Embed Size (px)
Citation preview

INTRODUCTION
A The subfamily Carolliinae (Phyllostomi-dae-Chiroptera) constitutes a group of frugiv-orous Neotropical bats widely distributedthroughout the New World. This subfamily iscomposed of two genera, Carollia and Rhino-phylla and is characterized by an intragenerickaryotypic stability, with multiple XX/XY1Y2and simple XX/XY chromosomal sexualdetermination systems (BAKER et al. 1989).
According to RUSSEL (1963), SOLARI (1994)and DELOBEL et al. (1998), the X chromo-some has an inactivation center responsiblefor a phenomenon called spreading effect ofthe chromatin inactivation, where it is proba-ble that inhibition occurs in genic transcrip-tion in the autosomal regions translocated tothe X chromosome in mammals.
In some species that present translocationsin sexual chromosomes, such as the mousewith Searle translocation (T(X;16) 16 H) inac-tivation of the sexual chromosomes occurstogether with the translocated autosomalregion (READER and SOLARI 1969; SOLARI
CARYOLOGIA Vol. 57, no. 1: 1-9, 2004
Meiotic analyses of the sex chromosomes inCarolliinae-Phyllostomidae (Chiroptera): NORseparates the XY1Y2 into two independent parts
RENATA C. RODRIGUES NORONHA1, *, CLEUSA Y. NAGAMACHI1, JULIO C. PIECZARKA1,SUELY MARQUES-AGUIAR2, MARIA DE FÁTIMA LIMA DE ASSIS3 and REGINA M. DE SOUZA BARROS1
1 Laboratório de Citogenética Animal, Departamento de Genética, CCB, UFPA, Av. Perimetral S/N, CEP 66.075-900,Belém, PA, Brasil.
2 Departamento de Zoologia, Museu Paraense Emilio Goeldi, Belém, Pará, Brasil.3 Instituto Evandro Chagas, Ananindeua, Pará, Brasil.
Abstract - Meiotic analyses of the sex chromosomes were made in two differentspecies belonging to the subfamily Carollinae (Phyllostomidae, Chiroptera): Rhino-phylla pumilio and Carollia perspicillata. These species have different sex chromo-some systems. Through banding techniques, chromosomes homologies betweenCarollia perspicillata and Rhinophylla pumilio were not found, possibly because Car-ollia species have highly rearranged chromosomes. Probably, Rhinophylla is betterassociated to Phyllostomini tribe. Sex vesicle pairing behavior at the meioticprophase was analyzed in both species. R. pumilio had a simple sex chromosomesystem (XX/XY), the XY pair showed an asynchronic behavior compared to theautosomal bivalents. The sex vesicle showes the X and Y pairing in the pseudoau-tosomal region. Moreover, a folding of the X-asynaptic axis on itself and con-densed differentiated axes were also identified. In C. perspicillata, with a multiplesex chromosome system (XY1Y2), the segments Xp-Y1 and Xq-Y2 paired inde-pendently from each other. Compared to the autosomes, the sex vesicle showed aprecocious pairing behavior. In C. perspicillata, a NOR stained with silver was local-ized in the X rearranged chromosome. Possibly, this region separates the XY1Y2trivalent in two independent parts: one involving the X and Y2 autosomal with tran-scriptional activity, and the other corresponding to the original X and Y sex pair.
Key words: Carolliinae, Meiosis, genic activity, translocations, NOR.
* Corresponding author: fax +55-21-91-211-1627; e-mail:[email protected]

1971). The hypothesis of inactivation of the Xchromosome assumes that in all heterogamet-ic organisms the X chromosome is normallydeactivated during a critical stage of sper-matogenesis. The same hypothesis suggeststhat the inactivation center of the X chromo-some (Xq3 in humans) leads to the inactiva-tion of X (sexual chromatin) in somatic cellsand to the formation of the sex vesicle in sper-matogenesis (SOLARI 1994).
