Embed Size (px)
Citation preview

1
Resursă educațională creată în cadrul programului Erasmus+ Parteneriate strategice în domeniul universitar prin proiectul:
”Online courses with videos for the field of veterinary communication dealing with prevention, diagnosis and treatment of diseases transferable from animals to humans”
Ref. nr. 2016-1-RO01-KA203-024732
MALARIA
Acest proiect a fost finanţat cu sprijinul Comisiei Europene.
Această publicaţie reflectă numai punctul de vedere al autorului şi Comisia nu este responsabilă pentru eventuala utilizare a informaţiilor pe care le conţine.

2
Autor: Liviu Miron
Coautori: Romania: Larisa Ivănescu, Dumitru Acatrinei, Olimpia Iacob, Lavinia Ciucă , Constantin Roman, Raluca Mîndru, Andrei Lupu, Andrei Cîmpan, Gabriela Martinescu, Elena Velescu, Mioara Calipsoana Matei, Doina Carmen Manciuc, Alina Manole, Doina Azoicai, Florentina-Manuela Miron, Gianina-Ana Massari, Anca Colibaba, Cintia Colibaba, Stefan Colibaba, Elza Gheorghiu, Andreea Ionel, Irina Gheorghiu, Carmen Antonita, Anais Colibaba Croația: Nenad Turk, Zoran Milas, Zeljana Kljecanin Franic
Lituania: Tomas Karalis, Rūta Karalienė, Virginija Jarulė, Leonora Norviliene , Donata Katinaite, Daiva Malinauskiene
Italia: Ilaria Pascucci, Ombretta Pediconi, Antonio Giordano

3
Copyright© 2016-019 Universitatea de Științe Agricole și Medicină Veterinară „Ion Ionescu de la Brad” (USAMV), Iasi (Romania). Toate drepturile rezervate
Universitatea de Științe Agricole și Medicină Veterinară „Ion Ionescu de la Brad” (USAMV), Iasi (Romania) este beneficiarul priectului Erasmus+ Online courses with videos for the field of veterinary communication dealing with prevention, diagnosis and treatment of diseases transferable from animals to humans (Ref. nr. 2016-1-RO01-KA203-024732) Nicio parte din acest volum nu poate fi copiată sau transmisă prin orice mijloace, electronice sau mecanice, inclusiv prin fotocopiere, fără permisiunea prealabilă scrisă a parteneriatului proiectului 2016-1-RO01-KA203-024732. MALARIA Online courses with videos for the field of veterinary communication dealing with prevention, diagnosis and treatment of diseases transferable from animals to humans 2016-1-RO01-KA203-024732 www.zoeproject.eu Erasmus+ Parteneriate strategice în domeniul universitar Parteneriat proiect:

4
CUPRINS
DEFINIȚIE ȘI ISTORIC / 5
ETIOLOGIE / 8
EPIDEMIOLOGIE / 16
PROFILAXIE / 58
TRATAMENT / 66
REFERINȚE BIBLIOGRAFICE / 69

5
DEFINITIE ȘI ISTORIC
Malaria este o boală cu o importanţă medicală foarte mare, produsă de protozoare parazite din genul Plasmodium, care sunt transmise la om prin mușcăturile femelei de țânțar Anopheles infectate, cu răspândire preponderent în regiunile tropicale şi subtropicale. Este maladia cu răspândirea cea mai largă de pe glob, sute de milioane de oameni fiind infectaţi anual în Africa, India, Asia de Sud-Est, Orientul Mijlociu, America Centrală şi de Sud, ceea ce face ca aproape 50% din populaţia globului să fie sub riscul infestării cu această boală.
ISTORIC De la sfârşitul pleistocenului, populaţiile de oameni migrând spre nord, au adus cu ei parazitul malariei; din epoca neolitică şi epoca bronzului rămăşiţile scheletice prezintă patologii de anemie cronică cauzată de infecţia cu Plasmodium falciparum. Prezenţa pe scară largă a talasemiei (anemia mediteraneeană) - boală genetică care conferă o anumită protecţie împotriva malariei, şi care indică o lungă perioadă din istorie pe parcursul căreia s-a intrat în contact cu agentul patogen al malariei, a fost semnalată încă din perioada Greciei antice şi a Romei antice. Introducerea agriculturii în jurul anului 7000 (Î.H.) a avut drept consecinţe creşterea stabilităţii populaţiilor şi ca urmare crearea unor condiţii favorabile pentru transmiterea malariei. Despăduririle au contribuit la menţinerea habitatutului necesar dezvoltării ţânţarilor Anopheles. În jurul anului 400, sunt semnalate o serie de schimbări ecologice, cu o treptată încălzire şi secetă în regiunea mediteraneană şi creşterea treptată a nivelului mării.
Prima menţiune literară despre existenţa unei febre de toamnă, este semnalată în Iliada lui Homer (800 sau 900 Î.H.), când Ahile se lupta cu Hector, iar regele Priam "vede un semn fatal înfipt pe ceruri care aduce o febră ucigătoare aupra oamenilor nenorociţi". Nu se poate şti cu exactitate dacă este vorba despre malarie, dar clar febra devastatoare este asociată cu perioada recoltei, care din punct de vedere ecologic este ideală pentru desfăşurarea ciclului biologic al ţânţarilor. Cert este că textele ulterioare confirmă că boala devine una semnificativă în Grecia antică. Hipocrate (460-377 Î.H) este primul care descrie

6
detaliat severitatea infecţiilor terţiene, asociate cu zonele umede, observând chiar splenomegalie (simptom în malaria cronică) ca manifestare întâlnită la persoanele care locuiau în zonele mlăştinoase. Medicii Praxagoras şi Heraclides au oferit descrieri similare asupra unei boli instalate în Grecia. Horace, Lucretius, Martial și Tacitus s-au numărat printre autorii latini care au menționat boala în Roma antică (Reiter, 2001). Secolul al doilea după Hristos vine cu o serie de scrieri ale lui Galenius şi Celsus care pe lângă simptome şi tratament, menţionează şi existenţa a trei specii de paraziți - P. falciparum, P. ovale și P. vivax frecvent prezente. În Evul mediu apar semnalări asupra unei perioade de răcire, astfel în anii 763-764 după Hristos, este descrisă prezenţa gheţii pe Marea Adriatică. Cu toate acestea armatele vizigoţilor, ostrogoţilor şi a altor barbari au fost nimicite de malarie şi mai mulţi papi au murit de malarie în călătoriile lor la Roma. Declinul malariei a început în a doua parte a secolului XIX. Danemarca a suferit epidemii devastatoare până în anii 1860, în special în mediul rural, apoi transmiterea s-a diminuat şi chiar a dispărut la începutul secolului. Situaţia a fost similară în Suedia, deşi cazuri izolate au fost raportate până în 1939, iar în Anglia a existat o scădere treptată până în 1880, după care a devenit rară, cu excepţia unei scurte perioade de după primul război mondial. Şi în Germania transmisia bolii s-a diminuat după anii 1880; ultima izbucnire având loc în Paris în 1865, în timpul construcţiei Grand Boulevards, în restul Franţei boala dispăruse, la fel ca şi în Elveţia. Declinul malariei în toate aceste ţări nu poate fi atribuit schimbărilor climatice pentru că a apărut în timpul unei faze de încălzire a temperaturilor. Alţi factori au care au fost implicaţi în declinul malariei includ: ameliorarea drenajului şi adoptarea de noi metode agricole care au ajutat la eliminarea habitatelor de ţânţari. Modificările demografice ca şi a condiţiilor de viaţă a oamenilor au avut un rol semnificativ, populaţiile scăzând în zona rurală pe măsură ce munca manuală a fost înlocuită de maşini, reducându-se astfel disponibilitatea oamenilor (faţă de animale) ca gazde pentru ţânţari şi a oamenilor ca gazde pentru parazit. Îmbunătăţirea serviciilor medicale şi utilizarea pe scară mai largă a chininei a redus rata de supravieţuire a parazitului malariei în gazda umană. În zona tropicală epidemiologia malariei este mult mai complexă decât în zona temperată. În Africa ecuatorială, în partea de nord a Indiei, Indonezia şi în partea de sud a Americii, transmiterea este stabilă, întrucât este destul de constantă de la an la an. Boala este endemică, dar epidemiile sunt mai puţin frecvente în zone ca India, Asia de Sud-Est şi America Centrală. Este numită instabilă întrucât transmisia variază mult de la un an la altul (Fig.1).

7
Fig.1. Zonele endemice pentru malarie (Sursa: Hay and Snow, 2006)
Malaria este endemic stabilă în regiunile în care anophelinele sunt antropofile şi au o rată ridicată de supravieţuire, temperatura şi umiditatea sunt în general ridicate şi există relativ puţină variaţie sezonieră. Boala este greu de controlat, deoarece transmisia este extrem de eficientă – majoritatea oamenilor infectându-se de mai multe ori pe an. Boala severă și decesul apar în general la copii şi la imigranţi. Temperatura joacă un rol important în transmiterea bolii, de ea depinzând incubaţia extrinsecă a parazitului în interiorul ţânţarului. Astfel vectorul Anopheles gambiae se găseşte atât la nivelul mării cât şi la 3000m deasupra mării dar malaria endemică dispare la peste 1.800 - 2.000 m (Reiter, 2001).

8
ETIOLOGIE, AGENT ETIOLOGIC, TAXONOMIE, DESCRIERE MORFOLOGICĂ ŞI CICLUL BIOLOGIC
ETIOLOGIE Malaria este o boală produsă de un protozoar din genul Plasmodium, ce infectază globulele roşii prin inocularea la om, de către femela ţânţar din genul Anopheles, în timpul hrănirii cu sânge (Fig. 2). Sunt cinci specii care parazitează la om şi pot fi transmise de la o persoană la alta: P. falciparum, P.vivax, P.ovale (cu două specii) și P.malariae. În ultimul timp au fost raportate din ce în ce mai des infecţii la om cu specia P. knowlesi care parazitează la maimuţă, în regiunile împădurite din Asia de Sud-Est şi în special în Insula Borneo.
Fig. 2. Infestarea intreritrocitară (Source: Boddey and Cowman, 2013)

9
TAXONOMIE Plasmodium este un protozoar ce aparţine familiei Plasmodiidae, Ordinul Haemosporidia și Încrengătura Apicomplexa care, alături de dinoflagellatele și ciliatele, formează Supraîncrengătura Alveolata, ce aparţine regnului Eukaryota. DESCRIERE MORFOLOGICĂ Parazitul malariei trece prin patru stadii de dezvoltare la om (schizonţi hepatici, apoi intraeritrocitar trofozoiţi, schizonţi şi gamonţi ) şi trei stadii de dezvoltare în interiorul vectorului ţânţar (oochinet, oochist şi sporozoit). Schizonţii se activează sub acțiunea anumitor factori, transformându-se în merozoiți, care pătrund în eritrocite, urmând următoarele faze:
• Faza de inel cu pecete: parazitul se găseşte sub forma unui inel, ocupând 1/3 din volumul total al eritrocitului. Protoplasma la capătul opus al nucleului este mai groasă și se colororează în albastru, iar nucleul în roșu. În mijloc apare o vacuolă incoloră plină cu lichid nuclear. Durata acestei faze este de aproximativ 12 ore la Plasmodium falciparum, variind în funcţie de specie .
• Faza de inel deformat: inelul se deformează în partea opusă nucleului. Se observă apariția unor granulații brune-închis, ce reprezintă pigment melanic rezultat din depunerea fierului din hemoglobină. Durata 12 ore.
• Faza de amibă de gradul I: se observă apariția unor pseudopode protoplasmatice în toate direcțiile, cu formă și mărime variate. Nucleul se mărește iar vacuola se micșorează sau dispare. Pigmentul melanic este abundent, în masa toată parazitului.
• Faza de amibă adultă de gradul II: parazitul se mărește ocupând tot eritrocitul, căpătând o formă mai regulată. Nucleul este mare. Pigmentul melanic abundent și răspândit în mod uniform. Durata ambelor stadii de amibă este de 16 ore.
• Faza de prerozetă: nucleul începe să se dividă, 2,3, 4 și 6 nuclei. • Faza de rozetă: nucleii sunt divizați la maximum (16-24). Uneori se observă și
diviziunea protoplasmei, ce înconjoară fiecare nucleu în parte, rezultând merozoiții cu diametru de 40-80μm. Aceștia se așează la periferia parazitului, pe mai multe planuri, dând rozetei aspectul de rozetă muriformă. Pigmentul melanic rămâne în centrul parazitului. Durata stadiului de prerozetă și rozetă este de 8 ore, întreg ciclul schizogonic durând 48 ore.

10
Formele sporogonice sunt reprezentate de gametociți. Într-un eritrocit hipertrofiat nedeformat, cu contur regulat, se observă hematozoarul de formă rotundă sau ovalară, cu protoplasma intens colorată în albastru, cu un nucleu mic și dens, colorat în roșu-purpuriu, fără vacuolă și cu granulații de pigment melanic foarte fine și răspândite uniform. Acesta este gametocitul femel sau macrogametocitul cu diametru de 7-14μm. Gametocitul mascul sau microgametocitul are protoplasma colorată în albastru-roșcat, cu un nucleu mare a cărui cromatină este difuză și cu pigment melanic de aspect grunjos, dispus în grămezi în jurul nucleului.În țânțar, microgameţii sunt lungi (15-25 μm în lungime) produse prin exflaglarea macrogameţilor fertilizaţi rotunzi pentru a forma oochineţii (15-20 x 2-5μm) care migrează prin peretele intestinal pentru a forma ovocite ovoide (până la 50μm în diametru ) pe suprafața exterioară. Oochistul produce mii de sporozoizi subțiri alungiți (~ 15 μm lungime), care în final infectează glandele salivare. CICLUL BIOLOGIC Ciclul biologic al malariei este unul din cele mai fascinante și complicate dintre toate organismele, constituind un interes deosebit mai ales pentru biologia moleculară, biologia celulară și imunologie. Transmiterea vectorială a malariei prin țânțarii din genul Anopheles a fost stabilită la sfârșitul secolului trecut de Grassi. Țânțarii implicați în transmiterea malariei sunt: A. hyrcanus, A. labranchiae, A. maculipennis maculipennis, A. maculipennis atroparvus, A. maculipennis messae, A. plumbeus, A. claviger, A. sacharovi. La noi în țară au fost semnalate speciile A. maculipennis, A. messae, A. atroparvus, A. sacharovi şi Anopheles labranchiae. Malaria la om, presupune o etapă asimptomatică în ficat și una simptomatică eritrocitară. Toate cercetările pentru descoperirea medicamentelor s-au concentrat pe etapa eritrocitară, care are un potențial limitat pentru profilaxia și blocarea transmiterii. O nouă tehnică poate fi folosită pentru studiul medicamentelor privind eficacitatea asupra stadiului hepatic, prin folosirea paraziților ce exprimă luciferază, prin bioluminescență. Majoritatea studiilor s-au realizat pe acțiunea medicamentelor asupra stadiilor eritrocitare, cunoscându-se puține medicamente ce distrug fazele hepatice. Cel mai cunoscut și eficient rămâne primaquine, singurul utilizat și pentru tratamentul formelor hipnozoite de Plasmodium vivax, care pot provoca recăderi după luni, chiar ani de la infecție. Problema este că primaquina produce anemie hemolitică la indivizii cu deficit de glucozo-6-fosfat (G6PD), acest deficit de enzimă fiind catacteristică mai ales în regiunile afectate de malarie din Africa, America de Sud și Asia (Adoubryn et al, 2004; Derbyshire, 2012).

11
CICLUL SPOROGONIC AL PARAZITULUI MALARIEI Prin schimbarea habitatului se produc pierderi mari pentru parazitul malariei, ce provoacă un dezechilibru în densitatea populației. Acest dezechilibru este compensat prin creșterea și reproducerea în nișe celulare ferite de răspunsurile imune ale gazdei (Frevert, 2004). În stadiul de ookinet și sporozoit, parazitul suferă pierderile cele mai mari, aceste etape desfășurându-se în vectorul țânțar (Silvie et al, 2008). Ookinetul se dezvoltă din zigot, în lumenul intestinului mijlociu al gazdei, fiind rezultatul fertilizării dintre un macrogametocit femel și un microgametocit mascul. Oochinetul traversează stratul de celule epiteliale ale intestinului pe partea apicală, ajungând în cele din urmă în lamina bazală. Ca răspuns la invazie, organismul își activează mecanismele de protecție, ceea ce duce la o scădere a densității populației parazitare. Oochineții se transformă în oochiști, aceasta fiind o etapă de dezvoltare extracelulară, în urma căreia se formarează sporozoiții ce sunt eliberați în cavitatea corpului țânțarului, invadând apoi glandele salivare (Al-Olayan EM et al, 2002; Vlachou D et al, 2006) (Fig.3). Pe parcursul acestei migrări are loc o scădere a densității populației parazitare, sporozoiții inoculați gazdei mamifer ajungând într-un timp foarte scurt în ficat. În 1948 Shortt şi Garnham au descoperit că sporozoiții infectează hepatocitele, dezvoltând aici o etapă înainte de a infecta eritrocitele. Etapa hepatică este asimptomatică, fiind de interes științific urmărindu-se producerea de vaccinuri, acum având loc și amplificarea de până la 10.000 de ori a numărului de paraziți culminând cu eliberarea de merozoiți exoeritrocitari. Merozoiții vor invada eritrocitele, inițiind astfel ciclul eritrocitar care este patogen (Vaughan et al, 2008). Astăzi este cunoscută expresia genelor și proteinele implicate în evoluția stadiilor din țânțar și faza pre-eritrocitară (Greenwood et al, 2005; Kappe et al, 2001; Lasonder, 2008). Descoperirea genei responsabilă de etapa hepatică a dus la formarea de sporozoiți atenuați, obținându-se un vaccin viu atenuat. Parazitul suferă reproducere sexuală o singură dată, în corpul țânțarului. Factorii care inițiază formarea gametociților nu sunt foarte bine cunoscuți. Ingestia de gametociți în timpul mesei de sânge, activează formarea de gameți (gametogeneză) în lumenul intestinal al țânțarului.

12
Fig. 3. Ciclul biologic al malariei (Sursa: Colecţie personală) O moleculă, derivată a acidului xanthurenic, împreună cu modificarea temperaturii și a pH-ului, pot declanșa gametogeneza de sex masculin sub formă de exflagelare (Billker et al, 1997; Billker et al, 1998). Proteina P48\45 este esențială în abilitatea gametului mascul de a fertiliza gameții de sex feminin (care are exprimată pe suprafață proteina P47). Fertilizarea are loc pentru a forma zigotul, care în urma meiozei se transformă în oochinet, care traversează mai multe celule epiteliale ale intestinului (Mueller et al, 2005). Oochineții traversează celulele epiteliale până în lamina bazală și se transformă în oochisturi, care au o creștere extensivă și în termen de 10-14 zile rezultă în producerea a mii de sporozoiți. Sporozoiții rezultați din oochisturi, ajung după o migrare prin hemolimfă, să se atașeze de partea bazală a celulelor acinare a glandei salivare. Sporozoiții traversează celulele acinare și pătrund în ductele glandelor salivare, de unde sunt transmiși la gazda mamifer în timpul reprizei de sânge (Pimenta et al, 1994; Robert and Boudin, 2003).
CICLUL SCHIZOGONIC ERITROCITAR AL PARAZITULUI MALARIE
Gazda mamifer este infectată cu sporozoiți din glandele salivare ale țânțarilor. Aproximativ 100 de sporozoiți sunt injectați de un singur țânțar (Medica and Sinnis, 2005). Aceștia rămân în derm o perioadă de timp, au o mișcare aparent aleatorie, până găsesc un vas de sânge, apoi migrează în ficat (Mota et al, 2001; Amino, 2007). După injectare

13
sporozoiți rămân în derm 1-3 ore apoi migrează spre ficat (Yamauchi, 2007). Nu toți sporozoiții ajung în circulație și ficat, o parte rămân cantonați la locul de injectare de unde probabil sunt eliminați prin fagocitoză. O parte ajung în sistemul limfatic, în ganglionii limfatici, unde este activat răspunsul celulelor T și CD8, care sunt capabili să elimine paraziții în curs de dezvoltare din ficat (Chakravarty et al, 2007; Amino et al, 2008). În piele sporozoiții migrează activ prin celule, ceea ce implică perturbarea membranei plasmatice a celulei gazdă, pentru traversare celulară fiind implicate cel puțin trei proteine SPECT-1, SPECT-2 și fosfolipază. Sporozoiții lipsiți de SPECT-1, SPECT-2 sunt imobilizați în derm ca urmare a afectării capacității de traversare a celulelor (Amino et al, 2006; Sturm et al, 2006; Ménard, 2007; Sturm et al, 2006; Aly et al, 2009). După inocularea sporozoiților de vectorul Anopheles, începe ciclul exoeritrocitar care durează în funcție de specie astfel: la Plasmodium falciparum – 6 zile; Plasmodium vivax – 8 zile; Plasmodium ovale – 9 zile; Plasmodium malariae – 15 zile. Unii sporozoiți ai Plasmodium vivax și Plasmodium ovale, pătrunși în hepatocite, rămân în stare latentă o lungă perioadă timp de câțiva ani si au fost numiţi hipnozoiți (Cogswell, 1992).
Fig.4. Ciclul de replicare asexuată a parazitului în interiorul celulelor roșii din sânge
(Sursa: Silvie et al, 2008) (1) Merozoiți extracelulari se atașează aleatoriu la suprafața eritrocitelor. (2) Apoi are loc orientarea capătului apical al merozoiților spre suprafața eritrocitelor. (3) Formarea de joncțiune Tight și merozoiții activi intră în eritrocite. (4) Invazia merozoiților apare prin formarea simultană a unei vacuole parazitofore. După finalizarea invaziei, merozoiții se transformă în inel. (5) Această etapă se caracterizează printr-o vacuolă digestivă mare, unde digestia hemoglobinei rezultă în formarea de pigment malaric, hemozoin. (6) Parazitul crește intraeritrocitar şi este numit trofozoit. Pigmentul hemozoin se acumulează în vacuola digestivă. (7) replicarea ADN-ului precede celula în devenire, process numit schizogonie. (8) merozoiții secretă exonema ce inițiază ieșirea din vacuola parazitoforă şi hematiile gazdei. (9) merozoiții ieșiți aderă la un eritrocit adiacent în câteva secunde. (1) inițierea unui nou ciclu eritrocitar (Fig. 4, 5).

14
Fig. 5. Variate stagii eritrocitare (1-2 - forma de inel; 3 - schizont matur; 4 - gametociţi) la Plasmodium vivax, coloraţia Giemsa, obiectiv x 1000 (Sursa: Colecţie personală) Formele sporogonice: gametociții (Fig. 6). Într-un eritrocit hipertrofiat nedeformat, cu contur regulat, se observă hematozoarul de formă rotundă sau ovalară, cu protoplasma intens colorată în albastru, cu un nucleu mic și dens, colorat în roșu-purpuriu, fără vacuolă și cu granulații de pigment melanic foarte fine și răspândite uniform. Acesta este gametocitul femel sau macrogametocitul. Gametocitul mascul sau microgametocitul are protoplasma colorată în albastru-roșcat, cu un nucleu mare a cărui cromatină este difuză și cu pigment melanic de aspect grunjos, dispus în grămezi în jurul nucleului (Field and Shute, 1956; Gratzer and Dluzewski, 1993; Moore et al, 2008). Inoculat la om parazitul suferă o serie de transformări, mai întâi în ficat (acesta constituie ciclul exoeritrocitar sau tisular), apoi pătrunde în globulele roșii unde se dezvoltă până duce la distrugerea globulelor roșii (ciclul eritrocitar sau asexuat). Parazitul eliberat din globulele distruse, parazitează din nou alte globule Tolle, 2009; Sturm et al, 2006) (Fig. 7).
a b Fig. 6. a. Schizonţi maturi şi merozoiţi Col. Giemsa x100, obiectiv de imersie, (Sursa: Colecţie personală) b. Stadii gametocitare (macrogametocit roz, microgametocit albastru) Col. Giemsa x100 (Sursa: Colecţie personală)

15
Țânțarul inoculează parazitul în forma de sporozoiți (formele infectante pentru om), care ajunși în ficat formează o masă plasmodială mare din care se desprind merozoiții ce ajung în sânge și pătrund în globulele roșii. În interiorul globulului roșu parazitul trece prin stadiul de amibă, apoi de rozetă, apoi distruge globulul roșu eliberând merozoiții, care vor parazita alte hematii în care se reface ciclul (Hafalla et al, 2011). Din ciclul eritrocitar mai rezultă gametociți (masculi, femeli) care ajungând în corpul țânțarului se unesc formând zigoți. În lumenul intestinal zigoții se transformă în oochineți din care se produc după o serie de transformări sporozoiții, localizați în glandele salivare de unde sunt inoculați la om (Vlachou, 2006).
Fig.7. Ciclul biologic la Plasmodium-vivax (Sursa: http://www.microbiologynotes.com/life-cycle-of-plasmodium-vivax/)
Femelele Anopheles care înțeapă un om bolnav, în al cărui sânge se găsesc gametociți și schizonți, aspiră odată cu sângele și un număr variabil de paraziți sub aceste două forme. Schizonții sunt digerați în stomacul țânțarilor, iar gametociții asigură continuarea ciclului sexuat sau sporogonic, din care rezultă sporozoiții. S-a demonstrat că unii țânțari conțin două gene care codifică carboxipeptidaza de la nivelul intestinului mijlociu, care sunt implicate în dezvoltarea parazitului la Anopheles gambiae (Lavazec and Bourgouin, 2008; Aly et al, 2009).

