Embed Size (px)
Citation preview

Minor pilin subunits are conserved in Vibrio cholerae type IV pili
Claudia Toma a;�, Hiromi Kuroki a, Noboru Nakasone a, Masahiko Ehara b,Masaaki Iwanaga a
a Department of Bacteriology, Faculty of Medicine, University of the Ryukyus, Okinawa 903-0215, Japanb Department of Bacteriology, Institute of Tropical Medicine, Nagasaki University, Nagasaki 852-8523, Japan
Received 8 October 2001; received in revised form 20 November 2001; accepted 13 December 2001
First published online 5 February 2002
Abstract
The nucleotide sequences of five open reading frames within the Vibrio cholerae NAGV14 type IV pilus gene cluster were determined.The genes showed high homology to the mannose-sensitive hemagglutinin (MSHA) pilus genes mshB, mshC, mshD, mshO and mshP. PCRanalysis showed that a MSHA-like gene cluster is highly conserved among different V. cholerae strains, with the exception of thepreviously reported major pilin subunit. Recombinant MshB and MshO proteins were purified and specific antiserum was raised to eachof them. Western blotting analyses showed that these antisera reacted with purified NAGV14 and MSHA pili. The results suggested thatMshB and MshO are minor components of the pilus fiber. Although there was no cross-reaction between the major pilin subunits ofNAGV14 and MSHA pili, minor components seemed to be highly homologous and immunologically cross-reactive. ß 2002 Federationof European Microbiological Societies. Published by Elsevier Science B.V. All rights reserved.
Keywords: Type IV pilus; Major pilin; Minor pilin; Vibrio cholerae
1. Introduction
Vibrio cholerae exists as more than 140 serogroups as-sociated with surface polysaccharides. Epidemic cholerahas been associated with serogroups O1 and O139 [1], inwhich the ability to produce cholera toxin is an essentialdeterminant of virulence. It has become clear that non-O1,non-O139 serogroups are also involved in acute secretorydiarrhea, which is called ‘cholera-like disease’. These se-rogroups that do not possess the CTX genetic element butthat can cause diarrhea in humans by an unknown mech-anism have been described as enteropathogenic V. cholerae[2]. A critical step in pathogenesis for all virulent strains islikely to be adhesion to and colonization of the intestinalmucosa [3]. This adhesion step is very important as atarget for investigation of pathogenic mechanisms andvaccine development, however, the adherence mechanismsutilized by V. cholerae are still unclear [4].
Type IV pili are thin, 6^7-nm ¢bers elaborated by awide variety of Gram-negative bacterial species. They are
implicated in adherence, bio¢lm formation, horizontal ge-netic transfer and twitching motility [3,5,6]. Three kinds oftype IV pilus gene clusters have been reported and genet-ically characterized for V. cholerae O1:1) the toxin-core-gulated pilus (TCP), which has been recently reported tobe a coat protein of a ¢lamentous bacteriophage [7], 2) themannose-sensitive hemagglutinin (MSHA) pilus [8], and 3)the PIL gene cluster [9]. The MSHA pilus is encoded in agene locus that contains 16 genes organized in secretoryand structural operons. Five of these genes encode type IVprepilin-like proteins [8]. Evidence for the presence of theTCP gene cluster in non-O1, non-toxigenic V. choleraestrains has been reported [10], however, no detailed inves-tigation about the presence of the MSHA and PIL geneclusters in these strains has been done.
We have puri¢ed and characterized a variety of type IVpili from V. cholerae [11^14], but only NAGV14 pili pu-ri¢ed from a V. cholerae O34 strain was identi¢ed as amediator of adhesion of the organisms to the rabbit intes-tine [14]. Although puri¢ed MSHA pili did not react withanti-NAGV14 pilus serum in Western blotting, whole celllysates of V. cholerae O1 strains reacted with this antise-rum, suggesting that an antigen cross-reacting with theNAGV14 pilus is present in V cholerae O1 strains [14].In a previous study, we analyzed the nucleotide sequence
0928-8244 / 02 / $22.00 ß 2002 Federation of European Microbiological Societies. Published by Elsevier Science B.V. All rights reserved.PII: S 0 9 2 8 - 8 2 4 4 ( 0 2 ) 0 0 2 7 3 - 0
* Corresponding author. Tel. : +81 (98) 895 1124;Fax: +81 (98) 895 1408.
