Embed Size (px)
Citation preview

at SciVerse ScienceDirect
Neuropharmacology 63 (2012) 1191e1199
Contents lists available
Neuropharmacology
journal homepage: www.elsevier .com/locate/neuropharm
Magnolol, a major bioactive constituent of the bark of Magnolia officinalis, inducessleep via the benzodiazepine site of GABAA receptor in mice
Chang-Rui Chen a,1, Xu-Zhao Zhou a,1, Yan-Jia Luo a, Zhi-Li Huang a,b,c,*, Yoshihiro Urade d, Wei-Min Qu a,c,*
aDepartment of Pharmacology, Fudan University, Shanghai, 200032, PR Chinab State Key Laboratory of Medical Neurobiology, Shanghai Medical College, Fudan University, Shanghai, 200032, PR Chinac Institute of Brain Science, Fudan University, Shanghai, 200032, PR ChinadDepartment of Molecular Behavioral Biology, Osaka Bioscience Institute, 6-2-4 Furuedai, Suita, Osaka 565-0874, Japan
a r t i c l e i n f o
Article history:Received 29 March 2012Received in revised form13 June 2012Accepted 14 June 2012
Keywords:EEGGABAA
MagnololSleepVLPOTMN
Abbreviations: EEG, electroencephalogram; BZ, benil; Mag, magnolol; VLPO, ventrolateral preoptic arnucleus.* Corresponding authors. Department of Pharmacol
of Medical Neurobiology, Shanghai Medical College200032, PR China. Tel.: þ86 21 54237043.
E-mail addresses: [email protected] (fudan.edu.cn (W.-M. Qu).
1 C.R.C. and X.Z.Z equally contributed to this work.
0028-3908/$ e see front matter � 2012 Elsevier Ltd.http://dx.doi.org/10.1016/j.neuropharm.2012.06.031
a b s t r a c t
Magnolol (6,60,7,12-tetramethoxy-2,20-dimethyl-1-beta-berbaman, C18H18O2), an active ingredient of thebark of Magnolia officinalis, has been reported to exert potent anti-epileptic effects via the GABAA
receptor. The receptor also mediates sleep in humans and animals. The aim of this study was to deter-mine whether magnolol could modulate sleep behaviors by recording EEG and electromyogram in mice.The results showed that magnolol administered i.p. at a dose of 5 or 25 mg/kg could significantly shortenthe sleep latency, increase the amount of non-rapid eye movement (non-REM, NREM) and rapid eyemovement (REM) sleep for 3 h after administration with an increase in the number of NREM and REMsleep episodes. Magnolol at doses of 5 and 25 mg/kg increased the number of bouts of wakefulness butdecreased their duration. On the other hand, magnolol increased the number of state transitions fromwakefulness to NREM sleep and subsequently from NREM sleep to wakefulness. Immunohistochemicalstudy showed that magnolol increased c-Fos expression in the neurons of ventrolateral preoptic area,a sleep center in the anterior hypothalamus, and decreased c-Fos expression in the arousal tuber-omammillary nucleus, which was located in the caudolateral hypothalamus. The sleep-promoting effectsand changes in c-Fos induced by magnolol were reversed by flumazenil, an antagonist at the benzodi-azepine site of the GABAA receptor. These results indicate that magnolol increased NREM and REM sleepvia the GABAA receptor.
� 2012 Elsevier Ltd. All rights reserved.
1. Introduction
Insomnia is a highly prevalent condition characterized byinability to sleep or a total lack of sleep. Insomnia afflicts 35% of thegeneral population worldwide. About 10e15% of insomnia patientsare diagnosed with moderate to severe disorders. This gives rise toemotional distress, daytime fatigue, and loss of productivity(Buckner et al., 2008). Among pharmacotherapeutic agents, non-benzodiazepine hypnotics (Z-drugs) are the first line of
nzodiazepines; Flu, flumaze-ea; TMN, tuberomammillary
ogy and State Key Laboratory, Fudan University, Shanghai
Z.-L. Huang), quweimin@
All rights reserved.
management for insomnia, followed by benzodiazepines (BZ),amitryptiline, and antihistamines. The traditional BZ and Z-drugsmodulate the GABAA receptors (Rudolph and Knoflach, 2011).Diazepam and zolpidem significantly increase total non-rapid eyemovement (non-REM, NREM) sleep, but they can also causea remarkable decrease in delta power activity during NREM sleep(van Lier et al., 2004). The discrepancy between the increase insleep continuity and the reduction of power in the lower EEGfrequencies caused by BZ in humans is consistent with findings inmice (Tobler et al., 2001). The adverse effects of BZ includedecreased psychomotor performance (e.g. next-day drowsiness),tolerance, dependency, and withdrawal symptoms. These havelimited their use, indicating that there is still a need for hypnoticsthat do not produce dependence or detrimental daytime conse-quences such as sedation and drowsiness. Over the past decades,the identification of separable key functions of GABAA receptorsubtypes has suggested that receptor subtype-selective compoundscould overcome the limitations of classical BZ (Rudolph andKnoflach, 2011).

Fig. 1. Chemical structure of magnolol.
C.-R. Chen et al. / Neuropharmacology 63 (2012) 1191e11991192
Magnolol (6,60,7,12-tetramethoxy-2,20-dimethyl-1-beta-berba-man, C18H18O2, see Fig. 1 for chemical structure) is the majorbioactive constituent of magnolia bark. Magnolol increases thethreshold of NMDA-induced seizures (Lin et al., 2005). It also delaysthe onset of myoclonic jerks and generalized clonic seizures anddecreases seizure stage and mortality (Chen et al., 2011). Magnololhas also been found to prolong the duration of sleeping timeinduced by pentobarbital in mice (Ma et al., 2009). These dataindicate that magnolol may have sleep-promoting effects.
The GABAA receptor is the main inhibitory neurotransmitterreceptor in the CNS. The fast-inhibitory activity of GABA is medi-ated by the GABAA receptor. It has been reported that magnololincreases the number of binding sites and affinity of the GABAAreceptor for GABA and then enhances chloride influx (Ai et al.,2001; Ma et al., 2009; Squires et al., 1999; Alexeev et al., 2012).These findings suggest that magnolol might enhance the inhibitoryaction of GABA on GABAA receptors by binding to the sites in GABAAreceptor. We hypothesized that the GABAA receptor might beresponsible for the sleep-promoting effects of magnolol.
In the present study, we showed that magnolol shortened sleeplatency, increased the number of bouts of NREM, and the amount ofNREM and REM sleep. Immunostaining showed that magnololincreased c-Fos expression in neurons of the ventrolateral preopticnucleus (VLPO), which is one of the sleep centers, and decreased itin the arousal histaminergic tuberomammillary nucleus (TMN).These findings suggest that magnolol may have applications in thetreatment of insomnia.
2. Methods and materials
2.1. Animals
Male SPF inbred C57BL/6J mice (weighing 20e28 g, 11e13 weeks old) wereobtained from the Laboratory Animal Center, Chinese Academy of Sciences(Shanghai, China). The animals were housed individually at a constant temperature(24� 0.5 �C) with a relative humidity of 60� 2% on an automatically controlled 12 hlight/dark cycle (lights on at 7:00 a.m.), and they had free access to food and water.All efforts were made to minimize animal suffering and to use only the number ofanimals required for the production of reliable scientific data. All experiments werecarried out in accordance with the National Institutes of Health Guide for the Careand Use of Laboratory Animals.