16
EPIDEMIOLOGIE
MODUL DE TRANSMITERE A MALARIEI Prin vectori - vectorul transmițător la om și în același timp gazdă obligatorie pentru stadiul sexuat al parazitului îl reprezintă țânțarii din genul Anopheles ( din aproximativ 400 de specii de pe glob, doar 30-40 pot fi în condiții naturale vectori pentru malarie). Perioada de formare a sporozoiților în glandele salivare variază între 7 și 30 de zile în funcție de specia parazitului și temperatura mediului. Prin transfuzii de sânge- malaria post-transfuzională cunoscută din 1911, se face de la purtătorii latenți, asimptomatici ai malariei. În cele mai multe cazuri transmiterea post-transfuzională s-a făcut cu P. malariae (care dă forme persistente cu purtători sanguini de durată) și mai rar cu P. vivax și P. falciparum. Perioada de contagiozitate a omului este perioada de timp cât gametociții infestanți ai parazitului malaric persistă în sângele bolnavilor şi variază cu specia de Plasmodium. Astfel gametociții de P. malariae pot persista în sânge chiar până la 53 de ani, P. vivax poate persista 1-3 ani, iar Pfalciparum până la 1 an. Femela de Anopheles rămâne infestantă toată durata vieții. Sunt posibile trei căi de transmitere a malariei de la bolnav sau purtătorul de plasmodii la persoanele sănătoase: în condiţii naturale prin înţepăturile ţânţarilor din genul Anopheles; intrauterin de la mama bolnavă la făt prin placentă sau în timpul naşterii; prin transfuzii - prin intermediul sângelui care conţine agentul patogen în timpul manipulării medicale sau efectuând injecţii în condiții aseptice cu ace contaminate. Transmiterea malariei se face în sezonul de activitate al țânțarilor, depinzând de intensitatea și durata transmiterii, precum şi de influența factorilor naturali și sociali cu efect pe termen lung.

17
VECTORII MALARIEI În majoritatea cazurilor, malaria este transmisă prin mușcătura femelei țânțar din genul Anopheles. Există peste 400 de specii diferite de țânțari ce aparţin genului Anopheles; în jur de 30 reprezintă vectori pentru malarie de o importanță majoră (Fig. 8). Intensitatea transmiterii depinde de factorii legați de parazit, vector, gazda umană și mediu. Fiecare specie de țânțar Anopheles are propriul habitat acvatic; de exemplu, unii preferă colecții mici de apă dulce, cum ar fi bălți și apa acumulată în urmele de copite, care sunt abundente în timpul sezonului ploios în țările tropicale. Înțelegerea biologiei și comportamentului țânțarilor Anopheles poate ajuta la înțelegerea modului în care este transmisă malaria și poate ajuta la elaborarea strategiilor de control adecvate. Factorii care afectează capacitatea unui țânțar de a transmite malarie includ susceptibilitatea sa înnăscută față de Plasmodium, alegerea gazdei și longevitatea sa. Factorii care ar trebui luați în considerare la proiectarea unui program de control includ susceptibilitatea vectorilor malarie la insecticide și locul preferat de hrănire și odihnă a țânțarilor adulți. Transmiterea este mai intensă în locurile în care durata de viață a țânțarilor este mai lungă (astfel parazitul are timp să-și finalizeze dezvoltarea în interiorul țânțarilor) și preferă hrănirea pe oameni mai degrabă decât pe animale. Durata lungă de viață și caracterul antrophofil a speciilor africane de ţânţari reprezintă principalul motiv pentru care aproape 90% din cazurile de malarie din lume se află în Africa. Transmiterea depinde și de condițiile climatice care pot afecta numărul și supraviețuirea țânțarilor, cum ar fi precipitațiile, temperatura și umiditatea. În multe locuri, transmiterea este sezonieră, cu vârf în timpul și imediat după sezonul ploios. Epidemiile de malarie pot apărea atunci când clima și alte condiții favorizează brusc transmiterea în zonele în care oamenii au imunitate mică sau deloc la malarie. Acestea pot apărea, de asemenea, atunci când persoanele cu imunitate scăzută se deplasează în zone cu transmitere intensă a malariei, de exemplu pentru a găsi un loc de muncă sau ca refugiați.

18
Fig. 8 Distribuția globală a vectorilor dominanți sau potențial importanți ai malariei (Proiecția Robinson) (Sursa: Kiszewksi et al, 2004)
Ţânţarii se găsesc răspândiţi în întreaga lume cu excepţia zonei arctice, şi din aproximativ 3500 de specii existente, trei sferturi provin din zonele tropicale şi subtropicale. Importanţa medicală a acestor insecte o reprezintă nevoia de a se hrăni cu sânge pentru a obţine proteina necesară maturării ouălor. Hrănirea se realizează cu ajutorul unei secreţii salivare complexe care facilitează transmiterea unor agenţi patogeni gazdei mamifer în timpul prelevării de sânge. Ecologia, comportamentul, supravieţuirea ţânţarilor şi dinamica transmiterii bolilor, sunt puternic influenţate de factorii climatici: temperatură, precipitaţii, umiditate, vânt, durata luminii zilei. De asemenea timpul de dezvoltare a agentului patogen în glandele salivare ale ţânţarilor este direct influenţat de temperatură (Reiter, 2001). Malaria este transmisă prin mușcătura țânțarilor femele din genul Anophleles. În Europa, țânțarii din complexul Anophleles maculipennis au fost incriminați ca vectori ai bolii. Prima persoană care a remarcat încă din 1926 existența unui complex de specii de maculipeni a fost Falleroni, care a descris diferențele morfologice privind corionul acestor specii. Există aproximativ 430 de specii înregistrate ca aparținând genului Anophleles, din care aproximativ 60 de specii sunt considerate vectori ai malariei (Reiter, 2001). Complexele de specii sunt grupuri de specii strâns legate între ele, care sunt foarte greu de diferențiat din punct de vedere morphologic. Anophles maculipennis Meigen, vectorul istoric al malariei în Europa și Orientul Mijlociu, a fost primul complex cu specii gemene, descoperit la țânțari (Falleroni, 1926; van Thiel, 1927). Căutarea a inclus studii de morfologie după ou (Falleroni, 1926; Falleroni, 1932; Corradetti, 1934; de Buck and

19
Swellengrebel, 1934; Hackett and Lewis, 1935; Korvenkontio et al, 1979), după cromozomi (Frizzi, 1952; Frizzi, 1953; Kitzmille, 1967; Stegnii, 1976; Stegnii and Kabanova, 1976), secvențe /DNJ (Marinucci et al, 1999; Proft et al, 1999; Linton et al, 2001; Linton et al, 2002; Linton et al, 2003; Sedaghat, 2003; Porter, 1991), ecologie (van Thiel, 1927; de Buck and Swellengrebel, 1934; Hackett and Missiroli, 1935), hibridizare (de Buck and Swellengrebel, 1934; Kitzmiller et al, 1967), larvă (Bates, 1939; Buonomini, 1940; Suzzoni-Blatger and Sevin, 1981; Boccolini et al, 1986; Suzzoni-Blatger et al, 1990; Romi, 2000), caracteristicile aripei (de Buck et al, 1933; Ungureanu and Shute, 1947; Sedaghat et al, 2003). După toate aceste studii s-a stabilit prezența a opt specii în cadrul complexului palearctic maculipennis: A. atroparvus van Thiel, A. beklemishevi Stegnii and Kabanova, A. labranchiae Falleroni, A. maculipennis, A. martinus Hingarev, A. melanoon Hackett, A. messeae Falleroniand, A. sacharovi Favre (White, 1978; de Zulueta et al, 1983; Cianchi et al, 1987; Ribeiro et al, 1988; Linton et al, 2002; Sedaghat et al, 2003; Knight and Stone, 1977; Kettle, 1995) (Fig. 9).
Fig.9. Speciile complexului Palearctic maculipennis (Sursa: https://ecdc.europa.eu/en/disease-vectors/facts/mosquito-factsheets/anopheles-sacharovi; http://mosquito-taxonomic-inventory.info/file/5212)
Toți membrii acestui complex sunt considerați vectori ai malariei în Europa. Având în vedere importanța medicală a complexului Maculipennis, diferențele privind capacitatea vectorială și distribuția speciilor, care sunt considerate a fi influențate la nivel mondial de încălzirea globală, devine necesară folosirea unor metode sigure de identificare a speciilor.
A. atroparvus
(van Thiel,1927) A. beklemishevi
(Stegnii and Kabanova, 1976) A. labranchiae
(Falleroni,1926) A. maculipennis (Meigen, 1818)
A. messae (Falleroni, 1926)
A. martinius (Shingarev,1926)
A.sacharovi (Favre, 1903)
Anopheles melanoon Hackett, 1934

20
Odată infectată cu Plasmodium, femela Anopheles va avea toată viața sporozoiți în glandele salivare, infectând o nouă gazdă la fiecare masă de sânge luată. Capacitatea vectorială este un indice în capacitatea de transmitere a vectorului malariei, fiind influențat acest indice de densitatea vectorului (infuențată de condițiile climatice: temperatură și umiditate; de condițiile geografice: variațiile sezoniere; și de preferința zoofilică sau antropofilică (Mullen and Durden, 2002; Marquardt, 2004; Toole, 2009). În lipsa gazdei preferențiale comportamentul de hrănire se poate modifica (Boete, 2006; Pages et al, 2007; Nitzulescu and Gherman, 1990). A. atroparvus, A. labranchiae și A. sacharovi au fost incriminați ca principali vectori ai malariei în regiunea Europei (Vicente et al, 2011). Durata ciclului gonotrofic (distanța dintre ponte) este de aproximativ 2 zile, iar durata evoluției sporozoiților este de aproximativ 12 zile, este nevoie de cel puțin 6 cicluri gonotrofice înainte ca femela să devină infectantă. Durata etapei sporogonice variază în funcție de specia de Plasmodium, astfel pentru P. falciparum între ingestia de gametociți și prezența de sporozoiți în glandele salivare, sunt necesare 12 zile la 25°C, 23zile la 20°C. Sub 18°C și peste 33°C ciclul încetează (Bruce-Chwatt and Zulueta, 1980). Distribuția globală a malariei depinde foarte mult de caracteristicile intrinseci ale vectorului, competența vectorului (capacitatea unei specii de Anopheles de a asigura dezvoltarea completă a parazitului) și capacitatea sa vectorială (Pratt and Moore, 1993). Incapacitatea parazitului malariei să se dezvolte în unele specii de țânțari ține de lipsa unor factori metabolici esențiali pentru dezvoltarea parazitului sau prezența unor toxine care inhibă dezvoltarea parazitului. Prezența vectorilor depinde în mare măsură de condițiile locale (Mouchet et al, 1999). Schimbările climatice cu creșterea temperaturii chiar cu 0,5°C pot duce la o creștere a populației de țânțari cu 30-100% (Patz and Olson, 2006). Acest fapt susține necesitatea raportării permanente a speciilor de Anopheles prezente și densitatea populațiior în vederea posibilității cuantificării măsurilor de control a malariei. În zonele muntoase din Africa, incidența malariei a crescut din 1970, corelându-se cu modificarea factorilor climatici, înregistrându-se o creștere a temperaturii (Pascual et al, 2006). Este importantă identificarea moleculară a speciilor complexului Maculipennis având în vedere importanța medicală, capacitatea vectorială și distribuția speciilor care se pare că este direct influențată de încălzirea globală (Sedaghat et al, 2003). Anopheles claviger are o distribuţie mai largă, de la zonele subcarpatice până în plină câmpie. Cuiburile larvare preferate se întâlnesc în dumbrăvi, livezi, în locuri mai ferite de insolaţie şi cu temperatură mai moderată. Larvele lui trăiesc şi în plină iarnă, sub gheaţă. Este un ţânţar destul de agresiv pentru om, atacându-l în păduri chiar în plină zi.

21
Anopheles hyrcanus este o specie foarte răspândită în Delta Dunării, dar şi în interiorul ţării, în colecţiile de apă salmastră. Centrul său de răspândire este în Extremul Orient, unde este vector pentru malarie, dar spre vest îşi pierde această capacitate.
Anopheles maculipennis, Anopheles messeae şi Anopheles sacharovi fac parte din grupul Maculipennis, care cuprinde anofeli cu aripi pătate, petele întunecate provenind din aglomerarea solzilor pe anumite zone de pe nervurile aripilor. Speciile din grupul Maculipennis se caracterizează printr-o plasticitate ridicată a larvelor, care se adaptează la cuiburi larvare destul de variate. Criteriul cel mai sigur pentru diferenţierea celor trei specii de anofeli din grupul Maculipennis, este oul. La fiecare specie, partea ventrală a oului, îndreptată îm sus în timpul plutirii pe apă, este ornamentată caracteristic. Anopheles maculipennis este o specie răspândită în toată țara, în apropierea pădurilor și pe văile râurilor. Apele dulci bine oxigenate reprezintă locul pentru depunerea pontei, cu o temperatura medie mai scăzută, ferite de lumina directă a soarelui. Femela este zoofilă şi ocazional antropofilă. În timpul iernii hibernarea este completă, cu concordanţă gonotrofică, în adăposturi ferite de oameni, în grajduri deschise, scorburi expuse frigului.
Anopheles messeae este răspândit în toate câmpiile joase din ţară şi în locurile cu ape stagnante (iazuri, lacuri, eleştee, treimile inferioare ale râurilor care de obicei au zone inundabile, zona inundabilă a Dunării). Cuiburile larvare sunt reprezentate de ape dulci, stagnante, întinse, cu fund puţin adânc, cu vegetaţie abundentă, puternic încălzite, bine însorite. Larvele se dezvoltă bine mai ales spre malul acestor ape şi la periferia insulelor de vegetaţie. Femela este predominant zoofilă dar se hrăneşte şi pe om. Ea are un somn hibernant complet, cu concordanţă gonotrofică, în adăposturi destul de expuse la frig .
Anopheles atroparvus este răspândit în toată ţara, în zonele cu ape mai mult sau mai puţin sărate. În acest sens larvele au o mare plasticitate de adaptare, de la cuiburi larvare cu apă dulce până la cele cu apă sărată ( 4-5%). Ele sunt destul de puţin sensibile faţă de temperaturi mai scăzute (Ungureanu and Shute, 1947). Femela iernează în adăposturi intradomestice şi peridomestice, ferite de frig, unde poate găsi hrană. Ea nu hibernează complet şi se trezeşte din când în când şi înţeapă omul sau vitele, dar este predominant antropofilă. Femela de Anopheles în general se împerechează o singură dată, și păstrează sperma în canalul spermatecilor de unde este folosită toată viața femelei pentru fertilizarea ouălor.

22
Pentru maturarea ouălor este necesară o priză de sânge, femela putând călători până la 3 km în căutarea gazdei, fiind atrasă de dioxidul de carbon emanat de aceasta (Smallegange RC et al, 2005). Dezvoltarea gemetociților (forma infectantă pentru om) se pare că este mult mai prielnică în interiorul speciei Anopheles gambiae, care este vectorul principal al malariei în Africa (Lacroix R et al, 2005). După împerechere sunt necesare 48h pentru maturarea ouălor, apoi ele sunt depuse pe apă, a cărei proprietăți diferă în funcție de specia de Anopheles (dimensiunea, expunerea la soare, colecție de apă naturală sau artificială, nepoluată). Femelele adulte depun 50-200 de ouă pe ovipoziție. Ouăle sunt așezate direct pe apă și pot pluti pe ambele părți. Ouăle nu sunt rezistente la uscare și ecloziune în decurs de 2-3 zile, deși eșantionarea poate dura până la 2-3 săptămâni în condiții mai scăzute de climă. Aceste caracteristici sunt pentru speciile de Anopheles din mediul rural și la periferia celui urban, riscul de transmitere a malariei fiind mult mai crescut în mediul rural (Pages F et al, 2007). Preferința pentru hrănire în interior (endofage) sau exterior (exofage), pentru mediul ambiental endofile sau exofile, variază de la o specie la alta, sau în cadrul aceleeași specii în funcție de zona geografică, caracteristici ce au un rol important în dezvoltarea unor strategii de luptă împotriva anofelilor (Pages F et al, 2007). Femelele se hrănesc din amurg până în zori, vârful de atac diferă cu specia, A. cruzii, A. bellator din America de Sud fiind diurne. Zborul speciilor din genul Anopheles este tăcut iar mușcătura este nedureroasă. Genul Anopheles cuprinde specii cu importanţă medicală deosebită, fiind reprezentat prin 500 de specii, dintre care 50 sunt vectori pentru malarie (Harbach, 1994). Adulţii se caracterizează din punct de vedere morfologic prin palpi la fel de lungi ca trompa la ambele sexe, scutel simplu şi abdomen lipsit de solzi. Hipopigiul este alcătuit din forcipule fără lobi, claspetele sunt prezente, paraproctele nu se chitinizează, iar falozomul este ornamentat adesea cu excrescenţe apicale. Larvele sunt lipsite de sifon. Ouăle sunt depuse izolat pe suprafaţa apei şi au o pereche de flotori laterali, mai dezvoltaţi la speciile de apă dulce şi mai mici la cele de apă sărată. Ciclul de dezvoltare de la ou la adult durează între 8 zile la 31°C și 20 de zile (Foster and Walker, 2009). Perioada dezvoltării agentului malariei în interiorul ţânţarului (incubaţie extrinsecă) variază între 10 şi 21 de zile în funcţie de specia de Plasmodium. Durata vieţii unui ţânţar în natură nu poate fi calculată exact, estimându-se că peste 20% din ţânţari ar supravieţui mai mult decât o perioadă de incubaţie extrinsecă de 14 zile. Astfel, se poate conluziona că nu densitatea populaţiei de ţânţari are importanţă, ci longivitatea acestora, scopul fiind scurtarea vieţii ţânţarilor cu ajutorul insecticidelor.

23
ANOPHELES MACULIPENNIS FEMELELE Culoarea este închisă sau maro mediu, deși varietatea culorii și a dimensiunii este mare. Caracteristica care diferențiază toate celelalte specii europene de Anopheles, sunt aripile cu solzi întunecați ce formează pete caracteristice. Trompa este de culoare maro închis iar palpii maxilari sunt de aceeași culoare și la fel de lungi ca și trompa. Vertexul are un smoc lung albicios îndreptat anterior. Scutumul are o bandă gri care se îngustează anterior. Părțile laterale ale scutumului sunt maro anterior și maro negricios în partea posterioară. Scutelul este de culoare maro cu solzi înguști aurii. Postnotul este maro deschis iar pleurele sunt maro închis. Femurul este maro închis pe partea dorsală și brun deschis pe partea ventrală, tibia este maro și mai deschisă la vârfuri, tarsul este maro închis. Aripile au solzi întunecați, distribuiți neuniform care formează pete negre aproape de RS și R2 3și M. La A. sacharovi se observă pete întunecate pe nervurile aripei și pe marginea posterioară. Abdomenul este maro sau brun negricios cu o dungă maro aurie (Capinera, 2002).
MASCULII La speciile de culicide din genul Anopheles, cel puţin cele prezente în ţara noastră, coxitul este lipsit de lobi. Pe el se inseră doi ( Anopheles plumbeus, A. hyrcanus şi grupul maculipennis) sau trei (A. claviger) spini coxali parabazali mari şi un spin coxal intern, a căror dispoziţie şi ramificare depinde de specie. Forma lobilor de pe coxite, ca şi ornamentarea cu spini şi peri variază cu specia. Baza coxitului se prelungeşte lateral prin trei apofize sau apodeme. Apodema externă şi cea postero-internă servesc la prinderea de segmentul 9. Apodema bazală sau placa bazală, pe care se inseră muşchii penieni şi anali, pătrunde în segmentele 9 şi 8, iar pe de altă parte se articulează cu paramerele segmentului 10.
LARVELE Foarte variabile în pigmentare și dimensiune, larvele din părțile nordice ale Europei, sunt de obicei mai mari, cu o capsulă a capului închisă, antena este aproape dreaptă, slab spiculată, aproape 2/3 din mărimea capului. Acul frontal (5-C la 7-C) este lung și plumos. Acul palmat pe segmentele abdominale I și II sunt rudimentare, dar bine dezvoltate pe segmentele III-VII. La anofeli există peri care se ramifică penat ca şi pe cap şi torace, spre deosebire de culicini, la care perii se ramifică de la bază. La speciile de Anopheles se mai pot observa câteva structuri caracteristice pe fiecare segment abdominal:

24
• plaga tergală chitinoasă, impară a cărei mărime variază cu specia; • plăcile accesorii, posterioare faţă de cea mediană, câte una, două sau trei pe fiecare
segment; • părul palmat, pe fiecare segment, reprezentând un sistem tensio-activ de plutire.
Aceşti peri pot fi folosiţi în diferenţierea speciilor din grupul Maculipennis şi a speciei Anopheles hyrcanus.
Al 8-lea segment abdominal, cu aceeaşi formă ca şi celelalte 7, constituie suportul unor structuri cu importanţă taxonomică. Pe pereţii laterali ai segmentului 8 se află pieptenii abdominali sau periile abdominale, câte unul pe fiecare parte. La anofeli sifonul lipseşte şi spiraculele (orificiile stigmatice) sunt menţinute de o farfurie spiraculară (Cosoroaba, 1992; Cosoroaba, 2000). ANOPHELES LABRANCHIAE (FELLERONI 1926) Larvele se dezvoltă în apă stagnantă,dulce sau sărată, în zonele de coastă din Europa, preferând căldura. Hibernarea se face ca femelă adultă la întuneric, în grajduri sau cavități naturale. Diapauza poate fi compltetă sau incompletă, existând prânzuri de sânge ocazionale. Femelele sunt în principal antropofage, ocazional hrănindu-se și pe animale domestice. Anopheles labranchiae este o specie endofilică, locuind în locuințe umane, grajduri, adăposturi de animale și ocazional în aer liber în adăposturi naturale (cavități de copac). Prezența larvelor a fost semnalată din aprilie până în octombrie. Zborul acestei specii este limitat la 2-5 km (Senevet and Andarelli, 1956). Anopheles labranchiae are o distribuție limitată în sudul și sud-estul Europei . Acesta a fost raportat în sud-estul Spaniei, Corsica, zonele de coastă din Italia, Sardinia, Sicilia. În Africa de Nord specia se găsește în Maroc, Algeria și Tunisia (Zahar, 1990). Anopheles labranchiae a înregistrat o creștere în Sardinia în ultimii 35 de ani. În 1940-1953 perioadă în care a fost eradicată malaria, Anopheles labranchiae a scăzut foarte mult, campania de eradicare constând în folosirea insecticidelor (DDT). Extinderea locuințelor urbane și modificările în practicile agricole au dus la o nouă creștere a populațiilor de Anopheles labranchiae. Sardinia este de interes biologic datorită poziției sale geografice centrale (lat 38-41 N, long 8-9E) în vestul Mării Mediterane, la jumătatea distanței dintre Africa și Europa. Sardinia este a doua insulă ca mărime din Marea Mediterană (250 x 120 km). Locurile de reproducere pentru Anopheles lambrachiae sunt foarte variate: mlaștini, râuri acoperite cu vegetație, găuri, canale, bazine, preferând locurile însorite. Anopheles labranchiae și Anopheles sacharovi sunt considerați a fi cei mai eficienți vectori în transmiterea malariei în regiunea palearctică

25
(Marchi, 1987). Principalele sit-uri de reproducere pentru Anopheles labranchiae în această regiune rămân câmpurile de orez, care continuă să fie o preocupare principală în agricultură. În Corsica, Anopheles labranchiae este în continuare vectorul principal al malariei, alături de Anophles sacharovi. Alte specii considerate posibili vectori în alte părți ale Europei și anume Anopheles algeriensis, Anopheles atroparvus, Anopheles claviger, Anopheles hyrcanus, Anopheles maculipennis SS, Anopheles marteri, Anopheles melanoon, Anopheles messae, Anopheles petragnani, Anopheles plumbeus, Anopheles superpictus, Anopheles subalpinus datorită caracterelor fiziologice, în special zoofilismul, li se atribuie un rol minim în transmiterea malariei (Toty C et al, 2010). Centrul Italiei a fost o zonă hiper-endemică pentru malarie până la mijlocul secolului XX, atunci când campania de eradicare a malariei a redus drastic numărul vectorilor incriminati în transmiterea malariei, Anophele labranchiae. Introducerea culturilor de orez a condus la o creștere substanțială a acestei specii, devenind îngrijorător pentru riscul re-emergenței malariei în această zonă. Modificarea utilizării terenurilor, variațiile climatice și demografice sezoniere au contribuit la intensificarea populațiilor de Anopheles labranchiae, mărind riscul de malarie autohtonă (Boccolini et al, 2012). În Maroc, Anopheles labranchiae este considerat principalul vector al malariei, ultimul caz de malaria autohtonă înregistrându-se în 2004. Controlul malariei include în această regiune folosirea metodei de combatere biologică și anume peștele larvivor (Gambusia holbrooki). Insecticidele sunt utilizate ca ultimă opțiune, ocupând totuși un loc important în cadrul Programului Național de control al malariei (NMCP). Insecticidul folosit pentru distrugerea larvelor de Anopheles labranchiae, recomandat de OMS este un organofosforic, temephos, concentrația de 0.125mg / L fiind cea mai mică concentrație cu 100% mortalitate (Faraj, 2010). Temephos este insecticidul folosit în distrugerea larvelor iar DDT-ul este utilizat în controlul adulților. Utilizarea pe scară largă a insecticidelor în agricultură poate constitui o problemă întrucât Anopheles labranchiae este frecvent întâlnit pe terenurile agricole. Anopheles labranchiae este singura specie din complexul maculipennis întâlnită în Africa de Nord. Anopheles labranchiae este pricipalul vector de transmisie a malariei și în Italia. În regiunea Toscana a fost înregistrat un caz de malaria autohtonă în 1997, în urma unui studiu, Anopheles labranchiae fiind singura specie de Anopheles identificată în regiune. Un studiu făcut pe acestă specie a demonstrat că în luna

26
august posibilitatea de transmisie a malariei este mare, îndeplinindu-se durata necesară dezvoltării parazitului în țânțar, 11 zile pentru Plasmodiumfalciparum și 10 zile pentru Plasmodium vivax la temperatura de 25°C, temperatura medie a zilei (Romi, 1999). Un articol din 1998 afirmă că în Spania, Anopheles labranchiae a dispărut (Eritja et al, 1998). Lipsa de similitudine a distribuției între Anophles atroparvus și Anophles labranchiae, pare a fi temperatura, ultimul preferând apele calde. Ambele specii preferă apele salmastre și lagune, dar Anopheles labranchiae preferă pentru depunerea ouălor, apele dulci. În Sardinia au fost găsite larve în aproape toate habitatele cu excepția celor foarte umbrite (Sinka et al, 2010). Femelele de A. labranchiae atacă cu ușurință omul, Romi şi colaboratorii au semnalat prezența sângelui uman la 86% - 90,7% femele hrănite Romi et al, 1997. Anopheles labranchiae poate fi atât endo cât și exofilic în funcție de habitatele disponibile (D’Alessandro et al, 1971). La fel ca A. atroparvus, A. labranchiae s-a dovedit a fi refractar la infecțiile cu tulpini africane de P. falciparum, deși la Centre de Production et d'Infection d'Anophèles (CEPIA) din Paris s-a demonstrat infecția A. labranchiae cu o tulpină africană de P. falciparum NF54 (Toty C et al, 2010). ANOPHELES ATROPARVUS (VAN THIEL 1927) Anopheles atroparvus își desfășoară activitatatea începând cu 15°C, când poate constitui risc în transmiterea malariei (Knottnerus, 2002). Preferă pentru reproducere apele salmastre sau dulci, bine luminate (Potugalia, Spania). Preferă apa relativ rece, larvele au fost identificate în mlaștini, șanțuri, bazine de inundații a solului, piscinele formate în albiile râurilor, cisterne pentru transport ciment, câmpuri de orez, fântâni și bazine dar și colecții mici de apă în anvelope uzate. Preferă situ-rile expuse la soare cu alge verzi filamentoase și alte plante acvatice. Anopheles atroparvus este o specie predominant zoofilică, preferând în ordine: iepurele, calul, vaca, porcul și oaia, fiind principalul vector al mixomatozei la iepure. Cu toate acestea a fost descris și ca antropofil fiind colectat în locuințe și capturi asupra oamenilor. Hibernarea se face în grajduri sau locuințe, ca femelă adultă, luând prânzuri periodice fără producție de ouă. Acest lucru a dus la răspândirea interioară de malarie în Olanda, Marea Britanie și alte părți ale Europei la începutul sec XX. Durata diapauzei depinde de lungimea zilei și de temperatură, astfel variază cu latitudinea și poate varia din septembrie până în octombrie în nordul Europei, sau din noiembrie până în februarie în sudul Europei. Este în general o specie de litoral, ce apare în sud-estul Portugaliei, de-a lungul coastelor Oceanului Atlantic, Marea Britanie și Marea Mediterană. În sudul și sud-estul Europei are o distribuție neuniformă. În Macedonia este larg răspândită în zona de câmpie, vizibil dominantă numai în zonele cu soluri saline/alcaline din Câmpia Pannonică. În Portugalia este cea mai comună și mai distribuită specie (Becker et al, 2003).

