-
www.elsevierhealth.com/journals/tube
Tuberculosis
Human immunity to M. tuberculosis: T cell subsetsand antigen
processing
W.H. Boom*, David H. Canaday, Scott A. Fulton, Adam J.
Gehring,Roxana E. Rojas, Marta Torres
Tuberculosis Research Unit (TBRU), Case Western Reserve
University and Division of Infectious Diseases,University Hospitals
of Cleveland, OH, USA
Summary A hallmark of M. tuberculosis infection is the ability
of most (9095%)healthy adults to control infection through acquired
immunity, in which antigenspecific Tcells and macrophages arrest
growth of M. tuberculosis bacilli and maintaincontrol over
persistent bacilli. In addition to CD4+ Tcells, other Tcell subsets
such as,gd, CD8+ and CD1-restricted T cells have roles in the
immune response to M.tuberculosis. A diverse T cell response allows
the host to recognize a wider range ofmycobacterial antigens
presented by different families of antigen-presentingmolecules, and
thus greater ability to detect the pathogen. Macrophages are
keyantigen presenting cells for T cells, and M. tuberculosis
survives and persists in thiscentral immune cell. This is likely an
important factor in generating this T celldiversity. Furthermore,
the slow growth and chronic nature of M. tuberculosisinfection
results in prolonged exposure to antigens, and hence further T
cellsensitization. The effector mechanisms used by T cells to
control M. tuberculosis arepoorly understood. To survive in
macrophages, M. tuberculosis has evolvedmechanisms to block immune
responses. These include modulation of phagosomes,neutralization of
macrophage effector molecules, stimulating the secretion
ofinhibitory cytokines, and interfering with processing of antigens
for T cells. Therelative importance of these blocking mechanisms
likely depends on the stage ofM. tuberculosis infection: primary
infection, persistence, reactivation or activetuberculosis. The
balance of the hostpathogen interaction in M. tuberculosisinfection
is determined by the interaction of T cells and infected
macrophages. Theoutcome of this interaction results either in
control of M. tuberculosis infection oractive disease. A better
understanding of this interaction will result in improvedapproaches
to treatment and prevention of tuberculosis.r 2003 Elsevier Science
Ltd. All rights reserved.
Natural history of M: tuberculosisinfection
Aerosolized M. tuberculosis bacilli are efficientlytransmitted
from person to person. Only small
numbers of bacilli need enter distal alveoli ofhuman lungs to
establish infection. In most persons,local innate immunity,
mediated primarily byalveolar macrophages, fails to control the
slowlyreplicating bacilli. As a result, the immune systemis exposed
to increasing amounts of mycobacterialantigen resulting in
development of adaptiveimmunity. In most healthy adults, adaptive
immu-nity mediated by T cells controls but does noteradicate M.
tuberculosis infection. Thus, ongoing
*Corresponding author. Tuberculosis Research Unit, CaseWestern
Reserve University, 10900 Euclid Ave., Cleveland, OH44106-4984,
USA. Tel.: +1-216-368-4844; fax: +1-216-368-2034E-mail address:
[email protected] (W.H. Boom).
1472-9792/03/$ - see front matter r 2003 Elsevier Science Ltd.
All rights reserved.doi:10.1016/S1472-9792(02)00054-9
Tuberculosis (2003) 83, 98106
-
protective immunity is required to maintain controlover
persistent bacilli, which are thought to bepresent in one-third of
the worlds population.Adaptive immunity also protects against
reinfec-tion. The latter is particularly important in areaswith
high levels of M. tuberculosis transmission.During the last 10
years great progress has been
made in understanding the human immune responseto M.
tuberculosis. The interaction of T cells andinfected macrophages is
central to protectiveimmunity to M. tuberculosis. CD4+ T cells have
anessential role but are supported by other T cellsubsets such as
CD8+, gd TCR+ Tcells (gd Tcells), andCD1 restricted T cells.
Antigens for many of these Tcells have been defined, and their
function in theimmune response is starting to be elucidated.
Insighthas been gained into mechanisms used by macro-phages to
control M. tuberculosis. How T cells helpthem perform this task
remains poorly understood.TNF-a, IL-12 and IFN-g are central
cytokines inregulatory and effector phases of the immuneresponse to
M. tuberculosis. Macrophages are notonly primary effector cells for
control of M.tuberculosis but also essential for processing
andpresentation of antigens to T cells. To survive (andthrive) in
macrophages, M. tuberculosis has evolvedmechanisms to block immune
responses. Theseinclude modulation of phagosomes, neutralizationof
macrophage effector molecules, inducing secre-tion of inhibitory
cytokines, and interference withprocessing of antigens for T
cells.The relative importance of different T cell
subsets and mechanisms employed by M. tubercu-losis to interfere
with macrophage and T cellfunction likely depends on the phase of
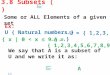
theinfection. As outlined in Fig. 1, after an initialinnate phase,
acute adaptive immunity develops tocontrol rapidly dividing
bacilli. This is followed by achronic memory immune phase necessary
forcontrol of persistent bacilli and surveillance for
possible re-infections. Immune failure during acuteor chronic
adaptive immunity results in clinicaltuberculosis, resulting in M.
tuberculosis spreadingto a new host. The balance of the
hostpathogeninteraction in M. tuberculosis infection is deter-mined
by the interaction of T cells and infectedmacrophages. The ability
of M. tuberculosis tomodulate antigen presenting and
microbicidalfunction of macrophages is essential for its survivalas
one of the most successful human pathogens.Current knowledge about
the role and function ofT cell subsets and the mechanisms used by
M.tuberculosis to interfere with the ability of macro-phages to
process T cell antigens and with T cellfunction, will be
reviewed.
The central role of CD4+ T cells inprotective immunity to M:
tuberculosis
Studies in humans and animal models demonstratethat acquired
immunity to M. tuberculosis requirescontributions by multiple T
cell subsets, whichinclude a dominant role for CD4+ T cells
andsignificant roles for CD8+ and gd Tcells.1 The reasonsfor
involvement of these multiple T cell subsets arenot known.
