Embed Size (px)
Citation preview

Mitochondrial genomes: anything goesGertraud Burger1, Michael W. Gray2 and B. Franz Lang1
1Canadian Institute for Advanced Research, Programme in Evolutionary Biology, Department de Biochimie, Universite de Montreal,
2900 Boulevard Edouard-Montpetit, Montreal, Quebec, Canada H3T 1J42Canadian Institute for Advanced Research, Programme in Evolutionary Biology, Department of Biochemistry and Molecular
Biology, Dalhousie University, Halifax, Nova Scotia, Canada B3H 1X5
Mitochondria have their own genetic system – a vesti-
gial genome originating from an endosymbiotic
a-proteobacterial ancestor. The genetic function of
mitochondrial DNA (mtDNA) is well-conserved, being
involved in a maximum of five mitochondrial processes:
invariantly in respiration and/or oxidative phosphoryl-
ation and translation, and also in transcription, RNA
maturation and protein import. Recent data from mito-
chondrial genomics research demonstrate that this gen-
etic conservatism contrasts with an often-perplexing
diversity in structure and gene expression mechanisms.
In addition to outlining the extraordinary diversity of
mtDNA, this review highlights the divergent trends in
mitochondrial genome evolution in the various eukary-
otic lineages, and examines the relationship between
mitochondrial and nuclear genome evolution in a given
organism.
Mitochondria, the morphologically distinctive double-membrane organelles of most cells, use electron transportcoupled with oxidative phosphorylation to generate ATP[1]. Although mitochondria have a central role in energytransduction (the powerhouse of eukaryotic cells), theseorganelles participate in several other important func-tions, including ion homeostasis, intermediary metab-olism and apoptosis. However, in only a few instances dothese pathways have components encoded by mitochon-drial DNA (mtDNA) (Figure 1); rather, most mitochondrialproteins are specified by nuclear genes, synthesized in thecytosol and imported into the organelle [2].
Mitochondria are ubiquitous throughout the eukaryoticdomain: with very few exceptions, eukaryotes cannot existwithout them. Eukaryotic lineages that lack both func-tional mitochondria and a mitochondrial genome do exist,but several of these ‘amitochondriate’ eukaryotes containDERIVED (see Glossary) mitochondria (e.g. hydrogeno-somes) that generate ATP anaerobically [3]. Otheramitochondriate species contain what appear to beremnant mitochondrial structures (of currently undefinedfunction), and/or express nucleus-encoded proteins typi-cally targeted to, and functioning in, mitochondria [4].These observations argue that the last common ancestor ofextant eukaryotes was a mitochondria-containing organ-ism, implying that PRIMITIVE amitochondriate eukaryotes(eukaryotes whose ancestors never had mitochondria)might not exist.
The limited genetic function of mtDNA is well-con-served across eukaryotes: mtDNA encodes a small numberof proteins whose mRNAs are translated by a distinctivemitochondrial protein-synthesizing system, some of whosecomponents are always (rRNAs), usually (tRNAs) oroccasionally (ribosomal proteins) specified by the mito-chondrial genome. In marked contrast to this geneticconservatism (as this review will emphasize), the mito-chondrial genome is characterized by an extreme and oftenbewildering diversity in structure and gene expressionmechanisms [5,6]. This dichotomy – genetic conservatismversus structural flamboyance – has long obscured theorigin of mitochondria and raised the question of whetherthis organelle arose more than once during eukaryotic cellevolution. The weight of evidence now firmly argues thatthe mitochondrion had a single origin, arising from aeubacterial symbiont whose closest contemporary rela-tives are found within the a-proteobacteria.
This review summarizes recent data from mitochon-drial genomics research. It aims to highlight the extra-ordinary structural diversity shown by mtDNA across theeukaryotic domain (Figure 2), and the exceptionallydivergent trends in mitochondrial genome evolution invarious eukaryotic lineages. Chloroplasts, a second type ofDNA-containing organelle in algae and plants, will not bediscussed. It is worth noting that chloroplasts, which arethought to be the product of a more recent endosymbioticevent than mitochondria, have genomes that showsignificantly less structural diversity than mitochondrialgenomes.
The many faces of mitochondrial genome architecture
Given that the mitochondrial genome of several animalsexists in the form of super-coiled DNA [7], and that mtDNAin most eukaryotes maps as a circle in restriction analysisand sequence assembly, it was assumed, until recently,that mtDNAs are generally circular molecules. However,there is now strong evidence that many, if not most,
Glossary
Derived: taxa or characters that have evolved far away from the primitive state.
In molecular phylogeny, long tree-branches indicate highly derived taxa.
Homologous: similar in sequence owing to common evolutionary origin.
Primitive: original or ancient state.
Monophyly: unique origin in a single ancestral species (a monophyletic
grouping includes the ancestor and all of its descendants).
Basal (early diverging): emerging early in the evolution of a clade (i.e. a deep
branch in a phylogenetic tree).
Corresponding author: Gertraud Burger ([email protected]).