27
Într-un studiu realizat în 2010, ce a urmărit identificarea celor mai frecvenți vectori din 49 de țări din Europa și Orientul Mijlociu, cuprinzând 2891 de locații, Anopheles atroparvus a fost semnalat în cele mai multe locații 1044 (Sinka et al, 2010). Anopheles atroparvus a fost principalul vector incriminat în transmiterea malariei în Insulele Britanice, fiind principala sursa de P. vivax. Pentru P. falciparum nu s-a stabilit dacă parazitul este capabil să finalizeze ciclul de viață și de a transmite infecția. Cele mai multe cercetări afirmă că Anopheles atroparvus este incapabil să transmnită tulpini tropicale de Plasmodium falciparum, dar poate transmite tulpini europene (Cambournac, 1994). Un studiu al lui Sousa menționează existența unui Anopheles atroparvus în Portugalia, capabil să transmită tulpini tropicale de P. falciparum (Sousa, 2008). Anopheles atroparvus este larg răspândit în Anglia, Scoția, Țara Galilor și Irlanda, alături de Anopheles messeae de care nu se poate diferenția morfologic decât în stadiul de ou (Snow, 1998). Anopheles atroparvus este larg răspândit în țările din bazinul mediteranean, dar datorită caracterului zoofilic nu reprezintă un risc major în transmiterea malariei, decât dacă populațiile ar avea densitate mare (Eritja et al, 1998). Distribuția lui variază în Europa, astfel este absent în unele regiuni mediteraneene, cum ar fi sudul Italiei, Grecia și Turcia (Vicente et al, 2011). Anopheles atroparvus, alături de Anopheles messae sunt considerați principalii vectori ai malariei în UK. Identificarea genetică pe baza secvenței ITS2 a fost confirmată de Linton et al. în 2002. Un studiu realizat în 2001, pe malurile râurilor Rin și Meuse, în partea de sud-vest a Țărilor de Jos a demonstrat că populațiile de Anopheles atroparvus, considerat principalul și singurul vector al malariei în zonă, a scăzut foarte mult, doar 4 din 150 de larve examinate au reprezentat A. atroparvus, restul fiind identificate prin PCR ca A. messeae (Takken, 2002). ANOPHELES SACHAROVI (FAVRE 1903) Această specie se distinge cel mai ușor din tot complexul maculipennis, prin culoarea luminoasă a mezonotului și prin banda mediană de pe scutum care la ceilalți membri ai complexului este absent. Părțile laterale ale scutumului sunt ,maro-gălbui ca și partea din mijloc. Petele de pe aripi sunt foarte puțin evidente, în comparație cu ceilalți membri ai complexului, la exemplarele vechi abia perceptibilă. Ouăle de A. sacharovi se disting prin lipsa flotoarelor iar larvele sunt mai mici decât ai celorlalți membri ai complexului (Becker et al, 2003). Anopheles sacharovi este considerat a fi cel mai eficient vector al malariei, vector primar în Turcia, Siria, Orientul Mijlociu, precum și Europa de Sud (Yaghoobi-Ershadi et al, 2001; Romi et al, 2002; Sedaghat et al, 2003).

28
Anopheles sacharovi este foarte plastic în alegerea habitatelor atât ca adult cât și ca larvă, se găsește în toate colecțiile de apă mică ce conține vegetație. Preferă habitatele cu apă dulce, dar a fost descris ca cel mai tolerant al salinității până la 20%, din tot grupul Maculipennis. Poate supraviețui în ape de până la 38-40°C, în ape stătătoare, putând face față și unui curent slab (Pener and Kitron, 1985). Această specie preferă sit-urile însorite, un habitat perfect fiind mlaștinile, dar a fost întâlnit și pe marginea râurilor, pâraie, izvoare, piscine și șanțuri. Ca urmare a campaniilor de eradicare și folosirea DDT-ului în trecut, s-a considerat caracterul zoofilic însă în prezent este apreciat un comportament antropofilic (Sinka et al, 2010). Spre deosebire de ceilalți membri ai complexului Maculipennis, A. sacharovi atacă pe toată durata zilei, deși intensitatea maximă este seara devreme între 20:00-22:00 (Alten et al, 2003). Hibernarea este incompletă, femela hrănindu-se de mai multe ori, fără să depună ponta (Manguin et al, 2008; Kasap, 1995). Anopheles sacharovi este vector important în Turcia, Siria, Israel, Iordania și Orientul Mijlociu (Zahar, 1990; Sedaghat et al, 2003). Pentru reproducere preferă apele mici cu vegetație acvatică. Anopheles sacharovi este considerat pricipalul vector în Balcani, în România a fost declarat dipărut după eradicarea malariei din aceste zone (Zahar, 1990), iar în Italia este întâlnit foarte rar (din cauza schimbărilor de mediu și a modificărilor practicilor agricole). Specia este încă abundentă în Turcia și nord-estul Greciei (Romi, 1999). Anopheles sacharovi este o specie care capătă repede rezistență la insecticide, cum arată un studiu realizat în 2012. Astfel nivelurile de iritabilitate descrise de OMS sunt DDT 4%, dieldrin 0,4%, malation 5%, fenitrotion 1%, permetrin 0,75%, iar pentru deltametrin 0,05%. Rezultatele arată că la doza de diagnosticare a insecticidelor pentru această specie s-a semnalat rezistență la DDT, toleranță la dieldrin și oarecum sensibil la fenitrothion, malathion, permetrin și deltametrin. DDT-ul a avut efectul cel mai iritant iar deltamethrin cel mai mic efect iritant (Vatandoost and Abai, 2012). În laborator, în diferite condiții ecologice, la altitudini de la 353 la 1126 m în provincia Sanliurfa din Turcia, Anopheles sacharovi nu a prezentat diferente semnificative de la ou la adult, sau diferențe de dezvoltare. Anopheles sacharovi este vectorul principal al malariei în Turcia și este, împreună cu Anopheles superpictus și Anopheles pulcherrimus, cel mai important vector al malariei în fosta Uniune Sovietică, deși A. messeae a fost implicat în renașterea malariei în Rusia și Ucraina. Anopheles maculipennis a fost considerat vector

29
principal al malarie pe coasta Mării Caspice și Anopheles sacharovi este considerat vector principal al malariei în platoul central al țării. Membrii acestui complex de specii sunt activi în nordul Iranului din mai până în septembrie, cu un vârf în iulie, dezvoltându-se bine în câmpurile de orez și apă curată, adulții găsindu-se în adăposturi de animale în proporție de 95%. Hrănirea cu sânge începe la ora 19.00 cu un vârf între 20.00-21.00, testul imunoenzimatic ELISA, arătând o preferință pentru vite, urmate într-o mică măsură de oi și păsări. Indexul antropofilic pentru această specie în nordul Iranului este de 1,7-4.9% (Djadid et al, 2007). Republica Azerbaidjan și Armenia a fost liberă de malaria mai bine de 30 de ani, până în 1994 când a fost reintrodusă, identificându-se ca și vectori A. sacharovi și A. superpictus. ANOPHELES MESSEAE (FALLERONI 1926) Anopheles messeae este cel mai răspândit membru a complexului Maculipennis, având o distribuție din Irlanda în Europa și Asia, China și Rusia. La A. messeae au fost identificate o serie de polimorfisme genetice, și s-au definit cinci haplotipuri separate în funcție de diferite zone geografice din distribuția sa; neputându-se demonstra dacă aceste polimorfisme au fost indicative de comportament alterat în aceste zone (Di Luca et al, 2004). Larvele sunt găsite în ape umbrite, foarte lin curgătoare sau stătătoare cum ar fi marginile lacurilor și mlaștinile. Larvele au fost găsite în sit-uri care conțin stuf, buruieni plutitoare acvatice, alge, apa limpede din lacuri mici și dune. Este specia care se găsește predominant în zonele inundabile ale marilor râuri ca Dunăre, Rhone, Save sau Rin. Este întâlnită foate rar sau aproape deloc de-a lungul coastelor și în regiunile muntoase (Becker et al, 2003). Anopheles messeae a fost găsit în locuințe umane, în interioare, scări, subsoluri, grajduri, în aer liber (scorburi, cavități subterane, iazuri) au fost colectate puține exemplare (Sinka et al, 2010). În Țările de Jos, A. messeae nu este considerat un pericol în transmiterea malariei datorită caracteristicilor zoofilice, hrănindu-se pe om doar dacă populațiile sunt foarte dense și animalele sunt puține (Takken et al, 2002). Spre deosebire de A. labranchiae și A. atroparvus, A. messeae intră în diapauză, hibernează ca femelă adultă preferând clădirile părăsite fără animale, lipsind hrănirea cu sânge până în primăvară, obținând energia din rezervele de grăsime (Jaenson et al, 1991). Anopheles messeae poate transmite malaria la temperaturi scăzute chiar de 4 °C, caz în care este necesar 44 de zile pentru ca țânțarul să devină infecțios. Astfel se explică cazurile din Scoția și Norvegia. În Marea Britanie, Anopheles messeae și Anopheles atroparvus sunt responsabile de cazurile de malarie semnalate (Knottnerus, 2002). Anopheles messae este întâlnit rar în apele ce conțin mai mult de 1,5g NaCl/l. Specia preferă locurile umbrite, fiind asociată cu A. typicus și A. melanoon (Zahar, 1990). Anopheles messeae este vector

30
important al malariei în Asia de Vest, recent a fost identificat ca vector principal al malariei renăscute din Ucraina și Rusia (Sedaghat et al, 2003). ANOPHELES MELANOON (HACKETT 1934) Anopheles melanoon este întâlnit în apa proaspătă și câmpurile de orez din nordul Italiei. De asemenea este întâlnit în zonele mlăștinoase, în apă stagnantă cu suprafață mare, margini de râuri și lacuri, iazuri și piscine. În Sardinia larvele pot fi găsite apă dulce, însorită în primăvară și umbroasă în timpul verii. De asemenea a fost raportată în câmpurile de orez (Becker et al, 2003). Hibernarea se face cu diapauză completă, fiind specie în principal zoofilică și ocazional doar se hrănește pe oameni atât în aer liber cât și în interior. A fost identificat în regiunea Anatolia din Turcia unde malaria este endemică, alături de A. sacharovi și A. maculipennis (Akiner and Cağlar, 2010). A. melanoon este o specie rară cu o distribuție limitată la sud-vestul și sudul Europei.
ANOPHELES MACULIPENNIS (MEIGEN 1818) Anopheles maculipennis își are habitatul de reproducere în apele reci, în regiuni montane, dar a fost întâlnit și aproape de mare în asociere cu Anopheles messeae în ape curgătoare. De asemenea a fost întâlnit în ape calde, de exemplu în Italia, Napoli (Zahar, 1990). Anopheles maculipennis a fost identificat ca vector major al malariei pe litoralul mării Caspice din Iran. Site-urile de reproducere caracteristice sunt zonele protejate ale marginilor cursului râului, câmpurile de orez și bazine artificiale. În regiunile montane din Europa Centrală, specia poate fi întâlnită la altitudini de 1000 m și mai mult, fiind singurul membru al complexului întâlnit în asemenea condiții. Altitudini de 2190 m și până la 2300 m sunt raportate de Bulgaria și Turcia. Hibernarea este completă dar poate fi de scurtă durată în zonele cu climă caldă. Femelele sunt în general zoofile, hrănindu-se în principal pe bovine dar și pe porci și pui, iar când acestea sunt în deficit se pot hrăni și pe oameni, atât în aer liber cât și în interior. Sunt specii predominant endofile, sit-urile de odihnă fiind grajdurile și locuințele. Distribuție: A. maculipennis S.S. este larg răspândit în întreaga Europă. Cu excepția sudului Peninsulei Iberice este înregistrat în aproape fiecare țară europeană. Se extinde spre est în sud-vestul Asiei și Golful Persic. Este considerată a fi o specie mai continentală cu cereri de umiditate considerabil mai mici decât cele ale unui A. messae și A. atroparvus (Becker et al, 2003).

31
ANOPHELES DACIAE A fost semnalată ca nouă specie ce aparține complexului Maculipennis de către Nicolescu şi colaboratorii în 2004, capturată pe coasta Mării Negre și în câmpiile adiacente Dunării, în sudul României (Nicolescu et al, 2004). Această specie este similară cu Anopheles messae (diferența a fost stabilită pe baza secvenței ITS2, stabilindu-se o diferență de 1.03% baze. Anopheles daciae a fost recent semnalat și în UK, dovedindu-se doi ani la rând că este vorba de o populație stabilă și nu există susceptibilitate de artefact (Linton et al, 2005). ANOPHELES SUBALPINUS (HACKETT ET LEWIS 1935) Anopheles subalpinus se reproduce în apele dulci sau ușor saline, ca în mlaștini însorite și iazuri cu vegetație, evitând locurile de reproducere umbrite. Este de asemenea întâlnit pe câmpurile de orez, primăvara preferând apele stagnante cu vegetație orizontală. În timpul verii ouăle sunt mai ușor de recunoscut, ca și în cazul lui A. typicus și A. messae, primăvara și toamna fiind de culoare neagră (Zahar, 1990). Sit-urile de reproducere regulate pot fi găsite până la o altitudine de 1200 m. Hibernarea la femele adulte este completă, în peșteri și clădiri abandonate. A. subalpinus este considerată ca fiind o specie puternic zoofilă, care rareori se hrănește pe oameni. Femelele sunt endofile și pot fi găsite în timpul zilei în număr mare în grajduri și hambare și mai rar în locuințe umane sau clădiri. Este o specie sudică europeană cu un interval între Peninsula Iberică, prin nord Țările mediteraneene în zona de câmpie din jurul Mării Caspice (Becker et al, 2003). ANOPHELES GAMBIAE Este considerat cel mai important vector al malariei de pe continentul African (Fig. 10). În 1960 a fost recunoscut existenţa unui complex Anopheles gambiae sau Anopheles gambiae sensu lato care cuprinde următoarele specii: Anopheles arabiensis, Anopheles bwambae, Anopheles melas, Anopheles merus, Anopheles quadriannulatus, Anopheles gambiae sensu stricto, Anopheles coluzzii, Anopheles amharicus. Luate individual speciile ce aparţin complexului sunt foarte dificil de diferențiat morfologic, putându-se într-o oarecare măsură să se facă după larvă sau femela adultă. În schimb trăsăturile comportamentale diferă, astfel Anopheles quadriannulatus este o specie de apă sărată și de apă minerală, A. melas și A. merus sunt specii de apă sărată, în timp ce restul sunt specii de apă dulce. Anopheles quadriannulatus este de preferinţă zoofil, hrănindu-se pe animale, în timp ce Anopheles gambiae sensu stricto se hrănește în general pe oameni, fiind considerat antropofil. Identificarea prin metode moleculare rămâne cea mai sigură, propunerile

32
echipei lui Fanello putând avea implicații importante pentru măsurile ulterioare de control (Fanello et al, 2002). Membrii complexului Anopheles gambiae sunt răspândiţi în Africa tropicală, la sud de deşertul Sahara, cu Anopheles arabiens care se extinde în sudul Arabiei. Anopheles gambiae s.s. este distribuit în Africa Subsahariană, inclusiv în Madagascar (OMS 1989). În habitatele deschise fără copaci se confruntă cu temperaturi mai ridicate la prânz decâz habitatele împădurite, astfel că efectul antropic are ca rezultat scurtarea ciclului gonotrofic al femelelor de Anopheles gambiae s-a scurtat cu 2,6 zile (52%) şi 2,9 zile (21%) în anotimpurile uscate şi respectiv ploioase, comparativ cu zonele împădurite (Patz and Olson, 2006).
Fig. 10. Anopheles gambiae (Sursa: https://cameronwebb.wordpress.com/tag/anopheles-gambiae/)
Femelele adulte se pot distinge morfologic, muşchii din apropierea gurii prelungindu-se fiind la fel de lungi ca şi trompa, iar în poziţie statică abdomenul este ţinut în aer (Foster and Walker, 2009). Culoarea variază de la maro deschis la gri, cu pete pale de culoare galben, alb sau crem şi pete întunecare pe aripi. Dimensiunile adulţilor sunt relativ mici variind între 2,8 şi 4,4 mm (Gillies and de Meillon, 1968).

33
Ouăle măsoară aproximativ 0,47 și 0,48 mm, sunt convexe pe faţa dorsală și concave pe cea ventrală, prezentând un desen poligonal. Sunt depuse individual pe apă, fiecare ou având flotoare pe ambele părți, rezistenţa la secetă este scăzută (Foster and Walker, 2009). Larvele de Anopheles gambiae trăiesc în mod obişnuit în locuri cu apă dulce, proaspătă, adâncă şi temporară cum ar fi depresiunile din sol, băltoace, piscine. Această caracteristică poate permite evitarea prădătorilor, deoarece larvele se pot dezvolta foarte repede (șase zile de la ou la adult în condiții optime), vegetaţia este foarte redusă ca urmare a naturii tmeporare a habitatelor, de asemenea alge plutitoare, câmpuri de orez şi situri lipsite de vegetaţie pot constitui habitate adecvate. Anopheles gambiae, se reproduce de obicei în locuri populate de oameni, după hrănire femela preferă să se odihnească pe pereţii din apropierea sau din interiorul locuinţelor. Acest comportament poate oferi o oportunitate de a eradica această specie din satele și casele din Africa prin utilizarea insecticidelor reziduale. Femelele își găsesc gazdele folosind o varietate de receptori senzoriali, răspund la mișcări, gradienți de dioxid de carbon și transpirație (Konate et al 1999; Meijerink et al, 2000; Roberts and Janovy, 2000).
EPIDEMIOLOGIA MALARIEI LA OM
SURSA DE AGENT PATOGEN Oamenii sunt cea mai importantă sursă de agenți patogeni, cu excepția Plasmodium malariae, care este comun oamenilor, maimuțelor africane și anumitor specii de maimuțe din America de Sud. Primatele non-umane sunt infectate în mod natural cu diferite specii ale parazitului, unele dintre ele fiind înrudite cu speciile umane (P. cynomolgi, P. simium, P. brasilianum, P. knowlesi). Acești paraziți trebuie monitorizați cu atenție deoarece au existat cazuri de infestări de laborator (experimentale) cu P. cynomolgi și infestări naturale (transmise prin țânțar de la animale la gazda umana) cu P. knowlesi și P. brasilianum, fapte care au făcut posibilă documentarea acestora specii ca o sursă de agenți patogeni pentru malaria umană (Heymann, 2012; Bocsan et al, 1999). Oamenii pot fi surse de agenți patogeni în trei situații: atunci când sunt bolnavi, purtători preinfecţioşi (viitorii bolnavi în timpul perioadei de incubație) sau purtători postinfecțioşi cronici. Perioada de incubație diferă în funcție de specie (10-12 zile pentru malaria produsă de P. falciparum, 13-15 zile când sunt implicați P. vivax și P. ovale și 27-42 zile pentru cazurile de malarie produse de P. malariae). Aceste perioade de timp pot fi prelungite (8-10 luni) dacă schemele de chimioprofilaxie nu sunt utilizate corect. De asemenea, perioada contagioasă (în timpul

34
căreia gametociţii sunt în sânge) variază în funcție de specia de Plasmodium și răspunsul la tratament: starea de purtător cronic poate fi prelungită până la 1-2 ani în infecțiile cu P. falciparum netratate, până la 4 ani în cazul infecției cu P. vivax și P. ovale și până la 30 de ani sau întreaga viață pentru infecția cu P. malariae (Heymann, 2012; Bocsan et al, 1999). O altă sursă importantă de agent patogen este reprezentată de țânțarii Anopheles.
MODURI ŞI CĂI DE TRANSMITERE Din cele peste 400 de specii de Anopheles, aproximativ 80-85 pot transmite malaria și doar 45 sunt considerate vectori de importanță majoră (Bocsan et al, 1999; Dondorp et al, 2017; Al-Eryani et al, 2016). În corpul țânțarilor, după trecerea prin ciclul sporogonic, gametociţii sunt transformaţi în sporozoiți, care vor fi inoculați prin mușcătură din glandele salivare ale țânțarilor direct la gazda umană receptivă. Ciclul sporogonic are o durată variabilă în funcție de specia de Plasmodium (pentru P. vivax - 10-14 zile, pentru P. falciparum - 16-23 zile și pentru P. malariae - 15-30 zile) și temperatura ambiantă (Bocsan et al, 1999). Malaria poate fi trasmisă atât direct, cât şi indirect. Modul direct poate fi realizat prin mușcătura de țânțar, prin transfuzii de sânge contaminat sau prin transplant medular. De asemenea, malaria poate fi transmisă de la mamă la făt în cazul anomaliilor placentare (malaria congenitală). Modul indirect implică utilizarea căilor de transmitere contaminate, în general a acelor și a seringilor contaminate de la utilizatorii de droguri intravenoase. Diseminarea agentului patogen are loc atât timp cât formele asexuale pot fi găsite în sângele circulant al sursei. În corpul subiectului receptiv, merozoţii continuă să se dezvolte în sângele periferic cu periodicitatea proprie, conducând la apariţia accesului de malarie. Pentru această categorie nu există recidivă deoarece nu există hipnozoiţi, dar devin infecțioase pentru țânțar datorită existenței gametociţilor. Cel mai frecvent agent patogen implicat în malaria transfuzională este P. malariae (care persistă asimptomatic pentru o perioadă lungă de timp), iar cea mai severă formă clinică a bolii este produsă de P. falciparum (Bocsan et al, 1999; Luca, 2002). Transmiterea malariei nu se produce la temperaturi ambientale sub 16°C (61°F) sau peste 33°C (61°F) şi la o altitudine de 2000 m peste nivelul mării deoarece dezvoltarea sporozoitului nu poate avea loc (Dondorp et al, 2017). Ca urmare, un rol important în transmiterea bolii îl are comportamentul vectorului şi modalitatea în care se realizează prânzul hematofag, cel mai frecvent prin muşcătură produsă seara, în timpul nopţii sau dimineaţa devreme. Un al factor cu rol major în transmiterea malariei este faptul că la gazda umană o populaţie de gametociţi apare după o serie de câteva cicluri asexuate pentru P. falciparum şi ceva mai devreme pentru P. vivax. Densitatea gametociţilor este mai crescută la pacienţii cu densitate mare a parazitului, la

35
indivizii imunosupresaţi (copiii) şi atunci când se utilizează antimalarice la care parazitul prezintă rezistenţă. Unele hemoglobinopatii genetice (exemple: siclemia, deficienţa de G6PD sau talasemia) conferă protecţie faţă de malaria cu P. falciparum (Dondorp et al, 2017; Abdul-Ghani et al, 2016).
RECEPTIVITATE Receptivitatea este generală, cu anumite particularităţi legate de rezistenţa faţă de malaria produsă de unele tipuri de Plasmodium (Bocsan et al, 1999). Răspunsul imun dezvoltat după infecţia naturală este de tip umoral şi celular, imunitatea umorală (anticorpi faţă de formele eritrocitare asexuate) constituind elementul cel mai important al protecţiei. Anticorpii apar precoce în cursul parazitemiei şi ating un titru maxim odată cu scăderea numărului de paraziţi circulanţi. În zonele endemice pentru malarie, imunitatea naturală pasivă, trasnplacentară (IgG de origine maternă), asigură protecţia nou-născutului până în jurul vârstei de 3 luni, după care nivelul acestor anticorpi se scade trepatat până la dispariţie. În primul an de viaţă al acestor copii, sistemul imunitar este expus continuu la contactul cu Plasmodium şi organismul îşi începe propria producţie de anticorpi (de tip IgM şi IgG) în prezenţa parazitemiei. Studiile epidemiologice arată că acest răspuns imun scade riscul apariţiei complicaţiilor severe şi al decesului prematur la copiii care au trăit primul an de viaţă în zone endemice, comparativ cu cei care au trăit primul an de viaţă în afara zonelor endemice (Bocsan et al, 1999; Luca, 2002). În populaţia adultă din aceste zone endemice se remarcă apariţia unor forme clinice mai uşoare ca urmare a imunizării oculte prin contactul repetat cu ţânţarul sursă de agent patogen pe parcursul anilor (Heymann, 2012). Studiile au arătat că un procent important din populaţiile indigene din Africa de Vest prezintă o rezistenţă naturală faţă de infecţia cu P. vivax, datorată absenţei antigenului Duffy de la nivel eritrocitar. Persoanele heterozigote care suferă de siclemie sunt relativ protejate faţă de formele severe de infecţie cu P. falciparum, nivelul parazitemiei fiind unul scăzut, însă homozigoţii au un risc crescut de a dezvolta forme grave de boală, adesea cu evoluţie spre deces (Heymann, 2012). La indivizii neimunizaţi (călători din zone indemne de malarie), boala este manifestă întotdeauna şi de cele mai multe ori îmbracă forme grave, iar imunitate după trecerea prin boală este tranzitorie şi nespecifică (Bocsan et al, 1999). O categorie specială de receptivi o constituie persoanele HIV pozitive din zonele endemice, care fac mai frecvent boala, dar care prezintă adesea un răspuns scăzut la terapia anti-malarică (Heymann, 2012).

