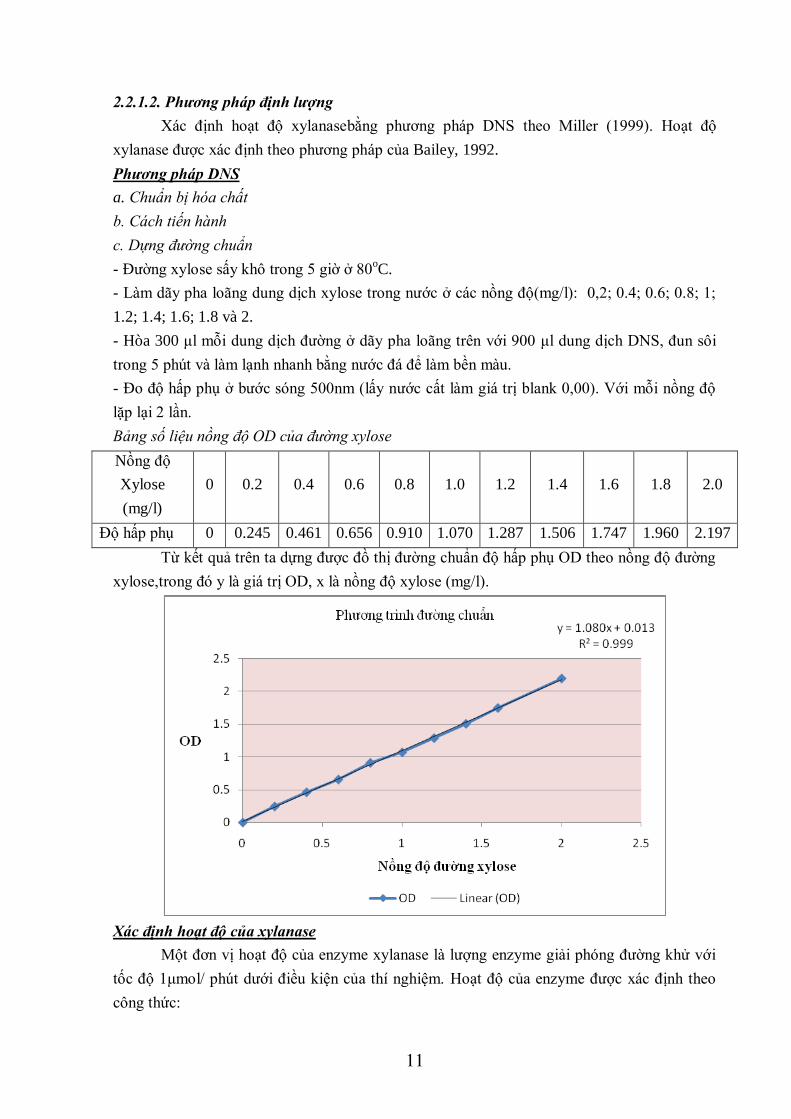
- Đường xylose sấy khô trong 5 giờ ở 80oC.
- Làm dãy pha loãng dung dịch xylose trong nước ở các nồng độ(mg/l): 0,2; 0.4; 0.6; 0.8; 1;
1.2; 1.4; 1.6; 1.8 và 2.
- Hòa 300 μl mỗi dung dịch đường ở dãy pha loãng trên với 900 μl dung dịch DNS, đun sôi
trong 5 phút và làm lạnh nhanh bằng nước đá để làm bền màu.
- Đo độ hấp phụ ở bước sóng 500nm (lấy nước cất làm giá trị blank 0,00). Với mỗi nồng độ
lặp lại 2 lần.
Bảng số liệu nồng độ OD của đường xylose
Nồng độ
Xylose
(mg/l)
0 0.2 0.4 0.6 0.8 1.0 1.2 1.4 1.6 1.8 2.0
Độ hấp phụ 0 0.245 0.461 0.656 0.910 1.070 1.287 1.506 1.747 1.960 2.197
Từ kết quả trên ta dựng được đồ thị đường chuẩn độ hấp phụ OD theo nồng độ đường
xylose,trong đó y là giá trị OD, x là nồng độ xylose (mg/l).
Xác định hoạt độ của xylanase
Một đơn vị hoạt độ của enzyme xylanase là lượng enzyme giải phóng đường khử với
tốc độ 1μmol/ phút dưới điều kiện của thí nghiệm. Hoạt độ của enzyme được xác định theo
công thức:

12
X × 1000 × 5 × 20
Hoạt độ U/ml hoặc (U/g) = ———————
150 × 10
Trong đó: - X là lượng đường khử tính theo đường chuẩn
- X × 1000 là lượng đường khử theo đường chuẩn đổi ra µl
(hoặc µg)
- 5 là lượng đường khử trong 1ml (hoặc 1g) phản ứng
- 20 là hoạt độ xylanase tính theo 1ml enzyme
- 150 là khối lượng phân tử của xylose
- 10 là thời gian ủ của phản ứng (phút)
2.2.2. Tuyển chọn chủng
2.2.3. Phƣơng pháp xác định khả năng sinh trƣởng
2.2.4. Phƣơng pháp phân loại
2.2.4.1. Phương pháp phân loại vi khuẩn dựa vào đọc trình tự ADN
a. Tách chiết ADN của vi khuẩn
b. Phản ứng khuếch đại ADN
c. Tinh sạch sản phẩm PCR.
d. Phản ứng khuếch đại ADN cho đọc trình tự
e. Tinh sạch sản phẩm PCR cho đọc trình tự
f. Phân tích trình tự và xây dựng cây phát sinh chủng loại
2.2.4.2. Quan sát hình thái
a. Hình thái tế bào và khuẩn lạc vi khuẩn
b. Quan sát hình thái xạ khuẩn
2.2.5. Nghiên cứu điều kiện nuôi cấy thích hợp cho khả năng tổng hợp xylanase ở các
chủng vi sinh vật nghiên cứu
2.2.5.1. Nghiên cứu điều kiện nuôi cấy giống thích hợp cho khả năng tổng hợp xylanase ở
các chủng nghiên cứu
a. Môi trường thích hợp
b. Nhiệt độ thích hợp
c. pH thích hợp
d. Thời gian thích hợp cho giống khởi động
2.2.5.2. Nghiên cứu điều kiện nuôi cấy xốp thích hợp cho khả năng tổng hợp xylanase ở
các chủng nghiên cứu
a. Lựa chọn cơ chất
b. Lựa chọn thời gian nuôi cấy
c. Lựa chọn độ ẩm
d. Lựa chọn tỷ lệ giống cấy
e. Lựa chọn nguồn cacbon bổ sung
f. Lựa chọn nguồn nitơ bổ sung

13
g. Lựa chọn nguồn khoáng
2.2.6. Thu hồi enzyme
2.2.6.1. Chiết enzyme
2.2.6.2. Thu hồi enzyme bằng các dung môi hữu cơ
2.2.6.3. Thu hồi enzyme bằng muối ammonisunfat
2.2.7. Tinh sạch xylanase của chủng 118
2.2.7.1. Tủa enzyme trong dung môi hữu cơ
2.2.7.2. Tủa enzyme trong muối ammonisunfat
2.2.7.3. Sắc ký trao đổi ion
2.2.8. Điện di
2.2.8.1. Điện di trên gel SDS-PAGE
2.2.8.2. Điện di trên gel hoạt tính
2.2.9. Nghiên cứu đặc tính enzyme
2.2.9.1. pH thích hợp cho hoạt động của enzyme
2.2.9.2.Nhiệt độ thích hợp cho hoạt động của enzyme
2.2.9.3. Khả năng bền nhiệt của enzyme
2.2.9.4. Ảnh hưởng của các ion kim loại đến hoạt độ enzyme
Chƣơng 3. KẾT QUẢ VÀ THẢO LUẬN
3.1. KẾT QUẢ TUYỂN CHỌN
3.1.1. Lựa chọn các chủng có hoạt độ xylanase cao
3.1.2. Lựa chọn các chủng vi sinh vật bền nhiệt có hoạt tính xylanase cao
Từ 26 chúng ban đầu chúng tôi chọn ra được hai chủng B2H2 và 118, là các chủng bền
nhiệt có hoạt độ xylanase cao nhất để tiến hành nghiên cứu sâu hơn.
