Embed Size (px)
Citation preview

METHODS TO ASSESS THE IMPACT OF EXTRACTION OF NON-TIMBER TROPICAL FOREST PRODUCTS ON PLANT POPULATIONS 1
P A M E L A H A L L A N D K A M A L J I T B A W A
Hall, Pamela, and Kamaljit Bawa (Biology Department, University of Massachusetts, Boston, MA 02215, U.S.A.). M~THODS TO ASSESS THE IMPACT OF EXTRACTION OF NON-TIMBER TROmCAL FOREST PRODUCTS ON PLANT POPULATIONS. Economic Botany 47:234-247. 1993. Thousands of plant and animal species in tropical regions provide a variety of non-timber products that are used by billions of people all over the world. Conservation and long term utilization of these species require that they be harvested on a sustainable basis. However, the extent to which non- timber forest products are exploited without adverse effects on natural populations is not known. There is in fact considerable evidence for non-sustainable harvest of non-timber products. We outline methods that may be used to assess the impact of harvest on population processes of the species that are being harvested. We present sampling protocols for rapid assessment as well as long term monitoring of populations. We briefly consider the limitations of these methods and suggest that the monitoring protocols we outline should be part of an overall management plan designed to extract and utilize non-timber tropical forest products on a long term basis.
Key Words: sustainability; non-timber tropical forest products; tropical forests; economic valuation; ecological methods.
Non- t imber forest products (NTFP) from tropical forests have become a subject of mount- ing international concern as a result of several factors which affect the availabil i ty o f many trop- ical species. First, continuing deforestation in the tropics threatens to el iminate hundreds, i f not thousands, o f species during the next few de- cades. Second, effective conservation and man- agement o f N T F P is viewed as a means of im- proving the rural economy and the well-being of indigenous societies that rely on N T F P for sub- sistence and cash income. Third, there is a grow- ing recognition that rural communit ies which rely on a variety of plant and animal species for their l ivelihoods should participate in conservation of tropical biodiversity. Fourth, many species that yield N T F P are assumed to be potential sources of new genes as well as new products, particularly drugs, and thus are valuable to the international agriculture and pharmaceutical industries. In short, the full economic valuation of endangered
Received 8 April 1993; accepted 5 May 1993.
tropical forest ecosystems is not possible without consideration of the value of non- t imber forest products.
Several lines of evidence suggest that in many cases the N T F P are not being exploited on a sustainable basis. For example, recent data from the state of Tami lnadu in India show decreases in bamboo yields (Table 1). Whether this decline is due to decreased yield in part icular areas or reduction in total area harvested is not known. Fernandes, Menon, and Viegas (1988) have com- piled extensive data on many N T F P harvested by indigenous societies in Orissa, another state in India; these data indicate a decline in yields. Again, in the absence of information about changes in the total area harvested, it is difficult to speculate on the reasons for the decline. Chambers, Saxena, and Shah (1989) examined government records of yields of N T F P and noted a decrease for several products in recent years. They attribute this decline to changes in the mar- ket structure and to profit incentives from the nationalization of trade in NTFP. Other expla- nations of reduced yields in India include defor-
Economic Botany 47(3) pp. 234-247. 1993 �9 1993, by The New York Botanical Garden, Bronx, NY 10458 U.S.A.

1993] HALL & BAWA: IMPACT OF EXTRACTION 235
estation, which reduces the area that can be har- vested, and growing human pressure on forests, which may cause unsustainable resource use. Chambers, Saxena, and Shah (1989) argue that the potential yields o f NTFP are far greater than realized yields, but they do not address the issue of sustainability.
The evidence for unsustainable yields is not confined to India. Reining et al. (199 l) show that two palm species collected and exported for floral arrangements are harvested on an unsustainable basis in the Maya Biosphere Reserve in Guate- mala, and Nepstad et al. (1992) present evidence for depletion of plant populations in Amazonian forests used by rubber tappers. Overharvesting is not restricted to products that are being col- lected for commercial markets; Browder (1992) cites many examples from South America of NTFP used primarily by local communities that are also being depleted by unsustainable extrac- tion. Browder also points out that once NTFP enter the commercial markets, their harvest can- not be sustained without external inputs (e.g., subsidies). The valuation models, however, in- corporate prices of NTFP based on market econ- omy, but assume no external subsidies for sus- tained production. This practice also contributes to unsustainable use of NTFP.
Although there is considerable evidence for overharvesting of NTFP, quantitative analyses of the effect of extraction on natural populations are lacking. Without such analyses, it is impos- sible to assess the effect of harvesting on deple- tion of NTFP in natural communities, nor is it possible to design appropriate conservation and management plans. In this paper, we describe methods to determine if plant species that yield NTFP are being exploited on a sustainable basis. We begin by defining sustainability in the sense it is used in the paper and then describe methods to monitor plant populations to assess the impact of harvest on the long term viability of popula- tion. We then point out the limitations of the methods, and we conclude with a plea for inte- grating ecological approaches into management goals that are often strictly based on economic considerations.
S U S T A I N A B I L I T Y
Our discussion is concerned with ecological rather than economic sustainability. Extraction o f NTFP may be economically sustainable if the value, adjusted for inflation, increases or remains
TABLE 1. BAMBOO YIELDS IN TAMIL NADU STATE,
INDIA (SOURCE: GOVERNMENT OF TAMIL NADU).
Y~r Q~ntity (MT) Value (lakhO
1985-1986 11 108 11.66 1986-1987 12 541 13.17 1988-1989 4672 7.36 1990-1991 2399 3.90
N.B. $1.00 ffi Rs 30.00 (1992 exchange ~tc). One lakh = 100000 Rs.
constant, but economic sustainability is not al- ways consistent with ecological sustainability. For example, overharvesting may lead to continuing decline of populations while persistent demand keeps the market value constant. As the popu- lations of NTFP species become depleted and the products become scarce, there may actually be an increase in economic return if the demand remains the same. Scarcity may also increase the cost of extraction, driving the price upward and reducing the demand (Homma 1992). Ultimate- ly, with complete resource depletion, there will be no sustainability of any kind, only local ex- tinctions of populations, and eventually extinc- tion of the species. In economic terms, the effects of unsustainable extraction take a long time to be detected, especially for long-lived trees.
