Embed Size (px)
Citation preview

Mannan and peptidoglycan induce COX-2 protein inhuman PMN via the mammalian target of rapamycin
Nieves Fern�ndez, Ana Gonz�lez, Isela Valera, Sara Alonso and Mariano S�nchez Crespo
Instituto de Biolog�a y Gen�tica Molecular, Consejo Superior de Investigaciones Cient�ficas, Valladolid, Spain
The induction of cyclooxygenase-2 (COX-2) protein expression was assessed in humanpolymorphonuclear leukocytes (PMN) stimulated via receptors of the innate immunesystem. Peptidoglycan (PGN) and mannan, and at a lower extent the bacteriallipoprotein mimic palmitoyl-3-cysteine-serine-lysine-4, induced COX-2 protein expres-sion. In contrast, lipoteichoic acid and muramyldipeptide were irrelevant stimuli. ThemRNA encoding COX-2 was present in resting PMN at an extent quite similar to thatdetected in stimulated PMN, whereas the expression of COX-2 protein wasundetectable. Treatment with the phosphatidylinositol 3-kinase inhibitor (PI3K)wortmaninn, the mammalian target of rapamycin (mTOR) inhibitor rapamycin, and thetranslation inhibitor cycloheximide blocked the induction of COX-2 protein in responsetomannan and PGN, whereas the transcriptional inhibitor actinomycin D did not show asignificant effect. These results disclose a capability of pathogen-associated molecularpatterns to induce the oxidativemetabolism of arachidonic acidmore robust than that ofPMN archetypal chemoattractants, since mannan and PGN make it coincidental therelease of arachidonic acid with a rapid induction of COX-2 protein regulated by asignaling cascade involving PI3K, mTOR, and the translation machinery. Thismechanism of COX-2 protein induction expression in PMN is substantially differentfrom that operative in mononuclear phagocytes, which is highly dependent ontranscriptional regulation.
Introduction
PMN are essential elements of the innate immunesystem due to their wide predominance in peripheralblood and their rapid mobilization into tissues in
response to microbial infection. PMN can be activatedby a variety of stimuli, which includes cytokines, IgGclass antibodies, complement factors, extracellularmatrix components, adhesion molecules, and micro-organism constituents. This group of stimuli is of chiefimportance, since early protection against pathogensrelies on the recognition by phagocytes of uniquepattern molecules, termed pathogen-associated mole-cular patterns (PAMP), which are recognized throughpattern-recognition receptors (PRR) by the host innateimmune system. The TLR family (for review see [1, 2]),the nucleotide-binding oligomerization domain (NOD)family proteins (for review see [3–5]), the lectin C-typereceptors such as the b-glucan receptor dectin-1 [6], andthemannose receptor (MR) family (for review see [7, 8])include a wide array of PRR able to interact with manystructural signatures expressed in microorganisms.
The study of the response of PMN to PAMP hasfocused on the induction of cytokines, the activation of
Correspondence: Dr. Mariano S�nchez Crespo, Instituto deBiolog�a y Gen�tica Molecular, C/ Sanz y For�s s/n, 47003-Valladolid, SpainFax: +34-983-184-800e-mail: [email protected]
Received 14/3/07Revised 24/5/07
Accepted 11/7/07
[DOI 10.1002/eji.200737262]
Key words:Endotoxin � Human� Innate immunity� Lipid mediators
� Neutrophils
Abbreviations: 4E-BP1: eIF4E-binding protein �AA: arachidonic acid �MDP:muramyldipeptide �MR:mannosereceptor � NOD: nucleotide–binding oligomerization domainfamily proteins � mTOR: mammalian target of rapamycin �PAF: platelet-activating factor � PAMP: pathogen-associatedmolecular patterns � PGN: peptidoglycan � PGRP: PGN-recog-nizing protein � PRR: pattern recognition receptors �UTR: untranslated region
Nieves Fern�ndez et al. Eur. J. Immunol. 2007. 37: 2572–25822572
f 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

oxidant production, the release of granular components,and the modulation of apoptosis [9–11]. However, fewdata are available regarding the activation of thearachidonic acid (AA) cascade, yet this seems of interestin view of the important role of eicosanoids inconnecting innate and adaptive immunity [12–15]and the attention paid to the pharmacological modula-tion of both the eicosanoid receptors and the enzymesinvolved in their biosynthesis in many clinical settings[16, 17]. Moreover, current understanding of PMNbiology has been modified by recent findings indicatingthat the life span of PMN can be prolonged byproinflammatory agonists [10], as well as by thedepiction of mechanisms of translational control ofthe expression of specific proteins, which endow thePMN with the potential for rapid protein synthesis fromconstitutive mRNA without requiring new transcriptgeneration [18–20]. The possibility that this mechanismcould be operative in PAMP-dependent responses andmight influence AA metabolism through the expressionof COX-2, the inducible isoform of COX, is a challenginghypothesis. It could provide a rationale for pharmaco-logical intervention with the macrolide antibioticrapamycin, an inhibitor of the mammalian target ofrapamycin (mTOR) [21], or alternatively, an explana-tion for some of the untoward effects of rapamycin.
In a previous study, we have observed thatpeptidoglycan (PGN) and mannan are potent stimulifor AA release and PGE2 production in human PMN [22].Since PGE2 can be produced both by COX-1, theconstitutive isoform of COX, and by COX-2, the inducibleisoform, we have addressed the effect of a set of PAMPsignatures on the expression of COX-2 in human PMN.We have found the presence of preformed mRNAencoding for COX-2 in resting PMN and an early andlasting induction of COX-2 protein in response to bothmannan and PGN. Since the expression of COX-2 proteinwas almost completely blocked by the inhibitor ofphosphatidylinositol 3-kinase wortmannin, by themTOR inhibitor rapamycin, and by the translationinhibitor cycloheximide, we propose the existence of amechanism of COX-2 induction expression in humanPMN mediated by the mTOR route.
Results
PAMP signatures are strong inducers of COX-2protein in human PMN
Since PMN have a limited life span and gene expressioninduction assays might entail long incubation periods,the appearance of apoptosis was assessed by looking atboth phosphatidylserine surface display and propidiumiodide staining. Less than 10% of total cells showed
positive staining with annexin V-FITC at the end of theisolation procedure, whereas this figure steadily in-creased along the incubation period to reach * 96%positive cells at 24 h. In contrast, propidium iodidestaining after 24 h of incubation only showed 18% ofpositive cells (Fig.1A), thus indicating that PMN isolatedby conventional procedures are viable and can be used inexperiments entailing long incubation periods, since nomassive late apoptosis was observed within 24 h.
Human PMN were found to contain high amounts ofCOX-2 mRNA, as judged from RT-PCR reactions carriedout in samples obtained both in resting cells and afterstimulation with agonists acting through a varied arrayof PRR. Of note, differences in the extent of COX-2mRNA expression could hardly be detected in RT-PCRreactions (Fig. 1B). These results were first observedwith primers selected from exons 5 and 7, but similarresults were obtained with primers selected from exons1 and 2 (see below). Real-time RT-PCR of COX-2 mRNAshowed a CT value of 18.33 � 0.73 in control cells, 16.09� 0.53 (mean � SD, n = 5) in PGN-treated PMN, and17.1 � 0.33 in mannan-treated PMN, i.e. a set of values,which after normalization for the expression of b-actinmRNA only represents a 2.46-fold increase for PGN- anda 1.8-fold induction for mannan-treated PMN. Thesefindings are different from those observed in macro-phages, where mannan and PGN increased COX-2mRNA 363- and 163-fold, respectively (Table 1), thusindicating either robust transcriptional activation ormRNA stabilization.
