Embed Size (px)
Citation preview

CELL BIOLOGY AND MORPHOGENESIS
Micropropagation of ornamental Prunus spp. and GF305 peach,a Prunus viral indicator
Anna Kalinina Æ Daniel C. W. Brown
Received: 26 July 2006 / Revised: 23 December 2006 / Accepted: 24 January 2007 / Published online: 24 February 2007
� Springer-Verlag 2007
Abstract A micropropagation approach was developed
for nine ornamental Prunus species, P. americana, P. cis-
tena, P. glandulosa, P. serrulata ‘Kwanzan’, P. laurocer-
asus, P. sargentii, P. tomentosa, P. triloba, P. virginiana
‘Schubert’, commercially important in North America, and
GF305 peach, commonly used for Prunus virus indexing.
The micropropagation cycle based on proliferation of
vegetative tissues includes establishment of tissue culture
through introduction of shoot meristems in vitro, shoot
proliferation, root induction and plant acclimatization steps
and can be completed in 5 months. A meristem steriliza-
tion protocol minimized bacterial and fungal contamina-
tion. Multiple shoot formation in ornamental Prunus was
obtained through the use of 1 mg l–1 6-benzyladenine. For
GF305 peach, alteration in the sugar composition, fructose
instead of sucrose, and addition of 1 mg l–1 ferulic acid had
a significant impact on the shoot proliferation rate and
maintenance of long-term in vitro culture. Rooting and
plant acclimatization conditions were improved using a
two-step protocol with a 4-day root induction in indole-3-
butiric acid (IBA)-containing media with consequent 3-
week root elongation in IBA-free media. One-month
incubation of rooted shoots in a vermiculite-based medium
resulted in additional shoot and root growth and provided
better acclimatization and plant recovery. The microprop-
agation approach can be used for maintenance of the clonal
properties for Prunus spp. as well as a protocol to support
meristem therapy against viral infection.
Keywords P. americana � P. cistena � P. glandulosa �P. serrulata ‘Kwanzan’ � P. laurocerasus � P. sargentii �P. tomentosa � P. triloba � P. virginiana ‘Schubert’ �Virus indexing � Meristem therapy
Abbreviations
BA 6-Benzyladenine
IBA Indole-3-butiric acid
GA Gibberellic acid
TDZ 1-phenyl-3-(1,2,3-thiadiazol-5-yl)urea
(thidiazuron)
NAA a-Naphthaleneacetic acid
FA Ferulic acid (trans-4-hydroxy-3-methoxycinnamic
acid)
MES 2-(N-Morpholino)-ethanesulphonic acid
MS Murashige and Skoog (1962) basic salt medium
(PhytoTechnology Laboratory, KS, USA)
QL Quoirin and Lepoivre (1977) basic salt medium
(PhytoTechnology Laboratory, KS, USA)
SAM Shoot apical meristem
AxM Axillary shoot meristem
MP Meristem proliferation medium
SP Shoot proliferation medium
RM Rooting medium
Introduction
The Prunus genus includes more than 200 species of
deciduous or evergreen trees and shrubs widely distributed
in temperate zones and are valued for their ornamental
Communicated by R.J. Rose.
A. Kalinina � D. C. W. Brown (&)
Southern Crop Protection and Food Research Centre,
Agriculture and Agri-Food Canada, 1391 Sandford Street,
London, ON N5V 4T3, Canada
e-mail: [email protected]
A. Kalinina
e-mail: [email protected]
123
Plant Cell Rep (2007) 26:927–935
DOI 10.1007/s00299-007-0315-x

features. Ornamental Prunus species, such as P. americana,
P. glandulosa, P. tomentosa and P. triloba, are grown for
their white, pink or red flowers, saucer-, bowl- or cup-shaped
with five petals, often in semi-double or double forms.
Some, such as P. serrulata ‘Kwanzan’ and P. sargentii, have
shiny and colourful bark and beautiful autumn leaf colour;
others, such as P. x cistena and P. virginiana ‘Schubert’,
have attractive purple or green waxy foliage. Dense, bushy
species, such as P. laurocerasus, are useful for groundcover
and green fences. The commercial importance of most or-
namental Prunus in the North American climate is based on
their attractive appearance, extensive vegetation, unpreten-
tious growth conditions and high adaptation abilities. GF305
peach (P. persica ‘GF305’) is commonly used for grafting
different commercially important peach varieties as well as
an indicator plant for virus indexing (Bernhard and Mare-
naud 1969; Gentit et al. 1998). More than 30 years before
establishment of molecular biology approaches the only way
to identify and characterize several degenerating Prunus
virus and virus-like diseases, such as Plum pox potyvirus,
Apple chlorotic leaf spot trichovirus, Prune dwarf ilavirus,
Prunus necrotic ring spot ilavirus, Tomato ring spot nepo-
virus, Peach latent mosaic viroid, European stone fruit
yellow phytoplasma and Peach asteroid spot agent, was
based on the symptom analysis of GF305 peach seedlings
(Desvignes 1976; EPPO Panel on Certification of Fruit
Crops 1992).