In Gerbillus sp. (RANTOMPONIRINA et al.1986) and Stenodermatinae (NORONHA et al.2001), careful description of the structure ofthe sexual pair in composite and multiple sys-tems during meiosis in male, showed the pres-ence of an intercalated heterochromaticregion that acts as an “isolator”, allowing theindependent behavior of the true (inactive)and autosomal (active) sexual segments. Insome species of mammals, the sex vesicle (XYbody) is associated with the nucleolar mater-ial in certain stages of the meiotic prophase,such as, for example, in the mouse Mus mus-culus (SOLARI, 1994). Nonetheless, few speciespresent the nucleolar organizer region (NOR)located in a sexual chromosome, such as thebat Carollia perspicillata (X chromosome)(VARELLA-GARCIA et al. 1989), Canis famil-iaris (PATHAK et al. 1982) and the primate Cal-lithrix jacchus (NAGAMACHI and FERRARI1986) and Hylobates syndactylus (LEDBETTER1981).
Through use of optical microscopy weintended to describe the meiotic behavior ofthe sex chromosomes comparing the two sys-tems of the chromosomal sex determination ofthe Carolliinae, Phyllostomid bats.
MATERIAL AND METHODS
The sample for this study is made up of 6 exem-plars (5M, 1F) of Carollia perspicillata and 2 (M) ofRhinophylla pumilio, collected at five localities inPará (Table 1).
� Submitted to the conventional meiosis tech-nique, following EICHER (1966): the cells werestained with Giemsa.
� Submitted to the synaptonemic complex tech-nique, following VERMA and BABU (1995): thesynaptonemic complexes were marked with sil-ver nitrate.
� The mitotic cells were extracted, according toFORD and HAMERTON (1956) and fibroblast celllines, tissue culture. The metaphasic chromo-somes were submitted to the NOR technique,following HOWELL and BLACK (1980); the nucle-olar organizer regions were marked with silvernitrate.The selected cells were photographed in a Carl
Zeiss III photomicroscope, 100X immersion lens,optovar 1,25, with a green filter, ASAs 3,2 (DIN 6)and 6,3 (DIN 9). The films employed were AgfaCopex Pan and Imagelink HQ, developed andfixed with Kodak D76 developer and Kodak fixer,respectively. The images were copied onKodabrome Print F3 paper (Kodak) and digitized.
RESULTS
Carollia perspicillata – This species present-ed 2n=20/21 and a multiple system of chromo-somal sexual determination of the XX/XY1Y2type. Analysis of the NORs in the mitotic cells,allowed us to identify them only in the sec-ondary interstitial constriction of the long arm
2 NORONHA, NAGAMACHI, PIECZARKA, MARQUES-AGUIAR, ASSIS and BARROS
Table 1 – Species identified, with references to the techniques applied and collection site.
Species N° (MPEG) N° (UFPA) Sex Collection site
Carollia perspicillataa 26435 AL 13 � M MPEG Park, Belém, Pará, Brazil26465 AL 46 � M Mosqueiro Island, Belém, Pará, Brazil26489 AL 71 � F Mosqueiro Island, Belém, Pará, Brazil26496 AL 77 �� M Campus of the UFPA, Belém, Pará, Brazil27499 AL 195� M Campus of the UFPA, Belém, Pará, Brazil27519 AL 215� M MPEG Park, Belém, Pará, Brazil
Rhinophylla pumiliob 26508 AL 93 � M Santa Rosa, Vigia, Pará, Brazil27279 AL129 � M Vila de Tacajós, Sta. Izabel, Pará, Brazil
a – species with multiple XY1Y2 sexual system. b – species with simple XY sexual system.The specimens were collected from natural populations and identified by Dr. Suely Marques-Aguiar, of the Zoology Department(MPEG) and are part of the collection at the Museu Paraense Emílio Goeldi.

of the X chromosome, with two stainings in thefemales (Fig. 1a) and one in the male (Fig. 1b).
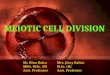
Through meiotic analysis of the male, 9autosomal bivalents and one sexual trivalentformed by XY1Y2 were observed. In the initialstages of meiosis a typically condensed sex vesi-cle is formed (Figs. 2a and 2b). In the diploteneand diacinesis, the formation of chiasmas in allthe extension of the 9 autosomal bivalents wasobserved (Fig. 2c); while in the sexual triva-
lent the formation of at least three chiasmas inthe distal half of the long arm of X with itshomologue Y2 (Fig. 2d) and point to pointpairing of Y1 with the extremity of the shortarm of X were visualized (Figs. 2e and 2f). Inmetaphase I, with 9 bivalents and one sexualtrivalent, one may observe a more precociousbehavior of the sexuals, with the Y1 open inrelation to the X axis, while Y2-X are stillpaired in a manner similar to that of the biva-
MEIOTIC ANALYSES OF THE SEX CHROMOSOMES IN CAROLLIINAE-PHYLLOSTOMIDAE 3
Fig. 1 – Ag-NOR staining in the interstitial region of the long arm of the X chromosome ofCarollia perspicillata, with 2n=20/21 XY1Y2system (parcial view of the cell). a) two stainingsin the females and b) one staining in the males.