3.2. PHÂN LOẠI
3.2.1. Chủng B2H2
3.2.1.1. Phân tích trình tự ADNr 16S
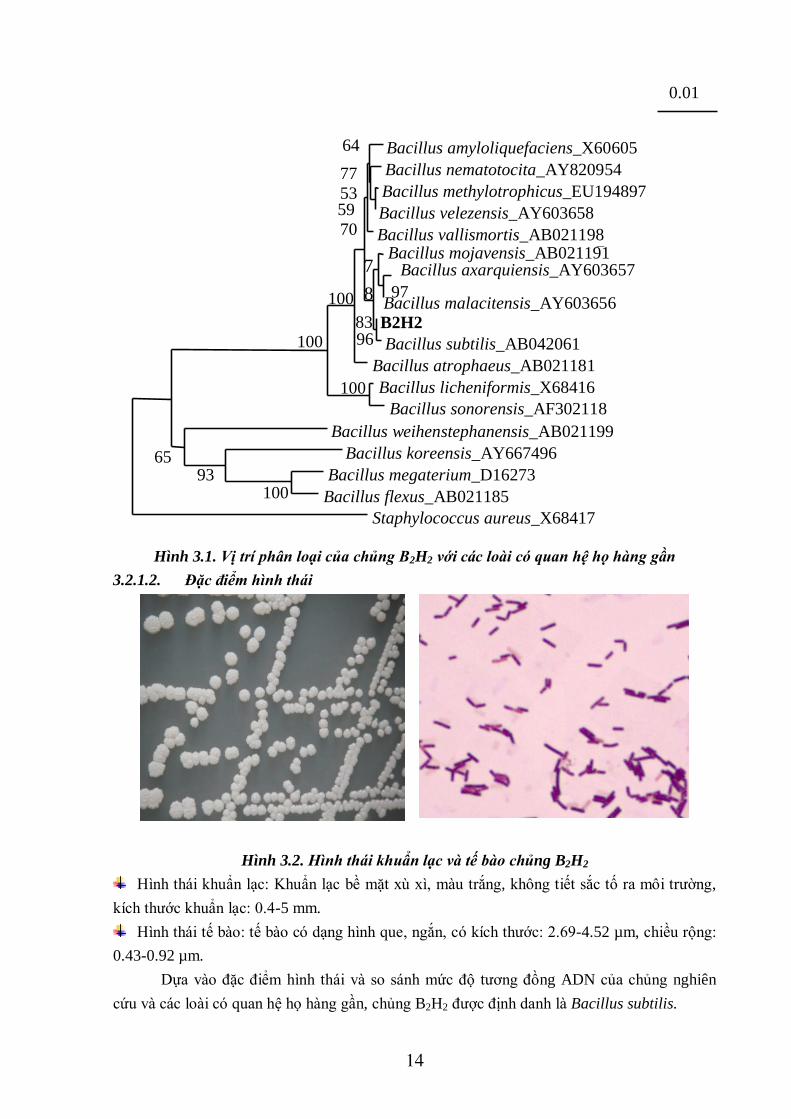
14
Hình 3.1. Vị trí phân loại của chủng B2H2 với các loài có quan hệ họ hàng gần
3.2.1.2. Đặc điểm hình thái
Hình 3.2. Hình thái khuẩn lạc và tế bào chủng B2H2
Hình thái khuẩn lạc: Khuẩn lạc bề mặt xù xì, màu trắng, không tiết sắc tố ra môi trường,
kích thước khuẩn lạc: 0.4-5 mm.
Hình thái tế bào: tế bào có dạng hình que, ngắn, có kích thước: 2.69-4.52 µm, chiều rộng:
0.43-0.92 µm.
Dựa vào đặc điểm hình thái và so sánh mức độ tương đồng ADN của chủng nghiên
cứu và các loài có quan hệ họ hàng gần, chủng B2H2 được định danh là Bacillus subtilis.
Staphylococcus aureus_X68417
Bacillus flexus_AB021185
Bacillus megaterium_D16273
Bacillus koreensis_AY667496
100
Bacillus weihenstephanensis_AB021199
93
Bacillus sonorensis_AF302118
Bacillus licheniformis_X68416
Bacillus atrophaeus_AB021181
Bacillus subtilis_AB042061
B2H2
Bacillus malacitensis_AY603656
Bacillus axarquiensis_AY603657 Bacillus mojavensis_AB021191
97
96
7
8
Bacillus vallismortis_AB021198
Bacillus velezensis_AY603658
Bacillus methylotrophicus_EU194897
Bacillus nematotocita_AY820954
59
Bacillus amyloliquefaciens_X60605
53
64
83
77
70
100
100
65
100
Z
0.01

15
3.2.2. Chủng 118
3.2.1.3. Phân tích trình tự ADNr 16S
Hình 3.3 .Vị trí phân loại của chủng XK-118 với các loài có quan hệ họ hàng gần
3.2.1.4. Đặc điểm hình thái
Kitasatosporia setalba_U93332
Streptomyces padanus_AF455813
Streptomyces gougerotii_AB184742
Streptomyces phaeoluteichromatogenes_AJ391814
XK-118
Streptomyces misionensis_EF178678
Streptomyces rajshahiensis_GQ500975
Streptomyces owasiensis_AB184515
88
53 80
83
Streptomyces levis_DQ442517
Streptomyces purpurascens_AB045888
Streptomyces viridochromogenes_DQ442555
Streptomyces ambofaciens_AB184182
Streptomyces flaveolus_AB184764
71
Streptomyces tendae_D63873
Streptomyces tritolerans_DQ345779
Streptomyces violaceoruber_AF503492
Streptomyces coelescens_AF503496
Streptomyces humiferus_AF503491
99
79
99
Streptomyces violaceorubidus_AJ781374
64
64
96
Streptomyces glaucescens_AB184843
Streptomyces pharetrae_AY699792
58
100
Streptomyces brunneogriseus_AB184509
Streptomyces eurythermus_D63870
Streptomyces ansochromogenes_AB184448
65
Streptomyces nodosus_AF114033
Streptomyces achromogenes_AY999846
66
76
56
100
58
55
66
82
0.01
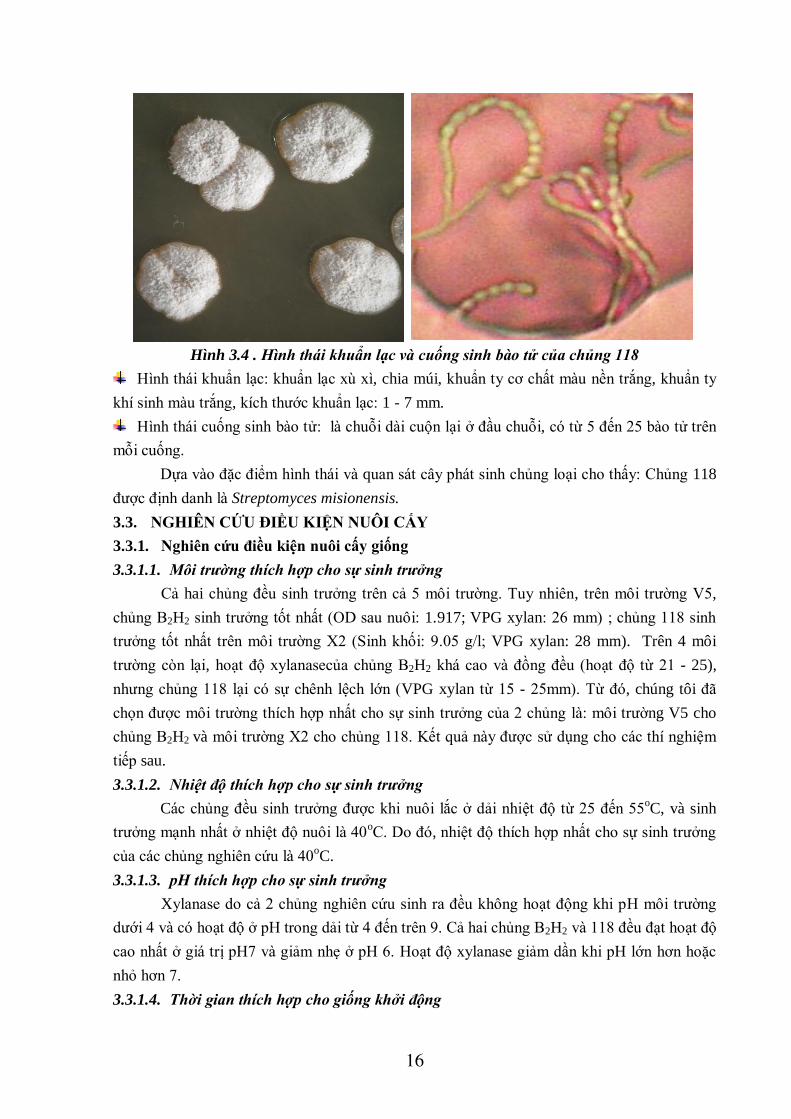
16
Hình 3.4 . Hình thái khuẩn lạc và cuống sinh bào tử của chủng 118
Hình thái khuẩn lạc: khuẩn lạc xù xì, chia múi, khuẩn ty cơ chất màu nền trắng, khuẩn ty
khí sinh màu trắng, kích thước khuẩn lạc: 1 - 7 mm.