Diversity in T cells that differ in antigenprocessing mechanisms
and molecules used forantigen presentation greatly expands the
repertoireof mycobacterial antigens recognized. The ability ofM.
tuberculosis to survive and persist in one of themajor
antigen-presenting cells, the macrophage,contributes further to
ready T cell activation. Theslow growth and chronic nature of M.
tuberculosisinfection results in prolonged exposure to a
largediversity of antigens. From the hosts perspective, adiverse T
cell repertoire allows recognition of a widerange of mycobacterial
antigens presented bydifferent families of antigen-presenting
molecules,
Bact
eria
l Loa
d
Innate (TST -)Adaptive (TST +)
Latent Infection (90%)
ReactivationDisease (5-10%)
1 ProgressiveDisease (< 5%)
Acute ChronicImm
unity
Figure 1 Natural history of M. tuberculosis infection. TST is
Tuberculin Skin Test.
Human immunity to M. tuberculosis 99
-
and thus greater ability to detect the pathogen. ForM.
tuberculosis to be such a successful persistentpathogen requires
that it evolve mechanisms tointerfere with T cell function.CD4+
abTCR+ T cells (CD4+ T cells) are central to
the human immune response to M. tuberculosis.The HIV pandemic
provides direct evidence thatloss of CD4+ T cell number and
function resulted inprogressive primary infection, reactivation of
en-dogenous M. tuberculosis and enhanced suscept-ibility to
re-infection.24 Early cell transfer studiesin mice established that
CD4+ T cells transferedprotection against M. tuberculosis.5 In vivo
deple-tion with anti-CD4 monoclonal antibodies con-firmed these
findings.6,7 Mice with deleted genesfor CD4 or MHC class II
molecules are markedlysusceptible to M. tuberculosis, firmly
establishing acentral role for CD4+ T cells in protection.8,9
Human CD4+ T cells activated by M. tuberculosisantigens and
specific CD4+T cells secrete themacrophage activating cytokines
IFN-g and TNF-a/b, are cytotoxic (CTL) for M. tuberculosisinfected
macrophages and help macrophages con-trol intracellular
mycobacteria.1018 As CTL, afteractivation by M. tuberculosis, CD4+
T cells expressgranzymes, Fas-L (CD95L), granulysin, and
perfor-in.19 However, its not clear if CTL function isassociated
with increased control of M. tubercu-losis in macrophages. CD4+ T
cells provide help forgd and CD8+T cells through IL-2 secretion.
All thesefunctions are likely important for CD4 Tcells role
inprotective immunity but may be expressed differ-entially during
the different stages of the infection.CD4+ T cells however, clearly
are critical at allstages of M. tuberculosis infection.The
importance of IFN-g was demonstrated by
the increased susceptibility to mycobacterial in-fection of
children with IFN-gR deficiency.20,21 IFN-gR knock-out mouse also
are markedly susceptibleto M. tuberculosis.22,23 How IF-g mediates
protec-tion is less clear. In humans, IFN-g alone isinefficient in
enhancing the ability of humanmacrophages to control M.
tuberculosis.24,25 Inmice, protective immunity by CD4+ T cells
cannotbe explained by IFN-g alone, since MHC class II andCD4
knock-out mice produce large amounts of thiscytokine yet fail to
control M. tuberculosis.8
Besides activating microbial killing mechanisms,IFN-g also is a
critical regulator of antigen-present-ing cell function by
increasing MHC and co-stimulatory molecule expression. This latter
activ-ity is essential for optimal T cell responses to
M.tuberculosis, and may represent the primary rolefor IFN-g in
mycobacterial immunity. In addition toIFN-g, TNFa and IL-12 have
been demonstrated inhuman and murine models to have critical roles
in
protective immunity to M. tuberculosis.26,27 IL-12functions to
increase IFN-g production. How TNF-gexerts its protective effect in
humans is not defined.The antigen repertoire for human M.
tuberculosis
specific CD4+ T cells is characterized by responsesto a large
number of antigens.17,28,29 CD4+T cellsrecognize mycobacterial
peptide fragments pre-sented to them by MHC class II molecules
onantigen-presenting cells such as macrophages. Nosingle
immunodominant antigen has emerged todate, but a number of antigens
have beenidentified that are recognized by a majority ofhealthy
tuberculin skin-test positive persons. Theseinclude the three
3032kDa 85 complex proteins,ESAT-6 and CFP-10, the 19 and 38 kDa
lipoproteinsand two recently described proteins of 32
(serineprotease) and 39 kDa.3036 The 3032 kDa 85A,B,Ccomplex
antigens are mycolyl transferases involvedin mycobacterial cell
wall synthesis, and arerestricted to mycobacterial species.37 85B,
themajor 30 kDa protein of M. tuberculosis, is recog-nized readily
by T cells from M. tuberculosisinfected persons, and a number of
MHC class IIrestricted epitopes have been mapped.38 Genes forESAT-6
and CFP-10 are present in M. tuberculosisbut deleted in all BCG
substrains (region ofdifference 1, RD-1, between M. tuberculosis
andM. bovis) and thus are of particular interest fordiagnostic
tests aimed at distinguishing thesemycobacterial species. Although
CD4+ T cell re-activity to the 19 and 38 kDa lipoproteins can
befound, these proteins have been disappointing inanimal vaccine
studies. They have, however, gainedpromise as stimuli of toll-like
receptors (TLR) onmacrophages. Proteins of 32 and 39 kDa
wereidentified by screening protein antigens of M.tuberculosis for
reactive T cells from healthytuberculin skin-test positive persons.
