Embed Size (px)
Citation preview

388 A.M. Adnan : Manajemen Musuh Alami Hama Utama Jagung
MANAJEMEN MUSUH ALAMI HAMA UTAMA JAGUNG
A.M. Adnan
Balai Penelitian Tanaman Serealia
ABSTRAK
Swasembada jagung sering terkendala oleh serangan hama dan penyakit tanaman. Hama penggerek batang jagung (Ostrinia furnacalis), penggerek tongkol (Helicoverpa armigera), ulat grayak (Spodopteram litura), lalat bibit (Atherigona sp.), belalang (Locusta migratoria), kutu daun, dan tikus (Rattus Sp.) merupakan organisme pengganggu tanaman (OPT) yang sering ditemui. Pemanfaatan musuh alami adalah salah satu cara pengendalian hama ini, namun petani seringkali tidak puas atas kinerja musuh alami. Salah satu faktor ketidakberhasilan penggunaan agen pengendali hayati pada areal pertanian adalah pengelolaan musuh alami yang tidak bijaksana. Jika faktor ini dapat ditangani dengan baik, maka kerusakan tanaman dapat ditekan yang pada akhirnya produksi dapat ditingkatkan dan ramah lingkungan. Pengelolaan musuh alami dimaksudkan untuk meningkatkan keefektifan agen pengendali hayati dalam mengendalikan serangga hama dan mengurangi penggunaan pestisida sintetik yang memiliki pengaruh negatif terhadap lingkungan.
Kata kunci: Manajemen, musuh alami, hama utama jagung
PENDAHULUAN
Kebutuhan akan jagung di Indonesia terus meningkat dari tahun ke tahun. Kurun waktu 2005 – 2009 tergambar bahwa konsumsi jagung nasional pada tahun 2005 sebesar 12,26 juta ton Pipilan Kering (PK) menjadi 12,50 juta ton PK pada tahun 2006, kemudian menjadi 13,29 ton PK, tahun 2007, 16,32 juta ton PK di tahun 2008, dan 17,66 juta ton PK di tahun 2009 (Tabel 1).
Sementara Produksi jagung nasional pada tahun 2005 sebesar 12,52 juta ton PK. Meskipun pada tahun 2006 produksinya sempat turun menjadi
11,61 juta ton PK, pada tahun 2007 mampu ditingkatkan kembali menjadi 13,28 juta ton PK. Selanjutnya terus meningkat, menjadi 16,32 juta ton PK pada tahun 2008 dan menjadi 17,66 juta ton PK pada tahun 2009.
Dengan demikian, terdapat surplus sebesar 0,26 juta ton PK pada tahun 2005, tetapi menjadi minus 0,89 juta ton PK pada tahun 2006. Karena produksi berhasil ditingkat kembali pada tahun 2007 sehingga terdapat surplus sebesar 0,07 juta ton PK. Selanjutnya menjadi surplus sebesar 1,66 juta ton PK pada tahun 2008 dan surplus sebesar 1,98 juta ton PK pada tahun 2009 (Tabel 1).
Tabel 1. Produksi dan Konsumsi Jagung, 2005 – 2009.
Uraian 2005 2006 2007 2008 2009 Produksi (ton PK) 12.523.894 11.609.463 13.287.527 16.317.252 17.659.067 Konsumsi (ton PK) 12.262.385 12.504.949 13.217.244 14.659.525 15.680.459 - Benih 92.463 77.790 87.128 85.327 82.620 - Pakan 3.530.000 3.640.000 3.810.000 4.230.000 4.540.000 - Konsumsi Langsung 864.014 894.435 947.696 964.367 990.261 - Tercecer, susut 1.252.389 1.160.946 1.328.753 1.631.725 1.765.907 - Lainnya 6.525.515 6.731.778 7.043.667 7.748.106 8.301.671 Surplus/Devisit (ton PK) 259.509 - 895.468 70.283 1.657.727 1.978.608
Sumber: BPS 2009

389 Seminar Nasional Serealia 2011
Oleh karenanya dalam kurun waktu 2010–2014, Program Kementerian Pertanian Republik Indonesia yang dituangkan dalam empat sukses Kementerian Pertanian menempatkan jagung sebagai salah satu komoditas prioritas untuk swasembada dan swasembada berkelanjutan. Hal ini mengindikasikan bahwa jagung masih menjadi komditas penting untuk pemenuhan pangan, pakan, dan industri di Indonesia.
Kendala mempertahankan swasembada jagung berkelanjutan adalah masalah abiotis dan biotis. Abiotis dapat berupa adanya perubahan iklim global dan kesuburan tanah, sementara biotis dapat berupa hama dan penyakit tanaman jagung (Swastika et al. 2004).
Serangga pengganggu tanaman jagung yang sering ditemui dipertanaman adalah penggerek batang jagung (Ostrinia furnacalis), penggerek tongkol (Helicoverpa armigera), ulat grayak (Spodopteram litura), lalat bibit (Atherigona sp.), belalang (Locusta migratoria), Kutu daun dan tikus (Rattus Sp.) (Metcal and Metcalf 1993). Kerusakan pada tanaman yang ditimbulkan oleh hama-hama ini berkisar antara 5 hingga 100%.
Pemanfaatan pestisida kimiawi untuk mengendalikan serangga hama tersebut telah mulai ditinggalkan karena petani merasakan adanya dampak negatif tergadap lingkungan dan mulai beralih dengan memanfaatkan biopestisida baik nabati maupun hayati. Pengendalian dengan memanfaatkan musuh alami telah banyak dilakukan seperti pengaplikasian Trichogramma spp,. Beauveria bassiana, Metharizium anisopliae, Cecopet (Euborellia annulata), dan nematoda Steinernema sp.
Aplikasi musuh alami untuk mengendalikan hama utama jagung terkadang dirasakan kurang efektif, hal tersebut dimungkinkan karena manajemennya tidak baik. Oleh karena itu, makalah ini akan membahas menajemen musuh alami hama utama jagung, sehingga dengan mengetahui cara pengelolaannya, maka keefektifan musuh alami dapat ditingkatkan.
KONDISI OPTIMAL BAGI HAMA UTAMA JAGUNG
Penggerek batang jagung (Ostrinia furnacalis)
Asia merupakan wilayah penyebaran O. furnacalis terutama ditemukan diseluruh Asia Tenggara, Asia Tengah, Asia Timur dan Australia (Mutuura dan Munroe 1970). Di Indonesia serangga ini menyebar luas di Papua, Nusa Tenggara, Sulawesi, dan Sumatera (Waterhouse 1993).
Fase paling merusak serangga hama ini adalah stadia larva dimana Larva O. furnacalis menyerang semua bagian tanaman jagung. Kehilangan hasil terbesar ketika kerusakan terjadi pada fase reproduktif (Kalshoven 1981). Serangga ini mempunyai karakteristik kerusakan pada setiap bagian tanaman jagung yaitu lubang kecil pada daun, lubang gorokan pada batang, bunga jantan, atau pangkal tongkol, batang dan tassel yang mudah patah, tumpukan tassel yang rusak, dan rusaknya tongkol jagung (Lee et al. 1980).
Imagonya mulai meletakkan telur pada tanaman yang berumur dua minggu. Puncak peletakan telur terjadi pada stadia pembentukan bunga jantan sampai keluarnya bunga jantan. Betina penggerek batang lebih suka meletakkan telur di bawah permukaan daun utamanya pada daun ke 5 sampai daun ke 9 (Legacion and Gabriel 1988). Jumlah telur yang diletakkan tiap kelompok beragam antara 30 sampai 50 butir atau bahkan lebih dari 90 butir (Kalshoven 1981). Seekor ngengat betina mampu meletakkan telur 300-500 butir.(Lee et al. 1980). Inang alternatif dari spesies ini adalah sorgum, kedelai, mangga, okra, tomat, tembakau, lada, tebu, kapas, jahe, dan rumput-rumputan (PGCPP 1987).
Kondisi lingkungan yang optimal untuk perkembangan hama ini adalah ada wilayah-wilayah dengan iklim tropis.
Penggerek tongkol jagung (Helicoverpa armigera Hbn.)
Fase paling merusak dari spesies ini juga adalah pada stadia larva. Larva