Hình thái cuống sinh bào tử: là chuỗi dài cuộn lại ở đầu chuỗi, có từ 5 đến 25 bào tử trên
mỗi cuống.
Dựa vào đặc điểm hình thái và quan sát cây phát sinh chủng loại cho thấy: Chủng 118
được định danh là Streptomyces misionensis.
3.3. NGHIÊN CỨU ĐIỀU KIỆN NUÔI CẤY
3.3.1. Nghiên cứu điều kiện nuôi cấy giống
3.3.1.1. Môi trường thích hợp cho sự sinh trưởng
Cả hai chủng đều sinh trưởng trên cả 5 môi trường. Tuy nhiên, trên môi trường V5,
chủng B2H2 sinh trưởng tốt nhất (OD sau nuôi: 1.917; VPG xylan: 26 mm) ; chủng 118 sinh
trưởng tốt nhất trên môi trường X2 (Sinh khối: 9.05 g/l; VPG xylan: 28 mm). Trên 4 môi
trường còn lại, hoạt độ xylanasecủa chủng B2H2 khá cao và đồng đều (hoạt độ từ 21 - 25),
nhưng chủng 118 lại có sự chênh lệch lớn (VPG xylan từ 15 - 25mm). Từ đó, chúng tôi đã
chọn được môi trường thích hợp nhất cho sự sinh trưởng của 2 chủng là: môi trường V5 cho
chủng B2H2 và môi trường X2 cho chủng 118. Kết quả này được sử dụng cho các thí nghiệm
tiếp sau.
3.3.1.2. Nhiệt độ thích hợp cho sự sinh trưởng
Các chủng đều sinh trưởng được khi nuôi lắc ở dải nhiệt độ từ 25 đến 55oC, và sinh
trưởng mạnh nhất ở nhiệt độ nuôi là 40oC. Do đó, nhiệt độ thích hợp nhất cho sự sinh trưởng
của các chủng nghiên cứu là 40oC.
3.3.1.3. pH thích hợp cho sự sinh trưởng
Xylanase do cả 2 chủng nghiên cứu sinh ra đều không hoạt động khi pH môi trường
dưới 4 và có hoạt độ ở pH trong dải từ 4 đến trên 9. Cả hai chủng B2H2 và 118 đều đạt hoạt độ
cao nhất ở giá trị pH7 và giảm nhẹ ở pH 6. Hoạt độ xylanase giảm dần khi pH lớn hơn hoặc
nhỏ hơn 7.
3.3.1.4. Thời gian thích hợp cho giống khởi động

17
Thời gian nuôi khởi động chủng B2H2 thích hợp nhất là 24h. Sau 24h mật độ tế bào
giảm dần. Thời gian nuôi khởi động chủng 118 thích hợp nhất là 42h, tại đó sinh khối thu
được là cao nhất.
3.3.2. Nghiên cứu điều kiện nuôi cấy xốp
3.3.2.1. Lựa chọn cơ chất
Các chủng vi khuẩn và xạ khuẩn đều sinh trưởng được trên 5 cơ chất xốp. Đối với
chủng vi khuẩn B2H2, hoạt độ xylanasethu được đạt cao nhất trên cơ chất malt (2154 U/g),
tiếp theo là trên cơ chất gạo (587 U/g). Đối với chủng xạ khuẩn 118, hoạt độ xylanase thu
được cao nhất trên cơ chất gạo (2386 U/g ), và thấp nhất trên cám gạo (184 U/g).
3.3.2.2. Lựa chọn độ ẩm thích hợp
Cả hai chủng vi khuẩn và xạ khuẩn đều tổng hợp enzyme xylanase tốt nhất ở độ ẩm
50%. Tuy nhiên ở chủng 118, khi độ ẩm đạt 40 và 60%, hoạt độ xylanase thu được vẫn khá
cao và xấp xỉ bằng hoạt độ ở độ ẩm 50%, còn chủng B2H2 thì hoạt độ xylanase chênh lệch
khá nhiều, khi độ ẩm là 40% hoạt độ xylanase thu được chỉ bằng 61% so với ở độ ẩm 50%.
3.3.2.3. Lựa chọn thời gian nuôi cấy
Đối với cả hai chủng thời gian nuôi cấy tốt nhất là 3 ngày, chủng B2H2 đạt hoạt tính
2154 U/g còn chủng 118 đạt hoạt tính cao nhất ở ngày nuôi cấy thứ 3 là 2181 U/g.
3.3.2.4. Lựa chọn tỷ lệ giống cấy thích hợp
Tỷ lệ giống cấy thích hợp nhất cho cả hai chủng vi khuẩn và xạ khuẩn là ở 15%. Với
tỷ lệ giống cấy lớn hơn hoặc nhỏ hơn, hoạt độ xylanase đều giảm.
3.3.2.5. Lựa chọn nguồn nitơ bổ sung
Đối với chủng vi khuẩn B2H2, tất cả các nguồn nitơ vô cơ như nitrat, nitrit, (NH4)2SO4
đều làm giảm khả năng sinh xylanase; ngược lại, hầu hết nguồn nitơ hữu cơ peptone, cao thịt,
cao men được chọn nghiên cứu (ngoại trừ ure) đều tăng cường khả năng sinh xylanase của
chủng (tăng khoảng 18%). Trong các nguồn nitơ được chọn thì cao thịt giúp tăng khả năng
tổng hợp enzyme xylanase cao nhât , tiêp đo la cao men; Nitrat và nitrit ức chế mạnh nhất khả
năng sinh trưởng và sinh enzyme của chủng này.
Đối với chủng xạ khuẩn 118, khả năng tổng hợp xylanase không chênh lệch nhiều khi
bổ sung thêm các nguồn nitơ khác nhau vào cơ chất nuôi cấy. Các nguồn nitơ tăng cường khả
năng sinh xylanase của chủng 118 là (NH4)2SO4, cao men, pepton và ure. Hoạt độ xylanase
đạt được cao nhất khi bổ sung nguồn nitơ là urê (tăng 20%), tiếp đó là cao thịt (tăng 19%);
Tuy nhiên, khi bổ sung 4% các nguồn nitơ khác như nitrat, nitrit, cao thịt thì hoạt độ xylanase
đều bị giảm, khoảng 15%.
3.3.2.6. Lựa chọn nguồn cacbon bổ sung
Khi bổ sung các nguồn cacbon khác nhau vào cơ chất nuôi cấy của cả 2 chủng vi
khuẩn và xạ khuẩn đã gây ra sự ức chế khả năng tổng hợp enzyme xylanase của 2 chủng
này. Hoạt độ xylanase của chủng vi khuẩn B2H2 giảm từ 2147 U/g xuống còn 1711 U/g khi
thêm 4% glucose vào cơ chất nuôi cấy. Đối với chủng xạ khuẩn 118, khi bổ sung thêm nguồn

18
cacbon hoạt độ xylanase thu được giảm mạnh (hoạt độ xylanasegiảm từ 2379 U/g xuống thấp
nhất còn 1679 (U/g).
3.3.2.7. Lựa chọn loại khoáng thích hợp
Chủng vi khuẩn B2H2 tổng hợp xylanase hiệu quả nhất khi sử dụng khoáng MA1 (hoạt
độ 3637 U/g) và thấp nhất khi sử dụng nước cất (hoạt độ đạt 1027 U/g). Chủng 118 tổng hợp
xylanase tốt nhất khi sử dụng khoáng MA3 (hoạt độ đạt 2609 U/g) và thấp nhất khi sử dụng
khoáng MA4 (1417 U/g).