E-mail address: [email protected] (C. Toma).
FEMSIM 1392 23-4-02 Cyaan Magenta Geel Zwart
FEMS Immunology and Medical Microbiology 33 (2002) 35^40
www.fems-microbiology.org

of the major pilin subunit of NAGV14 pilus (VcfA) andstudied the distribution of NAGV14 vcfA among V. cho-lerae strains by PCR. Only 4% of the V. cholerae non-O1,non-O139 strains tested were positive, while all V. choleraeO1 and all O139 strains were negative [15]. This resultsuggested that other pilin subunit(s) might be responsiblefor the cross-reactivity observed previously in the wholecell lysates of V. cholerae O1 strains [14]. In this study, wefurther analyzed the NAGV14 pilus gene cluster and in-vestigated the presence of cross-reactive minor pilin sub-units in NAGV14 and MSHA pili.
2. Materials and methods
2.1. Bacterial strains, vectors and media
V. cholerae O34 strain NAGV14 [14] was used for DNAsequencing. Escherichia coli XL-1 Blue and M15[pREP4]were used as recombinant host strains. V. cholerae O34strain 88UDT119, which also possesses NAGV14 pili[15], was used as a host strain for construction of a hyper-piliated strain. Other strains stocked in our laboratorywere used for PCR analysis. Bacteria were grown in Lu-ria^Bertani broth or agar. Media were supplemented, asnecessary, with ampicillin (50 or 100 Wg ml31) and/or ka-namycin (25 Wg ml31). The plasmids used in this studywere pCRR2.1 (Invitrogen), pGEX-3X (Pharmacia),pQE30 and pQE16 (Qiagen).
2.2. Nucleotide sequence analysis
DNA sequencing of the region downstream from vcfAwas performed using the LA PCR in vitro Cloning kit(Takara) and SalI-digested chromosomal DNA, followingthe manufacturer’s instructions as described previously[15]. Extension of DNA sequence was achieved with prim-er walking. For sequencing of the upstream region, a syn-thetic oligonucleotide primer (USV1, Table 1) was de-signed according to the known sequence of the regionupstream from mshB [16] and sequence of the PCR prod-uct obtained with primer HV8 (Table 1) was achieved.Automated dideoxy DNA sequencing was performed onboth strands using the ABI PRISM 310 Genetic Analyzerand BigDye Terminator Cycle Sequencing FS Ready Re-action kit (Applied Biosystems). The program BLAST1.4.9 [17] was used to search for homologous sequencesin the database. Primary amino acid alignments were per-formed using the CLUSTAL W program [18].
2.3. PCR analysis
The presence of a MSHA-like gene cluster in otherV. cholerae non-O1, non-O139 strains was investigated
using the primer combinations DSV 1/DSV6, DSV5/DSV4, and mshQ1/mshQ2. Three microliters of the super-natant of boiled bacterial suspension (ca. 108 bacteriaml31) was added to a 27-Wl PCR mixture consisting of:3 Wl of 10UPCR bu¡er, 25 mM MgCl2, 0.2 mM (of each)dNTP, 0.7 U of Taq DNA polymerase (Gibco BRL) and12.5 pM of each primer. PCR cycle times were as follows:93‡C for 1 min, 55‡C for 1 min, and 72‡C for 1 min (for25 cycles).
2.4. Expression and puri¢cation of recombinant proteins
Plasmid constructs encoding translational fusions be-tween the mouse dihydrofolic acid reductase (DHFR) pro-tein and the MshB or VcfA protein were made by cloninga 5P-terminally truncated version of the coding genes intothe pQE16 vector. PCR was used to amplify a segment ofthe mshB gene using primers BFbg and BRbg and a seg-ment of the vcfA gene by using primers AFbg and ARbg(Table 1). The PCR products were digested with BglIIrestriction enzyme and cloned into the BglII site ofpQE16. Expression of MshO was achieved by amplifyinga segment of the mshO gene with primers OSB and OAH,PCR product digestion with BamHI and HindIII andcloning into the appropriate restriction sites of thepQE30 vector. The recombinant plasmids were trans-formed into E. coli M15. Expression of the (His)6-taggedproteins was induced with 2 mM IPTG and cultured cellslysed with start bu¡er (8 M urea, O.1 M NaH2PO4, 0.01M Tris, 10 mM imidazole, pH 8.0). The soluble fractionwas collected and puri¢ed on a His Trap1 nickel column(Pharmacia) using 500 mM imidazole phosphate bu¡er aselution bu¡er.