390 A.M. Adnan : Manajemen Musuh Alami Hama Utama Jagung
spesies ini terdiri dari lima sampai tujuh instar, tetapi pada umumnya terdiri dari enam instar dengan pergantian kulit (moulting) setiap instarnya dua sampai empat hari. Larva serangga ini memiliki sifat kanibalisme sehingga hal ini merupakan salah satu faktor yang menekan perkembangan populasinya (CPC 2005).
Imago betina H. armigera meletakkan telur pada pucuk tanaman dan bilamana tongkol sudah mulai keluar maka telur tersebut diletakkan pada rambut jagung. Rata-rata produksi telur imago betina adalah 730 butir dengan masa oviposisi 10 sampai 23 hari.
Kondisi lingkungan seperti suhu dan kualitas makanan di alam menetukan periode perkembangan larva. Khususnya pada jagung, masa perkembangan larva pada suhu 24 sampai 27,2oC adalah 12,8 sampai 21,3 hari, telur menetas dalam tiga hari setelah diletakkan pada suhu 22,5oC dan dalam sembilan hari pada suhu 17oC Pada kondisi lingkungan mendukung, dan fase pupa bervariasi dari enam hari pada suhu 35oC sampai 30 hari pada suhu 15oC. (Kalshoven 1981).
Pada kondisi yang tidak memungkinkan H. armigera mengalami diapause atau sering disebut diapause pupa fakultatif. Diapause pupa dapat berlangsung beberapa bulan bahkan dapat lebih dari satu tahun.
Ulat grayak (Spodoptera litura.)
Asia Tenggara dan spesies ini juga terdistribusi luas keseluruh Asia tropis dan Asia Sub tropis, Australia, dan pulau-pulau di Pasifik merupakan wilayah penyebaran serangga hama ini (Kranz et al. 1977).
Spesies ini adalah serangga polipagous. Tanaman inangnya selain jagung adalah tomat, kapas, tembakau, padi, kakao, jeruk, ubi jalar, kacang tanah, jarak, kedelai, kentang, kubis, dan bunga matahari (Holloway 1989).
Larva yang terdiri dari 6 instar dan instar terakhir mempunyai berat mencapai 800 mg dan menghabiskan 80% dari total konsumsi makanannya
merupakan fase paling merusak (Kalshoven 1981). Larva bersembunyi dalam tanah pada siang hari dan baru aktif pada malam hari, kecuali S, exempta yang juga aktif pada siang hari.
Pada kondisi suhu 8oC, telur-telur serangga hama ini masih dapat menetas dan pada suhu 10oC, pupa masih masih dapat bertahan. Sementara stadia larva untuk berkembang secara optimal yaitu pada suhu antara 37oC sampai 40oC (Holloway 1989). Kondisi suhu ini sangat dimungkinkan bagi Spodoptera litura untuk berkembang dengan baik di wilayah Indonesia. Lalat Bibit (Atherigona sp)
Stadia larva merupakan fase merusak dari Atherigona sp. Larva berukuran panjang hingga sembilan mm yang berwarna putih krem pada awalnya dan selanjutnya menjadi kuning hingga kuning gelap. Pupa terdapat pada pangkal batang dekat atau di bawah permukaan tanah. Puparium berwarna coklat kemerah-merahan sampai coklat dengan ukuran panjang 4,1 mm. Imago betina mulai meletakkan telur tiga sampai lima hari setelah kawin dengan jumlah telur tujuh sampai 22 butir atau bahkan hingga 70 butir. Imago betina meletakkan selama tiga sampai tujuh hari. Lama hidup serangga betina dua kali lebih lama daripada jantan. Lama siklus hidup dari telur hingga dewasa (CPC 2005).
Kondisi iklim berupa kelembaban dan curah hujan tinggi mendukung perkembangan spesies ini. Jika kondisi sangat kering, telur akan gagal menetas atau larva mati sebelum dia mampu melakukan penetrasi batang. Penetasan dan aktivitas imago terjadi selama kondisi dingin dalam satu hari.
Kutu Daun (Aphis maidis)
Kutu daun membentuk koloni yang besar pada daun yang meliputi betina yang berproduksi secara partenogenesis (tanpa kawin). Seekor betina yang tidak bersayap mampu melahirkan rata-rata sebanyak 68,2 ekor nimfa, sementara betina bersayap 49 nimfa (Adams dan

391 Seminar Nasional Serealia 2011
Drew 1964). Lama hidup imago adalah 4-12 hari (Ganguli dan Raychaudhuri 1980). Umumnya, stadia nimfa terdiri dari empat instar (Kring 1985). Stadium nimfa terjadi selama 16 hari pada suhu 15oC, sembilan hari pada suhu 20oC, dan lima hari pada suhu 30oC. Serangga ini lebih senang berada pada suhu yang hangat dibandingkan pada suhu yang dingin. Mau dan Kessing (1992) melaporkan bahwa imago lebih aktif di lapangan pada suhu 17 oC dan 27oC.
BEBERAPA MUSUH ALAMI DAN KEFEKTIFANNYA
Musuh alami atau natural enemies berperan untuk mengendalikan populasi hama di pertanaman sehingga kerusakan yang ditimbulkan dapat ditekan atau bahkan dikurangi sehingga tidak melampaui ambang ekonomi. Musuh alami hama utama jagung dapat dilihat pada Tabel 2.
Tricogramma spp.
Serangga yang digolongkan sebagai musuh alami ini masuk kedalam famili Trichogrammatidae dari ordo Hymenoptera. Parasitoid telur ini mempunyai ukuran sangat kecil dengan panjang 0,1 – 0,5 mm bergantung spesies. Trichogramma ini umumnya memarasit telur dari ordo lepidoptera
meskipun juga dilaporkan dapat memarasit telur dari ordo Neuroptera dan Coleoptera (CPC 2005).
Parasitoid ini menghabiskan siklus hidupnya di dalam telur inangnya dan waktu yang dibutuhkan yaitu 7 hari pada suhu 28oC dan 8–9 hari pada musim hujan (Alba 1989). Selanjutnya Bayrami dan Kornosor (1998) melaporkan bahwa suhu optimal untuk perkembangan Trichogramma spp ini yaitu pada suhu antara 20–30oC dengan kelembaban nisbih 60%.
Parasitoid ini dapat diperbanyak pada telur Corcyra cephalonica dengan prosedur sebagai berikut: Parasitoid Trichogramma spp. dikumpulkan dari lapangan yaitu dengan mengambil telur Ostrinia furnacalis yang terparasit di pertanaman jagung petani dan di bawah ke laboratorium, diamati hingga parasitoid telur keluar. Perbanyakan Trichogramma ini dilakukan dengan menggunakan inang pengganti yaitu telur Corcyra cephalonica. Telur inang pengganti ini sebelum direkatkan pada pias kertas terlebih dahulu disinari ultra violet selama 30 menit dengan maksud embrio dalam telur tidak berkembang. Setelah itu telur direkatkan pada pias ukuran 7 x 2 cm dengan menggunakan lem povinal. Selanjutnya pias dimasukkan pada tabung reaksi
Tabel 2. Musuh Alami Hama Utama Jagung dan Stadia yang dipredasi atau diparasit.
Musuh Alami Serangga Target Stadia Trichogramma spp. Ostrinia furnacalis Telur Helicoverpa armigera Telur Atherigona sp. Telur Cecopet (Euborellia annulata) Ostrinia furnacalis Larva dan Pupa Helicoverpa armigera Telur dan Larva Baeuveria bassiana Ostrinia furnacalis Larva Helicoverpa armigera Larva Aphis sp. Nimfa dan Dewasa Spodoptera litura Larva Metarhizium anisopliae Spodoptera litura Larva Helicoverpa armigera
Ostrinia furnacalis Larva Larva
Nematoda Steinernema sp. Ostrinia furnacalis Larva Spodoptera litura Larva
Sumber: CPC 2005