36
Receptivitatea faţă de boală este infuenţată şi de funcţionarea normală a splinei, care are rol în apărarea organismului (în imunitate), dar şi de filtru pentru hematiile parazitate (sferice şi mai puţin deformabile). Persoanele cu asplenie, funcţională sau anatomică, fac forme cu progresie rapidă, o mare parte din celulele circulante în sângele periferic fiind parazitate. Recăderile se datorează formelor dormante pentru malaria cu P. vivax şi P. ovale, iar pentru infecţia cu P. malariae este implicată persistenţa parazitului în sânge pentru perioade lungi de timp (Bocsan et al, 1999).
FORMELE DE MANIFESTARE ALE MALARIEI ÎN POPULAŢIE Malaria poate fi continuă, sezonieră și intermitentă. Malaria sezonieră se caracterizează printr-un val de infecție în primăvară și un val în toamnă cu amplitudine mai mare. Malaria este sezonieră în regiunile unde condițiile climatice diferă de la un sezon la altul, permițând dezvoltarea periodică a parazitului malariei și a vectorului, populația indigenă neavând timp să dezvolte o imunitate suficientă, astfel grupurile de persoane cu risc de contractare a bolii sunt greu de definit și mortalitatea poate fi mai mare. În marea majoritate a cazurilor, epidemiile sunt legate de absența, scăderea sau pierderea imunității, ceea ce trebuie să constituie o alarmă pentru țara noastră, unde populația este lipsită complet de imunitate. Paraziții se dezvoltă în țânțar cu o viteză dependentă de temperatura ambiantă (astfel că la tropice în aproximativ 12 zile parazitul ajunge în glandele salivare fiind gata de infecție). La cealaltă extremă se află malaria stabilă caz în care transmisia este continuă sau regulată pe sezoane și toți sunt în curs de infecție sau reinfecție. Conform Organizaţiei Mondiale a Sănătăţii există cinci tipuri de focare de malarie (Tabelul I).
Table I. Tipuri de focare de malarie conform clasificării OMS (Source: Colofiţchi, 2007)
1. Focar natural restant inactiv
Focar în care transmiterea agentului patogen a fost oprită prin menținerea morbidității la nivelul infecțiilor anterioare
2. Focar natural restant activ Focar în care măsurile antimalarie au condus la o scădere considerabilă a transmiterii agentului patogen, dar transmiterea este încă efectuată.
3. Focar natural nou potenţial
Focar asanat, cu prezența cazurilor de malarie
4. Focar natural nou activ Focar în care transmiterea agentului patogen a fost efectuată de la un caz importat (poate deveni endemică)
5. Focar asanat Focar în care transmiterea agentului patogen a fost oprită pe o perioadă de timp egală sau mai mare de trei sezoane epidemice

37
DISTRIBUŢIA GEOGRAFICĂ Raportul prezentat de OMS privind malaria la nivel global arată că pentru anul 2017 situația este alarmantă deoarece mai puțin de jumătate din țările cu transmitere a malariei au reușit să atingă obiectivele privind reducerea ratei mortalității cauzate de infecția cu parazitul Plasmodium. În 2016 au fost înregistrate 216 milioane de cazuri de malarie la nivel mondial, cu 5 milioane mai mult comparativ cu 2015. Chiar dacă din anul 2010 a existat o scădere a cazurilor de malarie, începând cu 2014, ratele au stagnat sau tendința a fost ascendentă în unele regiuni unde a fost înregistrată o creștere a cazurilor de malarie. recorded (WHO World Malaria Report, 2017). Se estimează că în Africa sub-Saharană proporţia celor infectaţi cu diferite specii de Plasmodium a scăzut de la 17% în 2010 la 13% în 2015 (IC = 11–15%). Numărul persoanelor infectate cu parazit a scăzut de la 131 milioane (IC = 126–136 milioane) în 2010 la 114 milioane în 2015 (IC = 99–130 milioane) (WHO World Malaria Report, 2016). În cea mai mare parte a Africii sub-Sahariene, transmiterea este moderată spre crescută, cu o prevalenţă a malariei produse de P. falciparum de peste 50% (corespunzând unui număr de înţepături infectante = 10 - 1000/an, numit şi rata de inoclare entomologică). În afara Africii, malaria cu P. falciparum prezintă o endemicitate scăzută, cu o rată de prevalenţă de sub 10% (corespunzând unei rate de inoclare entomologică = sub 1/an). P. vivax foarte frecvent în Asia, unde este responsabil de aproximativ 50% din cazurile de malarie, precum şi în America Centrală şi de Sud, Africa de Nord şi Orientul Mijlociu. Această specie a parazitului este foarte rară în Africa de Vest, unde predomină cazurile de boală cu P. ovale (care circulă mai frecvent şi în Asia). Prevalenţa malariei cu P. malarie şi P. ovale au, în general, o frecvenţă mai scăzută comprativ cu celelalte specii (Dondorp et al, 2017). În anul 2015, la nivel mondial au fost estimate 212 milioane de cazuri noi de malarie (IC = 148–304 milioane). Majoritatea cazurilor au fost înregistrate în Regiunea Africană a WHO (90%), urmată de Regiunea Asia de Sud-Est a WHO (7%) şi Regiunea Mediteraneană a WHO (2%). Rata de incidenţă a malariei a scăzut cu 41% la nivel global în perioada 2000 - 2015 şi cu 21% între 2010 şi 2015. Cele mai multe dintre decesele produse de malarie au survenit în Regiunea Africană a WHO (92%), urmată de Regiunea Asia de Sud-Est a WHO (6%) şi Regiunea Mediteraneană a WHO (2%). Majoritatea deceselor s-au produs ca urmare a infecţiei cu P. falciparum (99%). S-a estimat că P. vivax a fost responsabil pentru 3.100 decese în anul 2015 (IC = 1800–4900), 86% fiind identificate în afara continentului african (WHO World Malaria Report, 2016).

38
Călătorii internaționali ar putea fi expuși riscului de infectare cu malarie în 91 de țări din întreaga lume, în special în Africa, Asia și America (Fig. 11). Dar toate aceste date nu prezintă o realitate de 100%, statisticile exacte cu privire la situația actuală a malariei în Africa nu sunt posibile din cauza problemelor întâlnite în procesul de obținere a datelor din sistemul de sănătate și din cauza faptului că multe tratamente și decese nu sunt înregistrate.
Fig. 11. Ţările endemic pentru malarie (Sursa: http://wwwnc.cdc.gov/travel/yellowbook/2012/chapter-3-infectious-diseases-related-to-travel/malaria.htm)
În Europa, malaria a fost eradicată, cu excepția Azerbaidjanului, Georgiei, Kyrgyzstanului, Tadjikistanului și Turciei. Se estimează că 25-30 de milioane de persoane călătoresc în fiecare an din Europa în zone cu transmitere a malariei. Pentru anul 2010, OMS a raportat 6.244 de cazuri de malarie în Europa, un numar mai mic față de anul 2000, când au fost raportate 14.703 de cazuri, dar cifrele reale au fost estimate a fi de șase ori mai mari. Cele mai recente date din Marea Britanie și Europa arată o creștere importantă a cazurilor de malarie, aceeași imagine fiind prezentă şi în SUA, unde s-a înregistrat o creștere cu 14% față de 2010 (Askling et al, 2012).

39
În 2011, 40 de cazuri de infecție cu Plasmodium vivax au fost raportate în Grecia, la pacienți fără antecedente de călătorie într-o zonă endemică de malarie, în cinci districte diferite, și anume Lakonia (n = 34), Attiki (n = 2), Evoia (n = 2), Viotia (n = 1) și Larissa (n = 1). Un grup de cazuri a fost identificat în zona de Evrotas, Lakonia unde 27 de cazuri au fost la cetățeni greci care locuiesc în zonă și 7 la imigranți din țări non-endemice. În plus, 23 de cazuri de P. vivax în Lakonia au fost raportate la muncitori agricoli migranți din țări endemice de malarie. La 22 iunie, Centrul Elen pentru Prevenirea și Controlul Bolilor (KEELPNO) a raportat primul caz de malarie locală, cauzată de Plasmodium vivax, în 2012 la Marathon, regiunea Attica, Grecia. Cazul a fost un bărbat de 78 de ani, la care simptomele au apărut pe data de 7 iunie, fără istoric de călătorie recentă în țări endemice de malarie. Regiunea Marathon, din Attica, a fost identificată ca fiind locul posibil de infecție. Un al doilea caz de malarie locală cu Plasmodium vivax a fost raportat de KEELPNO la 17 iulie. Acest caz a fost o femeie de 48 ani rezidentă în municipiul Evrotas, Lakonia. Pacientul a raportat debutul simptomelor la 29 iunie. Pacientul nu a călătorit într-o zonă endemică de malarie în ultimii 5 ani. Cele două cazuri locale de malarie Plasmodium vivax raportate în 2012 au fost în localități afectate în 2011. În acest moment se iau măsuri sezoniere (primăvară-toamnă) de prevenire a malariei care deocamdată nu reprezintă riscul de difuzare a bolii în alte țări, zonele afectate fiind agricole și nu turistice (ECDC, 2012). Cazuri de malaria autohtonă au fost semnalate și în Germania (Krüger et al 2001). Peste 5 milioane de migranți din Africa trăiesc în Europa, o treime din ei venind din Africa sub-sahariană. Ca urmare a focarului de malarie din Grecia din 2011, Centrul European de Prevenire și Control al Bolilor (ECDC) a organizat o conferință în Europa în ianuarie 2012 pentru a stabili riscul transmisiei parazitului Plasmodium vivax în Europa. În statele membre ale UE malaria a fost eradicată din 1975, deși vectorii, țânțari din genul Anopheles, sunt prezenți în continuare. În ultimii 10 ani transmiterea sporadică de malarie a fost semnalată în multe țări din Europa: Bulgaria, Franța, Germania, Grecia, Italia și Spania. Între 21 mai și 5 decembrie 2011 în Grecia au fost înregistrate 63 cazuri de malarie, Plasmodium vivax. Ca urmare a informațiilor disponibile, se pare că transmisia malariei în Grecia să fie ca urmare a introducerii parazitului anual prin intermediul imigranților. Creșterea numărului de cazuri autohtone semnalate în 2011, indică faptul că pot exista condiții favorabile pentru transmiterea locală susținută în zonele afectate.

40
Cele mai multe cazuri de malarie în Europa sunt reprezentate de migranții care locuiesc în Europa care şi-au vizitat rudele şi prietenii în țările lor (VFR), ei fiind mai vulnerabili decât turiștii. În Franța, Paris, un studiu realizat într-un spital situat într-un cartier vecin cu o comunitate africană, a prezentat 239 de cazuri de malarie importată. 81,2% au fost cazuri de VFR care nu au luat medicație profilactică înaintea întoarcerii în țara natală (majoritatea din Africa sub-sahariană și Comore) (Develoux et al, 2012). Datele prezentate de TropNetEurop până în 1999, arată că majoritatea cazurilor de malarie cu Pl. falciparum au fost importate din Africa de Vest, europenii prezentând multe complicații (Jelinek et al, 2002). Malaria a constituit o problemă majoră de sănătate în Franța, în special în Camargue, unde s-au înregistrat cazuri până la începutul secolului XX. Malaria a fost eradicată în Franța în 1943, ca urmare a drenării terenurilor mlăștinoase, a creșterii animalelor, a dezvoltării construcțiilor şi utilizării corespunzătoare a chininei. Studii entomologice recente au semnalat populații însemnate de țânțari din genul Anopheles, Anopheles hyrcanus considerându-se ca principal potențial vector al malariei, pe baza antropofilismului și a abundenței. Numărul de cazuri de malarie importate au crescut foarte mult începând cu 1970, o dată cu intensificarea călătoriilor internaționale, cu o medie de aproximativ 6.400 de cazuri pe an, pentru ultimii zece ani, în Franța (Poncon et al, 2008). Numărul cazurilor de malarie importată a crescut în Europa, fapt asociat cu creșterea imigranților stabiliți în țările non-endemice, dar care își vizitează des rudele din țările de origine. În cazul imigranților simptomele clinice sunt ușoare, asimptomatice sau întârziate, datorate semi-imunității dobândite după perioade lungi de expunere la malarie stabilă. Cazurile de malarie din rândul imigrantilor, precum și cazurile asimptomatice cu parazitemie microscopică, duc la creșterea riscului de transmitere şi de reintroducere a malariei în anumite zone în care sunt prezenți vectorii și condițiile climatice sunt favorabile. Malaria mai poate fi reintrodusă prin intermediul imigranților și prin metode ce nu implică prezența vectorului: transfuzii de sânge, transplant de organe, transmitere congenitală. Screening-ul malariei trebuie să fie efectuat în rândul imigranților recent sosiți din țări endemice de malarie, în scopul reducerii riscului de malarie clinică, dar și pentru a preveni transmiterea autohtonă de malarie în zonele în care a fost eradicată. În 2010 au fost prezenți 47.300.000 de oameni de origine străină în Uniunea Europeană (UE), corespunzător unui procent de 9,4 din totalul populației. Majoritatea dintre aceștia, 31,4 milioane, s-au născut în țări din afara UE, în timp ce 16 milioane s-au născut într-un alt stat membru al UE. Datele despre cei care provin din țări endemice de malarie sunt

41
insuficiente. Estimările indică faptul că mai mult de 5 milioane de imigranți africani ar putea trăi în Europa. Dintre aceștia, circa doua treimi sunt din Africa de Nord (Algeria, Maroc și Tunisia), iar restul sunt din Africa sub-sahariană (SSA), cea mai mare parte din Africa de Vest (Ghana, Nigeria și Senegal). Aproximativ 4 milioane provin din Asia de Sud-Est și aproape 2,2 milioane din America Latină. Cele mai multe cazuri au fost importate în Europa de Vest, Franța, Regatul Unit (UK), Germania și Italia, reprezentând mai mult de 70% din toate cazurile (Monge-Maillo and López-Vélez, 2012). Persoanele infestate cu agentul malariei prezintă febră şi frisoane, simptomele fiind asemănătoare gripei. Netratată boala poate produce complicaţii severe şi în unele cazuri decesul. Simptomele malariei apar după o perioadă de incubaţie de 7 zile sau mai mult în funcţie de specia de Plasmodium. Febra care apare la călători în termen de trei luni de la expunere, reprezintă o urgenţă medicală ce trebuie investigată imediat. Imunitatea gazdei umane este un alt factor important, în special în rândul adulților, în zone cu condiții moderate sau intense de transmitere. Imunitatea parțială se dezvoltă pe parcursul anilor de expunerea repetată la parazitul malariei și, deși nu asigură niciodată o protecție completă, reduce riscul unei forme severe de infecție. Din acest motiv, majoritatea deceselor cauzate de malarie în Africa apar la copiii mici, în timp ce în zonele cu o transmitere mai redusă și imunitate scăzută, toate grupurile de vârstă sunt expuse riscului. În România, malaria a fost eradicată în 1965, fiind considerată în 1967 de către OMS o țară liberă de malarie. Riscul de reemergenţă a malariei în România, îl constituie prezența vectorului Anopheles, care face parte din complexul Anopheles maculipennis, a agentului patogen în natură și modificarea factorilor climatici cu creşterea temperaturilor. Conform raportului din 2015 al ECDC, situația cazurilor de malarie în România a cunoscut o tendință ascendentă în ultimii ani. Pentru perioada 2004-2015, Spitalul de Boli Tropicale "Victor Babeş" a raportat 150 de cazuri de malarie în România. Dintre acestea, 99% provin de pe continentul african și doar un caz din Europa - Grecia, raportat în 2011. Plasmodium falciparum a fost înregistrat la 75 de cazuri, urmat de P. vivax, P. malariae și P. ovale, cu cel mai mic raport. În 2010 au fost 19 cazuri, în 2011 - 40 de cazuri, în 2012 - 32 de imbolnaviri, în 2013 - 43, iar în 2014 - 57 de cazuri de boală. În anul 2015, au fost raportate 30 de cazuri confirmate de malarie (reprezentând o incidență de 0,15 la 100.000 de locuitori, cu 37% mai puțin decât în 2014), dintre care 3 decese. Toate cazurile din 2015 au avut sursa de agent patogen pe continentul african, în special în Guineea Ecuatorială (6 cazuri) și Nigeria (6 cazuri), iar Plasmodium falciparum a fost
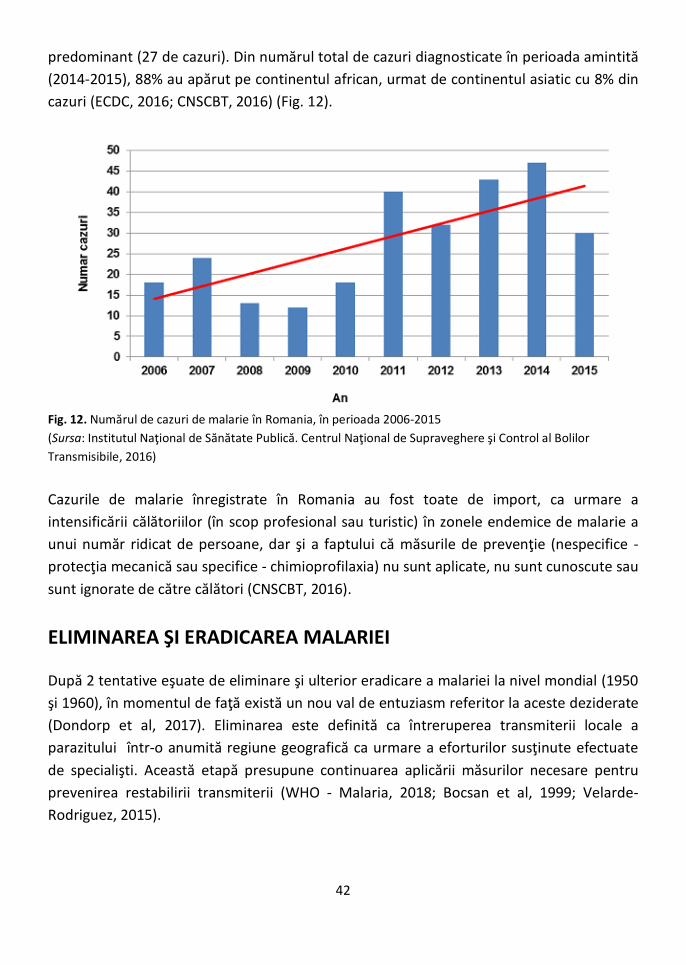
42
predominant (27 de cazuri). Din numărul total de cazuri diagnosticate în perioada amintită (2014-2015), 88% au apărut pe continentul african, urmat de continentul asiatic cu 8% din cazuri (ECDC, 2016; CNSCBT, 2016) (Fig. 12).
Fig. 12. Numărul de cazuri de malarie în Romania, în perioada 2006-2015 (Sursa: Institutul Naţional de Sănătate Publică. Centrul Naţional de Supraveghere şi Control al Bolilor Transmisibile, 2016)
Cazurile de malarie înregistrate în Romania au fost toate de import, ca urmare a intensificării călătoriilor (în scop profesional sau turistic) în zonele endemice de malarie a unui număr ridicat de persoane, dar şi a faptului că măsurile de prevenţie (nespecifice - protecţia mecanică sau specifice - chimioprofilaxia) nu sunt aplicate, nu sunt cunoscute sau sunt ignorate de către călători (CNSCBT, 2016).
ELIMINAREA ŞI ERADICAREA MALARIEI
După 2 tentative eşuate de eliminare şi ulterior eradicare a malariei la nivel mondial (1950 şi 1960), în momentul de faţă există un nou val de entuziasm referitor la aceste deziderate (Dondorp et al, 2017). Eliminarea este definită ca întreruperea transmiterii locale a parazitului într-o anumită regiune geografică ca urmare a eforturilor susţinute efectuate de specialişti. Această etapă presupune continuarea aplicării măsurilor necesare pentru prevenirea restabilirii transmiterii (WHO - Malaria, 2018; Bocsan et al, 1999; Velarde-Rodriguez, 2015).