2.5. Antisera production
Rabbits were immunized with 100 Wg of puri¢ed pro-teins every 2 weeks. One milliliter of antigen emulsi¢edwith an equal volume of Freund’s complete adjuvantwas injected subcutaneously into multisites. For the boost-ing injections, 100 Wg of antigen and Freund’s incompleteadjuvant was used.
2.6. Construction of hyperpiliated V. cholerae strain
A prepilin-expression vector was constructed to use forfurther transformation into V. cholerae strain 88UDT119as described by Ehara et al. [19]. Primers EXS14 andHV12 (Table 1) were used to amplify the vcfA gene byPCR, and the PCR product was ligated to pCRR2.1 vectorto obtain plasmid pNG14. The 550-bp EcoRI fragment ofpNG14 was subcloned into the EcoRI site of pGEX-3X toobtain pGV3. Plasmid pGV3 was electroporated into com-petent cells of strain 88UDT119.
FEMSIM 1392 23-4-02 Cyaan Magenta Geel Zwart
C. Toma et al. / FEMS Immunology and Medical Microbiology 33 (2002) 35^4036

2.7. NAGV14 and MSHA pili puri¢cation
Except for a few modi¢cations, NAGV14 pili were pu-ri¢ed essentially as described by Ehara et al. [19] fromV. cholerae 88UDT119 carrying plasmid pGV3. Brie£y,bacteria were grown in LB broth supplemented with 100Wg ampicillin ml31 and induced with IPTG for 4 h. Aheavy suspension of the harvest cells in 50 mM Tris^HCl bu¡er (pH 8.0) was agitated in a biomixer and cen-trifuged at 12 000Ug for 30 min. Solid ammonium sulfatewas added to the supernatant at a concentration of 30%saturation, kept at 37‡C overnight, and centrifuged. Thepellet was resuspended and applied to sucrose density gra-dient (10^60% (w/v) in Tris bu¡er) and centrifuged at150 000Ug for 3 h. Fractions rich in pili were pooledand regarded as puri¢ed pili. MSHA pili were puri¢edby the same procedure from V. cholerae Bgd17 carryingplasmid pVPP [19] with the exception that bacteria weregrown in LB broth supplemented with 50 Wg ampicillinml31.
2.8. SDS^PAGE and immunoblotting
SDS^PAGE and immunoblotting were carried out bythe method of Laemmli [20] and Towbin et al. [21], re-spectively. In Western blot analyses, the nitrocellulose
membranes were incubated with appropriate primary anti-bodies and secondary peroxidase-labelled antibodies.Bound antibodies were detected using 4-chloro-1-naph-thol. A prestained molecular marker (New England Bio-labs) was used as a standard.