392 A.M. Adnan : Manajemen Musuh Alami Hama Utama Jagung
berukuran 15 x 2 cm, di dalam tabung tersebut sudah terdapat telur Trichogramma yang siap menjadi imago, dan akan memarasit telur Corcyra cephalonica pada pias. Pada hari ketujuh atau delapan Trichogramma siap menjadi imago, kegiatan ini dilakuakn terus hingga diperoleh Trichogramma yang cukup untuk dilepas di lapangan (Nonci dan Masmawati 2005).
Kemampuan memarasit Trichogramma spp terhadap telur Ostrinia furnacalis dapat mencapai 81,98% (Tabel 3) ditingkat laboratorium. Pernah pula dilaporkan oleh Pabbage, et. al., 1999 bahwa Trichogramma spp. efektif memarasit penggerek batang jagung di laboratorium dengan tingkat parasitasi mencapai 97,68%.
Nonci et al. 2000 melaporkan bahwa tingkat parasitasi parasitoid telur penggerek batang di daerah-daerah
sentra produksi jagung di Sulawesi Selatan berkisar antara 71,56 ssampai 89,80%. Pengujian lain yang dilakukan oleh peneliti Balitsereal memperlihatkan bahwa parasitoid telur ini efektif mengendalikan penggerek batang di lapangan dengan parasitasi dapat mencapai 91,96% (Tabel 4).
Trichogramma spp. efektif pula dalam mengendalikan penggerek tongkol (Helicoverpa armigera). Keefektifannya tersebut berkisar antara 54,38% sampai 92,37% bergantung spesies di laboratorium (Tabel 5).
Parasitoid telur ini pula efektif dalam mengendalikan penggerek tongkol jagung (Helicoverpa armigera) di lapangan. Pabbage et al. 2000 melaporkan bahwa persentase telur penggerek tongkol yang terparasit di lapangan berkisar antara 71,67% sampai 100% (Tabel 6).
Tabel 3. Persentase telur penggerek batang Ostrinia furnacalis yang terparasit kelompok telur Trichogramma spp.
Pias Kelompok Telur O. furnacalis
Telur Penggerek batang Terparasit (%) Rata-rata (%) 2 5 8
15 ekor 42,64 58,61 35,06 45,44 30 ekor 80,95 85,67 66,81 77,81 45 ekor 79,82 83,64 82,48 81,98 Tanpa Trichogramma 0,0 0,0 0,0 0,0
Sumber: Nonci dan Pabbage (2000)
Tabel 4. Rata-rata tingkat parasitasi Trichogramma spp. terhadap telur O. furnacalis.
Perlakuan Tingkat Parasitasi pada pengamatan (%)
5 MST 6 MST 7 MST 8 MST 4, 5, 6, 7 dan 8 MST 0,00 15,28 25,63 54,24 4, 5, 6, dan 7 MST 5,28 0,00 31,43 51,98 5, 6, 7, dan 8 MST 13,50 10,29 0,00 91,96 6, 7, dan 8 MST 3,22 10,29 58,83 64,50 Berdasarkan gejala serangan di lapangan (populasi telur 1 kelompok/30 tanaman)
17,04 13,67 69,84 71,06
Kontrol (Tanpa Pelepasan) 17,62 6,40 60,24 33,33 Sumber: Nonci et al. 2001 Ket: MST: Minggu Setelah Tanam

393 Seminar Nasional Serealia 2011
Tabel 5. Daya parasitasi beberapa spesies Trichogramma spp. pada telur penggerek tongkol (Helicoverpa armigera) di Laboratorium.
Spesies Trichogramma
Telur penggerek
tongkol terparasit (%)
Telur penggerek tongkol terparasit mengeluarkan
Trichogramma (%)
Jumlah imago Trichogramma per
butir telur penggerek tongkol
T. bactrae-bactrae 73,33 75,31 2,41 T. australicum 90,44 51,53 1,79 T. japonicum (T) 54,38 48,56 1,84 T. bactrae fumata 95,18 89,81 1,32 T. japonicum (M) 72,75 70,59 1,35 T. evanescens 92,27 68,41 2,16
Sumber: Pabbage et al. 2000
Tabel 6. Rata-rata jumlah telur Helicoverpa armigera pada jambul per tongkol dan
persentase telur yang terparasit.
Perlakuan Jumlah Telur H. armigera per
tongkol (butir) Telur H. Armigera terparasit
(%) 56 HST 63 HST 56 HST 63 HST
5, 7, dan 9 MST 5,00 0,67 66,67 100 5 dan 7 MST 6,67 0,67 60,00 100 5 dan 9 MST 6,67 2,33 70,00 71,67 7 dan 9 MST 6,33 0,67 52,61 100 Berdasarkan keberadaan Telur H. Armigera
6,33 3,00 57,98 77,61
Kontrol 6,33 3,67 57,98 100 Sumber: Pabbage et al. 2000
Cecopet (Euborellia annulata)
Cecopet (E. annulata) merupakan predator larva dan pupa penggerek batang jagung dan predator telur dan larva penggerek tongkol. Telur serangga ini yang baru diletakkan berbentuk oval, mengkilap dan putih. Ukurannya bertambah setiap hari sampai pada hari ke empat sampai dua kali lipat ukurannya dari ukuran semula. Telur yang baru diletakkan berukuran +0,95 mm x 1,55 mm. Sebelum menetas, telur berbentuk mutiara, putih dan tembus cahaya serta perkembangannya menjadi embrio mulai tampak. (Javier and Morallo 1991).
Nimfa instar pertama adalah berwarna keputih-putihan tetapi beberapa jam kemudian akan berwarna gelap. Nimfa instar pertama dan kedua berkerumun sepanjang waktu. Setiap instar sama bentuk morfologinya pada tiap pergantian kulit dan yang bervariasi hanya ukurannya yang berbeda. Jenis
kelaminnya hanya dapat dibedakan pada fase dewasa. (Javier and Morallo 1991).
Selanjutnya Javier dan Morallo, 1991 melaporkan bahwa jumlah instar nimfa bervariasi tergantung pada ketersediaan makanan. Serangga yang makanannya kurang maka biasanya mengalami instar nimfa sampai lima sedangkan serangga yang makanannya tersedia cukup hanya sampai pada empat instar. Untuk menjadi imago, cecopet mengalami empat kali pergantian kulit.
Imago jantan dan bertina E. annulata berwarna hitam menkilap, antena terdiri dari 17 segmen dengan warna putih pada ujung antena. Imago betina meletakkan telur sampai 200 butir selama hidupnya. Tipe alat mulut menggigit mengunyah dan bermetamorfosis sederhana (Situmorang dan Gabriel 1988).
Perbanyakan cecopet ini relatif mudah dengan mengikuti tata cara sebagai berikut: Imago cecopet E. annulata yang telah diperoleh dari lapangan di masukkan ke dalam wadah

394 A.M. Adnan : Manajemen Musuh Alami Hama Utama Jagung
plastik yang berukuran 27 x 34 x 7 cm yang terlebih dahulu telah diberikan tanah dan pasir dengan perbandingan 3 : 1 setinggi 2 cm, kemudian di semprot dengan menggunakan sprayer untuk menjaga kelembaban media. E. annulata yang baru bertelur dipindahkan ke wadah yang baru, kemudian setiap pergantian kulit, dipindahkan ke wadah yang berbeda untuk digunakan.
Adnan dan Syamsuddin (2010) melaporkan bahwa mass produksi E. annulata dapat dilakukan dalam media percampuran antara biskuit anjing dan jenggel jagung dengan perbandingan yang paling baik adalah 60% biskuit anjing dan 40% janggel jagung (Tabel 7).
Pelepasan predator ini relatif efektif dalam menekan kerusakan
tanaman akibat hama penggerek batang (Gambar 1). Selanjutnya Adnan (2010) melaporkan bahwa pada pengamatan 10 minggu setelah tanam (MST) dan 12 MST, kerusakan tanaman dapat ditekan hingga 2,72 dan 4,22. Penekanan kerusakan tanaman ini dimungkinkan dengan adanya predator E. annulata yang dirilis sebanyak 10000 ekor/ha.
Kefektifan predator ini pula diindikasikan pada kurangnya jumlah lubang gerekan dan panjang gerekan pada pertanaman jagung sebagai akibat dari pelepasan agen pengendali hayati ini (Tabel 8). Adnan (2010) melaporkan bahwa pelepasan 10000 ekor/ha dapat menekan jumlah lubang gerekan dan panjang gerekan masing-masing 2,25 dan 2,30 cm.
Tabel 7. Rata-rata jumlah imago jantan dan betina predator cecopet (Euborellia annulata)
pada pengamatan 30 dan 60 hari setelah penempatan imago (HSPI).
Perlakuan HSPI
30 60 ♂ ♀ ♂ ♀
Kontrol (P0) 150,5 292,5 282,75 595,25 50% Biskuit Anjing + 50% Jenggel Jagung (P1) 240 695,25 480 1392 60% Biskuit Anjing + 40% Jenggel Jagung (P2) 242,25 697 483,75 1396 40% Biskuit Anjing + 60% Jenggel Jagung (P3) 238,75 690 469,75 1385
Sumber: Adnan dan Syamsuddin, 2010
Sumber: Adnan (2010)
Gambar 1. Persentase Tanaman Rusak akibat hama penggerek batang pada tiap perlakuan predator pada pengamatan 10 MST dan 12 MST.

