Embed Size (px)
Citation preview

Mitochondrial DNA from Prehistoric Canids Highlights RelationshipsBetween Dogs and South-East European Wolves
Fabio Verginelli,* Cristian Capelli,� Valentina Coia,� Marco Musiani,§ Mario Falchetti,kLaura Ottini,k Raffaele Palmirotta,*1 Antonio Tagliacozzo,{ Iacopo De Grossi Mazzorin,# andRenato Mariani-Costantini**Department of Oncology and Neurosciences, University G. d’Annunzio, and Center of Excellence on Aging (Ce.S.I.), G. d’AnnunzioFoundation, Chieti, Italy; �Institute of Legal Medicine, Catholic University of S. Cuore, Rome, Italy; �Department of Animal andHuman Biology, University ‘‘La Sapienza,’’ Rome, Italy; §Faculty of Environmental Design, University of Calgary, Calgary, Alberta,Canada; kDepartment of Experimental Medicine and Pathology (Section of History of Medicine), University La Sapienza, Rome,Italy; {Laboratory of Quaternary Paleontology and Archeozoology, Special Superintendence to the Luigi Pigorini NationalPrehistoric-Ethnographic Museum, Rome, Italy; and #Department of Cultural Heritage, University of Lecce, Lecce, Italy
The question of the origins of the dog has been much debated. The dog is descended from the wolf that at the end of the lastglaciation (the archaeologically hypothesized period of dog domestication) was one of the most widespread amongHolarctic mammals. Scenarios provided by genetic studies range from multiple dog-founding events to a single originin East Asia. The earliest fossil dogs, dated ’17–12,000 radiocarbon (14C) years ago (YA), were found in Europe and inthe Middle East. Ancient DNA (a-DNA) evidence could contribute to the identification of dog-founder wolf populations.To gain insight into the relationships between ancient European wolves and dogs we analyzed a 262-bp mitochondrialDNA control region fragment retrieved from five prehistoric Italian canids ranging in age from ’15,000 to ’3,000 14CYA. These canids were compared to a worldwide sample of 547 purebred dogs and 341 wolves. The ancient sequenceswere highly diverse and joined the three major clades of extant dog sequences. Phylogenetic investigations highlightedrelationships between the ancient sequences and geographically widespread extant dog matrilines and between the ancientsequences and extant wolf matrilines of mainly East European origin. The results provide a-DNA support for the involve-ment of European wolves in the origins of the three major dog clades. Genetic data also suggest multiple independentdomestication events. East European wolves may still reflect the genetic variation of ancient dog-founder populations.
Introduction
The dog (Canis familiaris L.), considered the first do-mestic animal, is descended from the grey wolf (Canis lu-pus L.), a highly mobile and adaptable carnivore that up tofew centuries ago was widespread throughout the Holarctic(Kurten 1968; Olsen 1985; Clutton-Brock 1995; Vila,Maldonado, and Wayne 1999). The places and dates of wolfdomestication are much debated. No dog images have beenidentified in the Franco-Cantabric cave art, which stronglysuggests that before ’16,000 radiocarbon (14C) yearsago (YA) the dog was still unknown in Western Europe(Delporte 1990; Nougier 1993; Clutton-Brock 1995). Theearliest dog fossils consist in two crania from Eliseyevichi1 (Central Russian Plain, 13–17,000 14C YA) and in a man-dible from Oberkassel (Germany, ’14,000 14C YA) (Nobis1979; Sablin and Khlopachev 2002). These represent robustwolf-sized dogs thought to have derived from the largenorthern Holarctic wolf formC. l. lupus. Remains of smallerdogs from Mesolithic (Natufian) cave deposits in the Mid-dle East (’12,000 14C YA) were interpreted as evidenceof independent domestication from a lighter SouthwestAsian wolf form, Canis lupus arabs (Davis and Valla1978; Dayan 1994; Clutton-Brock 1995; Tcherncov andValla 1997). Images in the Saharan and Iberic Epipaleo-lithic rock art and skeletal remains indicate that, by’10,000 14C YA, dogs were already present in areas as
far apart as North Africa, Western Europe, and North Amer-ica (Lindner 1950; Kurten and Anderson 1980; Clutton-Brock 1995; Schwartz 1997).
The striking phenotypic and genetic diversity of dogsclearly indicates that their founders were recruited froma large and varied wolf population (Clutton-Brock 1995;Wayne and Ostrander 1999; Parker et al. 2004). Mitochon-drial DNA (mtDNA) analyses have been used to elucidaterelationships between dog and wolf. Phylogenetic trees ofdog and wolf hypervariable region 1 (HVR1) sequencesshow that dogs group into several clades (Tsuda et al.1997; Vila et al. 1997; Leonard et al. 2002; Savolainenet al. 2002), a fact that clearly indicates independent originsfrom multiple wolf matrilines. Vila et al. (1997), usinga fixed substitution rate mtDNA molecular clock calibratedon a 1 million years age for wolf-coyote divergence (Kurtenand Anderson 1980), estimated that dogs and wolves sep-arated as early as 76–135,000 YA. Based on the same cal-ibration and focusing on worldwide HVR1 sequencevariation among dogs, Savolainen et al. (2002) proposedthat dogs diverged from wolves 40–15,000 YA and weremost probably domesticated in East Asia. This suggests thatthe available archaeozoological record could be biased to-ward West Eurasia because of the more extensive fieldworkconducted there. Concomitantly, Leonard et al. (2002)showed that pre-Columbian New World dog lineages de-rived from Eurasia and, more recently, Savolainen et al.(2004) provided substantial evidence that Australian din-goes originated from a population of East Asian dogs.The molecular time estimates of dog origins cited aboveare challenged by newly uncovered evidence showing thatclocks calibrated on phylogenetic substitution rates cannotbe applied to the dating of geologically recent divergenceevents (Ho et al. 2005).
1 Present address: Laboratory of Clinical Pathology, Regina ElenaCancer Institute, Rome, Italy.
Key words: dog, wolf, mtDNA, ancient DNA, archaeozoology,evolution.
E-mail: [email protected].
Mol. Biol. Evol. 22(12):2541–2551. 2005doi:10.1093/molbev/msi248Advance Access publication August 24, 2005
� The Author 2005. Published by Oxford University Press on behalf ofthe Society for Molecular Biology and Evolution. All rights reserved.For permissions, please e-mail: [email protected]
at UIC
Library, C
ollections Developm
ent on Novem
ber 15, 2014http://m
be.oxfordjournals.org/D
ownloaded from

For several millennia the breeding and geographic dis-tribution of dogs depended on man, while in the last fewcenturies human persecution caused the extinction or bot-tlenecking of most wolf populations (Vila et al. 1999; Randiet al. 2000; Flagstad et al. 2003). Thus, the genetics of dogsand wolves bears the historic influence of humans, a strongconfounding factor that can be circumvented only byancient DNA (a-DNA) studies (Hofreiter et al. 2001b;Leonard et al. 2002). A-DNA has shed novel light onthe evolutionary history of several species, notably includ-ing the Neanderthal, large Pleistocene carnivores, such asbears and hyenas, and domestic animals, such as horses,cattle, and native North American and Island SoutheastAsian dogs (Greenwood et al. 1999; Leonard, Wayne,and Cooper 2000; Hofreiter et al. 2001b, 2004; Troyet al. 2001; Vila et al. 2001; Jansen et al. 2002; Leonardet al. 2002; Savolainen et al. 2004).
Because of the geographic position and archaeozoo-logical record, Italy is a country of potential relevancefor dog evolutionary studies (Sauter 1948; Kurten 1968;Gamble 1986). The present study is centered on novel a-DNA evidence from five prehistoric Italian canids (PICs)ranging in age from ’15,000 to ’3,000 14C YA. The an-cient HVR1 sequences were pooled with the sequences of341 extant wolves and 547 purebred dogs retrieved fromdatabases. Our results provide novel insight into the rela-tionships between dogs and wolf populations.