3. Results
3.1. Nucleotide sequence analysis
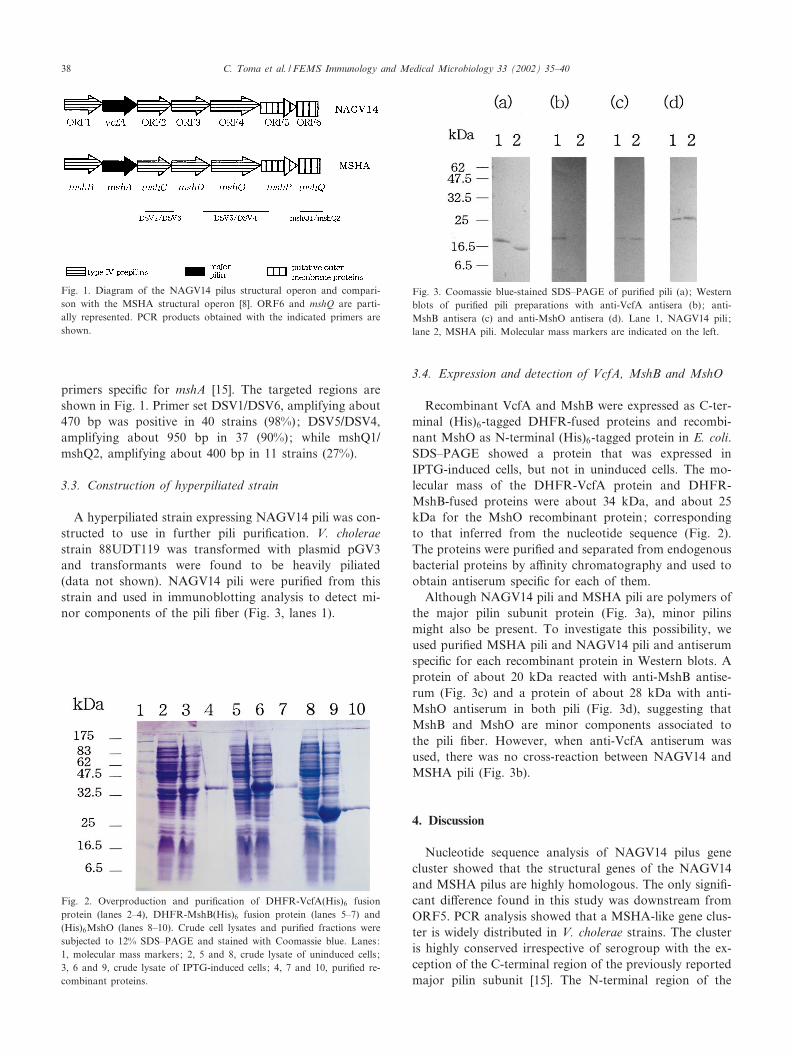
While analyzing the vcfA gene, which encodes the majorpilin subunit of NAGV14 pili, we obtained a PCR productof about 3000 bp using the SalI cassette included in theLA PCR in vitro Cloning kit [15]. The nucleotide sequenceof this fragment revealed ¢ve additional open readingframes (ORFs 2, 3, 4, 5 and 6) of which four were com-plete. The ORFs were in the same transcriptional orienta-tion as vcfA. ORFs 2, 3, 4 and 5 showed high homology toMsh proteins (Table 2). However, the 5P portion of ORF6showed no homology to MshQ. An ORF (ORF1) withhomology to MshB [16] was found upstream of vcfA (Ta-ble 2). The results supported our hypothesis about theoverall operon structure of the NAGV14 pilus gene locus,resembling the gene organization found in the MSHAgene locus [8] (Fig. 1).
All signal sequences of the type IV prepilin proteinsfound in the NAGV14 pilus cluster were identical to thehomologous protein in the MSHA locus, with the excep-tion of ORF2 signal sequence which was longer (15 resi-dues) than the MshC signal sequence (seven residues) [16].The sequence alignments using the CLUSTAL W programrevealed that ORF1 and ORF4 were almost identical toMshB and MshO, respectively, with four residues di¡erentfor ORF1 and two residues di¡erent for ORF4. The se-quence variations of ORF2, ORF3 and ORF5 were pri-marily located in the C-terminal half of the protein, andmore particularly within or around the conserved cysteineresidues. The GenBank accession numbers for the sequen-ces reported in this paper are AB050252 (upstream ofvcfA) and AB049152 (downstream of vcfA).