43
Eradicarea malariei reprezintă dispariţia tuturor cazurilor de boală (tipice sau atipice, manifeste sau subclinice) cauzate de Plasmodium, dar şi încetarea circulaţiei agentului patogen, ca urmare a eforturilor deliberate susţinute ale specialiştilor. Odată cu atingerea acestui obiectiv, măsurile intervenţionale nu mai sunt necesare (WHO - Malaria, 2018; Bocsan et al, 1999). Toate ţările care declară zero cazuri de boală 3 ani consecutiv sunt eligibile pentru a aplica la Organizaţia Mondială a Sănătăţii (OMS/WHO) în scopul obţinerii certificării eliminării malariei. În ultimii ani 7 ţări au atins acest obiectiv al certificării eliminării bolii: United Arab Emirates (2007), Morocco (2010), Turkmenistan (2010), Armenia (2011), Maldives (2015), Sri Lanka (2016) and Kyrgyzstan (2016). În perioada 2000 - 2015, 17 ţări au eliminat malaria (WHO - Malaria, 2018; Velarde-Rodriguez et al, 2015). "Strategia tehnică globală pentru malarie 2016-2030" a OMS - adoptată de World Health Assembly în mai 2015 - oferă un cadru tehnic pentru toate țările endemice pentru malarie. Acesta a fost conceput pentru a ghida și sprijini programele regionale și naționale care vizează controlul și eliminarea malariei. Strategia stabilește obiective globale ambițioase, dar realizabile, printre care: reducerea incidenței malariei cu cel puțin 90% până în 2030; reducerea ratelor de mortalitate cauzate de malarie cu cel puțin 90% până în 2030; eliminarea malariei în cel puțin 35 de țări până în 2030; prevenirea reemergentei malariei în toate țările fără malarie. Această strategie a fost rezultatul unui amplu proces de consultări care a avut loc în decurs de 2 ani și a implicat participarea a peste 400 de experți tehnici din 70 de state membre. Strategia se bazează pe trei piloni cheie: asigurarea accesului universal la prevenirea, diagnosticarea și tratarea malariei; accelerarea eforturilor pentru eliminarea bolii și obținerea statutului de țară fără malarie; și transformarea supravegherii malariei într-o intervenție de bază (WHO - Global technical strategy for malaria 2016-2030, 2015). În anii 1950-1960 campaniile de eradicare și control a malariei au fost un succes în țările cu climă temperată sau cu un număr redus de cazuri de malarie. Apariția rezistenței la insecticide a țânțarilor și la chlorochine a parazitului malariei au pus capăt epocii de eradicare a malariei, la aceasta contribuind şi infrastructura slabă din regiunile cu malarie, precum și diminuarea resurselor pentru cercetarea malariei (Lines, 1996; Carter and Mendis, 2002; Korenromp et al, 2003; ter Kuile et al, 2004). Conferința internațională din 1992, a restabilit controlul malariei ca prioritate de sănătate la nivel mondial. OMS a declarat în 2008 că sărăcia ar fi principalul factor de risc pentru malarie (Lucas and

44
McMichael, 2005). Finanțarea prevăzută pentru proiectul pe 2 ani (1997-1998) a reprezentat de 12 ori mai mult decât contribuția făcută de OMS în deceniul anterior. În 1998 campania Roll Back Malaria, a lansat parteneriate privind tehnicile de bază în strategiile de control a malariei. În 2000 Națiunile Unite au declarat 2001-2010 deceniul pentru Roll Back Malaria în țările în curs de dezvoltare, în special în Africa. În 2001 resursele pentru controlul malariei, au fost amplificate prin crearea Fondului Global pentru combaterea HIV, tuberculozei și malariei. Malaria a fost eliminată în 5 țări din Africa de nord, Tunisia, Libia, Egipt, Maroc, Algeria, unde malaria a fost produsă cel mai frecvent de P. vivax și transmisă de specii de țânțari ce au fost mai ușor de controlat; dar riscul reintroducerii malariei în aceste țări este continuu. În est, malaria este endemică în Madagascar, în insulele Comore (Republica Federală Islamică a Insulelor Comore) și colectivitatea franceză dinTeritoriul Mayotte și Zanzibar. În Mauritius, malaria a fost bine controlată din 1950, dar focarele ocazionale de malarie, cu specia P. vivax apar, ultimul ca urmare a unui ciclon în 1982. Din acel an a existat o scădere constantă a cazurilor și riscul este acum extrem de scăzut. Seychelles a fost indemnă de malarie din anul 1930, iar despre vectorii malariei se crede că nu mai există acolo (WHO/UNICEF - The Africa Malaria Report, 2003). Un studiu efectuat în Africa, în vestul Keniei, a arătat că principalul vector al malariei îl constituie Anopheles gambiae (Fillinger et al, 2004), iar reducerea mortalității la copii în această zonă s-a realizat cu ajutorul plaselor tratate cu insecticide (Nevill et al, 1996). În Asia, malaria reprezintă o problemă majoră în subregiunea Mediteranei de Est,în special în zonele în care, în ultimii 30 de ani, au existat situații de urgență din cauza distrugerii sistemelor de sănătate. Începând cu 1998-1999, finanţările regionale pentru controlul malariei au crescut. Un studiu realizat în 2007 a arătat eficacitatea uleiului de neem (copac din Asia de vest) ca larvicid în controlul malariei (Okumu et al, 2007).
Având în vedere rezistența la insecticidele sintetice formată în ultimii ani și efectul negativ al acestora asupra mediului, folosirea uleiului de neem poate constitui o variantă ieftină de control al larvelor. Folosit la diferite concentrații acesta s-a dovedit a fi un bun larvicid, astfel concentrația letală medie [LC (50)] din compoziția împotriva Anopheles stephensi, Culex quinquefasciatus și Aedes aegypti a fost identificată ca fiind 1,6, 1,8 și 1,7 ppm (Dua et al, 2009). Strategiile principale de control sunt accesul rapid la un tratament eficace şi folosirea plaselor tratate cu insecticid și biolarvicide (Mittal, 2003; WHO, 2006). Malaria cu P. vivax este endemică în Asia Centrală și Transcaucazia, iar malaria cu P. falciparum a reapărut în

45
Tadjikistan în 1990, astfel că începând din 2002 s-au organizat strategii de control a vectorilor. S-a reușit un control al epidemiilor de malarie, deşi raportul din 2003 arată o incidență a malariei de 10 ori mai mare decât în 1990. Asia de Sud-Est este regiunea cu rezistența cea mai mare a malariei la medicamente din lume, contribuind la reemergenţa malariei în multe zone, în special de-a lungul frontierelor internaționale. Toate țările din Asia de Sud-Est au adoptat strategii de control prin utilizarea plaselor tratate cu insecticide și larvicide. În Indonezia și Sri Lanka s-au înregistrat rezultate foarte bune, reducând incidența malariei la cel mai scazut nivel după 1967. Sub-regiunea Pacificului de Vest, unde malaria a devenit reemergentă ca urmare a declinului economic, a brasajului populațional pe scară largă și a deficiențelor din sistemul sanitar, strategiile de control implementate au dat rezultate bune. Transmiterea malariei se înregistrează în opt țări din America Centrală și Caraibe, și zona pădurilor tropicale din Amazon, datorată mișcărilor populației, mineritului și silviculturii. Toate aceste zone au adoptat strategii de control bazate pe utilizarea plaselor tratate cu insecticide și larvicide, reușindu-se ținerea sub control a epidemiilor de malarie. Utilizarea plaselor tratate cu insecticid a apărut ca o necesitate în urma rezistenței dobândite de Pl. falciparum la chloroquină (Pietra et al, 1991). Clorochina și-a păstrat eficacitatea pentru tratamentul și profilaxia malariei cu P. falciparum în America Centrală în nordul Canalului Panama, Republica Dominicană și Haiti, precum și pentru tratarea malariei cu P. vivax în întreaga regiune. O nouă strategie de creare a medicamentelor noi este folosirea de molecule hibride care permit producerea de molecule antipaludice cum sunt trioxaquinele, care sunt active asupra unor sușe de agenți infecțioși rezistenți la medicamentele clasice (Meunier and Robert, 2010). Primaquina este singurul medicament cu efect asupra formelor hepatice (hipnozoiți) dar folosirea ei este controversată pentru că produce hemoliză la persoanele cu deficit de glucoză-6-fosfat-dehidrogenază (G6PD) (Oliver et al, 2008; Bray et al, 2005. Primaquina poate fi utilizată profilactic în timpul călătoriilor și după întoarcerea din călătorii, precum și ca profilaxie la persoanele supuse unui risc ridicat de infecție cu P. vivax. De asemenea, se administrază primaquină persoanelor care au malarie cu P. vivax sau P. ovale, pentru a se evita riscul de recidivă mai mult sau mai puțin întârziată datorită hipnozoiților (Baird and Rieckmann, 2003; Baird et al, 2003; Baird and Hoffman, 2004; Ajdukiewicz, Ong, 2007; Carmona-Fonseca and Maestre, 2009). De exemplu, în Chimoio, Mozambique, între 2010 şi 2014, s-a înregistrat o tendinţă descendentă a ratelor de mortalitate, explicată parţial de utilizarea plaselor împotriva ţânţarilor tratate cu insecticid (Fig. 13) (Ferrão et al. 2017).

46
Fig. 13. Tendinţa mortalităţii prin malarie în Chimoio, Mozambique, pe luni şi ani (Sursă: Ferrão et al. 2017) Plățile internaționale pentru controlul malariei au crescut în ultimii opt ani, fiind estimate la 1,66 miliarde de dolari în 2011 și 1,84 miliarde de dolari în 2012. Finanțarea de la guvernul național a crescut în 2011 la 625 milioane de dolari. Cu toate acestea, fondurile disponibile în prezent pentru prevenirea și controlul malariei, sunt mult sub resursele necesare pentru a atinge obiectivele globale în ceea ce privește malaria. Un procent estimat de 5.1 miliarde dolari sunt necesari în fiecare an între 2011 și 2020, pentru a realiza accesul universal la intervenții în caz de malarie. În 2011, doar 2,3 miliarde dolari au fost disponibile, mai puțin de jumătate din ceea ce este necesar. Malaria rămâne totuși legată de sărăcie, rata de mortalitate cea mai ridicată, fiind înregistrată în țările cele mai sărace din Africa. Astfel obiectivele globale pentru controlul cazurilor și al deceselor datorate malariei, nu vor putea fi atinse decât dacă se înregistrează progrese în cele 17 țări luate în calcul, unde se înregistrează 80% din cazurile de malarie. Nigeria, Republica Democratică Congo, Republica Unită Tanzania, Uganda, Mozambic și Coasta de Fildeș reprezintă țările cu un număr estimat de 103 milioane (47%) de cazuri de malarie. În Asia de Sud-Est, a doua cea mai afectată de malarie regiune, India ocupă primul loc cu 24 milioane de cazuri pe an, urmată de Indonezia și Myanmar. Sistemele de supraveghere a malariei pot detecta aproximativ 10% din numărul total de cazuri de malarie la nivel mondial, în 41 de țări din întrega lume, neputându-se face o estimare fiabilă din cauza rapoartelor incomplete. Rezistența la medicamente a malariei este de interes global în lupta pentru controlul malariei cu P. falciparum (Girod et al,

47
2006). În patru țări din Asia de Sus-Est a fost raportată rezistența la artemisinină și în 64 de țări s-a raportat rezistența la insecticide. Fondurile alocate pentru controlul malariei au avut o creştere între 2004 și 2009 cu rezultate foarte bune, dar care s-au diminuat pentru perioada 2010-2011, ceea ce ar putea duce la pierderea câștigurilor de până acum, permiţând malariei un nou avânt. Astfel numărul plaselor tratate cu insecticide, pentru Africa sub-sahariană, a scăzut de la 145 milioane în 2010, la 66 milioane în 2012. Măsurile luate în ultimii opt ani, pentru controlul malariei, au dus la evitarea a 1 milion de decese. Nigeria și Republica Democrată Congo sunt cele mai afectate țări din Africa sub-sahariană, iar India este cea mai afectată din Asia de Sud-Est. Raportul privind malaria la nivel mondial din 2012 indică faptul că finanțarea internațională pentru malarie pare să fi ajuns la un platou cu mult sub nivelul necesar pentru a ajunge la atingerea obiectivelor stabilite. Un procent estimat de 5,1 miliarde de dolari sunt necesari în fiecare an, între 2011 și 2020, pentru a realiza intervențiile propuse în cele 99 de țări cu transmitere a malariei. În timp ce multe țări au crescut finanțarea internă pentru controlul malariei, finanțarea totală disponibilă la nivel mondial a rămas la nivelul de 2,3 miliarde de euro în 2011 - mai puțin de jumătate din ceea ce este necesar (WHO World Malaria Report, 2012).
Fig.14. Numărul estimat de decese cauzate de malarie, 2002-2012 (Sursa: https://www.statista.com/chart/1758/estimated-number-of-deaths-caused-by-malaria-worldwide/)

48
PATOGENEZĂ Malaria se dezvoltă în două faze: una care implică ficatul (faza exoeritrocitară) și una care implică globule roșii sau eritrocitele (fază eritrocitară). Când un țânțar infectat străpunge pielea unei persoane pentru a lua o masă de sânge, sporozoiţii din saliva țânțarilor intră în sânge și migrează spre ficat, unde infectează hepatocitele, înmulțindu-se asexuat și asimptomatic pentru o perioadă de 8-30 zile (Bledsoe GH, 2005). După o potențială perioadă de latenţă în ficat, aceste organisme se diferențiază pentru a da mii de merozoiți, care, după ruperea celulelor gazdă, scapă în sânge și infectează celulele roșii din sânge pentru a începe etapa eritrocitară a ciclului de viață (Bledsoe GH, 2005). Parazitul scapă din ficat nedetectat prin împachetarea în membrana celulară a celulei hepatice gazdă infectate (Vaughan AM et al, 2008). În interiorul celulelor roșii din sânge, paraziți se înmulțesc în continuare, din nou asexuat, evadand periodic din celulele gazdă pentru a invada celulele roșii sanguine tinere noi. Există mai multe astfel de cicluri de amplificare. Astfel, descrierile clasice ale valurilor de febră apar ca urmare a valurilor simultane de merozoiți care scapă și infectează celulele roșii din sânge (Bledsoe GH, 2005). Unii sporozoiţi ai P. vivax nu se dezvoltă imediat în merozoiţi, în fază exoeritocitară, ci produc hipnozoiţi care rămân latenți pentru perioade care variază de la câteva luni (7-10 luni în mod tipic) până la câțiva ani. După o perioadă dormantă, se reactivează și produc merozoiți. Hipnozoitii sunt responsabili de incubarea îndelungată și de recăderile tardive în cazul infecțiilor cu P. vivax (Richter et al, 2010), totuşi existența lor în infecţia cu P. ovale este incertă (White, 2011). Parazitul este relativ protejat de atacul sistemului imunitar al organismului deoarece pentru cea mai mare parte a ciclului său de viață uman se află în ficat și în celulele sanguine și este relativ invizibil pentru supravegherea imună. Cu toate acestea, celulele sanguine infectate care circulă sunt distruse în splină. Pentru a evita această soartă, parazitul P. falciparum exprimă proteine adezive pe suprafața celulelor sanguine infectate, determinând celulele sanguine să rămână ataşate la pereții vaselor mici de sânge, prin aceasta stopând parazitul de la trecerea prin circulația generală și splină (Tilley et al, 2011). Blocarea microvasculară provoacă simptome precum cele din malaria placentară (Mens et al, 2012). Celulele roșii din sânge sechestrate pot să traverseze bariera hemato-encefalică și să determine malaria cerebrală (Rénia et al, 2012).

49
MANIFESTĂRI CLINICE De obicei, pacienții cu malarie devin simptomatici la câteva săptămâni după infectare, dar expunerea anterioară a gazdei sau imunitatea gazdei împotriva malariei ar putea influența simptomatologia și perioada de incubație, care este tipică pentru fiecare specie Plasmodium. Un simptom important este febra tipică recurente (care poate fi periodică la 48 sau 72 de ore, în funcție de specie) (Fig. 15).
Fig.15. Simptomul malariei – febra recurentă tipică (Sursa: https://www.dailystar.co.uk/health/592904/Malaria-symptoms-tablets-treatment-prevention Paroxismul clasic începe cu o perioadă de frisoane care durează aproximativ 1-2 ore și sunt urmate de febră mare. Traspiratiile excesive cu care se confruntă pacientul sunt urmate de scăderea temperaturii corporale la normal sau sub normal. Practic toți pacienții cu malarie vin cu cefalee. Alte simptome includ oboseală, stare de rău, greață și vărsături, diaree, tuse, dificultăți de respirație, artralgie și mialgie (Dondorp et al, 2017; Nadjm et al, 2012; Bartoloni and Zammarchi, 2012) (Fig. 16, 17).
Fig.16. Simptomele malariei (Sursa: https://www.lybrate.com/topic/symptoms-of-malaria-and-chikunguniya/ dd771f92ae9155433ef28c1ecc43adc7)

50
Mulți pacienți, în special în timpul infecției precoce, nu prezintă paroxismul clasic, dar pot avea câteva mici vârfuri febrile pe zi. Periodicitatea febrei este specifică pentru fiecare specie (de exemplu 48 de ore pentru P. falciparum, P. vivax și P. ovale - așa-numita febră terțiană, 72 de ore pentru P. malariae - denumită și febră quartană), dar nu este evidentă în timpul infecția inițiale din cauza multiplelor elemente tinere care apare în sânge și adesea nu este observată în infecțiile cu P. falciparum. Pacienții cu infecții sincrone de lungă durată sunt mai predispuși să prezinte modele clasice de febră. În cazurile severe, de obicei cauzate de P. falciparum, pot apărea următoarele simptome: icter, anemie, insuficiență renală, șoc, sindrom de detresă respiratorie la adult, confuzie, comă, encefalopatie și acidoză (Dondorp et al, 2017; Nadjm et al, 2012; Bartoloni and Zammarchi, 2012) (Fig. 17).
Fig.17. Simptomele malariei (Sursa: http://zeenews.india.com/health/how-to-fight-dengue-malaria-flu-more-symptoms-prevention-2046312 Organizația Mondială a Sănătății (OMS) recomandă ca femeile gravide să evite călătoriile în zonele cu risc de malarie (WHO Information for travellers, 2018). Dacă se contracteză malaria în timpul sarcinii, mama şi copilul prezintă un risc crescut de a dezvolta complicaţii grave, cum ar fi: naştere prematură (naştere înainte de 37 de săptămâni de sarcină), greutate mică la naştere, creșterea limitată a copilului în uter, naşterea unui copil mort, avort spontan şi chiar moartea mamei.

51
DIAGNOSTIC Malaria trebuie recunoscută cu promptitudine pentru a trata pacientul în timp eficient și pentru a preveni răspândirea infecției în comunitate prin intermediul țânțarilor locali. Întârzierea diagnosticului și a instituirii tratamentului reprezintă o cauză principală de deces la bolnavii infestaţi cu malarie din Statele Unite. Malaria poate fi suspectată pe baza istoricului de călătorie al pacientului într-o zonă endemică de malarie, a simptomelor și a constatărilor la examenul clinic. Cu toate acestea, pentru ca diagnosticul să fie unul de certitudine ar trebui făcute, teste de laborator care demonstrezează prezenţa paraziților malariei. În cazul în care malaria nu mai este endemică într-o ţară, medicii ar putea să nu fie familiarizați cu boala. Clinicienii care văd un pacient cu malarie pot trece cu vederea considerarea malariei printre posibilele diagnostice și să nu comande testele de diagnostic necesare. Laboratorul poate să nu aibă experiență în ceea ce privește malaria și să nu detecteze paraziți la examinarea frotiurilor de sânge la microscop. DIAGNOSTICUL CLINIC Istoricul pacientului trebuie să includă întrebări despre: deplasarea recentă sau la distanță către o zonă endemică; statutul imunitar, vârsta și starea sarcinii; alergii sau alte afecțiuni medicale; medicamentele administrate în prezent. Majoritatea pacienților cu malarie nu au constatări fizice specifice, dar poate fi prezentă splenomegalia. Simptomele malariei sunt nespecifice și se pot manifesta ca o afecțiune asemanatoare gripei cu febră, cefalee, stare de rău, oboseală și dureri musculare. Unii pacienți cu malarie prezintă diaree și alte simptome gastro-intestinale. Persoanele imune pot fi complet asimptomatice sau pot prezenta o anemie ușoară. Subiectii non-imuni pot deveni rapid foarte bolnavi. În general, totuși, apariția periodicității febrei nu este un indiciu fiabil pentru diagnosticarea malariei. Malaria severă implică în primul rând infecția cu P. falciparum, deși moartea datorată rupturii splenice a fost raportată si la pacienții cu malarie non-P. falciparum. Malaria severă include malarie cerebrală (uneori cu comă); anemie severă; anomalii ale aparatului respirator (cu acidoză metabolică, detresa respiratorie asociată și edem pulmonar); semnele sindromului hiperpneumatic malaric (arsuri alarice, retragerea cutiei toracice, utilizarea mușchilor auxiliari pentru respirație și respirația anormal de profundă); insuficiență renală (de obicei reversibilă) (Dondorp et al, 2017; Bartoloni and Zammarchi, 2012). La copii, malaria are o evoluție mai scurtă, de multe ori progresând rapid până la

52
malarie severa. Copiii sunt mai predispuși să prezinte hipoglicemie, convulsii, anemie severă și moarte subită, dar sunt mai puțin susceptibili de a dezvolta insuficiență renală, edem pulmonar sau icter (Bartoloni and Zammarchi, 2012). În unele zone endemice ale malariei, transmiterea este atât de intensă încât o mare parte din populație este infectată, dar nu se îmbolnăvește de paraziți. Astfel de purtători s-au dezvoltat doar o imunitate suficientă pentru ai proteja de malarie, dar nu de infecțiile malariei. În această situație, găsirea de paraziți ai malariei la o persoană bolnavă nu înseamnă neapărat că boala este cauzată de paraziți. Malaria este suspectată clinic în primul rând pe baza prezenţei febrei sau a antecedentelor de febră. Tratamentul antimalaric ar trebui să fie limitat la cazurile cu teste pozitive, iar pacienții cu rezultate negative ar trebui verificaţi din nou pentru alte cauze comune de febră și trataţi în mod corespunzător. La pacienții suspecţi de malarie severă, precum și la grupurile cu risc ridicat, cum ar fi pacienții cu HIV/SIDA, absența sau întârzierea diagnosticului parazitologic nu ar trebui să oprească începerea imediată a tratamentului antimalaric. DIAGNOSTICUL DE LABORATOR Următoarele investigații de sânge trebuie efectuate:
• cultura de sange, concentrația hemoglobinei, numărul de trombocite, investigarea funcției hepatice și renale, concentrațiile de electroliți (în special sodiu), monitorizarea parametrilor care sugerează hemoliza (haptoglobină, dehidrogenază lactică, numărul de reticulocite);
• în cazuri selectate, se recomandă testarea rapidă a HIV; • numărul de celule albe din sânge: leucocitoza este prezentă la mai puțin de 5%
dintre pacienții cu malarie (în acest caz diagnosticul diferențial trebuie extins); • nivelul G6PD trebuie verificat dacă pacientul trebuie tratat cu primaquină; • nivelul de glucoză trebuie evaluat pentru a exclude hipoglicemia, dacă pacientul are
malarie cerebrală. Următoarele investigații imagistice pot fi luate în considerare:
• radiografie toracică, dacă sunt prezente simptome respiratorii; • Computer Tomografie cerebrală, dacă sunt prezente simptome ale sistemului
nervos central. Ar trebui efectuate teste specifice pentru malarie:
• centrifugarea microhematocritului (sensibilă, dar incapabilă de speciație); • coloranții fluorescenți / teste indicatoare ultraviolete (nu pot genera informații
despre specii);

53
• testele standard: frotiuri de sânge subțire (calitative) sau gros (cantitative) (1 frotiu negativ nu exclude malaria ca diagnostic și mai multe frotiuri trebuie examinate pe o perioadă de 36 de ore);
• alternativele la testarea frotiului de sânge (utilizate în cazul în care expertiza frotiului de sânge este insuficientă) sunt testele rapide de diagnostic, analiza PCR, amplificarea bazată pe secvența de acid nucleic (Bartoloni and Zammarchi, 2012; Greenwood et al, 2005).
Toți pacienții cu suspiciune de malarie ar trebui sa fie trataţi pe baza unui diagnostic confirmat prin examinare microscopică sau test de diagnosticare rapidă (RDT) (imunocromatografie) care urmăreşte detecţia antigenelor în sânge (Fig. 18). Totuşi, înainte de utilizarea pe scară largă a acestor teste rapide ar trebui îmbunătăţită acurateţea lor. Se recomandă ca toate RDT-urile să fie urmate de microscopie pentru a confirma diagnosticul și, dacă sunt pozitive, pentru a cuantifica proporția celulelor roșii din sângele infectat. Utilizarea acestor teste rapide poate reduce timpul necesar pentru diagnosticul malariei.
Fig. 18. Test Diagnostic Rapid pentru malarie (Sursa: http://adulldayatwork.blogspot.com/2011/10/25-october-2011-tuesday-rapid-malaria.html
Identificarea moleculară a parazitului malariei reprezintă metoda de diagnostic de certitudine, dar rezultatele testului PCR (Reacţie de Polimerizare în Lanţ) nu sunt întotdeauna disponibile suficient de repede pentru a fi de valoare în stabilirea diagnosticului de infecţie cu Plasmodium (Fig. 19). PCR-ul este cel mai util pentru confirmarea speciei de parazit după ce diagnosticul a fost stabilit fie prin microscopie în picătură groasă, fie prin RDT.

54
Fig. 19. Test PCR pentru identificarea malariei (Sursa: Ivanescu, 2015)
Serologia detectează anticorpi împotriva paraziților malariei, utilizând fie imunofluorescența indirectă (IFA), fie testul ELISA. Serologia nu detectează infecția curentă, ci măsoară mai degrabă expunerea din trecut. Paraziţii malariei pot fi identificați prin examinul microscopic hematologic (examenul de sânge periferic în picătură groasă) (Fig. 20).
Fig.20. Examenul de sânge în picătură groasă (Sursa: https://labmedicineblog.com/2018/06/19/hematology-case-study-a-51-year-old-woman-with-fever-and-chills/
Înainte de examinare, specimenul este colorat cel mai adesea cu coloraţie Giemsa (Fig. 21). Această tehnică rămâne standardul de aur pentru confirmarea de laborator a malariei. Totuși, depinde de calitatea reactivilor, de microscop și de experiența laboratorului.

55
Fig.21. Diagnosticul hematologic al malariei (Sursa: Personal collection)
Cazurile suspecte de malarie din Africa sunt din ce în ce mai mult verificate prin folosirea unui test rapid, înainte de prescrierea tratamentului antimalaric, asigurându-se astfel utilizarea eficientă a resurselor folosite pentru eliminarea paraziţilor, dar cauza principală a febrei individului rămâne totuşi necunoscută datorită potenţialului coinfecţiei cu o altă boală febrilă. Utilizarea pe scară largă a testelor rapide nu împiedică neapărat supraevaluarea cazurilor de malarie clinică sau gestionarea cazurilor sub-optimale a pacienților febrili. Într-o publicație recentă din jurnalul eLife, s-a prezentat o nouă abordare care permite deducerea prevalenței spațio-temporale atât a febrei malariei asociate cu Plasmodium falciparum, cât și a febrei nemalarice la copii din Africa sub-sahariană, din 2006 până în 2014. Se estimează că 35,7% din cazurile febrile raportate de autor au fost însoțite de infecţie malarică în 2014, dar numai 28,0% dintre acestea (10,0% din toate cazurile febrile) au fost cauzate de malarie. Cele mai multe cazuri febrile printre copiii cu test pozitiv pentru malarie sunt, prin urmare, cauzate de boli non-malarice (Fig. 22).

56
De asemene anemia sau scăderea concentrației hemoglobinei din sânge este frecvent asociată cu malaria. Anemia este foarte răspândită în Africa sub-sahariană, în special în rândul copiilor și femeilor de vârstă reproductivă, dar nu a fost cuantificată anterior pentru a stabili atât contribuția malariei, cât şi contribuţia altor cauze la instalarea anemiei (de exemplu, deficitul de fier alimentar, infecția helminților) (Dalrymple et al, 2017). În colaborare cu echipa ROAD-MAP, s-au colectat datele din sondajul făcut în gospodării în care respondenții sunt întrebați despre febra lor și istoricul tratamentului și primesc un diagnostic bazat pe paraziți malarie și măsurarea concentrației lor de hemoglobină din sânge. În plus, asociază datele sondajului cu variabilele sensibile la distanță colectate de ROAD-MAP, pentru a ajuta la predicția spațială a măsurătorilor de interes.
Fig.22. Proporția cazurilor de febră pozitivă pentru malarie din toate cazurile cu febră de toate cauzele, la copiii sub 5 ani (Sursa: https://elifesciences.org/articles/29198)
Astfel se poate s-a constatat că înainte de folosirea pe scară largă a testelor rapide, tratamentul antimalaric era administrat într-o proporţie mult mai ridicată, consemnându-se diagnosticul eronat al acestei boli. Mai mult, nu toți oamenii care fac malarie solicită tratament în instituțiile publice de sănătate. Unii primesc tratament în unități care nu raportează cazurile de malarie cum ar fi instituțiile medicale private, iar o parte dintre ei nu se tratează . Acest lucru contribuie la sub-raportarea cazurilor de malarie. Înaintea administrării tratamentului se recomandă testele de rezistență la medicament ce sunt efectuate în laboratoare specializate pentru a

57
evalua susceptibilitatea la compușii antimalariali ai paraziților recoltaţi de la un anumit pacient. Există două metode de laborator principale:
• teste in vitro: paraziții sunt crescuți în cultură în prezența concentrațiilor crescânde de medicamente; concentrația medicamentului care inhibă creșterea parazitului este utilizată ca punct final.
• caracterizarea moleculară: markerii moleculari evaluați prin PCR sau secvențierea genetică permit, de asemenea, prezicerea, într-o oarecare măsură, a rezistenței la unele medicamente.