Vaccine stu-dies in animal models by combining ESAT-6 with 85Band
32 kDa with the 39 kDa protein have beenpromising, warranting their
development for test-ing as subunit vaccines in humans. Access to
the M.tuberculosis genome will greatly facilitate theidentification
and expression of additional CD4+T cell antigens. Studies in human
populations withdifferent susceptibilities and responses to
M.tuberculosis will determine which antigens andfunctions are
critical for CD4+ T cells during thedifferent phase of M.
tuberculosis infection.
MHC class I restricted CD8+ T cells
CD8+abTCR+ T cells (CD8+T cells) are activated byM. tuberculosis
and BCG.3941 M. tuberculosis
100 W.H. Boom et al.
-
reactive CD8+ T cells are found among alveolar andperipheral T
cells of healthy tuberculin skin-testpositive persons.39 M.
tuberculosis activated CD8+ Tcells secrete IFN-g, but less than
CD4+ T cells.40 Theyexpress granzymes, Fas-L (CD95L), granulysin,
andperforin, which enables them to lyse infected macro-phages.19
CD8+ T cells can help macrophages controlintracellular
mycobacteria. Murine studies establisheda role for MHC-I restricted
CD8+ T cells in protectiveimmunity particularly during the later
stages ofinfection in the lung. To what extent protection byCD8+ T
cells depends on CTL activity is unclear. Forhumans, it is not
known at what stages of M.tuberculosis infection CD8+ T cells are
most critical.The majority of M. tuberculosis reactive CD8+
T cells recognize mycobacterial peptides in thecontext of MHC
class I molecules. Epitope mappingstudies, usually guided by
epitope predictionalgorithms, have defined HLA-A2 restricted
epi-topes on already defined M. tuberculosis antigenssuch as
ESAT-6, 85B, 85A, the 19k Da lipoprotein andCFP-10.4245 Others have
used a proteinomicapproach by eluting peptides from human MHCclass
I molecules (HLA-A2).46 This approach canidentify novel M.
tuberculosis antigens by deter-mining the epitopes presented by
MHC-I moleculeson infected cells. Whether the repertoire ofantigens
recognized by CD8+ and CD4+ T cellsoverlap or are distinct will be
determined as moreantigens for both T cell populations are defined
byproteinomic approaches. Class I and class II MHCmolecules have
distinct intracellular traffickingpathways and mechanisms for
loading of microbialpeptides. These different pathways have a key
rolein determining the repertoire of antigens recog-nized by CD4+
and CD8+ T cells.M. tuberculosis antigens can enter the MHC
class
I antigen processing pathway in human macro-phages through an
alternate pathway, that does notrequire traffic through the classic
ER-Golgi path-way.40 Traditionally, class I MHC molecules
presentmicrobial peptides derived from antigens present incytosol
either through de novo synthesis (viralantigens) or carried by
microbial agents capableof penetrating into the cytoplasm (e.g.
Listeriamonocytogenes, Leishmania etc.). Once in thecytosol,
antigens are cleaved by proteasomes andpeptide fragments delivered
to the endoplasmicreticulum, where they bind to newly
synthesizedclass I MHC molecules and are transported toplasma
membrane for recognition by CD8+ T cells.Particulate bacterial
antigens, such as M. tubercu-losis, can be processed for class I
MHC presentationby an alternate pathway that does not
requirepenetration into cytosol and thus providing anadditional
mechanism for activation of CD8+ Tcells.
cd T cells and other T cell populations
gd TCR expressing T cells (gd T cells) are character-ized by a
unique T cell antigen receptor (TCR)comprised of g and d chains.
The majority ofcirculating gd Tcells in adults express Vd9 (aka
Vg2)and Vd2 elements (Vd2+ T cells). M. tuberculosisbacilli readily
activate Vd2+ Tcells.47 Similar to CD4and CD8+ Tcells, Vd2+Tcells
secrete IFN-g, can lyseinfected macrophages and can help contain
myco-bacterial growth.17,48 Individuals sensitized tomycobacterial
antigens have a greater ability toactivate gd-T cells in response
to M. tuberculosisthan tuberculin negative persons.49,50
Vaccinationof adults with BCG increases in vitro expansion ofVg9
Vd2 T cells after stimulation with M. tubercu-losis antigens.51
Tuberculosis patients have adiminished ability to activate gd T
cells in responseto M. tuberculosis.50 gd T cells are found in
thelungs of tuberculosis patients, and there is acorrelation
between the absence or loss of Vg9Vd2 Tcells in blood and lung, and
extent of disease.52 In aprimate model gd T cell number and
reactivity tophosphate antigens increases during primary
my-cobacterial infection and during challenge afterBCG vaccination,
suggesting a role in protectiveimmunity.53
Vd2+ T cells react to small phosphate containingmolecules that
can be divided into two groups.There are nucleotide-conjugated
phosphatemolecules, such as TUBAg3-4, isopentenyl-ATP,and
pyrophosphate molecules such as TUBag1-2,isopentenyl pyrophosphate
[IPP], mono-ethylpyrophosphate (MEPP) and others (reviewed
inRef.5456). Recognition of phospho-molecules isTCR-dependent but
not restricted or dependenton any known MHC or MHC-like molecules.
Speci-ficity of prenylphosphate recognition depends onparticular
CDR3 regions in the Vg9 chain and theVd2 chain. The phosphoantigens
are intracellularand not secreted by M. tuberculosis.
Prenylpyrophosphates are ubiquitous precursors for cho-lesterol and
its derivatives, and for terpenoids.Discrimination between infected
and non-infectedcells may involve recognition of metabolic
inter-mediates produced by bacteria.57 Thus, despitesimilar
functions, Vd2+ T cells recognize an entirelydifferent range of
mycobacterial molecules thanCD4+ and CD8+ Tcells. How these
phospho-antigensare processed and presented to gd T cells (if at
all)by infected macrophages is unknown.58,59 Even ifthere is no
antigen processing, macrophages areimportant for activating gd T
cells because wheninfected they represent a concentrated source
ofphospho-antigen and because they enhance gdT cell responses by
providing co-stimulation.