3.2. PCR analysis
The presence of a MSHA-like gene cluster was exam-ined by PCR in 41 V. cholerae non-O1, non-O139 strainsthat were previously demonstrated to be negative with
Table 1Primers used in PCR, and protein expression
Namea Sequence (5P to 3P)b
USV1 CGTATAATGCACGCCGTACG
HV8 TTGCATGGCGCCTTTTAAGC
DSV1 CCTTATCGAACTGGTAGTGG
DSV6 CAATCATCGGCTTAACAGGC
DSV5 CTGGGTACGGATATTTCCTC
DSV4 CAGAGCATCATGATCAAACC
mshQ1 ACGCTAGTGGGATTGGGATG
mshQ2 GCATAAAGTTGTACAGTCCC
EXS14 GCTGCGTATTCACATGAAAG
HV12 CCCAAGCTTTTAGCAGCCACCTGCATACAC
BFbg CCGAGATCTATTGAGGGCGTTGCAGGGAGC
BRbg CCGAGATCTCTGCAAGGTTACGGTCACTCG
AFbg CCGAGATCTCAAGGCGATGCGCGAGAGGC
ARbg CCGAGATCTGCAGCCACCTGCATACACTG
OSB CCGGGATCCGGCGCGTGAAGCCTTAATCCAG
OAH CCCAAGCTTTTACGGTGTGTTGTAGACCAG
aAll primers were designed in this study with the exception of primersHV8 and HV12 [15].bUnderlined letters indicate restriction endonuclease sites.
Table 2Characteristics of the ORFs
ORF Number of bp Calculated molecular mass of product (Da) Homologous protein (percent of similarity)
ORF1 555 19745 MshB (88%)ORF2 513 17749 MshC (78%)ORF3 567 20260 MshD (89%)ORF4 771 27775 MshO (88%)ORF5 417 14986 MshP (78%)
FEMSIM 1392 23-4-02 Cyaan Magenta Geel Zwart
C. Toma et al. / FEMS Immunology and Medical Microbiology 33 (2002) 35^40 37
primers speci¢c for mshA [15]. The targeted regions areshown in Fig. 1. Primer set DSV1/DSV6, amplifying about470 bp was positive in 40 strains (98%); DSV5/DSV4,amplifying about 950 bp in 37 (90%); while mshQ1/mshQ2, amplifying about 400 bp in 11 strains (27%).
3.3. Construction of hyperpiliated strain
A hyperpiliated strain expressing NAGV14 pili was con-structed to use in further pili puri¢cation. V. choleraestrain 88UDT119 was transformed with plasmid pGV3and transformants were found to be heavily piliated(data not shown). NAGV14 pili were puri¢ed from thisstrain and used in immunoblotting analysis to detect mi-nor components of the pili ¢ber (Fig. 3, lanes 1).
3.4. Expression and detection of VcfA, MshB and MshO
Recombinant VcfA and MshB were expressed as C-ter-minal (His)6-tagged DHFR-fused proteins and recombi-nant MshO as N-terminal (His)6-tagged protein in E. coli.SDS^PAGE showed a protein that was expressed inIPTG-induced cells, but not in uninduced cells. The mo-lecular mass of the DHFR-VcfA protein and DHFR-MshB-fused proteins were about 34 kDa, and about 25kDa for the MshO recombinant protein; correspondingto that inferred from the nucleotide sequence (Fig. 2).The proteins were puri¢ed and separated from endogenousbacterial proteins by a⁄nity chromatography and used toobtain antiserum speci¢c for each of them.
Although NAGV14 pili and MSHA pili are polymers ofthe major pilin subunit protein (Fig. 3a), minor pilinsmight also be present. To investigate this possibility, weused puri¢ed MSHA pili and NAGV14 pili and antiserumspeci¢c for each recombinant protein in Western blots. Aprotein of about 20 kDa reacted with anti-MshB antise-rum (Fig. 3c) and a protein of about 28 kDa with anti-MshO antiserum in both pili (Fig. 3d), suggesting thatMshB and MshO are minor components associated tothe pili ¢ber. However, when anti-VcfA antiserum wasused, there was no cross-reaction between NAGV14 andMSHA pili (Fig. 3b).
4. Discussion
Nucleotide sequence analysis of NAGV14 pilus genecluster showed that the structural genes of the NAGV14and MSHA pilus are highly homologous. The only signi¢-cant di¡erence found in this study was downstream fromORF5. PCR analysis showed that a MSHA-like gene clus-ter is widely distributed in V. cholerae strains. The clusteris highly conserved irrespective of serogroup with the ex-ception of the C-terminal region of the previously reportedmajor pilin subunit [15]. The N-terminal region of the
Fig. 1. Diagram of the NAGV14 pilus structural operon and compari-son with the MSHA structural operon [8]. ORF6 and mshQ are parti-ally represented. PCR products obtained with the indicated primers areshown.