Materials and MethodsSpecimens and Radiocarbon Dating
PIC specimens were provided by the Luigi PigoriniNational Prehistoric-Ethnographic Museum (LPM), Rome,and by the Laboratory of Archaeozoology (LA), Archaeo-logical Superintendence of Rome. A-DNA could be re-trieved from five of eight tested specimens, includingPIC-1 (LPM), a mandibular ramus of large canid (no asso-ciated teeth) from the Late Glacial levels of the PalidoroUpper Paleolithic rock shelter, Latium, central Italy (Bietti1976–1977; Cassoli 1976–1977); PIC-2 (LPM), a fragmentof large canid metapodial from the Early Holocene ‘‘terrebrune’’ level of Romanelli Cave, a well-known Upper Pa-leolithic site in Apulia, southern Italy; PIC-3 (LPM), a frag-ment of large canid tibia also from the ‘‘terre brune’’ ofRomanelli Cave (Blanc 1920, 1928; Cassoli, Fiore, andTagliacozzo 1994); PIC-4 (LPM), a tibia from a relativelycomplete dog skeleton found at the Casal del Dolce Eneo-lithic site, Latium (Fiore and Tagliacozzo 1997); PIC-5(LA), a dog femur from the Vejano Recent Bronze Age site,Latium (De Grossi Mazzorin and Tagliacozzo 2000). PIC-1through -3 were associated with cold final Pleistocene faunaand Epigravettian Upper Paleolithic industry. These threePIC specimens were within the range of morphometricvariability of the Late Upper Pleistocene Italian wolves(Cassoli, Fiore, and Tagliacozzo 1994) but lacked diagnos-tic features that could exclude attribution to large proto-dogs (Sablin and Khlopachev 2002). PIC-4 and PIC-5 wereunambiguously attributable to medium-sized dogs, with nobreed differentiation evidenced after analysis of the PIC-4skeleton (Fiore and Tagliacozzo 1997; De Grossi Mazzorinand Tagliacozzo 2000). Specimens were radiocarbon dated
from purified collagen by using accelerator mass spec-trometry at BETA Analytic (Miami, Fla.). Reporteddates are 14C years before A.D. 1950, corrected for13C fractionation.
A-DNA Retrieval, Analysis, and Authentication
Bone surfaces were scraped with sterile blades, UVirradiated for 1 h, and microtrephined to obtain at least1 g of bone powder, stored in sealed 250 mg aliquots untiluse. Bone powder aliquots were washed overnight in0.5 M ethylenediaminetetraacetic acid (EDTA) (pH 8.0),digested at 37�C for 48 h in 600 ll of proteinase K(20 mg/ml in 0.5 M EDTA [pH 8.0]/5% sodium dodecylsulfate), and extracted as in Krings et al. (1997). Two nestedpartially overlapping polymerase chain reactions (PCRs)were used to amplify a 299-bp (with primers) canineHVR1 fragment encompassing the 262-bp segment (nucle-otide positions [nps] 15431–15687) utilized for phyloge-netic analyses. External (ext) and internal (int) primers,identified aligning 26 dog and 27 wolf haplotypes fromVila et al. (1997), and sizes of PCR products wereL15422extCTCTTGCTCCACCATCAGC-3#/H15548extTTATATGCATGGGGCAAACC-3#, 173 bp; L15426int-CTCCACCATCAGCACCC-3#/H15555intATGGGGCA-AACCATTAATG-3#, 157 bp; L15511extACTGTGCTA-TGTCAGTATCTCC-3#/H15691extTTGATGGTTTCTC-GAGCC-3#, 220 bp; and L15529intTCCAGGTAAACC-CTTCTTCCC-3#/H15692intGATGGTTTCTCGAGGCA-3#, 199 bp. PCRs were carried out in 40-ll volumes with 10mM Tris (pH 8.3), 1.5 mM MgCl2, 200 lM deoxynucleo-side triphosphates, 1.5 units of AmpliTaq Gold (AppliedBiosystems, Foster City, Calif.), 10 pM of each primer,and 4 ll of a-DNA extract. External PCR conditions were50 cycles of 50�C for 45 s, 72�C for 45 s, and 94�C for 30 s;for internal PCRs (19 cycles) annealing was at 55�C. Pri-mers produced no amplifications when tested on humanDNA; conversely, primers designed for human mtDNAdid not yield amplifications when tested on the a-DNAs.PCR products were directly sequenced on an ABI-PRISM310 Sequencer (Applied Biosystems). Contamination wasstrictly monitored with extraction and PCR blanks.
Some of the main a-DNA authentication criteria, in-cluding the most stringent (i.e., independent replication),were followed (Hofreiter et al. 2001a). All PIC specimenswere independently sampled, extracted, PCR amplified,and sequenced in Chieti (University G. d’Annunzio) andin Rome (Catholic University of S. Cuore). In both labora-tories, a-DNA extractions and amplifications were per-formed in separate dedicated spaces where no moderncanid samples had been manipulated (setup of PCR and se-quencing conditions on modern dog DNA was performedin a third laboratory). Reagents, bench surfaces, and non-disposable equipment were routinely sterilized under254-nm UV light. Chieti and Rome generated independentamplification products for the two partially overlappingHVR1 segments analyzed; consensus sequences were ob-tained by alignment of at least two independent direct se-quences. Identical sequences were replicated from the PIC-3 a-DNA extracts; discrepancies in 3, 2, 1, and 1 nt wereobserved for the PIC-1, PIC-2, PIC-4, and PIC-5 sequences,
2542 Verginelli et al.
at UIC
Library, C
ollections Developm
ent on Novem
ber 15, 2014http://m
be.oxfordjournals.org/D
ownloaded from

respectively. Nucleotide substitutions due to template dam-age occur randomly, and it is expected that multiple ampli-fications of the same fragment will not show the samerandom damage substitutions. Thus, additional amplifica-tions were performed to identify the reproducible variants.Irreproducible variants were always C to T or G to A sub-stitutions that most likely correspond to artifacts due to tem-plate damage (Krings et al. 1997; Hofreiter et al. 2001a).
Phylogenetic Analysis
Frequencies, geographic origins, and GenBank acces-sion numbers of the extant wolf and dog HVR1 sequencesused in this study are given in Tables S1–S4 (see the Sup-plementary Material online). Extant wolf sequences werefrom 341 individuals (Europe, 256; Asia, 66; America,19 [Tsuda et al. 1997; Vila et al. 1997; Randi et al.2000; Savolainen et al. 2002]). Extant dog sequences werefrom 547 purebred dogs, grouped according to continent asindicated by breed (Europe, 231; Asia, 255; Africa, 31;America, 30). Outbred dogs were not considered becauseof greater uncertainty about origins. PIC sequences (262 bp,nps 15431–15687) were pooled with unique GenBankHVR1 sequences (262–664 bp, nps 15431–15687/16039,Tables S1–S4, Supplementary Material online) of dogs(92 sequences), wolves (53 sequences), and coyotes (2sequences). To avoid collapse of previously identifieddog clades (Tsuda et al. 1997; Vila et al. 1997; Leonardet al. 2002; Savolainen et al. 2002, 2004), sequences wereevaluated along their full lengths, filling gaps with un-known nucleotides for shorter sequences. Arlequin 2000(Schneider, Roessli, and Excoffier 2000) was used to verifyhaplotype sharing between ancient and extant sequences andto perform Fu’s Fs tests of selective neutrality for identifiedclades. The phylogenetic trees were constructed throughNeighbor-Joining (NJ) algorithm using PHYLIP 3.6 Infer-ence Package (Felsenstein 2002). Distance matrixes werebased on the F84 model with a c-distribution for substitutionrate heterogeneity (Felsenstein 1984). The c-shape and a-transition/transversion parameters were set to 0.10 and9.94, respectively, as estimated using Tree-Puzzle 5.0(Schmidt et al. 2002). Significance was determined frombootstrap percentages obtained after replication of 100 trees.To obtain median-joining (MJ) networks of dog clades con-taining PICs, the DnaSP v.3 software (available at http://www.ub.es/dnasp/) was applied to sequence data to identifysegregating variants in the 262-bp PIC segment. Tables ofvariants were used in the MJ algorithm option of NET-WORK 4.000 (Bandelt, Forster, and Rohl 1999). The wolfMJ network was constructed as above, based on variantsin the 262-bp PIC segment (44 sequences including 16 fromEurope, 20 from Asia, 6 from America, and 2 shared be-tween Europe and Asia [Tsuda et al. 1997; Vila et al.1997; Randi et al. 2000; Savolainen et al. 2002]). To avoidhigh-dimensional cubes, rapidly mutating variants at nps15625 and 15643 were 10-fold down-weighted. Maximumparsimony (MP) algorithm was applied to purge the wolfMJ network of superfluous links. In all cases, default set-tings were r 5 2 and e 5 0 (most parsimonious pathway).