In ecological terms, however, extraction is considered sustainable if the harvest has no long term deleterious effect on the reproduction and regeneration of populations being harvested in comparison to equivalent non-harvested natural populations. Furthermore, sustainable harvest should have no discernable adverse effect on oth- er species in the community, or on ecosystem structure and function. These effects also can be determined by comparing harvested to unhar- vested systems. The adverse effects may be man- ifested in several ways. For example, harvesting of seeds and fruits decreases the availability of food for frugivore (fruit eating animals) popu- lations, limiting the number o f organisms and perhaps decreasing the diversity o f frugivores in a given community. These changes in turn may alter other trophic (food web) relationships and thus negatively affect other species in the com- munity. Even something as simple as removal of dead wood and leaves may devastate detriti- yore (decomposers, e.g., microbes, fungi, etc.) communities critical to the cycling o f the nutri- ents which support the vegetation in the forest.

236 ECONOMIC BOTANY [VOL. 47
ASSESSING SUSTAINABILITY
The methods used to assess ecological sustain- ability are more restricted than our definition of sustainability would indicate. Though biodiver- sity occurs at various levels of biological orga- nization-the gene, the population, the com- munity, the ecosystem--and extraction can impact all these levels, our ability to assess sus- tainability of harvesting activities depends upon our understanding of the dynamics of species, communities, and ecosystems. Current ecologi- cal theory provides us with a fairly robust un- derstanding of population dynamics of single species, since this area has been well studied, but our theoretical understanding of dynamics of communities and ecosystems is not as advanced. These systems are more complex and thus much harder to model and study, requiring greater time and effort and consideration of much larger spa- tial units. This problem is one reason for the focus on population level dynamics of single spe- cies adopted in the rest of this paper. Another reason is that so little is known at the population level about effects of harvesting NTFP on pop- ulations of the target species. For example, the quantity and manner in which NTFP are har- vested locally is not well documented except for a very few species. The discussion below there- fore is restricted to methods that one might use to evaluate the effects of harvest on population dynamics of the species in question. A further restriction is that only plant species are consid- ered.
The methods described below are derived from standard quantitative techniques employed to sample vegetation (Greig-Smith 1957; Kershaw 1973; Mueller-Dombois and Ellenberg 1974). Preliminary data from field studies indicate that the proposed methods have the potential to quantify the impact of NTFP collection on nat- ural populations (K. Murali and U. Aggarwal, pers. comm.).
In order to assess sustainability of harvesting a non-timber forest product, knowledge of the natural distribution, abundance, population structure and dynamics, and variation of these factors across a landscape is required for each species. Data concerning the distribution, abun- dance and population structure provide a static picture of the species populations. However, populations change over time, and sampling must continue in order to determine patterns of nat-
ural change in population structure. The effects of harvesting are superimposed upon the natural population dynamics. Since population charac- teristics, both static and dynamic, vary naturally over time, only direct comparison between nat- ural and harvested populations yields sufficient information to assess sustainability. Compara- tive sampling of natural populations and popu- lations subjected to different intensities of re- source extraction must be undertaken to determine the status of populations that are being harvested.
Identifying the distribution, abundance, pop- ulation structure (density, age/size distribution, number of reproductive adults), population dy- namics (mortality, recruitment, growth and re- productive rates) and the effect of harvesting on these parameters requires different sampling strategies. Collection of data on species abun- dance and distribution is an important prereq- uisite to investigating natural population dynam- ics and the impact of harvest on regeneration, since knowledge of these factors will determine the sampling design for gathering further infor- mation.
SAMPLING FOR DISTRIBUTION AND ABUNDANCE
The natural distribution and abundance of a species is partly a function of the spatial vari- ability in available habitats and the species' ca- pacity to colonize these habitats. Preferred hab- itats may be able to support population regeneration and therefore have persistent pop- ulations of a particular species, while other, mar- ginal habitats, may not support in situ regener- ation. However, if dispersal into marginal habitats from preferred habitats occurs faster than the loss of individuals due to natural mortality, then marginal habitats will have sustainable popula- tions. If the inverse is true, then marginal hab- itats will rapidly lose individuals, and the species will not persist in these areas.
Past and current harvesting practices may af- fect distribution and abundance of a species. For instance, if harvesting occurs in both preferred and marginal habitats, but reproduction and dis- persal is still sufficient to allow for the reestab- lishment of marginal habitat populations, then harvesting will be sustainable in both habitat types. When harvesting intensity is very great, however, there may not be sufficient propagules

1993] HALL & BAWA: IMPACT OF EXTRACTION 237
for dispersal into marginal habitats even i f re- generation occurs in the preferred habitat, and the populat ions will rapidly go extinct. Variat ion in effective dispersal from preferred to marginal habitats may occur naturally or may be con- trolled by harvesting practices. In order to dis- tinguish natural perturbations in distribution and abundance from those generated by harvesting pressures, simultaneous studies o f areas that en- compass both the range o f habitats which the species inhabits and the varying intensities of resource use should be undertaken. It is of par- amount importance to include a natural area which is not subject to human- induced distur- bance and which includes the range o f habitats used for harvesting; otherwise there will be no baseline data for determining the effects of har- vest.
M E T H O D S FOR SAMPLING DISTRIBUTION AND
ABUNDANCE
The best method for sampling a large, hetero- geneous area is to establish multiple transects, preferably parallel, which run along any topo- graphical, climatic or soil clines that exist. In addit ion, i f harvest intensity also varies spatially either along or across an environmental gradient, it is impor tant to establish paired transects be- tween harvested and undisturbed habitats in this area. A simple design calls for at least two, pref- erably three, transects 10 m in width and 1 km long, or 1.0 ha in total area. These transects must cover the environmental and harvest intensity variat ion to obtain some degree o f replication. A max imum of 5 transects will provide a good, block replicated sample. It is impor tant to sam- ple throughout the length of each transect for individuals of all age/size classes; however, small individuals will probably be more abundant than large individuals, so that a nested sampling de- sign should be used. For instance, tree-sized in- dividuals may be enumerated throughout the en- tire length o f each transect, saplings enumerated in l0 x 10 m plots placed every 100 m within the transect (a total of l 0 for 0.1 ha total area), and seedlings enumerated in 2 x 2 m plots placed within the sapling plots. For this initial enumer- ation, the largest individuals, for which the sam- ple size will probably be the smallest, should be permanently tagged, measured for size (e.g., di- ameter at breast height or dbh, height) and re-
productive condit ion recorded. Marking o f larger individuals will allow for easy relocation of ini- tially less well marked areas where smaller in- dividuals are sampled. Even i f an individual is not currently producing flowers or fruits, any ev- idence that such an individual has been or will shortly be reproductive should be recorded. For smaller individuals, the size and number could be recorded for the first enumerat ion so that it could be rapidly completed. Informat ion regard- ing topographic features, soil conditions, general description of the changes in habitat along the transect, as well as evidence of harvesting should be collected so that a prel iminary classification of sites according to variat ion in habitat and in- tensity of harvest can be accomplished.