Since recent studies have suggested that PMNrespond to chemical mediators by initiating the transla-tion of preformed mRNA [19–21], these findings wereconsidered as an indication that a similar molecularmechanism might be implicated in the regulation ofCOX-2 protein. Mannan and PGN produced a dose-dependent induction of the expression of COX-2 protein(Fig. 1C). Of note, this was also observed with platelet-activating factor (PAF) (Fig. 1D), which is the chemicalmediator first associated to this mechanism of signal-dependent translation of constitutive messenger RNA inPMN [19]. However, the level of COX-2 proteinexpression induced by PAF was somewhat lower,thereby suggesting a more robust COX-2 protein
Table 1. COX-2 mRNA expression in different cell typesa)
Stimulus PMN Monocytes Macrophages
Mannan 10 mg/mL 1.8 302 363
PGN 10 lg/mL 2.46 179 163
a) Results have been normalized to b-actin mRNA expressionlevels and are expressed as fold-induction upon resting cellvalues.
Eur. J. Immunol. 2007. 37: 2572–2582 Molecular immunology 2573
f 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu
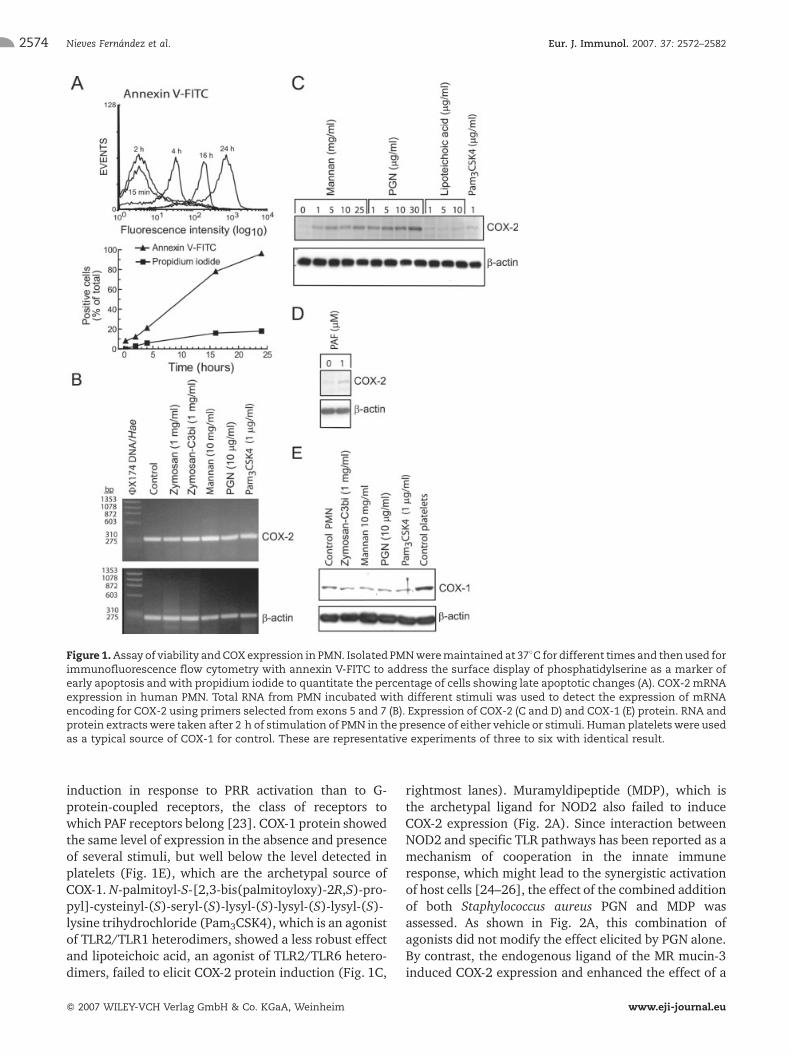
induction in response to PRR activation than to G-protein-coupled receptors, the class of receptors towhich PAF receptors belong [23]. COX-1 protein showedthe same level of expression in the absence and presenceof several stimuli, but well below the level detected inplatelets (Fig. 1E), which are the archetypal source ofCOX-1. N-palmitoyl-S-[2,3-bis(palmitoyloxy)-2R,S)-pro-pyl]-cysteinyl-(S)-seryl-(S)-lysyl-(S)-lysyl-(S)-lysyl-(S)-lysine trihydrochloride (Pam3CSK4), which is an agonistof TLR2/TLR1 heterodimers, showed a less robust effectand lipoteichoic acid, an agonist of TLR2/TLR6 hetero-dimers, failed to elicit COX-2 protein induction (Fig. 1C,
rightmost lanes). Muramyldipeptide (MDP), which isthe archetypal ligand for NOD2 also failed to induceCOX-2 expression (Fig. 2A). Since interaction betweenNOD2 and specific TLR pathways has been reported as amechanism of cooperation in the innate immuneresponse, which might lead to the synergistic activationof host cells [24–26], the effect of the combined additionof both Staphylococcus aureus PGN and MDP wasassessed. As shown in Fig. 2A, this combination ofagonists did not modify the effect elicited by PGN alone.By contrast, the endogenous ligand of the MR mucin-3induced COX-2 expression and enhanced the effect of a
Figure 1.Assay of viability andCOXexpression in PMN. Isolated PMNweremaintained at 37�C fordifferent times and thenused forimmunofluorescence flow cytometry with annexin V-FITC to address the surface display of phosphatidylserine as a marker ofearly apoptosis andwith propidium iodide to quantitate the percentage of cells showing late apoptotic changes (A). COX-2 mRNAexpression in human PMN. Total RNA from PMN incubated with different stimuli was used to detect the expression of mRNAencoding for COX-2 using primers selected from exons 5 and 7 (B). Expression of COX-2 (C and D) and COX-1 (E) protein. RNA andprotein extractswere taken after 2 h of stimulation of PMN in the presence of either vehicle or stimuli. Human plateletswere usedas a typical source of COX-1 for control. These are representative experiments of three to six with identical result.
Nieves Fern�ndez et al. Eur. J. Immunol. 2007. 37: 2572–25822574
f 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

suboptimal concentration of mannan (Fig. 2B, rightpanels).
PGN elicited detectable COX-2 protein induction assoon as 30 min after addition of the stimulus and thisremained almost unchanged from 1 to 18 h. A similartrend was observed for C3bi-coated zymosan andmannan, although a decreasing tendency was observedaround 18 h in response to these agonists (Fig. 2B). Todefine more widely the proinflammatory contextelicited by PRR in which COX-2 induction occurs, theexpression of other inflammatory proteins was assessedwith human Ab array membranes. The expression ofIL-1b, IL-8/CXCL8, and MIP-1b/CCL4 increased abovetheir basal expression level in response to mannan(Fig. 2C), thus agreeing with the induction of thesecytokines in human PMN stimulated with other agonists[19]. Taken together, the most likely interpretation ofthese results is that the MR might be involved in theresponse to mannan, and that PGN contains a structuralsignature not acting on NOD2 nor mimicked bylipoteichoic acid and Pam3CSK4, which could act viathe TLR route in combination with an additional catch-up receptor(s) and/or by an yet ill-defined TLR2-independent route.