3.4. THU HỒI ENZYME
3.4.1. Điều kiện thích hợp cho chiết enzyme
Trong 5 loại dịch chiết được sử dụng (đệm citrate 0.05M, pH 7.0; đệm Tris-HCl
0.05M, pH 7.0; Nước; đệm phosphat 0.05M, pH 7.0; dung dịch NaCl 0.09%), hiệu quả chiết
cao nhất ở lần chiết đầu tiên cho chủng B2H2 và 118 lần lượt là đệm phosphat 0.05M, pH7
(90.58%) và nước (81.12 %) hoặc đệm citrate 0.05 M, pH 7 (78.94%) đối với chủng 118. Tiến
hành chiết dịch enzyme lần 2, lần 3 bằng các dịch chiết trên chúng tôi thấy rằng kết quả sau 3
lần chiết tương tự kết quả sau lần chiết thứ nhất, loại dịch chiết cho hiệu quả cao nhất ở lần
chiết 1 sẽ cho kết quả cao nhất sau cả 3 lần chiết.
3.4.2. Thu hồi enzyme
Sau khi chiết dịch enzyme, tiến hành thu hồi enzyme bằng các dung môi hữu
cơ và bằng muối ammonisunfat. Các loại dung môi hữu cơ được sử dụng là cồn 100%, aceton
lạnh. Thu cặn, hòa tan cặn bằng nước, và xác định hoạt độ xylanase thu được.
Khi sử dụng cồn 100%, aceton lạnh và muối ammonisunfat để tủa dịch enzyme của 2
chủng vi sinh vật nuôi cấy trên dịch thể và trên xốp, hoạt độ xylanase thu được cao nhất khi
dịch enzyme được tủa trong aceton. Do đó, chúng tôi sẽ sử dụng aceton để tủa xylanase từ 2
chủng vi khuẩn và xạ khuẩn. Dịch enzyme sau khi tủa sẽ được dùng cho quá trình tinh sạch.
3.5. TINH SẠCH ENZYME
3.5.1. Tinh sạch enzyme xylanase chủng 118 theo phƣơng pháp sắc ký trao đổi ion
Chủng 118 được nuôi trên môi trường dịch thể X2. Sau 2 ngày, chúng tôi tiến hành thu
dịch enzyme, ly tâm ở 12.000 vòng/phút trong 10 phút, 4oC và tủa aceton như đã nêu trong
phần phương pháp.
Sử dụng mẫu enzyme thu được bằng phương pháp tủa trong aceton, chúng tôi tra 2ml
lên cột sắc ký trao đổi ion sepharose DEAE. Sau khi rửa cột bằng đệm 20mM Tris-HCl pH
8.0 với thể tích gấp 5 lần thể tích cột, chúng tôi tiến hành thôi protein bằng dung dịch NaCl
1000mM. Các phân đoạn protein thu được đem xác định hoạt độ xylanase. Kết quả được thể
hiện trong hình sau:
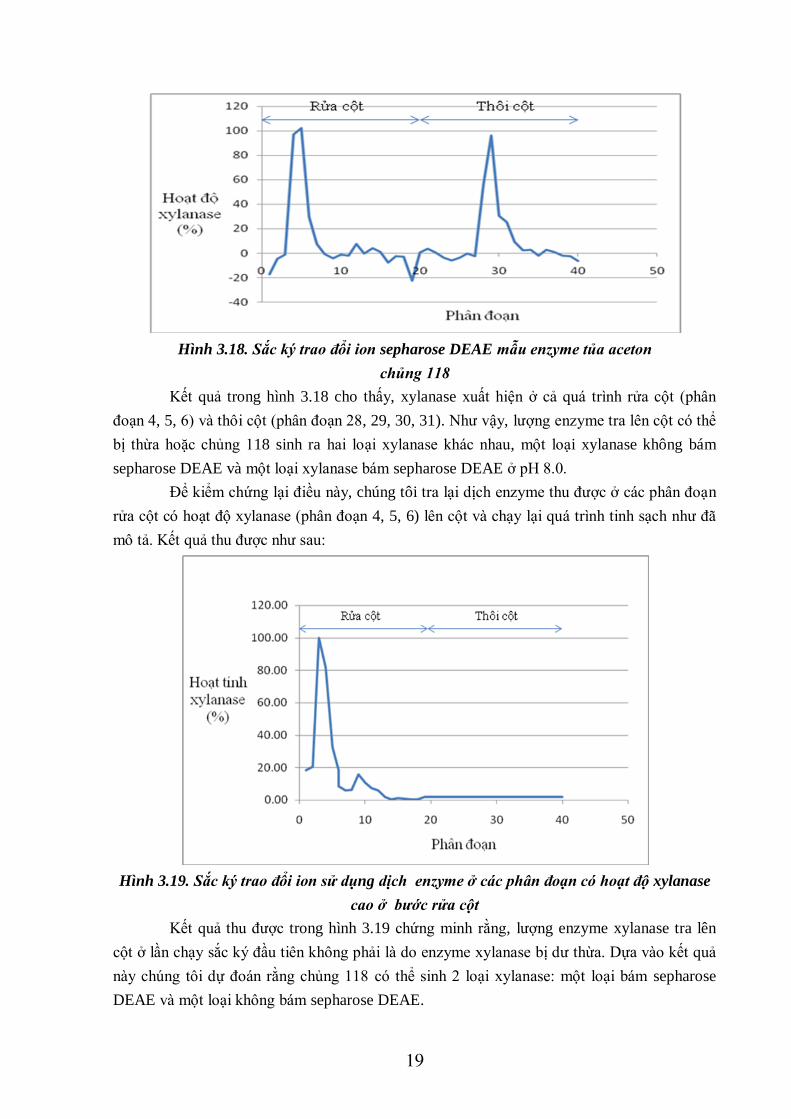
19
Hình 3.18. Sắc ký trao đổi ion sepharose DEAE mẫu enzyme tủa aceton
chủng 118
Kết quả trong hình 3.18 cho thấy, xylanase xuất hiện ở cả quá trình rửa cột (phân
đoạn 4, 5, 6) và thôi cột (phân đoạn 28, 29, 30, 31). Như vậy, lượng enzyme tra lên cột có thể
bị thừa hoặc chủng 118 sinh ra hai loại xylanase khác nhau, một loại xylanase không bám
sepharose DEAE và một loại xylanase bám sepharose DEAE ở pH 8.0.
Để kiểm chứng lại điều này, chúng tôi tra lại dịch enzyme thu được ở các phân đoạn
rửa cột có hoạt độ xylanase (phân đoạn 4, 5, 6) lên cột và chạy lại quá trình tinh sạch như đã
mô tả. Kết quả thu được như sau:
Hình 3.19. Sắc ký trao đổi ion sử dụng dịch enzyme ở các phân đoạn có hoạt độ xylanase
cao ở bước rửa cột
Kết quả thu được trong hình 3.19 chứng minh rằng, lượng enzyme xylanase tra lên
cột ở lần chạy sắc ký đầu tiên không phải là do enzyme xylanase bị dư thừa. Dựa vào kết quả
này chúng tôi dự đoán rằng chủng 118 có thể sinh 2 loại xylanase: một loại bám sepharose
DEAE và một loại không bám sepharose DEAE.

20
Tiếp theo, chúng tôi tiến hành kiểm tra hoạt tính xylanase trên gel hoạt tính. Dịch
enzyme xylanase thô và dịch enzyme tủa aceton được xử lý với 2-Mercaptoethanol (ME) hoặc
không được xử lý với ME được tra vào các giếng điện di, các mẫu enzyme này không qua xử
lý nhiệt. Kết quả được trình bày trong hình 3.20:
Hình 3.20. Điện di đồ trên gel hoạt tính các mẫu enzyme xylanase
1- Mẫu enzyme thô có ME; 2- Mẫu enzyme thô không có ME; 3- Mẫu enzyme tủa aceton
có ME; 4- Mẫu enzyme tủa aceton không có ME.
Từ hình 3.20 ta thấy- ở cả mẫu xylanase có ME và không có ME- chúng tôi đều thấy
xuất hiện 3 dải băng sáng giống nhau. Điều này cho thấy ME không ảnh hưởng đến xylanase
của chủng 118.
Mặt khác, quan sát bản gel điện di ta thấy, các băng sáng rất rộng, không tạo thành
vạch rõ rệt. Điều này có thể giải thích là do mẫu enzyme tra vào các giếng điện di không được
xử lý nhiệt nên protein chưa bị biến tính hoàn toàn, do đó, cùng một phân tử protein có thể tồn
tại dưới các dạng cấu trúc không gian khác nhau, mỗi cấu trúc không gian này tích điện âm
khác nhau trong môi trường có SDS. Vì vậy, khi điện di các cấu trúc không gian này sẽ bị
phân tán nên không nhìn rõ vạch trên bản gel.