Preliminary Analysis of Distribution and Abundance Data
Three types of information can be derived from this initial sampling of distr ibution and abun- dance. Though only static data on populat ions are available at this point, one can begin to iden- tify what are marginal versus preferred habitats for the species, determine how well the species was sampled within the transects, and assess the populat ion structure of the species. This infor- mat ion can be derived from these data by con- structing age/size distributions. Categories of habitat type and harvesting intensities can also be created from the descriptions that accompa- nied each transect. Further sampling should be guided by the distr ibution of populat ions within the transects to identify habitats or harvesting intensities within which the species is poorly rep- resented. In addi t ion inferences about popula- tion dynamics can be made from these distri- bu t ions , though on ly s ta t ic i n f o r m a t i o n on populat ion structure is provided. However, i f there is a suggestion that patterns of age/size dis- tr ibutions among habitats or ranges of harvesting intensities change over time, data must be col- lected at intervals in order to determine i f such changes are occurring.
I f habitats vary greatly and can be demarcated within the transects, separate distr ibutions for each habitat and within habitat type areas that vary in harvest intensity should be made. The construction of age/size distr ibutions for these categories will indicate how well the species was sampled within the transects. For instance, one
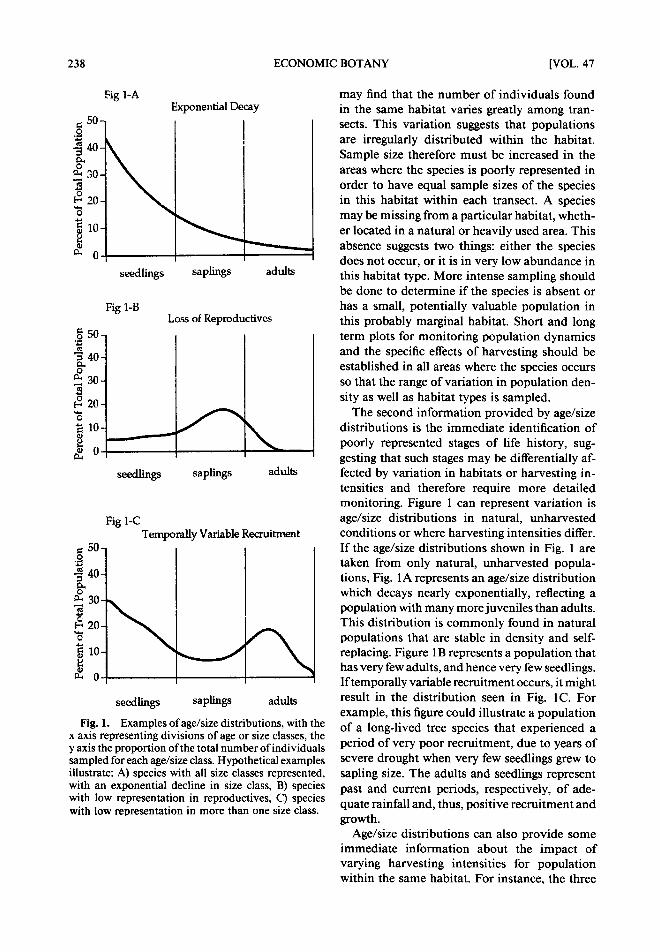
238 ECONOMIC BOTANY [VOL. 47
Fig 1-A
50-
~ 30-
~ 2 0 -
~ 10-
~ 0
seedlings
Exponential Decay
saplings adults
Fig 1-B
o ~ 50-
~40 - ~' 30-
~ 2 0 -
10-
d 0
Loss of Reproductives
seedlings saplings adults
Fig 1-C Temporally Variable Recruitment
50-
~ 40- o
30-
~ 2 0 - ~ ~
i 10-
~ 0
seedlings saplings adults
F i g . 1. Examples of age/size distributions, with the x axis representing divisions of age or size classes, the y axis the proportion of the total number of individuals sampled for each age/size class. Hypothetical examples illustrate: A) species with all size classes represented, with an exponential decline in size class, B) species with low representation in reproductives, C) species with low representation in more than one size class.
may find that the number of individuals found in the same habitat varies greatly among tran- sects. This variation suggests that populations are irregularly distributed within the habitat. Sample size therefore must be increased in the areas where the species is poorly represented in order to have equal sample sizes of the species in this habitat within each transect. A species may be missing from a particular habitat, wheth- er located in a natural or heavily used area. This absence suggests two things: either the species does not occur, or it is in very low abundance in this habitat type. More intense sampling should be done to determine if the species is absent or has a small, potentially valuable population in this probably marginal habitat. Short and long term plots for monitoring population dynamics and the specific effects of harvesting should be established in all areas where the species occurs so that the range of variation in population den- sity as well as habitat types is sampled.
The second information provided by age/size distributions is the immediate identification of poorly represented stages of life history, sug- gesting that such stages may be differentially af- fected by variation in habitats or harvesting in- tensities and therefore require more detailed monitoring. Figure 1 can represent variation is age/size distributions in natural, unharvested conditions or where harvesting intensities differ. If the age/size distributions shown in Fig. l are taken from only natural, unharvested popula- tions, Fig. 1A represents an age/size distribution which decays nearly exponentially, reflecting a population with many more juveniles than adults. This distribution is commonly found in natural populations that are stable in density and self- replacing. Figure 1B represents a population that has very few adults, and hence very few seedlings. If temporally variable recruitment occurs, it might result in the distribution seen in Fig. I C. For example, this figure could illustrate a population of a long-lived tree species that experienced a period of very poor recruitment, due to years of severe drought when very few seedlings grew to sapling size. The adults and seedlings represent past and current periods, respectively, of ade- quate rainfall and, thus, positive recruitment and growth.