2.2. Chemicals
Magnolol was purchased from the National Center for Safety Evaluation of Drugs(Beijing, China). It was shown to be 98% pure by high-performance liquid chroma-tography. Rabbit polyclonal anti-c-Fos antibody was purchased from Abcam (Cam-bridge, MA). Biotinylated donkey anti-rabbit IgG and avidinebiotin-peroxidasecame from Vector Laboratories (CA); flumazenil and 3,30-diamino-benzidine-tetra-hydrochloride (DAB) from SigmaeAldrich (SigmaeAldrich, St. Louis, MO, U.S.).Magnolol and flumazenil were dissolved in saline with 5% dimethyl-sulfoxide(DMSO).
2.3. Polygraphic recordings and vigilance state analysis
Under pentobarbital anesthesia (50 mg/kg, i.p.), mice were chronicallyimplanted with electrodes for polysomnographic recordings of EEG and electro-myogram (EMG). Two stainless steel screws (1 mm in diameter) were insertedthrough the skull into the cortex (antero-posterior, þ1.0 mm; lefteright, �1.5 mmfrom bregma or lambda) according to the atlas of Franklin and Paxinos (1997). These
served as EEG electrodes. Two insulated stainless steel, teflon-coated wires werebilaterally placed into both trapezius muscles. These served as EMG electrodes. Allelectrodes were attached to a microconnector and fixed onto the skull with dentalcement. The EEG and EMG recordings were carried out by means of a slip ringdesigned so that the behavioral movement of the mice would not be restricted. Aftera 10-day recovery period, the mice were housed individually in transparent barrelsand habituated to the recording cable for 3e4 days before polygraphic recording. Allmice that were subjected to EEG recordings received vehicle and drug treatment on2 consecutive days. On day 1, the mice were treated with vehicle (intraperitoneally)at 21:00 and the recordings made on that day served as the baseline data. On day 2,mice were treated with magnolol (intraperitoneally, in a volume of 10 ml/kg bodyweight) at 21:00, and EEG/EMG signals were recorded for 24 h.
2.4. Analysis of vigilance state
The EEG/EMG signals were amplified and filtered (EEG, 0.5e30 Hz; EMG,20e200 Hz), then digitized at a sampling rate of 128 Hz, and recorded usingSLEEPSIGN software as described before (Oishi et al., 2008; Qu et al., 2008). Thevigilance states were automatically classified off-line in 4 s epochs into REM sleep,NREM sleep, and wakefulness by SLEEPSIGN, according to the standard criteria (Quet al., 2010). As a final step, defined sleepewake stages were examined visually andcorrected if necessary.
2.5. Pharmacological treatments
Magnolol was prepared as described above immediately before use andadministered i.p. at 21:00 on the day of the experiment at a dose of 1, 5 or 25 mg/kg(n ¼ 5e7). All drugs were freshly prepared prior to use, and an injection volume(10 ml/kg) was kept constant for in vivo experiments. For baseline data, mice wereinjected i.p. with vehicle (10 ml/kg). To test receptor mechanisms, 30 min before theinjection of magnolol (25 mg/kg), mice were pretreated with flumazenil (Flu) i.p. at0.5 or 1 mg/kg. Diazepam at 6 mg/kg was injected as a positive control.
2.6. c-Fos immunohistochemistry
Nine groups of mice were used. One group was treated with vehicle; and theothers were injected i.p. with magnolol at doses of 1, 5, and 25 mg/kg and diazepamat 6 mg/kg. To test the receptor mechanisms, four groups of mice were used: Flu-mazenil (Flu) 1 mg/kg þ vehicle, Flu 1 mg/kg þ magnolol 25 mg/kg, Flu 0.5 mg/kg þ magnolol 25 mg/kg, and Flu 1 mg/kg þ diazepam 6 mg/kg groups. Mice werepretreated with Flu, and then the magnolol 25 mg/kg was given after 30 min. At60 min after the administration of magnolol, the animals were anesthetized with10% chloral hydrate and perfused via the heart with saline solution followed by ice-cold 4% paraformaldehyde (PFA) in 0.1 M phosphate buffer (pH 7.4). Their brainswere then removed, post-fixed in 4% PFA for 6 h, and immersed in 30% sucroseovernight. Thereafter, frozen sections were cut at 30 mm in coronal planes by use ofa freezing microtome (Leica Microsystems, Wetzlar, Germany). The sections werestored in a cryoprotectant solution at �20 �C for histological analysis. Immunohis-tochemistry was performed in accordance with the free-floating method describedearlier (Chen et al., 2011). Sections were fixed in 4% PFA for 10 min and incubatedwith 0.3% H2O2 for 15 min to quench the endogenous peroxidase activity. Thesections were next placed in blocking solution containing 10% normal goat serumwith 0.3% Triton X-100 in 0.01 M phosphate-buffered saline (PBS, pH 7.2) for 30 minat 37 �C and then incubated at 4 �C for 24 h with a rabbit polyclonal antibody againstc-Fos at a 1:5000 dilution in PBS containing 0.3% Triton X-100. On the second day,the sections were incubated with a 1:1000 dilution of biotinylated donkey anti-rabbit secondary antibodies for 30 min followed by a 1:200 dilution ofavidinebiotin-peroxidase for 1 h at 37 �C. The peroxidase reaction was visualizedwith 0.05% DAB in 0.1 M phosphate buffer and 0.01% H2O2. Sections were mounted,dehydrated, and cover slipped. As controls, adjacent sections were incubatedwithout primary antibody to confirm that no non-specific staining had occurred.
Digital images were viewed and captured using the Olympus DP 72 microscope(Olympus, Tokyo, Japan). Figures were assembled and adjusted for brightness andcontrast in Adobe Photoshop (Adobe Systems Inc, San Jose, CA, U.S.).
2.7. Statistical analysis
All data were expressed as the mean � SEM (n ¼ 5e7). Statistical analysis wasperformed with SPSS 17.0 (SPSS Inc., Chicago, IL, U.S.). Time-course of the hourlyamounts of each stage, histograms of the amounts of sleep and wakefulness, thenumber of transitions between sleep andwakefulness, and the number and durationof bouts of sleep and wakefulness were analyzed using the paired t-test, with eachanimal serving as its own control. For sleep latency, the total number of each vigi-lance stage during the 3 h immediately following drug treatment, one-way repeatedmeasures analysis of variance (ANOVA) was performed followed by the Fisherprobable least-squares difference (PLSD) test to determine whether the differencesamong groups were statistically significant. For the number of c-Fos immunoreac-tive neurons, one-way or two-way ANOVA was used, followed by PLSD test. Thesignificance level was set at P < 0.05 for all statistical tests.