Networks based on the 262-bp PIC segment werealso constructed by MP method using TCS version 1.13
(Clement, Posada, and Crandall 2000), where the probabil-ity of parsimony, calculated for pairwise differences up tothe 95% cutoff level, justifies the maximum number of mu-tational connections between pairs of sequences (indelsconsidered as a fifth state). Rooting weights were calculatedusing the algorithm implemented in the TCS program,based on the frequency of each candidate root haplotypeand on the number and frequencies of the haplotypes towhich it is connected (Castelloe and Templeton 1994).
Spatial structure in haplotype distribution was exam-ined with conventional F-statistics (Wright 1951) andhierarchical analysis of molecular variance (AMOVA;Excoffier, Smouse, and Quattro 1992). We calculated thematrixes of pairwise Fst values between sampling locations.Locations of dog and wolf sequences were assigned accord-ing to area of origin for the dog breed and/or according tothe references listed in Tables S1 and S2 (see the Supple-mentary Material online). Such locations were definedbased on the latitude and longitude of the center of thecountry (or continental area) of origin. In AMOVA, wetested statistical significance after 1,000 permutations forthe correlation of random genotypes within spatially clus-tered groups relative to the whole population. AMOVA uti-lizes both the frequency and sequence divergence betweengenotypes. We estimated the divergence between geno-types assuming a Tamura-Nei model of sequence evolutionand a gamma distribution of the substitution rates witha value of a 5 0.5 (Tamura and Nei 1993; Wakeley1993). A matrix of pairwise Euclidean distances betweenthe central locations for each group was also calculated(mean latitude and longitude for relevant sampling loca-tions). Finally, we obtained other matrixes by calculatingall pairwise geographic distances imposing a theoreticalpassage through the Bering Strait. Genetic and geographicmatrixes were tested for correlation using Mantel’s test(Liedloff 1999). Arlequin 2000 (Schneider, Roessli, andExcoffier 2000) and Genetix 4.05 (Belkhir et al. 2004) soft-ware were used for computations.
Results
We succeeded in retrieving 262-bp HVR1 sequencesfrom five PICs that included PIC-1 (Late Glacial, 14,670 6130 14C YA), PIC-2 (Early Holocene, 9,860 6 50 14C YA),PIC-3 (Early Holocene 9,670 6 40 14C YA), PIC-4 (Eneo-lithic, 4,110 6 40 14C YA), and PIC-5 (Recent Bronze Age,3,040 6 40 14C YA). These PICs yielded individual se-quences that varied by 2–11 nt at 13 polymorphic sites(all variants occur in GenBank wolf and/or dog sequences).Independent replication of these sequences strongly sup-ports their authenticity.
To highlight similarity relationships between PICsequences and modern wolf and dog HVR1 sequences(262–664 bp), we constructed an NJ tree rooted usingtwo coyote sequences. The NJ tree split dogs (92 haplo-types) and wolves (53 haplotypes) into nine clades, identi-fied with Roman numerals in figure 1. Clades I, II, IV, VI,and IX corresponded to previously defined dog-dominatedor dog-monophyletic clades (Tsuda et al. 1997; Vila et al.1997; Leonard et al. 2002; Savolainen et al. 2002, 2004).Clade I, the main dog clade, contained 56 dog haplotypes
Ancient DNA and Dog Origins 2543
at UIC
Library, C
ollections Developm
ent on Novem
ber 15, 2014http://m
be.oxfordjournals.org/D
ownloaded from

from Eurasia, the Americas, and Africa, 2 haplotypesshared between European wolves and dogs, and 3 Asianwolf haplotypes. Clade II comprised one West Asian andfive European dog haplotypes. Clade IV encompassed eightdog haplotypes found in Eurasian, American, and Africanbreeds. Clade VI, a dog-dominated clade comprising sev-eral wolves, included 15 dog haplotypes from Europe andAsia, 1 haplotype shared between European wolves anddogs, and 4 wolf haplotypes (three from Europe and onefrom Asia). Minor dog clade IX encompassed three EastAsian haplotypes. Clades III, V, VII, and VIII included onlywolves, with the exception of a dog haplotype embedded inclade VIII (Savolainen et al. 2002). Clade III included fiveEuropean wolf haplotypes (among which the single haplo-type of extant Italian/French wolves [Vila et al. 1997; Randiet al. 2000; Lucchini et al. 2002]). Minor clades V and VIIcontained three wolf haplotypes each (two from Asia andone from Europe [clade V] and two from America and onefrom Europe [clade VI]). Clade VIII, the main wolf clade,comprised 29 haplotypes from the whole wolf geographicrange and 1 East Asian dog haplotype. Wolf haplotypes W5(Mongolia), W10 (China), and W13 (China) were outsidedefined clades. Dog clades II, IV, and IX had bootstrap sup-port over 50%. Excluding wolf sequences in our bootstrapanalysis gave over 50% support also for dog clade VI (datanot shown). Because of extensive homoplasy, bootstrapsupport for clade I and for wolf clades could not be obtained(Savolainen et al. 2002).
The 262-bp PIC sequences clustered with major dogclades I, IV, and VI. PIC-1 (Late Glacial) and PIC-2 (EarlyHolocene) showed novel haplotypes. PIC-1 occupied asister branch of clade IV, PIC-2 clustered in clade VI,and PIC-3 (Early Holocene) clustered in clade I and resultedidentical with a reported a-DNA sequence retrieved frompre-European contact Alaskan dogs (Leonard et al. 2002;Savolainen et al. 2002). The PIC-3 sequence was sharedby several Asian dog haplotypes and by W51-A11, anintriguing haplotype found in a Bulgarian wolf and indog breeds with ancient Old and New World origins(Vila et al. 1997; Randi et al. 2000; Leonard et al. 2002;Savolainen et al. 2002). PIC-4 (Eneolithic) was sharedby four Old World dog haplotypes in clade IV andPIC-5 (Bronze Age) by five widespread dog haplotypesin clade I.
To define the evolutionary hierarchies of the haplo-types within major dog clades I, IV, and VI, we constructed
FIG. 1.—NJ tree of PIC and extant wolf and dog HVR1 sequences.Roman numerals designate sequence clades. Support values are indicatedat nodes when found in at least 50% of 100 bootstrap trees. To avoid col-lapse of previously identified clades, sequences were evaluated along theirfull lengths, filling gaps with unknown nucleotides for shorter sequences.Positions occupied by PICs are highlighted by color codes: PIC-1 se-quence, orange; PIC-2 sequence, brown; sequences sharing PIC-3, red; se-quences sharing PIC-4, green; sequences sharing PIC-5, blue. Extant wolf(gray) and dog (black) sequences are identified by the prefixes W (forwolves) and A–F (for dogs), followed by numbers (see online Supplemen-tary Material for GenBank accession numbers and references). Continentaldistribution of dog sequences is given after sequence designation in paren-theses as follows: A, Asia; EA, East Asia including Oceania; WA, WestAsia; EU, Europe; AM, Americas; AF, Africa. Wolf countries are also in-dicated after sequence designation.