Age/size distributions can also provide some immediate information about the impact of varying harvesting intensities for population within the same habitat. For instance, the three

1993] HALL & BAWA: IMPACT OF EXTRACTION 239
distributions in Fig. l could also represent the same species with A representing natural, un- harvested condition; B, intense, current har- vesting of adults; and C, an area of past intense harvesting o f adults. As the density of adults di- minishes to a point where they are difficult to find or locally extinct, harvesting may shift to a different location and the intensely harvested population will have a chance to recover as sap- lings grow to adult size and begin producing seed- lings. However, the loss o f recruitment from the period o f intense harvesting ofreproductives will remain evident in the population structure for some time. In this example, repeated, intense harvesting of adults at short term intervals would result in too few saplings to replace the harvested population, ultimately leading to extinction of the population.
The most intractable element of population dynamics to measure is temporal variability in mortality or recruitment, from either natural causes or human harvesting. This factor can only be investigated over long periods o f time. Again, using Fig. 1 as an example, these age/size distri- butions could represent a sequential pattern of change in population structure of a natural, un- harvested population. I f so, it will be very dif- ficult to distinguish the natural, temporal vari- ation in recruitment and mortality rates from any changes that are the result of harvesting. This example illustrates the importance of having multiple areas, particularly a control site where no harvesting occurs, in order to distinguish nat- ural from human-induced changes in age/size distribution. Therefore, it is important from the beginning to establish control plots that will con- tinue to address the issues o f natural, unhar- vested population dynamics long after the im- mediate information about harvesting impact has been evaluated.
EFFECTS OF HARVESTING ON POPULATION DYNAMICS
Harvesting can have both short and long term effects on population dynamics. Though every stage of the plant life cycle is ultimately critical to population maintenance, there may be prox- imate effects on particular stages o f the life his- tory for which the effect o f harvesting can be rapidly assessed. Immediate short term effects of harvesting can be seen in the decline of the har- vested product and changes in the density, growth rates or, if applicable, reproductive capacity of
the life history stage from which the product is harvested. Secondary short term effects include changes in population structure that result from immediate short term effect. For instance, har- vesting latex from large, mature trees may gen- erate an immediate effect of higher mortality rates among adults or minimally lower fruit produc- tion of heavily tapped trees. The lowered fruit production will have a secondary short term ef- fect on seedling recruitment and eventually a long term effect on the future density of adults. The final long term effect may be an overall lowering of density and a decrease in the average size and amount of reproductive output of adults. Low- ering the population size of a species may make it vulnerable to extinction by leaving it with less capacity to recover from catastrophic events such as prolonged droughts or tropical storms.
To distinguish short and long term effects o f harvesting on population dynamics monitoring populations should be done at two temporal stages: first rapidly assess the immediate short term impacts of harvesting on current population structure, and second, determine long term change in population dynamics (e.g., effects of temporal variation in recruitment and mortality on pop- ulation structure). The details o f sampling design vary depending on the life history of the species, the plant product that is being harvested and how it is harvested. For example, monitoring an an- nual herbaceous species for which the entire in- dividual is harvested requires a completely dif- ferent sampling design from monitoring a large tree species from which only fallen fruits are har- vested.
As with the analysis of distribution and abun- dance, all monitoring studies must include pop- ulations that are not subjected to harvesting and these populations must occur in the same range of habitats that harvested populations are found. In addition, monitoring of harvested populations must occur over a range of harvesting intensities. Immediate effects may be more obvious under intense harvesting, but both high and low levels of harvesting may result in the same secondary short term effects and eventually require similar long term management approaches to achieve sustainable harvest. By comparing the dynamics of harvested versus unharvested populations, and within the harvested populations, effects of vary- ing harvest intensities, short term proximate and long term effects of harvesting on population dy- namics can be distinguished.

240 ECONOMIC BOTANY [VOL. 47
Rapid Assessment First sampling for rapid assessment of har-
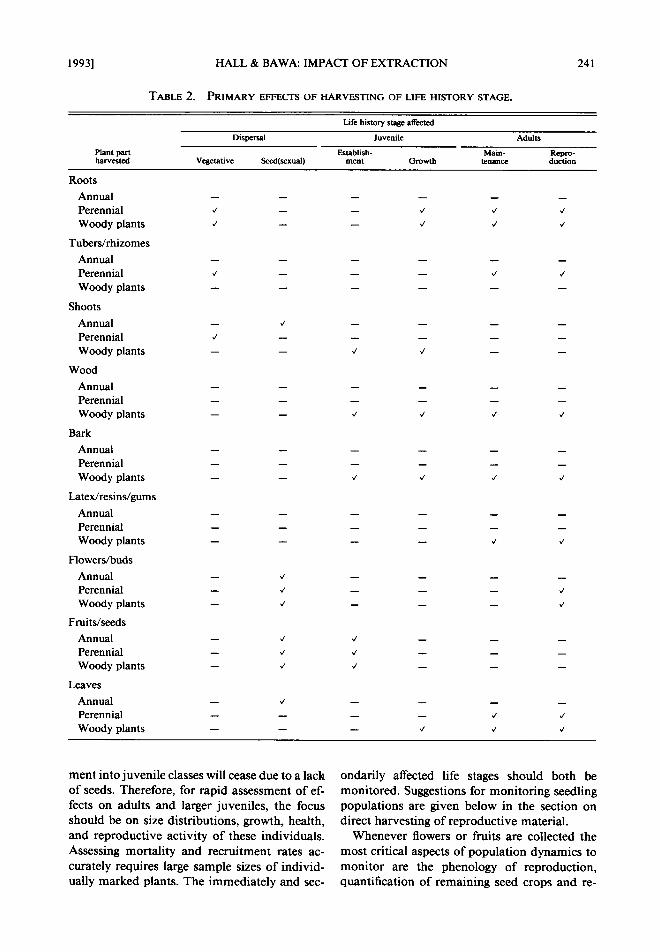
vesting effects should focus on the life stage that is being harvested (immediate short term effect of harvesting) and the life stage which is most strongly affected (secondary short term effect of harvesting). The plant part that is harvested will have different immediate effects on regeneration, and manifest secondary effects on different stages of the life cycle. Table 2 suggests a way to or- ganize the interactions among life forms, life his- tory stages, and harvesting. The life history stage that is most immediately affected by harvest and the strongest potential secondary effect on regen- eration are marked for each combination of plant part harvested versus life history stage for three plant life forms: annual herbs, perennial herbs, and woody species including shrubs and trees.
Sampling Designs for Rapid Assessment Annual Species. When annual herbs are har-
vested, complete destruction of the reproductive adult occurs. Therefore, there should be very strong effects of harvesting intensity on the pop- ulation dynamics of such a species because har- vesting eliminates at least a part of the current population. Regardless of what part of the plant is used, the most immediate impact of harvest will be on the dispersal of seeds. If plants are harvested before fruit set has occurred or seeds are the desirable material, then only those in- dividuals that escape detection or are purposely not collected will supply seeds for the next gen- eration. When all individuals of a population are harvested, marginal habitats may become the only seed source. Dispersal from marginal hab- itats may be severely limited and if so, the lack of dispersal will result in a rapid disappearance of the species from desirable, easily harvested habitats. If plants are harvested after fruit set (seeds are not of value), then seeds may have been adequately dispersed prior to or during the process of harvesting and the population may be self-perpetuating. Assessment of the secondary short term impact of harvesting should focus on identifying the seed source for subsequent years. Therefore, increased sampling intensity in mar- ginal habitats and measurement of the seeds re- maining within the harvested areas are suggest- ed.
Herbaceous Perennial Species. The harvesting of perennial herbs may also include complete
destruction of the adult, but many perennials are also harvested for their underground storage tis- sue. Even if adults do not die from harvesting of tubers or rhizomes, their reproductive capacity may be severely affected. While this effect may not hamper regeneration of storage tissue, over time there may be little replacement of individ- uals occurring. Sampling should therefore focus on mortality rates of harvested adults and on changes in reproductive capacity associated with harvesting intensity. Also, quantification of the amount of natural regeneration that occurs via vegetative reproduction versus seed production should be a major focus.
Woody Perennial Species. A larger array of materials can be harvested from shrubs and small and large tree species (woody perennials). Sam- piing to assess harvesting effects on woody pe- rennials will depend upon whether reproductive adults, juveniles, or both are being harvested. Because woody perennial species reach repro- ductive maturity at different sizes, harvesting of the same materials may affect their population dynamics in very different ways. For instance, bark may be harvested from juvenile and larger reproductives of large tree species but only from reproductives of shrubs and small tree species. Leaf material may only be easily harvested from juveniles of large tree species while leaves of shrubs and small tree species may be harvested from both juveniles and reproductives.
Harvesting of materials from adults will re- quire monitoring that focuses on the effects of harvest on reproductive output and increasing mortality rates due to disease associated with damage (e.g., cutting for harvest of firewood, res- ins, gums, etc.). For example, collection of fire- wood by removing branches but not cutting the main bole may require years for regrowth. Not only are resources probably diverted from re- production to regenerate branches, but the actual sites of reproduction may be lost and fruit pro- duction impossible until the branch has regrown. Constant pruning may lower reproductive out- put of affected individuals so severely that re- cruitment of new individuals into the population will not occur.
If juveniles are the focus of harvesting, then mortality and growth rates of these individuals need to be closely monitored. Constant removal of leaf material from juveniles, especially if young leaves are preferred, may not allow trees to reach reproductive maturity and eventually recruit-
1993] 241 HALL & BAWA: IMPACT OF EXTRACTION
T A B L E 2 . P R I M A R Y E F F E C T S O F H A R V E S T I N G O F L I F E H I S T O R Y S T A G E .
L i f e h i s t o r y s t a g e a f f e c t e d
D i s p e r s a l J u v e n i l e A d u l t s
P l a n t p a r t E s t a b f i s h - M a i n - R e p r o - h a r v e s t e d V e g e t a t i v e S e e d ( s e x u a l ) m e n t G r o w t h t e n a n c e d u c t i o n
Roots Annual Perennial Woody plants
Tubers/rhizomes
Annual Perennial Woody plants
Shoots
Annual Perennial Woody plants
Wood
Annual Perennial Woody plants
Bark Annual Perennial Woody plants
Latex/resins/gums
Annual Perennial Woody plants
Flowers/buds Annual Perennial Woody plants
Fruits/seeds Annual Perennial Woody plants
Leaves
Annual Perennial Woody plants
J D _ _ J j j
J m _ _ J J ~/
m J m _ _
J _ _ _ _ m
m m J J
m
m
m
m
B
B
D
m
D
m
m
m
m
D
D
, / J ~/ J
J J J J
m
n B
J , /
D
B
J . . . .
J - - _ _ _ _ J
,./ _ _ m m J
D
n
D
m
m
J J
J J
J J
w
m
J . . . .
- - - - - - 4 J
_ _ _ _ J J J
ment into juvenile classes will cease due to a lack of seeds. Therefore, for rapid assessment of ef- fects on adults and larger juveniles, the focus should be on size distributions, growth, health, and reproductive activity of these individuals. Assessing mortality and recruitment rates ac- curately requires large sample sizes of individ- ually marked plants. The immediately and sec-
ondarily affected life stages should both be monitored. Suggestions for monitoring seedling populations are given below in the section on direct harvesting of reproductive material.
Whenever flowers or fruits are collected the most critical aspects of population dynamics to monitor are the phenology of reproduction, quantification of remaining seed crops and re-

242 ECONOMIC BOTANY [VOL. 47
cruitment and establishment of seedlings. As stated in the sections on annual and herbaceous perennials, change in recruitment of seedlings is an impor tant potential secondary short term ef- fect that may occur and should be moni tored whenever materials that may affect reproductive capacity are harvested from adults. Harvesting of flowers or fruits may not directly affect the health or reproductive capacity o f an individual but i f all flowers or fruits are harvested, regen- eration will cease. Depending on the generation t ime of the species, the species and thus its prod- ucts may be rapidly lost from the forest.
For most species, seed production is in ap- parent great excess in comparison to that needed to replace the adult population. However, even though seed product ion is very high and natural mortal i ty may also be high, the rate of establish- ment o f seedlings and hence eventual regenera- t ion o f adults may be extremely sensitive to small changes in seed crop size and in mortal i ty rates among seedlings. Though it may appear that hu- mans are only harvesting seeds that would die under natural conditions, they may also be com- peting with animal (including insect) species that require the seed resource. This competi t ion may, in turn, lower the animal populat ions or increase their foraging ranges; furthermore, fruits or seeds that would have escaped predation in the absence o f human competi tors may be more effectively sought, resulting in much reduced recruitment rates o f seedlings to the population.
Under natural conditions, density dependent mortal i ty may occur among seedlings and be- cause turnover in seedlings is rapid, this effect can be assessed over a relatively short t ime in- terval. The effect of harvesting may be to lower local density and actually decrease the mortal i ty rates o f the remaining seeds and subsequent seedlings. Therefore, monitor ing populat ions from which seeds are removed should include sampling designs that take into account differ- ential density effects and whether changes in den- sity are a function of human or animal foraging. Again, actual mortal i ty and recruitment rates of seedlings are needed so large samples of individ- ually marked plants will be required. Also, be- cause morta l i ty or recruitment may be periodic or seasonal, more frequent enumerat ions are sug- gested in order to obtain information on how temporal variat ion in these parameters is affect- ed by temporal ly variable harvesting and inten- sity.
Quantification of phenological condit ion of adults should also be undertaken. This analysis can often be done with a simple categorical sys- tem that records the absence of flowers or fruits, and the proport ion of crown bearing flowers or fruits. Quantification of fruit production will usually require laying out seed traps which are arranged either around focal trees in a radial fashion or in a grid covering a given area. The distance to the nearest adult tree should be re- corded for each trap. Alternately, transects can be used, again either radiating from a focal in- dividual or arranged in a grid. However, seeds that are encountered should be marked in some manner so that the appearance of new seeds can be accurately detected.
Sampling intervals for all measures described above depend on the nature of the measure and the type of information required. For example, adult reproductive capacity will require pheno- logical information and therefore frequent sam- pling throughout the year. Monitoring adults or juveniles for growth and health will require es- tablishment of a permanent point of measure- ment on each individual and a s tandard method to assess size. Clear, precise, and reliably re- peated quantified data on health condit ions must also be collected. Monitoring for mortal i ty or recruitment may not require frequent sampling of larger individuals but is necessarily difficult to estimate in a short period of time.
Data Analysis for Rapid Assessment Because the type of data collected from mon-
itoring populations for immedia te and secondary effects o f harvesting will vary depending on what life stages are harvested, it is more difficult to generalize about how to handle these data and analyze them. However, both mortal i ty and re- crui tment rates are common to all types of mon- itoring and the proper way to calculate them is described below.
Mortal i ty and recruitment rates should be cal- culated as an exponential decline or increase in the size of an initial population. Annual mor- tality rates for a single populat ion (mi), as percent per year, are calculated as:
mi = 100[ln(no/(no - di))/ti]
where no is the number of trees at the first enu- meration, d i is the number o f trees that died during an interval and ti is the number o f years between enumerations. Therefore no - d i = ni,

1993] HALL & BAWA: IMPACT OF EXTRACTION 243
the number o f trees surviving to the ith year. This formula gives rise to a posit ive value for mortal i ty so that higher posit ive values imply higher mortal i ty rates.
Recrui tment rates are computed in a manner analogous to mortal i ty rates. The number of re- cruits are computed as a proport ion of the num- ber of trees initially recorded in the t ime period of interest plus the addit ional number of trees recruited during that t ime period. Recrui tment rate for a populat ion is defined as:
r~ = 100[ln((no + a~)/no)/td
where no is the number o f trees at the first enu- meration, a~ is the number of recruits added to the populat ion at the t ime of the second enu- merat ion and t~ is the length of the t ime between intervals in years. Therefore no + ai = ni, the size of the populat ion after recruitment without con- sidering mortal i ty in the initial population. This formula gives rise to a positive recruitment rate so that higher values imply higher recruitment rates. Calculation of mortal i ty and recruitment rates as proport ional changes in populat ion size and as an exponential function allows for direct comparisons among widely varying life forms and sampling designs.
It is important to analyze the quanti tat ive data on reproductive output, health and growth as a function of harvesting intensity. Populations in undisturbed areas, which have effectively zero harvesting intensity, must be included in the analysis. Both plant size and harvesting intensity may effect reproductive output, health, and growth, so these measures must be considered simultaneously. For instance, i f it is found that the mean number of fruits produced by trees which are intensively harvested for latex is sig- nificantly lower than for trees which are not sub- jected to harvesting, this result may be only a correlated effect because the mean tree size of harvested individuals may be lower than for un- harvested individuals. These types of interac- tions further emphasize the importance of ob- taining samples from unharvested populations.
Long Term Assessment
Long term assessment is needed in order to recognize how harvesting and secondary effects of harvesting will eventually alter populat ion dy- namics. Some of the long term effects may be quite unexpected because of the complex inter- actions among species, both animal and plant,
and humans within a community . For instance, the constant presence o f humans harvesting in an area may drive out animals that are impor tant dispersers of the harvested species. Initially, adult reproduction and seedling recruitment may con- tinue unabated, but i f the seeds that were dis- persed away from adults, as occurred when an- imal populations were at natural densities, were the greatest source of populat ion regeneration it may take a number of years to notice the loss o f effective sapling establishment and eventual growth of these individuals into adult size classes which are the focus of harvesting.
The sampling design for long term monitor ing of populat ion dynamics will bear many similar- ities regardless of the t ime scale over which enu- meration will take place. Maximizing sample sizes will involve increasing the number of individuals followed over t ime and maintaining as nearly equal a sample size as possible among different habitats and harvesting intensities. The actual amount of area enumerated is of much less im- portance than obtaining sufficient numbers of marked individuals o f the full range of age/size classes in different sites. As the length of the study increases, it can become difficult to accurately distinguish individuals i f they are not clear and permanently marked. Mortal i ty and recruitment rates can only be accurately measured by per- manently marking individuals. Mortal i ty rates of tree species may be very hard to measure as these rates are often quite low once individuals have reached relatively small size (between 1 and 2% per year for trees over 8 cm dbh). Therefore, in order to detect such low rates, a min imum of 100 trees from each site and size class o f interest must be marked and followed over time. A larger sample size is much more desirable as a popu- lation o f only 100 individuals with a mortal i ty rate of 1% per year results in the average obser- vation of only 1 death a year. In order to measure comparat ive recruitment rates over long periods, care must be taken in searching the same area for recuits upon each enumeration. Therefore, the area used for assessing recruitment must also be clearly and permanently marked. Increasing or changing the area from which recruitment is measured will not result in comparable recruit- ment rates.
As with monitoring for rapid assessment, the t ime between enumerat ions must be adjusted to reflect the life history of the species under in- vestigation. Measuring juveni le mortal i ty and re-

244 ECONOMIC BOTANY [VOL. 47
cmitment , especially of young seedlings, will re- quire more frequent enumerat ion for accurate est imates o f temporal variat ion in these rates. Growth o f established juveniles, and the growth and mortal i ty of adults may not require such frequent enumerat ion as these may occur slowly. It is also impor tant to measure growth rates at the same t ime o f the year as seasonal changes in water availabil i ty may result in changes in bole size not directly related to the net addi t ion of wood and thus growth rate measurements will be inaccurate. When measurement o f the repro- ductive phenology of adults is required, the t ime between enumerations should be sufficiently short to allow accurate measurement of the periods of reproductive activity. For many species once a month may be sufficient, but initially observa- tions should be more frequent during a repro- ductive episode so that a better sampling design can be developed. Sample sizes for growth, mor- tality, and reproductive phenology may vary, too; although all three may be o f importance to the species under investigation, not all measure- ments need to be taken on all individuals. Sub- sampling can be done and stratified by the de- mands of total number needed, number of habitats within the study site, the number o f har- vesting intensities being investigated, and the variat ion in the parameter measured.
Methods for Establishing Permanent Plots Establishing permanent plots for long term
monitor ing must be done in a manner that will allow unambiguous relocation and measurement of individuals and accurate determinat ion o f mortali ty, recruitment, and growth. Plots should be permanently marked, preferably at all four comers so that their boundaries are unambigu- ous. Subplots established for the enumerat ion of smaller individuals should also be unambigu- ously marked. Individuals must be permanent ly marked. A common practice is to use a luminum tags and nails on larger individuals (> 10 cm dbh) and small gauge wire for smaller individuals. Nursery tags or small metal tags on wire can be used for seedlings. Nails should not be placed at the point of measurement as deformation at the point o f entry often occurs and will interfere with future measurements o f diameter increment. For large trees with buttresses, it is much more con- venient to locate a tree when the tag is at eye level. Wire placed around smaller individuals should be sufficiently large to allow for growth
for many years so that reenumerat ion will not always include retagging all individuals. Precise maps need not be created, but the recording of tree posi t ion must be sufficient to allow accurate identification of recruits, particularly for the smaller size classes.
The point of measurement of dbh should be established at 1.3 m from the high ground side o f each plant. For plants with deformities, in- cluding buttresses, the point of measurement should be established at a fixed height above the deformity, usually at least 60 cm. The same height should be used for all types of deformity. Points of measurement should also be marked so that they can be relocated. The most convenient method is to paint the point o f measurement such that the upper edge o f the paint is where the tape is placed. Metal tapes should be used to measure diameter and the same units o f mea- surement should be used for each enumerat ion so that field measurements can be immedia te ly checked without the need o f calculation. Mea- surements should be done as accurately as pos- sible; there is no need to round off a number i f an exact value can be read. Field crews should ensure their precision of measurement by re- peatedly measuring the same tree on subsequent days until there is as little measurement error as feasible. Any categorical information, such as classification of light condit ions of individuals, quanti ty o f material harvested, or description o f habitat, must be not only clearly defined but also calibrated with each enumerat ion crew. Such ap- parently subjective data can be very useful and reliably collected i f sufficient care is taken to stan- dardize the categories both in written format and in the field. The initial enumerat ion will be the most t ime-consuming, but i f this enumerat ion is done with care and precision, all subsequent enu- merations will be faster and more accurate.
All information recorded should be tran- scribed as soon as possible into a format that allows for long term storage. Clear and detai led documentat ion of the keys of all coded variables and methods ofcoUecting that were used should accompany the data from all enumerations. I f at all possible, information should be scrutinized while still in the field and corrected since it is nearly impossible to correct field errors in the office. The last enumerat ion should always be brought to the field for the next enumerat ion. This information should not be used to bias new measurements, but without a list o f what the

1993] HALL & BAWA: IMPACT OF EXTRACTION 245
crew is expected to encounter, many mistakes can occur.
In principle, there is no problem with modi- fying sampling designs as newly collected infor- mation dictates. For instance, gradual loss of the establishment of saplings may only be recognized after years of enumeration. At this point more spatial information may be useful to understand how harvesting is affecting sapling establish- ment. Experiments which examine the relation- ship between density and distance dependent re- cruitment of seedlings and establishment o f saplings could be initiated. These experiments will require increasing sampling sizes to accom- modate new experimental protocols. However, if any changes in enumeration methodology are made, it is important to address how the changes affect the past collection of information and how to "map" the new onto the older information. What currently does not appear important may become essential in years to come. At the same time, in the interest of efficiency and rapidity, limiting sample sizes or abandoning certain enu- merations can be done to assure that sufficient numbers of different sites are studied. The strength of the comparative method is in the collection o f data in the same manner from many sites. The value of long term data increases with time.
Data Analysis for Long Term Assessment Many of the same techniques used to analyze
rapid assessment data are appropriate for long term assessment. However, temporal trends or variability will only become apparent after many years of enumeration yet may be of the greatest importance in determining what constitute sus- tainable harvests. It is important to establish a set of standard analyses that are performed after each enumeration. For instance, mortality and recruitment rates for size classes, habitats, and harvest intensities should be computed and graphed. Growth and reproductive rates as a function o f plant size, habitat, and harvest in- tensities should be graphed. Keeping records of these standard analyses, computed and displayed in the same manner, will allow recognition of subtle changes in population dynamics that can occur as a result of harvesting.
Rapid assessment of a population in which material from adults is being harvested may re- sult in an age/size distribution as show in Fig. 1B. However, with long term data, the popula-
tion may begin to shift towards the distribution illustrated in Fig. I C. I f such graphs are made with each enumeration, using the same scales and division of age/size classes, the investigator will be able to detect shifts in distribution and determine the rate at which they are occurring.
Data acquired from long term assessments will be composed of counts and frequencies o f indi- viduals in age/size classes within habitats, har- vest intensities, etc. Statistical methods such as log-linear analysis will help determine patterns of population dynamics and can be used to com- pare differences among sites, replicates or treat- ments. Log-linear analysis can also be used even when observations do not include marked fol- lowed individuals and therefore can allow in- vestigation of older perhaps incomplete data sets that may still provide useful information and help shape current sampling regimes.
