Embed Size (px)
Citation preview

MicroRNA-101 Regulates Expression of the v-fos FBJMurine Osteosarcoma Viral Oncogene Homolog (FOS)
Oncogene in Human Hepatocellular CarcinomaShuai Li,1* Hanjiang Fu,1* Yulan Wang,2 Yi Tie,1 Ruiyun Xing,1 Jie Zhu,1 Zhixian Sun,1 Lixin Wei,2 and Xiaofei Zheng1
MicroRNAs (miRNAs) have recently been proposed as a versatile class of molecules involved inregulation of various biological processes. Although there is emerging evidence that some mi-croRNAs can function as oncogenes or tumor suppressors, the specific role of miRNA in humanhepatocellular carcinoma (HCC) is unclear at this point. In this study, we examined the mi-croRNA expression profiles in a set of 20 human HCC specimens by miRNA microarray andquantitative real-time polymerase chain reaction. The results showed that among the 20 HCCsamples analyzed, microRNA-101 was significantly down-regulated twofold or more (twofold to20-fold) in 16 samples compared with the matching nontumoral liver tissues. Using both aluciferase reporter assay and Western blot analysis, we showed that microRNA-101 repressed theexpression of v-fos FBJ murine osteosarcoma viral oncogene homolog (FOS) oncogene, a keycomponent of the activator protein-1 (AP-1) transcription factor. Moreover, using a luciferaseexpression vector (pAP-1-Luc) driven by seven copies of an AP-1 cis-element, we observed thatmicroRNA-101 expression inhibited phorbol 12-myristate 13-acetate (PMA)–induced AP-1 ac-tivity. In in vitro Matrigel invasion and Transwell migration assays, enhanced microRNA-101expression inhibited the invasion and migration of cultured HCC cells, respectively. Thesefindings suggest that microRNA-101 may play an important role in HCC. Conclusion: Mi-croRNA-101, which is aberrantly expressed in HCC, could repress the expression of the FOSoncogene. (HEPATOLOGY 2009;49:1194-1202.)
Hepatocellular carcinoma (HCC) is the fourthmost common cause of death from cancer; thehighest age-standardized mortality rate is in
China (34.7/105, the second highest cancer killer since
the 1990s), which alone accounts for 53% of all livercancer deaths worldwide.1 Recently, the incidence ofHCC has been found to be increasing, particularly inmales in Western countries.2,3
Various molecular alterations occur in preneoplasticnodules and escalate in HCC, including dysregulation ofwell-known molecular pathways in carcinogenesis.4-8 Theimportant role of microRNAs (miRNAs) in those path-ways has recently been emphasized. MicroRNAs are smallconserved RNA molecules of approximately 22 nucleo-tides that negatively modulate gene expression in animalsand plants, primarily through base paring to the 3� un-translated region (UTR) of target messenger RNAs (mR-NAs); this leads to mRNA cleavage or translationrepression.9 More than 800 human miRNAs have beendescribed, and speculations about the total number ofhuman miRNAs exceed 1000. Increasing evidence showsthat expression of miRNA is deregulated in human can-cer.10,11 Specific overexpression or underexpression hasbeen shown to correlate with particular tumor types.12,13
For example, let-7, down-regulated in lung cancer,14 sup-presses Ras,15 miR-15 and miR-16, deleted or down-reg-ulated in leukemia,16 suppress BCL2,17 miR-17-5p andmiR-20a control the balance of cell death and prolifera-tion driven by the proto-oncogene c-Myc.18 Altered ex-
Abbreviations: AP-1, activator protein-1; DMEM, Dulbecco’s modified Eaglemedium; FBS, fetal bovine serum; HCC, hepatocellular carcinoma; HGF, hepato-cyte growth factor; miRNA, microRNA; mRNA, messenger RNA; pAP-1-Luc, lu-ciferase expression vector; PCR, polymerase chain reaction; PMA, phorbol 12-myristate 13-acetate; si-FOS, small interfering RNA against the FOS gene; siRNA,small interfering RNA; UTR, untranslated region.
From the 1Beijing Institute of Radiation Medicine, Beijing, China; and the2Department of Pathology, General Hospital of the People’s Liberation Army, Bei-jing, China.
*These authors contributed equally to this work.Received July 20, 2008; accepted November 18, 2008.Supported by grants from Chinese State Key Projects for Basic Research
(2006CB910407, 2002CB513103), National 863 projects (2006AA02Z127)and Chinese National Natural Science Foundation (30873008, 30870529).
Address reprint requests to: Xiaofei Zheng, M.D., or Lixin Wei, M.D., Address:Beijing Institute of Radiation Medicine, 27 Taiping Road, Beijing 100850,P.R.China (Xiaofei Zheng), or Department of Pathology, General Hospital of PLA,28 Fuxing Road, Beijing 100039, P.R. China (Lixin Wei). E-mail:[email protected] (Xiaofei Zheng) or [email protected] (Lixin Wei); fax:(86)-10-68214653 (Xiaofei Zheng).
Copyright © 2008 by the American Association for the Study of Liver Diseases.Published online in Wiley InterScience (www.interscience.wiley.com).DOI 10.1002/hep.22757Potential conflict of interest: Nothing to report.Additional Supporting Information may be found in the online version of this
article.