Fig. 2. Overproduction and puri¢cation of DHFR-VcfA(His)6 fusionprotein (lanes 2^4), DHFR-MshB(His)6 fusion protein (lanes 5^7) and(His)6MshO (lanes 8^10). Crude cell lysates and puri¢ed fractions weresubjected to 12% SDS^PAGE and stained with Coomassie blue. Lanes:1, molecular mass markers; 2, 5 and 8, crude lysate of uninduced cells ;3, 6 and 9, crude lysate of IPTG-induced cells ; 4, 7 and 10, puri¢ed re-combinant proteins.
Fig. 3. Coomassie blue-stained SDS^PAGE of puri¢ed pili (a); Westernblots of puri¢ed pili preparations with anti-VcfA antisera (b); anti-MshB antisera (c) and anti-MshO antisera (d). Lane 1, NAGV14 pili ;lane 2, MSHA pili. Molecular mass markers are indicated on the left.
FEMSIM 1392 23-4-02 Cyaan Magenta Geel Zwart
C. Toma et al. / FEMS Immunology and Medical Microbiology 33 (2002) 35^4038

mshQ gene seemed to be also variable among di¡erentstrains, since primers targeted to this region gave a pos-itive reaction only in 27% of the strains. Sequence varia-bility has been also observed in the type IV pilus-majorpilin subunit of Pseudomonas aeruginosa [22], E. coli [23]and Neisseria spp. [24]. However, the level of sequencediversity of the other pilus cluster genes have not beenanalyzed for these species.
Many organisms express type IV pili on their surfaces,but little is known about the biogenesis of these organelles.Type IV pili are usually composed of a single repeatingpilin protein. With the exception of Neisseria gonorrhoeae,which has an adhesin protein located at the tip of the pilus[25], and E. coli strains that express the R64 thin pilus,which has a minor pilus component [5]. In MSHA pili, thehemagglutinating activity was proved to be MshA-depen-dent because inframe mutation in the mshA gene wasfound to abolish this activity [16]. In N. gonorrhoeaetype IV pili, the major pilin subunit (PilE) is responsiblefor the pili mediated hemagglutination, while the tip-lo-cated adhesin PilC seemed to be crucial for bacteria tointeract with the epithelial cells [25]. Similarly, V. choleraetype IV pili might be composed of more than one kind ofpilin subunit, as was suggested from deduced amino acidsequence analyses by other investigators [16,18]. To verifythis hypothesis, we puri¢ed NAGV14 and MSHA pilifrom hyperpiliated strains to further use them in Westernblotting analyses.
Recombinant MshB and MshO proteins were puri¢edand speci¢c antiserum was raised to each of them. Westernblotting analyses showed that these antisera reacted withpuri¢ed NAGV14 and MSHA pili. The results suggestedthat MshB and MshO are minor components of the pili¢ber and supported the idea that the coding genes arehighly conserved among di¡erent strains. The results arequite consistent with the observations that mshB andmshO products have features of type IV prepilins. Theyhave putative type IV prepilin peptidase cleavage sites.PilD has been reported to be the prepilin peptidase essen-tial for MSHA pili production [9]. We also con¢rmed thepresence of pilD gene in V. cholerae NAGV14 strain byPCR and sequencing (data not shown). Although we didnot clarify the roles of these minor components, by anal-ogy with other pili they may act as adhesin, aid in initiat-ing new pili or regulate pilus length [26]. MshB and MshOare highly conserved among di¡erent strains, but theymight have di¡erent roles depending on the strain. Fim-brial shafts were shown to in£uence sugar-binding specif-icities of their FimH adhesin for E. coli type I pili [27].Therefore, certain domains of the minor component/smight be masked or might be presented di¡erently, de-pending on the context of the major pilin subunits. Thedi¡erence between the major pilin of MSHA andNAGV14 pili found in the C-terminal region might in£u-ence the recognition of the receptors on the epithelial cellsand explain why NAGV14 pili is adhesive. The fact that
MshB, a minor component of about 20 kDa, was piliassociated could also explain the cross-reaction found inV. cholerae O1 strains by Yamashiro and Iwanaga whenusing anti-NAGV14 pilus serum [14].