Tiếp theo, để biến tính hoàn toàn protein và kiểm tra hoạt tính xylanase, chúng tôi tiến
hành xử lý nhiệt các mẫu enzyme tủa aceton ở các nhiệt độ khác nhau, từ 55, 65, 75, 85 và
95oC trong 4 phút. Mẫu enzyme sau đó được xử lý tiếp với ME và tra vào các giếng điện di
trên gel hoạt tính. Kết quả thu được như trong hình 3.21:
Hình 3.21. Điện di trên gel hoạt tính mẫu enzyme xylanase tủa aceton
1 2 3 4
1 2 3 4 5 6
E2
E1
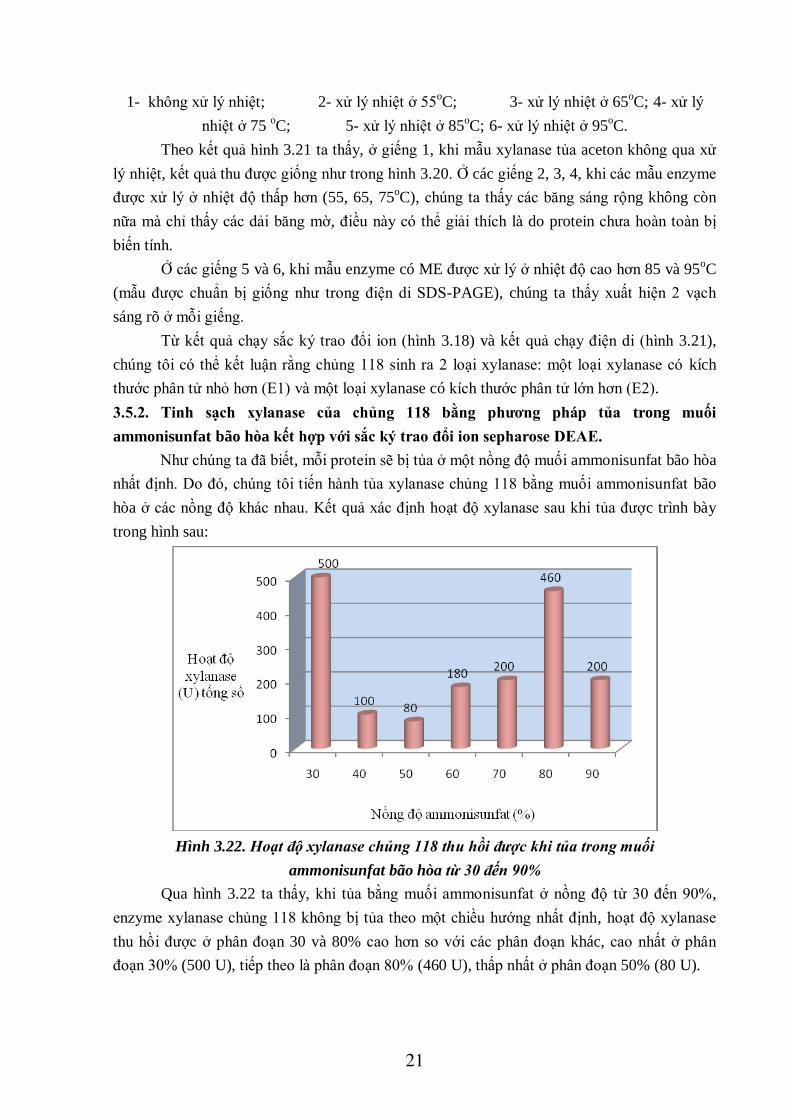
21
1- không xử lý nhiệt; 2- xử lý nhiệt ở 55oC; 3- xử lý nhiệt ở 65
oC; 4- xử lý
nhiệt ở 75 oC; 5- xử lý nhiệt ở 85
oC; 6- xử lý nhiệt ở 95
oC.
Theo kết quả hình 3.21 ta thấy, ở giếng 1, khi mẫu xylanase tủa aceton không qua xử
lý nhiệt, kết quả thu được giống như trong hình 3.20. Ở các giếng 2, 3, 4, khi các mẫu enzyme
được xử lý ở nhiệt độ thấp hơn (55, 65, 75oC), chúng ta thấy các băng sáng rộng không còn
nữa mà chỉ thấy các dải băng mờ, điều này có thể giải thích là do protein chưa hoàn toàn bị
biến tính.
Ở các giếng 5 và 6, khi mẫu enzyme có ME được xử lý ở nhiệt độ cao hơn 85 và 95oC
(mẫu được chuẩn bị giống như trong điện di SDS-PAGE), chúng ta thấy xuất hiện 2 vạch
sáng rõ ở mỗi giếng.
Từ kết quả chạy sắc ký trao đổi ion (hình 3.18) và kết quả chạy điện di (hình 3.21),
chúng tôi có thể kết luận rằng chủng 118 sinh ra 2 loại xylanase: một loại xylanase có kích
thước phân tử nhỏ hơn (E1) và một loại xylanase có kích thước phân tử lớn hơn (E2).
3.5.2. Tinh sạch xylanase của chủng 118 bằng phƣơng pháp tủa trong muối
ammonisunfat bão hòa kết hợp với sắc ký trao đổi ion sepharose DEAE.
Như chúng ta đã biết, mỗi protein sẽ bị tủa ở một nồng độ muối ammonisunfat bão hòa
nhất định. Do đó, chúng tôi tiến hành tủa xylanase chủng 118 bằng muối ammonisunfat bão
hòa ở các nồng độ khác nhau. Kết quả xác định hoạt độ xylanase sau khi tủa được trình bày
trong hình sau:
Hình 3.22. Hoạt độ xylanase chủng 118 thu hồi được khi tủa trong muối
ammonisunfat bão hòa từ 30 đến 90%
Qua hình 3.22 ta thấy, khi tủa bằng muối ammonisunfat ở nồng độ từ 30 đến 90%,
enzyme xylanase chủng 118 không bị tủa theo một chiều hướng nhất định, hoạt độ xylanase
thu hồi được ở phân đoạn 30 và 80% cao hơn so với các phân đoạn khác, cao nhất ở phân
đoạn 30% (500 U), tiếp theo là phân đoạn 80% (460 U), thấp nhất ở phân đoạn 50% (80 U).
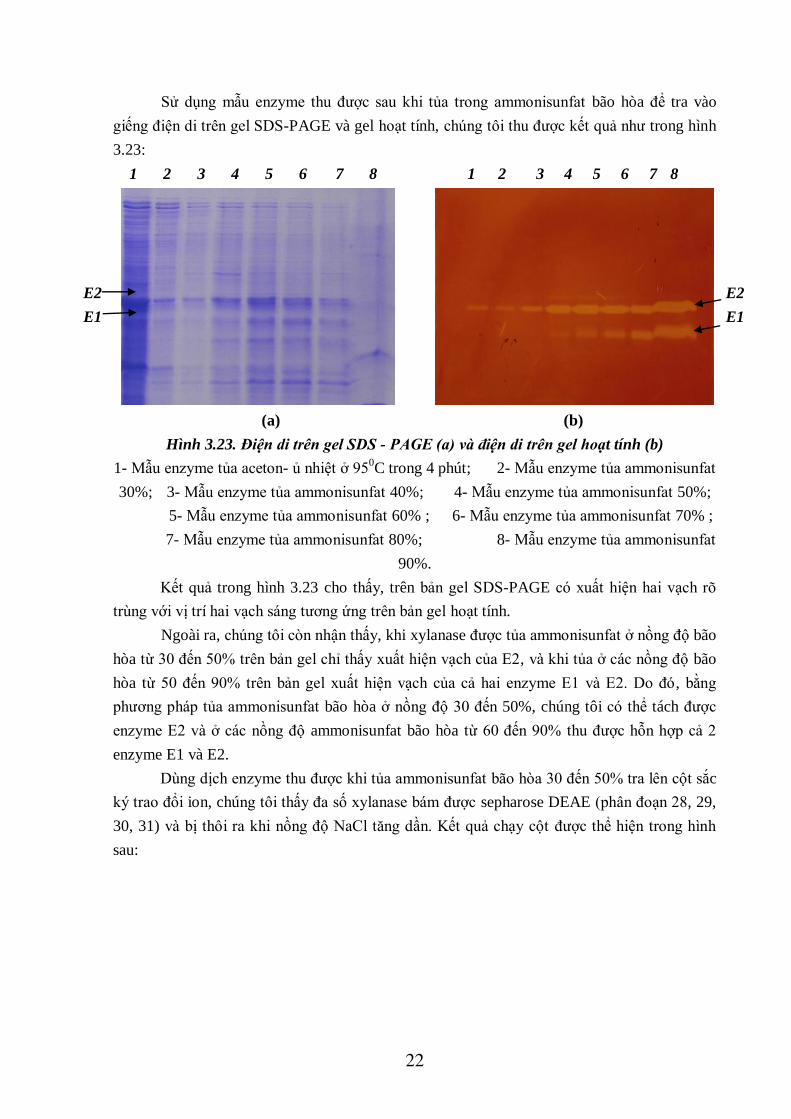
22
Sử dụng mẫu enzyme thu được sau khi tủa trong ammonisunfat bão hòa để tra vào
giếng điện di trên gel SDS-PAGE và gel hoạt tính, chúng tôi thu được kết quả như trong hình
3.23:
1 2 3 4 5 6 7 8 1 2 3 4 5 6 7 8
E2
E1
E2
E1
(a) (b)
Hình 3.23. Điện di trên gel SDS - PAGE (a) và điện di trên gel hoạt tính (b)
1- Mẫu enzyme tủa aceton- ủ nhiệt ở 950C trong 4 phút; 2- Mẫu enzyme tủa ammonisunfat
30%; 3- Mẫu enzyme tủa ammonisunfat 40%; 4- Mẫu enzyme tủa ammonisunfat 50%;
5- Mẫu enzyme tủa ammonisunfat 60% ; 6- Mẫu enzyme tủa ammonisunfat 70% ;
7- Mẫu enzyme tủa ammonisunfat 80%; 8- Mẫu enzyme tủa ammonisunfat
90%.