Human immunity to M. tuberculosis 101
-
One additional T cell subset that respondsto M. tuberculosis is
the CD1 restricted abTCR+ T cell.60 It is the least common T
cellsubset in human peripheral blood and lung. Inhumans, most of
these T cells express neitherCD4 or CD8 and are referred to as
doublenegative (DN) cells. A minority expresses CD8.CD1 is an
MHC-like molecule that associateswith b2-microglobulin that can be
induced bycytokines or constitutively expressed on
antigen-presenting cells such as dendritic cells. CD1molecules have
very little polymorphism andhave the unique ability to bind polar
non-peptide lipid antigens of M. tuberculosis suchas mycolic acids
and phosphatidyl-inositol-mannosides (PIMs). CD1 restricted T cells
secreteIFN-g, are cytotoxic for infected macrophages andcan help
macrophages control intracellular myco-bacteria.
Thus, the human T cell response to M. tubercu-losis is
characterized by the participation of multi-ple T cell subsets with
similar functions (Fig. 2).They secrete IFN-g and TNF-a, can lyse
infectedcells as CTL and can help macrophages control
M.tuberculosis growth. They differ markedly, how-ever in the range
of mycobacterial antigens theyrecognize and the antigen-processing
mecha-nisms used to process and present these antigensto their
TCRs. T cell subsets likely also differ instage of infection when
they are most active oressential, and in ability to enter sites
ofactive infection (granuloma, cavity, lymph node).Some have
suggested that gd T cells and CD1restricted T cells may provide a
link between theinnate and adaptive phases of the immuneresponse to
M. tuberculosis. CD4+ T cells areknown to have a key role through
all stages of M.tuberculosis infection but may differ as to
whencytokine secretion vs. CTL function is the mostimportant.
T cell inhibition by M: tuberculosis
The complexity and diversity of the adaptiveimmune response to
M. tuberculosis require devel-opment of mechanisms to interfere and
inhibit hostimmunity for pathogen survival. M. tuberculosisevolved
to survive and persist in key immuneeffector cells, the macrophage.
Macrophages read-ily take up microbial pathogens, have
effectivemicrobicidal mechanisms, process and presentantigens for T
cell recognition, and express thenecessary costimulatory molecules
to activateT cells.M. tuberculosis has evolved at least two
mechan-
isms to interfere with innate immune defenses ofmacrophages. M.
tuberculosis bacilli are taken upby receptor mediated phagocytosis
using a varietyof macrophage receptors including CR3, CR4
andmannose receptor. Mycobacteria remain withinphagosomes and do
not penetrate into cytoplasm.M. tuberculosis modulates its
phagosomal compart-ment by preventing fusion with acidic
lysosomalcompartments and actively excludes vesicularproton
ATP-ases, resulting in an elevated pH of6.36.5 (compared to the
normal lysosomal pH of4.5).61 IFNg can partially overcome M.
tubercu-losis mediated inhibition of phagosomal acidifica-tion. In
addition to modulation of the phagosome toprevent critical
proteases from attacking it, M.tuberculosis resists killing by
oxygen radical inter-mediates, through superoxide dismutases and
otherenzymes. M. tuberculosis is sensitive to nitric oxide(NO) made
by inducible nitric oxide synthase(iNOS), but whether enough iNOS
can be inducedin human macrophages to produce sufficient levelsof
NO remains controversial.M. tuberculosis also has a variety of
mechanisms
to interfere with adaptive immune function. First,molecules of
M. tuberculosis readily induce macro-phages to produce cytokines
that inhibit T cell
Phos. Ag
-TCRCD 4T cell
class II MHCclass IMHC
-TCRCD 8T cell
CD1 restricted T cellsDN TCR
M
T cell
-TCR
Figure 2 Many T cell subsets respond to M. tuberculosis infected
macrophages.
102 W.H. Boom et al.
-
function. These include inhibitory cytokines such asIL-10 and
TGFb, and are produced by the samecells producing pro-inflammatory
and Th-1 promot-ing cytokines such as IL-12, IL-15, TNFa, and
IL-1.Excess production of IL-10 and TGFb, as seen inactive
tuberculosis, inhibits the effects of the pro-inflammatory
cytokines and directly inhibits T cellfunction.Recent studies
suggest that recognition of patho-
gen-associated molecular patterns (PAMPs) by Toll-like receptors
(TLR) may be a primary signal forcytokine release by macrophages.
There are 10known TLRs of which macrophages express many.62
TLR-4 is the primary receptor for LPS from Gram-negative
bacteria, and TLR-2 recognizes Gram-positive bacteria.6365 TLR-2
and TLR-4 recognizeM. tuberculosis associated PAMPs, with
TLR-2recognizing mycobacterial lipoproteins such as19 kDa.65,66
Signalling through TLR results in macro-phage cytokine secretion,
which includes both pro-inflammatory and inhibitory cytokine
secretion.Whether specific M. tuberculosis PAMPs differ inthe range
or balance of pro-inflammatory andinhibitory cytokines they
stimulate, is not known.Second, active M. tuberculosis infection
is
associated with increased apoptosis of mycobac-terial
antigen-specific T cells.67 M. tuberculosis-specific T cell
apoptosis has a prolonged effect on Tcell responses to
mycobacterial antigens. Long (612 months) after initiation of TB
treatment andcontrol of bacterial replication, CD4+ T cellresponses
to proteins of M. tuberculosis remainmarkedly diminished compared
to responses tocontrol antigens such as tetanus toxoid or
themitogen PHA.68 This suggests that T cell apoptosisduring acute
infection can result in prolongeddefects in T cell repertoire and
function. Themolecules of M. tuberculosis and mechanismresponsible
for T cell apoptosis have not beendefined.