58
PROFILAXIE În 2016 guvernele ţărilor endemice de malarie şi a pertenerilor internaţionali, au investit peste 2,7 miliarde de dolari în lupta pentru controlul şi eradicarea malariei. Regiunea africană acaparează 74% din buget, procentajele scăzând pentru regiunile din Asia de Sud-Est (7%), Mediterana de Est și America (fiecare 6%) și Vestul Pacificului (4%). Guvernele ţărilor endemice au contribuit doar cu 800 milioane USD, ceea ce reprezintă doar 31% din suma investită, arătând că fără implicarea partenerilor din ţările non-endemice lupta contra eradicării malariei ar fi din start pierdută (Statele Unite ale Americii (SUA) a oferit un miliard de dolari (38%), urmând cu o contribuţia importantă şi Regatul Unit al Marii Britanii și Irlanda de Nord (Marea Britanie), precum și Franța, Germania și Japonia (WHO - World Malaria Report, 2017). La reuniunea anuală din 2010, "Strategia tehnică globală pentru malarie 2016-2030", a fost stabilit un buget anual până în anul 2020 pentru a reduce incidența cazurilor de malarie și a ratei mortalității cu cel puțin 40%, un buget care ajunge la suma de 6,5 miliarde de dolari pe an. Acest deficit este ingrijorator deoarece nu este suficient nici pentru menținerea nivelului atins până acum. Prevenţia de primă linie o constituie apărarea împotriva înţepăturii ţânţarului din genul Anopheles. Măsurile de profilaxie constau în dormitul sub plase tratate cu insecticid având o remanenţă de lungă durată și utilizarea de îmbrăcăminte de protecție și repelenţi împotriva insectelor. În anul 2016 ponderea gospodăriilor care folosesc plase tratate cu insecticide, a crescut la 80% faţă de 50% în 2010; dar periodic se produce un declin, atunci când ţările schimbă sau rotesc insecticidele folosind unele mai scumpe. Pulverizarea reziduală a interioarelor cu insecticide este o modalitate puternică de a reduce rapid transmiterea malariei. Potențialul său se realizează atunci când cel puțin 80% din casele din zonele vizate sunt pulverizate. Stropirea interioară este eficientă timp de 3-6 luni, în funcție de insecticidul utilizat și de tipul suprafeței pe care este pulverizată. În unele situații, sunt necesare mai multe runde de pulverizare pentru a proteja populația pentru întreg sezonul de malarie.

59
În funcţie de riscul transmiterii malariei în zona care urmează să fie vizitată şi de speciile de Plasmodium existente, călătorii pot de asemenea lua medicaţie profilactică (chimioprofilaxie) înainte, în timpul și la întoarcerea din călătorie. Unele categorii de călători, în special copiii mici, femeile însărcinate și persoanele cu un sistem imunitar slăbit, prezintă un risc crescut de a dezvolta forme grave, în cazul contactării bolii. La femeile gravide, malaria creşte riscul de deces matern, avort spontan, naşterea unui copil mort sau greutate mică la naştere, precum şi riscul asociat de deces neonatal. Din această cauză femeile însărcinate ar trebui să evite călătoriile în zonele endemice de malarie, iar părinții sunt sfătuiți să nu ia sugari sau copii de vârstă mică în zonele în care există riscul de infestare cu tulpina P. falciparum. Când călătoriile nu poate fi evitate, este foarte important să se ia măsuri de prevenţie eficiente împotriva malariei, chiar și atunci când călătoresc în zonele cu transmitere a malariei cauzate de P. vivax. În 2012, OMS a recomandat chemoprevența sezonieră a malariei ca o strategie suplimentară de prevenire a malariei pentru zonele din subregiunea Sahel din Africa. Strategia implică administrarea lunară de amodiaquine plus sulfadoxină-pirimetamină tuturor copiilor cu vârsta sub 5 ani în timpul sezonului cu transmitere ridicată.
Fig.23. Plase împotriva ţânţarilor tratate cu insecticide cu effect de lungă durată (Sursa: http://www.who.int/immunization/programmes_systems/interventions/malaria_llins/en/)

60
Se recomandă consultarea centrelor naționale de combatere a bolii, sau a altor instituții care pot furniza informații cu privire la măsurile prevenţie ce rebuiesc luate, înaintea călătoriei în ţările sau regiunile endemice de malarie. Prevenirea sau întârzierea apariţiei rezistenței la medicamente este esențială pentru succesul ambelor strategii naționale și globale de control și eventual de eradicare a malariei. Pentru a contribui la protejarea unor medicamente antimalarice actuale și viitoare, toate episoadele de malarie ar trebui tratate cu terapie combinată, și medicamentele antimalarice trebuie administrate în doze optime. Toate cazurile de suspiciune de malarie ar trebui confirmate printr-un test rapid de diagnosticare (RDT) sau test parazitologic. CONTROLUL VECTORILOR O mare parte din succesul în combaterea malariei se datorează controlului vectorilor. Controlul vectorial depinde în mare măsură de utilizarea piretroidelor, care sunt singura clasă de insecticide recomandată în prezent. În ultimii ani, rezistența țânțarilor la piretroizi a apărut în multe țări. În unele zone, a fost detectată rezistența la toate cele 4 clase de insecticide utilizate pentru sănătatea publică. Din fericire, această rezistență a fost asociată doar rareori cu scăderea eficacității în cazul utilizării plaselor de ţânţari tratate cu insecticide cu eficienţă îndelungată, care continuă să ofere un nivel substanțial de protecție în majoritatea situațiilor. Utilizarea prin rotație a diferitelor clase de insecticide utilizate în stropirea interioarelor este recomandată ca o abordare pentru gestionarea rezistenței insecticidelor. Cu toate acestea, zonele endemice de malarie din Africa sub-sahariană și India provoacă îngrijorări semnificative din cauza nivelurilor ridicate de transmitere a malariei și a raportărilor cu privire la răspândirea zonelor de rezistență la insecticide. Utilizarea a 2 insecticide diferite într-o plasă de țânțari oferă o oportunitate de reducere a riscului de dezvoltare și răspândire a rezistenței la insecticide; dezvoltarea acestor plase noi este o prioritate. Detectarea rezistenței la insecticide ar trebui să fie o componentă esențială a tuturor eforturilor naționale de control al malariei pentru a se asigura că se utilizează cele mai eficiente metode de control vectorial. Alegerea insecticidului pentru IRS ar trebui întotdeauna să fie informată de date recente, locale despre susceptibilitatea vectorilor țintă.

61
Ţânţarii aparţinând genului Anopheles se găsesc răspândiţi în toate lumea cu excepţia zonei arctice, speciile anopheline găsindu-se nu doar în zonele endemice de malarie ci şi în zonele unde malaria a fost eradicată. Pentru a evita riscul răspândirii unor boli vectoriale, trebuie controlat portajul vectorilor; astfel trebuie împiedicat transportul ţânţarilor prin comerţul cu pneuri sau containere. De asemenea persoanele (turişti şi imigranţi) infectaţi cu agenţi patogeni transmişi prin vectori trebuie diagnosticaţi şi trataţi de urgenţă, bolile transmise de tantari necunoscând granite, iar raspandirea lor este si o consecinta a mobilitatii si a globalizarii. Astfel că mobilitatea crescută a oamenilor și comerțul internațional joacă un rol mai important în diseminarea vectorilor și a agenților lor patogeni / paraziți decât creșterea temperaturilor (Yi H et al, 2014). Pentru perioada 2010-2016 datele furnizate din 76 de ţări endemice de malarie, arată că în 61 de ţări s-a raportat rezistenţa la cel puţin un insecticid. În 50 de ţări a fost raportată rezistenţa la două sau mai multe clase de insecticide. Rezistenţa la piretroide-singura clasă de insecticide utilizată în momentul de faţă în tratarea plaselor pentru ţânţari, este larg răspândită în toate zonele, în 2016 crescând procentajul la 81% a ţărilor ce au raportat rezistenţă la insecticide, faţă de 71% în 2010 (WHO - World Malaria Report, 2017). În ciuda rezistenţelor semnalate, plasele tratate cu insecticide rămân cel eficient instrument în lupta pentru prevenirea malariei, chiar şi în zonele unde s-au raportat rezistenţe la piretroide. PREVENŢIA MALARIEI LA OM Prevenţia şi controlul malariei include la acest moment trei direcţii:
• reducerea contactului între vector şi gazda umană, • prevenţia specială cu antimalarice şi • diagnosticul precoce şi tratamentul adecvat al episoadelor de malarie pentru
limitarea transmiterii (Heymann, 2012; Dondorp, 2017). PREVENŢIA GENERALĂ - MINIMIZAREA CONTACTULUI ÎNTRE GAZDA UMANĂ ŞI ŢÂNŢAR O metodă foarte importantă de prevenţie generală a malariei în zonele endemice este reprezentată de utilizarea plaselor de ţânţari tratate cu insecticide (ITN). Acestea necesită retratare după 3 spălări sau măcar o dată pe an. O variantă îmbunătăţită sunt plasele de ţânţari tratate cu insecticide cu durată lungă de acţiune (LLIN) care au încorporate în structura lor substanţe piretroide (Heymann, 2012; Ouattara et al, 2014). Noi speranţe se

62
conturează referitor la utilizarea unor plase de ţânţari cu două insecticide pentru scăderea riscului de apariţie a rezistenţei (Heymann, 2012). O altă metodă de prevenţie generală este pulverizarea cu insecticide reziduale în interior (IRS), care vizează populaţia adultă de ţânţari, şi are o rată ridicată de succes când sunt sprayate peste 80% din casele din zonă (WHO - Malaria, 2018; Heymann, 2012). Această metodă, care constituie o intervenţie de sănătate publică, are eficacitate maximă atunci când ţânţarii rămân în interior pe suprafeţe care pot fi pulverizate, atunci când persoanele sunt expuse în casă sau în aproprierea casei, când se aplică înainte de începerea sezonului de transmitere sau înainte de perioada de vârf a transmiterii (Heymann, 2012; Salam et al, 2014). În prezent există 12 insecticide recomandate de OMS pentru a fi utilizate în cadrul acestei măsuri, alegerea acestor substanţe fiind dependentă de susceptibilitatea insecticidului, particularităţile vectorului, de gradul de siguranţă pentru oameni şi mediu, de eficacitate şi de raportul cost-eficienţă. DDT-ul este încă utilizat pentru controlul speciilor de Anopheles, în acord cu Convenţia de la Stockholm privind poluanţii organici persistenţi, care interzic utilizarea DDT-ului, cu excepţia scopului de asigurare a sănătăţii publice (Heymann, 2012). Controlul stadiilor larvare prin eliminarea locurilor de reproducere ale ţânţarilor are valoare limitată în majoritatea zonelor unde persistă transmiterea malariei. O astfel de măsură este reprezentată de completarea şi drenarea sau creşterea vitezei de curgere a apei în canale naturale sau artificiale. Metodele chimice sau biologice de control, precum larvicidele bacteriene sau peştele larvivorus, pot fi dificil de pus în aplicare în zonele rurale, dar în unele zone africane urbane au fost înregistrate succese. Aceste metode reprezintă metode adjuvante utile în zonele aride, de coastă şi urbane sau pot constitui metode necesare pentru menţinerea unui nivel scăzut de receptivitate în zonele în care a fost realizată eliminarea malariei (Heymann, 2012; Bocsan et al, 1999). Utilizare preventivă intermitentă a unei doze complete de antimalaric la intervale predefinite în trimestrele 2 şi 3 de sarcină constituie o măsură eficientă de reducere a malariei la femeile gravide în zonele cu transmitere stabilă a P. falciparum, cu intensitate moderată până la intensă, precum Africa. Metoda are utilitate limitată în alte zone ale lumii, în care transmiterea este instabilă şi de mică intensitate. De asemenea, în zonele endemice, pentru această categorie de persoane este utilă utilizarea ITN şi LLIN pentru a diminua efectele nocive ale malariei în timpul sarcinii (Heymann, 2012; Mpogoro et al, 2014).

63
O altă măsură de prevenţie generală este reprezentată de educaţia pentru sănătate a persoanelor care călătoresc în zonele endemice pentru a cunoaşte regiunile în care pot fi expuse riscului, care sunt măsurile de protecţie care trebuie aplicate şi care este conduita în cazul apariţiei unui sindrom febril după întoarcerea acasă (Luca, 2002).
I. Prevenţia specială prin utilizarea de medicamente antimalarice Chimioprofilaxia înainte de călătorie trebuie să includă recomandări de utilizare a medicamentelor anti-malarice potrivite pentru destinaţia aleasă (dacă ele există), după cum urmează (Heymann, 2012; Bocsan et al, 1999; Luca, 2002):
• În zonele cu risc foarte scăzut de transmitere a malariei, chimioprofilaxia poate să nu fie recomandată, luând în considerare faptul că riscul de apariţie a reacţiilor adverse asociate antimalaricelor poate depăşi potenţialele beneficii. Călătorii trebuie însă să conştientizeze posibilitatea existenţei malariei dacă dezvoltă o boală febrilă.
• Chimioprofilaxia cu clorochină poate fi tuilizată în zonele în care apare doar malaria cu P. vivax şi acele zone în care P. falciparum este sensibil la clorochină. Se recomandă 5 mg bază /Kg /săptămână în doză unică sau 10 mg bază /Kg /săptămână, divizată în 6 doze /zi (Heymann, 2012; Luca, 2002).
• În zonele în care există risc de transmitere a P. vivax şi P. falciparum şi există rezistenţă la clorochină, chimioprofilaxia cu clorochină trebuie combinată cu proguanil (3 mg/ Kg /zi).
• În zonele cu risc crescut pentru malaria cu P. falciparum şi rezistenţă raportată la medicamentele antimalarice, chimioprofilaxia de elecţie se face cu atovaquon-proguanil (250 mg atovaquon şi 100 mg proguanil /zi), doxiciclină (1,5 mg/ Kg /zi) sau meflochină (5 mg / Kg / săptămână). Aceste recomandări sunt valabile şi pentru zonele cu risc moderat sau scăzut de malarie cu P. falciparum, dar cu nivele ridicate de rezistenţă la preparatele antimalarice (Heymann, 2012; Luca, 2002).
• Utilizarea antimalaricelor cu administrare zilnică (atovaquon-proguanil, clorochină, doxiciclină, proguanil) trebuie începută cu o zi înainte de sosirea în zona cu risc. Referitor la clorochină, administrarea trebuie începută cu o săptămână înainte de sosire, iar pentru mefloquină, cu 2-3 săptămâni înainte de plecare, pentru atingerea unor nivele de anticorpi ridicate, eficiente şi pentru a detecta eventualele efecte adverse înainte de plecare.
• Administrarea preventivă trebuie să fie luată cu regularitate strictă pe toată durata călătoriei în zonele cu risc de malarie şi trebuie continuată timp de 4 săptămâni după ultima posibilă expunere (Heymann, 2012; Luca, 2002).

64
II. Prevenţia specifică – dezvoltarea de vaccinuri Deşi există promisiuni ale unui preparat vaccinal împotriva malariei, la momentul actual nu există un vaccin licenţiat cu utilizare populaţională. Cele mai avansate sunt cercetările pentru elaborarea unui vaccin candidat împotriva P. falciparum, preparat cunoscut sub denumirea de RTS, S/AS01 (un vaccin recombinant ce utilizează proteine din stadiul pre-eritrocitic). Acest vaccin a fost evaluat în cadrul unui trial clinic extins, derulat în 7 ţări din Africa şi a fost apreciat pozitiv de către Agenţia Europeană a Medicamentului (European Medicines Agency) în iulie 2015 (WHO - Malaria, 2018; Dondorp, 2017; Umeh et al, 2014). Eficacitatea vaccinului a fost de 53% (IC95% = 28-69%) împotriva malariei clinic manifeste la copiii cu vârstă între 5 şi 17 luni, evaluată pe o durată de 8 luni. Pentru copilul mai mare eficacitatea acestui preparat a fost mai redusă. Alte vaccinuri candidate includ vaccinul preparat din parazit întreg atenuat (attenuated whole-parasite vaccine), care asigură un nivel de protecţie ridicat, dar ridică probleme de producere şi administrare. Alte tipuri de vaccinuri candidate au ca ţintă stadiile parazitului care se găsesc în circulaţia sanguină sau stadiile asexuale ale Plasmodium (Dondorp , 2017). OMS susţine foarte puternic aceste studii pilot ca fiind un pas important în obţinerea primului vaccin antimalarie din lume, astfel încât în octombrie 2015, a recomandat implementarea de studii pilot pentru vaccinul RTS, S/AS01 într-un număr limitat de ţări africane. În noiembrie 2016, OMS a anunţat că siguranţa şi eficacitatea vaccinului RTS, S/AS01 vor fi evaluate în cadrul a două studii pilot (trialuri clinice de fază III) derulate în 3 ţări ale Africii sub Sahariene, care vor debuta la începutul anului 2018 (WHO - Malaria, 2018). Acest proiect pilot ar putea deschide calea pentru o mai ampla dezvoltare a vaccinului dacă siguranța și eficacitatea acestuia vor fi considerate acceptabile. PREVENŢIA RECIDIVELOR INFECŢIEI CU P.VIVAX SAU P.OVALE Pentru a preveni recidivele de malarie cauzată de P.vivax sau P.ovale, atât la copii, cât și la adulți (cu excepția femeilor însărcinate şi a sugarilor cu vârstă sub 6 luni) se recomandă administrarea de primaquină, timp de 14 zile (0,25-0,5mg / kg corp) în toate mediile de transmitere. La pacienţii cu deficit de G6PD, prevenirea recidivelor se face cu primaquină în doză de 0,75mg / kg corp o dată pe săptămână, timp de 8 săptămâni, sub supraveghere medicală strictă deoarece primaquina poate prezenta un potențial hemolizant (WHO - Guidelines for the Treatment of Malaria, 2015). Artemisina și derivatele sale nu trebuie să fie utilizată ca monoterapie pe cale orală, deoarece aceastea favorizează dezvoltarea rezistenței la artemisină.

65
OMS recomandă monitorizarea regulată a eficacităţii medicamentelor antimalarice utilizate pentru a se asigura că tratamentele alese rămân eficiente. Aceste acţiuni trebuie implementate prin programele naționale pentru controlul malariei. Rezistența la medicamentele antimalarice reprezintă o problemă recurentă. Rezistența P. falciparum la generațiile anterioare de medicamente, cum ar fi clorochina și sulfadoxina-pirimetamina (SP), a devenit larg răspândită în anii 1950 și 1960, subminând eforturile de control al malariei și scăzând şansele de supraviețuire la copilul. OMS recomandă monitorizarea de rutină a rezistenței la medicamentele antimalarice și sprijină țările să își consolideze eforturile în acest domeniu. În ultimii ani, rezistența parazitară la artemisinină a fost detectată în 5 țări din subregiunea Mekong: Cambodgia, Republica Democrată Lao, Myanmar, Thailanda și Vietnam. Studiile au confirmat că rezistența la artemisinină a apărut independent în multe zone ale acestor subregiuni. În 2013, OMS a lansat răspunsul de urgență la rezistența la artemisinină (ERAR) din subregiunea Mekong, un plan de atac la nivel înalt pentru a limita răspândirea paraziților rezistenţi la medicamente și pentru a oferi instrumente de salvare a populației expuse riscului de infestare cu malarie. În paralel, în unele situații s-a raportat rezistență crescută la medicamentele asociate ACT. O nouă abordare a fost necesară pentru a ține pasul cu schimbarea peisajului malariei. În consecință, Comitetul consultativ pentru politica în domeniul malariei al OMS din septembrie 2014 a recomandat adoptarea obiectivului de eliminare a malariei cu P. falciparum în această subregiune până în 2030. OMS a lansat Strategia pentru eliminarea malariei în subregiunea Greater Mekong (2015-2030) 2015, care a fost aprobată de toate țările din subregiune. Cu orientare tehnică din partea OMS, toate țările din subregiune au elaborat planuri naționale de eliminare a malariei. Împreună cu partenerii, OMS va oferi sprijin permanent pentru eforturile de eliminare a malariei în toate țările, prin programul de eliminare a malariei în Mekong, care este o nouă inițiativă care a evoluat din ERAR (WHO - Strategy for malaria elimination in the Greater Mekong Subregion: 2015-2030, 2015).

66
TRATAMENT Malaria este o boală care poate fi în întregime prevenită și tratată. Obiectivul principal al tratamentului este de a asigura eliminarea rapidă și completă a parazitului Plasmodium din sângele pacientului, cu scopul de a preveni progresia spre forme complicate de malarie, precum și pentru a preveni infecţia cronică care duce la anemie severă. Din perspectiva sănătății publice, scopul tratamentului este de a reduce transmiterea infecției la alte persoane, prin reducerea rezervoarelor infecțioase, și de a preveni apariția și răspândirea rezistenței la medicamentele antimalarice. La pacienții cu suspiciune de malarie, înainte de începerea tratamentului , trebuie să se confirme diagnosticul parazitologic, fie prin microscopie, fie prin test de diagnosticare rapidă. Tratamentul bazat doar pe manifestările clinice, trebuie administrat doar în cazul în care testele de diagnosticare nu sunt accesibile în decurs de 2 ore de la prezentarea pacienților pentru tratament. Pentru a preveni complicațiile care amenință viața, tratamentul cu un medicament antimalaric eficient și sigur trebuie utilizat în 24 de ore de la debutul febrei. Malaria este tratată cu medicamente antimalarice, dar medicamentele alese depind de tipul și severitatea bolii. De asemenea, medicamentele împotriva febrei sunt folosite în mod obișnuit în malarie, dar efectele lor asupra rezultatelor nu sunt foarte clare (Meremikwu et al, 2012; Amadi et al, 2015). Malaria simplă sau necomplicată poate fi tratată cu medicamente pe cale orală. Artemisininele în combinație cu alte antimalarice (inclusiv: amodiachină, lumefantrină, meflochină sau sulfadoxină / pirimetamină), cunoscute sub numele de terapie combinată cu artemisinină (ACT), este cel mai eficient tratament pentru infecția cu P. falciparum care scade rezistența la oricare componentă medicamentoasă (WHO - Guidelines for the Treatment of Malaria, 2015; Amadi et al, 2015; Kokwaro, 2009). O altă combinație recomandată este dihidroartemisinina și piperaquina. ACT este eficientă în aproximativ 90% din cazuri atunci când este utilizată pentru tratamentul malariei necomplicate (WHO - Guidelines for the Treatment of Malaria, 2015; Amadi et al, 2015; Howitt et al, 2012). Pentru a trata malaria necomplicată în timpul sarcinii, OMS recomandă utilizarea chininei

67
plus clindamicină la începutul sarcinii (1-ul trimestru) şi a ACT în etapele avansate (al 2-lea şi al 3-lea trimestru) (WHO - Guidelines for the Treatment of Malaria, 2015; Piola et al, 2010; McGready et al, 2012). De obicei, infecțiile cu P. vivax, P. ovale sau P. malariae nu necesită spitalizare. Tratamentul cu P. vivax trebuie să includă atât tratamentul stadiilor din sânge (cu clorochină sau ACT), cât și curățarea formelor hepatice cu primachină (WHO - Guidelines for the Treatment of Malaria, 2015; Meremikwu et al, 2012). Tratamentul cu tafenoquină previne recidivele după confirmarea malariei P. vivax (Kokwaro, 2009; Rajapakse et al, 2015). În zonele cu transmitere scăzută, ar trebui să fie utilizată o singură doză mică de primaquină în tratamentul antimalaric, cu scopul de a reduce transmiterea infecției. Testarea pentru G6PD nu este necesară, pentru că o singură doză mică de primaquină este eficientă în blocarea transmisiei și este puțin probabil să provoace toxicitate gravă la persoanele cu deficit de G6PD. Infecția cu P. falciparum provoacă aproape întotdeauna malarie severă și complicată, dar celelalte specii provoacă de obicei doar boala febrilă (White, 2011). Malaria gravă și complicată reprezinta urgențe medicale care produc rate mari de mortalitate (între 10% și 50%) (Howitt et al, 2012). Malaria cerebrala este forma malariei severe și complicate cu cele mai grave simptome neurologice (Manyando et al, 2011). Tratamentul recomandat pentru malaria severă este utilizarea intravenoasă a medicamentelor antimalarice. Pentru malaria severă, artesunatul parenteral a fost superior chininei atât la copii, cât și la adulți (WHO - Guidelines for the Treatment of Malaria, 2015; Sinclair et al, 2012). Alte studii arată că derivații de artemisinină (artemether și arteether) au fost la fel de eficace ca și chinina în tratamentul malariei cerebrale la copii (Rajapakse et al, 2015). Este esențial ca în tratamentul inițial al malariei severe (cel puțin timp de 24 de ore) să se administreze imediat doze complete de tratament antimalaric eficace parenteral (sau rectal pentru copii sub vârsta de 6 ani). Aceasta trebuie urmată de o doză completă (administrare de 3 zile) de ACT eficient pe cale orală dacă pacientul poate tolera medicamentul. Două clase de medicamente sunt disponibile pentru tratamentul parenteral al malariei severe: derivații de artemisinină (artesunat sau artemether) și alcaloizii Cinchona (chinina și chinidina). Artesunatul parenteral este tratamentul de ales pentru toate cazurile de malarie severe. Un studiu clinic amplu randomizat privind tratamentul malariei severe cu P.falciparum a arătat o reducere importantă a ratei de mortalitate cu artesunat parenteral (intravenos sau intramuscular) în comparație cu chinina parenterală. Artesunatul a fost mai simplu și mai sigur de utilizat şi nu aprezentat o

68
creștere a cazurilor cu sechele neurologice la supraviețuitorii tratați (WHO - Guidelines for the Treatment of Malaria, 2015). Cele mai utilizate medicamente în tratamentul malariei sunt active împotriva formelor de paraziți din sânge (forma responsabilă pentru apariția bolii) și includ: chloroquina; atovaquone-proguanil (Malarone®); artemether-lumefantrine (Coartem®); mefloquina (Lariam®); chinina; chnidina; doxycyclina (utilizată în combinaţie cu chinina); clindamicina (utilizată în combinaţie cu chinina) şi artesunatul (Bartoloni and Zammarchi, 2011; Greenwood et al, 2005).