395 Seminar Nasional Serealia 2011
Tabel 8. Rata-rata jumlah lubang gerekan dan panjang gerekan (cm) pada tiap perlakuan pelepasan Euborellia.
Perlakuan Pengamatan
∑ Lubang Panjang Gerekan (cm) 0 ekor/ha (Kontrol) 12,00a 7,79a
1000 ekor/ha 9,50b 6,79b 5000 ekor/ha 5,50c 5,25c
10000 ekor/ha 2,25d 2,30d 20000 ekor/ha 2,50d 2,64d
Angka yang diikuti dengan huruf yang sama tidak bebeda nyata pada taraf 0,05 DMRT Sumber: Adnan (2010)
Tabel 9. Mortalitas larva O. furnacalis pada beberapa konsentrasi konidia diamati pada 4
interval pengamatan
Konsentrasi Cendawan (konidia/ml)
Mortalitas larva (%) larva instar ke-
2 3 4 5 Pada 1 HSI 5 x 107 7,5 5,0 7,5 22,5 5 x 106 2,5 5,0 12,5 10,0 5 x 105 5,0 7,5 5,0 2,5 5 x 104 2,5 7,5 2,5 2,5 5 x 103 2,5 2,5 2,5 5,0 Kontrol 0,0 0,0 0,0 0,0 Pada 3 HSI 5 x 107 30,0 17,5 37,5 32,5 5 x 106 10,0 10,0 17,5 10,0 5 x 105 12,5 17,5 20,5 17,5 5 x 104 5,0 10,0 17,0 10,0 5 x 103 10,0 5,0 10,0 7,5 Kontrol 0,0 0,0 0,0 0,0 Pada 5 HSI 5 x 107 45,0 45,0 50,0 47,5 5 x 106 30,0 37,5 40,0 27,5 5 x 105 25,0 37,5 35,0 17,5 5 x 104 7,5 17,5 22,5 10,0 10,0 5,0 17,5 7,5 Kontrol 0,0 0,0 0,0 0,0 Pada 7 HSI 5 x 107 62,5 55,0 57,5 55,0 5 x 106 45,0 30,0 30,0 27,5 5 x 105 35,0 47,5 47,5 20,0 5 x 104 17,5 12,5 12,5 10,0 5 x 103 10,0 12,5 12,5 2,5 Kontrol 0,0 0,0 0,0 0,0 HSI = Hari Setelah Inokulasi Sumber: Tandiabang et al. 2005
Beuveria bassiana
Pemanfaatan cendawan Beuveria bassiana sebagai agen pengendali hayati organisme pengganggu tanaman telah banyak dilakukan dan efektif terutama
dalam mengendalikan penggerek batang dan penggerek tongkol jagung. Cendawan ini menginfeksi serangga target melalui kontak kutikula, mulut, sistem pencernaan, dan pernafasan serangga (Tanada dan Kaya 1993).
396 A.M. Adnan : Manajemen Musuh Alami Hama Utama Jagung
Tabel 9 memperlihatkan bahwa konsentrasi 5 x 107 efektif untuk mengendalikan penggerek batang jagung dengan tingkat mortalitas dapat mencapai 62,5% sehingga dapat disimpulkan bahwa Beauvveria bassiana menjanjikan sebagai salah satu agen pengendali untuk penggerek batang jagung.
Faktor-faktor yang berpengaruh dalam perkembangan cendawan ini ada tiga komponen yang sangat menentukan yaitu patogen, lingkungan dan komponen makanan. Faktor-faktor lingkungan yang sangat berperan adalah suhu, kelembaban relatif dan intensitas cahaya. Untuk suhu, cendawan B. bassiana dapat berkembang pada 20oC – 30oC, tetapi perkembangan optimal untuk cendawan ini yaitu 23oC – 25oC. Suhu kritis yang dapat menghambat perkembangan agen pengendali ini adalah pada kondisi 5oC dan 35oC dan akan marti pada suhu 50oC. Kelembaban relatif yang optimum untuk perkembangan B. bassiana adalah 70 – 100% (Tandiabang et al. 2005).
Metarhizium anisopliae
Keefektifan cendawan M. Anisopliae ini dipengaruhi oleh konsentrasi konidia dan perkembangan larva O. furnacalis. Yasin et al. (2000)
melaporkan bahwa tingkat mortalitas larva penggerek batang instar II pada perlakuan M. Anisopliae konsentrasi 108 semakin meningkayt seiring dengan bertambahnya hari pengamatan yaitu 10 sampai 72,5%, sedangkan mortalitas larva instar II dan IV berturut-turut adalah 2,5 sampai 50% dan 0 sampai 35%.
Pengujian lapangan juga dilaporkan bahwa cendawan ini mampu mengendalikan penggerek batang terindikasi rendahnya kerusakan daun (13,25%) dan bunga jantan (5,305) dibandingkan dengan kontrol pada kerusakan daun dan bunga jantan berturut-turut adalah 24,25% dan 27% pada 6 Minggu Stelah Tanam (MST) (Baco dan Yasin 2000).
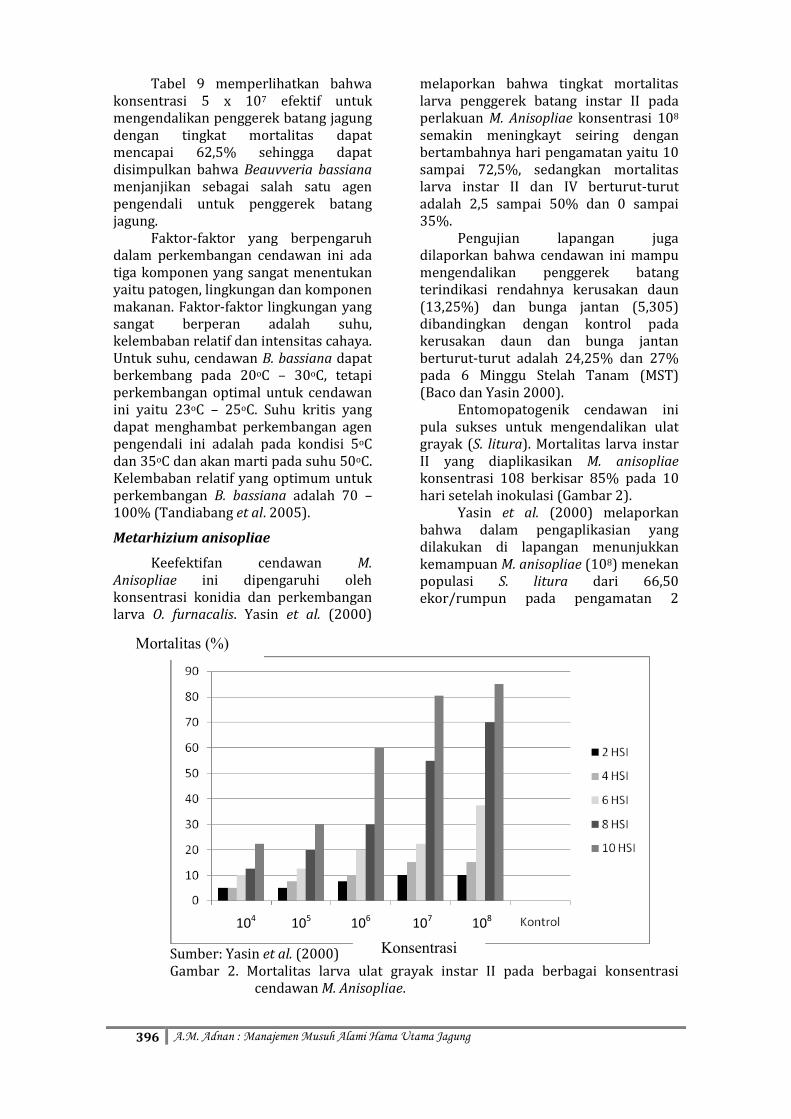
Entomopatogenik cendawan ini pula sukses untuk mengendalikan ulat grayak (S. litura). Mortalitas larva instar II yang diaplikasikan M. anisopliae konsentrasi 108 berkisar 85% pada 10 hari setelah inokulasi (Gambar 2).
Yasin et al. (2000) melaporkan bahwa dalam pengaplikasian yang dilakukan di lapangan menunjukkan kemampuan M. anisopliae (108) menekan populasi S. litura dari 66,50 ekor/rumpun pada pengamatan 2
Sumber: Yasin et al. (2000) Gambar 2. Mortalitas larva ulat grayak instar II pada berbagai konsentrasi
cendawan M. Anisopliae.
Konsentrasi
Mortalitas (%)
104 105 106 107 108
104