1194

pression of several miRNAs has also been described inHCC.19-23
In the current study, we compared the miRNA expres-sion profile from HCC tumor tissues and adjacent normalliver tissues. We found that miR-101 is underexpressed inHCC tumor tissues. Sequence analysis suggested a likelyinteraction between the 3� UTR of FOS mRNA andmiR-101. Indeed, we showed that miR-101 could repressthe expression of FOS oncogene using both a reporterassay and western blot analysis. Furthermore, overexpres-sion of miR-101 could inhibit the invasion and migrationof HepG2 and SMMC-7721 cells. These results provide anew pathway for regulation of FOS expression.
Patients and Methods
Patients and Tumor Characteristics. HCCs andsurrounding control tissue specimens were obtained from20 patients at General Hospital of the People’s LiberationArmy (Beijing, China) after surgical resection with in-formed consent. The tumor tissues and adjacent normaltissues were frozen in liquid nitrogen after resection. Nopatient in the current study received chemotherapy orradiation therapy before the surgery. Liver samples werefully clinically characterized (Table 1).
Cell Lines and Cultures. HepG2, HEK 293T, andSMMC-7721 cell lines were maintained in Dulbecco’smodified Eagle medium (DMEM) (GIBCO BRL, GrandIsland, NY) containing 10% fetal bovine serum (FBS)with 100 U/mL penicillin and 100 mg/mL streptomycinat 37°C with 5% CO2.
Vector Construction. For the expression of miR-101,genomic fragment of Homo sapiens miR-101 precursorwas amplified by polymerase chain reaction (PCR) usingthe primer pairs: 5�- acgtctccaaccagaaggtga-3� and 5�-ga-cagaggtgcagggtaagag-3�. The PCR product was clonedinto pcDNA3.0 (Invitrogen, Carlsbad, CA). 3� UTR seg-ment of FOS was subcloned into the pGL3 control vector(Promega, Madison, WI) immediately downstream of thestop codon of the luciferase gene. PCR with the appropri-ate primers also generated inserts with point substitutionsin the miRNA complementary sites. Wild-type and mu-tant inserts were confirmed by sequencing.
Quantitative Reverse Transcription PCR for Mi-croRNA. Total RNA was extracted from the patients’liver samples using Trizol (Invitrogen) according to themanufacturer’s protocol. For microRNA detection, totalRNA was polyadenylated by poly (A) polymerase (Am-bion, Austin, TX) first. Fifty microliters polyadenylationreaction was set up with 10 �g total RNA and 1 �L (2U)
Table 1. Patient Clinical Features and miRNA Expression Profile
PatientNo. Age Sex
Tumor Size(cm*cm*cm)
EdmondsonGrade HBsAg HCV-Ab Cirrhosis
Normalized miRNA Amount in Tumor Tissue Relative toAdjacent Normal Tissue 2���Ct
miR-101 miR-148a miR-182 miR-422b miR-451
1 66 F 15*7*5 3 Positive Negative No 0.05 0.14 54.5 0.09 0.052 39 M 12*11*6 3 Positive Negative No 0.11 0.26 7.11 0.09 0.033 47 M 5*5*4 2 Positive Negative Yes 0.25 0.57 0.28 0.04 0.024 52 M 8*7*4.5 2 Positive Negative No 0.28 0.25 0.04 0.2 0.065 52 M 7*7*7 3 Positive Negative Yes 0.29 0.08 0.87 0.18 0.076 51 M 2.5*2*2 2 Positive Negative Yes 0.24 0.4 59.3 0.31 0.217 59 M 8.8*8.5*8 2 Negative Negative No 0.12 2.36 18.4 2.93 0.178 34 M 10*7*7 3 Positive Negative Yes 0.07 0.11 5.73 0.04 0.039 50 M 2.5*2.5*2 3 Positive Negative No 0.22 0.7 52.7 0.2 0.03
10 33 M 10*8*7 4 Positive Negative Yes 0.26 0.21 0.25 0.06 0.00611 50 M 2.5*1.8*1.5 1 Negative Negative No 0.27 0.47 0.47 0.17 0.0612 52 M 10.5*8*5 3 Positive Negative Yes 0.15 0.1 1.83 0.03 0.0813 35 F 5*4*4 3 Positive Negative Yes 0.15 0.95 9.02 0.26 0.3414 63 M 14*10*10 2 Positive Negative Yes 0.14 0.18 1.02 0.19 0.0815 70 M 3.5*3.5*3 2 Positive Negative Yes 0.37 0.28 24.6 0.26 0.3116 64 M 8*5*4 2 Negative Positive No 1.29 0.64 0.15 0.46 25.317 50 F 6*5.5*2.5 1 Negative Negative No 0.81 0.008 657.1 0.01 0.3418 41 F 12*10*8 3 Positive Negative No 0.62 0.73 21.1 0.82 0.4919 51 M 2.5*2*2 2 Positive Negative Yes 0.63 0.97 84.4 0.88 0.0420 49 M 5*4*2.5 3 Positive Negative Yes 0.34 0.32 4.59 0.09 0.17
HBsAg indicates hepatitis B surface antigen; HCV-Ab, hepatitis C virus antibody; M, male; F, female; miRNA, microRNA. Relative quantification was performed bythe 2���Ct method with adjacent normal liver tissue sample as a calibrator. Data show the means from three independent analyzes. Every independent analyze wascarried out after the RNA extraction step. Total RNA was Poly-A tailed, reverse transcripted, and then real-time PCR tested. �CT obtained from real-time PCR was subjectto paired t test (�CT � CT miR � CT U6). The expression levels of miR-101, miR-148a, miR-422b, and miR-451 in tumor tissues were significantly lower than in adjacentnormal tissues (miR-101, P � 0.01, t � 7.76; miR-148a, P � 0.01, t � 4.60; miR-422b, P � 0.01, t � 6.95; miR-451, P � 0.01, t � 6.03). miR-182 expressionlevel in tumor tissues was significantly higher than in adjacent normal tissues (P � 0.01, t � �2.96).