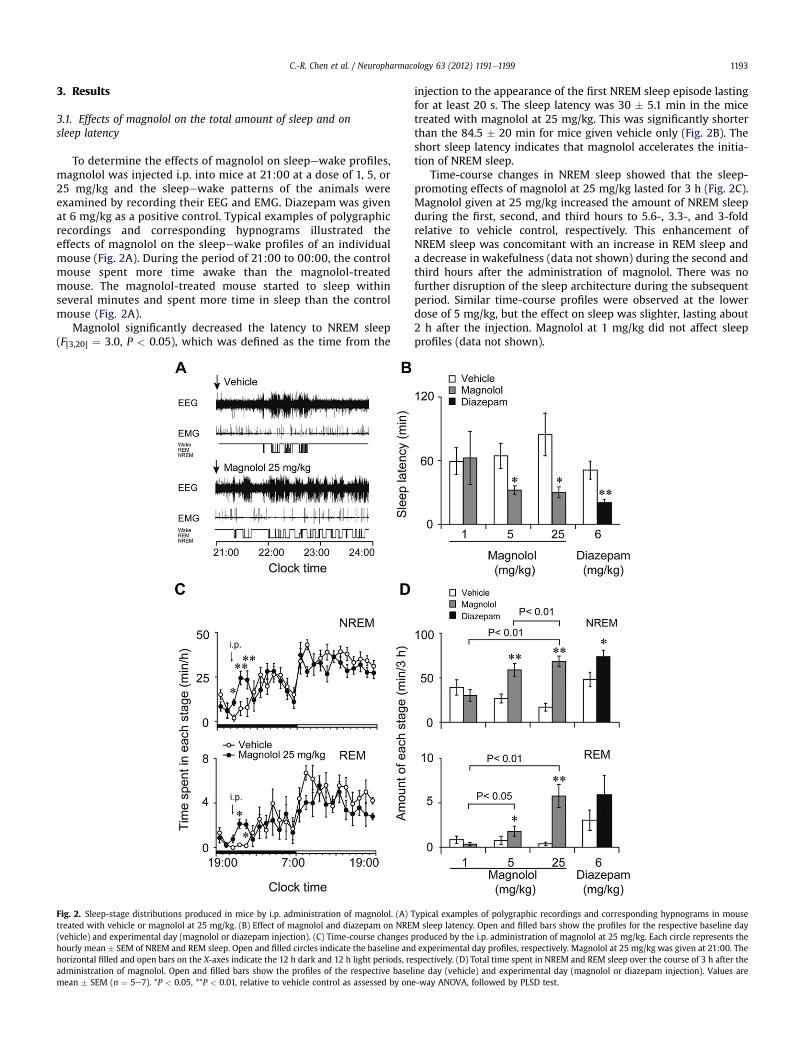
C.-R. Chen et al. / Neuropharmacology 63 (2012) 1191e1199 1193
3. Results
3.1. Effects of magnolol on the total amount of sleep and onsleep latency
To determine the effects of magnolol on sleepewake profiles,magnolol was injected i.p. into mice at 21:00 at a dose of 1, 5, or25 mg/kg and the sleepewake patterns of the animals wereexamined by recording their EEG and EMG. Diazepam was givenat 6 mg/kg as a positive control. Typical examples of polygraphicrecordings and corresponding hypnograms illustrated theeffects of magnolol on the sleepewake profiles of an individualmouse (Fig. 2A). During the period of 21:00 to 00:00, the controlmouse spent more time awake than the magnolol-treatedmouse. The magnolol-treated mouse started to sleep withinseveral minutes and spent more time in sleep than the controlmouse (Fig. 2A).
Magnolol significantly decreased the latency to NREM sleep(F[3,20] ¼ 3.0, P < 0.05), which was defined as the time from the
Fig. 2. Sleep-stage distributions produced in mice by i.p. administration of magnolol. (A) Ttreated with vehicle or magnolol at 25 mg/kg. (B) Effect of magnolol and diazepam on NRE(vehicle) and experimental day (magnolol or diazepam injection). (C) Time-course changes phourly mean � SEM of NREM and REM sleep. Open and filled circles indicate the baseline anhorizontal filled and open bars on the X-axes indicate the 12 h dark and 12 h light periods, readministration of magnolol. Open and filled bars show the profiles of the respective baselmean � SEM (n ¼ 5e7). *P < 0.05, **P < 0.01, relative to vehicle control as assessed by on
injection to the appearance of the first NREM sleep episode lastingfor at least 20 s. The sleep latency was 30 � 5.1 min in the micetreated with magnolol at 25 mg/kg. This was significantly shorterthan the 84.5 � 20 min for mice given vehicle only (Fig. 2B). Theshort sleep latency indicates that magnolol accelerates the initia-tion of NREM sleep.
Time-course changes in NREM sleep showed that the sleep-promoting effects of magnolol at 25 mg/kg lasted for 3 h (Fig. 2C).Magnolol given at 25 mg/kg increased the amount of NREM sleepduring the first, second, and third hours to 5.6-, 3.3-, and 3-foldrelative to vehicle control, respectively. This enhancement ofNREM sleep was concomitant with an increase in REM sleep anda decrease in wakefulness (data not shown) during the second andthird hours after the administration of magnolol. There was nofurther disruption of the sleep architecture during the subsequentperiod. Similar time-course profiles were observed at the lowerdose of 5 mg/kg, but the effect on sleep was slighter, lasting about2 h after the injection. Magnolol at 1 mg/kg did not affect sleepprofiles (data not shown).
ypical examples of polygraphic recordings and corresponding hypnograms in mouseM sleep latency. Open and filled bars show the profiles for the respective baseline dayroduced by the i.p. administration of magnolol at 25 mg/kg. Each circle represents the
d experimental day profiles, respectively. Magnolol at 25 mg/kg was given at 21:00. Thespectively. (D) Total time spent in NREM and REM sleep over the course of 3 h after theine day (vehicle) and experimental day (magnolol or diazepam injection). Values aree-way ANOVA, followed by PLSD test.
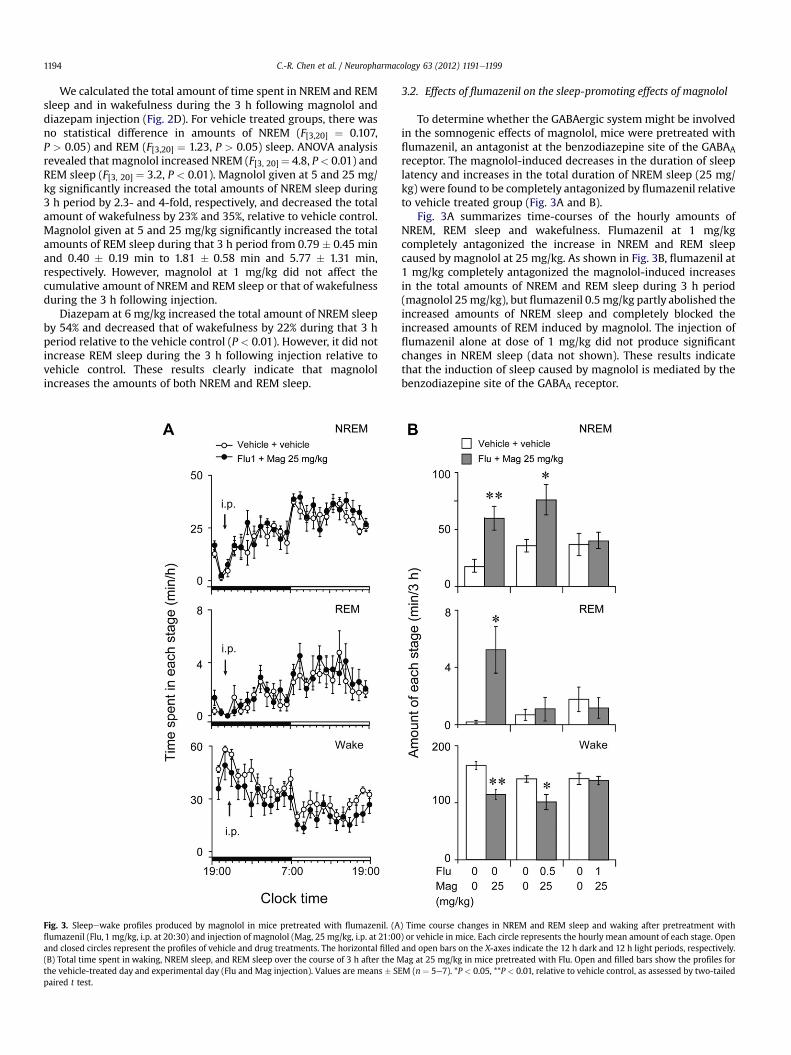
C.-R. Chen et al. / Neuropharmacology 63 (2012) 1191e11991194
We calculated the total amount of time spent in NREM and REMsleep and in wakefulness during the 3 h following magnolol anddiazepam injection (Fig. 2D). For vehicle treated groups, there wasno statistical difference in amounts of NREM (F[3,20] ¼ 0.107,P > 0.05) and REM (F[3,20] ¼ 1.23, P > 0.05) sleep. ANOVA analysisrevealed that magnolol increased NREM (F[3, 20]¼ 4.8, P< 0.01) andREM sleep (F[3, 20] ¼ 3.2, P < 0.01). Magnolol given at 5 and 25 mg/kg significantly increased the total amounts of NREM sleep during3 h period by 2.3- and 4-fold, respectively, and decreased the totalamount of wakefulness by 23% and 35%, relative to vehicle control.Magnolol given at 5 and 25 mg/kg significantly increased the totalamounts of REM sleep during that 3 h period from 0.79 � 0.45 minand 0.40 � 0.19 min to 1.81 � 0.58 min and 5.77 � 1.31 min,respectively. However, magnolol at 1 mg/kg did not affect thecumulative amount of NREM and REM sleep or that of wakefulnessduring the 3 h following injection.