Kết quả trong hình 3.23 cho thấy, trên bản gel SDS-PAGE có xuất hiện hai vạch rõ
trùng với vị trí hai vạch sáng tương ứng trên bản gel hoạt tính.
Ngoài ra, chúng tôi còn nhận thấy, khi xylanase được tủa ammonisunfat ở nồng độ bão
hòa từ 30 đến 50% trên bản gel chỉ thấy xuất hiện vạch của E2, và khi tủa ở các nồng độ bão
hòa từ 50 đến 90% trên bản gel xuất hiện vạch của cả hai enzyme E1 và E2. Do đó, bằng
phương pháp tủa ammonisunfat bão hòa ở nồng độ 30 đến 50%, chúng tôi có thể tách được
enzyme E2 và ở các nồng độ ammonisunfat bão hòa từ 60 đến 90% thu được hỗn hợp cả 2
enzyme E1 và E2.
Dùng dịch enzyme thu được khi tủa ammonisunfat bão hòa 30 đến 50% tra lên cột sắc
ký trao đổi ion, chúng tôi thấy đa số xylanase bám được sepharose DEAE (phân đoạn 28, 29,
30, 31) và bị thôi ra khi nồng độ NaCl tăng dần. Kết quả chạy cột được thể hiện trong hình
sau:
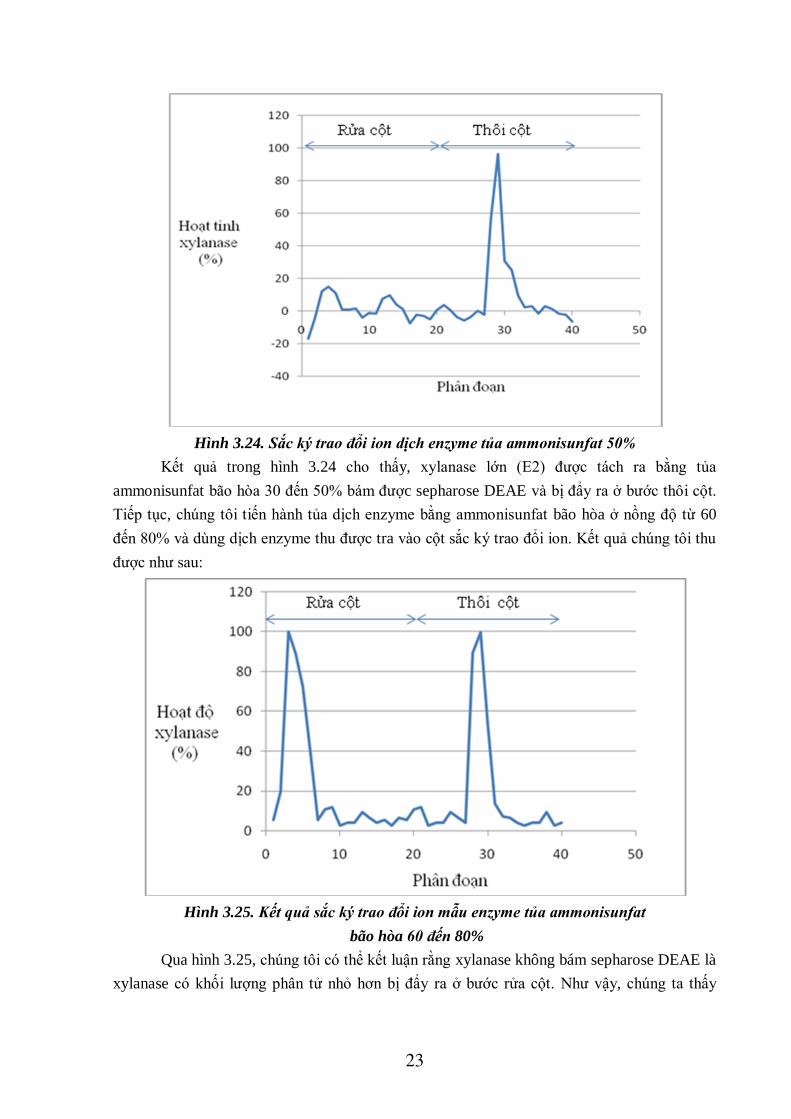
23
Hình 3.24. Sắc ký trao đổi ion dịch enzyme tủa ammonisunfat 50%
Kết quả trong hình 3.24 cho thấy, xylanase lớn (E2) được tách ra bằng tủa
ammonisunfat bão hòa 30 đến 50% bám được sepharose DEAE và bị đẩy ra ở bước thôi cột.
Tiếp tục, chúng tôi tiến hành tủa dịch enzyme bằng ammonisunfat bão hòa ở nồng độ từ 60
đến 80% và dùng dịch enzyme thu được tra vào cột sắc ký trao đổi ion. Kết quả chúng tôi thu
được như sau:
Hình 3.25. Kết quả sắc ký trao đổi ion mẫu enzyme tủa ammonisunfat
bão hòa 60 đến 80%
Qua hình 3.25, chúng tôi có thể kết luận rằng xylanase không bám sepharose DEAE là
xylanase có khối lượng phân tử nhỏ hơn bị đẩy ra ở bước rửa cột. Như vậy, chúng ta thấy

24
rằng có thể tách được 2 loại xylanase từ chủng 118 bằng phương pháp tủa trong
ammonisunfat bão hòa.
Để xác định trọng lượng phân tử của hai loại xylanase từ chủng 118, chúng tôi tiến
hành thí nghiệm chạy điện di trên gel SDS-PAGE. Các mẫu enzyme tủa ammonisunfat 80%
và enzyme E1, E2 được tra vào giếng điện di (thứ tự như trong hình 3.26).
Như vậy, bằng phương pháp tủa ammonisunfat bão hòa 30 đến 80% kết hợp sắc ký trao
đổi ion sepharose DEAE chúng tôi đã tách được hai loại xylanase từ chủng 118: một loại
xylanase bám sepharose DEAE có kích thước phân tử là 36 kDa và một loại xylanase không
bám sepharose DEAE có kích thước phân tử là 22 kDa (hình 3.26).
3.5. ĐẶC TÍNH ENZYME
Sau khi tách được 2 loại xylanase từ chủng 118 chúng tôi tiến hành xác định đặc tính
của 2 loại enzyme này.
3.6.1. Đặc tính enzyme xylanase từ chủng 118
3.6.1.1. pH thích hợp
Xylanase thô không hoạt động khi pH môi trường dưới 4 và có hoạt động ở pH trong
dải từ 4 đến 9 cao nhất ở pH6 và pH7. Đối với 2 enzyme xylanase tinh sạch (E1 và E2), E1 có
dải pH rộng hơn E2 và hoạt độ xylanaseđạt cực đại ở pH 6, và giảm nhẹ ở giá trị pH7. Đối với
các giá trị pH cao hơn (pH8 và pH9) hoạt độ của enzyme giảm mạnh, từ 92U/ml xuống 19
U/ml; Trong khi đó, hoạt độ của E2 cao nhất ở pH5. Ở pH6 và pH7 hoạt độ của enzyme giảm
tuy nhiên không giảm mạnh như E1. Ở pH3 và dưới 3 enzyme E2 không có hoạt động.