397 Seminar Nasional Serealia 2011
MST menjadi 7,00 ekor/rumpun pada 10 MST dengan kerusakan daun 32,50% (2 MST) menjadi 4,25% (10MST).
Keefektifan entomopatogen cendawan ini pula sangat dipengaruhi oleh kondisi lingkungan seperti curah hujan dan sinar matahari khususnya UV yang dapat merusak konidia cendawan (Tanada dan Kaya 1993; Wahyunendo 2002; Prayogo 2004; Suharsono dan Prayogo 2005).
Steinernema sp.
Steinernema sp. ini dikenal pula dengan nama nematoda entomopatogen atau nematoda patogen serangga. Nematoda ini dapat mengendalikan ordo lepidoptera, diptera, orthoptera, dan coleoptera (Kaya dan Gaugler 1993; Smart 1995). Kefektifan nematoda ini dalam mengendalikan serangga hama sangat dipengaruhi oleh besar pori tanah, kelembaban, suhu, ketersediaan air (Lewis 2001).
Dari ordo Lepidoptera seperti penggerek tongkol, penggerek batang jagung, dan ulat grayak sangat efektif dikendalikan dengan menggunakan nematoda ini. Glazers dan Navon (1990) menjelaskan bahwa mortalitas larva penggerek tongkol (H. armigera) dapat mencapai 98% (Gambar 3).
Adnan (2004) meneliti bahwa mortalitas penggerek batang jagung antara 50 sampai 73% dengan letal time 50% (LT50) berada pada 2,65 sampai 4,02 hari (Tabel 10). Ching et al.1998 pernah juga melaporkan bahwa instar ketiga dari nematoda entomopatogen memeliki poteginatas yang tinggi terhadap penggerek batang jagung (O. furnacalis). Tingkat patogenitas nematoda ini(LC50) pada instar 3, 4 dan 5 larva penggerek batang jagung adalah berturut turut 7,5, 8,7 dan 16,3 Ijs/ml. Nilai LT50 adalah 1,4, 2,4 dan 4,9 berturut-turut untuk larva instar 3, 4, dan 5.
0
20
40
60
80
100
1 2 3 4 5 6
Nematode Concentration (IJs/larva)
Inse
ct M
ort
ali
ty (
%)
Gambar 3. Mortalitas larrna H. Armigera setelah 48 jam dengan suhu 25oC pada
konsentrasi S. feltiae (Sumber : Glazers dan A. Navon 1990). Tabel 10. Rata-rata Mortalitas dan LT50 O. furnacalis pada konsentrasi 2000 IJs/ml
Steinernematid isolate Los Banos , Pengujian Laboratorium.
Instar Mortalitas (%)
LT50
(hari) Fiducial
Limit Garis Regresi Slope
Pertama 50 4.02 2.90-10.76 Y = 1.39 + 1.82X 1.82 Kedua 55 3.77 2.89-6.84 Y = 0.60 + 2.25X 2.25 Ketiga 63 3.03 2.34-4.72 Y = 1.05 + 2.12X 2.12 Keempat 73 2.65 2.07-3.64 Y = 0.89 + 2.28X 2.28 Kelima 63 2.86 2.30-3.91 Y = 0.35 + 2.53X 2.53
Sumber: Adnan (2004)
0 25 50 100 200 400

398 A.M. Adnan : Manajemen Musuh Alami Hama Utama Jagung
Demikian halnya dengan Ulat grayak yang relatif efektif dikendalikan dengan menggunakan nematoda patogen serangga ini. Adnan (2004) melaporkan bahwa mortalitas larva ulat grayak (S. litura) pada semua instar adalah 90% dengan LT50 berkisar antara 1,62 sampai 2,0 hari (Tabel 11).
Pada pengujian rumah kawat, nematoda patogen serangga ini memberikan mortalitas penggerek batang jagung berkisar antara 34 sampai 54% (Tabel 12), sementara itu mortalitas ulat grayak berkisar antara 36 sampai 58% (Tabel 13) dengan kondisi suhu
pada saat itu 28 – 29oC dengan kelembaban 80 – 83%.
MANAJEMEN MUSUH ALAMI HAMA UTAMA JAGUNG
Salah satu faktor ketidakberhasilan penggunaan agen pengendali hayati di lahan pertanian adalah manajemen dari musuh alami tersebut yang kurang bijaksana. Jika faktor ini dapat ditangani dengan baik, maka kerusakan tanaman dapat ditekan yang pada akhirnya produksi dapat ditingkatkan serta ramah lingkungan, karena manajemen atau pengelolaan musuh alami dimaksudkan untuk meningkatkan kesuksesan atau
Tabel 11. Rata-rata mortalitas dan LT50 Ulat Grayak yang diberi perlakuan 2000 IJs/ml
Steinernematid isolate Los Banos pada pengujian laboratorium.
Instar Mortalitas (%)
LT50
(hari) Fiducial
Limit Garis Regresi Slope
Pertama 90 1.87 1.51 – 2.22 Y = -0.60 + 3.38X 3.38 Kedua 90 1.67 1.33 – 1.99 Y = -0.50 + 3.43X 3.43 Ketiga 90 2.0 1.65 – 2.36 Y = -0.98 + 3.55X 3.55 Keempat 90 1.71 1.30 – 2.08 Y = 0.32 + 2.90X 2.90
Source: Adnan (2004)
Tabel 12. Pengaruh konsentrasi berbeda nematoda isolat NLB terhadap larva penggerek
batang jagung pada pengujian rumah kawat.
Konsentrasi Kerusakan Tanaman
pada 36 HST
Mortalitas Larva Larva yang Masih Hidup
Rata-rata Jumlah Nematoda dalam
tubuh larva N2000 8.2 345.48c 6.6.55b 31.133.95b N4000 7 424.47c 5.8.45b 42.27.77a N8000 7 545.48b 4.4.55c 43.147.50a
Insecticide 5 907.07a 1.71d 0c
Kontrol 9 88.37d 9.2.84a 0c Sumber: Adnan 2004
Tabel 13. Pengaruh konsentrasi berbeda nematoda isolat NLB terhadap larva ulat grayak
pada pengujian rumah kawat.
Konsentrasi Kerusakan Tanaman
pada 36 HST
Mortalitas Larva Larva yang Masih Hidup
Rata-rata Jumlah Nematoda dalam
tubuh larva N2000 5.8 365.48 c 6.4.55b 44.186.38b N4000 5.8 428.37c 5.8.84b 49.919.16b N8000 5 584.47b 4.2.45c 59.132.97a
Insecticide 3.4 965.47a 0.4.55d 0c
Kontrol 7.4 84.47d 9.2.45a 0c Sumber: Adnan 2004