Mechanisms involved in COX-2 induction inhuman PMN
Since PMN are terminally differentiated cells whichcontain regulators of transcriptional control and showsignal-dependent activation of mRNA translation [20],we put forward the hypothesis that COX-2 mRNA couldbe one of those mRNA controlled in the same manner,because abundant expression of COX-2 mRNA wasdetected in resting human PMN. Contrary to this view,one could argue that the calculation of the predictedsecondary structure energy of the 50- untranslatedregion (UTR) of COX-2 mRNA is - 36.94 kcal/mol, asjudged from the application of RNAfold software [27,28] to sequences available in data banks (Fig. 3A). Thisvalue is lower than that usually associated withtranscriptional regulation (-50 kcal/mol, [18]); how-ever, it fits well with those reported for many transcriptsdetected using cDNA library arrays which are regulatedat the transcriptional level in human monocytesadherent to P-selectin [21]. The presence of the 50-UTR in COX-2 transcripts in human PMN was confirmedby real-time PCR with a set of primers spanning the first20 nucleotides of exon 1 and exon 2 of COX-2, whichgave similar results to PCR reactions using the primersselected from exons 5 and 7 (Fig. 3B). Moreover, thepresence of four tracts of 5–8 consecutive pyrimidinebases (underlined in the sequence shown in Fig. 3A) isan additional feature strongly suggesting the possibilityof translational control by mTOR [19]. As shown in
Figure 2. Effect of different stimuli on COX-2 protein expres-sion. The effect of S. aureus PGN alone or in combination withMDP is shown in (A). The effect of the combined addition ofmannan and mucin-3 is shown in the right upper panel of (B).The time course of induction of COX-2 protein elicited by PGN,zymosan coated with C3bi, and mannan is shown in (B). Theeffect of 18 h of incubation with mannan on the expression ofseveral proinflammatory proteins assessed with the RayBioJ
Human Inflammation Antibody Array III is shown in (C). Blotsin (A) and (B) are representative of three independentexperiments. b-Actin expression was assayed to address theoccurrence of similar protein loading across the gels.
Eur. J. Immunol. 2007. 37: 2572–2582 Molecular immunology 2575
f 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu
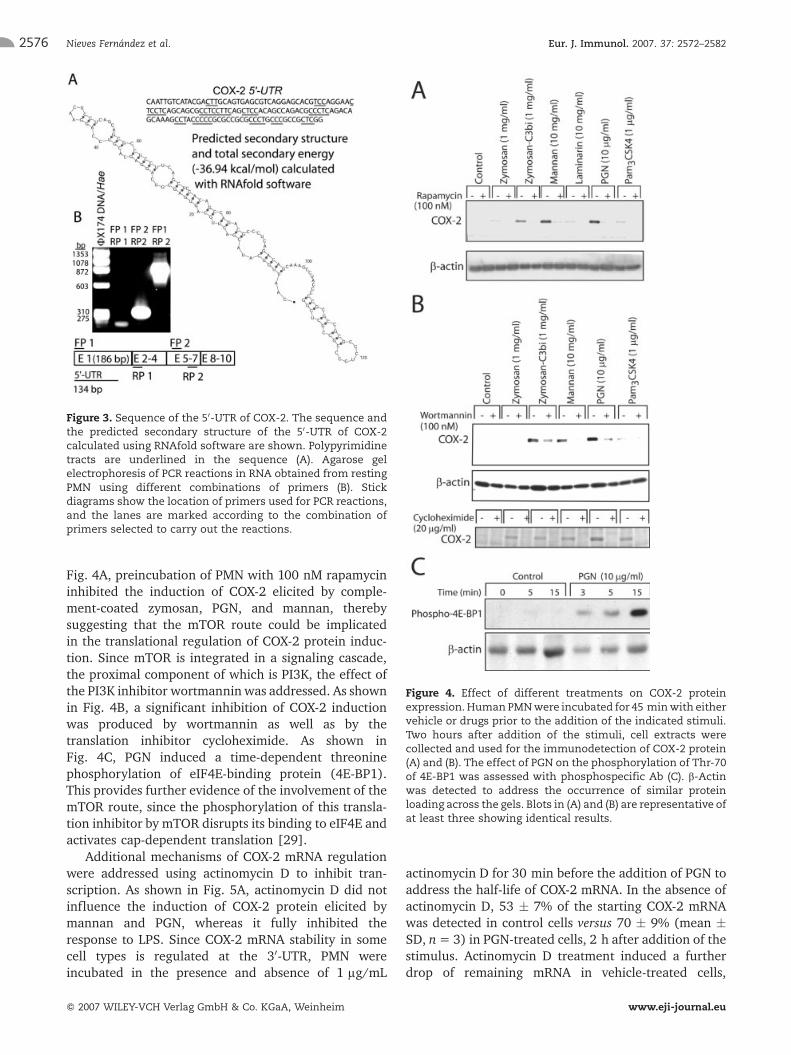
Fig. 4A, preincubation of PMN with 100 nM rapamycininhibited the induction of COX-2 elicited by comple-ment-coated zymosan, PGN, and mannan, therebysuggesting that the mTOR route could be implicatedin the translational regulation of COX-2 protein induc-tion. Since mTOR is integrated in a signaling cascade,the proximal component of which is PI3K, the effect ofthe PI3K inhibitor wortmannin was addressed. As shownin Fig. 4B, a significant inhibition of COX-2 inductionwas produced by wortmannin as well as by thetranslation inhibitor cycloheximide. As shown inFig. 4C, PGN induced a time-dependent threoninephosphorylation of eIF4E-binding protein (4E-BP1).This provides further evidence of the involvement of themTOR route, since the phosphorylation of this transla-tion inhibitor by mTOR disrupts its binding to eIF4E andactivates cap-dependent translation [29].
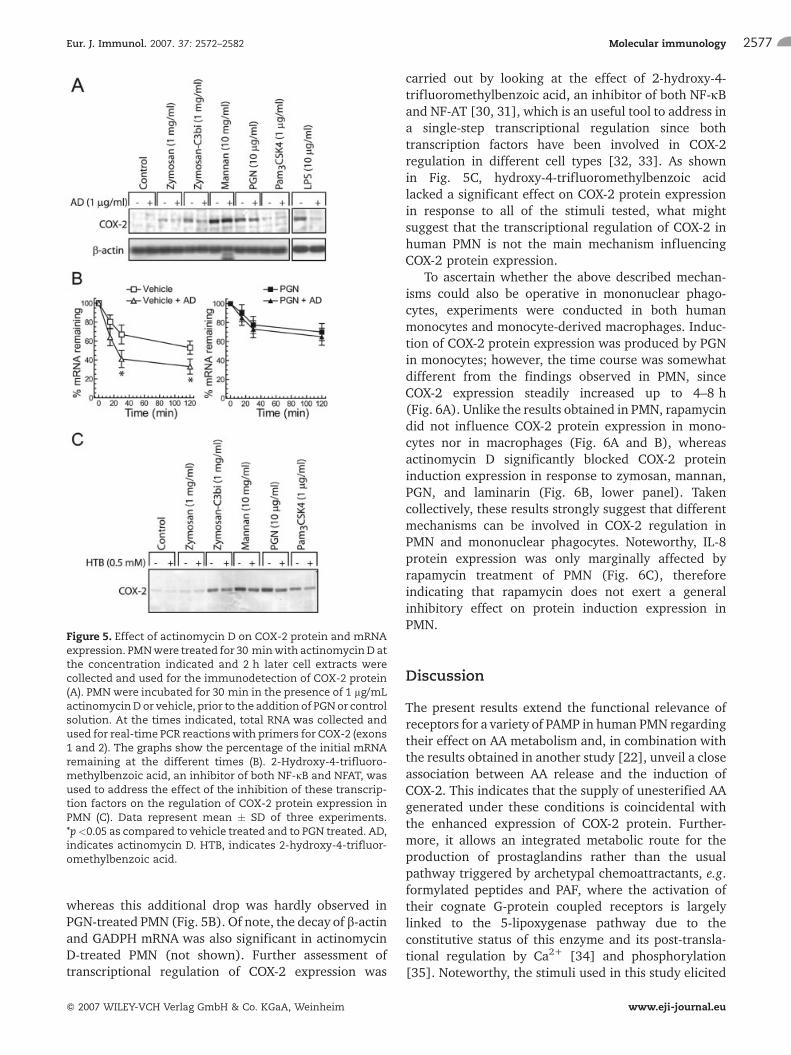
Additional mechanisms of COX-2 mRNA regulationwere addressed using actinomycin D to inhibit tran-scription. As shown in Fig. 5A, actinomycin D did notinfluence the induction of COX-2 protein elicited bymannan and PGN, whereas it fully inhibited theresponse to LPS. Since COX-2 mRNA stability in somecell types is regulated at the 30-UTR, PMN wereincubated in the presence and absence of 1 lg/mL
actinomycin D for 30 min before the addition of PGN toaddress the half-life of COX-2 mRNA. In the absence ofactinomycin D, 53 � 7% of the starting COX-2 mRNAwas detected in control cells versus 70 � 9% (mean �SD, n= 3) in PGN-treated cells, 2 h after addition of thestimulus. Actinomycin D treatment induced a furtherdrop of remaining mRNA in vehicle-treated cells,
Figure 3. Sequence of the 50-UTR of COX-2. The sequence andthe predicted secondary structure of the 50-UTR of COX-2calculated using RNAfold software are shown. Polypyrimidinetracts are underlined in the sequence (A). Agarose gelelectrophoresis of PCR reactions in RNA obtained from restingPMN using different combinations of primers (B). Stickdiagrams show the location of primers used for PCR reactions,and the lanes are marked according to the combination ofprimers selected to carry out the reactions.