Diazepam at 6 mg/kg increased the total amount of NREM sleepby 54% and decreased that of wakefulness by 22% during that 3 hperiod relative to the vehicle control (P < 0.01). However, it did notincrease REM sleep during the 3 h following injection relative tovehicle control. These results clearly indicate that magnololincreases the amounts of both NREM and REM sleep.
Fig. 3. Sleepewake profiles produced by magnolol in mice pretreated with flumazenil. (Aflumazenil (Flu, 1 mg/kg, i.p. at 20:30) and injection of magnolol (Mag, 25 mg/kg, i.p. at 21:00and closed circles represent the profiles of vehicle and drug treatments. The horizontal filled(B) Total time spent in waking, NREM sleep, and REM sleep over the course of 3 h after the Mthe vehicle-treated day and experimental day (Flu and Mag injection). Values are means � SEpaired t test.
3.2. Effects of flumazenil on the sleep-promoting effects of magnolol
To determine whether the GABAergic system might be involvedin the somnogenic effects of magnolol, mice were pretreated withflumazenil, an antagonist at the benzodiazepine site of the GABAAreceptor. The magnolol-induced decreases in the duration of sleeplatency and increases in the total duration of NREM sleep (25 mg/kg) were found to be completely antagonized by flumazenil relativeto vehicle treated group (Fig. 3A and B).
Fig. 3A summarizes time-courses of the hourly amounts ofNREM, REM sleep and wakefulness. Flumazenil at 1 mg/kgcompletely antagonized the increase in NREM and REM sleepcaused by magnolol at 25 mg/kg. As shown in Fig. 3B, flumazenil at1 mg/kg completely antagonized the magnolol-induced increasesin the total amounts of NREM and REM sleep during 3 h period(magnolol 25mg/kg), but flumazenil 0.5 mg/kg partly abolished theincreased amounts of NREM sleep and completely blocked theincreased amounts of REM induced by magnolol. The injection offlumazenil alone at dose of 1 mg/kg did not produce significantchanges in NREM sleep (data not shown). These results indicatethat the induction of sleep caused by magnolol is mediated by thebenzodiazepine site of the GABAA receptor.
) Time course changes in NREM and REM sleep and waking after pretreatment with) or vehicle in mice. Each circle represents the hourly mean amount of each stage. Openand open bars on the X-axes indicate the 12 h dark and 12 h light periods, respectively.ag at 25 mg/kg in mice pretreated with Flu. Open and filled bars show the profiles forM (n ¼ 5e7). *P < 0.05, **P < 0.01, relative to vehicle control, as assessed by two-tailed
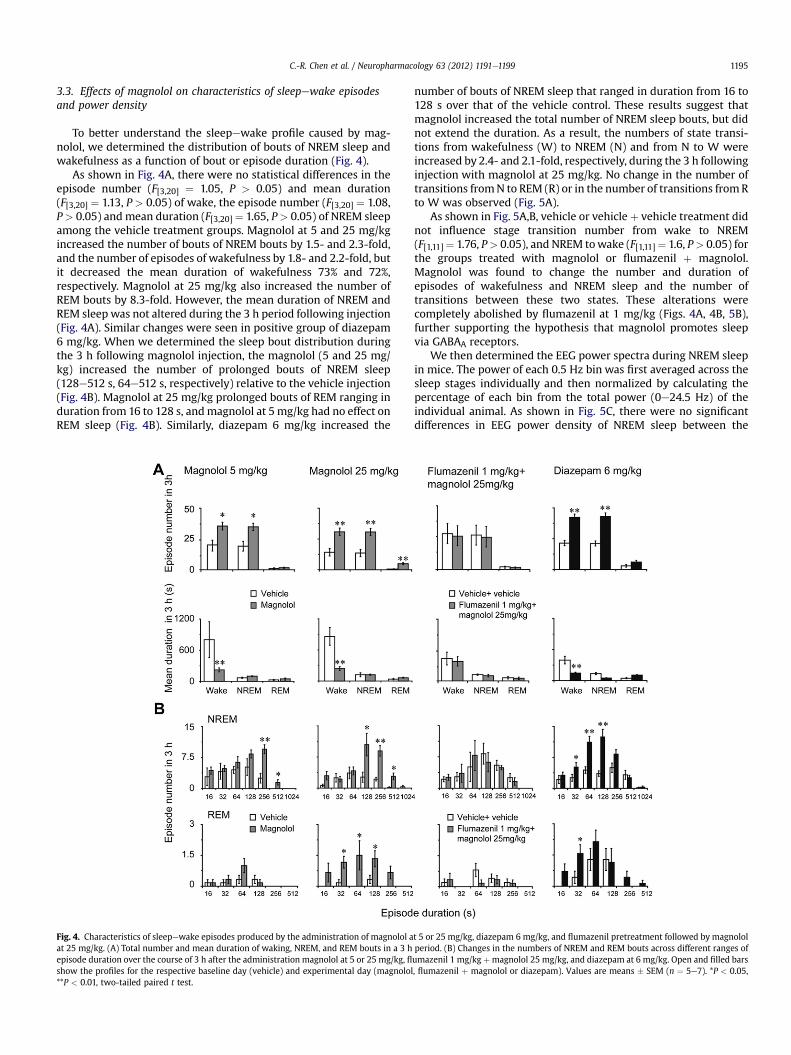
C.-R. Chen et al. / Neuropharmacology 63 (2012) 1191e1199 1195
3.3. Effects of magnolol on characteristics of sleepewake episodesand power density
To better understand the sleepewake profile caused by mag-nolol, we determined the distribution of bouts of NREM sleep andwakefulness as a function of bout or episode duration (Fig. 4).