399 Seminar Nasional Serealia 2011
mempertahankan keefektifan agen pengendali hayati dalam mengendalikan serangga hama dan mengurangi penggunaan pestisida sintetik yang memiliki pengaruh negatif terhadap lingkungan.
Manajemen Trichogramma spp. yang perlu diperhatikan agar keefktifannya dapat terjaga adalah suhu optimal (20 – 30oC) dengan kelembaban nisbi 60% dalam aplikasi dan tata cara pelepasannya. Pelepasan parasitoid ini ke pertanaman jagung didasarkan pada umur tanaman, hasil tangkapan ngengat, dan pengamatan populasi penggerek batang di lapangan. Upaya ini dilakukan agar pelepasan Trichogramma spp. sejalan dengan periode peletakan telur serangga target (O. furnacalis). Umur tanaman berkaitan dengan periode pelatakan telur penggerek batang. Imago penggerek batang meletakkan telur pada tanaman jagung dimulai pada saat tanaman berumur satu bulan, meskipun di lapangan umur tanaman 3 minggu
telah ditemukan koloni telur penggerek batang dan puncak peletakkan telur terjadi pada saat tanaman membentuk atau keluarnya bunga jantan (Nafus dan Schreiner 1987; Bayrami dan Kornosor 1998). Dengan demikian pelepasan parasitoid telur ini pada pertanaman sebaiknya dimulai pada saat tanaman berumur 3 atau 4 minggu.
Jumlah indivudu dan frekuensi pelepasan Trichogramma spp. mempengaruhi pula keefektifan agen pengendali hayati ini dalam mengendalikan serangga hama. Nonci et al. 2000 melaporkan bahwa pelepasan parasitoid telur Trichogramma spp. sebanyak 250.000 ekor/ha/aplikasi dengan frekwensi pelepasan empat kali dengan interval 1 minggu memebrikan tingkat parasitasi yang cukup tinggi (Gambar 4).
Demikian halnya untuk mengendalikan penggerek tongkol, pelepasan parasitoid telur Trichogramma spp. disesuaikan dengan
Keterangan:
P1 = Pelepasan pada 4, 5, 6, 7, dan 8 MST P4 = Pelepasan pada 6, 7, dan 8 MST P2 = Pelepasan pada 4, 5, 6, dan 7 MST P5 = Berdasarkan gejala serangan di lapangan P3 = Pelepasan pada 5, 6, 7, dan 8 MST MST = Minggu Setelah Tanam Sumber: Nonci et al. 2000
Gambar 4. Persentasi parasitasi Trichogramma spp. pada telur penggerek batang
diLapangan.
Konsentrasi
Parasitasi (%)

400 A.M. Adnan : Manajemen Musuh Alami Hama Utama Jagung
fase pembetukan tongkol jagung, karena prilaku imago betina H. armigera dalam meletakkan telur adalah ditempatkannya telur-telurnya secara individual pada rambut (silk) tongkol jagung, dan frekwensi pelepasan 2 sampai 3 kali (Tabel 5).
Cecopet (Euborellia annulata) sebagai predator ordo lepidoptera relatif lebih mudah untuk dikelola karena jumlah individu yang dibutuhkan untuk pelepasan relatif lebih sedikit hanya sekitar 10.000 ekor/ha dengan frekwensi aplikasi hanya 1 kali (Tabel 8 dan 9). Adnan (2010) menyarankan bahwa waktu aplikasi predatror E. annulata dilakukan pada sore hari.
Agen pengendali hayati yang juga perlu manajemen yang baik adalah cendawan entomopatogen atau cendawan patogen serangga baik Beauveeria bassiana maupun Metarhizium anisopliae. Kesuksesan kedua cendawan patogen serangga ini dalam mengendalikan hama dipengaruhi oleh suhu, kelembaban, sinar matahari, curah hujan, dan angin (Marcandier dan Khachatourians 1978; Tandiabang et al. 2005). Faktor-faktor tersebut yang perlu diperhatikan dalam aplikasi (waktu dan frekwensi) di lapangan.
Waktu aplikasi sangat mempengaruhi kefektifan cendawan patogen serangga. Setelah diaplikasikan, cendawan patogen serangga memerlukan kelembaban yang tinggi untuk tumbuh dan berkemabng (Lacey dan Goettel 1995; Mazet et al. 1996). Kelembaban udara yang tinggi diperlukan selama proses pembentukan germ tube sebelum terjadi penetrasi ke integumen serangga (Steinkraus dan Slaymaker, 1994; Arthurs dan Thomas, 2001). Kelembaban yang tinggi dia atas 90% selama 6 – 12 jam setelah inokulasi dibutuhkan cendawan untuk melakukan penetrasi ke dalam tubuh serangga (Hoddle 1999; Altre dan Vandenberg 2001; Cloyd 2003).
Waktu aplikasi perlu diperhatikan karena cendawan entomopatogen ini sangat rentan terhadap sinar matahari khususnya UV (Altre dan Vandenberg 2001; Cloyd 2003). Oleh karena itu, bila
cendawan diaplikasikan pada musim kemarau perlu dihindarkan dari sinar matahari langsung dan sebaiknya diaplikasikan pada saat kelembaban tinggi yaitu sore hari.
Frekuensi aplikasi cendawan entomopatogen baik Beauveria bassiana maupun Metarhizium anisopliae berpengaruh terhadap keefektifan hama. Hal ini karena Hal ini karena konidia yang diaplikasikan pada tahap awal (yang belum mampu menginfeksi hama sasaran) perlu digantikan oleh konidia yang diaplikasikan pada tahap selanjutnya. Frekuensi aplikasi dipengaruhi oleh kondisi cuaca, seperti curah, hujan, angin, dan sinar matahari. Aplikasi juga perlu memperhatikan stadia serangga hama di lapangan yang saling tumpang tindih (tidak seragam). Perubahan stadia instar (nimfa) akan mengakibatkan perubahan perilaku serangga yang akhirnya berpengaruh pada frekuensi aplikasi.
Frekuensi aplikasi perlu diketahui dengan tepat agar populasi hama di bawah nilai ambang kendali. Aplikasi M. anisopliae tiga kali berturut-turut selama 3 hari lebih efektif mengendalikan S. litura hingga menyebabkan kematian 86%, dibandingkan dengan aplikasi satu kali yang hanya menimbulkan kematian 40% (Prayogo dan Tengkano 2004). Sementara untuk B. bassiana dianjurkam hingga empat kali. Di daerah dengan curah hujan tinggi dianjurkan melakukan aplikasi berulang kali untuk menghindari kegagalan konidia dalam menginfeksi serangga (Hoddle 1999; Cloyd 2003).
Keberhasilan pengendalian hama dengan cendawan entomopatogen juga ditentukan oleh konsentrasi cendawan yang diaplikasikan (Hall 1980), yaitu kerapatan konidia dalam setiap mililiter air. Jumlah konidia berkaitan dengan banyaknya biakan cendawan yang dibutuhkan setiap hektar. Kerapatan konidia yang dibutuhkan untuk mengendalikan hama bergantung pada jenis dan populasi hama yang akan dikendalikan (Tohidin et al. 1993; Wikardi 1993).
Konsentrasi cendawan entomopatogen harus ditentukan secara