Review TRENDS in Genetics Vol.19 No.12 December 2003 709
http://tigs.trends.com 0168-9525/$ - see front matter q 2003 Elsevier Ltd. All rights reserved. doi:10.1016/j.tig.2003.10.012

Figure 1. Biological processes in mitochondria. The majority of mitochondrial functions is shown; only the mitochondrial matrix and inner membrane are depicted.
(For more details on mitochondrial metabolism and transport see [79].) Most mitochondrial components are encoded by the nucleus (blue); those highlighted in pink are
encoded by mtDNA in some eukaryotes but by the nuclear genome in other eukaryotes, whereas a small portion is invariably specified by mtDNA (orange). Processes that
have exclusively nucleus-encoded constituents are listed. Complexes I–V are involved in electron-transport and oxidative phosphorylation. TIM translocases are involved
in protein import and insertion into the inner membrane. Tat, Sec and Oxa1 translocases are involved in protein export from matrix and insertion into the inner membrane
(for a review see [2]). RNase P is a ribozyme that processes the 50-end of tRNAs (for recent results on fungal RNase P RNAs see [42]). It should be noted that the majority of
eukaryotes has a multi-subunit, rotenone-sensitive NADH dehydrogenase as depicted, with nad1, nad4 and nad5 invariantly encoded by mtDNA. However, in a few organ-
isms (yeast, Schizosaccharomyces and Plasmodium), this complex is replaced by a nucleus-encoded, single-polypeptide enzyme [80]. Abbreviation: EF-Tu, elongation
factor Tu.
TRENDS in Genetics
EF-Tu
RNAse P
Ribosome
Amino acid metabolism
Nucleotide metabolism
Lipid metabolism
DNA repair, replication, etc.
Fe-S synthesis
Heme synthesis
Ubiquinone synthesis
Complex IComplex II
Complex IIIComplex IV
Complex V
Cytochrome c
Sectranslocase
TIMtranslocases
Tattranslocase
Oxa1translocase
RNApolymerase Heme lyase
Co-factor synthesis
Signalling pathways
Chaperones
Carbohydrate metabolism
Proteases
Figure 2. Eukaryotic phylogeny. Schematic phylogenetic tree depicting relationships among eukaryotes based on evidence from both statistically well-supported molecular
phylogenies with different datasets [81], and from conjecture. Only the better-known, major organismal groups are shown, with taxa belonging to the same clade sharing
the same color shade. Broken lines indicate uncertain branching orders and tree topologies. For example, the Monophyly of apicomplexa plus ciliates and stramenopiles is
not well supported. Euglenozoa and jakobids have been suggested to be the most Basal (Early Diverging) lineages; however, the relative branching order at the base of the
tree (in excess of 1 billion years) remains largely unresolved.
TRENDS in Genetics
OchromonasCafeteria
AllomycesRhizopus
Saccharomyces
Phytophthora
Fungi
Stramenopiles
Amoebozoa
Apicomplexa
HarpochytriumGreen algaeand plants
Red algae
Arabidopsis
HomoFugu
MetridiumMonosiga
Marchantia
Scenedesmus
Chlamydomonas
EuglenaTrypanosoma
Cryptomonas
Porphyra
Holozoa
Reclinomonas
Acanthamoeba
Plasmodium
DictyosteliumPedinomonas
Cyanidioschyzon
JakobaEuglenozoaJakobids
Cryptophytes
Ciliates
Rhodomonas
Amoebidium
TetrahymenaParamecium
Review TRENDS in Genetics Vol.19 No.12 December 2003710
http://tigs.trends.com

circular-mapping mtDNAs consist primarily of linear,multimeric head-to-tail concatamers [8,9]. Linear map-ping (i.e. linear monomeric) mtDNA molecules havealso been detected in a few unrelated organisms, suchas ciliates, apicomplexa (Plasmodium and relatives),fungi, chlorophycean green algae (Chlamydomonas andrelatives), and several cnidarian animals. Notably,these linear molecules contain various specialized end-structures, such as covalently closed single-strandedDNA termini and terminally attached proteins; inaddition, they tend to have telomere-like repeats ofvarying length [10].
Mitochondrial genomes typically consist of a single‘chromosome’. Cases of mitochondrial genomes comprisingmultiple, circular-mapping molecules have been reportedin Spizellomyecs punctatus [11], a chytridiomycete fungus,and in the animals Globodera [12] and Dicyema [13]. Amost unusual and, indeed, unprecedented situation hasbeen identified in a single-celled protistan relative ofanimals, Amoebidium parasiticum [14], whose mtDNAcomprises a complex set of several hundred distinct typesof linear molecules [15] (Figure 3). These molecules carryat their termini a common pattern of short terminalrepeats, consisting invariably of the same ,40-bp term-inal sequence motif in inverted orientation, followedinternally by an ,100-bp repeat at one end, and adifferent ,65-bp repeat at the other end. The largenumber of distinct ‘chromosomes’ and the apparentabsence of centromeric structures characteristic of nuclearchromosomes raise questions about the mechanism ofconcerted replication and equal segregation of mtDNAduring mitochondrial division and subsequent celldivision in A. parasiticum. A similar dilemma exists inkinetoplastid protists (e.g. Trypanosoma), whose mito-chondria contain up to a few dozen gene-coding maxicirclesand up to several thousand minicircles that specify theguide RNAs involved in editing of mitochondrialmRNAs [16].