Figure 4. Effect of different treatments on COX-2 proteinexpression. Human PMNwere incubated for 45 minwith eithervehicle or drugs prior to the addition of the indicated stimuli.Two hours after addition of the stimuli, cell extracts werecollected and used for the immunodetection of COX-2 protein(A) and (B). The effect of PGN on the phosphorylation of Thr-70of 4E-BP1 was assessed with phosphospecific Ab (C). b-Actinwas detected to address the occurrence of similar proteinloading across the gels. Blots in (A) and (B) are representative ofat least three showing identical results.
Nieves Fern�ndez et al. Eur. J. Immunol. 2007. 37: 2572–25822576
f 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu
whereas this additional drop was hardly observed inPGN-treated PMN (Fig. 5B). Of note, the decay of b-actinand GADPH mRNA was also significant in actinomycinD-treated PMN (not shown). Further assessment oftranscriptional regulation of COX-2 expression was
carried out by looking at the effect of 2-hydroxy-4-trifluoromethylbenzoic acid, an inhibitor of both NF-jBand NF-AT [30, 31], which is an useful tool to address ina single-step transcriptional regulation since bothtranscription factors have been involved in COX-2regulation in different cell types [32, 33]. As shownin Fig. 5C, hydroxy-4-trifluoromethylbenzoic acidlacked a significant effect on COX-2 protein expressionin response to all of the stimuli tested, what mightsuggest that the transcriptional regulation of COX-2 inhuman PMN is not the main mechanism influencingCOX-2 protein expression.
To ascertain whether the above described mechan-isms could also be operative in mononuclear phago-cytes, experiments were conducted in both humanmonocytes and monocyte-derived macrophages. Induc-tion of COX-2 protein expression was produced by PGNin monocytes; however, the time course was somewhatdifferent from the findings observed in PMN, sinceCOX-2 expression steadily increased up to 4–8 h(Fig. 6A). Unlike the results obtained in PMN, rapamycindid not influence COX-2 protein expression in mono-cytes nor in macrophages (Fig. 6A and B), whereasactinomycin D significantly blocked COX-2 proteininduction expression in response to zymosan, mannan,PGN, and laminarin (Fig. 6B, lower panel). Takencollectively, these results strongly suggest that differentmechanisms can be involved in COX-2 regulation inPMN and mononuclear phagocytes. Noteworthy, IL-8protein expression was only marginally affected byrapamycin treatment of PMN (Fig. 6C), thereforeindicating that rapamycin does not exert a generalinhibitory effect on protein induction expression inPMN.
Discussion
The present results extend the functional relevance ofreceptors for a variety of PAMP in human PMN regardingtheir effect on AA metabolism and, in combination withthe results obtained in another study [22], unveil a closeassociation between AA release and the induction ofCOX-2. This indicates that the supply of unesterified AAgenerated under these conditions is coincidental withthe enhanced expression of COX-2 protein. Further-more, it allows an integrated metabolic route for theproduction of prostaglandins rather than the usualpathway triggered by archetypal chemoattractants, e.g.formylated peptides and PAF, where the activation oftheir cognate G-protein coupled receptors is largelylinked to the 5-lipoxygenase pathway due to theconstitutive status of this enzyme and its post-transla-tional regulation by Ca2+ [34] and phosphorylation[35]. Noteworthy, the stimuli used in this study elicited
Figure 5. Effect of actinomycin D on COX-2 protein and mRNAexpression. PMNwere treated for 30 minwith actinomycinD atthe concentration indicated and 2 h later cell extracts werecollected and used for the immunodetection of COX-2 protein(A). PMNwere incubated for 30 min in the presence of 1 lg/mLactinomycinDor vehicle, prior to the addition of PGNor controlsolution. At the times indicated, total RNA was collected andused for real-time PCR reactionswith primers for COX-2 (exons1 and 2). The graphs show the percentage of the initial mRNAremaining at the different times (B). 2-Hydroxy-4-trifluoro-methylbenzoic acid, an inhibitor of both NF-jB and NFAT, wasused to address the effect of the inhibition of these transcrip-tion factors on the regulation of COX-2 protein expression inPMN (C). Data represent mean � SD of three experiments.*p<0.05 as compared to vehicle treated and to PGN treated. AD,indicates actinomycin D. HTB, indicates 2-hydroxy-4-trifluor-omethylbenzoic acid.
Eur. J. Immunol. 2007. 37: 2572–2582 Molecular immunology 2577
f 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

the production of both LTB4 and PGE2 [22], thus fittingwell with the existence of two programs for AAmetabolism in PMN: (i) the constitutive metabolic routeconveying AA into the 5-lipoxygenase pathway, and (ii)the inducible route linked to COX-2 induction expres-sion. The response to PAMP, although showing somefeatures of the first program, fits best with the secondprogram. This fact is of functional relevance, since moststudies on AA release have been carried out withxenobiotics as calcium ionophores and phorbol esters, orcombining agonists of G-protein-coupled receptors with
substances producing a prolongation of Ca2+ transients,for instance cytochalasin B [36] and thapsigargin [37].On this basis, PGN- and mannose-containing molecularpatterns emerge as pathophysiologically relevant in-ducers of AA metabolism in human PMN, at a similarextent as the most functional stimuli so far tested, i.e.opsonized zymosan [38]. In contrast, our data do notsupport a role for lipoteichoic acid and MDP. In the lightof these findings, COX-2 induction expression uponmicrobial invasion seems to be a hallmark of host PMNresponse to microorganisms containing mannose, e.g.
Figure 6.Monocytes (A) andmonocyte-derived macrophages (B) were used to address the effect of rapamycin and actinomycin Don COX-2 protein induction expression. The upper panel in (A) shows the time course of COX-2 protein expression elicited by10 lg/mL PGN on monocytes. The effect of rapamycin on the induction of cytokines elicited by PGN and mannan in PMN wasaddressed using Human Cytokine Antibody Array III from RayBiotech (C). This array differs from the Human InflammationAntibody Array III in that it does not containMIP-1b/CCL4, thus explaining the absence of positive response for this chemokine inthese membranes, as compared to those used in the experiments shown in Fig. 2C.