69
REFERINȚE BIBLIOGRAFICE 1. Abdul-Ghani R, Mahdy MAK, Saif-Ali R, Alkubati SA, Alqubaty AR, Al-Mikhlafy
AA, Al-Eryani SM et al. Glucose-6-phosphate dehydrogenase deficiency among Yemeni children residing in malaria-endemic areas of Hodeidah governorate and evaluation of a rapid diagnostic test for its detection. Malari J 2016; 15:237. DOI 10.1186/s12936-016-1372-9.
2. Adoubryn KD, Ouhon J, Yapo CG, N’guettia KS, Kouakou KAE, Adou SJ et al. Efficacitѐ thѐrapeutique de la chloroquine dans le traitement du paludisme simple á Plasmodium falciparum de lꞌenfant dans lꞌest de la Côte-dꞌIvoire (1999-2002). Med Mal Infect 2004; 34 (3) : 127-131. https://doi.org/10.1016/j.medmal.2003.11.004.
3. Ajdukiewicz KMB, Ong ELC. Management of vivax malaria with low sensitivity to primaquine. J Infection 2007; 54 (3) : 209–211.DOI: https://doi.org/10.1016/j.jinf. 2006. 05.008.
4. Al-Olayan EM, Beetsma AL, Butcher GA, Sinden RE, Hurd H. Complete development of mosquito phases of the malaria parasite in vitro. Science 2002; 295 (5555) : 677-679. DOI: 10.1126/science.1067159.
5. Al-Eryani SMA, Kelly-Hope L, Harbach RE, Briscoe AG, Barnish G, Azazy A, McCall PJ. Entomological aspects and the role of human behaviour in malaria transmission in a highland region of the Republic of Yemen. Malari J 2016; 15 : 130. DOI 10.1186/s12936-016-1179-8.
6. D’Alessandro G, Bruno Smiraglia C, Lavagnino A. Further studies on the biology of Anopheles labranchiae labranchiae Falleroni in Sicily. World Health Organization, 1971, pp 1-11.
7. Alten B, Caglar SS, Simsek FM, Kaynas S. Effect of insecticide-treated bednets for malaria control in Southeast Anatolia-Turkey. J Vector Ecol 2003; 28 : 97-107.
8. Aly AS, Vaughan AM, Kappe SH. Malaria parasite development in the mosquito and infection of the mammalian host. Annu Rev Microbiol 2009; 63 : 195-221. doi: 10.1146/annurev.micro.091208.073403.
9. Amadi OK, Otuokere IE, Bartholomew CF. Synthesis, Characterization, in vivo Antimalarial Studies and Geometry. Optimization of Lumefantrine/Artemether Mixed Ligand Complexes. Research Journal of Pharmaceutical Dosage Forms and Technology 2015; 7 (1): 59-68.
10. Amino R, Giovannini D, Thiberge S, Gueirard P, Boisson B, Dubremetz JF et al. Host cell traversal is important for progression of the malaria parasite through the dermis to the liver. Cell Host Microbe 2008; 3 (2) : 88-96. doi: 10.1016/j.chom.2007.

70
11. Amino R, Thiberge S, Blazquez S, Baldacci P, Renaud O, Shorte S, Ménard R. Imaging malaria sporozoites in the dermis of the mammalian host. Nat Protoc 2007; 2 (7) : 1705-1712. DOI: 10.1038/nprot.2007.120.
12. Amino R, Thiberge S, Martin B, Celli S, Shorte S, Frischknecht F, Ménard R. Quantitative imaging of Plasmodium transmission from mosquito to mammal. Nat Med 2006; 12 (2) : 220-224. DOI: 10.1038/nm1350.
13. Askling H, Bruneel F, Burchard G, Castelli F, Chiodini PL, Grobusch M et al. Management of imported malaria in Europe. Malaria J 2012; 11: 328. https://doi.org/10.1186/1475-2875-11-328.
14. Akiner MM, Cağlar SS. Identification of Anopheles maculipennis group species using polymerase chain reaction (PCR) in the regions of Birecik. Beyşehir and Cankiri. Turkiye Parazitol Derg 2010; 34 (1) : 50-54.
15. Baird JK, Fryauff DJ, Hoffman SL. Primaquine for prevention of malaria in travelers. Clin Infect Dis 2003; 37 (12) : 1659-1667. DOI: 10.1086/379714
16. Baird JK, Rieckmann KH. Can primaquine therapy for vivax malaria be improved? Trends Parasitol 2003;19 (3) : 115-20.
17. Baird JK, Hoffman SL. Primaquine therapy for malaria. Clin Infect Dis 2004; 39 (9) : 1336-1345. DOI: 10.1086/424663.
18. Bartoloni A, Zammarchi L. Clinical aspects of uncomplicated and severe malaria. Mediterr J Hematol Infect Dis 2012; 4 (1): e2012026. doi: 10.4084/MJHID.2012.026.
22. Bates M. Variations in the antepalmate hairs of larvae ofthe Anopheles maculipennis group. Rivista di Malariologia 1939; 18 :299-312.
23. Becker N, Petric D, Boase C, Lane J, Zgomba M, Dahl C, Kaiser A. Mosquitoes and their control. Kluwer Academic / Plenum Publishers, New York, 2003.
24. Billker O, Lindo V, Panico M, Etienne AE, Paxton T, Dell A et al. Identification of xanthurenic acid as the putative inducer of malaria development in the mosquito. Nature 1998; 392 (6673) : 289-292. DOI: 10.1038/32667.
25. Billker O, Shaw MK, Margos G, Sinden RE. The roles of temperature, pH and mosquito factors as triggers of male and female gametogenesis of Plasmodium berghei in vitro. Parasitology 1997; 115 ( Pt 1) : 1-7.
26. Bledsoe GH. Malaria primer for clinicians in the United States. South Med J 2005; 98 (12): 1197–1204.
27. Boccolini D, Sabatini A, Coluzzi M. Valore diagnostico del numero dei rami delle setole antepalmate per l'identificazione del complesso Anopheles maculipennis. Ann Ist Super Sanita 1986; 22 : 201-204.
28. Boccolini D, Toma L, Di Luca M, Severini F, Cocchi M, Bella A et al. Impact of environmental changes and human-related factors on the potential malaria vector, Anopheles labranchiae (Diptera: Culicidae), in Maremma, Central Italy. J Med Entomol 2012; 49 (4) : 833-842. https://doi.org/10.1603/ME11252.
29. Bocşan IS, Rădulescu A, Brumboiu I, Şuteu O, Achimaş A. Epidemiologie practică pentru medici de familie. Editura Medicală Universitară Iuliu Haţieganu, Cluj Napoca, 1999, pg. 393-405. [Bocsan IS, Radulescu A, Brumboiu I, Suteu O, Achimas A. Practical epidemyology for general practitioners. Iuliu Hatieganu Medical University Publishing House, Cluj Napoca, 1999, pg. 393-405.]

71
30. Boddey JA and Cowman AF. Plasmodium nesting: remaking the erythrocyte from the inside out. Annu Rev Microbiol 2013; 67 : 243-269. doi: 10.1146/annurev-micro-092412-155730.
31. Boëte C (ed). Genetically Modified Mosquitoes for Malaria Control. Eurekah/Landes Bioscience, Georgetown, 2006.
32. Bogdan O, Marinica I. Hazardele climatice din zona temperată. Geneză și vulnerabilitate cu aplicații la România. Edit. Universitară "L. Blaga", Sibiu, 2007. [Climate risks in the temperate zone. Genesis and application vulnerability in Romania. “L. Blaga” University Publishing House, sibiu, 2007].
33. Bray PG, Martin RE, Tilley L, Ward SA, Kirk K, Fidock DA. Defining the role of PfCRT in Plasmodium falciparum chloroquine resistance. Mol Microbiol 2005; 56 (2) : 323-333. DOI: 10.1111/j.1365-2958.2005.04556.x.
34. Bruce-Chwatt LJ, de Zulueta J. The rise and fall of malaria in Europe. Oxford University Press, London, 1980, 240pp.
35. Buonomini G. L'esame delle setole antepalmate per lostudio della popolazione larvale dei focolai naturali di Anopheles maculipennis. Rivista di Parassitologia 1940; 6 : 163-174.
36. Cambournac FJ. Contribution to the history of malaria epidemiology and control in Portugal and some other places. Parassitologia 1994; 36 (1-2) : 215-222.
37. Capinera JL. Encyclopedia of Entomology. 2nd edition, Springer, New York, 2002, 4 vol, 4411pp.
38. Carmona-Fonseca J, Maestre A. Prevention of Plasmodium vivax malaria recurrence: efficacy of the standard total dose of primaquine administered over 3 days. Acta Trop 2009; 112 (2) : 188-192. doi: 10.1016/j.actatropica.2009.07.024.
39. Carter R, Mendis KN. Evolutionary and Historical Aspects of the Burden of Malaria. Clin Microbiol Rev 2002; 15 (4) : 564–594. doi: [10.1128/CMR.15.4.564-594.2002].
40. Chakravarty S, Cockburn IA, Kuk S, Overstreet MG, Sacci JB, Zavala F. CD8+ T lymphocytes protective against malaria liver stages are primed in skin-draining lymph nodes. Nat Med 2007; 13 (9) : 1035-41. DOI: 10.1038/nm1628.
41. Cianchi R, Sabatini A, Boccolini D, Bullini L,Coluzzi M. Electrophoretic evidence of reproductive isolation between sympatric populations of Anopheles melanoon and An. subalpinus. Third International Congress on Malaria and Babesiosis, International Laveran Foundation, 7–11 September, 1987, Annecy, France.
42. Cogswell FB. The hypnozoite and relapse in primate malaria. Clin Microbiol Rev 1992; 5 (1) : 26-35.
43. Colofiţchi A, Nicolescu G, Purcărea-Ciulacu V, Vladimirescu A, Coipan EC, Steriu D. Is malaria a re-emerging disease in Romania? 14th European Conference of the Society of vector Ecology, Bellinzona, Elvetia, 3 – 6 Septembrie 2003.
44. Corradetti A. Richerche sugli incroci tra le varietà di Anopheles maculipennis. Rivista di Malariologia 1934; 13 : 707–720.

72
45. Cosoroaba I. Entomologie veterinară. Editura Ceres, București, 1992 [Veterinary entomology. Ceres Publishing House, Bucharest, 1992].
46. Cosoroaba I. Parazitologie veterinară. Editura Mirton, Timisoara, 2000 [Veterinary parasitology. Mirton Publishing House, Bucharest, 2000].
47. Dalrymple U, Cameron E, Bhatt S, Weiss DJ, Gupta S, Gething PW. Quantifying the contribution of Plasmodium falciparum malaria to febrile illness amongst African children. eLlife 2017; 6: e29198. doi: 10.7554/eLife.29198.
48. de Buck A, Swellengrebel NH. Behaviour of Dutch Anopheles atroparvus and messeae in winter under artificial conditions. Rivista di Malariologia 1934; 8 : 404.
49. de Buck A, van der Torren G, Swellengrebel NH. Report for the year 1932 on investigation into the racial composition of Anopheles maculipennis in Holland. Rivista di Malariologia 1933; 12 : 265.
50. Derbyshire ER, Mazitschek R, Clardy J. Characterization of Plasmodium liver stage inhibition by halofuginone. Chem Med Chem 2012; 7 (5) : 844-849. doi: 10.1002/cmdc.201200045.
51. Develoux M, Le Loup G, Dautheville S, Belkadi G, Magne D, Lassel L et al. Malaria among immigrants, experience of a Parisian hospital (2006-2010). Bull Soc Pathol Exot 2012; 105 (2) : 95-102. doi: 10.1007/s13149-012-0217-1.
52. de Zulueta J, Ramsdale CD, Cianchi R, Bullini L, Coluzzi M. Observations on the taxonomic status of Anopheles sicaulti. Parassitologia 1983; 23 :73-92.
53. Di Luca M, Boccolini D, Marinucci M, Romi R. Intrapopulation polymorphism in Anopheles messeae (An. maculipennis complex) inferred by molecular analysis. J Med Entomol 2004; 41 : 582-586.
54. Djadid ND, Gholizadeh S, Tafsiri E, Romi R, Gordeev M, Zakeri S. Molecular identification of Palearctic members of Anopheles maculipennis in northern Iran. Malaria J 2007; 6 : 6. https://doi.org/10.1186/1475-2875-6-6.
55. Dondorp AM, von Seidlein L. Malaria. In: Cohen J, Powderly WG, Opal SM. (eds). Infectious Diseases. 4th edition, Vol I. Elsevier, 2017, p 1014-1025.
56. Dua VK, Pandey AC, Raghavendra K, Gupta A, Sharma T, Dash AP. Larvicidal activity of neem oil (Azadirachta indica) formulation against mosquitoes. Malar J 2009; 8 : 124. doi: 10.1186/1475-2875-8-124.
57. Eritja R, Aranda C, Padros J, GouIa M. Revised checklist of the Spanish mosquitoes. Acta Virologica Portuguesa 1998; 5 : 25.
58. Falleroni D. Zooprofilassi e sua applicazione in Ardea. (Argo Romano). Diverse razze di "A. maculipennis". Tipografia Cuggiani, Roma, 1932, pp. 48+ x.
59. Falleroni D. Fauna anofelica italiana e suo 'habitat' (paludi, risaie, canali). Metodi di lotta contro la malaria. Rivista di Malariologia 1926; 5 : 553-559.
60. Fanello C, Santolamazza F, della Torre A. Simultaneous identification of species and molecular forms of the Anopheles gambiae complex by PCR-RFLP. Med Vet Entomol 2002; 16 (4) : 461-464.
61. Faraj C, Adlaoui E, Elkohli M, Herrak T, Ameur B, Chandre F. Review of Temephos Discriminating Concentration for Monitoring the Susceptibility of Anopheles labranchiae (Falleroni, 1926), Malaria Vector in Morocco. Malar Res Treat 2010; 2010: 126085. doi: 10.4061/2010/126085.

73
62. Ferrão JL, Mendes JM, Painho M, Zacarias S. Malaria mortality characterization and the relationship between malaria mortality and climate in Chimoio, Mozambique. Malaria J 2017; 16 : 212. https://doi.org/10.1186/s12936-017-1866-0.
63. Field JW and Shute PG. The microscopic diagnosis of human malaria. II. A morphological study of the erythrocyticparasite. Government Press, Kuala Lumpur, Malaysia, 1956.
64. Fillinger U, Sonye G, Killeen GF, Knols BG, Becker N. The practical importance of permanent and semipermanent habitats for controlling aquatic stages of Anopheles gambiae sensu lato mosquitoes: operational observations from a rural town in western Kenya. Trop Med Int Health 2004; 9 (12) : 1274-1289. DOI: 10.1111/j.1365-3156.2004.01335.x.
65. Foster WA, Walker ED. Mosquitoes (Culicidae). In Mullen, G, Durden, L. (Eds.) Medical and veterinary entomology. 2nd edition. (pp. 207-259). Academic Press, Burlington, MA, 2009. 637 pp.
66. Frevert U. Sneaking in through the back entrance: the biology of malaria liver stages. Trends Parasitol 2004; 20 (9) : 417-424. doi:10.1016/j.pt.2004.07.007.
67. Frizzi G. Etude cytogénétique d'Anopheles maculipennis en Italie. Bull World Health Organ 1953; 9 : pp. 335-344.
68. Frizzi G. Nuovi contributi e prospetti di ricerca nel gruppo Anopheles maculipennis in base allo studio del dimorfismo cromosomico. Symposia Genetica 1952; 3 : pp. 231-265.
69. Gillies MT, de Meillon B. The Anophelini of Africa south of the Sahara (Ethiopian zoogeographical region). 2nd edition. Publ S Afr Inst Med Res 1968; 54 : l-343.
70. Girod R, Orlandi-Pradines E, Rogier C, Pagès F. Malaria transmission and insecticide resistance of Anopheles gambiae (Diptera: Culicidae) in the French Military Camp of Port-Bouët, Abidjan (Côte d’Ivoire): implications for vector control. J Med Entomol 2006; 43 (5) : 1082–1087.
71. Gratzer WB, Dluzewski AR. The red blood cell and malaria parasite invasion. Semin Hematol 1993; 30 (3) : 232-247.
72. Greenwood BM, Bojang K, Whitty CJ, Targett GA. Malaria. Lancet 2005; 365 (9469) : 1487–1498. doi: 10.1016/S0140-6736(05)66420-3.
73. Hackett LW, Lewis DJ. A new variety of Anopheles maculipennis in southern Europe. Rivista di Malariologia 1935; 14 : 377-383.
74. Hackett LW, Missiroli A. The varieties of Anopheles maculipennis and their relation to the distribution of malaria in Europe. Rivista di Malariologia 1935; 14 : 45-109.
75. Hafalla JC, Silvie O, Matuschewski K. Cell biology and immunology of malaria. Immunol Rev 2011; 240 (1) : 297-316. doi: 10.1111/j.1600-065X.2010.00988.x.
76. Hay S, Snow RW. The Malaria Atlas Project: Developing Global Maps of Malaria Risk. Plos Medicine 2006; 3 (12) : 2204-2208. https://doi.org/10.1371/journal.pmed.0030473.

74
77. Harbach RE. Review of the internal classification of the genus Anopheles (Diptera: Culicidae): the foundation for comparative systematics and phylogenetic research. Bull
a. Entomol Res 1994; 84: 331-342. 78. Heymann DL. Manual de Management al Bolilor Transmisibile. Un raport oficial
al American Public Health Association. Ediţia a 19-a . Editura Medicală Amaltea, Bucureşti, 2012, pag. 251-267. [Control Of Communicable Diseases Manual. An official report of the American Public Health Association. 19th Edition. Amaltea Medical Publishing House, Bucharest, 2012, pg. 251-267].
79. Howitt P, Darzi A, Yang GZ, Ashrafian H, Atun R, Barlow J et al. Technologies for global health. Lancet 2012; 380 (9840) : 507–535. doi: 10.1016/S0140-6736(12)61127-1.
80. Ivanescu Maria Larisa – Bioecologia si patogenitatea unor culicide . Teza de doctorat USAMV Iasi –Ro, 246 p., 2014 81. Ivanescu ML, Acatrinei D, Pavel I, Sulesco T, Miron L. PCR identification of five
species from the Anopheles maculipennis complex (Diptera: Culicidae) in North-Eastern Romania. Acta Parasitologica 2015; 60 (2) : 283-289. doi: 10.1515/ap-2015-0040.
82. Ivănescu ML, Bodale I, Florescu SA, Roman C, Acatrinei D, Miron L. Climate Change Is Increasing the Risk of the Reemergence of Malaria in Romania. Bio Med Res Internat 2016; http://dx.doi.org/ 10.1155/ 2016/8560519.
83. Jaenson TG, Ameneshewa B. Prehibernation diet and reproductive condition of female Anopheles messeae in Sweden. Med Vet Entomol 1991; 5 (2) : 243-252.
84. Jelinek T, Schulte C, Behrens R, Grobusch MP, Coulaud JP et al. Imported Falciparum malaria in Europe: sentinel surveillance data from the European network on surveillance of imported infectious diseases. Clin Infect Dis 2002; 34 (5) : 572-576. DOI: 10.1086/338235.
85. Kappe SH, Gardner MJ, Brown SM, Ross J, Matuschewski K, Ribeiro JM et al. Exploring the transcriptome of the malaria sporozoite stage. Proc Natl Acad Sci USA 2001; 98 (17) : 9895-9900. DOI: 10.1073/pnas.171185198.
86. Kasap H, Akbaba M, Kasap M, Lüleyap Ü, Alpaslan N, Alptekin D. A study of malaria andAnopheles sacharovi in Tuzla, Adana, Turkey.Acta Parasit Turcica 1995; 19 : 535-540.
87. Kettle DS (ed). Medical and veterinary entomology. 2nd ed. CAB International Publishing House, Wallingford, UK, 1995, 725 pp.
88. Kiszewski A, Mellinger A, Spielman A, Malaney P, Sachs SE, Sachs J. A global index representing the stability of malaria transmission. Am J Trop Med Hyg 2004; 70 (5) : 486-498.
89. Kitzmiller JB, Frizzi G, Baker R. Evolution and speciation within the maculipennis complex of the genus Anopheles. In Wright JW, Pal R (eds): Genetics of Insect Vectors of Disease. Elsevier Publishing, London, 1967, pp. 151-210.
90. Knight KL, Stone A. A catalog of the mosquitoes of the world. 2nd edition, Thomas say Foundation, 1977; 611 pp.

75
91. Knottnerus O.S. Malaria around the North Sea: a survey. In: Wefer G., Berger W.H., Behre KE., Jansen E. (eds) Climate Development and History of the North Atlantic Realm. Springer, Berlin, Heidelberg, 2002.
92. Kokwaro G. Ongoing challenges in the management of malaria. Malaria J 2009; 8 (Suppl. 1) : S2. doi:10.1186/1475-2875-8-S1-S2.
93. Konaté L, Zwetyenga J, Rogier C, Bischoff E, Fontenille D, Tall A et al. Variation of Plasmodium falciparum msp1 block 2 and msp2 allele prevalence and of infection complexity in two neighbouring Senegalese villages with different transmission conditions. Trans R Soc Trop Med Hyg 1999; 93 (Suppl 1) : 21-28.
94. Korenromp EL, Miller J, Cibulskis RE, Kabir Cham M, Alnwick D et al. Monitoring mosquito net coverage for malaria control in Africa: possession vs. use by children under 5 years. Trop Med Int Health 2003; 8 (8) : 693-703.
95. Korvenkontio P, Lokki J, Saura A, Ulmanen I. Anopheles maculipennis complex (Diptera: Culicidae) in northern Europe: species diagnosis by egg structure and enzyme polymorphism. J Med Entomol 1979; 16 : 169-170.
96. Krüger A, Rech A, Su XZ, Tannich E. Two cases of autochthonous Plasmodium falciparum malaria in Germany with evidence for local transmission by indigenous Anopheles plumbeus. Trop Med Int Health 2001; 6 (12) : 983-985.
97. Lacroix R, Mukabana WR, Gouagna LC, Koella JC. Malaria infection increases attractiveness of humans to mosquitoes. PLoS Biol 2005; 3 : 1590-1593. DOI: 10.1371/journal.pbio.0030298.
98. Lasonder E, Janse CJ, van Gemert GJ, Mair GR, Vermunt AM, Douradinha BG et al. Proteomic profiling of Plasmodium sporozoite maturation identifies new proteins essential for parasite development and infectivity. PLoS Pathog 2008; 4 (10) : e1000195. doi: 10.1371/journal.ppat.1000195.
99. Lavazec C, Bourgouin C. Mosquito-based transmission blocking vaccines for interrupting Plasmodium development. Microbes Infect 2008; 10 (8) : 845-849. doi: 10.1016/j.micinf.2008.05.004.
100. Lines J. The technical issues. In: Lengeler C, de Savigny D, Catani J (eds.). Net Gain: a new method of preventing malaria deaths. International Development Research Center, Ottawa, Canada and World Health Organization, Geneva, Switzerland, 1996.
101. Linton YM, Leel AS, Curtis C. Discovery of a third member of the Maculipennis Group in SW England. Europ Mosq Bull 2005; 19 : 5-9.
102. Linton YM, Harbach RE, Chang MS, Anthony TG, Matusop A. Morphological and molecular identity of Anopheles (Cellia) sundaicus (Diptera: Culicidae), the nominotypical member of a malaria vector species complex in Southeast Asia. Syst Entomol 2001; 26 :357-366.
103. Linton YM, Samanidou-Voyadjoglou A, Harbach RE. Ribosomal ITS2 sequence data for Anopheles maculipennis and An. messeae in northern Greece, with a critical assessment of previously published sequences. Insect Mol Biol 2002; 11 : 379-383.
104. Linton YM, Smith L, Koliopoulos G, Samanidou-Voyadjoglou A, Zounos AK, Harbach RE. Morphological and molecular characterization of Anopheles (Anopheles) maculipennis Meigen, type species of the genus and nominotypical member of the Maculipennis Complex. Syst Entomol 2003; 28 : 36-56.