2544 Verginelli et al.
at UIC
Library, C
ollections Developm
ent on Novem
ber 15, 2014http://m
be.oxfordjournals.org/D
ownloaded from
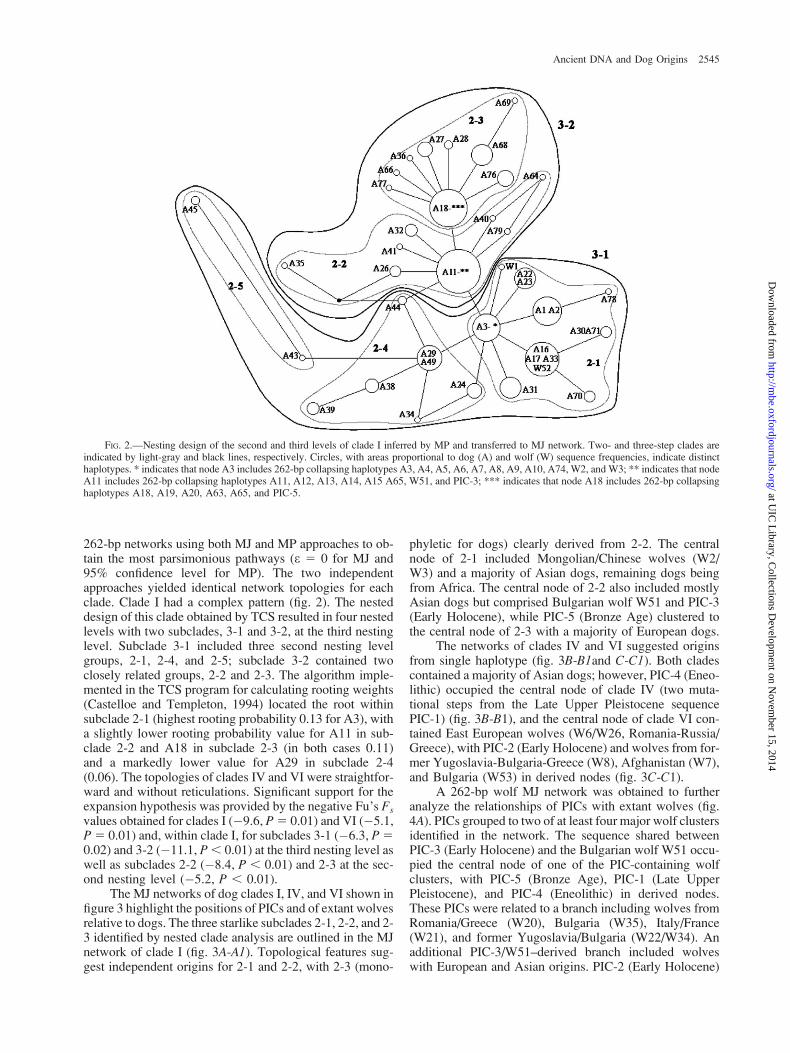
262-bp networks using both MJ and MP approaches to ob-tain the most parsimonious pathways (e 5 0 for MJ and95% confidence level for MP). The two independentapproaches yielded identical network topologies for eachclade. Clade I had a complex pattern (fig. 2). The nesteddesign of this clade obtained by TCS resulted in four nestedlevels with two subclades, 3-1 and 3-2, at the third nestinglevel. Subclade 3-1 included three second nesting levelgroups, 2-1, 2-4, and 2-5; subclade 3-2 contained twoclosely related groups, 2-2 and 2-3. The algorithm imple-mented in the TCS program for calculating rooting weights(Castelloe and Templeton, 1994) located the root withinsubclade 2-1 (highest rooting probability 0.13 for A3), witha slightly lower rooting probability value for A11 in sub-clade 2-2 and A18 in subclade 2-3 (in both cases 0.11)and a markedly lower value for A29 in subclade 2-4(0.06). The topologies of clades IV and VI were straightfor-ward and without reticulations. Significant support for theexpansion hypothesis was provided by the negative Fu’s Fs
values obtained for clades I (�9.6, P5 0.01) and VI (�5.1,P5 0.01) and, within clade I, for subclades 3-1 (�6.3, P50.02) and 3-2 (�11.1, P, 0.01) at the third nesting level aswell as subclades 2-2 (�8.4, P , 0.01) and 2-3 at the sec-ond nesting level (�5.2, P , 0.01).
The MJ networks of dog clades I, IV, and VI shown infigure 3 highlight the positions of PICs and of extant wolvesrelative to dogs. The three starlike subclades 2-1, 2-2, and 2-3 identified by nested clade analysis are outlined in the MJnetwork of clade I (fig. 3A-A1). Topological features sug-gest independent origins for 2-1 and 2-2, with 2-3 (mono-
phyletic for dogs) clearly derived from 2-2. The centralnode of 2-1 included Mongolian/Chinese wolves (W2/W3) and a majority of Asian dogs, remaining dogs beingfrom Africa. The central node of 2-2 also included mostlyAsian dogs but comprised Bulgarian wolf W51 and PIC-3(Early Holocene), while PIC-5 (Bronze Age) clustered tothe central node of 2-3 with a majority of European dogs.
The networks of clades IV and VI suggested originsfrom single haplotype (fig. 3B-B1and C-C1). Both cladescontained a majority of Asian dogs; however, PIC-4 (Eneo-lithic) occupied the central node of clade IV (two muta-tional steps from the Late Upper Pleistocene sequencePIC-1) (fig. 3B-B1), and the central node of clade VI con-tained East European wolves (W6/W26, Romania-Russia/Greece), with PIC-2 (Early Holocene) and wolves from for-mer Yugoslavia-Bulgaria-Greece (W8), Afghanistan (W7),and Bulgaria (W53) in derived nodes (fig. 3C-C1).
A 262-bp wolf MJ network was obtained to furtheranalyze the relationships of PICs with extant wolves (fig.4A). PICs grouped to two of at least four major wolf clustersidentified in the network. The sequence shared betweenPIC-3 (Early Holocene) and the Bulgarian wolf W51 occu-pied the central node of one of the PIC-containing wolfclusters, with PIC-5 (Bronze Age), PIC-1 (Late UpperPleistocene), and PIC-4 (Eneolithic) in derived nodes.These PICs were related to a branch including wolves fromRomania/Greece (W20), Bulgaria (W35), Italy/France(W21), and former Yugoslavia/Bulgaria (W22/W34). Anadditional PIC-3/W51–derived branch included wolveswith European and Asian origins. PIC-2 (Early Holocene)
FIG. 2.—Nesting design of the second and third levels of clade I inferred by MP and transferred to MJ network. Two- and three-step clades areindicated by light-gray and black lines, respectively. Circles, with areas proportional to dog (A) and wolf (W) sequence frequencies, indicate distincthaplotypes. * indicates that node A3 includes 262-bp collapsing haplotypes A3, A4, A5, A6, A7, A8, A9, A10, A74, W2, and W3; ** indicates that nodeA11 includes 262-bp collapsing haplotypes A11, A12, A13, A14, A15 A65, W51, and PIC-3; *** indicates that node A18 includes 262-bp collapsinghaplotypes A18, A19, A20, A63, A65, and PIC-5.
Ancient DNA and Dog Origins 2545
at UIC
Library, C
ollections Developm
ent on Novem
ber 15, 2014http://m
be.oxfordjournals.org/D
ownloaded from

was directly linked to the central node of the other PIC-containing wolf cluster, occupied by Romanian-Russian/Greek wolves (W6/W26). This cluster comprised wolvesfrom Afghanistan (W7), former Yugoslavia-Bulgaria-Greece (W8), China (W10), and Bulgaria (W53). Notably,the wolf clusters that contained PICs included all the 262-bp sequences shared between wolf and dog, five of which(i.e., W6/W26, W8, W51, W52, W53) are reported in EastEuropean wolves and one (i.e., W2/W3 [Mongolia/China])in Asian wolves.