3.6.1.2. Nhiệt độ thích hợp
Hoạt độ của xylanase thô và xylanase tinh sạch đều tăng dần khi nhiệt độ phản ứng tăng
từ 35 đến 50oC, tuy nhiên nhiệt độ thích hợp nhất cho hoạt động của cả 2 loại xylanase là
E2 (36 kDa)
E1 (22 kDa)
Hình 3.26. Điện di SDS-PAGE
M- marker (BSM0661; Biobasic);
1- Mẫu enzyme tủa
ammonisunfat 80%; 2- Dịch
xylanase E1 và 3- Dịch Xylanase
E2 thu được từ sắc ký trao đổi ion
DEAE.
M 1 2 3
10 kDa
40 kDa
30 kDa
20 kDa
50 kDa
70 kDa
100 kDa

25
50oC, hay nói cách khác hiệu suất phân giải xylan của enzyme xylanase đạt cực đại ở nhiệt độ
50oC. Ở các nhiệt độ cao hơn nhiệt độ hoạt động tối ưu, hoạt độ của cả 3 enzyme đều giảm.
3.6.1.3. Ảnh hưởng của các ion kim loại
Đối với enzyme xylanase thô, ion Fe2+
có khả năng làm tăng hoạt độ của xylanase
(tăng 28% so với ban đầu), trong khi tất cả các ion kim loại khác đều ức chế hoạt động của
enzyme này. Đối với 2 enzyme xylanase đã tinh sạch, Mn2+
và Fe2+
kích thích khả năng hoạt
động của chúng. Đối với E1, ion Fe2+
tăng cường hoạt động của xylanase nhiều nhất, tiếp theo
là Na+, Ca
2+ và Mn
2+, trong khi đó EDTA ức chế mạnh nhất. Đối với E2, ion Mn
2+ tăng
cường hoạt độ xylanase nhiều nhất, trong khi đó hầu hết các ion kim loại khác đều ức chế
hoạt động của enzyme này, đặc biệt Cu2+
ức chế mạnh nhất.
3.6.1.4. Khả năng chịu nhiệt của enzyme
Khi xử lý dịch enzyme xylanase từ chủng 118 ở 60oC trong các khoảng thời gian khác
nhau 10 phút, 20 phút và 30 phút, hoạt độ của cả enzyme xylanase thô và tinh sạch đều giảm.
Tuy nhiên, E2 có xu hướng bền nhiệt hơn so với enzyme xylanase thô và E1. Ở 60oC trong
10 phút, hoạt độ xylanase E2 còn 93%, trong khi enzyme E1 chỉ còn 50% và enzyme xylanase
thô chỉ còn 70%. Như vậy, enzyme E2 gần như vẫn giữ được hoạt độ khá cao so với dịch
enzyme ban đầu ở 60oC.
Hoạt độ của cả 2 enzyme khi được xử lý ở các nhiệt độ 70oC, 80
oC, 90
oC và 100
oC
giảm đi đáng kể. Ở 70oC, hoạt độ xylanase E1 và E2 còn lại lần lượt là 20% và 38% so với
mẫu enzyme không xử lý nhiệt. Hoạt độ cả hai enzyme gần như không còn ở 100oC.
3.6.2. Đặc tính xylanase từ B2H2
3.6.2.1. pH thích hợp
Xylanase do chủng B2H2 sinh ra không hoạt động ở pH dưới 4 và có hoạt động ở pH
trong dải từ 5 đến trên 9, và hoạt động tốt nhất ở pH 6. Nhìn chung, xylanase của chủng B2H2
hoạt động được trong dải pH từ 5 đến 9.
3.5.2.2. Nhiệt độ
Nhiệt độ thích hợp nhất cho hoạt động của enzyme xylanase của chủng B2H2 khi định
tính và định lượng đều là 50oC (Định tính: 33 mm, hoạt độ: 232 U/ml), hay nói cách khác
enzyme xylanase hoạt động tốt nhất ở nhiệt độ 50oC.
3.5.2.3. Bền nhiệt
Khi xử lý enzyme ở 60oC trong 10 phút, hoạt độ xylanase của chủng B2H2 giảm 10%
so với hoạt độ xylanaseban đầu. Tuy nhiên, hoạt độ của enzyme giảm mạnh khi xử lý ở nhiệt
độ cao hơn 70oC. Ở 80
oC, hoạt độ xylanase giảm hơn 70% so với hoạt độ xylanase ban đầu. Ở
100oC hoạt độ xylanase gần như không còn.
3.5.2.4. Ảnh hưởng của ion kim loại
Một số ion kim loại có khả năng tăng cường hoạt động của enzyme xylanase trong khi
một số lại ức chế. Ion Ca2+
tăng cường mạnh nhất hoạt độ xylanase (tăng 66% so với dịch
enzyme ban đầu), tiếp đó là Mn2+
và ion Fe2+
, ion Mg2+
làm giảm hoạt độ của enzyme mạnh
nhất (giảm 44%).

26
KÊT LUÂN VÀ KIẾN NGHỊ
1. Từ 26 chủng vi sinh vật được lưu giữ ở Bảo tàng giống Vi sinh vật đã chọn được 2 chủng
vi khuẩn và xạ khuẩn có khả năng sinh xylanase cao.
2. Kết hợp phân tích trình tự DNAr 16S và đặc điểm hình thái, chủng B 2H2 thuôc loài
Bacillus subtilis và chủng 118 thuộc loài Streptomyces misionensis.
3. Môi trương nuôi dich thê thich hơp cho giống khởi động của chung B 2H2 là V5 và chủng
118 là môi trường X2. Nhiệt độ nuôi cấy thích hợp cho cả hai chủng nghiên cứu là 40oC; pH
7.0; thời gian nuôi thích hợp cho giống khởi động của chủng B2H2 là 24h và chủng 118 là
42h.
4. Cơ chất thích hợp cho chủng B2H2 nuôi xốp là malt và chủng 118 là gạo. Độ ẩm nuôi cấy
thích hợp cho cả hai chủng là 50%, thời gian 3 ngày, tỷ lệ giống cấy 15%. Nguồn nitơ bổ sung
phù hợp nhất cho chủng B2H2 là cao thịt và urea cho chủng 118. Hỗn hợp khoáng thích hợp
nhất là MA1 cho chủng B2H2 và MA3 cho chủng 118. Dung dịch thích hợp cho chiết rút
enzyme là đệm 50mM phosphat pH 7.0 cho chủng B2H2 và nước cho chủng 118. Hiệu quả
thu hồi enzyme cao nhất cho cả hai chủng đạt được khi sử dụng aceton lạnh.
5. Tinh sạch và tách chiết được hai loại xylanase từ chủng 118 sử dụng phương pháp kết tủa
trong muối (NH4)2SO4 và sắc ký trao đổi ion Sepharose DEAE và bằng phương pháp điện di
trên gel SDS-PAGE, xác định được trọng lượng phân tử của hai loại xylanase tinh sạch là 22
kDa cho E1 và 36kDa cho E2.
6. pH thích hợp cho hoạt độ xylanase của chủng 118 là pH6 cho enzyme thô và E1; pH5 cho
E2. Nhiệt độ phản ứng thích hợp cho cả enzyme thô và tinh sạch là 50oC. Độ bền nhiệt là
60oC. Fe
2+ tăng cường hoạt độ xylanase cho enzyme thô và E1; Mn
2+ kích thích cho E2.
7. pH thích hợp cho enzyme xylanase của chủng B2H2 là pH6. Nhiệt độ phản ứng tối ưu là
50oC. Độ bền nhiệt là 60
0C. Ion kim loại làm tăng hoạt độ xylanase mạnh nhất là Ca
2+.
KIẾN NGHỊ
1. Tiếp tục nghiên cứu sâu hơn về đặc tính enzyme từ chủng 118
2. Tiếp tục tách chiết và tinh sạch xylanase từ chủng B2H2 và nghiên cứu đặc tính của enzyme
xylanase tinh sạch sinh ra từ chủng này.