Size versus content
The size of mtDNA in most eukaryotic phyla ranges from15–60 kbp; however, there are some notable exceptions.Among those organisms whose mtDNA has been comple-tely sequenced, the two extreme outliers are the apicom-plexan protists Plasmodium sp. (obligate parasitescausing malaria), which have a minuscule mitochondrialgenome of 6 kbp [17], and rice (Oryza sativa), whosemtDNA (at 490 kbp [18]) is ,80 times larger than that ofPlasmodium. The mapped, but as-yet unsequencedmtDNAs of cucurbit plants could be even larger than2000 kbp [19].
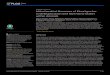
The coding capacity of mtDNA ranges from nearly100 genes in jakobid flagellates [20] to only five inPlasmodium, with the average across eukaryotes being40–50 genes. In contrast to their up-to-20-fold differ-ence in number, mitochondrial genes are involved, in atmost, five basic processes: invariantly in respirationand/or oxidative phosphorylation and translation, andoccasionally also in transcription, RNA maturation andprotein import (Figure 4).
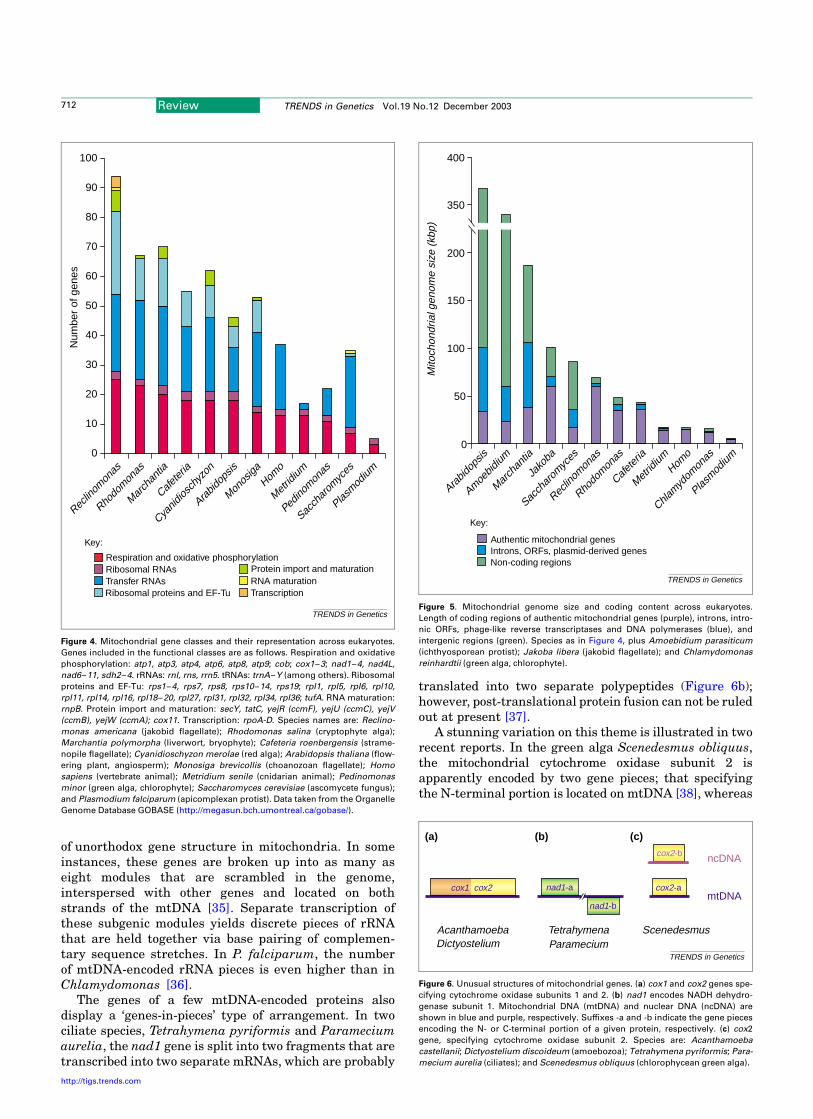
Somewhat counter-intuitively, there is no correlationbetween mtDNA size and gene content (Figure 5). In fact,the differences in size of mitochondrial genomes are mostlycaused by marked variations in the length and organiz-ation of intergenic regions, which, in some cases, consist ofextensive tandem-repeat arrays [21,22] or stem-loopmotifs [23]. Some of the repeat elements have beenproposed to be mobile and ‘selfish’ (e.g. [24]); others, toplay a role in recombination [25]. Another determinant ofmtDNA size is gene structure: a given gene product can beencoded by stretches of DNA of variable length owing tothe presence of introns (group I and II) of various size(0.15–4.00 kbp) and number (0 to.30). In the filamentousfungus Podospora anserina, introns account for up to 75%of the overall size of mtDNA [26].