Phylogeographic analyses were performed to testcorrelations between geographical locations and geneticrelatedness of the wolf and/or dog HVR1 sequences.Straight-line distances ranging from 3,666 to 13,697 kmseparated central locations for the dog and wolf sequencesfrom Europe, West Asia, East Asia, Africa, North America,and Central America that were analyzed in this study(Fig. S1A in the Supplementary Material online). Geneticrelatedness was mediated by distance. The matrix of pair-wise geographic distances calculated imposing a theoreticalpassage through the Bering Strait was correlated withthe matrix of pairwise Fst values between these groups(n5 6, r5 0.58, P, 0.05) (Fig. S1B in the SupplementaryMaterial online). Correlations were not significant whenplain Euclidean distances between locations were used.Our results did not support phylogeographic patterns withindog or wolf sequences analyzed independently, as wellas within clades or subclades, because no significant rela-tionships between genetic and geographic distances werefound.
Discussion
At the end of the last glaciation, that coincides with thearchaeologically hypothesized period of the origin of thedog, the wolf was one of the commonest among Holarcticmammals and could widely range across the NorthernHemisphere because low sea levels and prevalence ofsteppes/steppe-tundras facilitated its dispersal (Kurten1968; Kurten and Anderson 1980; Williams 1998). Givena probable late date for the diffusion of man in North Amer-ica (Dixon 2001), it is expected that wolves were initiallydomesticated in Eurasia, a view that agrees with geneticevidences (Leonard et al. 2002). Archaeozoological find-ings show that by 17–12,000 14C YA proto-dogs, probablyoriginating from different wolf populations, were alreadypresent in West Eurasia (Davis and Valla 1978; Nobis1979; Dayan 1994; Clutton-Brock 1995; Tcherncov andValla 1997; Sablin and Khlopachev 2002). Moleculartime estimates suggesting dog origins 76–135,000 YA or40–15,000 YA (Vila et al. 1997; Savolainen et al. 2002)need to be reevaluated based on recent findings showingthat molecular clocks cannot be used to date geologicallyrecent events, such as wolf-dog divergence (Ho et al. 2005).
To shed light on dog-founder wolf populations, wetook advantage of the unique insight provided by PICsequences. PIC-1 (14,670 6 130 14C YA) lived duringthe last Late Glacial Maximum (’22–14,000 14C YA),PIC-2 (9,860 6 50 14C YA) and PIC-3 (9,670 6 40 14CYA), almost synchronous and sympatric, lived shortly afterthe Younger Dryas, a cold interval following the initial
FIG. 3.—Phylogenetic 262-bp HVR1 networks of dog clades I, IV, and VI, containing PICs. Networks highlight the positions of PICs (red squareswith PIC designation) and extant wolves (triangles outlined in white, with sequence designation followed by country of origin). Circle areas are pro-portional to dog sequence frequencies (A–C) and to the number of unique haplotypes (262 bp or collapsed) they represent (A1–C1). Color codes indicatecontinental dog origins: yellow, Asia; blue, Europe; green, Africa; orange, Americas. Hypothetical haplotypes are represented by small white squares. Thestarlike subclades 2-1, 2-2, and 2-3, identified by nested clade analysis at the second nesting level are outlined.
2546 Verginelli et al.
at UIC
Library, C
ollections Developm
ent on Novem
ber 15, 2014http://m
be.oxfordjournals.org/D
ownloaded from
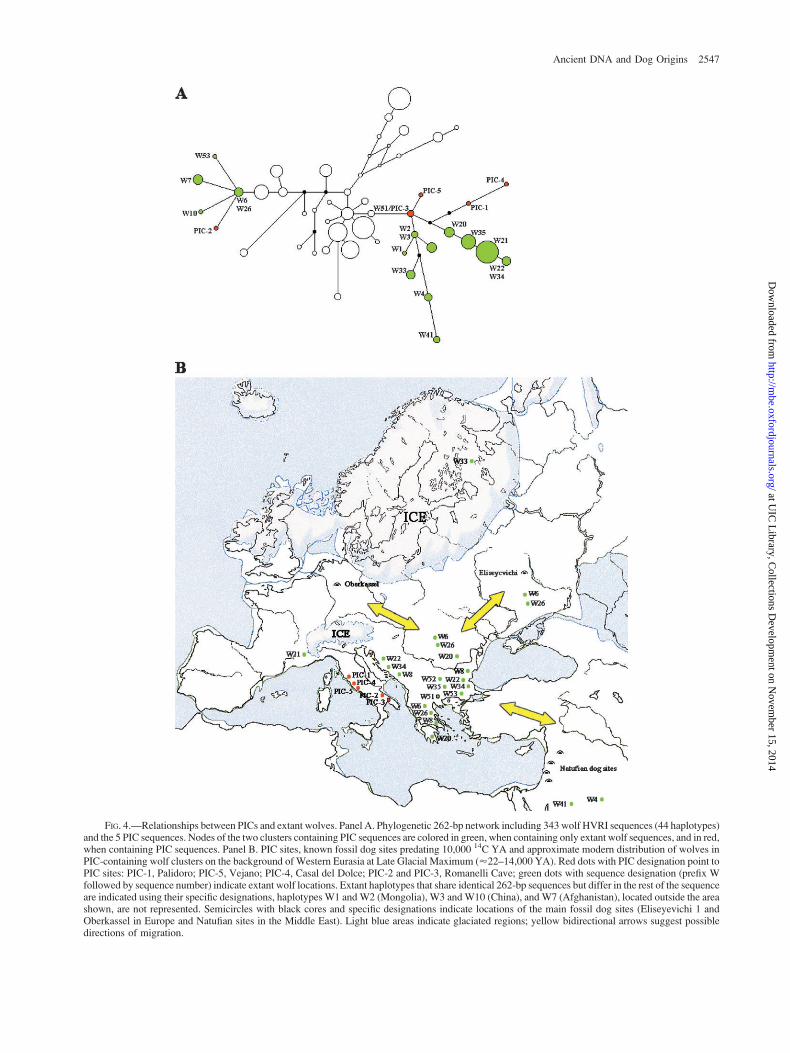
FIG. 4.—Relationships between PICs and extant wolves. Panel A. Phylogenetic 262-bp network including 343 wolf HVRI sequences (44 haplotypes)and the 5 PIC sequences. Nodes of the two clusters containing PIC sequences are colored in green, when containing only extant wolf sequences, and in red,when containing PIC sequences. Panel B. PIC sites, known fossil dog sites predating 10,000 14C YA and approximate modern distribution of wolves inPIC-containing wolf clusters on the background of Western Eurasia at Late Glacial Maximum (’22–14,000 YA). Red dots with PIC designation point toPIC sites: PIC-1, Palidoro; PIC-5, Vejano; PIC-4, Casal del Dolce; PIC-2 and PIC-3, Romanelli Cave; green dots with sequence designation (prefix Wfollowed by sequence number) indicate extant wolf locations. Extant haplotypes that share identical 262-bp sequences but differ in the rest of the sequenceare indicated using their specific designations, haplotypes W1 and W2 (Mongolia), W3 and W10 (China), and W7 (Afghanistan), located outside the areashown, are not represented. Semicircles with black cores and specific designations indicate locations of the main fossil dog sites (Eliseyevichi 1 andOberkassel in Europe and Natufian sites in the Middle East). Light blue areas indicate glaciated regions; yellow bidirectional arrows suggest possibledirections of migration.
Ancient DNA and Dog Origins 2547
at UIC
Library, C
ollections Developm
ent on Novem
ber 15, 2014http://m
be.oxfordjournals.org/D
ownloaded from

postglacial warming (Bjorck et al. 1996; Williams 1998).PIC-1 through -3 were within the range of morphometricvariability of the Late Upper Pleistocene Italian wolves(Cassoli, Fiore, and Tagliacozzo 1994) but could not be un-ambiguously assigned to wolves or wolf-sized proto-dogsbecause of the poor morphological resolving power of thefossil fragments (Sablin and Khlopachev 2002). However,despite intensive archaeological fieldwork, no proto-dogshave been identified in Italian Upper Paleolithic sites, whileat the end of the last glaciation, wolves were common inItaly and are archaeologically well documented (Kurten1968; Radmilli 1974; Cassoli, Fiore, and Tagliacozzo1994). PIC-4 (4,110 6 40 14C YA) and PIC-5 (3,040 640 14C YA), clearly identifiable as dogs, most likely reflectan indigenous Italian stock (De Grossi Mazzorin andTagliacozzo 1998).