Modulation of macrophage antigen-presenting cell function byM:
tuberculosis
Recent studies suggest an important third way forM. tuberculosis
to evade adaptive immune re-sponses, namely interfering with
antigen processingand presentation by macrophages. Processing
ofantigens is essential for T cells to recognize cellsinfected by
microbes and is regulated by complexcellular processes resulting in
peptide presentationby two distinct sets of molecules: MHC class II
forCD4+ and MHC class I for CD8+ T cells. Little is
known about inhibitors of antigen processing forMHC class I and
class I-like molecules. However,recent studies are starting to shed
light onmechanisms for inhibition of MHC class II antigenprocessing
and the molecule(s) of M. tuberculosisresponsible for this
inhibition.For class II MHC processing, antigens normally
are internalized by endocytosis or phagocytosis,and concentrated
within endosomes. As endosomesfuse with lysosomes, proteases break
downprotein into peptides. Class II MHC molecules areconcentrated
in a late endocytic compartment, theclass II MHC compartment
(MIIC). Class II MHCpeptide complexes are formed in these
compart-ments with peptides binding to class II MHCpromoted by the
acidic pH. Class II MHC moleculesare targeted for endosomes by
specific sequenceson the invariant chain associated with the a and
bchains of class II MHC as it emerges from endoplas-mic reticulum
(ER). Invariant chain contains a CLIPregion which blocks the class
II MHC peptide bindingsite. CLIP is exchanged for antigen peptide
withhelp from HLA-DM molecules. In murine macro-phages M.
tuberculosis phagosomes contain MHC IIcapable of presenting
mycobacterial peptide di-rectly to T cells.69
M. tuberculosis infected monocytes do notpresent tetanus toxoid
as well as uninfected cells.70
In murine macrophages, IL-6 secretion has beenshown under some
circumstances to inhibit T cellfunction, and M. tuberculosis has
been shown tointerfere with maturation of MHC class II and
withIFN-g signaling in THP-1 cells.7174 In murinemacrophages,
mycobacterial infection was asso-ciated with decreased CIITA (class
II transactivator)expression, resulting in decreased MHC class
IIlevels.75 Recent studies indicate that the 19 kDalipoprotein of
M. tuberculosis through TLR-2inhibits MHC class II expression and
antigenprocessing for CD4+ T cells in murine macro-phages.76 These
studies have been extended tohuman macrophages, and indicate that
M. tuber-culosis and the 19 kDa lipoprotein inhibit IFN-gmediated
regulation of human HLA-DR.77 Thisresults in decreased presentation
and thus activa-tion of M. tuberculosis specific CD4+ T
cellresponses. The ability to interfere with CD4+ T cellactivation
by hiding from the immune responsesis likely to be a major
mechanism used by M.tuberculosis to avoid detection and
eliminationduring the persistent phase of infection. Usingcarefully
designed assays to measure specific stagesof antigen processing for
MHC class II and Imolecules, it is likely that additional
mycobacterialmolecules will be identified that can block
antigenprocessing.
Human immunity to M. tuberculosis 103
-
Thus, M. tuberculosis has evolved a number ofmechanisms to
interfere with activation of bothinnate and adaptive phases of the
immune re-sponse. It is likely that the importance of
thesedifferent mechanisms will differ for each stage ofM.
tuberculosis infection. For example, suppressivecytokines and
apoptosis may be more importantduring phases when M. tuberculosis
is replicatingrapidly during primary and reactivation
phases,whereas inhibition of antigen processing may bethe most
important defense mechanisms during thepersistent phase of
infection. The balance betweenactivation of multiple T cell subsets
capable ofrecognizing a wide range of mycobacterial mole-cules in
the context of a wide range of antigen-presenting molecules and the
ability of M. tuber-culosis to interfere with immune recognition
and toblock the effector phase of adaptive immunitylikely
determines the outcome of M. tuberculosisinfection, which ranges
from active disease toindefinite persistence. Access to genomic
andproteonomic information of M. tuberculosiscoupled with increased
knowledge of the cellbiology of antigen processing and T cell
activationwill undoubtedly characterize further the molecu-lar
mechanims of the critical host pathogeninteraction in M.
tuberculosis infection: the inter-action of T cell subsets with
infected macrophages.
Acknowledgements
This work was supported by National Institutes ofHealth grants
A127243 and HL55967, and contractA195383 to the Tuberculosis
Research Unit.
References
1. Boom WH. The role of T cell subsets in M.
tuberculosisinfection. Infectious Agents Dis 1996;5:7381.
2. Raviglione MC, Snider Jr. DE, Kochi A. Global epidemiologyof
tuberculosis. Morbidity and mortality of a worldwideepidemic [see
comments]. JAMA 1995;273:2206.
3. Barnes PF, Bloch AB, Davidson P, Snider D. Tuberculosis
inpatients with human immunodeficiency virus infection. NEngl J Med
1991;324:164450.
4. Hopewell PC. Impact of human immunodeficiency virusinfection
on the epidemiology, clinical features, manage-ment, and control of
tuberculosis. Clin Infect Dis 1992;15:5407.
5. Orme IM, Collins FM. Protection against
Mycobacteriumtuberculosis infection by adoptive transfer. J Exp
Med1983;158:7483.
6. Muller I, Cobbold S, Waldmann H, Kaufmann SH.
Impairedresistance to Mycobacterium tuberculosis infection
afterselective in vivo depletion of L3T4+ and Lyt-2+ Tcells.
InfectImmun 1987;55(9):203741.
7. Pedrazzini T, Hug K, Louis JA. Importance of L3T4+ and Lyt-2+
cells in the immunologic control of infection withMycobacterium
bovis strain bacillus Calmette-Guerin inmice. Assessment by
elimination of T cell subsets in vivo.J Immunol
1987;139(6):20327.
8. Caruso AMN, Serbina N, et al. Mice deficient in CD4 Tcells
have only transiently diminished levels of IFN-gamma,yet succumb to
tuberculosis. J Immunol 1999;162(9):540716.
9. Ladel CH, Daugelat S, et al. Immune response to
Mycobac-terium bovis bacille Calmette Guerin infection in
majorhistocompatability complex I- and II-deficient knock-outmice:
contribution of CD4+ and CD8+ T cells to acquiredresistance. Eur J
Immunol 1995;25:37784.