401 Seminar Nasional Serealia 2011
tepat untuk mendapatkan hasil pengendalian yang optimal. Kerapatan konidia M. anisopliae yang tepat untuk mengendalikan ulat grayak (S. litura) adalah 108/ml (Gambar 1) dan kerapatan konidia B. bassaiana untuk mengendalikan O. furnacalis adalah 5 x 107/ml (Tabel 10).
Demikian halnya dengan nematoda patogen serangga (Steinernema sp.), temperatur kelembaban, dan UltraViolet mempengaruhi keberhasilan nematoda ini dalam mengendalikan serangga hama utamanya di lapangan saat diaplikasikan. Ada dua cara yang umumnya digunakan dalam aplikasi nematoda patogen serangga yaitu aplikasi melalui tanah dan aplikasi melalui daun. Aplikasi melalui permukaan tanah paling umum digunakan dan sangat efektif mengendalikan serangga tanah, karena kedua organisme tersebut memiliki habitat yang sama (Adnan dan Burhanuddin 2007).
Aplikasi melalui daun juga dapat dilakukan untuk mengendalikan serangga-serangga hama di atas permukaan tanah. Aplikasi melalui daun ini perlu memperhatikan kondisi lingkungan utamanya suhu, kelembaban dan UV. Nematoda entomopatogen kurang efektif dalam mengendalikan hama pada suhu lingkungan di atas 32oC dan tereksopos oleh radiasi ultra violet. Pengaruh efek sinar matahari dapat dikurangi dengan memberikan proteksi pada nematoda patogen serangga (Steinernema carpocapse) yaitu Flourecent Brightener Tinopal dan Viral Enhancer Blankhopor BBH (Nickle dan Shapiro 1994). Hal lain yang perlu diperhatikan dalam aplikasi adalah kelembaban. Kelembaban di atas 80% dipersyaratkan agar nematoda patogen serangga dapat berkembang dan efektif untuk mengendalikan hama (Baur et al. 1997). Oleh karena itu, nematoda entomopatogen direkomendasikan untuk diaplikasikan pada kondisi lingkungan dengan suhu dibawah 32oC, tidak tereksopos lama dibawah sinar matahari, dan kelembaban lebih dari 80% (sore hari).
PENUTUP
Aplikasi musuh alami untuk mengendalikan hama utama jagung adakalanya kurang efektif. Salah satu faktor ketidakberhasilan penggunaan agen pengendali hayati di areal pertanian adalah manajemen musuh alami tersebut. Jika faktor ini dapat ditangani dengan baik, maka kerusakan tanaman dapat ditekan yang pada akhirnya produksi dapat ditingkatkan dan ramah lingkungan. Pengelolaan musuh alami dimaksudkan untuk meningkatkan keefektifan agen pengendali hayati dalam mengendalikan serangga hama dan mengurangi penggunaan pestisida sintetik yang memiliki pengaruh negatif terhadap lingkungan.
Pengelolan musuh alami, akan berhasil jika memperhatikan jenis hama, jenis musuh alami, tata cara aplikasi, lingkungan berupa suhu, kelembaban, curah hujan, sinar matahari, dan angin.
DAFTAR PUSTAKA
Adams, J.B. and M.E. Drew. 1964. Grain aphids in Brunswick. II. Comparative development in the greenhouse of three aphid species on four kinds of grasses. Canadian Journal of Zoology, 42:741-744.
Adnan, A.M., 2004. Isolation, characterization, and efficacy of entomopathogenic nematides as biological control agent against Asian Corn Borer (Ostrinia furnacalis) and cutworm (Spodoptera litura). University of The Philippines Los Banos. Dissertation.
Adnan, A.M., 2010. Pemanfaatan predator cecopet Eiborellia annulata untuk mengendalikan penggerek batang jagung (Ostrinia furnacalis). Laporan Akhir Tahun TA. 2010. Balai Penelitian Tanaman Serealia.
Adnan dan Burhanuddin, 2007. Bioecology of entomopathogenic nematode (Steinernemadae; Rhabditids). Prosiding Seminar

402 A.M. Adnan : Manajemen Musuh Alami Hama Utama Jagung
Ilmiah dan Pertemuan Tahunan XVIII PEI dan PFI. Hal: 199-320
Adnan dan Syamsuddin. 2009. Alba M.C., 1989. Eggs parasitoids of
lepidoptera pest economic importance in the Philippines. Biological Control of Pest. Biotrop Special Publ. No. 36: 123-139.
Altre, J.A. and J.D. Vandenberg. 2001a. Penetration of cuticle and proliferation in himolymph by Paecilomyces fumosoroseus isolates that differ in virulence against lepidopteran larvae. J. Invertebr. Pathol. (78): 78-85.
Arthurs, S. and M.B. Thomas. 2001. Effects of temperature and relative humidity on speculation of Metarhizium anisopliae var. acridum in mycosed cadavers of Schistocerca gregaria. J. Invertebr. Pathol. (78): 59-65.
Badan Pusat Statistik, 2009. Luas panen, produktivitas, dan produksi Jagung. www.bps.go.id/
Baco dan Yasin, 2000. Pengendalian penggerek batang jagung (Ostrinia furnacalis) dengan predator dan patogen. Laporan Tahunan Penelitian Hama dan Penyakit. Balitjas. 2001.
Baur M.E, H.K. Kaya, and Bruce E.T., 1997. Efficacy of a dehydrate steinernematid nematod against black cutworm (Lepidoptera; Noctuidae) and diomand back moth (Lepidoptera: Plutellidae). J. Econ. Entomology 90(5): 1200-1206
Bayrami dan Kornosor, 1998. Biological feature of Trichogramma evanescens Westwood (Hymenoptera; Trichogrammatidae) on the eggs of Sesamia nonagrioides Lefebre (Lepidoptera; Noctuidae). Http:// www.infolomd.at.
Cloyd, R. 2003. The entomopathogen Verticillium lecanii. Midwest
Biological Control News. University of Illions. http://www. extension.umn.Edu.
Crop Protection Compendium, 2005. CABI
Ganguli, R.N. and D.N. Raychaudhuri. 1980. Studies on Rhopalosiphum maidis Fitch (Aphididae: Homoptera)-a formidable pest of Zea mays (maize), in Tripura. Science and Culture, 46(7):259-261.
Glazers dan A. Navon, 1990. Activity and persistance of entomopathogenic tested against Heliothis armigera (Lepidoptera; Noctuidae). J. Econ. Entomol. 83(5): 1795-1800.
Hall, R.A. 1980. Control of aphids by the fungus, Verticillium lecanii: Effect of spore concentration. Entomol. Experiment. App. (27): 1-5.
Hoddle, M.S. 1999. The biology and management of silverleaf whitefly Bemisia argentifolii Bellows and Perring (Homoptera: Aleyrodidae) on greenhouse grown ornamentals.http://www.biocontrol.ucr.edu/ bemisia.html# verticillium [5 April 2005].
Holloway JD, 1989. The moths of Borneo: family Noctuidae, trifine subfamilies: Noctuinae, Heliothinae, Hadeninae, Acronictinae, Amphipyrinae, Agaristinae. Malayan Nature Journal, 42(2-3):57-228.
Javier P.A. and Morallo Rejesus B., 1991. Selective Toxicity of Insecticide to the Earwig Eauborellia annulata fabricus (Dermaptera: Anisolabididae) Predatory to the Asian Corn Borer Ostrinia furnacalis Guenne. The Philiphine Agriculturist.
Kalshoven, LGE. 1981. Pests of Crops in Indonesia. PT. Ichtiar. Baru-Van- Hoeve. Jakarta. 701 p.
Kaya, H.K dan R. Gaugler, 1993. Entomopathogenic nematodes.