The high genetic diversity of PIC-1, PIC-2, and PIC-3is clearly demonstrated by their dispersal across the threemajor dog clades and suggests that these early PICs wereconnected with a large and varied population. This earlydiversity fits with the variation of the later PIC dogs, butthe fact that two of the three Late Pleistocene/Early Holo-cene PIC sequences were novel, while both the later PICdog sequences were found in modern dogs, suggests thatmuch of the ancient variability has been lost (Vila et al.1999; Flagstad et al. 2003). Loss of genetic diversity isclearly evident when comparing the Late Pleistocene/EarlyHolocene PICs with extant Italian/French wolves, charac-terized by a single HVR1 haplotype (Vila et al. 1997; Randiet al. 2000; Lucchini et al. 2002). This could reflect severebottlenecking during the Holocene and suggests thatcaution is necessary in drawing historical inferences frommodern population data, as previously noted for otherUpper Pleistocene carnivores (Leonard, Wayne, andCooper 2000; Hofreiter et al. 2004).
PIC HVR1 sequences were compared to a large sam-ple of wolf and dog sequences worldwide. As observed bySavolainen et al. (2002), the interposition of wolf and dogclades, the presence of wolves within dog clades, and thelack of bootstrap support for clade I (the main dog clade)and for wolf clades indicated that wolf/dog taxonomic sta-tus was independent of HVR1 lineage. Significant bootstrapsupport was obtained for dog clades II, IV, and IX, as ob-served for corresponding clades by Savolainen et al. (2002),and, excluding wolves, for dog clade VI.
The complex pattern of the clade I network was com-patible with a minimum of two independent foundingevents, for subclades 3-1 and 3-2 identified at the third nest-ing level by nested clade analysis. At the second nestinglevel, 3-1 encompassed subgroups 2-1, 2-4, and 2-5, while3-2 included subgroup 2-2 and derived 2-3. The starlikeshape of 2-1, 2-2, and 2-3 and the significantly negativeFu’s Fs test values obtained for subclades 3-1 and 3-2 atthe third nesting level and subclades 2-2 and 2-3 at the sec-ond nesting level indicate sudden population expansionevents. The presence of Asian but not European wolvesand dogs in the starlike node of 2-1 suggests an Asian or-igin, as previously observed (Savolainen et al. 2002). How-ever, the position of PIC-3 and W51/A11 (the haplotypeidentical between a Bulgarian wolf and ancient breeds withOld and New World origins [Randi et al. 2000]) suggests
that 2-2 derived from European wolves. Furthermore, a Eu-ropean origin for dog-monophyletic subclade 2-3, derivedfrom 2-2, is strongly supported by the central position of theBronze Age dog PIC-5, together with a majority of Euro-pean dogs. The networks of the two other major dog cladesIV and VI are compatible with single origins that are linkedto European wolves by the positions of PICs and of EastEuropean wolves. In fact, the Eneolithic dog PIC-4 joinedthe central node of clade IV, distantly related to a node oc-cupied by the Late Upper Pleistocene PIC-1 sequence,while Romanian-Russian/Greek wolves clustered to thecentral node of clade VI, with the other Early Holocene se-quence, PIC-2, in derived position. The apparent contrastbetween the central positions occupied by PICs and EastEuropean wolves and the Asian origins of the majorityof dogs could reflect confounding effects of artificial breed-ing practices (there is evidence that the European dogbreeds, stringently selected after the formalization of thebreed concept in the 19th century, were derived from sharedstock [Parker et al. 2004]). Although not containing PICs, itis worth mentioning that the composition of the remainingminor dog clades II and IX suggests recent geographicallydefined origins in Europe and Asia, respectively.
The above-discussed links between PICs and East Eu-ropean wolves are supported by the wolf phylogenetic net-work. In this respect, it is intriguing that the two wolfclusters containing PICs hold all the 262-bp HVR1 se-quences shared between dog and wolf (five reported in EastEuropean wolves, one in wolves from Mongolia/China).Some of these shared sequences were previously attributedto dog gene flow into wolves (Randi et al. 2000). However,maternal introgressions of extant dogs into wolves follow-ing natural hybridization appear to be rare because of phy-siological, behavioral, and ecological barriers (Vila andWayne 1999). The interpretation of the dog-shared EastEuropean wolf sequences as representative of true ancientEuropean wolf genetic variation is supported by the closerelationship between PIC-2 and the Romanian-Russian/Greek wolf sequence W6/W26 and by the fact that theBulgarian wolf sequence W51 contains PIC-3.
We further investigated phylogeographic patternsamong global wolf and/or dog HVR1 sequences. Geo-graphic distances were not correlated with genetic distancevalues for dog or wolf sequences analyzed independently,even when analyses were restricted to clades and subclades.Lack of phylogeographic patterns in wolves, probablyreflecting their high mobility, was also found by Vilaet al. (1999). On the other hand, we found that geographicdistances calculated imposing a theoretical passage throughthe Bering Strait were correlated with genetic distancesamong European, West Asian, East Asian, African, NorthAmerican, and Central American dogs and wolves analyzedtogether, suggesting genetic differentiation related toisolation by geographic distance (Mayr 1942, 1963;Wright 1943).
Our data are in agreement with the notion that wolf anddog populations have been connected through Bering in thepast (Clutton-Brock 1995). The nonrandom geographic dis-tribution of wolf and dog HVR1 sequences could reflectphylogeographic patterns preceding the reproductive isola-tion of dogs from wolves. Our findings would also support
2548 Verginelli et al.
at UIC
Library, C
ollections Developm
ent on Novem
ber 15, 2014http://m
be.oxfordjournals.org/D
ownloaded from

the hypothesis of multiple independent origins of dogsand/or of frequent interbreeding between early proto-dogsand wolves throughout a vast geographic range (Sablin andKhlopachev 2002). In fact, genetic separation between dogsand wolves is likely to have occurred only after the Neo-lithic agropastoral revolution (’8,000 YA) that resultedin incompatibility between wolves and humans becauseof the presence of livestock (Lindner 1950; Clutton-Brock1995; Sablin and Khlopachev 2002).
The last glaciation could have restricted Upper Pleis-tocene carnivores such as bears and hyenas to refugia(Hofreiter et al. 2004) but could have facilitated the dis-persal of a highly mobile and adaptable predator such asthe wolf, which can thrive in open steppe environments(Kurten 1968). The mtDNA haplotypes of the extant EastEuropean wolves may represent a relic of the last glaciation.In fact, the relationships between PICs and East Europeanwolves are consistent with the paleogeographic background(Fig. 4B). At Last Glacial Maximum (’22–14,000 14CYA), because of the extensive regression of the EasternMediterranean Sea, Italy participated in a vast temperatesteppe province extending eastward into the Danubianbasin and southeastward, through a land bridge across theBosphorous, into West Asia (Williams 1998). Due to thepresence of ice sheets to the north, this southern steppe, thatlinked the distant locations of the earliest known dog sitesin Europe and in the Middle East (Davis and Valla 1978;Nobis 1979; Benecke 1994; Dayan 1994; Tcherncov andValla 1997; Sablin and Khlopachev 2002), was the mainroute of faunal and human migration into Europe and WestAsia (Sauter 1948; Kurten 1968; Gamble 1986). As shownin figure 4B, the distribution of the PIC-related wolfhaplotypes bridges the geographic gap between the earliestfossil dog sites.