Nieves Fern�ndez et al. Eur. J. Immunol. 2007. 37: 2572–25822578
f 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

Candida albicans, Leishmania donovani, and Pneumo-cistis carinni, b-glucan, e.g. Candida albicans, and PGN, acomponent mainly found in Gram-positive bacteria,where it accounts for around 90% w/w of the cell wall.This fact helps explain why PGN can reach very highconcentrations in tissues infected by Gram-positivebacteria [39] and be a major etiological factor in thepathogenesis of pyogenic inflammation or in bacterial-infection-induced inflammatory diseases such as psor-iasis [40]. LPS and adenosine have been found to induceCOX-2 protein expression in human PMN previously[41–43], yet these are agonists distinct from those usedin the present experiments, because LPS is a typicalactivator of the TLR4/NF-jB route and adenosine actsthrough the adenosine receptor A2A.
Regulation of COX-2 protein expression is a tightlyregulated process involving both transcriptional andpost-transcriptional events, which may show cell type-dependent mechanisms. Regarding post-transcriptionalmechanisms, regulation by both 50- and 30-UTR seemspossible. The structural hallmarks of 50-UTR implicatedin post-transcriptional control are a 50-terminal oligo-pyrimidine tract and a stable secondary structuredefined by calculated stabilities of –50 kcal/mol orgreater [18, 44]. Whereas the secondary structureenergy of the 50-UTR of COX-2 is more consistent withtranscriptional control, the presence of polypyrimidinetracts and/or as yet unidentified binding proteins maymake COX-2 mRNA responsive to translational controlvia 50-UTR in PMN. In addition, the detection of COX-2mRNA in resting PMN, but not the protein, the effectof rapamycin at the concentrations tested [45–48],and the phosphorylation of 4E-BP1, which allowseIF4E to recognize the 7-methyl cap of mRNA and theinitiation of translation, support post-transcriptionalregulation.
Regulation of COX-2 protein by the 30-UTR has beendescribed at least in monocytes, macrophages, andneoplastic cells since COX-2 mRNA contains 23 copies ofthe AUUUA RNA instability element, which makes thismRNA short-lived and suitable for post-transcriptionalregulation by mechanisms dependent on at least thep38-MAP kinase [49] and the PI3K routes [50].Interestingly, a recent report using a preparation ofplatelets and monocytes has shown that efficientinduction of COX-2 protein requires combination ofsignals involving activation of jB-driven transcriptionalactivation and stabilization of COX-2 mRNA by silencingthe AU-rich mRNA element [51]. Whereas cells with alasting life span might need combination of mechanismsfor a timely regulation of COX-2 protein expression, theexistence of rapid regulatory mechanisms in short-livedcells such as PMN seems most appropriate.
Using actinomycin D to block transcription we havefound a stability of COX-2 mRNA in PMN above that
reported for human monocytes [49] and alveolarmacrophages [50]. Moreover, studies in these cellsrequired preincubation with LPS for periods of 4 to 24 hto allow induction of COX-2 mRNA, whereas in thepresent study the preincubation period could besuppressed due to the presence of preformed COX-2mRNA. Since both the p38-MAP kinase and the PI3Kroutes may be triggered by PGN, it cannot be ruled outthat mRNA stability could be an additional mechanisminvolved in the regulation of the expression of COX-2protein in PMN, because COX-2 mRNA shows a lowerdecay in PGN-treated PMN than in vehicle-treated cells.However, our findings cannot be uniquely explained onthis basis, since COX-2 mRNA is detected in resting PMNat a level not found in monocytes and macrophages, andthe induction of COX-2 protein in PMN shows a morerapid time course than that observed in monocytesunder identical conditions [41]. Unlike actinomycin D,all of the drugs acting on the cascade involving PI3K,mTOR, and the translation machinery showed an almostcomplete inhibition of COX-2 protein expression, thussuggesting that COX-2 is another element of the array ofconstitutive messenger RNA translated by proinflam-matory stimuli in human PMN [19].
TheMR seems to be themost likely PRR for mannose-containing molecular patterns [22], but regarding PGN,three different types of receptors could be taken intoaccount. In a previous study, we have confirmed theexpression of TLR2, TLR1, TLR6, NOD2, and PGN-recognizing protein (PGRP)-S [22], all of which havesome binding capacity for PGN or its elementarybuilding blocks. As regards the TLR2 route, PGN is anarchetypal ligand [39], even though this view has beenchallenged recently after a report has shown that theability of PGN to activate TLR2 might be lost afterremoval of lipoproteins and lipoteichoic acids [52],thereby pointing to the intracellular receptors NOD1 andNOD2 as the actual PGN receptors in view of theircapacity to interact with peptidic moieties of diamino-pimelic acid-type PGN, the type detected in Gram-negative bacteria, and MDP, respectively. However, thelack of COX-2 protein induction by MDP is an argumentagainst a chief involvement of NOD2, although a morecomplex mechanism involving cooperation of receptorscould be operative.
The possible involvement of PGRP-S is a challenginghypothesis within a rapidly moving field. Recent studieshave disclosed the role of PGRP-S in infection defense onboth intracellular [53] and extracellular bacteria, sincePGPR-S is present in neutrophil extracellular traps andcan bind to both Gram-positive and Gram-negative PGN[54]. On this basis, it seems likely that notions alreadyestablished in insect paradigms, for instance, the PGNcatch-up receptor function of Drosophila PGPR-SA,could be extended to the human model [55]. Irrespec-
Eur. J. Immunol. 2007. 37: 2572–2582 Molecular immunology 2579
f 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

tive of the PRR involved, the present data disclose acentral role for mannan- and PGN-based signatures inthe induction of inflammatory responses in human PMNby showing a strong AA-releasing activity coupled toCOX-2 induction and to the production of otherinflammatory proteins. Translation of preformed mRNAby an mTOR-dependent route seems to be the mostlikely mechanism explaining the induction of some ofthose proteins.
Materials and methods
Reagents
Mucin-3, zymosan particles, soluble mannan from Sacchar-omyces cerevisiae, muramyldipeptide (N-acetylmuramyl-L-ala-nyl-D-isoglutamine), Staphylococcus aureus PGN, and lipotei-choic acid were from Sigma Chemical (St. Louis, MO). Thesynthetic palmitoylated mimic of bacterial lipopeptidesN-palmitoyl-S-[2,3-bis(palmitoyloxy)-2R,S)-propyl]-cysteinyl-(S)-seryl-(S)-lysyl-(S)-lysyl-(S)-lysyl-(S)-lysine trihydrochlor-ide (Pam3CSK4) was from Invivogen. Rabbit Ab reactive to 4E-BP1 phosphorylated at Thr-70 was from Cell SignalingTechnology (#9455).
Cell purification and culture
Human PMN were isolated from buffy coats of healthyvolunteer donors by centrifugation into Ficoll cushions andsedimentation in Dextran T500. Monocytes were isolated bycentrifugation onto Ficoll cushions and adherence to plasticdishes. Differentiation of monocytes into macrophages wascarried out by culture of adhered monocytes for 2 weeks inPrimaria six-well dishes (BD Biosciences), in the presence of5% human serum. Since the experimental design could entaillong incubation periods with regard to the life span of PMN,the occurrence of constitutive apoptosis was addressed bydetecting the surface display of phosphatidylserine with theannexin V-FITC apoptosis detection kit of BD PharMingen todetect early apoptosis, andwith propidium iodide vital stainingto assess late apoptosis. The samples were analyzed usingFACScan and CellQuest program (Becton Dickinson). Theprotocol of study has been approved by the BioethicalCommittee of the Spanish Research Council and Centro deHemoterapia y Hemodonaci�n de Castilla y Le�n.