In this paper, we demonstrated for the ¢rst time thatV. cholerae type IV MSHA pili and NAGV14 pili, are nothomopolymers of a single subunit protein, but have otherminor components associated with the pili ¢ber. There-fore, the heteropolymeric nature of the Neisserial pili re-ported by Rudel et al. [25] also seemed to be applicable forV. cholerae pili. The results presented here have importantimplications for our understanding of type IV pilus struc-ture, and the role of pilin variation in pathogenic mecha-nism.
Acknowledgements
This work was supported by a Grant-in Aid for Scien-ti¢c Research from the Ministry of Education, Culture,Sports, Science and Technology, Japan.
References
[1] Albert, M.J. (1994) Vibrio cholerae O139 Bengal. J. Clin. Microbiol.32, 2345^2349.
[2] Sharma, C., Thungapathra, M., Ghosh, A., Mukhopadhyay, A.K.,Basu, A., Mitra, R., Basu, I., Bhattacharya, S.K., Shimada, T.,Ramamurthy, T., Takeda, T., Yamasaki, S., Takeda, Y. and Nair,G.B. (1998) Molecular analysis of non-O1, non-O139 Vibrio choleraeassociated with an unusual upsurge in the incidence of cholera-likedisease in Calcutta, India. J. Clin. Microbiol. 36, 756^763.
[3] Strom, M.S. and Lory, S. (1993) Structure^function and biogenesis ofthe type IV pili. Annu. Rev. Microbiol. 47, 565^596.
[4] Kirn, T.J., La¡erty, M.J., Sandoe, C.M.P. and Tayler, R.K. (2000)Delineation of pilin domains required for bacterial association intomicrocolonies and intestinal colonization by Vibrio cholerae. Mol.Microbiol. 35, 896^910.
[5] Yoshida, T., Furuya, N., Ishikura, M., Isobe, T., Haino-Fukushima,K., Ogawa, T. and Komano, T. (1998) Puri¢cation and character-ization of thin pili of Incl1 plasmids ColIb-P9 and R64: formation ofPilV-speci¢c cell aggregates by type IV pili. J. Bacteriol. 180, 2842^2848.
[6] Merz, A.J., So, M. and Sheetz, M.P. (2000) Pilus retraction powersbacterial twitching motility. Nature 407, 98^101.
[7] Karaolis, D.K.R., Somara, S., Maneval Jr, D.R., Johnson, J.A. andKaper, J.B. (1999) A bacteriophage encoding a pathogenicity island,a type-IV pilus and a phage receptor in cholera bacteria. Nature 399,375^379.
[8] Marsh, J.W. and Taylor, R.K. (1999) Genetic and transcriptionalanalyses of the Vibrio cholerae mannose-sensitive hemagglutinintype 4 pilus gene locus. J. Bacteriol. 181, 1110^1117.
[9] Fullner, K.J. and Mekalanos, J.J. (1999) Genetic characterization ofa new type IV-A pilus gene cluster found in both classical and El Torbiotypes of Vibrio cholerae. Infect. Immun. 67, 1393^1404.
[10] Novais, R.C., Coelho, A., Salles, C.A. and Vicente, A.C.P. (1999)Toxin-co-regulated pilus cluster in non-O1, non-toxigenic Vibrio cho-lerae : evidence of a third allele of pilin gene. FEMS Microbiol. Lett.171, 49^55.
[11] Iwanaga, M., Nakasone, N. and Ehara, M. (1989) Pili of Vibrio
FEMSIM 1392 23-4-02 Cyaan Magenta Geel Zwart
C. Toma et al. / FEMS Immunology and Medical Microbiology 33 (2002) 35^40 39

cholerae O1 biotype El Tor: a comparative study on adhesive andnon-adhesive strains. Microbiol. Immunol. 33, 1^9.
[12] Nakasone, N. and Iwanaga, M. (1990) Pili of Vibrio cholerae non-O1.Infect. Immun. 58, 1640^1646.