In conclusion, the data show that the prehistoric canidsof the Italian peninsula were genetically diverse and notclosely related to the extant Italian wolves. Genetic dataobtained comparing the ancient sequences with extantdog and wolf sequences and early archaeozoological evi-dences concur in suggesting that Late Glacial/Early Holo-cene wolf populations of the West Eurasian steppes (thatstretched over South-Eastern Europe and West Asia) con-tributed to the origins of the dog (Davis and Valla 1978;Nobis 1979; Benecke 1994; Dayan 1994; Tcherncov andValla 1997; Sablin and Khlopachev 2002). Genetic dataalso suggest multiple independent Asian and European do-mestication events. In spite of the severe global decline ofwolves, extant East European wolf populations may stillcarry genetic signatures of dog-founder populations anddeserve dedicated study and conservation efforts.
The question of dog origins is geographically, genet-ically, and archaeologically complex and clearly requiresadditional multidisciplinary studies. In this respect, furthera-DNA evidence from relevant areas of the world shouldallow to better understand the evolution of wolves anddogs.
Supplementary Material
The following tables are available online: Table S1(frequencies, geographic origins, GenBank accession
numbers, and references of the extant wolf and coyoteHVR1 sequences [262–664 bp, nps 15431–15687/16039]);Table S2 (frequencies, geographic origins, GenBank acces-sion numbers, and references of the purebred dog HVR1 se-quences [262–664 bp, nps 15431–15687/16039]); Table S3(frequencies and geographic origins of unique 262-bp HVR1sequences [nps 15431–15687] of extant wolves [excludingsequences shared with dogs]); and Table S4 (frequenciesand geographic origins of unique 262-bp HVR1 sequences[nps 15431–15687] of purebred dogs [including sequencesshared with wolves]).
The following figures are available online: Fig. S1 (A,global map with sampling locations for the dog and wolfHVR1 sequences studied; B, plot of the genetic distancevs. the geographic distance between dog and wolf popula-tions sampled in Europe [EU], West Asia [WA], East Asia[EA], Africa [AF], North America [NA], and CentralAmerica [CA]).
The sequences reported in this paper have been de-posited in the GenBank database (accession numbersAY741666–AY741670).
Acknowledgments
This work was supported by the Italian National Re-search Council (CNR, Project Beni Culturali-Archivio Bio-logico coordinated by A. Guarino) and by the Ministerodell’Istruzione, dell’Universita e della Ricerca (MIUR),Progetti di Ricerca di Interesse Nazionale (PRIN) grants2000 and 2003, coordinated by L. R. Angeletti. We thankCarles Vila, University of Uppsala, Sweden; GiovanniDestro-Bisol, Department of Animal and Human Biology,University La Sapienza, Rome; and two anonymous re-viewers for critical suggestions. We acknowledge the helpof Associazione per lo Studio degli Animali Estinti(ASAE) (www.asae-onlus.org).
Literature Cited
Bandelt, H. J., P. Forster, and A. Rohl. 1999. Median-joining net-works for inferring intraspecific phylogenies. Mol. Biol. Evol.16:37–38.
Belkhir, K., P. Borsa, L. Chikhi, N. Raufaste, and F. Bonhomme.2004. GENETIX 4.05, logiciel sous Windows TM pour lagenetique des populations. Laboratoire Genome, Populations,Interactions, CNRS UMR 5171, Universite de Montpellier II,Montpellier, France.
Benecke, N. 1994. Der Mensch und seine Haustiere. Theiss,Stuttgart.
Bietti, A. 1976–1977. The excavations 1955–59 in the UpperPaleolithic deposit of Palidoro (Rome, Italy): a brief generalintroduction. Quaternaria 19:149–155.
Bjorck, S., B. Kromer, S. Johnsen et al. (11 co-authors). 1996.Synchronized terrestrial-atmospheric deglacial records aroundthe North Atlantic. Science 274:1155–1160.
Blanc, G. A. 1920. Grotta Romanelli. I. Stratigrafia. Arch. Antro-pol. Etnol. (Firenze) 50:65–103.
———. 1928. Grotta Romanelli. II. Dati ecologici e paletnologici.Arch. Antropol. Etnol. (Firenze) 58:365–431.
Cassoli, P. F. 1976–1977. Upper Paleolithic fauna at Palidoro(Rome): 1955 excavations. Quaternaria (Rome) 19:187–196.
Cassoli, P. F., I. Fiore, and A. Tagliacozzo. 1994. Butcheringand differentiated introduction of main mammals in the
Ancient DNA and Dog Origins 2549
at UIC
Library, C
ollections Developm
ent on Novem
ber 15, 2014http://m
be.oxfordjournals.org/D
ownloaded from

Epigravettian levels of the Romanelli Cave. Anthropozoologica25–26:309–318.
Castelloe, J., and A. R. Templeton. 1994. Root probabilities forintraspecific gene trees under neutral coalescent theory.Mol. Phylogenet. Evol. 3:102–113.
Clement, M., D. Posada, and K. A. Crandall. 2000. TCS: a computerprogram to estimate gene genealogies. Mol. Ecol. 9:1657–1659.
Clutton-Brock, J. 1995. Origins of the dog: domestication andearly history. Pp. 8–20 in J. Serpell, ed. The domestic dog:its evolution, behaviour and interactions with people. Cam-bridge University Press, Cambridge.
Davis, S. J. M., and F. R. Valla. 1978. Evidence for the domes-tication of the dog 12.000 years ago in the Natufian of Israel.Nature 276:608–610.
Dayan, T. 1994. Early domesticated dogs of the Near East.J. Archaeol. Sci. 21:633–640.
De Grossi Mazzorin, J., and A. Tagliacozzo. 2000. Morphologicaland osteological changes in the dog from the Neolithic to theRoman period in Italy. Pp. 141–161 in S. J. Crockford, ed.Dogs through time: an archaeological perspective. IntenationalCongress of the International Council for Archaeozoology,BAR International Series 889, Victoria, BC.
Delporte, H. 1990. L’image des animaux dans l’art prehistorique.Picard, Paris.
Dixon, E. J. 2001. Human colonization of the Americas: timing,technology and process. Quat. Sci. Rev. 20:277–299.
Excoffier, L., P. E. Smouse, and J. M. Quattro. 1992. Analysis ofmolecular variance inferred from metric distances among DNAhaplogroups: applications to human mitochondrial DNArestriction data. Genetics 131:479–491.
Felsenstein, J. 1984. Distance methods for inferring phylogenies:a justification. Evolution 38:16–24.
———. 2002. PHYLIP (phylogeny inference package). Version3.6a3. University of Washington, Seattle, Wash.
Fiore, I., and A. Tagliacozzo. 1997. I reperti osteologici animalidelle tombe 1 e 8 (Area A) e 4 (Area C) della necropoli eneo-litica: il contributo della tafonomia. Pp. 249–272 in A. Zarattiniand L. Petrassi, eds. Casale del Dolce. Ambiente, economia ecultura di una comunita preistorica della Valle del Sacco.Ministero per i Beni Culturali e Ambientali, SoprintendenzaArcheologica per il Lazio, Roma.
Flagstad, Ø., C. W. Walker, C. Vila, A.-K. Sundqvist, B.Fernholm, A. K. Hufthammer, Ø. Wiig, I. Koyola, and H.Ellegren. 2003. Two centuries of the Scandinavian wolf pop-ulation: patterns of genetic variability and migration during anera of dramatic decline. Mol. Ecol. 12:869–880.
Gamble, C. 1986. The Palaeolithic settlement of Europe. Cam-bridge University Press, Cambridge.
Greenwood, A. D., C. Capelli, G. Possnert, and S. Paabo. 1999.Nuclear DNA sequences from Late Pleistocene megafauna.Mol. Biol. Evol. 16:1466–1473.
Ho, S. Y. W., M. J. Phillips, A. Cooper, and A. J. Drummond.2005. Time dependency of molecular rate estimates and sys-tematic overestimation of recent divergence times. Mol. Biol.Evol. 22:1561–1568.
Hofreiter, M., V. Jaenicke, D. Serre, A. von Haeseler, andS. Paabo. 2001a. DNA sequences from multiple amplificationsreveal artifacts induced by cytosine deamination in ancientDNA. Nucleic Acids Res. 29:4793–4799.
Hofreiter, M., D. Serre, H. N. Poinar, M. Kuch, and S. Paabo.2001b. Ancient DNA. Nat. Rev. Genet. 2:353–359.