Unusual mitochondrial gene structures
In addition to genes split by introns, mitochondria showother, more unusual gene organizations, one of which isgene fusion. In two amoebozoan protists, Acanthamoebacastellanii [27] and Dictyostelium discoideum [28], thegenes coding for subunits 1 and 2 of cytochrome oxidase(cox1 and cox2, respectively) are immediately adjacent toone another and in the same transcriptional orientation inthe mtDNA, forming a single contiguous ORF (Figure 6a).The existence of a bi-cistronic mRNA has been shown inAcanthamoeba, but it is not clear at this time whether afusion protein is initially synthesized as well [29].Chimeric genes, often containing recognizable pieces of aknown gene (or genes) within an otherwise unassignedORF, have been associated with cytoplasmic male sterilityin plants, a phenotype that is probably attributable toalterations in electron-transport-coupled oxidative phos-phorylation in pollen-producing cells [30]. In other cases(e.g. [31,32]), the functional consequences (if any) ofreported gene fusions are unknown.
Genes that exist in pieces, which were first described forthe mitochondrial rRNA genes in the ciliate protozoonTetrahymena pyriformis [33,34] and the green algaChlamydomonas reinhardtii [35], represent another type
Figure 3. Mitochondrial genome architectures. DNA molecules are mostly circular-
supercoiled in humans, and linear-monomeric in the ciliate Tetrahymena pyrifor-
mis (reviewed in [10]). In the fungus Spizellomyces punctatus, mtDNA consists of
three types of circular-mapping molecules [6], whereas several hundred types of
linear-monomeric molecules comprise the mitochondrial genome in the ichthyos-
porean protist Amoebidium parasiticum [15]. Triangles represent terminal repeat
motifs of 32 bp in T. pyriformis mtDNA and 40 bp in A. parasiticum mtDNA. Filled
circles and diamonds represent sub-terminal repeats, which are ,100-bp and
,65-bp long, respectively, in A. parasiticum mtDNA.
TRENDS in Genetics
Human mtDNA
Spizellomyces mtDNA
Tetrahymena mtDNA
Amoebidium mtDNA10 kbp
Review TRENDS in Genetics Vol.19 No.12 December 2003 711
http://tigs.trends.com
of unorthodox gene structure in mitochondria. In someinstances, these genes are broken up into as many aseight modules that are scrambled in the genome,interspersed with other genes and located on bothstrands of the mtDNA [35]. Separate transcription ofthese subgenic modules yields discrete pieces of rRNAthat are held together via base pairing of complemen-tary sequence stretches. In P. falciparum, the numberof mtDNA-encoded rRNA pieces is even higher than inChlamydomonas [36].
The genes of a few mtDNA-encoded proteins alsodisplay a ‘genes-in-pieces’ type of arrangement. In twociliate species, Tetrahymena pyriformis and Parameciumaurelia, the nad1 gene is split into two fragments that aretranscribed into two separate mRNAs, which are probably
translated into two separate polypeptides (Figure 6b);however, post-translational protein fusion can not be ruledout at present [37].
A stunning variation on this theme is illustrated in tworecent reports. In the green alga Scenedesmus obliquus,the mitochondrial cytochrome oxidase subunit 2 isapparently encoded by two gene pieces; that specifyingthe N-terminal portion is located on mtDNA [38], whereas
Figure 4. Mitochondrial gene classes and their representation across eukaryotes.
Genes included in the functional classes are as follows. Respiration and oxidative
phosphorylation: atp1, atp3, atp4, atp6, atp8, atp9; cob; cox1–3; nad1–4, nad4L,
nad6–11, sdh2–4. rRNAs: rnl, rns, rrn5. tRNAs: trnA–Y (among others). Ribosomal
proteins and EF-Tu: rps1–4, rps7, rps8, rps10–14, rps19; rpl1, rpl5, rpl6, rpl10,
rpl11, rpl14, rpl16, rpl18–20, rpl27, rpl31, rpl32, rpl34, rpl36; tufA. RNA maturation:
rnpB. Protein import and maturation: secY, tatC, yejR (ccmF), yejU (ccmC), yejV
(ccmB), yejW (ccmA); cox11. Transcription: rpoA-D. Species names are: Reclino-
monas americana (jakobid flagellate); Rhodomonas salina (cryptophyte alga);
Marchantia polymorpha (liverwort, bryophyte); Cafeteria roenbergensis (strame-
nopile flagellate); Cyanidioschyzon merolae (red alga); Arabidopsis thaliana (flow-
ering plant, angiosperm); Monosiga brevicollis (choanozoan flagellate); Homo
sapiens (vertebrate animal); Metridium senile (cnidarian animal); Pedinomonas
minor (green alga, chlorophyte); Saccharomyces cerevisiae (ascomycete fungus);
and Plasmodium falciparum (apicomplexan protist). Data taken from the Organelle
Genome Database GOBASE (http://megasun.bch.umontreal.ca/gobase/).