10. Boom WH, Wallis RS, Chervenak KA. Human
Mycobacteriumtuberculosis-reactive CD4+ T-cell clones:
heterogeneity inantigen recognition, cytokine production, and
cytotoxicityfor mononuclear phagocytes. Infect Immun
1991;59(8):273743.
11. Haanen JB, de-Waal-Malefijt R, Res PC, Kraakman EM,Ottenhoff
TH, de-Vries RR, Spits H. Selection of a human Thelper type 1-like
T cell subset by mycobacteria. J Exp Med1991;174(3):58392.
12. Hansen PW, Petersen CM, Povlsen JV, Kristensen T.
Cytotoxichuman HLA class II restricted purified protein
derivativereactive T lymphocyte clones. Scand J
Immunol1987;25:295303.
13. Mustafa AS, Godal T. BCG-induced suppressor T cells
optimalconditions for in vitro induction and mode of action. Clin
ExpImmunol 1985;62(3):47481.
14. Kumararatne DS, Pithie AS, Drysdale P, Gaston JS,
KiesslingR, Iles PB, Ellis CJ, Innes J, Wise R. Specific lysis
ofmycobacterial antigen-bearing macrophages by class II
MHC-restricted polyclonal T cell lines in healthy donors orpatients
with tuberculosis. Clin Exp Immunol 1990;80(3):31423.
15. Ottenhoff TH, Ab BK, Van-Embden JD, Thole JE, Kiessling
R.The recombinant 65-kD heat shock protein of Mycobacter-ium bovis
Bacillus Calmette-Guerin/M. tuberculosis is atarget molecule for
CD4+ cytotoxic T lymphocytes that lysehuman monocytes. J Exp Med
1988;168(5):194752.
16. Hancock GE, Cohn ZA, Kaplan G. The generation of
antigen-specific, major histocompatability complex-restricted
cyto-toxic T lymphocytes of the CD4+ phenotype. J Exp
Med1989;169:90919.
17. Tsukaguchi K, Balaji KN, et al. CD4+ alpha-beta T cell
andgamma delta T cell responses to Mycobacterium tubercu-losis:
similarities and differences in antigen recognitioncytotoxic
effector function and cytokine production.J Immunol
1995;154:178696.
18. Tsukaguchi K, de Lange B, et al. Differential regulation
ofIFN-gamma, TNF-alpha, and IL-10 production by CD4(+)alphabetaTCR+
T cells and vdelta2(+) gammadelta T cells inresponse to monocytes
infected with Mycobacterium tuber-culosis-H37Ra. Cell Immunol
1999;194(1):1220.
19. Canaday DH, Wilkinson RJ, Li Q, Harding CV, Silver RF,
BoomWH. CD4+ and CD8+ Tcells kill intracellular M. tuberculosisby a
perforin and FAS/FASL independent mechanism.J Immunol
2001;167:273442.
20. Jouanguy E, Altare F, et al.
Interferon-gamma-receptordeficiency in an infant with fatal bacille
Calmette-Guerininfection. N Engl J Med 1996;335(26):195661.
21. Jouanguy E, Lamhamedi-Cherradi S, et al. Partial
interferon-gamma receptor 1 deficiency in a child with tuber-culoid
bacillus Calmette-Guerin infection and a sibling withclinical
tuberculosis. J Clin Invest 1997;100(11):265864.
104 W.H. Boom et al.
-
22. Flynn JL, Chan J, et al. An essential role for
interferongamma in resistance to Mycobacterium tuberculosis
infec-tion. J Exp Med 1993;178(6):224954.
23. Cooper AM, Dalton DK, et al. Disseminated tuberculosis
ininterferon-gamma gene-disrupted mice. J Exp Med
1993;178:22437.
24. Douvas GS, Looker DL, et al. Gamma interferon activateshuman
macrophages to 4become tumoricidal and leishma-nicidal but enhances
replication of macrophage-associatedmycobacteria. Infect Immun
1985;50(1):18.
25. Silver RF, Li Q, et al. Lymphocyte-dependent inhibition
ofgrowth of virulent Mycobacterium tuberculosis H37Rv withinhuman
monocytes: requirement for CD4 + T cells in purifiedprotein
derivative-positive, but not in purified proteinderivative-negative
subjects. J Immunol 1998;160(5):240817.
26. de Jong R, Altare F, Haagen IA, Elferink DG, Boer T,
vanBreda Vriesman PJ, Kabel PJ, Draaisma JM, van Dissel JT,Kroon
FP, Casanova JL, Ottenhoff TH. Severe mycobacterialand Salmonella
infections in interleukin-12 receptor-defi-cient patients. Science
1998;280:14358.
27. Keane J, Gershon S, Wise RP, Mirabile-Levens E, Kasznica
J,Schwieterman WD, Siegel JN, Braun MM. Tuberculosisassociated with
infliximab, a tumor necrosis factor alpha-neutralizing agent. N
Engl J Med 2001;345:1098104.
28. Havlir DV, Wallis RS, Boom WH, Daniel TM, Chervenak K,Ellner
JJ. Human immune response to Mycobacteriumtuberculosis antigens.
Infect Immun 1991;59(2):66570.
29. Schoel B, Gulle H, Kaufmann SH. Heterogeneity of
therepertoire of T cells of tuberculosis patients and
healthycontacts to Mycobacterium tuberculosis antigens separatedby
high-resolution techniques. Infect Immun 1992;60(4):171720.
30. Wiker HG, Harboe M. The antigen 85 complex: a majorsecretion
product of Mycobacterium tuberculosis. MicrobiolRev
1992;56(4):64861.
31. Sorensen AL, Nagai S, Houen G, Andersen P, Andersen
A.Purification and characterization of a low-molecular-mass Tcell
antigen secreted by Mycobacterium tuberculosis. InfectImmun
1995;63:17107.