As shown in Fig. 4A, there were no statistical differences in theepisode number (F[3,20] ¼ 1.05, P > 0.05) and mean duration(F[3,20] ¼ 1.13, P > 0.05) of wake, the episode number (F[3,20] ¼ 1.08,P> 0.05) andmean duration (F[3,20] ¼ 1.65, P> 0.05) of NREM sleepamong the vehicle treatment groups. Magnolol at 5 and 25 mg/kgincreased the number of bouts of NREM bouts by 1.5- and 2.3-fold,and the number of episodes of wakefulness by 1.8- and 2.2-fold, butit decreased the mean duration of wakefulness 73% and 72%,respectively. Magnolol at 25 mg/kg also increased the number ofREM bouts by 8.3-fold. However, the mean duration of NREM andREM sleep was not altered during the 3 h period following injection(Fig. 4A). Similar changes were seen in positive group of diazepam6 mg/kg. When we determined the sleep bout distribution duringthe 3 h following magnolol injection, the magnolol (5 and 25 mg/kg) increased the number of prolonged bouts of NREM sleep(128e512 s, 64e512 s, respectively) relative to the vehicle injection(Fig. 4B). Magnolol at 25 mg/kg prolonged bouts of REM ranging induration from 16 to 128 s, andmagnolol at 5mg/kg had no effect onREM sleep (Fig. 4B). Similarly, diazepam 6 mg/kg increased the
Fig. 4. Characteristics of sleepewake episodes produced by the administration of magnolol aat 25 mg/kg. (A) Total number and mean duration of waking, NREM, and REM bouts in a 3 hepisode duration over the course of 3 h after the administration magnolol at 5 or 25 mg/kg, flshow the profiles for the respective baseline day (vehicle) and experimental day (magnolol**P < 0.01, two-tailed paired t test.
number of bouts of NREM sleep that ranged in duration from 16 to128 s over that of the vehicle control. These results suggest thatmagnolol increased the total number of NREM sleep bouts, but didnot extend the duration. As a result, the numbers of state transi-tions from wakefulness (W) to NREM (N) and from N to W wereincreased by 2.4- and 2.1-fold, respectively, during the 3 h followinginjection with magnolol at 25 mg/kg. No change in the number oftransitions fromN to REM (R) or in the number of transitions fromRto W was observed (Fig. 5A).
As shown in Fig. 5A,B, vehicle or vehicle þ vehicle treatment didnot influence stage transition number from wake to NREM(F[1,11]¼ 1.76, P> 0.05), and NREM towake (F[1,11]¼ 1.6, P> 0.05) forthe groups treated with magnolol or flumazenil þ magnolol.Magnolol was found to change the number and duration ofepisodes of wakefulness and NREM sleep and the number oftransitions between these two states. These alterations werecompletely abolished by flumazenil at 1 mg/kg (Figs. 4A, 4B, 5B),further supporting the hypothesis that magnolol promotes sleepvia GABAA receptors.
We then determined the EEG power spectra during NREM sleepin mice. The power of each 0.5 Hz bin was first averaged across thesleep stages individually and then normalized by calculating thepercentage of each bin from the total power (0e24.5 Hz) of theindividual animal. As shown in Fig. 5C, there were no significantdifferences in EEG power density of NREM sleep between the
t 5 or 25 mg/kg, diazepam 6 mg/kg, and flumazenil pretreatment followed by magnololperiod. (B) Changes in the numbers of NREM and REM bouts across different ranges ofumazenil 1 mg/kg þmagnolol 25 mg/kg, and diazepam at 6 mg/kg. Open and filled bars, flumazenil þ magnolol or diazepam). Values are means � SEM (n ¼ 5e7). *P < 0.05,
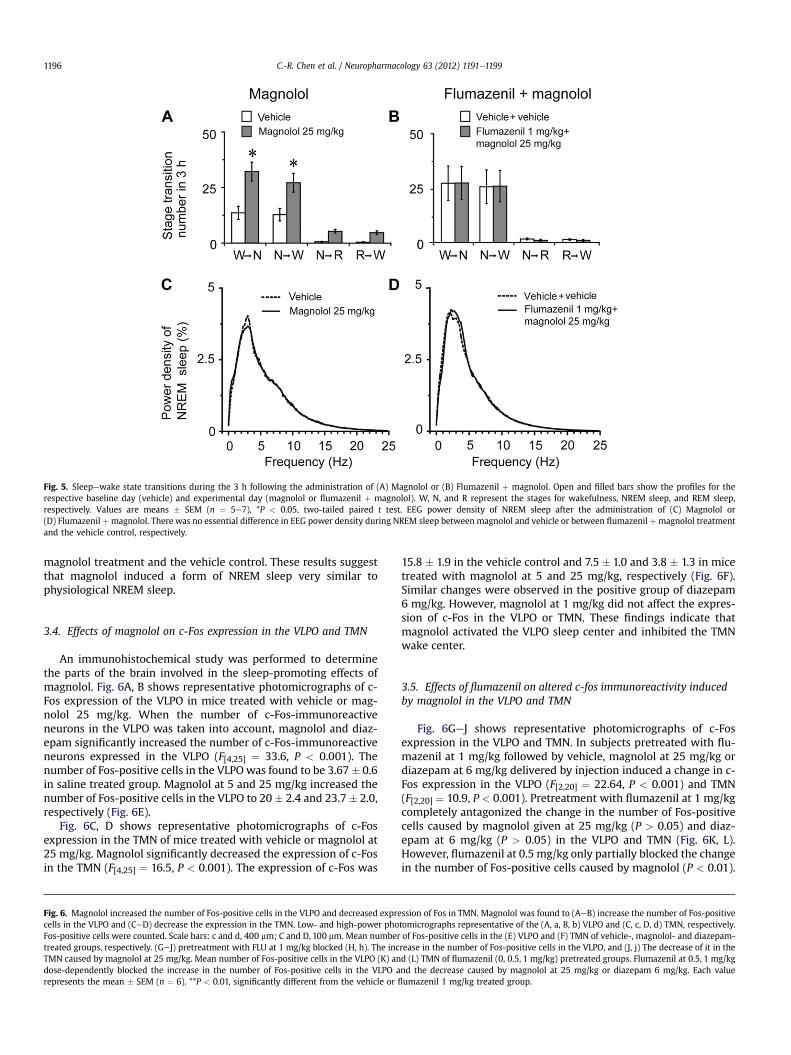
Fig. 5. Sleepewake state transitions during the 3 h following the administration of (A) Magnolol or (B) Flumazenil þ magnolol. Open and filled bars show the profiles for therespective baseline day (vehicle) and experimental day (magnolol or flumazenil þ magnolol). W, N, and R represent the stages for wakefulness, NREM sleep, and REM sleep,respectively. Values are means � SEM (n ¼ 5e7), *P < 0.05, two-tailed paired t test. EEG power density of NREM sleep after the administration of (C) Magnolol or(D) Flumazenil þmagnolol. There was no essential difference in EEG power density during NREM sleep between magnolol and vehicle or between flumazenil þmagnolol treatmentand the vehicle control, respectively.
C.-R. Chen et al. / Neuropharmacology 63 (2012) 1191e11991196
magnolol treatment and the vehicle control. These results suggestthat magnolol induced a form of NREM sleep very similar tophysiological NREM sleep.