Propagation of most Prunus spp. and selection of new
cultivars based on traditional crossing, seed and clonal (stem
cuttings) propagation are time and labour consuming and
often applicable for only a few species (Matt and Jehle
2005). Moreover, most ornamental Prunus do not produce
pollen or viable generative organs. Micropropagation tech-
niques for sour and sweet cherries (Borkowska 1983;
Hammat and Grant 1996; Matt and Jehle 2005), cherry and
peach rootstocks (Dalzotto and Docampo 1997; Marino
1997; Radice et al. 1999), plum (Mante et al. 1989, 1991;
Gonzalez Padilla et al. 2003) and peach (Hammerschlag
et al. 1987) have been described. There are only a few re-
ports on ornamental Prunus spp., P. tomentosa, P. fructi-
cosa, P. virginiana ‘Carrington’ and P. pensylvanica (e.g.
Pruski et al. 2000, 2005). Protocols for P. tomentosa and P.
virginiana ‘Carrington’ described a short-term (3-month)
tissue culture maintenance, terminated by rooting and had
low plant recovery efficiency. Most protocols are optimized
only for particular cultivar, and it is not clear if they are
applicable to other cultivars or species. Micropropagation of
other commercially used ornamental species, such as P.
americana, P.x cistena, P. glandulosa, P. serrulata
‘Kwanzan’, P. laurocerasus, P. sargentii and P. triloba,
have not yet been reported. There are a few reports on
propagation of GF305 peach confirming that it was a re-
calcitrant species under in vitro conditions, allowing only 2–
3 months of subculture (Lansac et al. 1998). The purpose of
this investigation was to develop a reliable system for mi-
cropropagation of nine ornamental Prunus spp., which have
a high commercial importance in North America and GF305
peach, which is a well-known species used as an indicator of
Prunus diseases for virus-free certification of plant material.
Materials and methods
Plant material
Nine ornamental Prunus cultivars, P. americana Marshall
(American plum), P.x cistena Koehne (P. cerasifera
‘Atropurpurea’ · P. pumila) (Purpleleaf sand cherry), P.
glandulosa Thunb. (Flowering almond), P. serrulata Lindl.
‘Kwanzan’, P. laurocerasus L. (Cherry laurel), P. sargentii
Rehder (Sargent cherry), P. tomentosa Thunb. (Nanking
cherry), P. triloba Lindl. (Flowering almond) and P. vir-
giniana L. ‘Schubert’ (Choke cherry) (Encyclopaedia of
Garden Plants 1997), obtained from nurseries in the
Niagara region (Ontario, Canada), and seeds of P. persica
L. (Batsch) ‘GF305’, obtained from the INRA research
station in Bordeaux, France, were used for establishing
in vitro cultures.
Establishment of Prunus in vitro culture
Shoot apical meristems (SAM) derived from greenhouse-
grown 1-year-old trees of P. americana, P.x cistena
(Fig. 1a), P. glandulosa, P. tomentosa, P. triloba, P. vir-
giniana ‘Schubert’ and GF305 peach (Fig. 1g), and axillary
shoot meristems (AxM) obtained from branches of field-
grown trees of P. serrulata ‘Kwanzan’, P. sargentii and P.
laurocerasus were used to establish in vitro cultures. Wood
sticks were forced to shoot using a bud-breaking medium
based on the half-strength MS basic salt medium (Mu-
rashige and Skoog 1962; PhytoTechnology Laboratory,
KS, USA), supplemented with 6-benzyladenine (BA),
indole-3-butiric acid (IBA) and gibberellic acid (GA)
(Table 1). Shoot tip explants with several leaf primordia
2–5 mm long were surface sterilised using 70% ethanol for
30 s followed by immersion in 15% Bleach and 0.02%
Tween-20 solution on a shaker for 15 min and then washed
with sterile distilled water. Explants were placed on mer-
istem proliferation media (MP1-5, Table 2). After 4 weeks,
explants were transferred into shoot proliferation media
(SP1 or SP2, Table 2) and subcultured each month.
Rooting in vitro and plant acclimatization
Shoot explants obtained from proliferating 4-week-old
Prunus tissue culture, 1–2 cm long, were placed in a
928 Plant Cell Rep (2007) 26:927–935
123

vertical position into different rooting media (RM1-8, Ta-
ble 3) and incubated for 4 weeks under three conditions
with different duration periods (Table 4). Experiment 1
included shoot incubation in RM1 or RM5 for 28 days;
experiment 2 included a 4-day incubation in RM1 or RM5
and subsequent transfer of shoots into RM2 or RM6;
experiment 3 included a 4-day incubation in RM3 or RM7
and subsequent transfer of shoots into RM4 or RM8.
Successfully rooted explants were transferred into a ver-
miculite-based medium (medium size vermiculite, Therm-
O-Rock East Inc., PA, USA) containing 1/2 MS salts and
vitamins (see vitamins used in the Table 1) without sugar,
growth regulators and Bactoagar, dispensed in Magenta
vessels and incubated for an additional month to adapt
newly formed roots to soil conditions. After 1 month,
plants were transferred into pots with ProMix ‘BX’ soil
mixture (ProMix ‘BX’, Premier Horticulture Inc., Canada),
placed in a growth chamber (+25 ± 2�C, 16 h photoperiod,
800–850 lmol m–2 s–1, 75% humidity), continuously irri-
gated with 1/5 MS salts solution and maintained in the light
chamber for an additional month before transfer into the
greenhouse.