HEPATOLOGY, Vol. 49, No. 4, 2009 LI, FU, ET AL. 1195

poly (A) polymerase according to the manufacturer’s pro-tocol. The reaction was incubated at 37 °C for 60 min-utes. After incubation, poly (A)-tailed total RNA wasrecovered by phenol/chloroform extraction and ethanol pre-cipitation. Reverse transcription was performed using 1 �gpoly (A)-tailed total RNA and 1 �g reverse transcriptionprimer (5�-gcgagcacagaattaatacgactcactatagg(t)18VN-3�)with 1 �L ImPro-II Reverse Transcriptase (Promega) ac-cording to the manufacturer’s protocol. Quantitative PCRwas performed as described in the method of QuantitectSYBR Green PCR Kit (Qiagen, Hilden, Germany) withMx3000p (Stratagene, La Jolla, CA) supplied with analyticalsoftware. One primer of microRNA amplification is mi-croRNA specific (micorRNA-101 5�-tacagactgtgataactgaag-3�; microRNA-148a 5�-tcagtgcactacagaactttg-3�; mi-croRNA-182 5�-tttggcaatggtagaactcaca-3�; microRNA-422b 5�-ctggacttggagtcagaaggcc-3�; microRNA-451 5�-aaaccgttaccattactgagt-3�), and the other is a universal primer(5�-gcgagcacagaattaatacgac-3�). U6 small nuclear RNA lev-els were used for normalization (U6-F 5�-cgcttcggcagca-catatacta-3�; U6-R 5�-cgcttcacgaatttgcgtgtca -3�). GenEXsoftware (TATAA Biocenter, Munich, Germany) was em-ployed to analyze real-time PCR data. Paired Student t testwas performed to determine the difference of miRNA’s ex-pression levels observed between noncancerous and cancertissues.
MiRNAs, Small Interfering RNAs, and Transfec-tion. The miR-101 duplex and small interfering RNAs(siRNAs) against FOS were designed and synthesized byGenePharma (GenePharma, Shanghai, China). The se-quences of miRNA duplex and siRNA are presented inSupporting Table 1. Small interfering RNAs and miR-NAs transfection were performed using Lipofectamine2000 (Invitrogen). In brief, cells were plated in six-wellplate to 50% confluence. For each well, 5 �L siRNA ormiRNA (20 �M) was added into 250 �L Opti-MEMmedium (GIBCO BRL), 5 �L lipofectamine 2000 into250 �L Opti-MEM medium, and then mixed siRNA ormiRNA with lipofectamine 2000. The mixture was addedto cells and incubated for 6 hours before replacing themedium. Total RNAs and protein were prepared 48hours after transfection and were used for quantitativereverse transcription PCR or Western blotting analysis.
Western Blot. Total cell lysate was prepared in 1�sodium dodecyl sulfate buffer. Proteins at the sameamount were separated by sodium dodecyl sulfate poly-acrylamide gel electrophoresis and transferred onto poly-vinylidene fluoride membranes. After probing with anti-FOS (sc-52; Santa Cruz Biotechnology, Inc., Santa Cruz,CA) or anti-�-actin antibody (Beijing Zhongshan Bio-technology, Beijing, China), antigen–antibody complex
was visualized by enhanced chemiluminescence reagentsSupersignal (Pierce, Rockford, IL). For quantification,Quantity One software (Bio-Rad Laboratories, Hercules,CA) was used.
Luciferase Reporter Assay. Adherent HepG2 cellswere grown in 10% FBS in DMEM to 80%-90% conflu-ence in 24-well plates. Cells were cotransfected with 100ng firefly luciferase reporter vector containing the FOS3�UTR (named pGL3-FOS-3�UTR) or FOS 3�UTRmutant (named pGL3-FOS-3�UTRmut) and 8 ng of thecontrol vector containing Renilla luciferase, pRL-TK(Promega), in a final volume of 0.5 mL using lipo-fectamine 2000 (Invitrogen). Firefly and Renilla lucif-erase activities were measured consecutively using dual-luciferase assays (Promega) 48 hours after transfection.
Promotor Activity Assay. Cells were grown on 24-well plates at 70% to 80% confluence and then trans-fected with 100 ng luciferase expression vector (pAP-1-luc; Stratagene) and 10 pmol negative control, miR-101duplex, or siRNA against the FOS gene (si-FOS). Ap-proximately 48 hours after transfection, cells were treatedwith either DMSO or phorbol 12-myristate 13-acetate(PMA) (40 ng/mL) for 2 hours. Cells were lysed and Lucactivity was measured using a luciferase assay kit (Pro-mega). Luc activity of individual samples was normalizedagainst total protein.
Transwell Cell Invasion and Migration Assay. Forinvasion assay, SMMC-7721 or HepG2 transfectants wereserum-starved for 18 hours in DMEM containing 0.1%FBS. Serum-starved cells were trypsinized and resuspendedin DMEM containing 0.1% FBS, and 2 � 105 cells wereadded to the upper chamber of each well (6.5 mm in diam-eter, 8-�m pore size; Corning, NY) coated with 30 mg/cm2
Matrigel (ECM gel, Sigma-Aldrich, St. Louis, MO). Me-dium containing 0.1% FBS and supplemented with hepato-cyte growth factor (HGF) (20 ng/mL) was placed in thelower compartment of the chamber. After 24 hours at 37°C,cells on the upper membrane surface were removed by care-ful wiping with a cotton swab, and the filters were fixed bytreatment with 95% ethanol for 30 minutes and stained with0.2% crystal violet solution for 30 minutes. Invasive cellsadhering to the undersurface of the filter were then counted(five high-power fields/chamber) using an inverted micro-scope. The migration assay is the same with invasion assayexcepting no matrigel was used and the permeating time forcells was 12 hours.