32. Dillon DC, Alderson MR, Day CH, Bement T, Campos-Neto
A,Skeiky YA, Vedvick T, Badaro R, Reed SG, Houghton R.Molecular and
immunological characterization of Mycobac-terium tuberculosis
CFP-10, an immunodiagnostic antigenmissing in Mycobacterium bovis
BCG. J Clin Microbiol2000;38:328590.
33. Skeiky YA, Lodes MJ, Guderian JA, Mohamath R, Bement
T,Alderson MR, Reed SG. Cloning, expression, and immunolo-gical
evaluation of two putative secreted serine proteaseantigens of
Mycobacterium tuberculosis. Infect Immun1999;67:39984007.
34. Dillon DC, Alderson MR, Day CH, Lewinsohn DM, Coler R,Bement
T, Campos-Neto A, Skeiky YA, Orme IM, Roberts A,Steen S, Dalemans
W, Badaro R, Reed SG. Molecularcharacterization and human T-cell
responses to a memberof a novel Mycobacterium tuberculosis mtb39
gene family.Infect Immun 1999;67:294150.
35. Surcel HM, Troye-Blomberg M, Paulie S, Andersson G, MorenoC,
Pasvol G, Ivanyi J. Th1/Th2 profiles in tuberculosis, basedon the
proliferation and cytokine response of bloodlymphocytes to
mycobacterial antigens. Immunology1994;81:1716.
36. Harris DP, Vordermeier HM, Friscia G, Roman E, Surcel
HM,Pasvol G, Moreno C, Ivanyi J. Genetically permissiverecognition
of adjacent epitopes from the 19-kDa antigenof Mycobacterium
tuberculosis by human and murineT cells. J Immunol
1993;150(11):504150.
37. Belisle JT, Vissa VD, et al. Role of the major antigen
ofMycobacterium tuberculosis in cell wall biogenesis.
Science1997;276(5317):14202.
38. Silver RF, Wallis RS, Ellner JJ. Mapping of T cell
epitopesof the 30-kDa alpha antigen of Mycobacterium bovisBCG in
PPD-positive individuals. J Immunol 1994;154:4665.
39. Tan JS, Canaday DH, Boom WH, Balaji KN, Schwander SK,Rich
EA. Human alveolar T lymphocyte responses toMycobacterium
tuberculosis antigens: role for CD4+ andCD8+ cytotoxic T cells and
relative resistance of alveolarmacrophages to lysis. J Immunol
1997;159:2907.
40. Canaday DH, Ziebold C, Noss EH, Chervenak KA, Harding
CV,Boom WH. Activation of human CD8+ ab TCR+ cells byMycobacterium
tuberculosis via an alternate class IMHC antigen-processing
pathway. J Immunol 1999;162:3729.
41. Turner J, Dockrell H. Stimulation of human peripheral
bloodmononuclear cells with live Mycobacterium bovis BCGactivates
cytolytic CD8+ T cells in vitro. Immunology1996;87(3):33942.
42. Mohagheghpour N, Gammon D, et al. CTL response
toMycobacterium tuberculosis: identification of an immuno-genic
epitope in the 19-kDa lipoprotein. J Immunol1998;161(5):24006.
43. Lalvani A, Brookes R, et al. Human cytolytic and
interferongamma-secreting CD8+ T lymphocytes specific for
Mycobac-terium tuberculosis. Proc Natl Acad Sci USA
1998;95(1):2705.
44. Smith SM, Brookes R, et al. Human CD8+ CTL specific for
themycobacterial major secreted antigen 85A. J
Immunol2000;165(12):708895.
45. Geluk A, van Meijgaarden KE, et al. Identification of
majorepitopes of Mycobacterium tuberculosis AG85B that
arerecognized by HLA-A*0201-restricted CD8+ T cells in
HLA-transgenic mice and humans. J Immunol 2000;165(11):646371.
46. Flyer DC, Ramakrishna V, Miller C, et al., Identification
bymass spectrometry of CD8(+)-T-cell Mycobacterium tuber-culosis
epitopes within the Rv0341 gene product. InfectImmun
2002;70:292632.
47. Havlir DV, Ellner JJ, Chervenak KA, Boom WH.
Selectiveexpansion of human gamma delta T cells by
monocytesinfected with live Mycobacterium tuberculosis. J Clin
Invest1991;87(2):72933.
48. Dieli F, Troye-Blomberg M, Ivanyi J, Fournie JJ, Krensky
AM,Bonneville M, Peyrat MA, Caccamo N, Sireci G, Salerno
A.Granulysin-dependent killing of intracellular and extracel-lular
Mycobacterium tuberculosis by Vgamma9/Vdelta2 Tlymphocytes. J
Infect Dis 2001;184:10825.
49. Kabelitz D, Bender A, Schondelmaier S, Schoel B, KaufmannSH.
A large fraction of human peripheral blood gamma/delta+ T cells is
activated by Mycobacterium tuberculosisbut not by its 65-kD heat
shock protein. J Exp Med1990;171(3):66779.
50. Barnes PF, Grisso CL, Abrams JS, Band H, Rea TH, Modlin
RL.Gamma delta T lymphocytes in human tuberculosis. J InfectDis
1992;165(3):50612.
51. Hoft DF, Brown RM, et al. Bacille Calmette-Guerin
vaccina-tion enhances human gamma delta T cell responsiveness
tomycobacteria suggestive of a memory-like phenotype.J Immunol
1998;161(2):104554.
52. Li B, Rossman MD, et al. Disease-specific changes
ingammadelta T cell repertoire and function in patientswith
pulmonary tuberculosis. J Immunol 1996;157(9):42229.
Human immunity to M. tuberculosis 105
-
53. Shen Y, Zhou D, Qiu L, Lai X, Simon M, Shen L, Kou Z, WangQ,
Jiang L, Estep J, Hunt R, Clagett M, Sehgal PK, Li Y, ZengX, Morita
CT, Brenner MB, Letvin NL, Chen ZW. Adaptiveimmune response of
Vgamma2Vdelta2+ T cells duringmycobacterial infections. Science
2002;295:22558.