TRENDS in Genetics
Respiration and oxidative phosphorylation
Key:
Ribosomal RNAsTransfer RNAsRibosomal proteins and EF-Tu
RNA maturationProtein import and maturation
Transcription
Num
ber
of g
enes
100
90
80
70
60
50
40
30
20
10
0
Reclin
omon
as
Rhodo
monas
March
antia
Cafeter
ia
Cyanid
iosch
yzon
Arabid
opsis
Monos
iga
Homo
Metridi
um
Pedino
monas
Sacch
arom
yces
Plasmod
ium
Figure 5. Mitochondrial genome size and coding content across eukaryotes.
Length of coding regions of authentic mitochondrial genes (purple), introns, intro-
nic ORFs, phage-like reverse transcriptases and DNA polymerases (blue), and
intergenic regions (green). Species as in Figure 4, plus Amoebidium parasiticum
(ichthyosporean protist); Jakoba libera (jakobid flagellate); and Chlamydomonas
reinhardtii (green alga, chlorophyte).
TRENDS in Genetics
0
50
Non-coding regionsIntrons, ORFs, plasmid-derived genesAuthentic mitochondrial genes
Key:
100
150
200
350
400
Mito
chon
dria
l gen
ome
size
(kb
p)
Arabid
opsis
Amoebid
ium
March
antia
Jako
ba
Sacch
arom
yces
Reclin
omon
as
Rhodo
monas
Cafeter
ia
Metridi
umHom
o
Chlamyd
omon
as
Plasmod
ium
Figure 6. Unusual structures of mitochondrial genes. (a) cox1 and cox2 genes spe-
cifying cytochrome oxidase subunits 1 and 2. (b) nad1 encodes NADH dehydro-
genase subunit 1. Mitochondrial DNA (mtDNA) and nuclear DNA (ncDNA) are
shown in blue and purple, respectively. Suffixes -a and -b indicate the gene pieces
encoding the N- or C-terminal portion of a given protein, respectively. (c) cox2
gene, specifying cytochrome oxidase subunit 2. Species are: Acanthamoeba
castellanii; Dictyostelium discoideum (amoebozoa); Tetrahymena pyriformis; Para-
mecium aurelia (ciliates); and Scenedesmus obliquus (chlorophycean green alga).
TRENDS in Genetics
ncDNA
mtDNAnad1-b
cox1 cox2 cox2-a
TetrahymenaParamecium
AcanthamoebaDictyostelium
Scenedesmus
nad1-a
cox2-b
(a) (b) (c)
Review TRENDS in Genetics Vol.19 No.12 December 2003712
http://tigs.trends.com

the C-terminal portion is believed to be nucleus-encoded[39] (Figure 6c). In the green algae Chlamydomonas andPolytomella, the corresponding two protein portions areencoded by separate nuclear genes, which are synthesizedin the cytoplasm and imported into the mitochondrion [39].A second recent example of a split gene with one-half inmtDNA and the other half in nuclear DNA is rpl2 inflowering plants [40].
Finally, gene simplification is another eccentricphenomenon seen in mitochondrial genomes. Examplesinclude the truncated tRNAs (lacking one, or more, helicalarms) found in the mitochondria of several animal lineages[7], the structurally streamlined mitochondrial rRNAs inmost animals [7] and some protists [41], and mitochondrialRNase P RNAs in fungi [42]. In certain organisms(e.g. in the fungus Harpochytrium [43]), a similartrend is observed for mitochondrion-encoded proteingenes, whose coding regions are often shortened in theC-terminal portion compared with their more primitivecounterparts.
Events implicated in changes in the overall organization
of mitochondrial genomes
Changes in mtDNA architecture have been associatedwith autonomously replicating mitochondrial plasmids,which are common in fungi but which also occur in severalplant and protist taxa. The well-characterized mitochon-drial plasmids in fungi are primarily monomeric linearmolecules with inverted terminal repeats [44]. Insertion ofsuch linear plasmids into mtDNA probably triggersconversion of genome conformation from circular mappingto monomeric linear, as observed in maize cytoplasmicmale sterility [45]. Stable transitions of mtDNA topologyfrom circular to linear have occurred within relativelyshort phylogenetic distances, as seen in yeasts [10], goldenalgae [46] and chytridiomycete fungi [43,47].
Changes in mtDNA that lead to size expansion aremainly caused by an accumulation of repeat sequences andAT-rich intergenic spacers, mobile elements and introns.Even within a genus, the length of non-coding intergenicregions in mtDNA can vary considerably. For example,mtDNA size in fission yeast species varies between 17 and$80 kbp [48], with multiple arrays of intergenic repeatsaccounting for most of the variation. By contrast, rampantsize-expansion of plant mtDNAs seems to have arisenprincipally through acquisition of foreign DNA of chlor-oplast, nuclear, viral and as-yet-unknown origin [49],involving active DNA uptake [50] with master circles andsubcircles constantly recombining at a low level to createmixed populations of subgenomic DNA molecules (‘sub-limons’ [25]). In turn, size reduction of mtDNAs oftenoccurs by gene transfer to the nucleus (Figure 7); examplesof this phenomenon in flowering plants are especiallynotable in terms of both number and frequency [51–53].Other strategies of mtDNA size reduction include intronloss, and elimination of intergenic spacers, sometimesto such a degree that coding regions of adjacent genesoverlap (e.g. [7,27]).