Statistical Analysis of Data. Unless stressed, all val-ues are reported as means � standard deviation. Differ-ences were assessed by two-tailed Student t test usingExcel software. P � 0.05 was considered statistically sig-nificant.
1196 LI, FU, ET AL. HEPATOLOGY, April 2009

Results
MiR-101 Is Underexpressed in Human HCC. Tis-sue samples of 20 primary HCCs including cancerous andadjacent normal liver tissues were obtained from patientswho underwent hepatectomy (Table 1). Two microarrayhybridization studies were performed on a pair of tumor-derived and matching adjacent normal liver–derivedRNA (patient 20; see Supporting Methods). The miR-NAs that are differentially overexpressed in HCC in-
cluded miR-21, miR-182, and miR-200c. Of themiRNAs that had decreased expression, the greatestchanges were noted for miR-101, miR-148a, miR-422b,and miR-451 (Fig. 1). Several studies that investigated theexpression of the entire microRNAome in various humansolid tumors such as lung cancer, breast cancer, and pitu-itary adenoma have revealed differences in miR-101 ex-pression between neoplastic and normal tissues. 24-27 Weemployed real-time PCR to analyze the expression of ma-ture miR-101 in other 19 primary HCCs. The results
Fig. 1. MicroRNA expression profiling of adjacent normal liver and HCC tumor tissue (patient 20). MicroRNAs were detected by microarray. Aheatmap was generated from the average of normalized log-transformed fluorescent intensity for each data set.
HEPATOLOGY, Vol. 49, No. 4, 2009 LI, FU, ET AL. 1197

showed that among the 19 HCC samples analyzed, miR-101 was significantly underexpressed twofold or more(twofold to 20-fold) in 15 samples (numbers 1-15) com-pared with the matching nontumoral liver tissues (Table1). Moreover, four miRNAs were found to be signifi-cantly up-regulated (miR-182) or down-regulated (miR-148a, miR-422b, and miR-451) in HCC tissuescompared with the adjacent normal liver tissues (Table 1).
Interaction of miR-101 with the 3� UTR of the FOSmRNA. To find target genes of miR-101, we analyzedcandidates by using the TargetScan bioinformaticstools.28 The analysis revealed the FOS gene 3� UTR har-bors one putative binding site for miR-101 and, impor-tantly, the site is conserved across various species (Fig.2A). To investigate the potential interaction experimen-tally, the human FOS 3� UTR was subcloned after thefirefly luciferase open reading frame (ORF) and cotrans-fected into HepG2 cells with the miR-101–expressingvector (pcDNA3.0-miR101). pcDNA3.0-miR101 pro-duced a 40% decrease in relative luciferase activity com-pared with pcDNA3.0-transfected cells (Fig. 2B). An
analogous reporter with point substitutions disruptingthe target sites (Fig. 2A) was also cotransfected withpcDNA3.0-miR101. There was no decrease in relativeluciferase activity compared with pcDNA3.0-transfectedcells (Fig. 2B). Moreover, we designed a synthetic RNAduplex mimicking the miR-101 molecule. The MiR-101duplex produced a 60% decrease in relative luciferase ac-tivity compared with negative control (Supporting Fig.1). Some other miRNAs that have putative target sites onFOS 3� UTR, such as miR-29a, were also used for lucif-erase assay by cotransfection of the pGL3-FOS-3� UTRvector and miRNA’s expressing vector into HepG2 cells.No decreases in relative luciferase activity were producedby any of these miRNAs (Supporting Fig. 2).
FOS Is a Potential Target of miR-101. To learnwhether miR-101 can affect endogenous FOS proteinlevel, we next examined the effect of this miRNA in cul-tured tissue cells treated with the FOS-inducing PMA.29
Western blot from protein extracts obtained from theHEK 293T cells after treatment of PMA revealed dramat-ically reduced activation of FOS after pcDNA3.0-
Fig. 2. MicroR-101 inhibited FOS expression. (A; upper panel) The target site of miR-101 in FOS 3� UTR is conserved among mammalian species(shown in red). (A; lower panel) Predicted duplex formation between miR-101 and the targeted FOS 3� UTR. The FOS 3� UTR mutant is identical tothe wild-type, except that it had four point substitutions (red) disrupting pairing to miR-101 seed. (B) pGL3-Fos-3�UTR reporter plasmid in which theluciferase coding sequence had been fused to the 3� UTR of FOS was cotransfected into HepG2 cells with pcDNA3.0 (gray columns) orpcDNA3.0-miR101 (white columns). Luciferase activity was normalized relative to a simultaneously transfected Renilla expression plasmid. The 3�UTR-Mut indicates the introduction of alterations into the seed complementary sites shown in Fig. 2A. Results of the mean of triplicate assays withstandard deviation of the mean are presented. *P � 0.05. (C) Western blots of FOS from HEK 293T cells after PMA induction. Cells were transfectedwith pcDNA3.0 or pcDNA3.0-miR101; after 48 hours, PMA (40 ng/mL) was added. Cells were harvested 2 hours later, and 30 �g of whole-cell lysatewas loaded into each lane. A �-actin antibody was used in a reprobing as a loading control. (D) Real-time reverse transcription PCR of FOS in HEK293T cells transfected with the pcDNA3.0 or pcDNA3.0-miR101 vector for 48 hours and thereafter treated with PMA (40 ng/ml) for 1 hour. Datawere normalized to the level of glyceraldehyde 3-phosphate dehydrogenase mRNA. Results of the mean of triplicate quantitative PCR assays withstandard deviation of the mean are presented.