Immunoblots of COX-2
The amount of protein in each cell lysate sample was assayedusing the Bradford reagent and 50 lg of protein of each samplewere loaded on each lane of a 10% SDS/PAGE gel. Proteinswere transferred to nitrocellulose membranes using a semi-drytransfer system. The membranes were blocked with dry milkand used for immunoblot with a goat polyclonal antiserum(SC-1745) from Santa Cruz Biotechnology. This was followedby incubation with donkey anti-goat IgG-horseradish per-oxidase-conjugated Ab. Detection was performed using theAmersham (Little Chalfont, UK) ECL system.
Assay of proinflammatory protein release by PMN
Culture supernatants were incubated with RayBioJ humanInflammation Antibody Array III and Human Cytokine Anti-body Array III from RayBiotech, and developed according tothe manufacturer's instruction. For this purpose, membraneswere incubated with medium obtained from the incubation ofPMN with mannan and PGN for 18 h, followed by washing,incubation with HRP-conjugated streptavidin, and detectionreactionwith the ECL system. Blot densities were assayed usingQuantity OneJ software from Bio-Rad laboratories (Hercules,CA), and normalized with the positive controls incorporatedinto the membranes. Only variations upon control valuesshowing larger than twofold increase on basal densities wereconsidered positive.
RT-PCR assays for COX-2
Total cellular RNA was extracted by the TRIzol method (LifeTechnologies, Grand Island, NY). cDNA first strand wassynthesized from total RNA by reverse transcription reaction.The reaction mixture contained 0.2 mg/mL total RNA, 2.5 lLH2O, 20 U of RNasin ribonuclease inhibitor, 4 lL buffer 5X,2 lL DTT 0.1 M, 4 lL dNTP 2.5 mM, 1 lL hexanucleotide0.1 mM, and 200 U of Moloney-murine leukemia virus reversetranscriptase. The reaction was carried out at 37�C for 60 minin a volume of 20 lL. The cDNA was amplified by PCR in areaction mixture containing 2 lg of cDNA template, 10 lLH2O, 2.5 lL buffer 10X, 0.75 lL MgCl2 50 mM, 1.0 lL dNTP2.5 mM, 1.25 lL of each forward and reverse primers and0.25 lL of Taq DNA polymerase 5 U/lL. The amplificationprofile for detection included 1 cycle of initial denaturation at94�C for 5 min, 30 cycles of denaturation at 94�C for 30 s,primer annealing at 60�C for 30 s, and extension at 72�C for30 s; 1 cycle of final extension at 72�C for 7 min. Theexpression of b-actin was used as a control for the assay ofconstitutively expressed genes. PCR products were identifiedby automatic sequencing of the DNA eluted from the agarosegel by excision of the band under UV light followed bypurification using a QIAquick PCR purification kit (Qiagen,Valencia, CA). The oligonucleotide primers used in the PCRreactions were selected from different exons and are shown inTable 2.
Table 2. Oligonucleotide primers used for the detection of themRNA encoding for COX-2a)
Exon Orientation Sequence (50–30) Position
1 Forward CAATTGTCATACGACTTGCA 1-20
2 Reverse GTGGGAACAGCAAGGATTTG 187-206
5 Forward TTCAAATGAGATTGTGGAAA 611-619
7 Reverse AGATCATCTCTGCCTGAGTA 896-916
a) Selected from GeneBank AC: NM_000963 [57].
Nieves Fern�ndez et al. Eur. J. Immunol. 2007. 37: 2572–25822580
f 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

Real-time RT-PCR of COX-2
Genomic DNA was removed from purified RNA by treatmentwith DNase (Turbo-DNA freeTM, Ambion) and used for RTreactions. The resulting cDNA was amplified in a PTC-200apparatus equipped with a Chromo4 detector (Bio-Rad) usingSYBR Green I mix containing HotStart polymerase (ABgene).The primers for this reaction were selected in exon 1 and exon2with the purpose of detecting the 50-UTR of COX-2 transcriptsin exon 1. Cycling conditions were as follows: 94�C for 5 min,followed by 40 cycles of 94�C for 30 s, 60�C for 30 s, andextension at 72�C for 30 s. b-Actin and GAPDH were used ashousekeeping genes to assess the relative abundance of COX-2mRNA, using the comparative CT (circle threshold) method forrelative expression. This method allows quantifying therelative expression for a given cDNA using the formula2–DCT, where DCT = DCT
COX-2 - DCThousekeeping gene. Thus, 1
arbitrary unit (AU) corresponds to the expression of thehousekeeping gene and the results of relative expressionchanges between treatments can be expressed by the ratio2–DDCT [56]. This quantitative approach was initially applied tothe study of COX-2 mRNA stability with actinomycin D;however, whereas the expression of mRNA of both b-actin andGADPH did not show significant changes upon stimulationwith PGNandmannan, thesemRNAwere significantly reducedupon actinomycin D treatment. For this reason, results werefinally expressed as percentage of the initial COX-2 mRNAaccording to the formula: (amount of mRNA at a time/amountof mRNA at time 0) � 100.
Acknowledgements: This work was supported bygrants from Plan Nacional de Salud y Farmacia (grantSAF2004–01232), Junta de Castilla y Le�n (GrantCSI05C05), Red Brucella, Red Respira, and Red Tem�ticade Investigaci�n Cardiovascular from Instituto de SaludCarlos III. N.F. is under contract within the Ram�n yCajal Program (Ministerio de Educaci�n y Ciencia ofSpain and Fondo Social Europeo), A.G.V. was therecipient of a grant from Instituto de Salud Carlos III,I.V. was the recipient of a grant from Banco deSantander-Central-Hispano.
References
1 Akira, S. and Takeda, K., Toll-like receptor signaling. Nat. Rev. Immunol.2004. 4: 499–511.
2 Takeda, K. and Akira S., Toll-like receptors in innate immunity. Int.Immunol. 2005. 17: 1–14.
3 Inohara, N., Chamaillard, M., McDonald, C. and Nu�ez, G., NOD-LRRproteins: role in host-microbial interactions and inflammatory disease.Annu. Rev. Biochem. 2005. 74: 355–383.
4 Ting, J. P. and Davis, B. K., CATERPILLER: A novel gene family important inimmunity, cell death, and diseases. Annu. Rev. Immunol. 2005. 23: 387–414.
5 McDonald, C., Inohara, N. and Nu�ez, G., Peptidoglycan signaling ininnate immunity and inflammatory disease. J. Biol. Chem. 2005. 280:20177–20180.
6 Taylor, P.-R., Brown, G. D. Reid, M., Willment, J. A., Martinez-Pomares,L., Gordon, S. and Wong, S. Y., The b-glucan receptor, dectin-1, is
predominantly expressed on the surface of cells of the monocyte/macrophage and neutrophil lineages. J. Immunol. 2002. 169: 3876–3882.
7 East, L. and Isacke, C. M., The mannose receptor family. Biochim. Biophys.Acta 2002. 1572: 364–386.
8 Taylor, P. R., Gordon, S. and Mart�nez-Pomares, L., The mannosereceptor: linking homeostasis and immunity through sugar recognition.Trends Immunol. 2005. 26: 104–110.
9 Hayashi, F., Means, T. K. and Luster, A. D., Toll-like receptors stimulatehuman neutrophil function. Blood 2003 2003. 102: 2660–2669.