403 Seminar Nasional Serealia 2011
Annual Review Entomology. 38z; 181-206
Kranz, J., H. Schumutterer, and W. Koch (Eds.) 1977. Diseases, Pests, and Weeds in Tropical Crops. Berlin and Hamburg, Germany: Verlag Paul Parley
Kring, T.J. 1985. Key and diagnosis of the instars of the corn leaf aphid Rhopalosiphum maidis (Fitch). Southwestern Entomologist, 10(4):289-293.
Legacion, D.M. and B.P. Gabriel. 1988. Note: oviposition of Asiatic corn borer moths on corn plants. Philippine Agriculturist, 71(3):375-378.
Lee YB, CY, Hwang, KM, Choi, and JY, Shim, 1980. Studies on the bionomics of the oriental corn borer Ostrinia furnacalis (Guenee). Korean Journal of Plant Protection, 19(4):187-192.
Lewis, E.E. 2001. Behavioural Biology. In Entomopathogenic nematodes edited by R. Gaugler. Pp. 205-223
Lacey, L.A. and M.S. Goettel. 1995. Current developments in microbial control of insect pests and prospects for the early 21 century. Entomophaga (40): 3-27.
Mazet, I., J.C. Pendland, and D.G. Boucias. 1996. Dependence of Verticillium lecanii (Fungi:Hypomycetes) on high humidity for infection and sporulation using Myzus persicae (Homoptera:Aphididae) as host. Environ. Entomol. (15): 380-382.
Marcandier, S. dan S.S. Khachatourians, 1978. Susceptibility of the migratory grasshopper Melanophus sanguininipes Fab (Orthoptera; Acrididae) to Beauveria bassiana Bals (Hyphomycetes), influence of relatif humidity. The Canadian Entomologist, 119 (10: 9011-907).
Mau, R.F.L. and J.L.M. Kessing. 1992. Rhopalosiphum maidis (Fitch). Honolulu, Hawaii: Hawaii
Entomology Extension Service. http://www.extento. hawaii.edu/.
Metcal RL. and R.A. Metcalf, 1993. Destructive and useful insect, their habits, and their control. Fifth Edition.Mc. Grow-Hill. Inc.
Mutuura A, and E. Munroe, 1970. Taxonomy and distribution of the European corn borer and allied species: genus Ostrinia (Lepidoptera:Pyralidae). Memoirs of the Entomological Society of Canada No.71, 112 pp.
Nafus dan Schreiner. 1987. Location of Ostrinia furnacalis Gueene. Eggs and larvae on sweet corn in relation to plant growth. Journal of econ entomol, 84(2). 411-416
Nickle W.R dan M. Shapiro, 1994. Effect of eiigth brigthteners as solar radiation protectans for Steinernema carpocapse, All Strain. Supplement to journal of Nematology 26 (48). P. 82-784
Nonci, N, J. Tandiabang, Masmawati, A Muis, 2000. Inventarisasi musuh alami penggerek batang jagung (Ostrinia furnacalis) pada sentra produksi jagung di Sulawesi Selatan. Penelitian Pertanian. Pusat Penelitian Tanaman Pangan. Badan Penelitian dan Pengembangan Pertanian. Hal 38-49.
Nonci dan Masmawati, 2005. Kemampuan jelajah Trichogramma evanescens Westwood parasitoid telur penggerek batang (Ostrinia furnacalis). Prosiding Seminar dan Lokakarya Nasional. Pusat Penelitian dan Pengembangan Tanaman Pangan. Badan Penelitian dan Pengembangan Pertanian. Makassar, 23-25 September 2005. Hal: 562-566.
Nonci dan Pabbage, 2000. Tingkat parasitasi Trichogramma evanescens Westwood pada telur penggerek batang jagung Ostrinia furnacalis Guenee di rumah kaca. Hasil penelitian hama dan penyakit 1999-2000. Balai Penelitian

404 A.M. Adnan : Manajemen Musuh Alami Hama Utama Jagung
Tanaman Jagung dan Serealia Lain. Badan Penelitian dan Pengembangan Pertanian.
Pabbage, M.S., N. Nonci, D. Baco. 2000. Kefektifan Trichogrammatoidae bactrae fumata dalam pengendalian penggerek tongkol jagung Helicoverpa armigera (Lepidoptera; Noctuidae) di laboratorium. Hasil Penelitian Hama dan Penyakit 1999/2000. Balai Penelitia Tanaman Jagung dan Serealia Lainnya. Badan Penelitian dan Pengembangan Pertanian.
Prayogo, Y. 2004. Keefektifan lima jenis cendawan entomopatogen terhadap hama pengisap polong kedelai Riptortus linearis (L.) (Hemiptera: Alydidae) dan dampaknya terhadap predator Oxyopes javanus Thorell (Araneida: Oxyopidae). Tesis. Departemen Hama Penyakit Tanaman, Sekolah Pascasarjana, Institut Pertanian Bogor. 51 hlm.
Prayogo, Y., W. Tengkano, dan Suharsono. 2002a. Jamur entomopatogen pada Spodoptera litura dan Helicoverpa armigera. Seminar Hasil Penelitian Tanaman Kacangkacangan dan Umbi-umbian. Malang, 2526 Juni 2002. Balai Penelitian Tanaman Kacang-kacangan dan Umbi-umbian, Malang. 15 hlm.
Philippine-German Crop Protection Programme (PGCPP), !987. Integrated Pest Management. Corn. Bureau Of Plant Industry, Department of Agriculture. San Andres, Malate, Manila. Philippines.
Situmorang J. and B.P. Gabriel, 1988. Biologi of two predatory Earwigs, Nala lividipes (Dotour) (Dermaptera: Labiduridae) and Euborellian annulata (Fabricus) (Dermaptera: Anisolabididae). Philipphine Entomol. 7 (3): 215-238.
Smart, G.C., Jr. 1995. Entomopathogenic nematodes for biological control of insects. Supplement to J. Nematol. 27(48): 529-534
Steinkraus, D.C. and P.H. Slaymaker. 1994. Effect of temperature and humidity on formation, germination, and infectivity of conidia of Neozygites fresenii (Zygomycetes: Neozygitaceae) from Aphis gossypii (Homoptera: Aphididae). J. Invertebr. Pathol. (64): 130-137.
Suharsono dan Y. Prayogo. 2005. Pengaruh lama pemaparan pada sinar matahari terhadap viabilitas jamur entomopatogen Verticillium lecanii. Jurnal Habitat XVI(2): 122-131.
Swastika, Dewa K.S., F. Kasim, W. Sudana, R. Hendayani, K. Suhariyanto, R.V. Gerpacio, P.L. Pingali. 2004. Maize in Indonesia, Production system, constraints, research priorities. CIMMYT.
Tanada, Y. and H.K. Kaya. 1993. Insect Pathology. Academic Press, Inc., California.
Tandiabang, J., Y. Said, M.S. Saenong, 2005. Resensi teknologi penelitian Beauveria bassiana Vuill untuk penanganan OPT jagung. Prosiding Seminar dan Lokakarya Nasional. Pusat Penelitian dan Pengembangan Tanaman Pangan. Badan Penelitian dan Pengembangan Pertanian. Makassar, 23-25 September 2005.
Tohidin, A.T. Lisrianto, dan B.P. Machdar. 1993. Daya bunuh jamur entomopatogen Beauveria bassiana (Balsamo) Vuillemin (Moniliales: Moniliacea) terhadap Leptocoriza acuta Thunberg (Hemiptera: Alydidae) di rumah kaca. hlm. 135-143. Dalam E. Martono, E. Mahrub, N.S. Putra, dan Y. Trisetyawati (Ed.). Prosiding Simposium Patologi Serangga I. Yogyakarta, 12-13 Oktober 1993. Universitas Gadjah Mada, Yogyakarta.

405 Seminar Nasional Serealia 2011
Waterhouse DF, 1993. The major arthropod pests and weeds of agriculture in Southeast Asia. The major arthropod pests and weeds of agriculture in Southeast Asia., v + 141 pp.; [ACIAR Monograph No. 21]; 3 pp. of ref.
Wahyunendo, Y.D. 2002. Sporulasi cendawan entomopatogen Beauveria bassiana (Bals.) Vuill. pada berbagai media alami dan viabilitasnya di bawah pengaruh suhu dan sinar matahari [Skripsi]. Bogor. Institut Pertanian Bogor. Jurusan Hama dan Penyakit Tumbuhan.
Wikardi, E.A. 1993. Teknik perbanyakan Beauveria bassiana dan aplikasinya
di lapang. hlm. 205-214. Dalam E. Martono, E. Mahrub, N.S. Putra, dan Y. Trisetyawati (Ed.). Prosiding Simposium Patologi Serangga I. Yogyakarta, 12-13 Oktober 1993. Universitas Gadjah Mada, Yogyakarta
Yasin, M., S. Mas’ud, A.H. Talanca, D. Baco, 2000. Pengaruh lama penyimpanan cendawan Beauveria bassiana dalam pengendalian penggerek batang jagung (Ostrinia furnacalis. Hasil Penelitian Hama dan Penyakit 2000. Balai Penelitia Tanaman Jagung dan Serealia Lainnya. Badan Penelitian dan Pengembangan Pertanian.





