3.4. Effects of magnolol on c-Fos expression in the VLPO and TMN
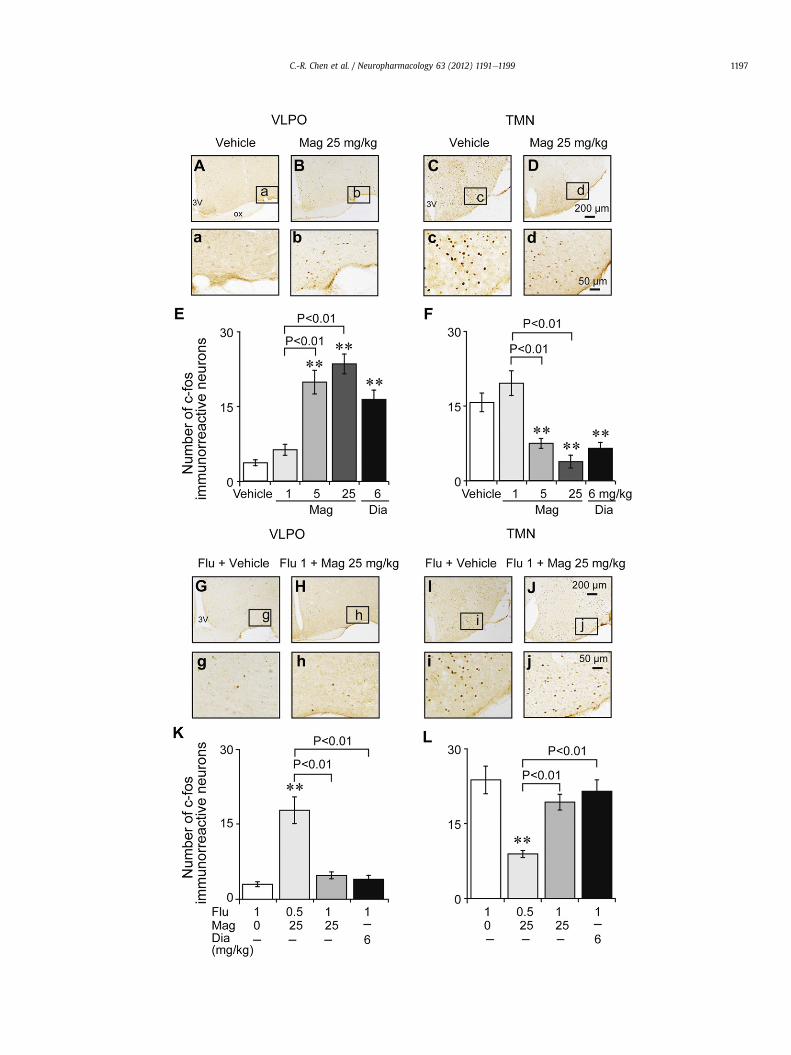
An immunohistochemical study was performed to determinethe parts of the brain involved in the sleep-promoting effects ofmagnolol. Fig. 6A, B shows representative photomicrographs of c-Fos expression of the VLPO in mice treated with vehicle or mag-nolol 25 mg/kg. When the number of c-Fos-immunoreactiveneurons in the VLPO was taken into account, magnolol and diaz-epam significantly increased the number of c-Fos-immunoreactiveneurons expressed in the VLPO (F[4,25] ¼ 33.6, P < 0.001). Thenumber of Fos-positive cells in the VLPO was found to be 3.67 � 0.6in saline treated group. Magnolol at 5 and 25 mg/kg increased thenumber of Fos-positive cells in the VLPO to 20 � 2.4 and 23.7 � 2.0,respectively (Fig. 6E).
Fig. 6C, D shows representative photomicrographs of c-Fosexpression in the TMN of mice treated with vehicle or magnolol at25 mg/kg. Magnolol significantly decreased the expression of c-Fosin the TMN (F[4,25] ¼ 16.5, P < 0.001). The expression of c-Fos was
Fig. 6. Magnolol increased the number of Fos-positive cells in the VLPO and decreased exprecells in the VLPO and (CeD) decrease the expression in the TMN. Low- and high-power phoFos-positive cells were counted. Scale bars: c and d, 400 mm; C and D, 100 mm. Mean numbertreated groups, respectively. (GeJ) pretreatment with FLU at 1 mg/kg blocked (H, h). The incTMN caused by magnolol at 25 mg/kg. Mean number of Fos-positive cells in the VLPO (K) andose-dependently blocked the increase in the number of Fos-positive cells in the VLPO arepresents the mean � SEM (n ¼ 6). **P < 0.01, significantly different from the vehicle or fl
15.8 � 1.9 in the vehicle control and 7.5 � 1.0 and 3.8 � 1.3 in micetreated with magnolol at 5 and 25 mg/kg, respectively (Fig. 6F).Similar changes were observed in the positive group of diazepam6 mg/kg. However, magnolol at 1 mg/kg did not affect the expres-sion of c-Fos in the VLPO or TMN. These findings indicate thatmagnolol activated the VLPO sleep center and inhibited the TMNwake center.
3.5. Effects of flumazenil on altered c-fos immunoreactivity inducedby magnolol in the VLPO and TMN
Fig. 6GeJ shows representative photomicrographs of c-Fosexpression in the VLPO and TMN. In subjects pretreated with flu-mazenil at 1 mg/kg followed by vehicle, magnolol at 25 mg/kg ordiazepam at 6 mg/kg delivered by injection induced a change in c-Fos expression in the VLPO (F[2,20] ¼ 22.64, P < 0.001) and TMN(F[2,20] ¼ 10.9, P < 0.001). Pretreatment with flumazenil at 1 mg/kgcompletely antagonized the change in the number of Fos-positivecells caused by magnolol given at 25 mg/kg (P > 0.05) and diaz-epam at 6 mg/kg (P > 0.05) in the VLPO and TMN (Fig. 6K, L).However, flumazenil at 0.5 mg/kg only partially blocked the changein the number of Fos-positive cells caused by magnolol (P < 0.01).
ssion of Fos in TMN. Magnolol was found to (AeB) increase the number of Fos-positivetomicrographs representative of the (A, a, B, b) VLPO and (C, c, D, d) TMN, respectively.of Fos-positive cells in the (E) VLPO and (F) TMN of vehicle-, magnolol- and diazepam-rease in the number of Fos-positive cells in the VLPO, and (J, j) The decrease of it in thed (L) TMN of flumazenil (0, 0.5, 1 mg/kg) pretreated groups. Flumazenil at 0.5, 1 mg/kgnd the decrease caused by magnolol at 25 mg/kg or diazepam 6 mg/kg. Each valueumazenil 1 mg/kg treated group.
C.-R. Chen et al. / Neuropharmacology 63 (2012) 1191e1199 1197

C.-R. Chen et al. / Neuropharmacology 63 (2012) 1191e11991198
These findings indicate that the GABAA receptor mediates theability of magnolol to activate the VLPO sleep center and inhibit theactivity of the TMN arousal system.
4. Discussion
The present study clearly showed that magnolol can shortensleep latency and increase the amounts of NREM and REM sleep.Magnolol also increased the number of NREM sleep episodes andthe number of transitions between wakefulness and NREM sleepstates. Flumazenil, a competitive inhibitor of the activity at the BZrecognition site on the GABAA/BZ ligandereceptor complex,reversed the sleep-promoting effects of magnolol. These resultssuggest that magnolol might share a similar mechanism of actionwith BZ.
GABA is the most abundant inhibitory neurotransmitter in themammalian central nervous system. It plays a key role in neuronalfiring patterns and in the activity of neuronal networks. Themodulators of GABAA receptors have been the focus of severaltherapies aimed at insomnia (Zammit, 2007). Several in vivo andin vitro studies support our findings that magnolol might elicitsleep-promoting effects by acting on the GABAA receptor. Magnololwas found to decrease the release of many neurotransmitters, suchas acetylcholine (Ach) and 5-hydroxytryptamine (5-HT) (Kuribaraet al., 2000; Tsai et al., 1995; Xu et al., 2008). However, 5-HT andcholinergic receptor subtype antagonists could not block theinhibitory effects of magnolol on 5-HT or Ach release (Kuribaraet al., 2000; Tsai et al., 1995). Therefore, the effects of magnololon GABA receptor seem to be primary, while its impact on otherneurotransmission pathways might be secondary.