10 Francois, S., El Benna, J., Dang, P. M., Pedruzzi, E., Gougerot-Pocidalo,M. A. and Elbim, C., Inhibition of neutrophil apoptosis by TLR agonists inwhole blood: involvement of the phosphoinositide 3-kinase/Akt and NF-jBsignaling pathways, leading to increased levels of Mcl-1, A1, andphosphorylated Bad. J. Immunol. 2005. 174: 3633–3342.
11 Bellocchio, S., Moretti, S. M., Perruccio, K., Fallarino, F., Bozza, S.,Montagnoli, C., Mosci, P. et al., TLRs govern neutrophil activity inaspergillosis. J. Immunol. 2004. 173: 7406–7415.
12 Peters-Golden, M., Canetti, C., Mancuso, P. and Coffey, M.-J.,Leukotrienes: underappreciated mediators of innate immune responses.J. Immunol. 2005. 174: 589–594.
13 Serezani, C. H., Aronoff, D. M., Jancar, S., Mancuso, P. and Peters-Golden, M., Leukotrienes enhance the bactericidal activity of alveolarmacrophages against Klebsiella pneumoniae through the activation ofNADPH oxidase. Blood 2005. 106: 1067–1075.
14 Harizi, H. and Gualde, N., The impact of eicosanoids on the crosstalkbetween innate and adaptive immunity: the key roles of dendritic cells.Tissue Antigens 2005. 65: 507–514.
15 Luster, A. D. and Tager, A. M., T-cell trafficking in asthma: lipid mediatorsgrease the way. Nat. Rev. Immunol. 2004. 4: 711–724.
16 Kunikata, T., Yamane, H., Segi, E., Matsuoka, T., Sugimoto, Y., Tanaka,S., Tanaka, H. et al., Suppression of allergic inflammation by theprostaglandin E receptor subtype EP3. Nat. Immunol. 2005. 6: 524–531.
17 Fukunaga, K., Kohli, P., Bonnans, C., Fredenburgh, L. E. and Levy, B. D.,Cyclooxygenase 2 plays a pivotal role in the resolution of acute lung injury. J.Immunol. 2005. 174: 5033–5039.
18 Gingras, A. C., Raught, B. and Sonenberg, N., mTOR signaling totranslation. Curr. Top. Microbiol. Immunol. 2004. 279: 169–197.
19 Lindemann, S.W., Yost, C. C., Denis, M.M,McIntyre, T.M.,Weyrich, A. S.and Zimmerman, G. A., Neutrophils alter the inflammatory milieu bysignal-dependent translation of constitutive messenger RNAs. Proc. Natl.Acad. Sci. USA 2004. 101: 7076–7081.
20 Yost, C. C., Denis, M. M., Lindemann, S., Rubner, F. J., Marathe, G. K.,Buerke,M., McIntyre, T. M. et al., Activated polymorphonuclear leukocytesrapidly synthesize retinoic acid receptor-a: a mechanism for translationalcontrol of transcriptional events. J. Exp. Med. 2004. 200: 671–680.
21 Mahoney, T. S., Weyrich, A. S., Dixon, D. A., McIntyre, T. M., Prescott, S.M. and Zimmerman, G. A., Cell adhesion regulates gene expression attranslational checkpoints in humanmyeloid leukocytes. Proc. Natl. Acad. Sci.U A 2001. 98: 10284–10289.
22 Valera, I, Gonz�lez Vigo, A., Alonso, S., Barbolla, L. S�nchez Crespo, M.and Fern�ndez, N., Peptidoglycan and mannose-based molecular patternstrigger the arachidonic acid cascade in human polymorphonuclearleukocytes. J. Leukoc. Biol. 2007. 81: 925–933.
23 Honda, Z., Nakamura, M., Miki, I., Minami, M., Watanabe, T., Seyama,Y., Okado, H. et al., Cloning by functional expression of platelet-activatingfactor receptor from guinea-pig lung. Nature 1991. 349: 342–346.
24 Netea, M. G., Ferwerda, G., de Jong, D. J., Jansen, T., Jacobs, L., Kramer,M., Naber, T. H. et al., Nucleotide-binding oligomerization domain-2modulates specific TLR pathways for the induction of cytokine release. J.Immunol. 2005. 174: 6518–6523.
25 Uehori, J., Fukase, K., Akazawa, T., Uematsu, S., Akira, S., Funami, K.,Shingai, M. et al., Dendritic cell maturation induced by muramyl dipeptide(MDP) derivatives: monoacylated MDP confers TLR2/TLR4 activation. J.Immunol. 2005. 174: 7096–7103.
26 Uehara, A., Yang, S., Fujimoto, Y., Fukase, K., Kusumoto, S., Shibata, K.,Sugawara, S. and Takada, H.,Muramyldipeptide and diaminopimelic acid-containing desmuramylpeptides in combination with chemically synthe-
Eur. J. Immunol. 2007. 37: 2572–2582 Molecular immunology 2581
f 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu

sized Toll-like receptor agonists synergistically induced production ofinterleukin-8 in a NOD2- and NOD1-dependent manner, respectively, inhuman monocytic cells in culture. Cell. Microbiol. 2005. 7: 53–61.
27 Zuker, M. and Stiegler, P., Optimal computer folding of large RNAsequences using thermodynamic and auxiliary information. Nucleic AcidsRes. 1981. 9: 133–148.
28 Zuker, M., Mfold web server for nucleic acid folding and hybridizationprediction. Nucleic Acids Res. 2003. 31: 3406–3415.
29 Gingras, A. C., Raught, B., Gygi, S. P., Niedzwiecka, A., Miron,M., Burley,S. K., Polakiewicz, R. D. et al., Hierarchical phosphorylation of thetranslation inhibitor 4E-BP1. Genes Dev. 2001. 15: 2852–2864.
30 Bay�n, Y., Alonso, A. and S�nchez Crespo, M., 4-Trifluoromethylderivatives of salicylate, triflusal and its main metabolite 2-hydroxy-4-trifluoromethylbenzoic acid, are potent inhibitors of nuclear factor jBactivation. Br. J. Pharmacol. 1999. 126: 1359–1366.
31 Aceves, M., Due�as, A., G�mez, C., San Vicente, E., S�nchez Crespo, M.and Garc�a-Rodr�guez, C., A new pharmacological effect of salicylates:Inhibition of NFAT1 function. J. Immunol. 2004. 173: 5721–5729.
32 Fernandez de Arriba, A., Cavalcanti, F., Miralles, A., Bayon, Y., Alonso, A.Merlos, M., Garcia-Rafanell, J. and Forn, J., Inhibition of cyclooxygenase-2 expression by 4-trifluoromethyl derivatives of salicylate, triflusal, and itsdeacetylated metabolite, 2-hydroxy-4-trifluoromethylbenzoic acid. Mol.Pharmacol. 1999. 55: 753–760.
33 Duque, J., Fresno, M. and I�iguez, M. A., Expression and function of thenuclear factor of activated T cells in colon carcinoma cells: involvement inthe regulation of cyclooxygenase-2. J. Biol. Chem. 2005. 280: 8686–8693.
34 Rouzer, C.-A. and Samuelsson, B., Reversible, calcium-dependentmembrane association of human leukocyte 5-lipoxygenase, Proc. Natl.Acad. Sci. USA 1987. 84: 7393–7397.
35 Werz, O., Klemm, J., Samuelsson, B. and R�dmark, O., 5-Lipoxygenase isphosphorylated by p38 kinase-dependent MAPKAP kinases, Proc. Natl. Acad.Sci. USA 2000. 97: 5261–5266.