1198 LI, FU, ET AL. HEPATOLOGY, April 2009
miR101 transfection (Fig. 2C). In contrast, pcDNA3.0-transfected cells showed no reduction in FOS activation.Because inhibition of expression by miRNAs may also bemediated by mRNA degradation,30,31 We examinedwhether the PMA-induced elevation in FOS mRNA lev-els might be affected by pcDNA3.0-miR101. Figure 2Dshows that the FOS mRNA expression is not changed,indicating that reduction of FOS levels by miR-101 oc-curs by translational suppression rather than by mRNAdegradation. The PMA-induced expression of FOS genewas also inhibited at the protein level when HEK 293Tcells were transfected with miR-101 duplex (SupportingFig. 3).
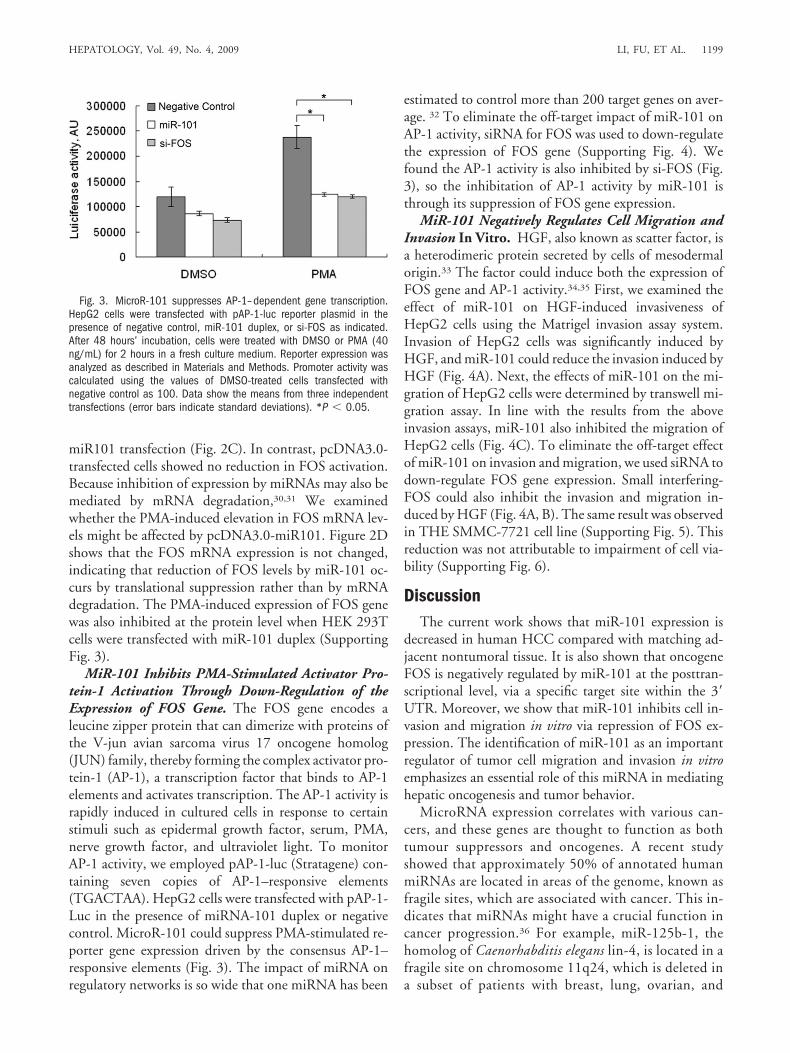
MiR-101 Inhibits PMA-Stimulated Activator Pro-tein-1 Activation Through Down-Regulation of theExpression of FOS Gene. The FOS gene encodes aleucine zipper protein that can dimerize with proteins ofthe V-jun avian sarcoma virus 17 oncogene homolog(JUN) family, thereby forming the complex activator pro-tein-1 (AP-1), a transcription factor that binds to AP-1elements and activates transcription. The AP-1 activity israpidly induced in cultured cells in response to certainstimuli such as epidermal growth factor, serum, PMA,nerve growth factor, and ultraviolet light. To monitorAP-1 activity, we employed pAP-1-luc (Stratagene) con-taining seven copies of AP-1–responsive elements(TGACTAA). HepG2 cells were transfected with pAP-1-Luc in the presence of miRNA-101 duplex or negativecontrol. MicroR-101 could suppress PMA-stimulated re-porter gene expression driven by the consensus AP-1–responsive elements (Fig. 3). The impact of miRNA onregulatory networks is so wide that one miRNA has been
estimated to control more than 200 target genes on aver-age. 32 To eliminate the off-target impact of miR-101 onAP-1 activity, siRNA for FOS was used to down-regulatethe expression of FOS gene (Supporting Fig. 4). Wefound the AP-1 activity is also inhibited by si-FOS (Fig.3), so the inhibitation of AP-1 activity by miR-101 isthrough its suppression of FOS gene expression.