76
105. Luca M. Malaria. În: Ivan A (ed.): Tratat de epidemiologie a bolilor transmisibile. Editura Polirom, Iaşi, 2002, pag. 676-681. [Malaria. In: Ivan A (ed.): Communicable diseases epidemiology handbook. Polirom Publishing House, Iasi, 2002, pg. 676-681].
106. Lucas RM, McMichael AJ. Association or causation: evaluating links between "environment and disease". Bull World Health Organ 2005; 83 (10) : 792–795.
107. Manguin S, Garros C, Dusfour I, Harbach RE, Coosemans M. Bionomics, taxonomy, and distribution of the major malaria vector taxa of Anopheles subgenus Cellia in Southeast Asia: an updated review. Infect Genet Evol 2008; 8 (4) : 489-503. doi: 10.1016/j.meegid.2007.11.004.
108. Manyando C, Kayentao K, D'Alessandro U, Okafor HU, Juma E, Hamed K. A systematic review of the safety and efficacy of artemether-lumefantrine against uncomplicated Plasmodium falciparum malaria during pregnancy. Malaria J 2011; 1 (11) : 141. doi: 10.1186/1475-2875-11-141.
109. Marchi A, Munstermann LE. The mosquitoes of Sardinia: species records 35 years after the malaria eradication campaign. Med Vet Entomol 1987; 1 (1) : 89-96.
110. Marinucci M, Romi R, Mancini P, Di Luca M, Severini C. Phylogenetic relationships of seven Palearctic members of the maculipennis complex inferred from ITS2 sequence analysis. Insect Mol Biol 1999; 8 (4) : 469–480. 10.1046/j.1365-2583.1999.00140.x.
111. Marquardt WC, Kondratieff BC. Biology of disease vectors. 2nd edition. Elsevier Academic Press, 2004. 816pp.
112. McGready R, Lee SJ, Wiladphaingern J, Ashley EA, Rijken MJ, Boel M et al. Adverse effects of falciparum and vivax malaria and the safety of antimalarial treatment in early pregnancy: a population-based study. Lancet Infect Dis 2012; 12 (5) : 388-396. doi: 10.1016/S1473-3099(11)70339-5.
113. Medica DL, Sinnis P. Quantitative dynamics of Plasmodium yoelii sporozoite transmission by infected anopheline mosquitoes. Infect Immun 2005; 73 (7) : 4363-4369. DOI: 10.1128/IAI.73.7.4363-4369.2005.
114. Meijerink J, Braks MAH, Brack AA, Adam W, Dekker T, Posthumus MA et al. Identification of Olfactory Stimulants for Anopheles gambiae from Human Sweat Samples. J Chem Ecol 2000; 26 : 1367–1382.
115. Ménard D, Andrianina NNH, Ramiandrasoa Z, Randriamanantena A, Rasoarilalao N, Jahevitra M. Randomized clinical trial of artemisinin versus non-artemisinin combination therapy for uncomplicated falciparum malaria in Madagascar. Malaria J 2007; 6 : 65. doi:10.1186/1475-2875-6-65.
116. Mens PF, Bojtor EC, Schallig HDFH. Molecular interactions in the placenta during malaria infection. Eur J Obstet Gynecol Reprod Biol 2012; 152 (2): 126–132. doi: 10.1016/j.ejogrb.2010.05.013.
117. Meremikwu MM, Odigwe CC, Akudo Nwagbara B, Udoh EE. Antipyretic measures for treating fever in malaria. Cochrane Database Syst Rev 2012; 9 : CD002151. DOI: 10.1002/14651858.CD002151.pub2.

77
118. Meunier B, Robert A. Heme as Trigger and Target for Trioxane-Containing Antimalarial Drugs. Acc Chem Res 2010; 43 (11) : 1444-1451. DOI: 10.1021/ar100070k
119. Mittal PK. Biolarvicides in vector control: challenges and prospects. J Vector Borne Dis 2003;40 (1-2) : 20-32.
120. Monge-Maillo B, López-Vélez R. Migration and Malaria in Europe. Mediterr J Hematol Infect Dis 2012; 4 (1) : e2012014. doi: [10.4084/MJHID.2012.014].
121. Moore RB, Oborník M, Janouskovec J, Chrudimský T, Vancová M, Green DH et al. A photosynthetic alveolate closely related to apicomplexan parasites. Nature 2008; 451 (7181) : 959-963. doi: 10.1038/nature06635.
122. Mota MM, Pradel G, Vanderberg JP, Hafalla JC, Frevert U, Nussenzweig RS et al. Migration of Plasmodium sporozoites through cells before infection. Science 2001; 291 (5501) : 141-414. DOI: 10.1126/science.291.5501.141.
123. Mouchet J, Carnevale P, Coosemans M, Fontenille D, Ravaonjanahary C, Richard A et al. Typologie du paludisme en Afrique. Cahiers Santé 1999; 3 : 220–238.
124. Mpogoro FJ, Matovelo D, Dosani A, Ngallaba S, Mugono M, Mazigo HD. Uptake of intermittent preventive treatment with sulphadoxine-pyrimethamine for malaria during pregnancy and pregnancy outcomes: a cross-sectional study in Geita district, North-Western Tanzania. Malaria J 2014; 13:455. doi.10.1186/1475-2875-13-455.
125. Mueller AK, Labaied M, Kappe SH, Matuschewski K. Genetically modified Plasmodium parasites as a protective experimental malaria vaccine. Nature 2005; 433 (7022) : 164-167. DOI: 10.1038/nature03188.
126. Mullen G, Durden L. Medical and Veterinary Entomology. 1st edition, Academic Press, 2002. 597pp.
127. Nadjm B, Behrens RH. Malaria: An update for physicians. Infect Dis Clin N Am 2012; 26 (2) : 243–259. doi: 10.1016/j.idc.2012.03.010.
128. Nevill CG, Some ES, Mung'ala VO, Mutemi W, New L, Marsh K et al. Insecticide-treated bednets reduce mortality and severe morbidity from malaria among children on the Kenyan coast. Trop Med Int Health 1996; 1 (2) : 139-146.
129. Nicolescu G, Linton YM, Vladimirescu A, Howard TM, Harbach RE. Mosquitoes of the Anopheles maculipennis group (Diptera; Culicidae) in Romania, with the discovery and formal recognition of a new species based on molecular and morphological evidence. Bull Entomol Res 2004; 94: 525–535. G. doi.org/10.1079/BER2004330.
130. Nitzulescu V, Gherman I. Entomologie medicală. Editura Academiei Romane, 1990, 117-129. [Medical entomology. Romanian Academy Publishing House, 1990, 117-129.]
131. Okumu FO, Knols BGJ, Fillinger U. Larvicidal effects of a neem (Azadirachta indica) oil formulation on the malaria vector Anopheles gambiae. Malaria J 2007; 6 : 63. doi:10.1186/1475-2875-6-63.
132. Oliver M, Simon F, de Monbrison F, Beavogui AH, Pradines B, Ragot C et al. New use of primaquine for malaria. Med Mal Infect 2008; 38 (4) : 169-179. doi: 10.1016/j.medmal.2008.01.011.

78
133. Ouattara AF, Dagnogo M, Constant EA, Kone M, Raso G, Tanner M, et al. Transmission of malaria in relation to distribution and coverage of long-lasting insecticidal nets in central Cote d'Ivoire. Malar J 2014; 13 : 109. doi: 10.1186/1475-2875-13-109.
134. Pages F, Orlandi-Pradines E, Corbel V. Vecteurs du paludisme: biologie, diversité, contrôle et protection individuelle Vectors of malaria: biology, diversity, prevention, and individual protection. Med Mal Infect 2007; 37 : 153–161.
135. Pascual M, Ahumada JA, Chaves LF, Rodó X, Bouma M. Malaria resurgence in the East African highlands: temperature trends revisited. Proc Natl Acad Sci USA 2006; 103 (15) : 5829-5834. DOI: 10.1073/pnas.0508929103.
136. Patz JA, McGeehin MA, Bernard SM, Ebi KL, Epstein PR, Grambsch A et al. The potential health impacts of climate variability and change for the United States: executive summary of the report of the health sector of the US National Assessment. Environ Health Perspect 2000; 108 (4) : 367–376.
137. Patz JA, Olson SH. Malaria risk and temperature: Influences from global climate change and local land use practices. Proc Natl Acad Sci USA 2006; 103 (15) : 5635–5636. doi: 10.1073/pnas.0601493103.
138. Patz JA, Frumkin H, Holloway T, Vimont DJ, Hains A. Climate change: challenges and opportunities for global health. JAMA 2014; 312 (15) : 1565-1580. doi: 10.1001/jama.2014.13186.
139. Pener H and Kitron U. Distribution of mosquitoes (Diptera: Culicidae) in northern Israel: a historical perspective. I. Anopheline mosquitoes. J Med Entomol 1985; 22 (5) : 536-543.
140. Pietra Y, Procacci PG, Sabatinelli G, Kumlien S, Lamizana L, Rotigliano G. Impact de l'utilisation des rideaux imprégnés de perméthrine sur le paludisme dans une zone rurale de haute transmission au Burkina Faso. Bull Soc Pathol Exot 1991; 84 : 375–385.
141. Pimenta PFP, Orfano AS, Bahia AC, Duarte APM, Ríos-Velásquez CM, Melo FF et al. An overview of malaria transmission from the perspective of Amazon Anopheles vectors. Mem Inst Oswaldo Cruz 2015; 110 (1) : 23–47. doi:10.1590/0074-02760140266.
142. Piola P, Nabasumba C, Turyakira E, Dhorda M, Lindegardh N, Nyehangane D et al. Efficacy and safety of artemether-lumefantrine compared with quinine in pregnant women with uncomplicated Plasmodium falciparum malaria: an open-label, randomised, non-inferiority trial. Lancet Infect Dis 2010; 10 (11) : 762-779. doi: 10.1016/S1473-3099(10)70202-4.
143. Ponçon N1, Tran A, Toty C, Luty AJ, Fontenille D. A quantitative risk assessment approach for mosquito-borne diseases: malaria re-emergence in southern France. Malar J 2008; 7 : 147. doi: 10.1186/1475-2875-7-147.
144. Porter CH, Collins FH. Species-diagnostic differencesin a ribosomal internal transcribed spacer from the sibling species Anopheles freeborni and Anopheles hermsi (Diptera:Culicidae). Am J Trop Med Hyg 1991; 45 : 271-279.
145. Pratt H, Moore C. Mosquitoes of public health importance and their control. CDC Department of Health and Human Services. Public Health Service, Centers

79
for Diseases Control and Prevention, Public Health Practice Program Office, Atlanta, Georgia, 1993.
146. Proft J, Maier WA, Kampen H. Identi®cation of six sibling species of the Anopheles maculipennis complex (Diptera: Culicidae) by a polymerase chain reaction assay. Parasitol Res 1999; 85: 837-843.
147. Rajapakse S, Rodrigo C, Fernando SD. Tafenoquine for preventing relapse in people with Plasmodium vivax malaria. Cochrane Database Syst Rev 2015; (4): 1–55. doi: 10.1002/14651858.CD010458.pub2.
148. Reiter P. Climate change and mosquito-borne disease. Environ Health Perspect 2001; 109 (Suppl 1) : 141-161. DOI: 10.1289/ehp.01109s1141.
149. Rénia L, Howland SW, Claser C, Charlotte Gruner A, Suwanarusk R, Hui Teo T, Russell B, Ng LF. Cerebral malaria: mysteries at the blood-brain barrier. Virulence 2012; 3 (2) : 193–201. doi: 10.4161/viru.19013.
150. Ribeiro H, Ramos HC, Pires CA, Capela RA. An annotated checklist of the mosquitoes of continental Portugal (Diptera, Culicidae). Actas III Congreso Ibérico de Entomologia 1988; 3 : 233-254.
151. Richter J, Franken G, Mehlhorn H, Labisch A, Häussinger D. What is the evidence for the existence of Plasmodium ovale hypnozoites? Parasitol Res 2010; 107 (6): 1285–1290. https://doi.org/10.1007/s00436-010-2071-z.
152. Robert V, Boudin C. Biology of man-mosquito plasmodium transmission. Bull Soc Pathol Exot 2003; 96 (1) : 6-20.
153. Roberts LS, Janovy J Jr. Foundations of Parasitology - Phylum Apicomplexa: Gregarines, Coccidia, and Related Organisms. 6th edition. McGraw-Hill, Boston, MA, 2000; 117–139.
154. Romi R, Pierdominici G, Severini C, Tamburro A, Cocchi M, Menichetti D, et al. Status of malaria vectors in Italy. J Med Entomol 1997; 34 : 263-271.
155. Romi R. Anopheles labranchiae, an important malaria vector in Italy, and other potential malaria vectors in Southern Europe. European Mosquito Bulletin 1999; 4 : 8-10.
156. Romi R, Boccolini D, Di Luca M, La Rosa G, Marinucci M. Identification of the sibling species of the Anopheles maculipennis complex by heteroduplex analysis. Insect Mol Biol 2000; 9 : 509-513.
157. Romi R, Boccolini D, Hovanesyan I, Grigoryan G, Di Luca M, Sabatinelli G. Anopheles sacharovi (Diptera: Culicidae): A reemerging malaria vector in the Ararat Valley of Armenia. J Med Entomol 2002; 39 (3) : 446-450.
158. Salam RA, Das JK, Lassi ZS, Bhutta ZA. Impact of community-based interventions for the prevention and control of malaria on intervention coverage and health outcomes for the prevention and control of malaria. Infect Dis Poverty 2014; 3 : 25. doi: 10.1186/2049-9957-3-25.
159. Sarneckis K. Mosquitoes in Constructed Wetlands. Government of South Australia, 2002. Available from: https://epdf.tips/mosquitoes-in-constructed-wetlands.html.
160. Sedaghat MM, Linton Y-M, Nicolescu G, Smith L, Koliopoulos G, Zounos AK et al. Morphological and molecular characterization of Anopheles (Anopheles)

80
sacharovi Favre, a primary vector of malaria in the Middle East. Syst Entomol 2003; 28 : 241-256.
161. Senevet G, Andarelli L. Les Anophèles d’Afrique du Nord et du Bassin méditérranéen. Encyclopédie Entomologique XXXIII. 1st edition. Paul Lechevalier Publishing House, Paris, 1956.
162. Silvie O, Mota MM, Matuschewski K, Prudencio M. Interactions of the malaria parasite and its mammalian host. Curr Opin Microbiol 2008; 11 (4) : 352-359. doi: 10.1016/j.mib.2008.06.005.
163. Sinka ME, Bangs MJ, Manguin S, Coetzee M, Mbogo CM, Hemingway J et al. The dominant Anopheles vectors of human malaria in Africa, Europe and the Middle East: occurrence data, distribution maps and bionomic précis. Paras Vect 2010; 3 : 117. doi:10.1186/1756-3305-3-117.
164. Sinclair D, Donegan S, Isba R, Lalloo DG. Artesunate versus quinine for treating severe malaria. Cochrane Database Syst Rev 2012; (6) : CD005967. doi: 10.1002/14651858.CD005967.pub4.
165. Smallegange RC, Qiu YT, van Loon JJ, Takken W. Synergism between ammonia, lactic acid and carboxylic acids as kairomones in the host-seeking behaviour of the malaria mosquito Anopheles gambiae sensu stricto (Diptera: Culicidae). Chem Senses 2005; 30 (2) : 145-152. DOI: 10.1093/chemse/bji010.
166. Snow K. Distribution of Anopheles mosquitoes in the British Isles. Europ Mosq Bull 1998; 1 : 9-13.
167. Sousa CA. Malaria vectorial capacity and competence of Anopheles atroparvus Van Thiel, 1927 (Diptera: Culicidae): implications for the potential re-emergence of malaria in Portugal. PhD Thesis, New University of Lisbon, Portugal, 2008, 187 pp.
168. Stegnii VN. Revealing of chromosome races in malarialmosquitoes Anopheles sacharovi (Diptera: Culicidae). Tsitologiya 1976; 18 : 1039-1041 (in Russian).
169. Stegnii VN, Kabanova VM. Cytological study of indigenous populations of the malaria mosquito in the territory of the USSR. I. Identification of a new species of Anopheles in the maculipennis complex by the cytodiagnostic method. Meditsinskaya Parazitologiya I Partazitarnye Bolezni 1976; 45: 192-198 (in Russian). English translation: Mosquito Systematics 1978; 10 : pp. 1-12).
170. Sturm A, Amino R, van de Sand C, Regen T, Retzlaff S, Rennenberg A et al. Manipulation of host hepatocytes by the malaria parasite for delivery into liver sinusoids. Science 2006; 313 (5791) : 1287-1290. DOI: 10.1126/science.1129720.
189. Suzzoni-Blatger J, Sevin A. Étude de la chétotaxie larvaire du complexe maculipennis (Diptera: Culicidae) dans la région Toulousaine. Ann Parasitol Hum Comp 1981; 57 : 640-654.
190. Suzzoni-Blatger J, Cianchi R, Bullini L, Coluzzi M. Le complexe maculipennis: critéres morphologiques et enzym-atiques de détermination. Ann Parasitol Hum Comp 1990; 65 : 37-40.
191. Takken W, Geene R, Adam W, Jetten TH, van der Velden JA. Distribution and dynamics of larval populations of Anopheles messeae and A. atroparvus in the

81
delta of the rivers Rhine and Meuse, The Netherlands. Ambio 2002; 31 (3) : 212-218.
192. ter Kuile FO, Parise ME, Verhoeff FH, Udhayakumar V, Newman RD, van Eijk AM et al. The burden of co-infection with human immunodeficiency virus type 1 and malaria in pregnant women in sub-saharan Africa. Am J Trop Med Hyg 2004; 71 (2 Suppl) : 41-54.
193. Tilley L, Dixon MW, Kirk K. The Plasmodium falciparum-infected red blood cell. Int J Biochem Cell B 2011; 43 (6): 839–842. doi: 10.1016/j.biocel.2011.03.012.
194. Tolle MA. Mosquito-borne diseases. Curr Probl Pediatr Adolesc Health Care 2009; 39 (4) : 97-140. doi: 10.1016/j.cppeds.2009.01.001.
195. Toty C, Barré H, Le Goff G, Larget-Thiéry I, Rahola N, Couret D et al. Malaria risk in Corsica, former hot spot of malaria in France. Malaria J 2010; 9 : 231 doi:10.1186/1475-2875-9-231.
196. Umeh R, Oguche S, Oguonu T, Pitmang S, Shu E, Onyia JT, et al. Immunogenicity and safety of the candidate RTS,S/AS01 vaccine in young Nigerian children: A randomized, double-blind, lot-to-lot consistency trial. Vaccine 2014; 32 (48) : 6556-6562. doi: 10.1016/j.vaccine.2014.07.067.
201. Ungureanu E, Shute PG. The value of the wing scales asan aid to the taxonomy of adult Anopheles maculipennis. Proceedings of the Royal Entomological Society of London,Series B, Taxonomy, 1947; 16 : 79-85.
202. Vatandoost H; Abai MR. Irritability of malaria vector, Anopheles sacharovi to different insecticides in a malaria-prone area. Asian Pac J Trop Med 2012; 5 (2) : 113-116. https://doi.org/10.1016/S1995-7645(12)60007-8.
203. Vaughan AM, Aly ASI, Kappe SHI. Malaria parasite pre-erythrocytic stage infection: Gliding and Hiding. Cell Host Microbe 2008; 4 (3) : 209–218. doi: 10.1016/j.chom.2008.08.010.
204. van Thiel PH. Sur l'origine des variations de taille de l'Anopheles maculipennis dans les Pays-Bas. Bull Soc Pathol Exot 1927; 20 : 66-390.
205. Velarde-Rodriguez M, Van den Bergh R, Fergus C, Casellas A, Sanz S, Cibulskis R, et al. Origin of malaria cases: a 7-year audit of global trends in indigenous and imported cases in relation to malaria elimination. Global Health Action 2015; 8: 29133. doi.org/10.3402/gha.v8.29133.
206. Vicente JL, Sousa CA, Alten B, Caglar SS, Falcutá E, Latorre JM et al. Genetic and phenotypic variation of the malaria vector Anopheles atroparvus in southern Europe. Malaria J 2011; 10: 5. https://doi.org/10.1186/1475-2875-10-5.
207. Vlachou D, Schlegelmilch T, Runn E, Mendes A, Kafatos FC. The developmental migration of Plasmodium in mosquitoes. Curr Opin Genet Dev 2006; 16 (4) : 384-391. DOI: 10.1016/j.gde.2006.06.012.
208. White NJ. Determinants of relapse periodicity in Plasmodium vivax malaria. Malaria J 2011; 10 : 297. Available from:
a. https://malariajournal.biomedcentral.com/track/pdf/10.1186/1475-2875-10-297. 209. White GB. Systematic reappraisal of the Anopheles maculipennis complex.
Mosquito Systematics 1978; 10 : 3-44.

82
210. Zahar AR. Vector bionomics in the epidemiology and control of malaria. Part II. The WHO European Region and the WHO Eastern Mediterranean Region. Vol. II. Applied field studies. 1st edition. World Health Organisation, Geneva; 1990.
211. Yaghoobi-Ershadi MR, Namazi J, Piazak N. Bionomics of Anopheles sacharovi in Ardebil province, northwestern Iran during a larval control program. Acta Tropica 2001; 78 : 207-215. DOI: 10.1016/S0001-706X(01)00080-8.
212. Yamauchi LM, Coppi A, Snounou G, Sinnis P. Plasmodium sporozoites trickle out of the injection site. Cell Microbiol 2007; 9 (5) : 1215-1222. DOI: 10.1111/j.1462-5822.2006.00861.x.
213. Yi H. Effects of global warming on mosquitoes & mosquito‐borne diseases and the new strategies for mosquito control. Entomol Res 2014; 44 (6) : 215-235 . DOI: 10.1111/1748-5967.12084.
214. *** World Health Organisation. Information for travellers. 2018. [Online]. Available from: http://www.who.int/malaria/travellers/en/.
215. *** World Health Organization. Malaria. [Online]. Available from: http://www.who.int/mediacentre/factsheets/fs094/en/.
216. *** European Centre for Diseases Prevention and Control. Annual Epidemiological Report 2015. Malaria Stocholm, ECDC, 2016.
217. *** European Centre for Diseases Prevention and Control. Epidemiological update: Malaria in Greece, 20 July 2012. In: Communicable Disease Threats Report. Week 31, 29 July-4 August 2012. Available from:
a. https://ecdc.europa.eu/sites/portal/files/media/en/publications/Publications/CDTR_ECDC_20120803.pdf.
218. *** Institutul Naţional de Sănătate Publică. Centrul Naţional de Supraveghere şi Control al Bolilor Transmisibile. Analiza evoluţiei bolilor transmisibile aflate în supraveghere. Raport pentru anul 2015. Bucureşti, 2016, pg. 48-50. [National Institute of Public Health. National Center for Communicable Disease Control and Control. Analysis of the evolution of communicable diseases under surveillance. Report for the year 2015. Bucharest, 2016, pp. 48-50].
219. *** World Malaria Report 2017. World Health Organization, Geneva; 2017. Licence: CC BY-NC-SA 3.0 IGO. Available from:
a. http://apps.who.int/iris/bitstream/handle/10665/259492/9789241565523-eng.pdf?sequence=1.
220. *** World Malaria Report 2016. World Health Organization, Geneva; 2016. Licence: CC BY-NC-SA 3.0 IGO. Available from:
a. http://apps.who.int/iris/bitstream/handle/10665/252038/9789241511711-eng.pdf?sequence=1.
221. *** World Health Organization . World Malaria Report 2012. World Health Organization, WHO Press, Geneva; 2012. Available from:
222. http://www.who.int/malaria/publications/world_malaria_report_2012/en/ 223. *** World Health Organisation. Global technical strategy for malaria 2016-
2030. WHO Press, Geneva, Switzerland, 2015. Available from: a. https://www.who.int/malaria/publications/atoz/9789241564991/en/ 224. ***World Health Organization. Guidelines for the Treatment of Malaria. 3rd
edition. WHO Press, Geneva, Switzerland, 2015. Available from:

83
a. http://apps.who.int/iris/bitstream/handle/10665/162441/9789241549127_eng.pdf?sequence=1.
225. *** World Health Organisation. Strategy for malaria elimination in the Greater Mekong Subregion: 2015-2030. WHO Press, Geneva, Switzerland, 2015. Available from:
226. http://iris.wpro.who.int/bitstream/handle/10665.1/10945/9789290617181_eng.pdf.
227. *** World Health Organization. Pesticides and their application : for the control of vectors and pests of public health importance. 6th edition. WHO Press, Geneva, 2006. Available from: http://www.who.int/iris/handle/10665/69223.
228. *** World Health Organization. The Africa Malaria Report 2003. World Health Organization/UNICEF, 2003. Available from:
a. http://apps.who.int/iris/bitstream/handle/10665/67869/WHO_CDS_MAL_2003.1093.pdf?sequence=1.

84
SOURCES FOR FIGURES AND TABLES - ADDITIONAL REFERENCES:
1. http://www.microbiologynotes.com/life-cycle-of-plasmodium-vivax/
2. https://ecdc.europa.eu/en/disease-vectors/facts/mosquito-factsheets/anopheles-sacharovi
3. http://mosquito-taxonomic-inventory.info/file/5212 4. https://cameronwebb.wordpress.com/tag/anopheles-
gambiae/ 5. http://wwwnc.cdc.gov/travel/yellowbook/2012/chapter-3-
infectious-diseases-related-to-travel/malaria.htm 6. https://www.statista.com/chart/1758/estimated-number-
of-deaths-caused-by-malaria-worldwide/ 7. https://www.dailystar.co.uk/health/592904/Malaria-
symptoms-tablets-treatment-prevention 8. https://www.lybrate.com/topic/symptoms-of-malaria-and-
chikunguniya/dd771f92ae9155433ef28c1ecc43adc7 9. http://zeenews.india.com/health/how-to-fight-dengue-
malaria-flu-more-symptoms-prevention-2046312. 10. http://adulldayatwork.blogspot.com/2011/10/25-october-
2011-tuesday-rapid-malaria.html 11. https://labmedicineblog.com/2018/06/19/hematology-
case-study-a-51-year-old-woman-with-fever-and-chills/ 12. https://elifesciences.org/articles/29198 13. http://www.who.int/immunization/programmes_systems/i
nterventions/malaria_llins/en/