[13] Yamashiro, T., Nakasone, N. and Iwanaga, M. (1993) Puri¢cationand characterization of pili of a Vibrio cholerae non-O1 strain. Infect.Immun. 61, 5398^5400.
[14] Yamashiro, T. and Iwanaga, M. (1996) Puri¢cation and character-ization of a pilus of a Vibrio cholerae strain: a possible colonizationfactor. Infect. Immun. 64, 5233^5238.
[15] Kuroki, H., Toma, C., Nakasone, N., Yamashiro, T. and Iwanaga,M. (2001) Gene analysis of Vibrio cholerae NAGV14 pilus and itsdistribution. Microbiol. Immunol. 45, 417^424.
[16] Jonson, G., Lebens, M. and Holmgren, J. (1994) Cloning and se-quencing of Vibrio cholerae mannose-sensitive haemagglutinin pilingene: localization of mshA within a cluster of type 4 pilin genes. Mol.Microbiol. 13, 109^118.
[17] Altschul, S.F., Madden, T.L., Scha¡er, A.A., Zhang, J., Zhang, Z.,Miller, W. and Lipman, D.J. (1997) Gapped BLAST and PSI-BLAST: a new generation of protein database search programs. Nu-cleic Acids Res. 25, 3389^3402.
[18] Thompson, J.D., Higgins, D.G. and Gibson, T.J. (1994) CLUSTALW: improving the sensitivity of progressive multiple sequence align-ment through sequence weighting, position-speci¢c gap penalties andweight matrix choice. Nucleic Acids Res. 22, 4673^4680.
[19] Ehara, M., Iwami, M., Ichinose, Y. and Hirayama, T. (2000) Devel-opment of hyper¢mbriated strains of Vibrio cholerae O1. Microbiol.Immunol. 44, 439^446.
[20] Laemmli, U.K. (1970) Cleavage of structural proteins during theassembly of the head of bacteriophage T4. Nature 227, 680^685.
[21] Towbin, H., Staehelin, T. and Gordon, J. (1979) Electrophoretictransfer of proteins from polyacrylamide gels to nitrocellulose sheets:procedure and source applications. Proc. Natl. Acad. Sci. USA 76,4350^4354.
[22] Hahn, H.P. (1997) The type-4 pilus is the major virulence-associatedadhesin of Pseudomonas aeruginosa ^ a review. Gene 192, 99^108.
[23] Blank, T.E., Zhong, H., Bell, A.L., Whittam, T.S. and Donnenberg,M.S. (2000) Molecular variation among type IV pilin (bfpA) genesfrom diverse enteropathogenic Escherichia coli strains. Infect. Immun.68, 7028^7038.
[24] Nassif, X., Lowy, J., Stenberg, P., O’Gaora, P., Ganji, A. and So, M.(1993) Antigenic variation of pilin regulates adhesion of Neisseriameningitidis to human epithelial cells. Mol. Microbiol. 8, 719^725.
[25] Rudel, T., Scheuerp£ug, I. and Meyer, T.F. (1995) Neisseria PilCprotein identi¢ed as type-4 pilus tip-located adhesin. Nature 373,357^359.
[26] Thanassi, D.G. and Hultgren, S.J. (2000) Assembly of complex or-ganelles: pilus biogenesis in Gram-negative bacteria as a model sys-tem. Methods 20, 111^126.
[27] Madison, B., Ofek, I., Clegg, S. and Abraham, S.N. (1994) Type 1¢mbrial shafts of Escherichia coli and Klebsiella pneumoniae in£uencesugar-binding speci¢cities of their FimH adhesins. Infect. Immun. 62,843^848.
FEMSIM 1392 23-4-02 Cyaan Magenta Geel Zwart
C. Toma et al. / FEMS Immunology and Medical Microbiology 33 (2002) 35^4040



![Vibrio cholerae - As Biological Weapons · Vibrio cholerae - as biological weapons Cholera (Vibrio cholerae) ± MDNREUR ELRORJLF]QD miotr Daniszewski Department of Invertebrate Zoology](https://img.pdfslide.net/doc/110x75/60abe1c9df963953a24dc9d3/vibrio-cholerae-as-biological-weapons-vibrio-cholerae-as-biological-weapons.jpg)