MiR-101 Negatively Regulates Cell Migration andInvasion In Vitro. HGF, also known as scatter factor, isa heterodimeric protein secreted by cells of mesodermalorigin.33 The factor could induce both the expression ofFOS gene and AP-1 activity.34,35 First, we examined theeffect of miR-101 on HGF-induced invasiveness ofHepG2 cells using the Matrigel invasion assay system.Invasion of HepG2 cells was significantly induced byHGF, and miR-101 could reduce the invasion induced byHGF (Fig. 4A). Next, the effects of miR-101 on the mi-gration of HepG2 cells were determined by transwell mi-gration assay. In line with the results from the aboveinvasion assays, miR-101 also inhibited the migration ofHepG2 cells (Fig. 4C). To eliminate the off-target effectof miR-101 on invasion and migration, we used siRNA todown-regulate FOS gene expression. Small interfering-FOS could also inhibit the invasion and migration in-duced by HGF (Fig. 4A, B). The same result was observedin THE SMMC-7721 cell line (Supporting Fig. 5). Thisreduction was not attributable to impairment of cell via-bility (Supporting Fig. 6).
DiscussionThe current work shows that miR-101 expression is
decreased in human HCC compared with matching ad-jacent nontumoral tissue. It is also shown that oncogeneFOS is negatively regulated by miR-101 at the posttran-scriptional level, via a specific target site within the 3�UTR. Moreover, we show that miR-101 inhibits cell in-vasion and migration in vitro via repression of FOS ex-pression. The identification of miR-101 as an importantregulator of tumor cell migration and invasion in vitroemphasizes an essential role of this miRNA in mediatinghepatic oncogenesis and tumor behavior.
MicroRNA expression correlates with various can-cers, and these genes are thought to function as bothtumour suppressors and oncogenes. A recent studyshowed that approximately 50% of annotated humanmiRNAs are located in areas of the genome, known asfragile sites, which are associated with cancer. This in-dicates that miRNAs might have a crucial function incancer progression.36 For example, miR-125b-1, thehomolog of Caenorhabditis elegans lin-4, is located in afragile site on chromosome 11q24, which is deleted ina subset of patients with breast, lung, ovarian, and
Fig. 3. MicroR-101 suppresses AP-1–dependent gene transcription.HepG2 cells were transfected with pAP-1-luc reporter plasmid in thepresence of negative control, miR-101 duplex, or si-FOS as indicated.After 48 hours’ incubation, cells were treated with DMSO or PMA (40ng/mL) for 2 hours in a fresh culture medium. Reporter expression wasanalyzed as described in Materials and Methods. Promoter activity wascalculated using the values of DMSO-treated cells transfected withnegative control as 100. Data show the means from three independenttransfections (error bars indicate standard deviations). *P � 0.05.
HEPATOLOGY, Vol. 49, No. 4, 2009 LI, FU, ET AL. 1199

cervical cancers. MiR-101, which is located at fragilesite and cancer-associated genomic region, has beenshown to be down-regulated in many different solidtumors, including breast, lung, and pituitary adeno-ma.24-27 Thus, it is likely that miR-101 plays a funda-mental role in tumor cell behavior and malignanttransformation, and our findings may potentially berelevant to other tumors in which miR-101 is under-expressed.
FOS has been implicated as a key contributor to HCCpathogenesis. Arbuthnot et al.37 reported that c-myc andc-fos mRNA, as well as their protein products, are in-creased in human liver cancers. Wang et al.38 have alsofound an apparently higher expression of N-ras and c-fosin human hepatoma than in its adjacent liver tissue. Re-cently, Feng et al.39 reported the positive rates and signalintensity of c-fos, and some other proteins in HCC aresignificantly higher than those in pericarcinomatous tis-sues. Yuen et al.40 found that expression of c-fos was sig-nificantly higher in tumor tissue than in nontumor tissue(91% versus 0%, P � 0.0001). Our observations suggestthat down-regulation of miR-101 in HCC tissues maycontribute to the high expression level of FOS protein inHCC.
The activation of the AP-1 family of transcription fac-tors, including c-fos and c-jun family members, is one ofthe earliest nuclear events induced by growth factors thatstimulate extracellular signal-regulated kinases. In the caseof c-fos, the activation of extracellular signal-regulatedkinases leads to an increased expression of FOS mRNA(Fig. 5). We therefore have a potential autoregulatoryloop whereby growth factors activate FOS and the tran-scriptional activation of AP-1 while increasing expressionof miR-101 (via 5� AP-1 sites), which as shown here canact as a functional brake on FOS expression, a neat systemfor preventing transcriptional overshoot. However, theamount of miR-101 in HepG2 cells has no change whentreated with HGF or PMA (data not shown).
One hallmark of metastasis and invasive growth is thetransition of tumour cells from an epithelial to a mesen-chymal morphology, known as the epithelial–mesenchy-mal transition (EMT). Both c-fos and c-jun can induceepithelial–mesenchymal transition, which is associatedwith loss of cell polarity, in mammary epithelial cells.41,42
However, only the overexpression of c-fos is able to pro-mote invasive growth in collagen gels, which indicatesthat c-fos might have a more important role than c-junduring late-stage tumorigenesis.41 The importance of c-fos in tumor invasion has been supported in vivo, becausethe progression of chemically induced papillomas to in-vasive squamous cell carcinomas is impaired in c-fos–de-ficient mice.43 In the current study, we found that miR-
Fig. 4. MicroR-101 suppressed invasion and migration of HepG2 cellsby blocking FOS expression. (A) Cell invasion evaluated using Matrigelinvasion chambers. Mock-transfected (no miRNA or siRNA) cells or cellstransfected with negative control siRNA, miR-101 duplex, or FOS siRNAwere plated 48 hours after transfection on six-well transwell plates. Cellswere added to the upper chamber of each well. After incubation for 24hours, cells that invaded through the pores to the under surface of themembrane were fixed, stained, and counted. Five random microscopicfields were counted for each treatment. (B) Invading cell numbers are theaverage count of five random microscopic fields. Each bar represents themean � standard deviation of the counts from a single representativeexperiment. **P � 0.01; ***P � 0.001. (C) MicroR-101 blocks cellmigration as assayed in transwell chambers, with incubation for 12 hoursand HGF (20 ng/ml) as chemoattractant. Cells that migrated the filterwere fixed and stained with crystal violet. Representative images of thelower surface of the membrane are shown. (D) Quantification of the cellmigration results from three experiments. Values shown are mean �standard deviation from a representative experiment. ***P � 0.001.