Hofreiter, M., D. Serre, N. Rohland, G. Rabeder, D. Nagel, N.Conard, S. Munzel, and S. Paabo. 2004. Lack of phylogeog-raphy in European mammals before the last glaciation. Proc.Natl. Acad. Sci. USA 101:12963–12968.
Jansen, T., P. Forster, M. A. Levine, H. Oelke, M. Hurles, C.Renfrew, J. Weber, and K. Olek. 2002. Mitochondrial DNA
and the origins of the domestic horse. Proc. Natl. Acad.Sci. USA 99:10905–10910.
Krings, M., A. Stone, R. W. Schmitz, H. Krainitzki, M. Stoneking,and S. Paabo. 1997. Neanderthal DNA sequences and theorigin of modern humans. Cell 90:19–30.
Kurten, B. 1968. Pleistocene mammals of Europe. Weidenfeld &Nicholson, London.
Kurten, B., and E. Anderson. 1980. Pleistocene mammals of NorthAmerica. Columbia University Press, New York.
Leonard, J. A., R. K. Wayne, and A. Cooper. 2000. Populationgenetics of Ice Age brown bears. Proc. Natl. Acad. Sci.USA 97:1651–1654.
Leonard, J. A., R. K. Wayne, J. Wheeler, R. Valadez, S. Guillen,and C. Vila. 2002. Ancient DNA evidence for Old Worldorigin of New World dogs. Science 298:1613–1616.
Liedloff, A. 1999. Mantel version 2.0 Mantel non-parametric testcalculator. School of Natural Resource Sciences, QueenslandUniversity of Technology, Brisbane, Australia.
Lindner, K. 1950. La succession des civilisations et le developpe-ment de la technique des armes. Pp. 22–116 in K. Lindner, ed.La chasse prehistorique. Payot, Paris.
Lucchini, V., E. Fabbri, F. Marucco, L. Ricci, L. Boitani, and E.Randi. 2002. Noninvasive molecular tracking of colonizingwolf (Canis lupus) packs in the western Italian Alps. Mol. Ecol.11:857–868.
Mayr, E. 1942. Systematics and the origin of species. ColumbiaUniversity Press, New York.
———. 1963. Animal species and evolution. Harvard UniversityPress, Cambridge.
Nobis, G. 1979. Der alteste Haushund lebte vor 14,000 Jahren.Umschau 19:610.
Nougier, L.-R. 1993. Art prehistorique mondial. Librairie GeneralFran1aise, Paris.
Olsen, S. J. 1985. Origins of the domestic dog. University ofArizona Press, Tucson.
Parker, H. G., L. V. Kim, N. B. Sutter, S. Carlson, T. D. Lorentzen,T. B. Malek, G. S. Johnson, H. B. DeFrance, E. A. Ostrander,and L. Kruglyak. 2004. Genetic structure of the purebreddomestic dog. Science 304:1160–1164.
Radmilli, A. M. 1974. Gli scavi nella Grotta Polesini a PonteLucano di Tivoli e la piu antica arte del Lazio. Origines—Studie materiali pubblicati a cura dell’Istituto Italiano di Preistoria eProtostoria, Sansoni, Firenze.
Randi, E., V. Lucchini, M. F. Christensen, N. Mucci, S. M. Funk,G. Dolf, and V. Loeschcke. 2000. Mitochondrial DNA analysisin Italian and East European wolves: detecting the conse-quences of small population size and hybridization. Conserv.Biol. 14:464–473.
Sablin, M. V., and G. A. Khlopachev. 2002. The earliest ice agedogs: evidence from Eliseevichi 1. Curr. Anthropol. 43:795–799.
Sauter, M. R. 1948. Prehistoire de la Mediterranee. Payot, Paris.Savolainen, P., T. Leitner, A. N. Wilton, E. Matisoo-Smith, and
J. Lundeberg. 2004. A detailed picture of the origin of the Aus-tralian dingo, obtained from the study of mitochondrial DNA.Proc. Natl. Acad. Sci. USA 101:12387–12390.
Savolainen, P., Y. P. Zhang, J. Luo, J. Lundeberg, and T. Leitner.2002. Genetic evidence for an East Asian origin of domesticdogs. Science 298:1610–1613.
Schmidt, H. A., K. Strimmer, M. Vingron, and A. von Haeseler.2002. TREE-PUZZLE: maximum likelihood phylogeneticanalysis using quartets and parallel computing. Bioinformatics18:502–504.
Schneider, S., D. Roessli, and L. Excoffier. 2000. Arlequin ver.2000: a software for population genetics data analysis.Genetics and Biometry Laboratory, University of Geneva,Switzerland.
2550 Verginelli et al.
at UIC
Library, C
ollections Developm
ent on Novem
ber 15, 2014http://m
be.oxfordjournals.org/D
ownloaded from

Schwartz, M. 1997. A history of dogs in the early Americas. YaleUniversity Press, New Haven, Conn.
Tamura, K., and M. Nei. 1993. Estimation of the number of nu-cleotide substitutions in the control region of mitochondrialDNA in humans and chimpanzees. Mol. Biol. Evol. 10:512–526.
Tcherncov, E., and F. R. Valla. 1997. Two new dogs, and otherNatufian dogs, from the southern Levant. J. Archaeol. Sci.24:65–95.
Troy, C. S., D. E. McHugh, J. F. Bailey, D. A. Magee, R. T.Loftus, P. Cunningham, A. T. Chamberlain, B. C. Sykes,and D. G. Bradley. 2001. Genetic evidence for Near-Easternorigins of European cattle. Nature 410:1088–1091.
Tsuda, K., Y. Kikkawa, H. Yonekawa, and Y. Tanabe. 1997. Ex-tensive interbreeding occurred among multiple matriarchalancestors during the domestication of dogs: evidence frominter- and intraspecies polymorphisms in the D-loop regionof mitochondrial DNA between dogs and wolves. GenesGenet. Syst. 72:229–238.
Vila, C., I. R. Amorim, J. A. Leonard, D. Posada, J. Castroviejo, F.Petrucci-Fonseca, K. A. Crandall, H. Ellegren, and R. K.Wayne. 1999. Mitochondrial DNA phylogeography and pop-ulation history of the grey wolf Canis lupus. Mol. Ecol.8:2089–2103.
Vila, C., J. A. Leonard, A. Gotherstrom, S. Marklund, K.Sandberg, K. Liden, R. K. Wayne, and H. Ellegren. 2001.
Widespread origins of domestic horse lineages. Science 291:474–477.
Vila, C., J. E. Maldonado, and R. K. Wayne. 1999. Phylogeneticrelationships, evolution, and genetic diversity of the domesticdog. J. Hered. 90:71–77.
Vila, C., P. Savolainen, J. E. Maldonado, I. R. Amorim, J. E. Rice,R. L Honeycutt, K. A. Crandall, J. Lundeberg, and R. K.Wayne. 1997. Multiple and ancient origins of the domesticdog. Science 276:1687–1689.
Vila, C., and R. K. Wayne. 1999. Hybridization between wolvesand dogs. Conserv. Biol. 13:195–198.
Wakeley, J. 1993. Substitution rate variation among sites in hyper-variable region 1 of human mitochondrial DNA. J. Mol. Evol.37:613–623.
Wayne, R. K., and E. A. Ostrander. 1999. Origin, genetic diversity,and genome structure of the domestic dog. Bioessays21:247–257.
Williams, M. A. J. 1998. Quaternary environments. EdwardArnold, London.
Wright, S. 1943. Isolation by distance. Genetics 28:114–138.———. 1951. The genetical structure of populations. Ann. Eugen.
15: 323–354.
William Martin, Associate Editor
Accepted August 16, 2005
Ancient DNA and Dog Origins 2551
at UIC
Library, C
ollections Developm
ent on Novem
ber 15, 2014http://m
be.oxfordjournals.org/D
ownloaded from





