54. Rojas R, Boom WH. Gamma delta T cells and HIV-1
infection.In: Cellular aspects of HIV infection. Cytometric
cellularanalysis series, New York, NY: p. 147205. 2001.
Wiley-LissPress.
55. Constant P, Davodeau F, Peyrat MA, et al. Stimulation
ofhuman gamma delta T cells by nonpeptidic mycobacterialligands.
Science 1994;264(5156):26770.
56. Tanaka Y, Morita C, Tanaka Y, Nieves E, Brenner MB, BloomBM.
Natural and synthetic non-peptide antigens recognizedby human gamma
delta T cells. Nature 1995;375:1558.
57. Sicard H, Fournie JJ. Metabolic routes as targets
forimmunological discrimination of host and parasite. InfectImmun
2000;68(8):43757.
58. Morita CT, Beckman EM, Bukowski JF, et al.
Directpresentation of nonpeptide prenyl pyrophosphate antigensto
human gamma delta T cells. Immunity 1995;3(4):495507.
59. Lang F, Peyrat MA, Constant P, et al. Early activation
ofhuman V gamma 9V delta 2 Tcell broad cytotoxicity and
TNFproduction by nonpeptidic mycobacterial ligands. J
Immunol1995;154(11):598694.
60. Ulrichs T, Porcelli SA. CD1 proteins: targets of T
cellrecognition in innate and adaptive immunity. Rev Immuno-genet
2000;2:41632.
61. Sturgill-Koszycki S, Sclesinger P, Chakraborty P, et al.
Lack ofacidification in Mycobacterium phagosomes produced
byexclusion of the vesicular proton-ATPase.
Science1994;263:678.
62. Medzhitov R, Janeway Jr. CA. Innate immunity: the virtuesof
a nonclonal system of recognition. Cell 1997;91:2958.
63. Yoshimura A, Lien E, et al. Recognition of
Gram-positivebacterial cell wall components by the innate
immunesystem occurs via Toll-like receptor. J Immunol
1999;1633(1):15.
64. Takeuchi O, Hoshino K, Kawai T, Sanjo H, Takada H, Akira
S.Differential roles of TLR2 and TLR4 in recognition of
Gram-negative and Gram-positive bacterial cell wall
components.Immunity 1999;11:44351.
65. Means TK, Wang S, Lien E, Yoshimura A, Golenbock D, FentonM.
Human toll-like receptors mediate cellular activationby
Mycobacterium tuberculosis. J Immunol 1999;163:39207.
66. Brightbill HD, Libraty DH, Krutzik SR. Host Defense
Mechan-isms Triggered by Microbial Lipoproteins through the
Toll-like Receptors. Science 1999;285:7326.
67. Hirsch CS, Toossi Z, Johnson JL, et al. Augmentation
ofapoptosis and interferon-gamma production at sites ofactive
Mycobacterium tuberculosis infection in humantuberculosis. J Infect
Dis 2001;183:77988.
68. Hirsch CS, Toossi Z, Vanham G, et al. Apoptosis and T
cellhyporesponsiveness in pulmonary tuberculosis. J Infect
Dis1999;179:94553.
69. Ramachandra L, Noss EN, Boom WH, Harding CV. Processingof M.
tuberculosis antigen 85B involves intra-phagosomalformation of
peptide: MHC-II complexes and is inhibited bylive bacilli that
decrease phagosome maturation. J Exp Med2001;194:142132.
70. Gercken J, Pryjma J, et al. Defective antigen presentationby
Mycobacterium tuberculosis-infected monocytes. InfectImmun
1994;62(8):34728.
71. VanHeyningen TK, Collins HL, et al. IL-6 produced
bymacrophages infected with Mycobacterium species sup-presses T
cell responses. J Immunol 1997;158(1):3307.
72. Hmama Z, Gabathuler R, Jefferies WA, de Jong G, Reiner
NE.Attenuation of HLA-DR expression by mononuclear phago-cytes
infected withMycobacterium tuberculosis is related tointracellular
sequestration of immature class II heterodi-mers. J Immunol
1998;161:488293.
73. Hussain S, Zwilling BS, Lafuse WP. Mycobacterium
aviuminfection of mouse macrophages inhibits IFN-gamma
Januskinase-STAT signaling and gene induction by down-regulationof
the IFN-gamma receptor. J Immunol 1999;163:20418.
74. Ting LM, Kim AC, Cattamanchi A, Ernst JD.
Mycobacteriumtuberculosis inhibits IFN-g transcriptional
responseswithout inhibiting activation of STAT1. J Immunol
1999;163:3898906.
75. Wojciechowski W, DeSanctis J, et al. Attenuation of MHCclass
II expression in macrophages infected with Mycobac-terium bovis
bacillus Calmette-Guerin involves class IItransactivator and
depends on the Nramp1 gene. J Immunol1999;163(5):268896.
76. Noss EH, Sellati TJ, Radolf JD, Belisle J, Golenbock DT,
BoomWH, Harding CV. Toll-like receptor 2-dependent inhibition
ofmacrophage class II MHC expression and antigen processingby 19 kD
lipoprotein of M. tuberculosis. J Immunol2001;167:9108.
77. Gehring A, Rojas R, Canaday D, et al. Inhibition of
humanMHC-II antigen processing by 19 kD lipoprotein ofM.
tuberculosis. Submitted.
106 W.H. Boom et al.
Human immunity to M. tuberculosis: T cell subsets and antigen
processingNatural history of M.tuberculosis infectionThe central
role of CD4+ T cells in protective immunity to M.tuberculosisMHC
class I restricted CD8+ T cellsgammadelta T cells and other T cell
populationsT cell inhibition by M.tuberculosisModulation of
macrophage antigen-presenting cell function by
M.tuberculosisAcknowledgementsReferences