Embed Size (px)
Citation preview

Instructions for use
Title Melilotus 属の種間雑種に関する育種学的基礎研究 : 第Ⅸ報 種間雑種、Melilotus taurica×M. alba, M.taurica×M. wolgica, および M. dentata×M. taurica の細胞学的研究
Author(s) 佐野, 芳雄; 喜多, 富美治
Citation 北海道大学農学部邦文紀要, 9(1), 1-9
Issue Date 1973-12-15
Doc URL http://hdl.handle.net/2115/11849
Type bulletin (article)
File Information 9(1)_p1-9.pdf
Hokkaido University Collection of Scholarly and Academic Papers : HUSCAP

Melilotus属の種間雑種に関する育種学的基礎研究
第 IX報種間雑種,Melilotus taurica x M. alba,
λ1. taωゴcax M. wolgicaおよび M.dentata
xM. tauricaの細胞学的研究1)
佐野芳雄
(北海道大学農学部育種学教室)
喜多富美治
(北海道大学農学部附属農場)
Studies of Interspecific Hybrids in the Genus Melilotω
from the Plant Breeding Standpoint
IX. Cytological Studies of the Interspecific Hybrids,
Melilotus taurica x M. alba, M. taurica x M.
wolgica, and M. dentata x M. taurica
Yoshio SANO and Fumiji KITA
(Plant Breeding Institute, and Experimental Farm,
Faculty of Agriculture, Hokkaido University, Sapporo, Japan)
Received, January 12, 1973
1. 緒言
Melilotus属すなわち Sweetcloverの種間交雑によ
る育種の基礎研究として,著者らは種間相互の細胞学的
関係を明らかにする目的で,一連の交雑実験を遂行中で
ある。
Eumelilotus亜属には 9種が分類され,これらの間に
おける種間雑種 F¥の多くは幼苗時に葉緑素欠乏を呈し,
これら種間の隔離機構の重要な要因の 1っと考えられて
いる (SMITH1954)。
さらに,これらいくつかの種聞には相互転Illiに由来す
る染色体構造差の存在することが報告され (BRINGHU・
RST 1951, JARANOWSKI 1961,喜多 1962),EUlIle-
lilotus lI!!属の系統発生上,現!I!tある問題を提起しつつ
ある。
著者らは,EUlIlelilotus直属に含まれる多くの系統を
用いて種間交雑を行ない,いくつかの組合せにおいて種
1) 北海道大学農学部附属農場業績
間雑種 F1を得開花登熟まで育成することが出来た。本
報においては,そのうち 4種 6系統聞の交雑より得られ
た種間雑種 Flについて詳細な観察を行ない, これら種
聞の細胞学的関係に関し若干の興味ある結果が得られた
のでここに取纏め報告する。
なお本実験に供試した材料は DR.H. J. GORZ2)およ
び DR.G. A. STEVENSON3)より提供せられたもので,
ここに記して深甚なる謝意を表する。また実験遂行と取
纏めに関し;種々懇篤な御教示をl!易わった北海道大学農学
部教授・高橋高右衛門博士ならびに文部教官・新関稔氏
に対し心より深く感謝の意、を表するとともに,実験遂行
上御協力し、ただし、た文部技官・渡会寓治・飛渡正夫・功、
川ß~閣の 3 氏に心から謝意を表する次第である。
Il. 実験材料および方法
Eumelilotus亜属には 9種が含まれるが,本実験には
そのうち 4種 (M.alba, M. taurica, 1'v1. wolgicaおよ
2) Crops Research Division, ARS, USDA, Lincoln Nebr 3) Research Station, Canada Depertment of Agricu¥ture, Brandon, Manitoba.
l

2 北海道大学農学部邦文紀要第 9巻第 1号
Table 1. Number of seeds and seedlings obtained from crosses between
pairs of sp巳ciesin the subgenus EUlllelilotus
cross
M. taurica 70 xM. alba Cumino
N1. taurica 70 xM. alba Denta
M. taurica P. I. 314094xM. alha Cumino
NI. taurica 70 xM. wolgica 58-254
λ1. dentata 58-190 xM. taurゴca 70
N1. dentata 58-190 xM. taul・ica P. I. 314094
び M.dentata) 6系統を供試した。これらの系統のうち,
M. alha Cuminoおよび M.alba Dentaはともにリ1.
alba>くM.dentata)XM. albaの戻し交雑法により M.