36 Garc�a, C., Montero, M., Alvarez, J. and S�nchez Crespo, M., Biosynthesisof platelet-activating factor (PAF) induced by chemotactic peptide ismodulated at the lyso-PAF:acetyl-CoA acetyltransferase level by calciumtransient and phosphatidic acid. J. Biol. Chem. 1993. 268: 4001–4008.
37 Garc�a Rodr�guez, C., Montero, M., Alvarez, J., Garcia-Sancho, J. andS�nchez Crespo, M., Dissociation of platelet-activating factor productionand arachidonate release by the endomembrane Ca2+-ATPase inhibitorthapsigargin. Evidence for the involvement of a Ca2+-dependent route ofpriming in the production of lipid mediators by human polymorphonuclearleukocytes. J. Biol. Chem. 1993. 268: 24751–24757.
38 Rubin, B. B., Downey, G. P., Koh, A., Degousee, N., Ghomashchi, F.,Nallan, L., Stefanski, E. et al., Cytosolic phospholipase A2a is necessary forplatelet-activating factor biosynthesis, efficient neutrophil-mediated bacter-ial killing, and the innate immune response to pulmonary infection: cPLAA2a does not regulate neutrophil NADPH oxidase activity. J. Biol. Chem.2005. 280: 7519–7529.
39 Dziarski, R. and Gupta, D., Staphylococcus aureus peptidoglycan is a Toll-like receptor 2 activator: a reevaluation. Infect. Immun. 2005. 73:5212–5216.
40 Baker, B.-S, Powles, A. and Fry, L., Peptidoglycan: a major aetiologicalfactor for psoriasis? Trends Immunol. 2006. 27: 545–551.
41 Maloney, C. G., Kutchera, W. A., Albertine, K. H., McIntyre, T. M.,Prescott, S. M. and Zimmerman, G. A., Inflammatory agonists inducecyclooxygenase type 2 expression by human neutrophils. J. Immunol. 1998.160: 1402–1410.
42 Nagano, S., Otsuka, T., Niiro, H., Yamaoka, K., Arinobu, Y., Ogami, E.,Akahoshi, M. et al., Molecular mechanisms of lipopolysaccharide-induced
cyclooxygenase-2 expression in human neutrophils: involvement of themitogen-activated protein kinase pathway and regulation by anti-inflam-matory cytokines. Int. Immunol. 2002. 14: 733–740.
43 Pouliot, M., Fiset, M. E., Masse, M., Naccache, P. H. and Borgeat, P.,Adenosine up-regulates cyclooxygenase-2 in human granulocytes: impact onthe balance of eicosanoid generation. J. Immunol. 2002. 169: 5279–5286.
44 Brown, E. J. and Schreiber, S. L., A signaling pathway to translationalcontrol. Cell 1996, . 86: 517–520.
45 Beretta, L., Gingras, A. C., Svitkin, Y. V., Hall, M. N. and Sonenberg, N.,Rapamycin blocks the phosphorylation of 4E-BP1 and inhibits cap-dependent initiation of translation. EMBO J. 1996. 15: 658–664.
46 von Manteuffel, S. R., Gingras, A. C., Ming, X. F., Sonenberg, N. andThomas, G., 4E-BP1 phosphorylation is mediated by the FRAP-p70s6kpathway and is independent of mitogen-activated protein kinase. Proc. Natl.Acad. Sci. USA 1996. 93: 4076–4080.
47 Graves, L. M., Bornfeldt, K. E., Argast, G. M., Krebs, E. G., Kong, X., Lin, T.A. and Lawrence, J. C. Jr., cAMP- and rapamycin-sensitive regulation of theassociation of eukaryotic initiation factor 4E and the translational regulatorPHAS-I in aortic smooth muscle cells. Proc. Natl. Acad. Sci. USA 1995. 92:7222–7226.
48 Lin, T. A., Kong, X., Saltiel, A. R., Blackshear, P. J. and Lawrence, J. C. Jr.,Control of PHAS-I by insulin in 3T3-L1 adipocytes. Synthesis, degradation,and phosphorylation by a rapamycin-sensitive and mitogen-activatedprotein kinase-independent pathway. J. Biol. Chem. 1995. 270:18531–18538.
49 Dean, J. L., Brook, M., Clark, A. R. and Saklatvala J., p38 mitogen-activated protein kinase regulates cyclooxygenase-2 mRNA stability andtranscription in lipopolysaccharide-treated humanmonocytes. J. Biol. Chem.1999. 274: 264–269.
50 Monick, M. M., Robeff, P. K., Butler, N. S., Flaherty, D. M., Carter, A. B.,Peterson, M. W. and Hunninghake, G. W., Phosphatidylinositol 3-kinaseactivity negatively regulates stability of cyclooxygenase 2 mRNA. J. Biol.Chem. 2002. 277: 32992–33000.
51 Dixon, D. A., Tolley, N. D., Bemis-Standoli, K., Martinez, M. L., Weyrich,A. S., Morrow, J. D., Prescott, S. M. and Zimmerman, G. A., Expression ofCOX-2 in platelet-monocyte interactions occurs via combinatorial regulationinvolving adhesion and cytokine signaling. J. Clin. Invest. 2006. 116:2727–2738.
52 Travassos, L. H., Girardin, S. E., Philpott, D. J., Blanot, D., Nahori, M. A.,Werts, C. and Boneca, I. G., Toll-like receptor 2-dependent bacterial sensingdoes not occur via peptidoglycan recognition. EMBO Rep. 2004. 10:1000–1006.
53 Dziarski, R., Platt, K. A., Gelius, E., Steiner, H. and Gupta, D., Defect inneutrophil killing and increased susceptibility to infection with nonpatho-genic gram-positive bacteria in peptidoglycan recognition protein-S (PGRP-S)-deficient mice. Blood 2003. 102: 689–697.
54 Cho, J. H., Fraser, I. P., Fukase, K., Kusumoto, S., Fujimoto, Y., Stahl, G. L.and Ezekowitz, R. A., Human peptidoglycan recognition protein-S is aneffector of neutrophil-mediated innate immunity. Blood 2005. 106:2551–2558.
55 Kang, D., Liu, G., Lundstrom, A., Gelius, E. and Steiner, H., Apeptidoglycan recognition protein in innate immunity conserved frominsects to humans. Proc. Natl. Acad. Sci. USA 1998. 95: 10078–10082.
56 Pfaffl, M. W.A new mathematical model for relative quantification in realtime RT-PCR. Nucleic Acids Res. 2001. 29: 2003–2007.
57 Jones, D. A., Carlton, D. P., McIntyre, T. M., Zimmerman, G. A., andPrescott, S. M., Molecular cloning of human prostaglandin endoperoxidesynthase type II and demonstration of expression in response to cytokines. J.Biol. Chem. 1993. 268: 9049–9054.
Nieves Fern�ndez et al. Eur. J. Immunol. 2007. 37: 2572–25822582
f 2007 WILEY-VCH Verlag GmbH & Co. KGaA, Weinheim www.eji-journal.eu










![Target of Rapamycin Signaling in Plant Stress …Update on Target of Rapamycin Signaling in Plant Stress Responses Target of Rapamycin Signaling in Plant Stress Responses1[OPEN] Liwen](https://img.pdfslide.net/doc/110x75/5f05e4b57e708231d4153f1e/target-of-rapamycin-signaling-in-plant-stress-update-on-target-of-rapamycin-signaling.jpg)












![Mannan, M. Sam [Resume]](https://img.pdfslide.net/doc/110x75/589aefbd1a28abf6458bd237/mannan-m-sam-resume.jpg)