Media and growth conditions
All solid media consisted of 0.8% Bactoagar (Fisher, USA)
and 0.5 g l–1 MES (2-(N-Morpholino)-ethanesulphonic
acid); pH was adjusted to 5.7–5.8 by 1 M KOH before
autoclaving. For GF305 peach shoot proliferating culture,
instead of MS, QL basic salt medium (Quoirin and Le-
poivre 1977; PhytoTechnology Laboratory, KS, USA) was
used supplemented with 15 g l–1 fructose and 1 mg l–1
ferulic acid (trans-4hydroxy-3-methoxycinnamic acid, FA)
(Tables 2, 3). Vitamins and growth regulators were filter-
sterilized and added to the autoclaved media when re-
quired. Media were dispensed into 100 · 25-mm Petri
plates (40 ml per plate) or Magenta vessels (Magenta,
Chicago, Il, USA), 60 ml per vessel when specified. Tis-
sues were grown in a culture room at +25 ± 2�C, 16 h light
photoperiod with light provided by full-spectrum fluores-
cent lamps (General Electric, ‘‘Starcoat’’, F32T8/SP41,
Canada) at an intensity of 300–350 lmol m–2 s–1. All
chemical products were purchased from Sigma, USA, ex-
cept when specified.
Results
Establishment of in vitro meristem proliferation system
for ornamental Prunus and GF305 peach
The SAM and AxM sterilization protocol reduced the
frequency of bacterial or fungal contamination of green-
house and field-grown material to a very low level (we
estimated less than 1% visual culture contamination). SAM
of P.x cistena (Fig. 1b) and P. triloba had a high prolif-
eration rate in MP1 containing 0.5 mg l–1 BA, 0.5 mg l–1
IBA and 2 mg l–1 GA, respectively 94 and 66% (Table 5).
SAM and AxM of P. serrulata ‘Kwanzan’, P. americana
and P. virginiana ‘Schubert’ proliferated in MP2 containing
Fig. 1 P.x cistena (a-f) and
GF305 peach (g-l)micropropagation system. a, gA shoot tip with SAM. b, hSAM proliferation in tissue
culture. c, i Multiple shoot
formation. d, j Root induction.
e, k Plant acclimatization in
Magenta boxes. f, l Recovered
plant. Bars: 1 cm
Table 1 Medium used for excised dormant Prunus bud-wood
Components Concentration (mg l–1)
MS salts 1/2
Vitamins
Glycine 2.0
Myo-inositol 100
Nicotinic acid 0.5
Pyridoxine 0.5
Thiamin-HCl 1.0
Ascorbic acid 2.0
Growth regulators
BA 2.0
IBA 0.1
GA 5.0
MS Macro and microelements from Murashige and Skoog (1962)
(PhytoTechnology Laboratory, KS, USA)
Plant Cell Rep (2007) 26:927–935 929
123
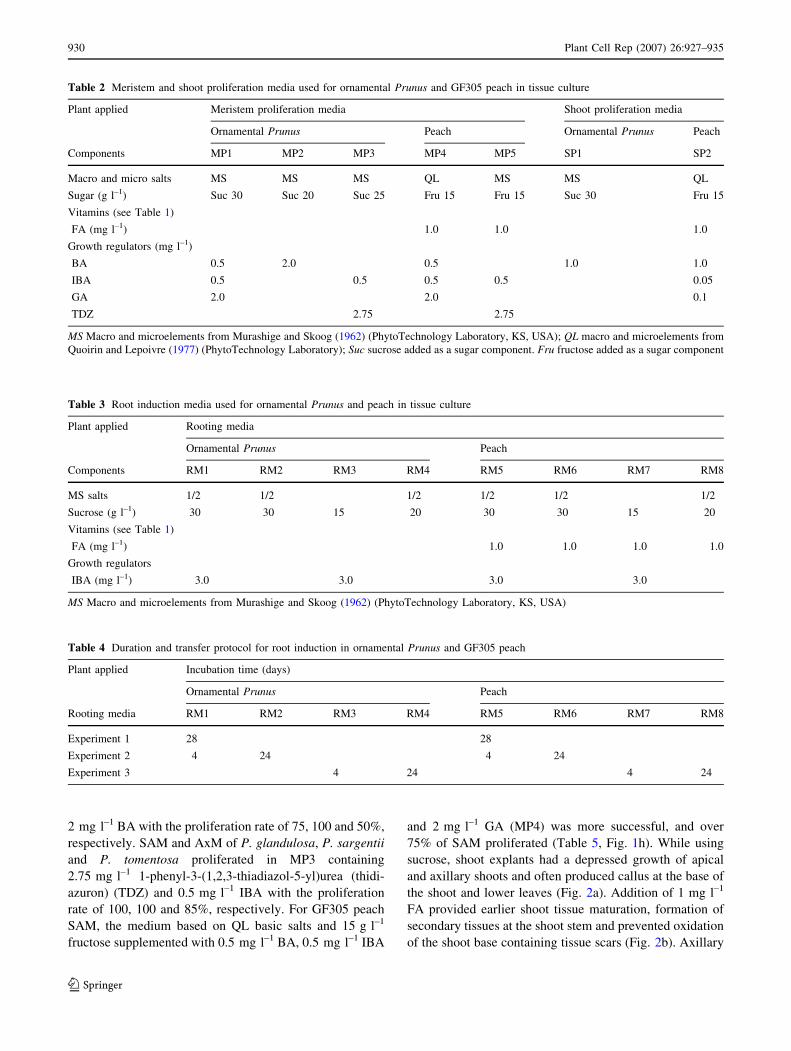
2 mg l–1 BA with the proliferation rate of 75, 100 and 50%,
respectively. SAM and AxM of P. glandulosa, P. sargentii
and P. tomentosa proliferated in MP3 containing
2.75 mg l–1 1-phenyl-3-(1,2,3-thiadiazol-5-yl)urea (thidi-
azuron) (TDZ) and 0.5 mg l–1 IBA with the proliferation
rate of 100, 100 and 85%, respectively. For GF305 peach
SAM, the medium based on QL basic salts and 15 g l–1