GABAA receptors are ligand-gated chloride channels assembledfrom a family of 16 homologous subunits including a1e6, b1e3, g1e3,d, ε, p, and q. The most widely distributed GABAA receptors in thehuman brain are composed of two a1, two b2, and one g2 subunit(Enna and Mohler, 2007; Hadingham et al., 1995; Olsen andSieghart, 2008; Ymer et al., 1990). The GABAA receptor can bemodulated by allosteric regulators such as the classical agonist BZ.The pharmacological properties of GABAA receptors are largelydetermined by their subunit composition (Rudolph et al., 2001). BZaction requires the presence of both a and g subunits, but notb subunit (Wafford et al., 1993;Wieland et al., 1992). Magnolol mostefficiently modulates GABAA receptors containing a and b subunits(Ai et al., 2001; Taferner et al., 2011), but not g subunit (Ai et al.,2001). In addition, magnolol also selectively up-regulates theexpression of the GABAA receptor a subunit, elevates both thenumber of binding sites and the affinity of [3]H-muscimol’s bindingto GABAA receptor a-subunit, and increases Cl� influx (Ai et al.,2001; Squires et al., 1999). However, magnolol showed no effecton the abundance of g subunits (Ai et al., 2001; Ma et al., 2009).These results suggest that hypnotic effect of magnolol might bemediated by the GABAA receptor a subunit.
EEG delta wave activity has been used as an indicator of thedepth of NREM sleep (Tobler et al., 2001). Although diazepam andmagnolol promote sleep, diazepam suppresses the EEG delta waveactivity of NREM sleep, whereasmognolol does not.Why the effectsof magnolol on sleep are different from those of benzodiazepinesstill remains unknown. Probably, they act at different GABAAreceptor a subunits. Diazepam binds to GABAA receptors containinga1, a2, a3, and a5 subunits, but its ability to inhibit REM sleep and toalter EEG spectra density was found to be mediated by a2-con-taining GABAA receptors (Kopp et al., 2004), but not by a1- or a3-containing GABAA receptors (Kopp et al., 2003; Tobler et al., 2001). Itwas reported that magnolol most efficiently modulated receptorscontaining a1, a2, a3 and a5 subunits (Ai et al., 2001; Taferner et al.,2011). In addition, magnolol increased the number of binding sites
and the binding affinity on a1b2g2 and a1b2 combinations in in vitrosaturation binding assays (Ai et al., 2001). Honokiol, another activeingredient of the Chinese herb houpo also does not alter the EEGpower density of NREM sleep but increases NREM sleep withouteffects on REM sleep (Qu et al., 2012). Honokiol enhanced chloridecurrents through GABAA receptors of seven different subunitcompositions and was most effective at those containinga3b2> a2b2> a1b2> a1b1 subunits (Taferner et al., 2011), indicatingthat magnolol and honokiol may more effective on a1 and a3,respectively, than a2-containing receptors, and thus have lessinfluence on EEG delta activity of NREM sleep. Taken together, thesefindings indicate that sleep promotion by magnolol, honokiol andbenzodiazepine is mediated by different GABAA receptor subunits.No alterations of honokiol and magnolol on delta power density ofNREM sleep suggest that they may be suitable for the treatment ofinsomnia because they increase sleep similar to physiological sleep.
Fos expression can be utilized as biomarker of neural activationby magnolol. The VLPO is a crucial site for the regulation of sleep.The VLPO contains one essential population of sleep-promotingneurons, which are more active during sleep, as indicated by theexpression of c-Fos (Gong et al., 2000; Lu et al., 2000; Saper et al.,2005; Sherin et al., 1996). GABAergic neurons are found to beclustered in the region of the VLPO. They make up roughly 80% ofthe neurons of the VLPO (Sherin et al., 1998). The sleep-positiveneurons in the VLPO innervate the histaminergic neurons in theTMN (Chou et al., 2002; Sherin et al., 1996). These histaminergiccones are closely linked to the transition between arousal andNREM sleep (Huang et al., 2006, 2007). Histaminergic output fromthe TMN is thought to play an important role in the mediation offorebrain arousal (Lin et al., 1986; Monti, 1993). Inhibition of theseneurons by the VLPO may play a major role in causing sleep. In thepresent study, we observed that magnolol increased c-Fos proteinexpression in the VLPO and decreased expression in the TMN,indicating that the hypnotic effects of magnolol may be mediatedby activation of the sleep center VLPO and inhibiting the neurons ofthe TMN.
Conflicts of interest
Except as noted in the acknowledgment, all authors declare thatno financial support or compensation has been received from anyindividual or corporate entity for any research or professionalservice at any time during the past 3 years. All authors declare noconflict of interest.
Acknowledgments
We are grateful to Dou Yin and to Yi-Ying Li for excellent tech-nical support. This study was supported in part by grants-in-aid forscientific research from National Natural Science Foundation ofChina (30970955, 30901797, 31070957, 31171049, 31171010,31121061), Shanghai Committee of Science and Technology(10XD1400400, 10441901600), National Basic Research Program ofChina (2009CB5220004, 2011CB711000), Shanghai LeadingAcademic Discipline Project (B119), China National Science andTechnology Major Project for Drug Discovery (2009ZX09303-006),and Program of Basic and Applied Researches for Innovations inBio-oriented Industry of Japan.
References
Ai, J., Wang, X., Nielsen, M., 2001. Honokiol and magnolol selectively interact withGABAA receptor subtypes in vitro. Pharmacology 63, 34e41.
Alexeev, M., Grosenbaugh, D.K., Mott, D.D., Fisher, J.L., 2012. The natural productsmagnolol and honokiol are positive allosteric modulators of both synaptic andextra-synaptic GABA(A) receptors. Neuropharmacology 62, 2507e2514.

C.-R. Chen et al. / Neuropharmacology 63 (2012) 1191e1199 1199
Buckner, J.D., Bernert, R.A., Cromer, K.R., Joiner, T.E., Schmidt, N.B., 2008. Socialanxiety and insomnia: the mediating role of depressive symptoms. DepressAnxiety 25, 124e130.
Chen, C., Tan, R., Qu, W., Wu, Z., Wang, Y., Urade, Y., Huang, Z., 2011. Magnolol,a major bioactive constituent of the bark of Magnolia officinalis, exerts antiep-ileptic effects via the GABA/benzodiazepine receptor complex in mice. Br. J.Pharmacol. 164, 1534e1546.
Chou, T.C., Bjorkum, A.A., Gaus, S.E., Lu, J., Scammell, T.E., Saper, C.B., 2002. Afferentsto the ventrolateral preoptic nucleus. J. Neurosci. 22, 977e990.
Enna, S.J., Mohler, H., 2007. The GABA Receptors (NJ).Franklin, K.B.J., Paxinos, G., 1997. The Mouse Brain in Stereotaxic Coordinates.
Academic, San Diego.Gong, H., Szymusiak, R., King, J., Steininger, T., McGinty, D., 2000. Sleep-related c-Fos
protein expression in the preoptic hypothalamus: effects of ambient warming.Am. J. Physiol. Regul. Integr. Comp. Physiol. 279, R2079eR2088.
Hadingham, K.L., Wafford, K.A., Thompson, S.A., Palmer, K.J., Whiting, P.J., 1995.Expression and pharmacology of human GABAA receptors containing gamma 3subunits. Eur. J. Pharmacol. 291, 301e309.