Rearrangements through insertions, deletions orrecombination follow special rules owing to the multi-copy character of mitochondrial genomes. In several
eukaryotic groups, mitochondrial genomes are distributedover more than one mitochondrion per cell, and themtDNA copy number ranges from around ten to severalthousand per cell (often varying with growth conditions).The multiplicity of mtDNA molecules within the cellimplies that mitochondrial mutations might accumulategradually, without immediate deleterious impact. Thiscould partly explain the accelerated evolutionary rates ofmtDNA relative to nuclear DNA that are primarilyobserved in animals (e.g. vertebrates and insects),fungi (e.g. yeasts and fission yeasts) and certain protists(e.g. ciliates and slime moulds).
In human somatic cells, it has been well-documentedthat mtDNA accumulates point mutations and largedeletions, predominantly in brain and heart muscle ofolder individuals (.40 years) [54]. This high incidence ofmutations has been attributed to several factors, includingslipped mispairing at direct sequence repeats, topoisome-rase cleavage of mtDNA, the absence of excision repair,and incomplete repair of damage caused by free-radicalsproduced as a by-product of oxidative phosphorylation[55]. However, as there are few comparative biochemicalstudies in other eukaryotes, the specific mechanismsunderlying the particularly high occurrence of mitochon-drial mutations in human versus other eukaryotesremains largely unknown.
Exceptional horizontal transfer of mitochondrial genes
Although the evolutionary transfer of genes from organellegenomes is well-documented (Figure 7), there are fewreports to-date of the transfer of genetic material into themitochondrial genome and, especially, between mitochon-drial genomes. Only in flowering plants (angiosperms)does incorporation of nuclear and, particularly, chloroplastDNA sequences into mtDNA occur frequently; suchtransfer is not evident among non-angiosperm land plantsor in green algae [49]. Exchange of mobile elements (groupI and group II introns) between mtDNA and non-mtDNAsources has been inferred on the basis of sequencecomparisons [56–58], whereas mtDNA-to-mtDNA trans-fer is suggested by the presence of highly similar introns atidentical positions in Homologous mitochondrial genes ofdistantly related species [59–61]. A few cases have beendescribed that involve transfer of coding regions betweenmitochondrial genomes; one of these examples involvesfungi (probably from two different classes) and results in ahybrid atp6 gene [62].
Recently, widespread horizontal transfer of mitochon-drial genes (both respiratory chain and ribosomal protein)has been reported to occur between distantly relatedangiosperms, even between dicotyledons and monocotyle-dons [63]. The genomic consequences of these mtDNA-to-mtDNA transfers include gene duplication, recapture ofgenes previously lost through transfer to the nucleus and,in one particularly notable case, a chimeric, half-monocot–half-dicot rps11 gene. It is estimated that these transferevents occurred between 30 and 140 million years ago inindividual cases. It would seem that transfer of geneticmaterial into angiosperm mtDNA occurs relatively fre-quently on an evolutionary timescale, regardless ofwhether such transfer emanates from the chloroplast
Review TRENDS in Genetics Vol.19 No.12 December 2003 713
http://tigs.trends.com

genome of the same species or from the mtDNA of adifferent plant species. Successful functional transfer ofgenes into angiosperm mtDNA might be facilitated by theexpansive, largely non-coding nature of this genome [64],whose expression seems little constrained by gene locationand overall organization.
Are evolutionary trends similar in mitochondrial and
nuclear genomes?
With the recent availability of nuclear genome data fromseveral eukaryotes, we are now in a position to compareevolutionary trends for the mitochondrial and nucleargenomes of a given organism. One readily measurableparameter is the proportion of non-coding sequence in thetwo genomes. For example, human mtDNA is tightlypacked with intron-less genes [65], whereas humannuclear genes contain a large number of introns separatedby extensive intergenic regions [66,67]. By contrast, genesin yeast mtDNA are intron-rich and separated by long non-coding stretches [68], whereas yeast nuclear DNA har-bours closely spaced and mostly intron-less genes [69].Finally, in the pufferfish Fugu, gene density is high in bothgenomes [70,71]. Comparison of the relative gene numbersin the two genomes shows similarly unrelated variations,implying that the evolutionary trends shaping codingcapacity and density in the nuclear versus mitochondrialgenome proceed independently from one another in a givenorganism.
Another issue that can now be investigated is whetherthe propensity to insert foreign DNA is the same for themitochondrial genome and nuclear genome of a givenorganism. In flowering plants, acquisition of foreign DNAis rampant both in the nucleus and in mitochondria (butnot in chloroplasts), whereas in yeast, DNA escapes frommitochondria to the nucleus (but not in the opposite
direction) at a notably high frequency [72]. Moreover, inprimates, nuclear genomes contain substantial numbers ofmtDNA insertions [73], but the mitochondrial genome inthese cases is free of foreign DNA. These contrastingexamples illustrate the concept that evolution of thenuclear and organellar genomes of a given eukaryote canproceed in very different directions.
Does a parasitic lifestyle accelerate mtDNA evolution?