fructose supplemented with 0.5 mg l–1 BA, 0.5 mg l–1 IBA
and 2 mg l–1 GA (MP4) was more successful, and over
75% of SAM proliferated (Table 5, Fig. 1h). While using
sucrose, shoot explants had a depressed growth of apical
and axillary shoots and often produced callus at the base of
the shoot and lower leaves (Fig. 2a). Addition of 1 mg l–1
FA provided earlier shoot tissue maturation, formation of
secondary tissues at the shoot stem and prevented oxidation
of the shoot base containing tissue scars (Fig. 2b). Axillary
Table 2 Meristem and shoot proliferation media used for ornamental Prunus and GF305 peach in tissue culture
Plant applied Meristem proliferation media Shoot proliferation media
Ornamental Prunus Peach Ornamental Prunus Peach
Components MP1 MP2 MP3 MP4 MP5 SP1 SP2
Macro and micro salts MS MS MS QL MS MS QL
Sugar (g l–1) Suc 30 Suc 20 Suc 25 Fru 15 Fru 15 Suc 30 Fru 15
Vitamins (see Table 1)
FA (mg l–1) 1.0 1.0 1.0
Growth regulators (mg l–1)
BA 0.5 2.0 0.5 1.0 1.0
IBA 0.5 0.5 0.5 0.5 0.05
GA 2.0 2.0 0.1
TDZ 2.75 2.75
MS Macro and microelements from Murashige and Skoog (1962) (PhytoTechnology Laboratory, KS, USA); QL macro and microelements from
Quoirin and Lepoivre (1977) (PhytoTechnology Laboratory); Suc sucrose added as a sugar component. Fru fructose added as a sugar component
Table 3 Root induction media used for ornamental Prunus and peach in tissue culture
Plant applied Rooting media
Ornamental Prunus Peach
Components RM1 RM2 RM3 RM4 RM5 RM6 RM7 RM8
MS salts 1/2 1/2 1/2 1/2 1/2 1/2
Sucrose (g l–1) 30 30 15 20 30 30 15 20
Vitamins (see Table 1)
FA (mg l–1) 1.0 1.0 1.0 1.0
Growth regulators
IBA (mg l–1) 3.0 3.0 3.0 3.0
MS Macro and microelements from Murashige and Skoog (1962) (PhytoTechnology Laboratory, KS, USA)
Table 4 Duration and transfer protocol for root induction in ornamental Prunus and GF305 peach
Plant applied Incubation time (days)
Ornamental Prunus Peach
Rooting media RM1 RM2 RM3 RM4 RM5 RM6 RM7 RM8
Experiment 1 28 28
Experiment 2 4 24 4 24
Experiment 3 4 24 4 24
930 Plant Cell Rep (2007) 26:927–935
123

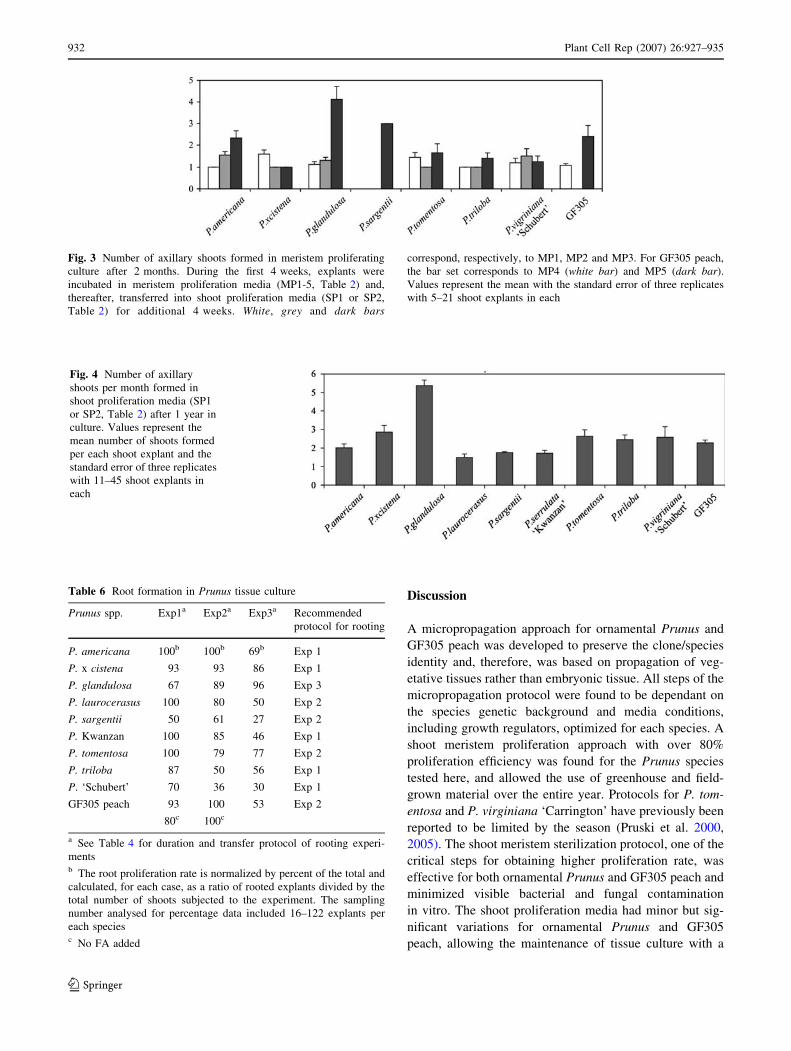
shoots were formed on proliferated shoot explants only
during the second passage on proliferation media SP1 and
SP2, respectively, for ornamental Prunus and peach (Ta-
ble 2). Particularly, P. glandulosa shoot explants had a
high axillary shoot formation rate of 4.1 ± 0.6 shoots per
explant within first 2 months (Fig. 3), whereas, the rest of
the species required additional subculturing on SP1 or SP2
(Fig. 1c, i). The number of axillary shoots formed per ex-
plant per month was evaluated within the first and the
second year after establishment of shoot meristem culture
and was stable for all investigated Prunus spp. (Fig. 4).