One of the most useful applications o f long term assessment data of marked individuals is the construction of population matrix models which can directly assess population stability. This assessment is accomplished by computa- tion of age/size class specific survivorship, fe- cundity and transition rates from one age/size class to the next (Caswell 1989). These models can be used to investigate how population age/ size structure may change when subjected to dif- ferent harvesting or management regimes and help determine the most critical life history stage of the population. Hence, they have immediate predictive power. They can also be amended to incorporate new data that is continuously ac- quired from long term assessment, thereby im- proving the description of population dynamics and the predictive power o f the models.
LIMITATIONS OF METHODS
There are several caveats to the methods de- scribed in the preceding section. First, the meth- ods outlined are intended to monitor the effects of harvest on the short and long term fate of single-species populations. However, in the ab- sence of strict controls, a situation not unusual in tropical field ecology, any adverse effects on reproduction, regeneration or population struc- ture cannot be directly attributed to extraction. Population decline, in particular, can occur due to a number of biotic as well as abiotic causes (see below).
A second caveat is that population dynamics of a particular species varies over time and space.

246 ECONOMIC BOTANY [VOL. 47
As a result the models described here are not predictive and lack generalization. Caution must be exercised in attributing similar changes in de- mographic parameters of populations at different sites to the same causal agent.
Third, population dynamics of a given species is influenced by other species in the community. In tropical forests, apart from competition and predators, mutualists play a critical role in the life cycles of plants. Thus, changes in populations of animals that act as pollinators, seed dispersal agents and herbivores, as argued earlier, can in- fluence the population dynamics of plant species as well as several communi ty level processes. Thus, the observed changes in population of spe- cies may not be a direct consequence of extrac- tion of that particular species.
Finally, in most tropical forests, many species of plants and animals are harvested simulta- neously. Thus communi ty level processes may be changing without a detectable short term effect on the population dynamics of individual spe- cies. Conversely, changes in population dynam- ics of a given species may have a negligible effect on the overall structure, composition and dy- namics of the community.
CONCLUDING REMARKS
We have proposed methods to assess the im- pact of harvesting NTFP on the population dy- namics of the target species. The use of methods described here might provide insights into the impact of harvesting practices on depletion of extracted populations, but these methods them- selves do not provide prescriptions to remedy situations where NTFP have been over-exploit- ed. Unfortunately, evidence suggests that ex- traction is non-sustainable in many tropical ar- eas. Once the over-exploitation has been detected, the area may be allowed to recover or other man- agement interventions made, depending upon the severity of depletion and the overall degradation of the habitat.
The utility of our methods to assess sustain- ability will also depend upon management goals. For example, if areas from which NTFP are ex- tracted are large, and if the products are extracted on a rotational basis, depleted populations can recover between rotation cycles. In small areas, in the absence of rotation cycles, intensive sam- pling to assess sustainability for continued yields would be unnecessary. At the moment, however, there are few, if at all any sites where manage-
ment goals for the sustained extraction of NTFP are known or specified.
Finally, ecological sustainability cannot be considered in isolation. Economic considera- tions have and will remain paramount in land use patterns. Increasing commercialization of NTFP may lead to total depletion or domesti- cation. In the latter case, the issue of ecological sustainability becomes a moot point except in cases where individuals with access to the com- mons continue to exploit NTFP. Conversely, ecological conditions may be such that the eco- nomic exploitation is infeasible. Thus, once again the methods proposed here may serve as a di- agnostic tool that might be integrated into either a conservation or a management plan.
ACKNOWLEDGMENTS
We thank Elizabeth Taylor for her suggestions to improve the manu- script. This research has been supported by grants from the MacArthur Foundation, Pew Charitable Trust's Pew Scholars Program in Conser- vation and the Environment, and Conservation, Food and Health Foun- dation.
LITERATURE CITED
Browder, J. O. 1992. Social and economic con- straints on the development of market-oriented ex- tractive reserves in Amazon rain forests. Pages 33- 42 in D. C. Nepstad and S. Schwartzman, eds., Non-timber products from tropical forests: evalu- ation of a conservation and development strategy. Advances in Economic Botany 9. The New York Botanical Garden, Bronx, NY.
Caswell, H. 1989. Matrix population models. Sin- auer Associates, Inc. Publishers, Sunderland, MA.
Chambers, R. C., N. C. Saxena, and T. Shah. 1989. To the hands of the poor: water and trees. Westview Press, Boulder, CO. 273 pp.
Fernandes, W., G. Menon, and P. Viegas. 1988. For- ests, environment and tribal economy: deforesta- tion, impoverishment and marginalisation in Oris- sa. Indian Social Institute. Tribes of India Series 2. Indian Social Institute, New Delhi. 363 pp.
Greig-Smith, P. 1957. Quantitative plant ecology. Academic Press, New York.
Homma, A. K.O. 1992. The dynamics of extraction in Amazonia: a historical perspective. Pages 23-31 in D. C. Nepstad and S. Schwartzman, eds., Non- timber products from tropical forests: evaluation of a conservation and development strategy. Ad- vances in Economic Botany 9. The New York Bo- tanical Garden, Bronx, NY.
Kershaw, K.A. 1973. Quantitative and dynamic plant ecology. Second edition. Edward Arnold, London.
Mueller-Dombois, D., and H. Ellenberg. 1974. Aims

1993] HALL & BAWA: IMPACT OF EXTRACTION 247
and methods of vegetation ecology. John Wiley and Sons, New York.
Nepstad, D. C., L F. Brown, L. Luz, A. Alechandra, and V. Viana. 1992. Biotic impoverishment of Amazonian forests by tappers, loggers and cattle ranchers. Pages 1-14 in D. C. Nepstad and S. Schwartzman, eds., Non-timber products from tropical forests: evaluation of a conservation and development strategy. Advances in Economic Bot- any 9. The New York Botanical Garden, Bronx, NY.
Reining, C. C. S., R. M. Heinzman, M. C. Madrid, S. L6pez, and A. Sol6rzano. 1991. The non-timber forest product industries of the Maya Biosphere Reserve, Guatemala: results of ecological and so- cio-economic surveys. Unpublished report. Con- servation International Foundation, Washington, DC.





