1200 LI, FU, ET AL. HEPATOLOGY, April 2009

101 could inhibit the HGF-induced invasion andmigration in the HCC cell line through repressing theexpression of the FOS oncogene. These findings implythat miR-101 might be a good candidate for anticancertherapy.
In conclusion, miR-101, a microRNA underexpressedin HCC, inhibits the expression of the FOS oncogeneposttranscriptionally by binding to the 3� UTR of theFOS mRNA, thereby reducing HGF-induced cell inva-sion and migration.
References1. Pisani P, Parkin DM, Bray F, Ferlay J. Erratum: estimates of the worldwide
mortality from 25 cancers in 1990. Int J Cancer 1999;83:18-29; 870-873.2. Taylor-Robinson SD, Foster GR, Arora S, Hargreaves S, Thomas HC.
Increase in primary liver cancer in the UK, 1979-94. Lancet 1997;350:1142-1143.
3. El-Serag HB, Mason AC. Rising incidence of hepatocellular carcinoma inthe United States. N Engl J Med 1999;340:745-750.
4. El-Serag HB, Rudolph KL. Hepatocellular carcinoma: epidemiology andmolecular carcinogenesis. Gastroenterology 2007;132:2557-2576.
5. Breuhahn K, Vreden S, Haddad R, Beckebaum S, Stippel D, Flemming D,et al. Molecular profiling of human hepatocellular carcinoma defines mu-tually exclusive interferon regulation and insulin-like growth factor II over-expression. Cancer Res 2004;64:6058-6064.
6. Wurmbach E, Chen Y, Khitrov G, Zhang W, Roayaie S, Schwartz M, et al.Genome-wide molecular profiles of HCV-induced dysplasia and hepato-cellular carcinoma. HEPATOLOGY 2007;45:938-947.
7. Lee JS, Thorgeirsson SS. Comparative and integrative functional genomicsof HCC. Oncogene 2006;25:3801-3809.
8. Lemmer ER, Friedman SL, Llovet JM. Molecular diagnosis of chronic liverdisease and hepatocellular carcinoma: the potential of gene expression pro-filing. Semin Liver Dis 2006;26:373-383.
9. Bartel DP. MicroRNAs: genomics, biogenesis, mechanism, and function.Cell 2004;116:281-297.
10. Croce CM, Calin GA. miRNAs, cancer, and stem cell division. Cell 2005;122:6-7.
11. Gregory RI, Shiekhattar R. MicroRNA biogenesis and cancer. Cancer Res2005;65:3509-3512.
12. Lu J, Getz G, Miska EA, Alvarez-Saavedra E, Lamb J, Peck D, et al.MicroRNA expression profiles classify human cancers. Nature 2005;435:834-838.
13. Volinia S, Calin GA, Liu CG, Ambs S, Cimmino A, Petrocca F, et al. AmicroRNA expression signature of human solid tumors defines cancer genetargets. Proc Natl Acad Sci U S A 2006;103:2257-2261.
14. Takamizawa J, Konishi H, Yanagisawa K, Tomida S, Osada H, Endoh H,et al. Reduced expression of the let-7 microRNAs in human lung cancers inassociation with shortened postoperative survival. Cancer Res 2004;64:3753-3756.
15. Johnson SM, Grosshans H, Shingara J, Byrom M, Jarvis R, Cheng A, et al.RAS is regulated by the let-7 microRNA family. Cell 2005;120:635-647.
16. Calin GA, Dumitru CD, Shimizu M, Bichi R, Zupo S, Noch E, et al.Frequent deletions and down-regulation of micro- RNA genes miR15 andmiR16 at 13q14 in chronic lymphocytic leukemia. Proc Natl Acad Sci U SA 2002;99:15524-15529.
17. Cimmino A, Calin GA, Fabbri M, Iorio MV, Ferracin M, Shimizu M, etal. miR-15 and miR-16 induce apoptosis by targeting BCL2. Proc NatlAcad Sci U S A 2005;102:13944-13949.
18. O’Donnell KA, Wentzel EA, Zeller KI, Dang CV, Mendell JT. c-Myc-regulated microRNAs modulate E2F1 expression. Nature 2005;435:839-843.
19. Kutay H, Bai S, Datta J, Motiwala T, Pogribny I, Frankel W, et al. Down-regulation of miR-122 in the rodent and human hepatocellular carcino-mas. J Cell Biochem 2006;99:671-678.
20. Murakami Y, Yasuda T, Saigo K, Urashima T, Toyoda H, Okanoue T, etal. Comprehensive analysis of microRNA expression patterns in hepato-cellular carcinoma and non-tumorous tissues. Oncogene 2006;25:2537-2545.
Fig. 5. A schematic diagram illustrating how miR101 can modulate (FOS) mRNA translation through interactions with the 3� UTR.