The highest rate of axillary shoot formation per month,
5.37 ± 0.3 shoots per explant, was observed in P. glan-
dulosa and less so, 2.01 ± 0.2 to 2.65 ± 0.6, in P. ameri-
cana, P. x cistena, P. tomentosa, P. triloba, P. virginiana
‘Schubert’ and GF305 peach. Whereas, in P. serrulata
‘Kwanzan’, P. laurocerasus and P. sargentii the rate was
within 1.5 ± 0.2 to 1.8 ± 0.2 (Fig. 4).
Root induction and plant recovery in ornamental
Prunus and GF305 peach
Several different media (RM1-8, Table 3) and approaches
(Experiments 1–3, Table 4) were applied to optimize a
rooting in vitro protocol for ornamental Prunus spp. and
GF305 peach. Successful root induction and elongation
was obtained in experiment 1 for P. americana, P.x cistena
(Fig. 1d), P. serrulata ‘Kwanzan’, P. triloba and P. vir-
giniana ‘Schubert’ using RM1 medium containing 3 mg l–1
IBA (experiment 1, Table 6). In P. laurocerasus, P. sar-
gentii and P. tomentosa, roots formed after a 4-day
induction in RM1 and a 3-week elongation in RM2 without
IBA (experiment 2). For P. glandulosa, roots successfully
formed using a 4-day induction in RM3, MS salts-free
medium contained 3 mg l–1 IBA, followed by root elon-
gation in RM4 without IBA (as outlined for experiment 3).
GF305 peach formed roots with a high efficiency (Fig. 1j)
with conditions used in experiment 2, i.e. a 4-day root
induction period in RM5 with 3 mg l–1 IBA, 1 mg l–1 FA
and 30 g l–1 sucrose instead of fructose and a subsequent
root elongation in RM6 without IBA (Table 6). The num-
ber of roots formed per shoot explant and the timing of root
formation was strongly dependant on the species and media
(Fig. 5). In general, roots appeared between 8 and 16 days
post induction. Early rooting species were P. americana,
P.x cistena, P. tomentosa, P. triloba, P. virginiana
‘Schubert’ and GF305 peach. With some delay, roots were
formed by P. serrulata ‘Kwanzan’, P. sargentii and
P. laurocerasus. Mostly, these latter species formed callus
at the explant base before root initiation.
Acclimatization to greenhouse conditions was based
on a two-step protocol. Firstly, rooted microshoots were
transferred into a vermiculite-based medium in Magenta
vessels to adapt roots to soil conditions for 1 month
(Fig. 1e, k). Then plantlets were exposed briefly to
ambient air conditions in the tissue culture room and
subsequently moved into pots. The survival rate was
generally high and species dependant (Table 7). P.x cis-
tena, P. laurocerasus, P. sargentii, P. serrulata ‘Kwan-
zan’, P. tomentosa and GF305 peach had more than 80%
survival rate in soil. In contrast, P. americana, P. glan-
dulosa, P. triloba and P. virginiana ‘Schubert’ were af-
fected by low humidity and had a reduced efficiency.
After 1–3 months cultivation in soil, plants produced
axillary shoots and had the same morphological tree-like
structure including an apical dominance growth pattern
and axillary branches (Fig. 1f, l).
Table 5 Meristem proliferation rate after 1 month incubation
Prunus spp. MP1 MP2 MP3 Recommended medium
for meristem proliferation
P. americana 25a 100 30 MP2
P. x cistena 94 13 29 MP1
P. glandulosa 37 76 100 MP3
P. laurocerasus NA NA NA MP1
P. sargentii 0 0 100 MP3
P. ‘Kwanzan’ 48 75 21 MP2
P. tomentosa 80 28 85 MP3
P. triloba 66 20 3 MP1
P. ‘Schubert’ 45 50 23 MP2
MP4 MP5
GF305 peach 76 62 MP4
NA Not attempteda The shoot proliferation rate is normalized by percent of the total and
calculated, for each case, as a ratio of explants with shoots prolifer-
ated divided by the total number of explants subjected to the exper-
iment. The sampling number analysed for percentage data included
25–88 explants per each species
Fig. 2 Influence of sugar composition on growth and development of
GF305 shoots in tissue culture. a Incubation of explants in shoot
proliferation medium containing 30 g l–1 sucrose. b Multiple shoot
formation in SP2 medium containing 15 g l–1 fructose and 1 mg l–1
FA. Bars: 1 cm
Plant Cell Rep (2007) 26:927–935 931
123
Discussion
A micropropagation approach for ornamental Prunus and
GF305 peach was developed to preserve the clone/species
identity and, therefore, was based on propagation of veg-
etative tissues rather than embryonic tissue. All steps of the
micropropagation protocol were found to be dependant on
the species genetic background and media conditions,
including growth regulators, optimized for each species. A
shoot meristem proliferation approach with over 80%
proliferation efficiency was found for the Prunus species
tested here, and allowed the use of greenhouse and field-
grown material over the entire year. Protocols for P. tom-
entosa and P. virginiana ‘Carrington’ have previously been
reported to be limited by the season (Pruski et al. 2000,
2005). The shoot meristem sterilization protocol, one of the
critical steps for obtaining higher proliferation rate, was
effective for both ornamental Prunus and GF305 peach and
minimized visible bacterial and fungal contamination
in vitro. The shoot proliferation media had minor but sig-
nificant variations for ornamental Prunus and GF305
peach, allowing the maintenance of tissue culture with a
Fig. 3 Number of axillary shoots formed in meristem proliferating
culture after 2 months. During the first 4 weeks, explants were
incubated in meristem proliferation media (MP1-5, Table 2) and,
thereafter, transferred into shoot proliferation media (SP1 or SP2,
Table 2) for additional 4 weeks. White, grey and dark bars
correspond, respectively, to MP1, MP2 and MP3. For GF305 peach,
the bar set corresponds to MP4 (white bar) and MP5 (dark bar).