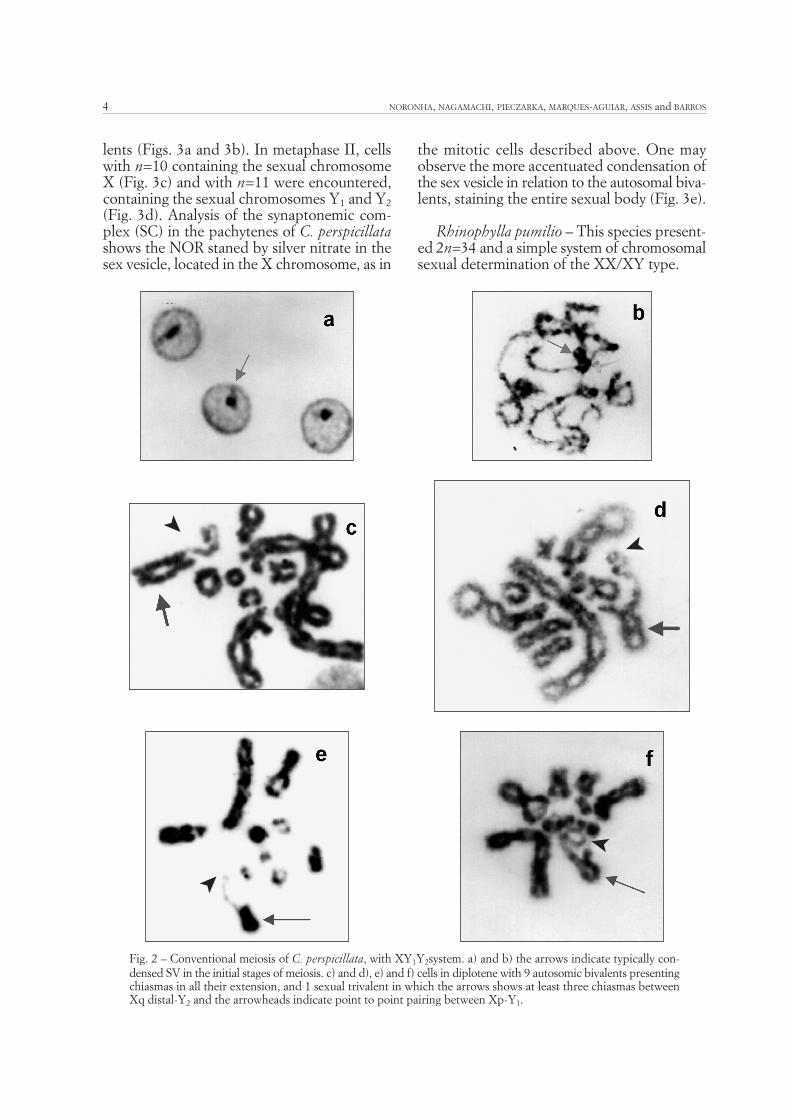
lents (Figs. 3a and 3b). In metaphase II, cellswith n=10 containing the sexual chromosomeX (Fig. 3c) and with n=11 were encountered,containing the sexual chromosomes Y1 and Y2(Fig. 3d). Analysis of the synaptonemic com-plex (SC) in the pachytenes of C. perspicillatashows the NOR staned by silver nitrate in thesex vesicle, located in the X chromosome, as in
the mitotic cells described above. One mayobserve the more accentuated condensation ofthe sex vesicle in relation to the autosomal biva-lents, staining the entire sexual body (Fig. 3e).
Rhinophylla pumilio – This species present-ed 2n=34 and a simple system of chromosomalsexual determination of the XX/XY type.
4 NORONHA, NAGAMACHI, PIECZARKA, MARQUES-AGUIAR, ASSIS and BARROS
Fig. 2 – Conventional meiosis of C. perspicillata, with XY1Y2system. a) and b) the arrows indicate typically con-densed SV in the initial stages of meiosis. c) and d), e) and f) cells in diplotene with 9 autosomic bivalents presentingchiasmas in all their extension, and 1 sexual trivalent in which the arrows shows at least three chiasmas betweenXq distal-Y2 and the arrowheads indicate point to point pairing between Xp-Y1.
➤
➤
➤
➤
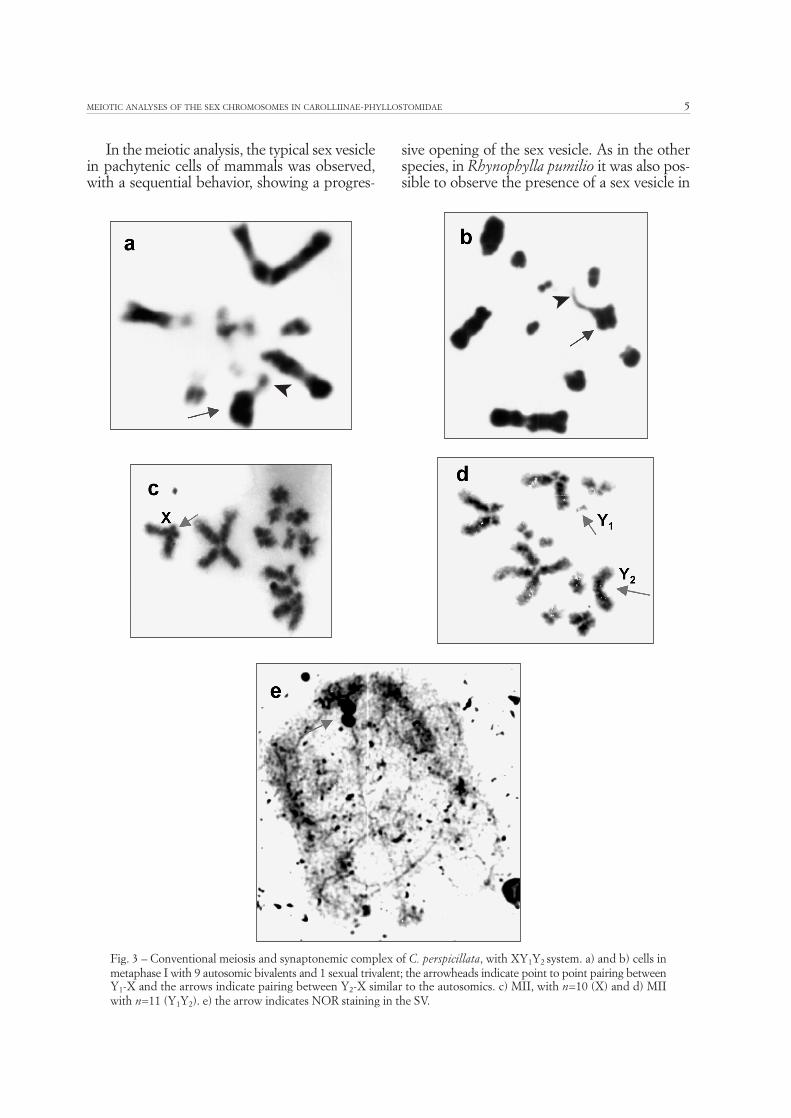
In the meiotic analysis, the typical sex vesiclein pachytenic cells of mammals was observed,with a sequential behavior, showing a progres-
sive opening of the sex vesicle. As in the otherspecies, in Rhynophylla pumilio it was also pos-sible to observe the presence of a sex vesicle in
MEIOTIC ANALYSES OF THE SEX CHROMOSOMES IN CAROLLIINAE-PHYLLOSTOMIDAE 5
Fig. 3 – Conventional meiosis and synaptonemic complex of C. perspicillata, with XY1Y2 system. a) and b) cells inmetaphase I with 9 autosomic bivalents and 1 sexual trivalent; the arrowheads indicate point to point pairing betweenY1-X and the arrows indicate pairing between Y2-X similar to the autosomics. c) MII, with n=10 (X) and d) MIIwith n=11 (Y1Y2). e) the arrow indicates NOR staining in the SV.
➤
➤
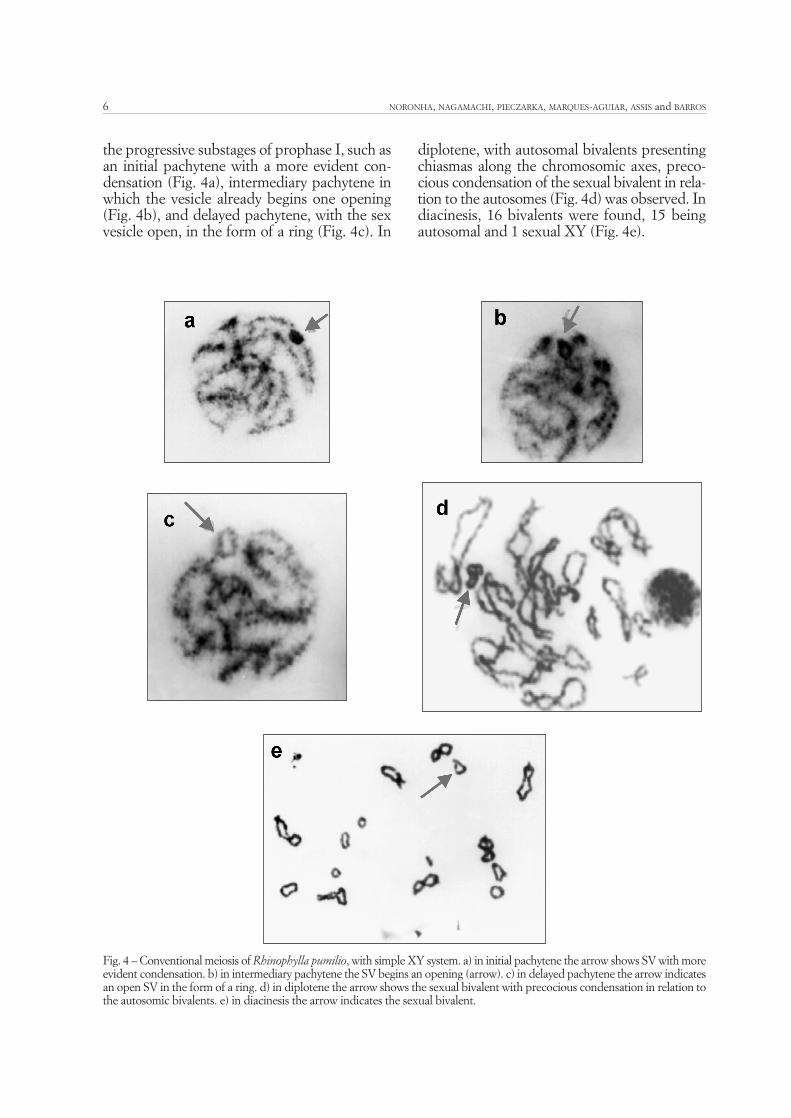
the progressive substages of prophase I, such asan initial pachytene with a more evident con-densation (Fig. 4a), intermediary pachytene inwhich the vesicle already begins one opening(Fig. 4b), and delayed pachytene, with the sexvesicle open, in the form of a ring (Fig. 4c). In
diplotene, with autosomal bivalents presentingchiasmas along the chromosomic axes, preco-cious condensation of the sexual bivalent in rela-tion to the autosomes (Fig. 4d) was observed. Indiacinesis, 16 bivalents were found, 15 beingautosomal and 1 sexual XY (Fig. 4e).
6 NORONHA, NAGAMACHI, PIECZARKA, MARQUES-AGUIAR, ASSIS and BARROS
Fig. 4 – Conventional meiosis of Rhinophylla pumilio, with simple XY system. a) in initial pachytene the arrow shows SV with moreevident condensation. b) in intermediary pachytene the SV begins an opening (arrow). c) in delayed pachytene the arrow indicatesan open SV in the form of a ring. d) in diplotene the arrow shows the sexual bivalent with precocious condensation in relation tothe autosomic bivalents. e) in diacinesis the arrow indicates the sexual bivalent.

DISCUSSION
Rhynophylla pumilio and Carollia perspicil-lata, despite being in the same subfamily havedifferent sexual systems: simple XX/XY andmultiple of type XY1Y2, respectively. TheXY1Y2 system in males of C. perspicillata orig-inated after the translocation of one autosometo the X. Thus, the autosomal homologuebegins to act as sexual, calling Y2 and Y1 is thereal one of the system. The occurrence of mul-tiple sexual systems in independent lineageswithin the family Phyllostomidae, involves dif-ferent autosomes in translocation to the X.This indicates that the sexual systems arosemore than once during the evolutionary histo-ry of the family.
STOCK (1975), compared the karyotypes ofC. castanea with C. perspicillata. They observedhomology in the morphology of all the chro-mosomes among the two species. It was possi-ble to identify the ancestral autosome of C.castanea, which was translocated to C. perspic-illata, which suggests an autosome-X fusion,leading to the formation of the multipleXY1Y2system, in C. perspicillata.
Nucleolar Organizer Region – Analysis ofthe distribution of the NORs in C. perspicilla-ta showed little variability, which was foundonly in the interstitial region of the long arm ofX (Fig. 1). Staining was not observed in the Y2homologue to autosome-X, indicating thestaining of the nucleolar organizer region onlyin the two original X chromosomes of the C.perspicillata females. According to FAGUNDES(1993), in the majority of the cells analyzed infemale rodents, only one of the X chromo-somes presented a positive staining in theNORs, although in a few metaphases the twoX were marked. These data confirm thehypothesis of the incomplete inactivation ofone of the X chromosomes of the female mam-mals, in which some regions escape thisprocess, and remain active during the entirecellular cycle. According to YONENAGA et al.(1983), the two Xs may be genic activity at thesame time.
According to GOODPASTURE and BLOOM(1975), there is one NOR in the telomere ofthe long arm of Y in C. castanea. Thus, it isprobable that there was a loss of this NORduring evolution, leading to a disparity of
genic activity (DNAr) in males of C. perspicil-lata with only one staining in X.
According to HSU et al. (1968), the originalNOR may have been located in the terminalportion of the long arm of X in C. castanea,thus, after fusion with one autosome, NORbecomes interstitial (Xq) in C. perspicillata;however, GOODPASTURE and BLOOM (1975),observed NOR staining in the distal segmentof the short arm of X rearranged in C. cas-tanea. The data obtained in this paper agreewith the staining of terminal Xp NOR of theancestral species C. castanea observed byGOODPASTURE and BLOOM (1975), suggestingan alternative hypothesis in the evolution of Xin C. perspicillata, in which the autosome wastranslocated to the short arm of X. After thisrearrangement the Xp de C. castanea begins tobe Xq, originating the interstitial Xq NOR ofC. perspicillata.
Meiosis – In Rhinophylla pumilio with sim-ple XY sexual system, in the pachytenic cellsthis pair formed a typically condensed sex vesi-cle with precocious behavior in relation to theautosomes, as in the majority of the eutherianmammals. In C. perspicillata, with a multiplesystem, one may note a formation of typical SVin the pachytenes, and, beginning in the moreadvanced stages, such as diplotene and diaci-nesis, the sexual trivalent (XY1Y2) presentsdifferences in behaviors of the axes of the orig-inal XYs in relation to the translocated auto-some. Based on visualization of the formationof at least three chiasmas in the autosome-Xpairing (Fig. 2d), one may prove genic activityin this region of the sexual trivalent, as well asin the autosomal bivalents, with chiasmas atvarious chromosomic points.
It is known that the autosome translocatedto the X of C. perspicillata is separated by asecondary constriction, in which the NOR, ofthe original X axis is located. From this data,one may propose a hypothesis similar to theone applied for groups of mammals that pre-sent autosome-sexual rearrangements, inwhich the original segment of X is separatedby a heterochromatic block of the autosomalsegment. This block probably impedes the dif-fusion of inactivation to the autosomal seg-ment of X, during meiosis (PATHAK et al. 1973;STOCK 1975; KASAHARA and DUTRILLAUX1983; DELOBEL et al. 1998; NORONHA et al.
MEIOTIC ANALYSES OF THE SEX CHROMOSOMES IN CAROLLIINAE-PHYLLOSTOMIDAE 7

2001). The same hypothesis applies to theNOR of C. perspicillata, which impedes thespread of inactivation and maintains the sexu-al trivalent (XY1Y2) in two independent partsduring meiosis, as attested by genic activity inthe region of autosomal pairing and conden-sation in the sexual segment.
In C. perspicillata, instead of heterochro-matin, there is possibly a nucleolar organizerregion, which blocks the diffusion of inacti-vation to the autosome, impeding its progres-sion to the segments of the translocated auto-somes, allowing the production of viablegametes.
According to DELOBEL et al. (1998), thepulling of the autosomes to within the SVmake them hypercondensed, as happens withthe sexual chromosomes. This explain thedegeneration of the spermatocyts, suggestingautosomal inactivation through a diffusioneffect of inactivation. At no moment we detectdegeneration of the spermatocyts of C. per-spicillata but instead the presence ofmetaphase II with differences in the haploidnumber, justified by the segregation of the twoYs to the pole opposite X, assuring the pro-duction of balanced gametes. In C. perspicilla-ta, the autosome translocated to X continuesits original genic activity, without interfering inthe function of sexual determination.
Synaptonemic complex – Analysis of thebehavior of the sexual axes through the SC inC. perspicillata was not as instructive as con-ventional meiosis, due to NOR staining in X,making the delineation of chromosomic axesdifficult. In any case, the differentiation of thecondensation of the sex vesicle related to theautosomes was observed. The NORs wereobserved in initial pachytene, linked to theaxis of the X chromosome. In this fashion, thischromosome was easily recognized in the sexvesicle of C. perspicillata. According to PATTONand GARDNER (1971), the NORs began todegrade with the progress of pachytene, dis-connecting from the chromosomic axes andappearing as points scattered throughout thecell. Our data confirms the initial phase ofpachytene observed in figure 3e, in which theNOR stays intact in the X chromosome, cor-roborating the mitotic results, in which only insexual chromosome (X) is found stainingnucleolar organizer region.
Acknowledgements – The authors are gratefulto CAPES, CNPq, UFPA, Brazil, for financialsupport. We also thank the students and techni-cians from Laboratório de Citogenética-UFPA,Emilio Goeldi Museum and Evandro ChagasInstitute who contributed to this research, espe-cially to the Chiroptera Team (Nelson, Luis andAdailton).
REFERENCES
BAKER R.J., HOOD C.S. and HONEYCUTT R.L., 1989– Phylogenetic relationships and classification ofthe higher categories of the New World bat fami-ly Phyllostomidae. Systematic Zoology, 38: 228-238.
DELOBEL B., DJLELATI R., GABRIEL-ROBEZ O., CRO-QUETTE M.F., ROUSSEAUX-PREVOST R.,ROUSSEAUX J., RIGOT J.M. and RUMPLER Y.,1998 – Y-autosome translocation and infertility:usefulness of molecular, cytogenetic and meioticstudies. Human Genetics, 102: 98-102.
EICHER E., 1966 – An improve air-drying techniquefor recovery of all stages of meiosis in the mam-malian testis. Mammal. Chrom. Newsl. 20: 74.
FAGUNDES V., CHRISTOFF A.U. and YONENAGA-YASSUDA Y., 1998 – Extraordinary chromosomalpolymorphism with 28 different karyotypes in theneotropical species Akodon cursor (Muridae, Sig-modontinae), one of the smallest diploid numberin rodents (2n=16, 15 and 14). Hereditas, 129:263-274.
FORD C.E. and HAMERTON J.L., 1956 – A colchicine,hypotonic-citrate, squash sequence for mammalianchromosomes. Staining Technology, 31: 247-251.
GOODPASTURE C. and BLOOM S.E., 1975 – Visual-ization of nuclear organizer regions in mammalianchromosomes using silver staining. Chromoso-ma, 53:37-50.
HOWELL W.M. and BLACK D.A., 1980 – Controlledsilver-staining of nucleolar organizer regions withprotective colloidal developer: a 1-step method.Experientia, 36: 1014-1015.
HSU T.C., BAKER R.J. and UTAKOJI T., 1968 – Themultiple sex chromosome system of Americanleaf-nosed bats (Chiroptera, Phyllostomidae).Cytogenetics, 7: 27-38.