It is tempting to speculate that mtDNA might be undermore relaxed selection in parasitic lineages, leading to theemergence of exotic mitochondrial genome features, asdescribed earlier. Indeed, mtDNA with the smallestnumber of genes is found in Plasmodium, and the mostextreme case of mRNA editing occurs in trypanosomes,both of which are obligate parasites. Somewhat surpris-ingly, this correlation does not withstand careful scrutiny.For example, Prototheca wickerhamii, a non-photosyn-thetic chlorophycean alga and (mild) human pathogen, hasone of the most primitive mitochondrial genomeswithin this group of green algae [38,74,75]. Bycontrast, several free-living, photosynthetic relativeshave highly derived mtDNAs: Scenedesmus obliquushas an unusual translation code and substantiallyfewer mtDNA-encoded genes than P. wickerhamiimtDNA [38]. In addition, Pedinomonas minor [21]and Chlamydomonas reinhardtii (and its close rela-tives) have mtDNAs that are typical cases of reducedmitochondrial genomes [76]. Other examples of non-pathogens with highly derived mtDNAs are Euglenagracilis [77], baker’s and fission yeast (and its closerelatives), animals in general, and Amoebidium para-siticum [15], a saprotrophic ectosymbiont that isfrequently found on crustaceans and insects thatinhabit fresh water ponds.
Figure 7. Fate of proto-mitochondrial genes. Early gene migration has taken two main routes. (a) The a-proteobacterial symbiont underwent massive gene loss during tran-
sition to an organelle. Among the genes essential for mitochondrial functions, the majority was transferred to the nucleus, with a small portion remaining in the evolving
mitochondrial DNA (mtDNA). (b) The hypothetical primitive nucleus of the host [either an amitochondriate eukaryote (see text) [82] or an Archaea-related organism [83])
acquired several hundred symbiont genes, some of which were substituted for nuclear genes. In some instances, nucleus-acquired DNA from other sources replaced a
mitochondrial function initially encoded by the a-proteobacterial symbiont. For example, mtDNA in jakobid flagellates contains genes specifying the components of a typi-
cal eubacteria-like mitochondrial RNA polymerase, whereas in all other eukaryotes, this enzyme has a T3-phage-like structure and is encoded by the nuclear genome [84].
Genes are colour-coded as follows: red, a-proteobacterial genes; blue, nuclear genes of the original host; yellow, a-proteobacterial genes that replaced host genes; green,
foreign genes acquired from sources other than the genome of the mitochondrial ancestor.
TRENDS in Genetics
α-Proteobacterialendosymbiont
Acquisition ofsymbiont genes and foreign genes;
gene substitutions
Gene export and lossMitochondrion
Primitive nucleus Nucleus
Modern eukaryote
Primitive eukaryote
(b)
(a)
Review TRENDS in Genetics Vol.19 No.12 December 2003714
http://tigs.trends.com

Why do we see the highly derived mtDNAs in somespecies but not in others? One possible explanation is thatderived mitochondrial genomes are a consequence ofaccelerated growth rates (i.e. they tend to occur in‘weeds’, that is, organisms with short generation times).Some such species (e.g. yeasts) might be highly specializedto thrive in specific environments, whereas other organ-isms of this type (such as chlamydomonad algae) occurmore widely, dominating over other species through moreefficient propagation.
Mitochondrial DNA variability: implications for gene
expression
In our quest to understand the role of genome organizationin gene expression, a comparative mitochondrial genomicsapproach is only the first step. Complete sequencing ofmtDNA enables the identification of genes and permits oneto make inferences about unusual modes of geneexpression (e.g. split and divergently transcribed genes,or fused ORFs). However, only systematic and compre-hensive analysis of the transcriptome (the portion of thegenome that is transcribed) will reveal: (i) whether allidentified genes are active, (ii) whether co- or post-transcriptional processes, such as RNA editing, alterencoded genetic information on the way to its expression,and (iii) whether RNA species (perhaps having regulatoryfunctions) are synthesized in addition to the usualinformational and structural RNAs. Identification of agene, even one having all the hallmarks of a functionalunit, is no guarantee that it is active, as exemplified by thesilent cox2 gene in soybean mtDNA [78]. Even wheretranscription of a protein-coding gene is shown, definitiveevidence of function ultimately requires identification ofthe corresponding protein. This criterion has beensatisfied in very few cases, although direct analysis ofthe mitochondrial proteome is rapidly gaining favour.
Concluding remarks
Comparative mitochondrial genomics, especially in pro-tists, has revealed an often perplexing diversity in overallorganization and mode of expression of mtDNA. Thecomparative approach has also amply demonstrated thatthis extreme structural divergence co-exists with theconserved functional role of the mitochondrial genome.In this respect, mtDNA is an evolutionary playground parexcellence. No correlation is evident in the mode and tempoof evolutionary change in the mitochondrial and nucleargenomes across the broad spectrum of eukaryotic lineages.