Values represent the mean with the standard error of three replicates
with 5–21 shoot explants in each
Fig. 4 Number of axillary
shoots per month formed in
shoot proliferation media (SP1
or SP2, Table 2) after 1 year in
culture. Values represent the
mean number of shoots formed
per each shoot explant and the
standard error of three replicates
with 11–45 shoot explants in
each
Table 6 Root formation in Prunus tissue culture
Prunus spp. Exp1a Exp2a Exp3a Recommended
protocol for rooting
P. americana 100b 100b 69b Exp 1
P. x cistena 93 93 86 Exp 1
P. glandulosa 67 89 96 Exp 3
P. laurocerasus 100 80 50 Exp 2
P. sargentii 50 61 27 Exp 2
P. Kwanzan 100 85 46 Exp 1
P. tomentosa 100 79 77 Exp 2
P. triloba 87 50 56 Exp 1
P. ‘Schubert’ 70 36 30 Exp 1
GF305 peach 93 100 53 Exp 2
80c 100c
a See Table 4 for duration and transfer protocol of rooting experi-
mentsb The root proliferation rate is normalized by percent of the total and
calculated, for each case, as a ratio of rooted explants divided by the
total number of shoots subjected to the experiment. The sampling
number analysed for percentage data included 16–122 explants per
each speciesc No FA added
932 Plant Cell Rep (2007) 26:927–935
123

stable proliferation rate over a 2-year period. Particularly,
alteration in the sugar composition, fructose instead of
sucrose, and addition of 1 mg l–1 FA prevented shoot
necrosis and withering and had a significant impact on the
shoot rate and survival of GF305 peach tissue culture. A
treatment with antioxidant compounds such as ascorbic
acid, citric acid, activated charcoal, FA, polyvinilpyrroli-
done and silver nitrate has been reported for several mi-
cropropagation systems (Komalavalli and Rao 2000;
Huang et al. 2002; Wen and Deng 2005). Formation of
morphologically well-developed axillary shoots was ob-
tained at the optimal BA concentration of 1 mg l–1. The
same BA level was also suggested for P. tomentosa and P.
virginiana ‘Carrington’ by Pruski et al. (2000, 2005).
Treatments with elevated BA concentrations and TDZ
promoted the shoot number per explant but decreased the
shoot length and negatively affected shoot development
(Pruski et al. 2000; Sarwar and Skrivin 1997; our obser-
vations).
Ex vitro root induction by application of 2 mg l–1 IBA
has been reported for some Prunus spp. such as P. tom-
entosa, P. fruticosa, P. virginiana ‘Carrington’ and P.
pensylvanica (Pruski et al. 2000, 2005). Our observations
confirmed this approach for P. glandulosa, P. tomentosa
and P. virginiana ‘Schubert’ (data not shown), whereas,
root induction in P. americana, P.x cistena, P. serrulata
‘Kwanzan’, P. sargentii, P. laurocerasus and P. triloba
was only achieved by use of an in vitro protocol. Most
Prunus spp. produced a high number of roots in IBA-
containing media, but roots were often shorter and thicker
and did not form lateral roots (data not shown). Rooting
was improved using the two-step protocol with a 4-day root
induction in IBA-containing media with a subsequent 3-
week root elongation in IBA-free media. As shoot height
and number of leaves had been reported to be critical
factors for acclimatization of plum plantlets (Gonzalez
Padilla et al. 2003), an additional 1-month incubation of
rooted shoots in the vermiculite-based medium provided
extensive shoot and root growth and resulted in better
acclimatization for most Prunus spp.