KASAHARA S. and DUTRILLAUX B., 1983 – Chromo-some banding patterns of four species of bats,with special reference to a case of X-autosometranslocation. Annals Genetics, 26: 197-201.
LEDBETTER D.H., 1981 – NOR-bearing Y chromo-some in a primate, Hylobates (Symphalangus)syndactylus. Cytogenetics Cell Genetics, 29: 250-252.
8 NORONHA, NAGAMACHI, PIECZARKA, MARQUES-AGUIAR, ASSIS and BARROS

NAGAMACHI C.Y. and FERRARI I., 1986 – Cytoge-netic studies of Callitrix jacchus (Callithricidae,Platyrrhini) from two diferent sites in Brazil: II.NOR on the Y chromosome. Mammalian Chro-mosomes Newsletter, 27: 81-86.
NORONHA R.C.R., NAGAMACHI C.Y., PIECZARKAJ.C., MARQUES-AGUIAR S., BARROS R.M.S., 2001– Sex-autosome translocations: meiotic behavioursuggests an inativation block with permanenceof autosomal gene activity in phyllostomid bats.Caryologia, 54: 267-277.
PATHAK S., HSU T.C. and UTAKOJI T., 1973 – Rela-tionships between patterns of chromosome bandingand DNA synthetic sequences: a study on the chro-mosomes of the Seba’s fruit bat Carollia perspicil-lata. Cytogenetic Cell Genetics, 12: 157-164.
PATHAK S., VAN TUINEN P. and MERRY D.E., 1982 –Heterochromatin, synaptonemal complex, andNOR activity in the somatic and germ cells of amale domestic dog, Canis familiaris (Mammalia,Canidae). Cytogenetics Cell Genetics, 34: 112.
PATTON J.C. and GARDNER A.L., 1971 – Parallelevolution of multiple sex-chromosome systems inthe Phyllostomatid bats, Carollia and Choeronis-cus. Experientia, 27: 105-106.
RANTOMPONIRINA C., VIEGAS-PEQUIGNOT E.,DUTRILLAUX B., PETTER F. and RUMPLER Y.,1986 – Synaptonemal complexes in Gerbillidae:probable role of intercalated heterochromatin ingonosome-autosomes translocations. CytogeneticsCell Genet., 43: 161.
READER C. and SOLARI A.J., 1969 – The histologyand cytology of the seminiferous epithelium ofthe maouse with Searle’s X-autosome transloca-tion. Acta Physiol. Lat. Am., 19: 249.
RUSSEL L.B., 1963 – Mammalian X-chromosomeaction: inactivation limited in spread and inregion of origin. Science, 140: 976.
SOLARI A.J., 1971 – The behaviour of chromosomalaxes in Searle’s X autosome translocation. Chro-mosoma, 34: 99.
–, 1994 - Sex chromosomes and sex determination invertebrates. CRC Press.
SOLARI A.J. and PIGOZZI, 1994 – Fine structure ofthe XY body in the XY1Y2 trivalent of the batArtibeus lituratus. Chromosome Research, 2: 53-58.
STOCK A.D., 1975 – Chromosome banding patternhomology and its phylogenetic implications in thebat genera Carollia and Choeroniscus. Cytoge-netic Cell Genetics, 14: 34-41.
VARELLA-GARCIA M., MORIELLE-VERSUTE E. andTADDEI V.A., 1989 – A survey of cytogenetic dataon brazilian bats. Revista Brasileira de Genética,12: 761-793.
VERMA R.S. and BABU A., 1995 – Human Chromo-somes: Principles and techniques. Mcgrew-Hill,inc. Health professions division.
YONENAGA Y., 1972 – Polimorfismos cromossômicosem roedores brasileiros. Tese de Doutoramento,Departamento de Biologia, Instituto de Bio-ciências, USP, São Paulo.
YONENAGA Y., ASSIS M.F.L., KASAHARA S., L’AB-BATE M. and SOUZA M.J., 1983 – Nucleolar orga-nizer regions in Akodon arviculoides (Cricetidae,Rodentia): evidence for the activity of rDNAgenes in both X chromosomes of females. Cyto-genet. Cell Genet., 35: 143-147.
Received November 5, 2002; accepted January 30, 2003
MEIOTIC ANALYSES OF THE SEX CHROMOSOMES IN CAROLLIINAE-PHYLLOSTOMIDAE 9