dentataより‘lowcoumarin gene'を導入した育成品
種であるが他の系統はし、ずれも野生系統である。
これらの系統を用い 1971年および 1972年の両年冬季
に温室内で交配を行なった。用いた種および系統,交西日
花数,得られた種子数,自家受精した個体数および得ら
れた種間雑種目の個体数を示すと Table1のごとくで
ある。
得られた種間雑種 F1~'i\'、ずれも幼苗時にかなりの程
度の葉緑素欠乏を呈し,種間雑種であることが容易に判
定出来た。 いずれの種間雑種 Flも世代短縮の目的で長
日処理下で栽培管理され,順調に成育し開花登熟に達
した。
交配の方法および減数分裂の観察の方法は前報 (1973)
に従った。
III. 実験結果
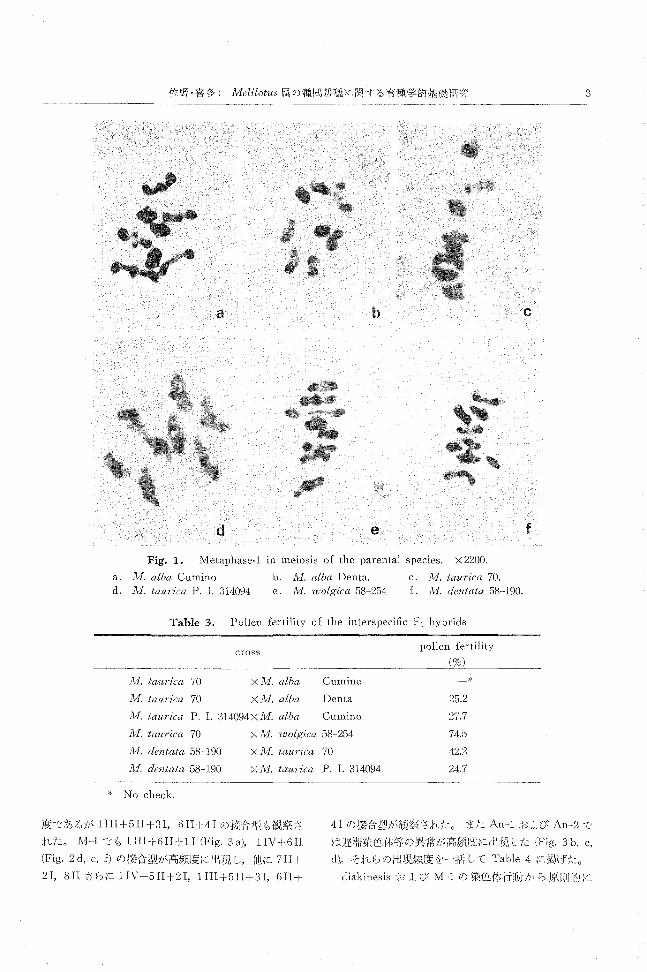
1. 親の種の細胞学的観察
各親の種の花粉稔率は Table2に示されたごとくい
ずれも極めて高かった。また花粉母細胞の減数分裂にお
ける M-1では規則的に 8IIを形成し (Fig.1), 他の時
期においても異常は観察されなかった。これらのことよ
り,本実験に供試した各親の種はし、ずれも細胞学的に安
定していると考えられる。
しかしながら, 1¥1. alba Cuminoおよび M.alba
Denta はともに M.dentataより‘lowcoumarin gene'
を導入された育成品種である。またこれら 2系統と M.