In summary, the micropropagation cycle based on pro-
liferation of vegetative tissues includes establishment of
tissue culture through introduction of shoot meristems
in vitro with reduced fungal and bacterial contamination,
meristem and shoot proliferation, root induction and plant
acclimatization steps (Fig. 6). The complete cycle from
introduction of meristems into tissue culture to recovery of
greenhouse grown plants requires about 5 months. The
overall efficiency for plant recovery during the minimum
time period required per each step depends on the genotype
and ranges from 2.38 to 0.51 (Table 8). The maximum
efficiency occurred for P. glandulosa and P.x cistena, 2.38
and 2.0, respectively; the second group including P. tom-
entosa, GF305 peach, P. laurocerasus and P. serrulata
Fig. 5 Numbers of adventitious roots formed in Prunus spp. after a
4-week incubation in root induction media (RM1-8, Table 3). White,
grey and dark bars correspond to experiment 1, experiment 2 and
experiment 3 (Table 4). For GF305 peach, the second bar set
corresponds to experiments 1 and 2 without FA in the media. Values
represent the mean number of roots formed per each shoot explant
with the standard error of three replicates with 5–22 shoot explants in
each
Table 7 The survival rate of Prunus tissue culture-derived plantlets
after rooting and acclimatization for greenhouse conditions
Prunus spp. Number of plants Survival rate
P. americana 38 29a
P. x cistena 50 80
P. glandulosa 67 46
P. laurocerasus 12 92
P. sargentii 14 79
P. ‘Kwanzan’ 33 85
P. tomentosa 60 84
P. triloba 22 57
P. ‘Schubert’ 38 57
GF305 peach 52 83
a The survival rate is normalized by percent of the total and calcu-
lated, for each case, as a ratio of rooted plants growing in ProMix soil
(ProMix’BX’, Premier Horticulture Inc., Canada) after 1 month di-
vided by the total number of plantlets subjected to the experiment
Plant Cell Rep (2007) 26:927–935 933
123

Fig. 6 Five-month Prunus micropropagation cycle. All stages of the
protocol are genotype dependent; the efficiency of meristem
proliferation ranges 0.5–1.0 explants surviving; shoot proliferation
ranges 1.5–5.4 shoots formed per explant; root induction ranges 0.6–
1.0 explants rooted and acclimatization ranges 0.3–0.9 plantlets
recovered. Bars: 1 cm
Table 8 The calculated efficiency of the micropropagation cycle based on the efficiency of each step of the protocol
Prunus spp. Micropropagation steps Total efficiency
Meristem proliferation Shoot proliferation Rooting Acclimatization
P. americana 1.0a 2.1b 1.0c 0.29d 0.61e
P. x cistena 0.94 2.86 0.93 0.8 2.0
P. glandulosa 1.0 5.37 0.96 0.46 2.38
P. laurocerasus NA 1.49 1.0 0.92 1.37
P. sargentii 1.0 1.76 0.61 0.79 0.85
P. ‘Kwanzan’ 0.75 1.72 1.0 0.85 1.10
P. tomentosa 0.85 2.64 1.0 0.84 1.88
P. triloba 0.66 2.45 0.87 0.57 0.8
P. ‘Schubert’ 0.5 2.58 0.7 0.57 0.51
GF305 peach 0.76 2.28 1.0 0.83 1.44
The efficiency of each step was calculated at the minimum time period used for each step, i.e meristem proliferation, shoot proliferation, rooting
and acclimatizationa Value calculated from Table 5 divided by 100%b Value taken from Fig. 4c Value calculated from Table 6 divided by 100%d Value calculated from Table 7 divided by 100%e Total efficiency is calculated by multiplication of the efficiency of each step of the protocol
934 Plant Cell Rep (2007) 26:927–935
123

‘Kwanzan’ had an efficiency of 1.1–1.9 and the third group
ranges from 0.5–0.8 for P. sargentii, P. triloba, P. ameri-
cana and P. virginiana ‘Schubert’. In addition, Prunus
tissue culture at the shoot proliferation stage can be ex-
tended for at least 2 years without visible changes in the
shoot formation rate and shoot development properties. An
extended shoot proliferation phase can result in an im-
proved plant multiplication rate. For example, in P. glan-
dulosa, if a 1-month proliferation phase was used then 2.38
plants are recovered, whereas, in a 3-month proliferation
phase up to 70 plants can be recovered. The micropropa-
gation approach could also be used for other applications
such as genetic transformation. The high rate of multiple
shoot proliferation in most Prunus spp. and fast root
induction protocols might be useful for micropropagation
and recovery of putative transformants. Another applica-
tion of the micropropagation cycle is that recovered orna-
mental Prunus plants could be used as part of a protocol for
self-rooted or grafted viral-free material in nurseries and
orchards. Manganaris et al. (2003) has showed that an
in vitro based meristem-tip culture/heat therapy protocol
was effective for virus elimination. The established pro-
tocols for tissue culture and plant recovery of P. glandul-
osa, P. tomentosa, P. serrulata ‘Kwanzan’ and GF305
peach plantlets might be useful for propagation of the plant
material for virus indexing in vitro culture, greenhouse or
field conditions.
Acknowledgments This work was supported by an NSERC re-
search fellowship for Canadian Government Laboratories for A.K.
The authors thank Paul Moote for technical assistance and Alex
Molnar for picture graphic design.
References
Bernhard R, Marenaud C (1969) Une methode plus sensible
d’indexage sur pecher. Phytopathol Mediterr II 3:209–218
Borkowska B (1983) Micropropagation of sour cherry culti-
var—Schattenmorelle. Fruit Sci Rep 10:59–66
Dalzotto A, Docampo DM (1997) Micropropagation of rootstock
from the Marianna-2624 plum (Prunus cerasifera x Prunusmunsoniana) and the pixy plum (P. insititia L.) under controlled
conditions. Phyton Int J Exp Bot 60:127–135
Desvignes C (1976) The virus diseases detected in greenhouse and in
field by the peach seedling GF305 indicator. Acta Hortic
67:315–322
Encyclopaedia of Garden Plants (1997) 1st edn. DK Publishing Inc.,
New York, pp 836–840
EPPO Panel on Certification of Fruit Crops. EPPO Council (EPPO/
EPPO) (1992)
Gentit P, Cornaggia D, Desvignes JC (1998) Identification and
comparison of different Prunus phytoplasma diseases by index-
ing on GF305 peach seedlings in the greenhouse. Acta Hortic
472:723–726
Gonzalez Padilla IM, Webb K, Scorza R (2003) Early antibiotic
selection and efficient rooting and acclimatization improve the
production of transgenic plum plants (Prunus domestica L.).