HEPATOLOGY, Vol. 49, No. 4, 2009 LI, FU, ET AL. 1201

21. Meng F, Henson R, Wehbe-Janek H, Ghoshal K, Jacob ST, Patel T.MicroRNA-21 regulates expression of the PTEN tumor suppressor gene inhuman hepatocellular cancer. Gastroenterology 2007;133:647-658.
22. Ladeiro Y, Couchy G, Balabaud C, Bioulac-Sage P, Pelletier L, RebouissouS, et al. MicroRNA Profiling in hepatocellular tumors is associated withclinical features and oncogene/tumor suppressor gene mutations. HEPA-TOLOGY 2008;47:1955-1963.
23. Varnholt H, Drebber U, Schulze F, Wedemeyer I, Schirmacher P, DienesHP, et al. MicroRNA gene expression profile of hepatitis C virus-associatedhepatocellular carcinoma. HEPATOLOGY 2008;47:1223-1232.
24. Yanaihara N, Caplen N, Bowman E, Seike M, Kumamoto K, Yi M, et al.Unique microRNA molecular profiles in lung cancer diagnosis and prog-nosis. Cancer Cell 2006;9:189-198.
25. Iorio MV, Ferracin M, Liu CG, Veronese A, Spizzo R, Sabbioni S, et al.MicroRNA gene expression deregulation in human breast cancer. CancerRes 2005;65:7065-7070.
26. Mattie MD, Benz CC, Bowers J, Sensinger K, Wong L, Scott GK, et al.Optimized high-throughput microRNA expression profiling providesnovel biomarker assessment of clinical prostate and breast cancer biopsies.Mol Cancer 2006;5:24.
27. Bottoni A, Zatelli MC, Ferracin M, Tagliati F, Piccin D, Vignali C, et al.Identification of differentially expressed microRNAs by microarray: a pos-sible role for microRNA genes in pituitary adenomas. J Cell Physiol 2007;210:370-377.
28. Lewis BP, Shih IH, Jones-Rhoades MW, Bartel DP, Burge CB. Predictionof mammalian microRNA targets. Cell 2003;115:787-798.
29. Hill CS, Wynne J, Treisman R. Serum-regulated transcription by serumresponse factor (SRF): a novel role for the DNA binding domain. EMBOJ 1994;13:5421-5432.
30. Lim LP, Lau NC, Garrett-Engele P, Grimson A, Schelter JM, Castle J, etal. Microarray analysis shows that some microRNAs downregulate largenumbers of target mRNAs. Nature 2005;433:769-773.
31. Bagga S, Bracht J, Hunter S, Massirer K, Holtz J, Eachus R, et al. Regula-tion by let-7 and lin-4 miRNAs results in target mRNA degradation. Cell2005;122:553-563.
32. Zamore PD, Haley B. Ribo-gnome: the big world of small RNAs. Science2005;309:1519-1524.
33. Nakamura T, Nishizawa T, Hagiya M, Seki T, Shimonishi M, SugimuraA, et al. Molecular cloning and expression of human hepatocyte growthfactor. Nature 1989;342:440-443.
34. Fabregat I, de Juan C, Nakamura T, Benito M. Growth stimulation of ratfetal hepatocytes in response to hepatocyte growth factor: modulation ofc-myc and c-fos expression. Biochem Biophys Res Commun 1992;189:684-690.
35. Rahmani M, Nadori F, Durand-Schneider AM, Lardeux B, Bernuau D.Hepatocyte growth factor activates the AP-1 complex: a comparison be-tween normal and transformed rat hepatocytes. J Hepatol 1999;30:916-925.
36. Calin GA, Sevignani C, Dumitru CD, Hyslop T, Noch E, Yendamuri S, etal. Human microRNA genes are frequently located at fragile sites andgenomic regions involved in cancers. Proc Natl Acad Sci U S A 2004;101:2999-3004.
37. Arbuthnot P, Kew M, Fitschen W. c-fos and c-myc oncoprotein expressionin human hepatocellular carcinomas. Anticancer Res 1991;11:921-924.
38. Wang Z, Xiang Q, Li D, Li S. Correlation between gene expression andchromatin conformation of c-fos and N-ras in human liver and hepatoma.Chin Med Sci J 1991;6:6-8.
39. Feng DY, Zheng H, Tan Y, Cheng RX. Effect of phosphorylation ofMAPK and Stat3 and expression of c-fos and c-jun proteins on hepatocar-cinogenesis and their clinical significance. World J Gastroenterol 2001;7:33-36.
40. Yuen MF, Wu PC, Lai VC, Lau JY, Lai CL. Expression of c-Myc, c-Fos,and c-jun in hepatocellular carcinoma. Cancer 2001;91:106-112.
41. Reichmann E, Schwarz H, Deiner EM, Leitner I, Eilers M, Berger J, et al.Activation of an inducible c-FosER fusion protein causes loss of epithelialpolarity and triggers epithelial-fibroblastoid cell conversion. Cell 1992;71:1103-1116.
42. Fialka I, Schwarz H, Reichmann E, Oft M, Busslinger M, Beug H. Theestrogen-dependent c-JunER protein causes a reversible loss of mammaryepithelial cell polarity involving a destabilization of adherens junctions.J Cell Biol 1996;132:1115-1132.
43. Saez E, Rutberg SE, Mueller E, Oppenheim H, Smoluk J, Yuspa SH, et al.c-fos is required for malignant progression of skin tumors. Cell 1995;82:721-732.
1202 LI, FU, ET AL. HEPATOLOGY, April 2009





