taurzcaとの交雑において,その種間雑種目はし、ずれも
開花登熱に達するが, 1¥よ albaの他の系統と M.taurica
との交雑では既に報告されたとおり (SMITH1954,
no. of no. of no. of seedlings
flowers seeds selfs hybrids
27 10 4 6
34 3 2 1
8 6 3 3
83 27 21 6
17 4 2 2
20 5 2 2
Table 2. Pollen fertility of the parental
species and strains
species and strain
M. alba Cumino
Denta
M. taurica 70
P. I. 314094
M.ωolgica 58-254
M. dentata 58-190
po llen fertili ty
(%)
99.3
97.7
98.4
98.1
99.2
98.7
KITA 1965)いずれも発芽後まもなく強度の葉緑素欠
乏のため枯死した。
著者らは予め M.alba Cuminoおよび M.alba
Dentaの2系統と M.albaの他の系統とが同じ染色体
構造をもつかどうかを検討するため,これら 2系統を含
む多くの M.albaの系統を用い,M. suaveolens, M.
officinalis, M. polonicaの3種との問で交雑実験を行
なった。詳しくは続報に譲るが,この交雑実験の結果よ
り染色体構造に関し系統間差異は存在しないと推定され
た。しかしながら厳密には, M. albaの系統間雑種を作
出しその染色体行動を検討する必要があると考える。
2. 種間雑種の細胞学的観察
各交雑組合せの種間雑種目の花粉稔率を一括して
Table 3に示した。 M.tauricaXM. wolgica F1は
74.5'70 とかなり高い稔性を示したが, f也の種間雑種目
はL、ずれも著しい花粉不稔を示した。
次に減数分裂の染色体行動の観察を行なったが1¥主
taurica x M. albaの3組合せの Flは¥,、ずれも各々 煩似
の異常を示した。すなわち, diakinesisではlIV十6II
(Fig. 2 b, c)の接合型が高頻度に出現し他に 1III十6II
十1I, 8IIおよび 7II十21が出現した。また極めて低頻
佐野・喜多 hleWotus慌の種関紙稔に関する育種さ罪的基礎研究
a b
d e
Fig. 1. M巴taphase“ inmeiosis of the parental species. X 2200.
a. 1vI. alba Cumino b. 1¥1I. alba Denta. c. lvI. taurica 70
d. 1vI. taurica P. 1. 314094 e. 1vI. wolgica 58-254 f. 1¥1I. dentata 58-190.
Table 3. Pollen fertility of the interspecific F1 hybricls
A1. taurica 70
λ/l. taurica 70
cross
X JIll. alba Cumino
×ル1.alba Denta
λ1. taurica P. 1. 314094XJvI. alba Cumino
1V1. taU1・ica70 X JIlI. wolgica 58-254
lVI. dentata 58-190
lvf. dentata 58-190
* No check
X 1V1. taurica 70
X JIl1. taurica P. 1. 314094
pollen fertility
25.2
27.7
74.5
42.2
24.7
*
3
C
f
度であるが 1III+5 TI 十:n, 611 十 41 の接合型もfI~祭さ
れた。 M-1でも l1II十6II十11(Fig. 3 a), 1 1V -ト6II
(Fig. 2 d, e, f)の接合型が高頻度iこ山現し,他に 7II十
21, 8 IIさらiこlIV十5II十21,1 III十5II十31,6 II十
41の銭合型が観察された。 また1¥11-1および1¥11-2で
は遅滞染色体等の異常が高頻度にU¥現した (Fig.3 b, c,
dlo それらの出現頻度を一括して Table4に掲げた。
diakil1esisおよび M-1の染色体行動から原則的に
4 北海道大学農学部邦文京己要 第 9巻第 1号
b
d e
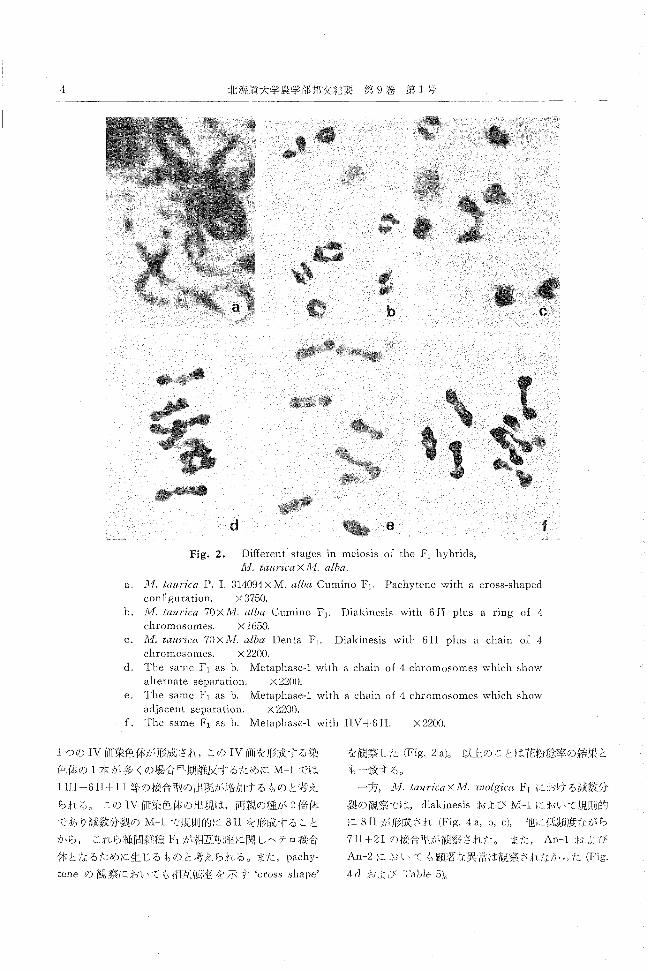
Fig. 2. Different stages in meiosis of the f<¥ hybrids, 11,1. tallrica X 1i1. alba
c
f
a. 1'vl. taurica P. 1. 314094xM. alba Cumino F)・ Pachytenewith a cross-shapecl
con五guration. X 3750
11. 1¥;1. taurica 70 X lvI. alba Cumino F 1. Diakinesis with fi 11 plus a ring 01 4
chromosomes. X 1650目
c. 1v1. taurica 70X1i1. alba Denta F)目 Diakinesiswith 6 II plus a chain of 4
chromosomes. X 2200.
<1. The same F1 as b‘ Metaphase-l with a chain of 4 chromosomes which show
alternate separation. X 2200.
e. The same F) as b. Metaphase-l with a chain of 4 chromosomes which show
acljacent separation. く2200.
f. The same F) as b. Metaphase-l with IIV十fi11. X 2200.
tつの IV仰i染色体が形成され,この IV耐nを形成する染 を観察した (Fig.2 a)。 以 tのことは花粉稔率の結果と
色体の 1木が多くの場合早期離反するために M-}では も一致する。
1 III十fiII十11等の媛合型のIt¥現が増加するものと考え 一方 11,1.tau円"caXJvl.wolgica F)における減数分
られる。 この IVfI而染色体の出現は,両親の種が 2倍休 裂の観察では, diakinesisおよび M-lにおいて規則的
であり減数分裂の M-lで規則的に 8IIを形成すること に 8IIが形成され (Fig.4 a, b, c), 他に{氏頻度ながら
から, ζれら種間雑種 F)が相互転践に関しヘテロ桜合 7II+21の抜合引が観察された。 また An-lおよび
休となるために生じるものと考えられる。主た, pachy回 An-2においても顕著な異J訟は観祭されなかった (Fig.
teneの観察におし、ても相互転肢を示す 'crossshape' 4 dおよび TabJe5)0

護
cross
A1. taun:c日
70 ×
JIlf. alba Cumino
λ1. taurica 70 ×
li1. alba Denta
1¥If. taurica
佐野・高多 lI/[el.:lotus属の種間雑税に関する153-穂学的基礎研究 5
a
C
Table 4.
stagc
b Fig. 3. Different stag巴 inmeiosis of
the F 1 hybrids, 1¥1[, Lauru:a X
み[.alba. X 2200
a. Af. taurica 70 Xル1.alba Cumino Fl・
Metaphase-1 showing 1 III十6II十11
con五guration.
b ル1. tau rica P. 1. 314094 X 2¥:[, alba
Cumino Fl. Anaphase-1 showing 7
9 separa tion.
c. The same Fl as a. Anaph乱回目1show-
ing a lagging chromosome.
d. The same Fl as a. Anaphase-2 show-
ing lagging chromosomes
Chromosome conngurations at cliakinesis ancl m巴taphase-1
ancl thcir clistribution in later stages in the Fl hybricls, 11レf
Laurica X JI. alba
frcquency of PMCs with
lIV+ 1III十 1 III十 1 III十8II 7II十 6II十 nor. abnor.