Huang, Z.L., Mochizuki, T., Qu, W.M., Hong, Z.Y., Watanabe, T., Urade, Y., Hayaishi, O.,2006. Altered sleep-wake characteristics and lack of arousal response to H3receptor antagonist in histamine H1 receptor knockout mice. Proc. Natl. Acad.Sci. U. S. A. 103, 4687e4692.
Huang, Z.L., Urade, Y., Hayaishi, O., 2007. Prostaglandins and adenosine in theregulation of sleep and wakefulness. Curr. Opin. Pharmacol. 7, 33e38.
Kopp, C., Rudolph, U., Keist, R., Tobler, I., 2003. Diazepam-induced changes on sleepand the EEG spectrum in mice: role of the alpha3-GABA(A) receptor subtype.Eur. J. Neurosci. 17, 2226e2230.
Kopp, C., Rudolph, U., Low, K., Tobler, I., 2004. Modulation of rhythmic brain activityby diazepam: GABA(A) receptor subtype and state specificity. Proc. Natl. Acad.Sci. U. S. A. 101, 3674e3679.
Kuribara, H., Kishi, E., Hattori, N., Okada, M., Maruyama, Y., 2000. The anxiolyticeffect of two oriental herbal drugs in Japan attributed to honokiol frommagnolia bark. J. Pharm. Pharmacol. 52, 1425e1429.
Lin, J.S., Sakai, K., Jouvet, M., 1986. Role of hypothalamic histaminergic systems inthe regulation of vigilance states in cats. C R Acad. Sci. III 303, 469e474.
Lin, Y.R., Chen, H.H., Ko, C.H., Chan, M.H., 2005. Differential inhibitory effects ofhonokiol and magnolol on excitatory amino acid-evoked cation signals andNMDA-induced seizures. Neuropharmacology 49, 542e550.
Lu, J., Greco, M.A., Shiromani, P., Saper, C.B., 2000. Effect of lesions of the ventro-lateral preoptic nucleus on NREM and REM sleep. J. Neurosci. 20, 3830e3842.
Ma, H., Kim, C.S., Ma, Y., Nam, S.Y., Kim, D.S., Woo, S.S., Hong, J.T., Oh, K.W., 2009.Magnolol enhances pentobarbital-induced sleeping behaviors: possibleinvolvement of GABAergic systems. Phytother Res. 23, 1340e1344.
Monti, J.M., 1993. Involvement of histamine in the control of the waking state. LifeSci. 53, 1331e1338.
Oishi, Y., Huang, Z.L., Fredholm, B.B., Urade, Y., Hayaishi, O., 2008. Adenosine in thetuberomammillary nucleus inhibits the histaminergic system via A1 receptorsand promotes non-rapid eye movement sleep. Proc. Natl. Acad. Sci. U. S. A. 105,19992e19997.
Olsen, R.W., Sieghart, W., 2008. International Union of Pharmacology. LXX. Subtypesof gamma-aminobutyric acid(A) receptors: classification on the basis of subunitcomposition, pharmacology, and function. Update Pharmacol. Rev. 60, 243e260.
Qu, W.M., Huang, Z.L., Xu, X.H., Matsumoto, N., Urade, Y., 2008. Dopaminergic D1and D2 receptors are essential for the arousal effect of modafinil. J. Neurosci. 28,8462e8469.
Qu, W.M., Xu, X.H., Yan, M.M., Wang, Y.Q., Urade, Y., Huang, Z.L., 2010. Essential roleof dopamine D2 receptor in the maintenance of wakefulness, but not inhomeostatic regulation of sleep, in mice. J. Neurosci. 30, 4382e4389.
Qu, W.M., Yue, X.F., Sun, Y., Fan, K., Chen, C.R., Hou, Y.P., Urade, Y., Huang, Z.L., 2012.Honokiol promotes non-rapid eye movement sleep via the benzodiazepine siteof the GABA(A) receptor in mice. Br. J. Pharmacol.
Rudolph, U., Crestani, F., Mohler, H., 2001. GABA(A) receptor subtypes: dissectingtheir pharmacological functions. Trends Pharmacol. Sci. 22, 188e194.
Rudolph, U., Knoflach, F., 2011. Beyond classical benzodiazepines: novel therapeuticpotential of GABAA receptor subtypes. Nat. Rev. Drug Discov. 10, 685e697.
Saper, C.B., Scammell, T.E., Lu, J., 2005. Hypothalamic regulation of sleep andcircadian rhythms. Nature 437, 1257e1263.
Sherin, J.E., Elmquist, J.K., Torrealba, F., Saper, C.B., 1998. Innervation of histamin-ergic tuberomammillary neurons by GABAergic and galaninergic neurons in theventrolateral preoptic nucleus of the rat. J. Neurosci. 18, 4705e4721.
Sherin, J.E., Shiromani, P.J., McCarley, R.W., Saper, C.B., 1996. Activation of ventro-lateral preoptic neurons during sleep. Science 271, 216e219.
Squires, R.F., Ai, J., Witt, M.R., Kahnberg, P., Saederup, E., Sterner, O., Nielsen, M.,1999. Honokiol and magnolol increase the number of [3H] muscimol bindingsites three-fold in rat forebrain membranes in vitro using a filtration assay, byallosterically increasing the affinities of low-affinity sites. Neurochem. Res. 24,1593e1602.
Taferner, B., Schuehly, W., Huefner, A., Baburin, I., Wiesner, K., Ecker, G.F., Hering, S.,2011. Modulation of GABAA-receptors by honokiol and derivatives: subtypeselectivity and structure-activity relationship. J. Med. Chem. 54, 5349e5361.
Tobler, I., Kopp, C., Deboer, T., Rudolph, U., 2001. Diazepam-induced changes insleep: role of the alpha 1 GABA(A) receptor subtype. Proc. Natl. Acad. Sci. U. S. A.98, 6464e6469.
Tsai, T.H., Chou, C.J., Chen, C.F., 1995. Glucuronidation of magnolol assessed usingHPLC/fluorescence. Planta Med. 61, 491e492.
van Lier, H., Drinkenburg, W.H., van Eeten, Y.J., Coenen, A.M., 2004. Effects ofdiazepam and zolpidem on EEG beta frequencies are behavior-specific in rats.Neuropharmacology 47, 163e174.
Wafford, K.A., Bain, C.J., Whiting, P.J., Kemp, J.A., 1993. Functional comparison of therole of gamma subunits in recombinant human gamma-aminobutyric acidA/benzodiazepine receptors. Mol. Pharmacol. 44, 437e442.
Wieland, H.A., Luddens, H., Seeburg, P.H., 1992. A single histidine in GABAAreceptors is essential for benzodiazepine agonist binding. J. Biol. Chem. 267,1426e1429.
Xu, Q., Yi, L.T., Pan, Y., Wang, X., Li, Y.C., Li, J.M., Wang, C.P., Kong, L.D., 2008. Anti-depressant-like effects of the mixture of honokiol and magnolol from the barksof Magnolia officinalis in stressed rodents. Prog. Neuropsychopharmacol. Biol.Psychiatry 32, 715e725.
Ymer, S., Draguhn, A., Wisden, W., Werner, P., Keinanen, K., Schofield, P.R.,Sprengel, R., Pritchett, D.B., Seeburg, P.H., 1990. Structural and functionalcharacterization of the gamma 1 subunit of GABAA/benzodiazepine receptors.EMBO J. 9, 3261e3267.
Zammit, G.K., 2007. The prevalence, morbidities, and treatments of insomnia. CNSNeurol. Disord. Drug Targets 6, 3e16.





