References
TIẾNG VIỆT
1. Phạm Thị Trân Châu, Phan Tuấn Nghĩa (2007), Công nghệ sinh học Enzym và ứng dụng, Nhà
xuất bản Giáo dục (16).
2. Nguyễn Sỹ Lê Thanh, Quyền Đình Thi (2009), ―Nhân dòng và phân tích trình tự gen mã hóa
xylanase từ A. niger DSM1957”, Báo cáo Hội nghị Công nghệ Sinh học toàn quốc, Nhà xuất
bản Đại học Thái Nguyên, tr. 701 – 704 (6).

27
3. Ngô Tự Thành, Bùi Thị Việt Hà, Vũ Minh Đức, Chu Văn Mẫn (2007), ―Nghiên cứu hoạt độ
enzym ngoại bào của một số chủng Bacillus mới phân lập và khả năng ứng dụng chúng trong
xử lý nước thải‖, Tạp chí Khoa học ĐHQGHN, Khoa học Tự nhiên và Công nghệ, 25 (2009),
tr. 101-106.
4. Đặng Thị Thu, Lê Ngọc Tú, Tô Kim Anh, Phạm Thu Thủy, Nguyễn Xuân Sâm (2004), Công
nghệ Enzyme, Nhà xuất bản Khoa học và Kỹ thuật.
5. Trân Đinh Toai, Trần Thị Hồng, Lê Minh Trí, Đỗ Trung Sỹ, Hoàng Thị Bích, Nguyễn Thị Hà
Giang (2008), ―Nghiên cưu sư dung cellulase tach tư actinomycetes đê xư ly phê thai nông
nghiêp‖ , Hóa sinh và sinh học phân tử phục vụ nông, sinh, y học và công nghệ thực phẩm,
Nhà xuất bản Khoa hoc va Ky thuât , Hà Nội, tr. 901 – 903.
6. Nguyễn Quốc Trị, Nguyễn Văn Hải (2001), ―Tóm tắt những nghiên cứu về enzyme trong thức
ăn cho lợn‖, Viện chăn nuôi - Bộ nông nghiệp và Phát triển nông thôn.
7. Lê Ngọc Tú, La Văn Chứ, Đặng Thị Thu, Nguyễn Thị Thịnh, Bùi Đức Lợi, Lê Doãn Diên
(2005), Hóa sinh công nghiệp, Nhà xuất bản Khoa học và Kỹ thuật.
TIẾNG ANH
8. Amani M.D., El A. and Amany S. Y. (2007), ―Xylanase production by Bacillus pumilus:
Optimization by Statistical and Immobilization Methods‖, Journal of Agriculture and
Biological Sciences, 3(6), pp. 727-732.
9. Annane T., Anu H. and Juha R. (1994), ―Three – dimensional structure of endo–1,4–β –
xylanase II from Trichoderma reesei: two conformational states in the active site‖, The
Excellence in the Life Sciences Journal, 13 (11), pp. 2493-2501
10. Ashwani S., Neelam G., Jitender S., Kalika K., Ramesh C. K., Vijay K. G. (2007),
―Optimization of xylanase production using inexpensive agro-residues by alkalophilic
Bacillus subtilis ASH in solid-state fermentation‖, World Journal of Microbiology and
Technology, 24, pp. 633-640.
11. Chantasingh D., Pootanakit K., Champreda V., Kanokratana P., Eurwilaichitr L. (2005),
―Cloning, expression, and characterization of a xylanase 10 from Aspergillus terreus
(BCC129) in Pichia pastoris‖, Protein Expression and Purification, 46, pp. 143 – 149.
12. Chenyan Z., Yongtao W., Zhenhua L., Guanhua F., Duan L., Weiyun G., Huigen F.and Wu
W. (2009), ―Acidic xylanase II from Aspergillus usamii: Efficient expression in Pichia
pastoris and mutational analysis‖, Life Science and Technology, 9, pp. 1472-6750.
13. Cohen M.F., Hideo Y., and Mark M. (2004), ―Bioremediation of soils by plant–microbe
systems‖, International Journal of Green energy, 1, pp. 301–312.
14. Cristobal N. A., Gerardo G. S., PLilia A. B., Raul R. H., Jose L. M. H. and Juan C. C. E.
(2008),‖ Perspectives of Solid State Fermentation for Production of Food Enzymes‖,
American Journal of Biochemistry and Biotechnology, 4 (4), pp. 354-366.
15. Degefu Y. (2003), Cloning and characterization of xylanase gene from phytopathogenetic
fungi with a special reference to Helminthosporium turcicum, the cause of the northern leaf
blight of maize, University of Helsinki.

28
16. Frederic D. L. E., Virgine R., Josette L. B., Brirgit Q., and Jean M. P. (2004), ―Acidophilic
adaptation of family 11 endo-β-1,4-xylanase: Modiling and mutational analysic‖, Protein
Science, 13, pp. 1209-1218.
17. Ghose T. K. (1987), ―Measurement of cellulase activities‖, Pure and Applied Chemistry,
59(2), pp. 257-268.
18. Jane F.S. (2006), ―Mapping of residues involved in the interaction between the Bacillus
subtilis xylanase A and proteinaceous wheat xylanase inhibitors‖, Protein Engineering,
Design & Selection. 19, pp. 205-210.
19. Jiang Z. Q., Deng W., Li X. T., Ai Z. L., and Kusakebe L. T. (2005), ―Structure, Function
and Applications of Industrial enzymes‖, Industrial enzymes, 4, pp. 67-69.
20. Julio P., Andrew P. M. (2007), Industrial Enzymes: Structure, Function and Applications ,
University of Helsinki, pp. 65-97.
21. Ken K. Y. W., Larry U. L. T, and John N.S. (1988), ―Multiplicity of β-1,4-xylanase in
microorganisms: Functions and application‖, Microbiological reviews, pp. 305-317.
22. Lee Y. E., Pyung O. L. (2004), ―Purification and Characterization of Two Thermostable
Xylanases from Paenibacillus sp. DG , 22‖, Journal of Microbiology and Biotechnology, 14
(5), pp. 1014 - 1021.
23. Maheswari M. U., Chandra T. S. (2000), ―Production and potential applications of a xylanase
from a new strain of Streptomyces cuspidosporus‖, World Journal of Microbiology and
Biotechnology, 16, 257 - 263.
24. Masood S. M., Tahir N., Zulfiqar A. and Muhammad T. S. (2008), ―Xylanases and their
application in baking industry‖, Food technology and Biotechnology, 46, p. 22-31.
25. Nicemol J. and Parukuttyamma P. (2006), ―Influence of mode of fermentation on production
of polygalacturonase by a novel strain of Streptomyces lydicus‖, Food Technology and
Biotechnology, 44, pp. 263–267.
26. Nyilas I., Acs K., Papp T., Nagy E., Vagvolgyi C. (2005), ―Agrobacterium tumefaciens –
mediated transformation of Mucor circinellnoides‖, Fonia Microbiology, 50 (5), pp. 415 –
420.
27. Sapre M.P., Jha H. and Patil M.B. (2005), ―Purification and characterization of a
thermoalkalophilic xylanase from Bacillus sp.”, World Journal of Microbiology &
Biotechnology, 21, pp. 649 - 654.
28. Susana R. C., Sanroman M. A. (2006), ―Application of solid-state fermentation to food
industry—A review‖, Journal of Food Engineering, 76, pp. 291 - 302.
29. Taechowisan T., Peberdy J.F., Lumyong S. (2003), ―Chitinase production by endophytic
Streptomyces aureofaciens CMUAc130 and its antagonism against phytopathogenic fungi‖,
Annals of Microbiology, 53 (4), pp. 447-461.

29
30. Ufuk B., Sebnem Y., Ferda G., Aysegul E. (2001), ―An endo-1,4-xylanase from Rhizopus
oryzae: production, partial purification and biochemical characterization‖, Journal of Applied
Sciences Research, 6 (9), pp. 1373-1378.
31. Yin et al., Agric J. (2010), Food Chemistry, 58, pp. 557–562 .
32. Waksman S. A. (1961), The Actinomycetes : Classification, identification and descriptions
of genera and species, The Williams & Wilkins Co., Baltimore.
33. www.bioclub.forumotion.com/t22,topic
34. www.mt.lhu.edu.vn
35. www.ncfar.org
36. www.sciencedirect.com/science
37. www.sigmaaldrich.com
38. www.tailieu.vn
39. www.vi.wikipedia.org
40. www.wattpad.com





