Acknowledgements
This project was supported by grants from the Canadian Institutes forHealth Research (MSP14226, MOP42475 and MOP4124). Salary andinteraction support from the Canadian Institute for Advanced Research(G.B., M.W.G. and B.F.L.) and the Canada Research Chairs Programme(M.W.G. and B.F.L.) is gratefully acknowledged.
References
1 Saraste, M. (1999) Oxidative phosphorylation at the fin de siecle.Science 283, 1488–1493
2 Herrmann, J.M. (2003) Converting bacteria to organelles: evolution ofmitochondrial protein sorting. Trends Microbiol. 11, 74–79
3 Embley, T.M. et al. (2003) Mitochondria and hydrogenosomes are two
forms of the same fundamental organelle. Philos. Trans. R. Soc. Lond.B Biol. Sci. 358, 191–201
4 Roger, A.J. and Silberman, J.D. (2002) Cell evolution: mitochondria inhiding. Nature 418, 827–829
5 Gray, M.W. et al. (1999) Mitochondrial evolution. Science 283,1476–1481
6 Lang, B.F. et al. (1999) Mitochondrial genome evolution and the originof eukaryotes. Annu. Rev. Genet. 33, 351–397
7 Boore, J.L. (1999) Animal mitochondrial genomes. Nucleic Acids Res.27, 1767–1780
8 Bendich, A.J. (1993) Reaching for the ring: the study of mitochondrialgenome structure. Curr. Genet. 24, 279–290
9 Bendich, A.J. (1996) Structural analysis of mitochondrial DNAmolecules from fungi and plants using moving pictures and pulsed-field gel electrophoresis. J. Mol. Biol. 255, 564–588
10 Nosek, J. and Tomaska, L. Mitochondrial genome diversity: evolutionof the molecular architecture and replication strategy. Curr. Genet.(in press)
11 Burger, G. and Lang, B.F. (2003) Parallels in genome evolution inmitochondria and bacterial symbionts. IUBMB Life 55, 205–212
12 Armstrong, M.R. et al. (2000) A multipartite mitochondrial genome inthe potato cyst nematode Globodera pallida. Genetics 154, 181–192
13 Watanabe, K.I. et al. (1999) Mitochondrial genes are found onminicircle DNA molecules in the mesozoan animal Dicyema. J. Mol.Biol. 286, 645–650
14 Lang, B.F. et al. (2002) The closest unicellular relatives of animals.Curr. Biol. 12, 1773–1778
15 Burger, G. et al. (2003) Unique mitochondrial genome architecture inunicellular relatives of animals. Proc. Natl. Acad. Sci. U. S. A. 100,892–897
16 Morris, J.C. et al. (2001) Replication of kinetoplast DNA: an update forthe new millennium. Int. J. Parasitol. 31, 453–458
17 Feagin, J.E. (2000) Mitochondrial genome diversity in parasites. Int.J. Parasitol. 30, 371–390
18 Notsu, Y. et al. (2002) The complete sequence of the rice (Oryza sativaL.) mitochondrial genome: frequent DNA sequence acquisition andloss during the evolution of flowering plants. Mol. Genet. Genomics268, 434–445
19 Ward, B.L. et al. (1981) The mitochondrial genome is large andvariable in a family of plants (cucurbitaceae). Cell 25, 793–803
20 Lang, B.F. et al. (1997) An ancestral mitochondrial DNA resembling aeubacterial genome in miniature. Nature 387, 493–497
21 Turmel, M. et al. (1999) The complete mitochondrial DNA sequences ofNephroselmis olivacea and Pedinomonas minor. Two radicallydifferent evolutionary patterns within green algae. Plant Cell 11,1717–1730
22 Lunt, D.H. et al. (1998) Mitochondrial DNA variable number tandemrepeats (VNTRs): utility and problems in molecular ecology. Molec.Ecol. 7, 1441–1455
23 Paquin, B. and Lang, B.F. (1996) The mitochondrial DNA of Allomycesmacrogynus: the complete genomic sequence from an ancestral fungus.J. Mol. Biol. 255, 688–701
24 Paquin, B. et al. (2000) Double-hairpin elements in the mitochondrialDNA of Allomyces: evidence for mobility. Mol. Biol. Evol. 17,1760–1768
25 Andre, C. et al. (1992) Small repeated sequences and the structure ofplant mitochondrial genomes. Trends Genet. 8, 128–132
26 Cummings, D.J. et al. (1990) The complete DNA sequence of themitochondrial genome of Podospora anserina. Curr. Genet. 17,375–402
27 Burger, G. et al. (1995) The mitochondrial DNA of the amoeboidprotozoon, Acanthamoeba castellanii: complete sequence, gene contentand genome organization. J. Mol. Biol. 245, 522–537
28 Ogawa, S. et al. (2000) The mitochondrial DNA of Dictyosteliumdiscoideum: complete sequence, gene content and genome organiz-ation. Mol. Gen. Genet. 263, 514–519
29 Lonergan, K.M. and Gray, M.W. (1996) Expression of a continuousopen reading frame encoding subunits 1 and 2 of cytochrome c oxidasein the mitochondrial DNA of Acanthamoeba castellanii. J. Mol. Biol.257, 1019–1030
30 Hanson, M.R. (1991) