Embed Size (px)
Citation preview

Toxicology 132 (1999) 85–98
Review
Metallothionein in radiation exposure:its induction and protective role
Lu Cai a,b, Masahiko Satoh c, Chiharu Tohyama c, M. George Cherian a,*a Department of Pathology, The Uni6ersity of Western Ontario, London, Ontario, Canada N6A 5C1
b Institute of Radiation Medicine, Norman Bethune Uni6ersity of Medical Sciences, Changchun, People’s Republic of Chinac En6ironmental Health Sciences Di6ision, National Institute for En6ironmental Studies, Onogawa 16-2, Tsukuba,
Ibaraki 305-0053, Japan
Received 4 September 1998; accepted 20 November 1998
Abstract
Since its discovery about 40 years ago, there has been a wide interdisciplinary research interest in metallothionein(MT) on its physiological and toxicological aspects. Functionally, MT is involved not only in metal detoxification andhomeostasis, but also in scavenging free radicals during oxidative damage. Among over 4500 publications which canbe retrieved by Medline search, only about 50 reports have been published on the relationship of MT with ionizingand UV radiation. In this review, we have evaluated critically the published data on the induced synthesis of MT byradiation, and the potential functions of MT in radiation induced cell damage. MT mRNA expression or MTsynthesis was found to be induced by exposure of cells in vitro or tissues in vivo to ionizing or UV radiation. In mostof the studies in animals and tissue cultures, high doses of ionizing radiation were used to induce MT, and, therefore,it is difficult to extrapolate these results to low level of repeated exposures to radiation in humans. Induced synthesisof MT is considered as one of the mechanisms involved in the adaptive response to low dose radiation exposure. Thepresence of MT in normal cells may provide protective effects from radiation-induced genotoxicity and cytotoxicity.However, in tumor cells, the presence of MT can result in drug and radiation resistance as well. These effects aremodulated by other cellular factors, besides MT, such as antioxidants, and by the cell cycle stages in cell proliferationand differentiation. © 1999 Elsevier Science Ireland Ltd. All rights reserved.
Keywords: Metallothionein; Radiation protection; UV radiation; Ionizing radiation; Review
* Corresponding author. Tel.: +1-519-6612030; fax: +1-519-6613370; e-mail: [email protected].
0300-483X/99/$ - see front matter © 1999 Elsevier Science Ireland Ltd. All rights reserved.
PII: S 0300 -483X(98 )00150 -4

L. Cai et al. / Toxicology 132 (1999) 85–9886
1. Introduction
Metallothioneins (MT) are a family of low-molecular weight (about 6–7 kDa), cysteine-rich(30%) intracellular proteins with high affinity forboth essential (zinc, Zn and copper, Cu) andnon-essential (cadmium, Cd and mercury, Hg)metals. It has been 40 years since MT, a Cd- andZn-binding protein, was isolated first from horsekidney (Margoshes and Vallee, 1957). During thelast four decades, there have been several reportson the function and structure of MT, but thenumber of publications on MT and radiationexposure is few. However, radiation is used widelyfor cancer treatment and the role of MT inductionin radiation exposure is of great interest. In thisreview article, we have tried to evaluate criticallythe published data on MT and radiation.
There are four isoforms of MT: MT-I, -II, -IIIand -IV in the mammalian organs. Among them,MT-I and -II are the most predominant isoformsand have been well-documented. The two majorisoforms of MT, MT-I and -II, contain two dis-tinct adamantine-like metal–thiolate clusters, onewith four and another with three metal ions. Theisoforms, MT-I and -II are expressed ubiquitouslyin most mammalian organs, and regulated co-or-dinately, whereas the MT-III and -IV isoforms areexpressed specifically in the brain and insquamous epithelia, respectively (Chan andCherian, 1993; Cherian and Chan, 1993; Sato andBremner, 1993). MT-I and -II mRNA and proteinare inducible not only by metals, but also bymany non-metallic compounds including ethanol,alkylating agents, and physical or chemical oxida-tive stress conditions (Cherian and Chan, 1993;Sato and Bremner, 1993). In addition to its princi-pal role in detoxification of potentially toxicheavy metals, and in regulation of the homeosta-sis of essential trace metals, MT can also act as anantioxidant and a free radical scavenger (Cherianand Chan, 1993; Sato and Bremner, 1993;Cherian and Ferguson, 1997). The antioxidant orfree radical scavenging activity of MT has beencompared with reduced glutathione (Thornalleyand Vasak, 1985; Miura et al., 1997). The targeteddisruption of MT-I and -II genes in MT-null micemade them more sensitive to Cd and anticancer
drug cytotoxicity (Masters et al., 1994; Kondo etal., 1995). Therefore, induction of MT synthesismay be one of the important defense mechanismsagainst Cd toxicity and oxidative damage (Ko-ropatnick and Cherian, 1988). The cellular dam-age of radiation is mainly an oxidative damagedue to the formation of several types of oxygenfree radicals including hydroxyl and superoxideradicals by radiolysis of water in cells. Severalreports have shown both induced synthesis of MTand a protective role for MT against free radicalsduring radiation exposure. Since UV radiation-mediated cell injury may also be involved in oxy-gen radical generation (Black, 1987; Hanada etal., 1992) we describe both ionizing radiation andUV radiation in this review.
Radiotherapy is widely used in the treatment ofvarious types of cancer, and recent reports suggestthat the sensitivity of tumors to radiation maydepend on their MT levels (Cherian and Chan,1993; Kondo et al., 1995; Cherian and Ferguson,1997). Therefore, we have reviewed also the po-tential role of MT in treatment of cancer withradiation.
2. Induction of MT synthesis by radiation
2.1. Ionizing radiation
The induced synthesis of MT in different or-gans after whole body irradiation has been re-ported in rats (Shiraishi et al., 1983, 1986;Togami, 1987; Hayashi, 1988; Shiraishi et al.,1989) and in mice (Matsubara et al., 1986,1987a,b; Koropatnick et al., 1989). In certainstudies, both MT protein and mRNA contentswere found to be induced in a dose-dependentmanner in mice (Koropatnick et al., 1989). Allthese reports are summarized in Table 1.
Induction of MT synthesis or mRNA expres-sion varied in different organs after radiationexposure, and was unrelated to the radiosensitiv-ity of the organs but may depend on animalspecies. For instance, MT mRNA expression wasincreased markedly in the liver, kidney and thy-mus, but not in other tissues (the brain, spleen,lung, heart and testis) of the rat irradiated with

L. Cai et al. / Toxicology 132 (1999) 85–98 87
Table 1Induction of MT proteins or mRNA by ionizing radiation (IR) or UV radiation (UV)
Tissues/cells Proteins/mRNA Dose range ReferencesAnimal speciesTreatments
In 6i6oRat, mouseINRa Liver MT proteins 1–80 Gy Shiraishi et al., 1983, 1986; Togami,
1987; Matsubara et al., 1987a,b;Koropatnick et al., 1989
MT mRNA 2–80 Gy Hayashi, 1988; Shiraishi et al.,1989; Koropatnick et al., 1989
Kidney MT proteinsINR 15–30 GyRat, mouse Shiraishi et al., 1986MT mRNA 2 or 20 Gy Hayashi, 1988; Shiraishi et al., 1989
Thymus MT mRNARat 2 or 20 GyINR Shiraishi et al., 1989MouseINR T-tumorb MT protein 5–20 Gy Shibuya et al., 1995, 1997
Liver MT protein 4.7 T Satoh et al., 1996SMFc MouseSkin cells MT protein 2 or 3 MEDdHuman Jasani et al., 1993; Anstey et al., 1996UV
In 6itroV79 cells MT proteinsHamster 0.3 Gy×60INR Osmak and Uzarevic, 1991
HumanINR Hela cells MT proteins 0.5 Gy×30 Osmak, 1993SSK cells MT proteinsMouse 6 Gy×15INR Eichholtz-Wirth et al., 1993CNS MT protein, mRNAINR 30 GyHuman Hammond et al., 1997S49 cells MT mRNA 5.9–23 J/m2UV Lieberman et al., 1983
HamsterUV V79 cells MT mRNA 1–17 J/m2 Fornace et al., 1988a,bFibroblasts MT mRNA 20 and 55 J/m2Human Rimoldi et al., 1992UV
a INR, ionizing radiation.b T-tumor, transplanted tumor.c SMF, static magnetic field.d MED, minimal erythema dose.
2.0 and 20.0 Gy X-irradiation (Shiraishi et al.,1983, 1986, 1989). In mice, MT protein content ormRNA expression was increased only in the liverafter whole body exposure to g-irradiation, butnot in other tissues (Matsubara et al., 1986,1987a,b; Koropatnick et al., 1989). The kinetics ofincreased MT protein or mRNA expression werealso dependent on the species. In rats the MTproteins reached maximal levels at 72 h after 20Gy irradiation both in the liver and kidney, whilemRNA reached a plateau at 9 h after irradiation,and these changes were not dose-dependent (Shi-raishi et al., 1986, 1989). In mice, the maximallevels of both MT protein and mRNA were de-tected at 9 h after irradiation (Koropatnick et al.,1989). Species differences in the induction of MTsynthesis in certain organs were not specific forradiation exposure but has been reported forother MT inducers (Choudhuri et al., 1993; Henryet al., 1994).
Because reports on radiation-induced MT syn-thesis in cultured cells are limited in terms ofidentical experimental conditions, the results ob-tained so far provide only fragmentary informa-tion, and are not always consistent. Singleexposure of rodent cell lines (Chinese hamsterV79 lung fibroblasts, B16F1 mouse melanomaand NIH 3T3 mouse embryonic cells), or humancells (IGR human melanoma cells and HL-60human promyelocytic leukaemia cells) to 2–10Gy X-rays did not increase either MT proteincontent or mRNA expression (Fornace et al.,1988b; Koropatnick et al., 1989). However, withmultiple doses of g-rays (0.3 Gy/day, 5 days/weekfor 12 weeks) MT protein synthesis was enhancedin V 79 cells (Osmak and Uzarevic 1991; Osmak1993). When mouse fibrosarcoma cells (SSK) wereirradiated with fractionated doses of g-rays (6 Gyper exposure, total dose of 90 Gy at a dose rate of1.2 Gy/min) a high level of MT protein was found

L. Cai et al. / Toxicology 132 (1999) 85–9888
(Eichholtz-Wirth et al., 1993) (Table 1). The re-sults suggested that multiple or fractionated mod-erate radiation doses could be more effective ininducing MT synthesis in cultured cells than asingle large dose of radiation.
Unlike MT-I or -II, the MT-III isoform wasnot induced by ionizing radiation. In primarycultured cells of the central nervous system (CNS)from human, after exposure to ionizing radiation,there was a marked induction of total MTproteins with enhanced MT-I mRNA expressionbut without any change in MT-III mRNA expres-sion (Hammond et al., 1997).
2.2. UV radiation
UV radiation seems to be more effective thanionizing radiation in inducing MT protein contentor mRNA expression (Table 1). Lieberman et al.(1983) presented the first evidence for UV-activa-tion of MT mRNA expression in S49 cells byDNA demethylation. This was confirmed in laterstudies using Chinese hamster V79 lung fibroblastcells, human HeLa cells and primary human skinfibroblast cells (Fornace et al., 1988a,b; Stein etal., 1989; Rimoldi et al., 1992; Jasani et al., 1993).Furthermore, MT protein synthesis has been re-ported in human tissues which were exposed toUV radiation (Jasani et al., 1993; Anstey et al.,1996; Nishimura et al., 1997).
2.3. Mechanisms for radiation-induced MTprotein synthesis and mRNA expression
The mechanisms involved in induction of MTsynthesis by X-, g- and UV-radiation are verysimilar in the published work, and it is difficult todiscuss them separately. Therefore, we have dis-cuss them together in this section. Radiation-in-duced MT synthesis was not mediated either bymetal redistribution in organs or release of gluco-corticoids from radiation-damaged tissues (Shi-raishi et al., 1986; Koropatnick et al., 1989).Increase in MT synthesis after radiation exposuremay be due to the interaction of different tissuefactors and may involve indirect mechanisms. Al-though the results from a group of mice exposedto radiation showed a marked MT induction, the
individual MT-I mRNA accumulation in the livershowed a large variation among mice, suggestingthat the induction may not be a direct effect ofradiation exposure (Koropatnick et al., 1989). Inthe same experiment, radiation did not induceMT-I mRNA accumulation in cultured humanand rodent cells (Koropatnick et al., 1989). Induc-tion of MT synthesis after radiation exposure maybe mediated by the following indirect pathways.
2.3.1. Induction of MT protein synthesis ormRNA expression by oxidati6e stress
Both MT protein content and mRNA expres-sion were induced after exposure of rats or miceto chemicals which can produce oxygen free radi-cals (Hidalgo et al., 1988; Bauman et al., 1991;Sato and Bremner, 1993). Thus, the formation oflipid peroxides and oxidative cell damage may beone of the mechanisms for radiation-induced MTprotein synthesis (Hidalgo et al., 1988). OxidativeDNA damage was considered as another interme-diate step in inducing MT mRNA expression byUV radiation (Fornace et al., 1988b; Stein et al.,1989). However, the inability of cultured cells invitro to induce MT protein content or mRNAexpression after a single large dose of ionizingradiation may be due to high cytotoxic effects(Stein et al., 1989).
2.3.2. Induction of MT protein synthesis andmRNA expression by cytokines
The production of cytokines such as inter-leukin-1 (IL-1), IL-6, IL-8, tumor necrosis factor(TNF), and interferon (IFN) is one of the re-sponses of radiation exposure (Neta and Oku-nieff, 1996). The induction of MT synthesis bycytokines has been widely documented both invivo and in vitro (Sato et al., 1993). This mayexplain the reason for radiation-induced MTprotein synthesis or mRNA expression in vivo,but not in vitro. In vivo radiation exposure maykill the most radiosensitive cells, but the radiore-sistant cells may produce cytokines resulting ininduction of MT. In vitro, at high doses of radia-tion exposure, the cellular metabolic activities incultured cells may be inhibited or the cells may bekilled. Therefore, multiple or fractionated moder-ate radiation exposure may be more effective in

L. Cai et al. / Toxicology 132 (1999) 85–98 89
production of free radicals and cytokines to in-duce MT synthesis. In a recent study, culturedCNS cells, which are relatively radioresistant,were used to investigate MT induction by ionizingradiation. The results showed that 30 Gy g-raysinduced marked MT protein synthesis andmRNA expression without showing cytotoxic ef-fects, while 60 Gy g-rays did not induce MTprotein and mRNA expression due to cytotoxiceffect (Hammond et al., 1997). In addition,Kramer et al. (1993) demonstrated that UV irra-diated cells could release into the culture mediumvarious factors that can induce the transcriptionof several UV-induced genes including MT IIA,when added to non-irradiated cultured cells.These results show an indirect mechanism of MTsynthesis after exposure to UV radiation.
2.3.3. Induction of MT protein synthesis andmRNA expression by signal transduction
Radiation is able to initiate a cascade of signaltransduction factors to stimulate gene expression.One of the examples is the changes in intracellularcalcium which plays an important role in thecellular response to ionizing radiation (Hallahanet al., 1994), and also in the induction of MTsynthesis (Xiong et al., 1992; Shiraishi andWaalkes, 1994). In addition, protein kinase C(PKC) plays a role in the transcriptional regula-tion of MT synthesis. Heme-hemopexin has beenknown to induce MT synthesis through a mecha-nism associated with PKC (Ren and Smith, 1995).Moreover, a PKC activator such as phorbol estercan induce MT gene expression through the AP-1activation mechanism while PKC inhibitors suchas H7 and chelerythrine can inhibit Cd-inducedMT synthesis (Yu et al., 1997), suggesting a rolefor PKC in the transcriptional regulation of MTsynthesis. Direct activation of PKC by UV radia-tion can occur in a variety of cell types withinseconds to minutes after exposure (Haimovitz-Friedman et al., 1996), and UV-induced MTmRNA expression may also depend on the PKCpathway (Hansen et al., 1997). Therefore, theradiation induced MT protein synthesis and geneexpression may depend on the cell types where thesignal transduction response such as the PKCpathway responds to radiation exposure.
3. Protective role of MT from radiation-inducedinjury
The rationale for a protective role of MT in thelethal effects of ionizing radiation and UV-radia-tion was developed based on the following obser-vations: (a) sulfhydryl agents, such as cysteine,cysteamine and glutathione, can protect mam-malian cells from radiation-induced DNA damageand cell death, and MT with a high cysteinecontent (approximately 30%) can also providesimilar protection (Cai et al., 1996); (b) ionizingradiation causes about 10% of the DNA damagedue to direct energy absorption and subsequentcleavage of C�H bonds from short-lived carbonradicals which can react with oxygen to formperoxides and oxygen radicals. With free radicalattack the sulfhydryl groups in MT could bereleased from metal binding to restore the originalDNA structure in radiation-induced damage(Greenstock et al., 1987); and (c) hydroxyl radi-cals contribute to about 90% of low-linear energytransfer-radiation-induced DNA damage (Hall,1994), and these free radicals could be inactivatedby Zn-MT.
3.1. Radioprotecti6e effect of pre-induced MT
3.1.1. In 6i6o studiesAs early as in 1982, Matsubara et al. (1982)
reported the protective effects of Zn pretreatmenton radiation damage. They observed that injec-tion of Zn a few days prior to irradiation reducedthe mortality in mice and speculated that it wasdue to the induction of MT synthesis. Subsequentexperiments confirmed that preinduction of MTproteins could reduce the damage caused by ioniz-ing radiation (Matsubara et al., 1986, 1987a,b,1988; Matsubara, 1988; Satoh et al., 1989; Satoet al., 1990; Koterov et al., 1993). A number ofchelates of essential metals can also provide radio-protection (Sorenson et al., 1995). In order todetermine a direct protective effect of MT inradiation injury, other MT inducers includingtoxic metals, stress and hypothermia were testedand were also shown to protect from radiationeffects, suggesting a potential role for MT inradio-protective effects (Table 2).

L. Cai et al. / Toxicology 132 (1999) 85–9890
Table 2Radioprotection by pretreatment with MT inducers
Treatments ReferencesEndpoints for protection
In 6i6oMortalityZn Matsubara et al., 1986, 1987b
Matsubara et al., 1987a, 1988; Matsubara, 1988Body weight lossCataract Kobayashi et al., 1989Skeletal malformationCd Michel and Fritz-Niggli, 1986; Ota et al., 1996
Matsubara et al., 1987b; Koterov et al., 1993MortalityGracheva et al., 1989Cytotoxicity of bone marrow cellsMatsubara et al., 1987bLeukocyte reduction
Skin cell damage Hanada et al., 1991Leukocyte reductionBismuth Satoh et al., 1989; Miura et al., 1998
Kagimoto et al., 1991Thymoma inductionSatoh et al., 1989; Kagimoto et al., 1991; Miura et al., 1998MortalityKagimoto et al., 1991; Miura et al., 1998Cytotoxicity of bone marrow
MortalityManganese Matsubara et al., 1987a,b, 1988; Matsubara 1988; Ogata and Izoumo1990
Magnesium Mortality Ogata and Izoumo, 1990Sato et al., 1990Skin cell damageAAa
Skin cell damageHypothermia Ota et al., 1996Hanada et al., 1995Skin cell damage1,25(OH)2D3b
Cytokines Neta and Okunieff, 1996
In 6itroBakka et al., 1982Human epithelial cellsCd
WHFIB cells, V79 cells Greenstock et al., 1987HeLa cells Renan and Dowman, 1989
Hanada et al., 1991, 1992MGH-U1 cellsV79 cell Dudek et al., 1993NB1RGB cellsZn Kobayashi et al., 1994
a AA, Aloe arborescens.b 1,25(OH)2D3, 1,25-dihydroxyvitamin D3.
3.1.2. In 6itro studiesBakka et al. (1982) demonstrated a radioprotec-
tive effect in two different cell lines (a humanepithelial line derived from normal skin and amouse fibroblast line) with high contents of MT,after the addition of Cd salts (Table 3). It is alsoreported that tumor cells exposed to MT inducingagents acquired radioresistance (Renan and Dow-man, 1989). Radiation-induced chromosomalaberrations were prevented in rabbit lymphocytes,not only when MT was induced in vitro by addi-tion of Zn to the cell culture medium, but alsowhen MT was induced in the cells by injection ofZn salts to rabbits before the cells were collected(Cai and Cherian, 1996).
In clones of V79 Chinese hamster lung fibrob-lasts, mouse keratinocytes and human cells the
MT level was correlated with the increased cellu-lar resistance to UVA and UVB radiation(Hanada et al., 1991; Dudek et al., 1993).Kobayashi et al. (1994) demonstrated that amongfibroblasts from normal adults (HS-K) andneonatal (NB1RGB) human skins, and kerati-nocytes either from human skin (SV40-HSK) orfrom new-born Balb/c mouse skin (Pam 212),NB1RGB cells contained 4–30 times more MTthan other cells. The rate of [3H]thymidine incor-poration into NB1RGB cells was also higher thanother cells after exposure to UVB radiation. Fur-thermore, NB1RGB cells in which the MT con-tent was elevated by dexamethasone (1 mM) or Zn(7 mg/ml) treatment were more resistant to UVBirradiation than untreated cells. These results sug-gested that MT levels in cells may play an impor-
L. Cai et al. / Toxicology 132 (1999) 85–98 91
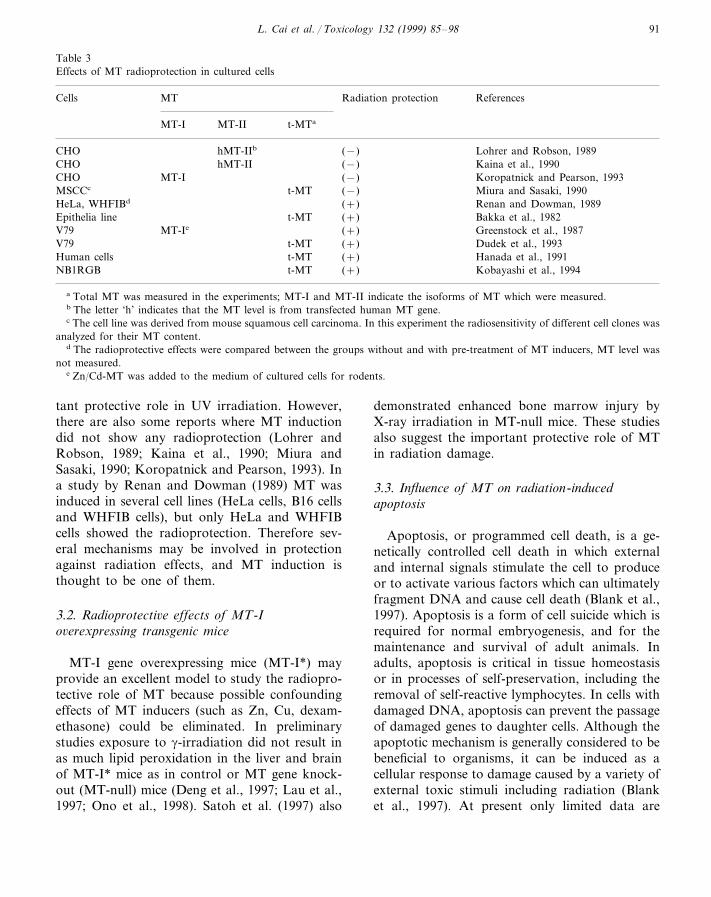
Table 3Effects of MT radioprotection in cultured cells
Cells Radiation protectionMT References
MT-II t-MTaMT-I
CHO hMT-IIb (−) Lohrer and Robson, 1989hMT-IICHO (−) Kaina et al., 1990
MT-I (−)CHO Koropatnick and Pearson, 1993MSCCc t-MT (−) Miura and Sasaki, 1990HeLa, WHFIBd (+) Renan and Dowman, 1989
t-MT (+)Epithelia line Bakka et al., 1982MT-IeV79 (+) Greenstock et al., 1987
V79 t-MT (+) Dudek et al., 1993t-MT (+)Human cells Hanada et al., 1991t-MTNB1RGB (+) Kobayashi et al., 1994
a Total MT was measured in the experiments; MT-I and MT-II indicate the isoforms of MT which were measured.b The letter ‘h’ indicates that the MT level is from transfected human MT gene.c The cell line was derived from mouse squamous cell carcinoma. In this experiment the radiosensitivity of different cell clones was
analyzed for their MT content.d The radioprotective effects were compared between the groups without and with pre-treatment of MT inducers, MT level was
not measured.e Zn/Cd-MT was added to the medium of cultured cells for rodents.
tant protective role in UV irradiation. However,there are also some reports where MT inductiondid not show any radioprotection (Lohrer andRobson, 1989; Kaina et al., 1990; Miura andSasaki, 1990; Koropatnick and Pearson, 1993). Ina study by Renan and Dowman (1989) MT wasinduced in several cell lines (HeLa cells, B16 cellsand WHFIB cells), but only HeLa and WHFIBcells showed the radioprotection. Therefore sev-eral mechanisms may be involved in protectionagainst radiation effects, and MT induction isthought to be one of them.
3.2. Radioprotecti6e effects of MT-Io6erexpressing transgenic mice
MT-I gene overexpressing mice (MT-I*) mayprovide an excellent model to study the radiopro-tective role of MT because possible confoundingeffects of MT inducers (such as Zn, Cu, dexam-ethasone) could be eliminated. In preliminarystudies exposure to g-irradiation did not result inas much lipid peroxidation in the liver and brainof MT-I* mice as in control or MT gene knock-out (MT-null) mice (Deng et al., 1997; Lau et al.,1997; Ono et al., 1998). Satoh et al. (1997) also
demonstrated enhanced bone marrow injury byX-ray irradiation in MT-null mice. These studiesalso suggest the important protective role of MTin radiation damage.
3.3. Influence of MT on radiation-inducedapoptosis
Apoptosis, or programmed cell death, is a ge-netically controlled cell death in which externaland internal signals stimulate the cell to produceor to activate various factors which can ultimatelyfragment DNA and cause cell death (Blank et al.,1997). Apoptosis is a form of cell suicide which isrequired for normal embryogenesis, and for themaintenance and survival of adult animals. Inadults, apoptosis is critical in tissue homeostasisor in processes of self-preservation, including theremoval of self-reactive lymphocytes. In cells withdamaged DNA, apoptosis can prevent the passageof damaged genes to daughter cells. Although theapoptotic mechanism is generally considered to bebeneficial to organisms, it can be induced as acellular response to damage caused by a variety ofexternal toxic stimuli including radiation (Blanket al., 1997). At present only limited data are

L. Cai et al. / Toxicology 132 (1999) 85–9892
available on influence of MT on radiation-in-duced apoptosis. In MT-I* and MT-null mousemodels, a high incidence of apoptosis in the thy-mus of MT-null mice was found in response towhole-body irradiation, as compared to MT-I*and control mice (Deng et al., 1997). This wasconfirmed by a recent study where pre-inductionof MT by Cd or b-thujaplian, a non-metal MTinducer, provided a marked protection from UV-induced apoptosis of rat keratinocytes both invivo and in vitro (Baba et al., 1998).
The incidence of apoptotic cell death in thetestis of MT-null mice was also higher than thatof MT-I* mice when they were exposed to 2–5Gy radiation (Cai and Cherian, 1997). When theradiation dose was increased from 5 to 10 or 20Gy, MT-I* mice showed a higher incidence oftesticular apoptotic cell death that was mainlydistributed in the epithelium where meiosis hadoccurred. These results suggest that MT may actin different ways to modulate radiation-inducedapoptosis. It could act as an antioxidant to reducethe formation of reactive oxygen radicals and theoxidative damage to DNA resulting in a lowincidence of apoptosis. In addition, MT may acti-vate certain signals to increase apoptosis in thepre-meiotic cells by preventing the passage ofDNA-damaged cells to S-phase in the cell cycle.Further experiments are needed to understand therole of MT on modulation of radiation-inducedapoptosis.
3.4. Influencing factors on MT radioprotection
Although a number of in vivo and in vitrostudies showed the protective role of MT induc-tion in tissues or cells against ionizing or UVradiation-induced damage (Table 2), there are fewreports which did not show any increased radiore-sistance in cells containing high MT. These appar-ent inconsistencies may be due to a number offactors.
In most experimental animals such as rodents,radiation effects on different organs can be iden-tified depending on the magnitude of the irradia-tion doses. For example, cerebrovascular effectsoccur at very high doses, while gastrointestinaleffects and hematopoietic effects occur at interme-
diate doses (5–12 Gy) and low doses (2.5–5 Gy),respectively, for rodents. The lethal dose (LD50)of radiation for humans is very similar to the dosethat causes hematopoietic effects (Hall, 1994). Instudies in vivo (Matsubara et al., 1986, 1987a,b),induction of MT synthesis by various inducerscould reduce the lethality in rats after exposure toa lethal dose (6–8 Gy), suggesting that the radio-protective effects of MT was mainly due to pro-tection of the hematopoietic system (Kagimoto etal., 1991). The induction of MT in bone marrowby compounds such as bismuth nitrate or subni-trate can protect mice from radiation effects (Sa-toh et al., 1989; Miura et al., 1998). Therefore, thecorrelation of MT induction in the liver afterinjection of Cd with the survival rate of radiationmay not be significant at different time intervals,while the correlation of Cd induced MT in bonemarrow with the rate of survival may be signifi-cant (r=0.96) (Koterov et al., 1993). Thus thetissue-specificity of different MT inducers shouldbe considered to determine their radioprotectiveeffect. For example, although there was a 6-foldincrease in MT in mouse liver, the pretreatmentwith manganese chloride did not protect againstacute radiation injury to skin and intestinal cryp-tic cells because there was no increase in MTcontent in the skin and intestine (Murata et al.,1995).
The subcellular localization of MT in cells maybe another factor to determine whether MT canprotect from radiation cytotoxicity or genotoxic-ity. The NIH3T3 cell line transfected with themouse-MT-I gene showed elevated cytoplasmicexpression of MT and these cells had a highresistance to the cytotoxic effects of tert-butylhydroperoxide but not to the DNA-damagingeffects (Schwarz et al., 1994). In contrast, a cloneof V79 cells with a high localization of MT in thenucleus was resistant to H2O2-induced DNA dam-age (Chubatsu and Meneghini, 1993). In rat ker-atinocytes treatment with Cd increased bothmRNA expression and cytoplasmic MT levels,which showed protection from subsequent UV-in-duced cell damage (Hanada et al., 1991, 1998).The reactive oxygen species generated by UVradiation are thought to be responsible for DNAdamage in skin cells. In keratinocytes UV radia-

L. Cai et al. / Toxicology 132 (1999) 85–98 93
tion can cause a shift in Cd-induced MT fromthe cytoplasm into the nucleus within 12 h ofexposure to UV radiation (Hanada et al., 1998).In these cells, MT may provide a protective ef-fect at an early stage from UV cytotoxic effect,and at a late stage from UV genotoxic effect.Thus the cytoplamic and nuclear localization ofMT in different types of cells may be anotherimportant factor to be considered in the protec-tive effect of MT from radiation injury. Thismay explain some of the differences in the pro-tective effect of MT in radiation-induced celldamage in different cell types (summarized inTable 3).
Another factor to be considered in cellular ra-dioprotection is the glutathione (GSH) level.Hepatic MT concentrations are high in the fetaland neonatal livers of mammals (Cherian, 1993).While the GSH concentration is low in fetalliver, and the maximal GSH synthesis occursonly after the first 2 weeks of postnatal lifewhen the MT level is low (Wirth andThorgeirsson, 1978). Thus the role of endoge-nous MT or GSH as an antioxidant or as a freeradical scavenger in certain cell lines may de-pend on the cell differentiation or the develop-mental stage of the tissues. When fibroblastsfrom adult (HS-K) and neonatal (NB1RGB) hu-man skin were exposed to UV radiation,[3H]TdR incorporation into the cells was inhib-ited in a dose-dependent manner (Kobayashi etal., 1994). A high level of endogenous GSH waspresent in HS-K cells while NB1RGB cells con-tained high MT levels but these cells were simi-lar in sensitivity to UV irradiation. Whenendogenous GSH in both cells were reduced toless than 10% of normal levels by buthioninesulfoximine (BSO), the inhibition curves of[3H]TdR incorporation by UV radiation were af-fected by the GSH depletion only in the HS-Kcells, but not in NB1RGB cells. When boththese cells were treated with MT inducer theinhibition of [3H]TdR incorporation by UV ra-diation was modified, suggesting the importantprotective role of MT in NB1RGB cells, andalso the protective role of MT in HS-K cells.These results suggest that the sensitivity of skincells to UVB irradiation was not always associ-
ated with their endogenous GSH levels but MTmay also play a major role in protection againstUVB irradiation (Kobayashi et al., 1994). In an-other study using human lung carcinoma cells(Biaglow et al., 1983) GSH depletion by BSOresulted in some radiosensitization. However, ifBSO pretreated cells were exposed to diethyl-maleate (DEM) a few minutes prior to X-rayirradiation much more sensitization occurred.Because DEM can bind with all free sulfhydrylgroups, thiols other than GSH in the cells mayalso play a role in radiation-induced damage.Liebmann et al. (1995) showed that enhancedGSH peroxidase expression protected cells fromhydroperoxides, but not from radiation. There-fore, in certain cells such as NB1RGB, MT mayplay an important role in protection against ra-diation damage, and in other cells such as HS-K, GSH may be involved in radioprotection(Michel and Balla, 1987; Astor et al., 1988;Mitchell et al., 1988). In the experiments whereMT induction did not provide the radioresis-tance, the amount of endogenous GSH was notdetermined (Koropatnick et al., 1989; Lohrerand Robson, 1989; Kaina et al., 1990; Miuraand Sasaki, 1990; Koropatnick and Pearson,1993). Therefore, the possibility that reactiveoxygen species are co-ordinately scavenged byboth endogenous GSH and MT should be inves-tigated in further studies.
Cytokines which can induce MT protein syn-thesis can also protect against radiation damage,especially in the immunological and hematopoi-etic systems (Neta and Okunieff, 1996). In cer-tain studies, where animals were exposed tovarious stress conditions to induce MT synthe-sis, a role for cytokines in radioprotection can-not be ruled out, especially in radiation-inducedbone marrow cytotoxicity (Matsubara et al.,1987b, 1988; Matsubara, 1988). Injection of Cdinto recipient mice leads to survival of trans-planted exogenous bone marrow and to thestimulation of colony formation after irradia-tion, suggesting the importance of microenviron-ments derived from Cd treatment, possibly theinvolvement of cytokines in addition to MT, inthe radioprotection (Izotova et al., 1992).

L. Cai et al. / Toxicology 132 (1999) 85–9894
3.5. MT in tumors and radiation treatment
Earlier studies reported that certain tumor cellshave increased content of MT and show resis-tance against anticancer drugs and radiation(Kelley et al., 1988; Renan and Dowman, 1989;Miura and Sasaki, 1990; Cherian, 1993; Cherianand Chan, 1993; Eichholtz-Wirth et al., 1993;Kondo et al., 1995). Therefore, altering MT levelsin tumors may modulate the resistance to anti-cancer drugs and radiation. For example, in tu-mor-bearing mice when the tumor MT level wasincreased by injection of Zn, the antitumor activ-ity of X-irradiation was decreased (Shibuya et al.,1997). The increased resistance of cultured tumorcells to g-radiation was shown by pretreatmentwith MT inducing agents (Renan and Dowman,1989). Exposure to g-radiation itself can induceMT synthesis in certain tumor cells, resulting inresistance to cisplatin, and probably to subse-quent radiation treatment (Eichholtz-Wirth et al.,1993; Shibuya et al., 1995). Thus a high expres-sion of MT can be considered as one of thefactors in radiation resistance of certain tumors.
Attempts have been made to reduce MT induc-tion in tumor cells during radiation and cisplatintreatment with propargyl glycine (PPG), an irre-versible inhibitor of the enzyme cystathionase(Satoh et al., 1993, 1994). In hepatocytes cysteinefor MT synthesis can be generated from methion-ine by a cystathionase pathway and this can beinhibited by PPG both in vivo and in vitro. Whentumor-bearing mice were injected with PPG, thecontent of MT in the tumor cells was decreased,and both cisplatin and radiation treatments wereeffective in reducing tumor weight (Satoh et al.,1993, 1994; Shibuya et al., 1997). Thus the antitu-mor activity of X-radiation may be affected bychanges in MT content in certain tumors.
In other studies certain chemicals such as bis-muth salts, which do not induce MT synthesis intumor cells but can induce in kidney and bonemarrow, have been used to protect these organsduring radiation treatment without compromisingthe antitumor activity (Satoh et al., 1989). Inaddition, bismuth salts can also depress thethymic lymphomas caused by X-irradiation(Kagimoto et al., 1991). These results suggest
that bismuth salts can prevent both acute toxiceffects and tumorigenicity of ionizing radiationby inducing MT synthesis only in target tissuessuch as bone marrow (Satoh et al., 1989; Kagi-moto et al., 1991; Miura et al., 1998).
Thus bismuth salts and PPG can be consideredas promising adjuncts in cancer radiation treat-ment to overcome the radio-resistance of tumors,and also to protect the sensitive organs fromacute radiation toxicity.
4. Possible role of MT in low-doseradiation-induced adaptive response
Pre-exposure of human lymphocytes or mam-malian tissues (Cai and Jiang, 1995; Cai andWang, 1995) to low-dose ionizing radiation(LDR) can render these cells or tissues moreresistant to DNA or chromosomal damage by asubsequent high-dose radiation (HDR), and thisresistance is known as an adaptive response.These cells also become cross-resistant to chemi-cal mutagens (bleomycin, mitomycin C (MMC)and cisplatin) (Cai and Jiang, 1995; Cai andWang, 1995). In addition, pre-treatment with lowconcentrations of chemical mutagens such asbleomycin, MMC, H2O2, actinomycin D, MMSand MNNG (Cai and Jiang, 1995; Cai andWang, 1995) could also induce an adaptive re-sponse to subsequent HDR exposure. The resultssuggested that there may be certain commonmechanisms involved in the induction of theadaptive response by low doses of ionizing radia-tion and chemical mutagens. For example, stimu-lation by oxidative radical scavengers orantioxidants after exposure to LDR or other mu-tagens has been documented, and it has beenshown that the protein synthesis is required forthis adaptive response (Cai and Wang, 1995; Caiand Jiang, 1995). The induction of MT has beensuggested as one of the mechanisms for the adap-tive response in LDR exposure where it may actas a free radical scavenger (Cai and Cherian,1996). However, more work is needed to under-stand the potential role of MT in the adaptiveresponse.

L. Cai et al. / Toxicology 132 (1999) 85–98 95
5. Summary
Most of the published reports on the inductionof MT after exposure to ionizing and UV radia-tion are on rodent experimental models and cellsin culture. It is difficult to extrapolate these resultsdirectly to human exposure conditions. However,in general, these studies suggest that repeated lowdoses of exposure are better than a large singleradiation exposure to induce MT synthesis. It isunclear whether radiation itself can induce MTsynthesis or these effects are secondary due to theformation of free radicals or release of cytokines.The protective effect of MT in radiation injuryhas been demonstrated both in vivo and in cellsafter pre-induction of MT by various agents. Thisradioprotective effect may be either due to MTitself or may be modulated by other cellular fac-tors. The protective role of MT in radiation in-duced apoptosis is also unclear. The transgenicmouse models where the MT genes are deleted, orthe cells derived from them, provide an excellentopportunity to study the specific effects of MT inradiation injury. The role of MT in radiationexposure may be important not only in radiopro-tection but also in the treatment of certain typesof cancers with both drugs and radiation. Thepresence of MT may be one of the several factorsinvolved in the radioresistance of tumor tissueand the adaptive response in LDR. More studiesare required to understand the exact role of MTin radiation-induced injury, especially at lowdoses of long-term exposure to radiation.
Acknowledgements
Dr M. George Cherian was a visiting scientistat the National Institute for Environmental Stud-ies at Tsukuba and was supported by a Scienceand Technology Agency (STA) Fellowship.
References
Anstey, A., Marks, R., Long, C., et al., 1996. In vivo photoin-duction of metallothionein in human skin by ultravioletirradiation. J. Pathol. 178, 84–88.
Astor, M.B., Anderson, M.E., Meister, A., 1988. Relationshipbetween intracellular GSH levels and hypoxic cell ra-diosensitivity. Pharmacol. Ther. 39, 115–121.
Baba, T., Nakano, H., Tamai, K., et al., 1998. Inhibitoryeffect of b-thujaplicin on ultraviolet B-induced apoptosis inmouse keratinocytes. J. Invest. Dermatol. 110, 24–28.
Bakka, A., Johnsen, A.S., Endresen, L., Rugstad, H.E., 1982.Radioresistance in cells with high content of metalloth-ionein. Experientia 38, 381–383.
Bauman, J.W., Liu, J., Liu, Y.P., Klaassen, C.D., 1991. In-crease in metallothionein produced by chemicals that in-duce oxidative stress. Toxicol. Appl. Pharmacol. 110,347–354.
Biaglow, J.E., Clark, E.P., Epp, E.R., Morse-Guadio, M.,Varnes, M.E., Mitchell, J.B., 1983. Nonprotein thiols andradiation response of A549 human lung carcinoma cells.Int. J. Radiat. Biol. 44, 489–495.
Black, H.S., 1987. Potential involvement of free radical reac-tions in ultraviolet light-mediated cutaneous damage. Pho-tochem. Photobiol. 46, 213–221.
Blank, K.R., Rudoltz, M.S., Kao, G.D., Muschel, R.J.,McKenna, W.G., 1997. Review: the molecular regulationof apoptosis and implications for radiation oncology. Int.J. Radiat. Biol. 71, 455–466.
Cai, L., Cherian, M.G., 1996. Adaptive response to ionizingradiation-induced chromosome aberrations in rabbitlymphocytes: effect of pre-exposure to zinc, and coppersalts. Mutat. Res. 369, 233–241.
Cai, L., Cherian, M.G., 1997. Metallothionein in normal andabnormal spermatogenesis in mice. 4th Int. Metalloth-ionein Meeting, Kansas City, MO, Abstract No. 100.
Cai, L., Jiang, J., 1995. Mild hyperthermia can induce adapta-tion to cytogenetic damage caused by subsequent X irradi-ation. Radiat. Res. 134, 26–33.
Cai, L., Wang, P., 1995. Induction of a cytogenetic adaptiveresponse in germ cells of irradiated mice with very low-dose rate of chronic g-irradiation and its biological influ-ence on radiation-induced DNA or chromosomal damageand cell killing in their male offspring. Mutagenesis 10,95–100.
Cai, L., Koropatnick, J., Cherian, M.G., 1996. Metalloth-ionein and DNA damage. In: Gong, S.L. (Ed.), BiologicalEffects of Low Level Ionizing Radiation and MolecularBiology Research. Norman Bethune University of MedicalSciences, Changchun, pp. 77–88.
Chan, H.M., Cherian, M.G., 1993. Metallothionein isoformsin different mammalian species. Toxicologist 13, 165.
Cherian, M.G., 1993. Nuclear and cytoplasmic localization ofmetallothionein inn human liver during development andin tumor cells. In: Suzuki, K.T., Imura, N., Kimura, M.(Eds.), Metallothionein III, Biological Roles and MedicalImplications. Birkhauser, Basel, pp. 175–188.
Cherian, M.G., Chan, H.M., 1993. Biological functions ofmetallothionein: a review. In: Suzuki, K.T., Imura, N.,Kimura, M. (Eds.), Metallothionein III, Biological Rolesand Medical Implications. Birkhauser, Basel, pp. 87–109.

L. Cai et al. / Toxicology 132 (1999) 85–9896
Cherian, M.G., Ferguson, P.J., 1997. Metallothionein in cyto-toxicity and genotoxicity of metals. In: Hadjiliadis, N.D.(Ed.), Cytotoxic, Mutagenic and Carcinogenic potential ofHeavy Metals Related to Human Environment, NATOASI Series, 2. Environment, vol. 26. Kluwer, Dordrecht,pp. 217–230.
Choudhuri, S., McKim, J.M., Klaassen, C.D., 1993. Differen-tial expression of the metallothionein gene in liver andbrain of mice and rats. Toxicol. Appl. Pharmacol. 119,1–10.
Chubatsu, L.B., Meneghini, R., 1993. Metallothionein protectsDNA from oxidative damage. Biochem. J. 291, 193–198.
Deng, D.X., Cai, L., Cherian, M.G., 1997. Metallothioneinand apoptosis in thymus after radiation. Fundam. Appl.Toxicol. 36, 254.
Dudek, E.J., Peak, J.G., Roth, R.M., Peak, M.J., 1993. Isola-tion of V79 fibroblast cell lines containing elevated metal-lothionein levels that have increased resistance to thecytotoxic effects of ultraviolet-A radiation. Photochem.Photobiol. 58, 836–840.
Eichholtz-Wirth, H., Reidel, G., Hietel, B., 1993. Radiation-induced transient cisplatin resistance in murine fibrosar-coma cells associated with elevated metallothioneincontent. Br. J. Cancer 67, 1001–1006.
Fornace, A.J. Jr., Alamo, I. Jr., Hollander, M.C., 1988. DNAdamage-inducible transcripts in mammalian cells. Proc.Natl. Acad. Sci. USA 85, 8800–8804.
Fornace, A.J. Jr., Schalch, H., Alamo, I. Jr., 1988. Coordinateinduction of metallothioneins I and II in rodent cells byUV irradiation. Mol. Cell. Biol. 8, 4716–4720.
Gracheva, N.I., Kapitskaia, M.Z., Krutilina, R.I., Pereverzev,A.E., Stepan’ian, L.I., Tomilin, N.V., 1989. In vivo induc-tion of the metallothionein gene by cadmium ions and itseffect on endogenous colony formation in the spleen ofsublethally irradiated mice. Radiobiologiia 29, 737–741.
Greenstock, C.L., Jinot, C.P., Whitehouse, R.P., Sargent,M.D., 1987. DNA radiation damage and its modificationby metallothionein. Free Radic. Res. Commun. 2, 233–239.
Haimovitz-Friedman, A., Kolesnick, R.N., Fuks, Z., 1996.Modulation of the apoptotic response: potential for im-proving the outcome in clinical radiotherapy. Semin. Ra-diat. Oncol. 6, 273–283.
Hall, E.J., 1994. Radiobiology for the Radiologist. J.B. Lip-pincott, Philadelphia, PA.
Hallahan, D.E., Bleakman, D., Virudachalam, S., et al., 1994.The role of intracellular calcium in the cellular response toionizing radiation. Radiat. Res. 138, 392–400.
Hammond, R.R., Cai, L., LeBlanc M., Cherian, M.G., 1997.Induction of metallothionein expression and apoptotic celldeath in human primary CNS cultures by ionizing radia-tion. 4th Int. Metallothionein Meeting, Kansas City, MO,Abstract No. 168.
Hanada, K., Gange, R.W., Siebert, E., Hasan, T., 1991.Protective effects of Cd chloride against UVB injury inmouse skin and in cultured human cells: a possible role ofCd-induced metallothionein. Photodermatol. Photoim-muunol. Photomed. 8, 111–115.
Hanada, K., Baba, T., Hashimoto, I., Fukui, R., Watanabe,S., 1992. Possible role of cutaneous metallothionein inprotection against photo-oxidative stress-epidermal local-ization and scavenging activity for superoxide and hy-droxyl radicals. Photodermatol. Photoimmunol.Photomed. 9, 209–213.
Hanada, K., Sawamura, D., Nakano, H., Hashimoto, I., 1995.Possible role of 1,25-dihydroxyvitamin D3-induced metal-lothionein in photoprotection against UVB injury in mouseskin and cultured rat keratinocytes. J. Dermatol. Sci. 9,203–208.
Hanada, K., Tamai, K., Sawamura, D., Hashimoto, I., Mura-matsu, T., 1998. Dynamic changes in intracellular locationof metallothionein in rat keratinocytes after ultraviolet-Birradiation. J. Invest. Dermatol. 110, 98–100.
Hansen, C., Ablett, E., Green, A., et al., 1997. Biphasicresponse of the metallothionein promoter to ultravioletradiation in human melanoma cells. Photochem. Photo-biol. 65, 550–555.
Hayashi, H., 1988. Increase of hepatic and renal metalloth-ionein mRNA level induced by whole body irradiation.Nippon Igaku Hoshasen Gakkai Zasshi 48, 1467–1471.
Henry, R.B., Liu, L., Choudhuri, S., Klaassen, C.D., 1994.Species variation in hepatic metallothionein. Toxicol. Lett.74, 23–33.
Hidalgo, J., Campmany, L., Borras, M., Garvey, J.S., Ar-mario, A., 1988. Metallothionein response to stress in rats:role in free radical scavenging. Am. J. Physiol. 255, 518–524.
Izotova, N.I., Alekseeva, L.V., Pereverzev, A.E., Tomilin,N.V., 1992. The effect of cadmium on the survivability ofcolony-forming haematopoietic cells in mice exposed toX-ray irradiation. Tsitologiia 34, 105–109.
Jasani, B., Anstey, A., Marks, R., Lond, C.C., Pearse, A.D.,1993. Wild type p53 and metallothionein are expressedsimultaneously in UV irradiated skin: a possible link tophotocarcinogenesis. J. Invest. Dermatol. 101, 422.
Kagimoto, O., Naganuma, A., Imura, N., Toge, T., Niwa, O.,Yokoro, K., 1991. Effect of the administration of bismuthnitrate on radiogenic thymoma induction in mice. J. Ra-diat. Res. 32, 417–428.
Kaina, B., Lohrer, H., Karin, M., Herrlich, P., 1990. Overex-pressed human metallothionein IIA gene protects Chinesehamster ovary cells from killing by alkylating agents. Proc.Natl Acad Sci USA 87, 2710–2714.
Kelley, S.L., Basu, A., Teicher, B.A., Hacker, M.P., Hamer,D.H., Lazo, J.S., 1988. Overexpression of metallothioneinconfers resistance to anticancer drugs. Science 241, 1813–1815.
Kobayashi, S., Kasuya, M., Ishii, Y., et al., 1989. Effect ofzinc on rat X-ray cataract. J. Eye 6, 1249–1252.
Kobayashi, S., Hirota, Y., Sayato-Suzuki, J., et al., 1994.Possible role of metallothionein in the cellular defensemechanism against UVB irradiation in neonatal humanskin fibroblasts. Photochem. Photobiol. 59, 650–656.
Kondo, Y., Woo, E.S., Michalska, A.E., Choo, K.H., Lazo,J.S., 1995. Metallothionein null cells have increased sensi-tivity to anticancer drugs. Cancer Res. 55, 2021–2023.

L. Cai et al. / Toxicology 132 (1999) 85–98 97
Koropatnick, J., Cherian, M.G., 1988. Exposure to differentforms of Cd in mice: differences in metallothionein anda-fetoprotein mRNA induction in liver and kidney. J.Biochem. Toxicol. 3, 159–172.
Koropatnick, J., Pearson, J., 1993. Altered cisplatin and cad-mium resistance, and cell survival in Chinese hamsterovary cells expressing mouse metallothionein. Mol. Phar-macol. 44, 44–45.
Koropatnick, J., Leibbrand, M., Cherian, M.G., 1989. Organ-specific metallothionein induction in mice by X-irradiation.Radiat. Res. 119, 356–365.
Koterov, A.N., Sazykin, AIu., Filippovich, I.V., 1993. Theconnection between the content of metallothioneins inbone marrow and liver and the survivability of irradiatedmice after Cd chloride administration. Radiobiologiia 33,122–127.
Kramer, M., Sachsenmaier, C., Herrlich, P., Rahmsdorf, H.J.,1993. UV irradiation-induced interleukin-1 and basicfibroblast growth factor synthesis and release mediate partof the UV response. J. Biol. Chem. 268, 6734–6741.
Lau, J.C., Cai, L., Cherian, M.G., 1997. Effect of g-irradiationin metallothionein synthesis and lipid peroxidation intransgenic mice. Fundam. Appl. Toxicol. 36, 299.
Lieberman, M.W., Beach, L.R., Palmiter, R.D., 1983. Ultravi-olet radiation-induced metallothionein-I gene activation isassociated with extensive DNA demethylation. Cell 35,207–214.
Liebmann, J., Fisher, J., Lipschultz, C., Kuno, R., Kaufman,D.C., 1995. Enhanced glutathione peroxidase expressionprotects cells from hydroperoxides but not from radiationor doxorubicin. Cancer Res. 55, 4465–4470.
Lohrer, H., Robson, T., 1989. Overexpression of metalloth-ionein in CHO cells and its effect on cell killing by ionizingradiation and alkylating agents. Carcinogenesis 10, 2279–2284.
Margoshes, M., Vallee, B. L., 1957. A cadmium bindingprotein from equine kidney cortex. J. Am. Chem. Soc. 79,4813–4814.
Masters, B.A., Kelly, E.J., Quaife, C.J., Brinster, R.L.,Palmiter, R.D., 1994. Targeted disruption of metalloth-ionein-I and II genes increases sensitivity to Cd. Proc.Natl. Acad. Sci. USA 91, 584–588.
Matsubara, J., 1988. Metallothionein induction: a measure ofradioprotective action. Health. Phys. 55, 433–436.
Matsubara, J., Ishioka, K., Egawa, S., Inada, T., Machida, K.,1982. Protective effect of zinc against lethality of theirradiated mice. Proc. Soc. Radiol. Prot. 1, 358–369.
Matsubara, J., Shida, T., Ishioka, K., Egawa, S., Inada, T.,Machida, K., 1986. Protective effect of Zn against lethalityin irradiated mice. Environ. Res. 41, 558–567.
Matsubara, J., Tajima, Y., Karasawa, M., 1987. Promotion ofradioresistance by metallothionein induction prior to irra-diation. Environ. Res. 43, 66–74.
Matsubara, J., Tajima, Y., Karasawa, M., 1987. Metalloth-ionein induction as a potent means of radiation protectionin mice. Radiat. Res. 111, 267–275.
Matsubara, J., Tajima, Y., Ikeda, A., Kinoshita, T., Shi-moyama, T., 1988. A new perspective of radiation protec-tion by metallothionein induction. Pharmacol. Ther. 39,331–333.
Michel, C., Balla, I., 1987. Interaction between radiation andcadmium or mercury in mouse embryos during organogen-esis. Int. J. Radiat. Biol. 51, 1007–1019.
Michel, C., Fritz-Niggli, H., 1986. Teratogenic interactionsbetween cadmium and radiation in mice. Experientia 42,80–81.
Miura, M., Sasaki, T., 1990. Relationship between radiosensi-tivity and metallothionein content in clones from a mousesquamous cell carcinoma. Radiat. Res. 123, 171–175.
Miura, T., Muraoka, S., Ogiso, T., 1997. Antioxidant activityof metallothionein compared with reduced glutathione.Life Sci. 60, PL301–309.
Miura, N., Satoh, M., Imura, N., Naganuma, A., 1998. Pro-tective effect of bismuth nitrate against injury to the bonemarrow by g-irradiation in mice: possible involvement ofmetallothionein synthesis. J. Pharmacol. Exp. Ther. 286,1427–1430.
Mitchell, J.B., Biaglow, J.E., Russo, A., 1988. Role of glu-tathione and other endogenous thiols in radiation protec-tion. Pharmacol. Ther. 39, 269–274.
Murata, R., Nishimura, Y., Hiraoka, M., Abe, M., Satoh, M.,1995. Manganese chloride treatment does not protectagainst acute radiation injury of skin or crypt cells. Radiat.Res. 143, 316–319.
Neta, R., Okunieff, P., 1996. Cytokine-induced radiation pro-tection and sensitization. Semin. Radiat. Oncol. 6, 306–328.
Nishimura, N., Tohyama, C., Nishimura, H., Adelson D.L.,Reeve, V.E., 1997. UVB-induced immunosuppression andcutaneous metallothionein (MT) expression. 4th Int.Metallothionein Meeting, Kansas City, MO, Abstract No.169.
Ogata, H., Izoumo, Y., 1990. Mortality reduction in miceadministered a single abundant dose of zinc, manganese ormagnesium after irradiation by g-rays at sublethal doses.Radioisotopes 39, 573–576.
Ono, S.I., Cai, L., Cherian, M.G., 1998. Induction of brainmetallothionein by g-irradiation in transgenic mice andlipid peroxidation. Radiat. Res. 150, 52–57.
Osmak, M., 1993. Multifactorial molecular mechanisms areinvolved in resistance of preirradiated human cervix car-cinoma cells to cis-dichlorodiammineplatinum (II) and vin-cristine. Neoplasma 40, 97–101.
Osmak, M., Uzarevic, B., 1991. Mechanisms involved in resis-tance of preirradiated Chinese hamster V79 cells to cyto-toxic drugs are multifactorial. Res. Exp. Med. Berl. 191,413–421.
Ota, T., Hanada, K., Hashimoto, I., 1996. The effect of coldstress on UVB injury in mouse skin and cultured kerati-nocytes. Photochem. Photobiol. 64, 984–987.
Ren, Y., Smith, A., 1995. Mechanism of metallothionein generegulation by heme-hemopexin. Roles of protein kinase C,reactive oxygen species, and cis-acting elements. J. Biol.Chem. 270, 23988–23995.

L. Cai et al. / Toxicology 132 (1999) 85–9898
Renan, M.J., Dowman, P.I., 1989. Increased radioresistance oftumor cells exposed to metallothionein-inducing agents.Radiat. Res. 120, 442–455.
Rimoldi, D., Flessate, D.M., Samid, D., 1992. Changes in geneexpression by 193- and 248-nm excimer laser radiation incultured human fibroblasts. Radiat. Res. 131, 325–331.
Sato, M., Bremner, I., 1993. Oxygen free radicals and metal-lothionein. Free Radic. Biol. Med. 14, 325–337.
Sato, Y., Ohta, S., Shinoda, M., 1990. Studies on chemicalprotectors against radiation. XXXI. Protective effects ofAloe arborescens on skin injury induced by X-irradiation.Yakugaku Zasshi 110, 876–884.
Sato, M., Sasaki, M., Hojo, H., 1993. Induction of metalloth-ionein synthesis by oxidative stress and possible role inacute phase response. In: Suzuki, K.T., Imura, N.,Kimura, M. (Eds.), Metallothionein III, Biological Rolesand Medical Implications. Birkhauser, Basel, pp. 125–140.
Satoh, M., Miura, N., Naganuma, A., Matsuzaki, N., Kawa-mura, E., Imura, N., 1989. Prevention of adverse effects ofg-ray irradiation after metallothionein induction by bis-muth subnitrate in mice. Eur. J. Cancer Clin. Oncol. 25,1727–1731.
Satoh, M., Kloth, D.M., Kadhim, S.A., et al., 1993. Modula-tion of both cisplatin nephrotoxicity and drug resistance inmurine bladder tumor by controlling metallothionein syn-thesis. Cancer Res. 53, 1829–1932.
Satoh, M., Cherian, M.G., Imura, N., Shimizu, H., 1994.Modulation of resistance to anticancer drugs by inhibitionof metallothionein synthesis. Cancer Res. 54, 5255–5257.
Satoh, M., Tsuji, Y., Watanabe, Y., et al., 1996. Metalloth-ionein content increased in the liver of mice exposed tomagnetic fields. Arch. Toxicol. 70, 315–318.
Satoh, M., Shibuya, K., Tohyama, C., 1997. Enhanced bonemarrow injury by X-ray irradiation in metallothionein-Iand -II null mice. 4th Int. Metallothionein Meet., KansasCity, MO, 1997, Abstract No. 141.
Schwarz, M.A., Lazo, J.S., Yalowich, J.C., et al., 1994. Cyto-plasmic metallothionein overexpression protects NIH 3T3cells from tert-butyl hydroperoxide toxicity. J. Biol. Chem.269, 15234–15238.
Shibuya, K., Satoh, M., Muraoka, M., Watanabe, Y., Oida,M., Shimizu, H., 1995. Induction of metallothionein syn-thesis in transplanted murine tumors by X irradiation.Radiat. Res. 143, 54–57.
Shibuya, K., Cherian, M.G., Satoh, M., 1997. Sensitivity toradiation treatment and changes in metallothionein synthe-
sis in a transplanted murine tumor. Radiat. Res. 148,235–239.
Shiraishi, N., Waalkes, M.P., 1994. Enhancement of metal-lothionein gene expression in male Wister (WF/NCr) ratsby treatment with calmodulin inhibitors: potential rolecalcium regulatory pathways in metallothionein induction.Toxicol. Appl. Pharmcol. 125, 97–103.
Shiraishi, N., Aono, K., Utsumi, K., 1983. Increased metal-lothionein content in rat liver induced by X-irradiation andexposure to high oxygen tension. Radiat. Res. 95, 298–302.
Shiraishi, N., Yamamoto, H., Takeda, Y., et al., 1986. In-creased metallothionein content in rat liver and kidneyfollowing X-irradiation. Toxicol. Appl. Pharmacol. 85,128–134.
Shiraishi, N., Hayashi, H., Hiraki, Y., et al., 1989. Elevationin metallothionein messenger RNA in rat tissues afterexposure to X-irradiation. Toxicol. Appl. Pharmacol. 98,501–506.
Sorenson, J.R., Soderberg, L.S., Chang, L.W., 1995. Radia-tion protection and radiation recovery with essential metal-loelement chelates. Proc. Soc. Expt. Biol. Med. 210,191–204.
Stein, B., Rahmsdorf, H.J., Steffen, A., Litfin, M., Herrlich,P., 1989. UV-induced DNA damage is an intermediate stepin UV-induced expression of human immunodeficiencyvirus type 1, collagenase, c-fos, and metallothionein. Mol.Cell Biol. 9, 5169–5181.
Thornalley, P.J., Vasak, M., 1985. Possible role for metalloth-ionein in protection against radiation induced oxidaticstress. Kinetics and mechanism of its reaction with super-oxide and hydroxyl radicals. Biochim. Biophys. Acta 827,36–44.
Togami, I., 1987. A factor of increase of hepatic metalloth-ionein content in normal and adrenolectomized rats in-duced by whole body irradiation. Nippon Igaku HoshasenGakkai Zasshi 47, 69–73.
Wirth, P., Thorgeirsson, S.S., 1978. Glutathione synthesis anddegradation in fetal and adult rat liver and Novik offhepatoma. Cancer Res. 38, 2861–2865.
Xiong, X., Arizono, K., Garrett, S.H., Brady, F.O., 1992.Induction of zinc metallothionein by calcium ionophore invivo and in vitro. FEBS Lett. 299, 192–196.
Yu, C.W., Chen, J.H., Lin, L.Y., 1997. Metal-induced metal-lothionein gene expression can be inactivated by proteinkinase C inhibitor. FEBS Lett. 40, 69–73.
.





