Plant Cell Rep 22:38–45. doi:10.1007/s00299-003-0648-z
Hammat N, Grant NJ (1996) Micropropagation of mature British wild
cherry. Plant Cell Tissue Organ Cult 47:103–110. doi:10.1007/
BF02318945
Hammerschlag FA, Bauchan GR, Scorza R (1987) Factors influenc-
ing in vitro multiplication and rooting of peach cultivars. Plant
Cell Tissue Organ Cult 8:235–242. doi:10.1007/BF00040950
Huang LC, Lee YL, Huang BL, Kuo CI, Shaw JF (2002) High
polyphenol oxidase activity and low titratable acidity in
browning bamboo tissue culture. In vitro Cell Dev Biol Plant
38:358–65
Komalavalli A, Rao M (2000) In vitro micropropagation of Gymnemasylvestre—a multipurpose medicinal plant. Plant Cell Tissue
Organ Cult 61:97–105. doi:10.1023/A:1006421228598
Lansac M, Chalak L, Cardona B, Sorbier A, Bodin-Ferri M, Dosba F,
Labonne G, Quiot L, Quiot JB (1998) In vitro inoculation of Prunusspecies with plum pox potyvirus. Acta Hortic 472:455–459
Manganaris GA, Economou AS, Boubourakas IN, Katis NI (2003)
Elimination of PPV and PNRSV through thermotherapy and
meristem-tip culture in nectarine. Plant Cell Rep 22:195–200.
doi:10.1007/s00299-003-0681-y
Mante S, Scorza R, Cordts JM (1989) Plant regeneration from
cotyledons of Prunus persica, Prunus domestica, and Prunuscerasus. Plant Cell Tissue Organ Cult 19:1–11. doi:10.1007/
BF00037771
Mante S, Morgens PH, Scorza R, Cordts JM, Callahan AM (1991)
Agrobacterium mediated transformation of plum (Prunusdomestica L.) hypocotyl slices and regeneration of transgenic
plants. Biotechnology 9:853–857
Marino G (1997) The influence of ethylene on in vitro rooting of GF-
677 (Prunus persica x Prunus amygdalis) hybrid peach root-
stock. In vitro Cell Dev Biol Plant 33:26–29
Matt A, Jehle JA (2005) In vitro plant regeneration from leaves and
internode sections of sweet cherry cultivars (Prunus avium L.).
Plant Cell Rep 24:486–476. doi:10.1007/s00299-005-0964-6
Murashige T, Skoog F (1962) A revised medium for rapid growth and
bioassay with tobacco tissue cultures. Physiol Plant 15:473–497
Pruski KW, Lewis T, Astatkie T, Nowak J (2000) Micropropagation
of Chokecherry (Prunus virginiana L.) and Pincherry (P.pensylvanica L.) cultivars. Plant Cell Tissue Organ Cult
63:93–100. doi:10.1023/A:1006461423373
Pruski KW, Astatkie T, Nowak J (2005) Tissue culture propagation of
Mongolian cherry (Prunus fruticosa) and Nanking cherry
(Prunus tomentosa). Plant Cell Tissue Organ Cult 82:207–211.
doi:10.1007/s11240-004-7836-6
Quoirin M, Lepoivre P (1977) Improved media for in vitro culture of
Prunus spp. Acta Hortic 78:437–442
Radice S, Perelman PE, Caso OH (1999) Clonal propagation of three
rootstocks of the genus Prunus for the ‘Flooding Pampa’. Phyton
Int J Exp Bot 64:149–156
Sarwar M, Skrivin RM (1997) Effect of thiadizuron and 6-
benzylaminopurine on adventitious shoot regeneration from
leaves of three strains of ‘McIntosh’ apple (Malus x domesticaBorkh) in vitro. Sci Hortic 68:95–100
Wen XP, Deng XX (2005) Micropropagation of chestnut rose (Rosaroxburghii Tratt) and assessment of the genetic stability in
in vitro plants using RAPD and AFLP markers. J Hortic Sci
Biotechnol 80:54–60
Plant Cell Rep (2007) 26:927–935 935
123







![ECPGR Priority Descriptors for Peach · DRAFT VERSION 1 - AUGUST 2013 ECPGR Priority Descriptors for Peach [Prunus persica (L.) Batsch] Daniela Giovannini1, Alessandro Liverani1,](https://img.pdfslide.net/doc/110x75/5c658aac09d3f28c6e8cee11/ecpgr-priority-descriptors-for-draft-version-1-august-2013-ecpgr-priority.jpg)

![ECPGR Priority Descriptors for Peach...DRAFT VERSION 1 - AUGUST 2013 ECPGR Priority Descriptors for Peach [Prunus persica (L.) Batsch] Daniela Giovannini1, Alessandro Liverani1, Daniele](https://img.pdfslide.net/doc/110x75/5e6219720040950705465a26/ecpgr-priority-descriptors-for-peach-draft-version-1-august-2013-ecpgr-priority.jpg)



