total 6II 5II十2I 6II+1I 5II十31 21 41
同一一 一一 一一
Diakinesis 89 10 4 20 10 つゐJ 135
Metaphase-1 50 7 110 2 14 26 b 214
Anaphase町1 73 77 150
Anaphase之 87 63 150
Dial王meslS 64 4 16 4 88
Metaphase同1 15 2 19 3 3 42
Anaphase-1 51 11 62
Anaphase悶2 29 10 39
Diakinesis 38 4 6 48
P. 1. 314094 Mεtaphase-1 26 1 40 1 7 9 3 87 × Anaphase-1 58 18 76 λ1. alba
Cumino Anaphase-2 39 17 56
6
a
C
北海道大学農学部邦文紀要第 9巻第 1号
b
品
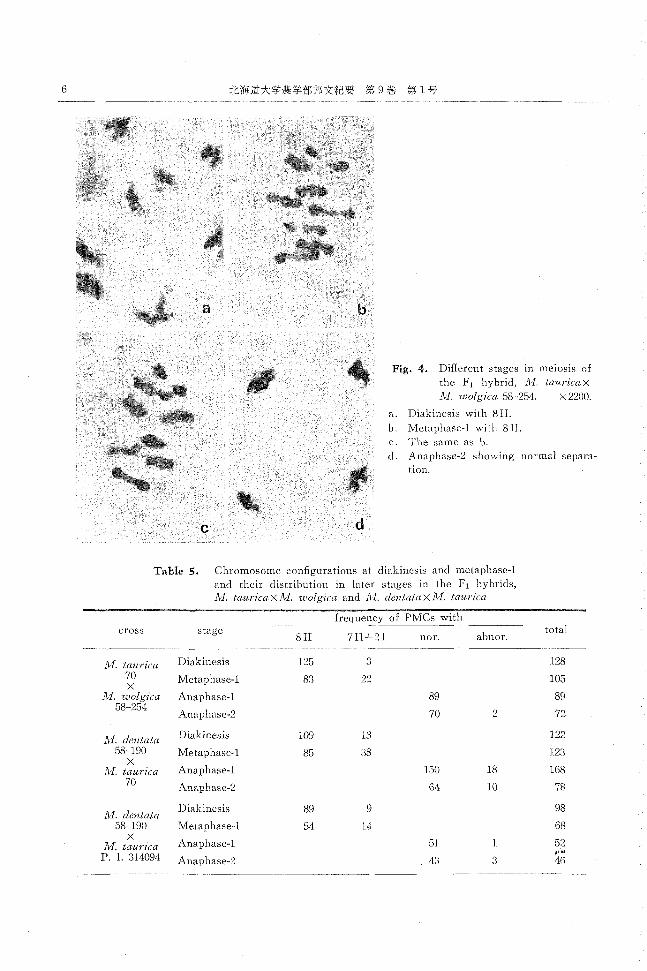
Fig. 4. Different stages in meiosis of
the F1 hybrid, lvI. tauricax
lvI. wolgica 58-254. x 2200.
a. Diakinesis with 8II.
b. Metaphase-1 with RIl.
c. The same as b.
d. Anaphase-2 showing normal separa-
tlOn
Table 5. Chromosome con五gurationsat diakinesis and metaphase-l
and their distribution in Jater stages in the F1 hybricls, Aよ ta川・icaXli1.'lι叫'gicaancl lvI. dentata X ivL taurica
frequency of P乱ICswith cross stage
abnor. total
8Il 7II ト21 nor.
lvI. taurica Diakinesis 125 3 128
70 Metaphase-1 ×
83 22 105
lvI. wο19u:a Anaphase-1 89 89 58-254
Anaphase-2 70 2 72
1',1. dentata Diakinesis 109 13 122
58-190 ×
Metaphase幽 85 38 123
kI. taU1ゴca Anaphase働1 150 18 168 70 Anaphase-2 64 10 78
λ1. dentata Diakinesis 89 9 98
58-190 Metaphase欄1 54 14 68 ×
Anaphase問lλ,1. taurica 51 52
P. 1. 314094 Anaphase-2 43 3 戸46対

佐野・喜多 Melilotl付属の種間雑種に~)する育穂学的基礎研究 7
8
c
さらに lvl.dentata X 1¥;[. tauricaの2組合せの Fl
では,花粉稔率I主極めて低かったにもかかわらず減数分
裂におけ』る染色体行動は概して正常であった。すなわ
ち, diakinesisおよび M-lでは高頻度で 8IIが出現し
(Fig. 5 a, b, c), An…1および An-2においても正常に分
離するものが多く観察された (Fig.5 dおよび Table5)。
これらのことより ,lvI. taω-icaxlvI. wolgica Flお
よび lVI.dentata X lvI. taurica F 1の染色体行動はとも
に概ね正常であると考えられる。
IV. 論 議
b
d
Fig. 5. Di妊erentstages in meiosis of
the FJ hybrids, M. dentataX 1¥1. taurica. X 2200
a目 1'v[.dentata 58-190xM. taurica 70 Fl
Diakinesis showing 8 II configuration.
b. The same F1 as a. Metaphase-1
showing 8 II con五guration.
c. l'vI. dentata 58-190xl¥1. taurica P. 1.
:114094 F J• M巴taphase-1 showing 8 II
configuration
d. The sam巴 F1as a. Anaphase-1 show伺
ing normal separation.
altissima, lvI. dentata,λ1. taurica, M. wolgica)およ
び GROUPC (M. officinalis)に分演した。その後,M.
officinalisおよび M.hirsutaはともに GROUPAに分
類されるのが妥当であることが報告された (WEBSTER
1955,喜多 1962)。
これより先, SMITH (1943)は GROUPA に含まれる
M. albaと GROUPB に含まれる 1¥1.dentataの種間
雑種 FJは著しい葉緑素欠乏を長し発芽後まもなく枯死
するが,接木によりその Fl植物を開花登熟まで育成す
ることに成功した。 さらに BRINGHURST(1951)は,
この種間雑種を供試して (M.alba>くM.dentata)>くM.
Eumelilotus illi.属における種間雑穏 F1には,幼商時に albaの減数分裂における pachyteneの観察より‘cross
J種々の程度の葉緑素欠乏が出現し,このことは Eumeli司 shape'の出現することを認め, 111. albaと 1'11.dentata
lotus亜属に含まれる種聞にIjJiJ!く隔離機構のうちの大き の種間には相互転践に由来する染色体構造差の存在する
な要因の 1っと考えられる。 ことを示唆した。
SMITH (1954)は Eumelilotus頂属の 8徳のrJJで広 木実験に供試した 111.alba Cuminoおよび M.alba
汎な交雑を行い,交~\jf和合性と種間雑種 F1 に出現する Denta はともに,前述したようにこの (M.albaxM.
3主総素欠乏の程度によりこれら 8種を GROUPA (M. dentata)XM. albaの後代より作出した‘lowcoumarin
alba, 1'vI. tolonica, lv[. suaveolens), GROUP B (1¥1. gene'てど M.albaに遵入した育成品種であり,他の M

8 北海道大学農学部邦文紀要第 9巻第 1号
albaの系統とは若干性質を異にしている。すなわち,M.
albaと M.tauricaの種間雑種目は著しい葉緑素欠乏
により発芽後まもなく枯死するが,ヨギ;実験より明らかな
ごとく M.alha Cuminoおよび λ1.alba Denta 2系
統と M.tau円;caとの交雑において, その縄問雑極 Fl
はし、ずれもかなりの程度の葉緑素欠乏を示すが開花主主熟
まで生育を続ける。このことは,M. alha Cuminoおよ
び M.alba Denta 2系統が M.dentataより遺伝子の
導入されたことに原因していると考えられる。また先に
述べたように,多少の疑問は残るが,M. alba Cumino
および M.alba Den taはともに M.albaの他の系統
と同ーの染色体構造をもっと考えられる。このことと木
実験より得られた M.taurica X M. alba F 1の結果を
考え合せると M.tauricaと M.albaの種間には 2つ
の非相同染体聞に起る相互転座に由来する染色体儲造差
が存在すると考えられる。さらに M.taurica X M.
wolgica Flおよび M.dentataXM. taurica Flの結
果より ,M. taurica, M.日 olgica,および M.dentata
の3積の聞には細胞学的差異はなし、と考えられる。以上
のことを総合すると, M. alhaと M.dentataの間に
は当然相互転座に由来する染色体構造差の存在すること
が推定されるが,このことは BRINGHURST(1951)の報
告に一致する。
しかしながら,九1.taurica, M. wolgicaおよび λf
dentataの 3績と M.albaの聞に存在する相互転座由
来の染色体構造差が同一起源のものかどうかに関しては
疑問が残る。すなわち,花粉稔率の結果より M.taurica
と M.wolgicaに存在する M.albaとの聞の染色体構
造差は同一起源であると考えられるが,M. dentataX
M. taurica F 1の花粉稔率が極めて低かったことより,
M. dentataと M.tauricaに存在する M.albaとの
聞の染色体構造差が切断点を異にする相互転~ーによるも
のすなわち同一起源ではない可能性のあることが示唆さ
れる。 しかしながら, この低稔性の原因が遺伝子支配
によることも当然考えられることより ,M. dentataの
染色体構造に関しては今後さらに検討する必要があると
考える。いずれにしても,M. taurica, M. wolgicaお
よび M.dentataの3種と M.albaとの聞には 2つの
非相同染色体聞における 1つの相互転座に由来する染色
体構造差が存在すると考えられる。
v.摘要
Melilotus属 (Sweetclover)の Eumelilotus亜属に
含まれる 4種 (M.alba, M. taurica, M. wolgicaおよ
び M.dentata) 6系統闘で交雑を行い,得られた種間雑
穣 Flに関し主として細胞学的な研究結栄を取縛め報告
した。結果を契約すると次のごとくである。
1. 111. dentataより‘lowcoumarin gene'の導入
された λ1.alha Cuminoおよび M.alha Den taの
2系統はともに M.tauricaと容易に交雑出来,それら
の積問雑種 Flはかなりの程度の葉緑素欠乏を呈するが
いずれも開花登熟まで)i児育した。
2. M‘ tauricaxM. alba Flの減数分裂における
diakinesisおよび M-1において, 1つの IVIi而染色体が
出現した。 このことより,これらの積間雑種 Flは相互
転座に関しヘテロ接合体であることが明らかとなった。
3. M. taurica X M.叩 olgicaFlおよび M.dentata
xM. tau門;caF1の減数分裂における染色体行動はし、ず
れも概ね正常であった。 したがって,11よ taurica,M.
wolgicaおよび M.dentataの 3種の聞には細胞学的
差異はなし、と考えられる。
4. 以上のことより ,M. taU/・ica,M. wolgicaおよ
び M.dentataの3積と M.alhaの穐間には相互転座
に由来する染色体構造差が存在すると考えられる。
参考文献
1) BRINGHUST, R. S. (1951): Genetic analysis of
chlorophyll c1eficiency in Melilotus albaXM.
dentata hybrids with some observations on
meiotic irregularities. Summaries of Doctoral
Dissertations司 Univ.Wis. 11: 96-97.
2) GORZ, H. .r., and W. K. SMITH (1961): Use of
diverse germ plasm in the improvement of
some forag巴 legumes. Germ PlaslIl Resouces.
149-159.
3) J ARANOWSKI, J. K. (1961): Semisterility in
the interspeci五chybrid, Melilotus polonicαxM. alba. Amer. J. Bot. 48: 28-35.
4)喜多富美治 (1962): Melilotus属の種間雑種に関
する育種学的基礎研究,第 I報種間雑種 Melilotus
alba x M. hirsutaの細胞遺伝. 北大邦文紀要,第
4巻,第 1号 67-74.
5) KIT A, F. (1965): Studies on the genus Melilotus
(Sweetclover) with special reference to inter-
relationships among species from a cytological
point of view. J. Fac. Agri. Hokkaido Univ.
54: 22-121.
6)喜多富美治・佐野芳雄 (1972): Melilotus属の種間
雑種に関する育種学的基礎研究,第 VII報種間雑
種 Melilotusofficinalis P. 1. 178985xM. alba var.
Cuminoの細胞遺伝. 北大農場報告,第 18号 1-6.

佐野・喜多 Melilotus属の種間雑種に関する育種学的基礎研究 9
7)佐野芳雄・喜多富美治 (1973): Melilotus属の種間 the species belonging to the genus Melilotus. The
雑種に関する育種学的基礎研究,第 VIII報 種 間 雑 results obtained are summarised as fullows:
種 MelilotusalhaXM. suaveolens, M. polonicax 1. Generally, the F1 hybrids between M. alba
/0.1. slluveolensおよび /0.1.hirsuta Xλ1. suαveolens and M. taurica cannot grow to flowering stage due
の細胞学的研究. 北大邦文紀要, 第 8巻, 第4号 to chlorophyll deficiency. However, using two low
411-419. coumarin varieties of M. alba, Cumino and Denta
8) SHASTRY, S. V. S目, W. K. SMITH, and D. C. which are d巴rivativesof the crosses ofω;[. albα×
COOPER (1960) : Chromosome di妊erentiationin M. dentata)XM. alba, the Fl hybrids from the
several species of Melilotus. Amer.]. Bot. 47: crosses of M. tauricaXM. alb叫 inthis experiment
61:3-621. showed moderate chlorophyll de五cientin the s何 dl・
9) SMITH, W. K. (1943): Propagation of chloro・ ingstage and were able to grow to maturity.
phyll-cieficient sweetclover hybrids as grafts. 2. In the interspeci五c hybrids, M. tauricaX
J Heredity 34: 135-140. M. alba, a ring or chain of 4 chromosomes was
10) (1948): Transfer from Melilotus observed at diakinesis and metaphase-1, which
dentata to M. alba of the genes for reduction indicated that these F1 hybrids were heterozygous
in coumarin content. Genetics, 33: 124-125. for a reciprocal translocation.
11) (1954): Viability of interspecific 3. In the two interspeci五CF1 hybrids, M. taurica
hybrids in Melilotus. Gen巴tics39: 266-279. xM. wolgica and M. dentataXM. taurica, chro・12) WEBSTER, G. T. (1955): Interspeci五chybrid- mosome behaviors in the course of meiosis were
ization of Meliloω.5 alba x M. officinalis using regular. These observations indicate that there is
embryo culture. Agron..T. 47・138-142. no structural difference of chromosomes among
three species, M. tau門ica,M. wolgica, and M.
Summary dentata・
Three interspeci五C F1 hydrids, fo.!lelilotus tallrir:a 4. Henceforce, it is considered cytologically that
x /0.1. alba, /0.1. tallrica X M.τf)olgica, and /0.1. d,仰 tata M. alba differs from the three species, M. taurica,
xl¥1. tallrica were studied in order to make clear M. wolgica, and M. dentata by one reciprocal
the picture of the cyto¥ogica¥ re¥ationships among translocation.




![[成果情報名]種子が少なく果皮が薄いレモン新品 …...[成果情報名]種子が少なく果皮が薄いレモン新品種「イエローベル」 [要約]レモン新品種「イエローベル」は、道谷系「ビラフランカ」の自然交雑実生から選](https://img.pdfslide.net/doc/110x75/5e63b29daf863c2ab405f894/oefcoeoecoeefff-oefcoeoecoeefffciffffi.jpg)









![は虫類 両生類 魚類nagorep.com/gairai.pdfハナガメ、スウィンホーキノボリトカゲ、 ハナガメ×[ニホンイシガメ・ミナミイシガメ・クサガメ]の交雑種](https://img.pdfslide.net/doc/110x75/5e6c9395dcabbc3ab4333673/ee-ce-ee-ffffffffoeff.jpg)



![症例報告 体表の自壊した腫瘤に対する Mohs ペーストの有用性 · ─ 53 ─ 広島県獣医学会雑誌 №29(2014) [症例2]雑種犬,17 歳,未去勢オス,体重16kg.](https://img.pdfslide.net/doc/110x75/60dd48b7300f075902666c17/c-eeec-mohs-fffoec.jpg)








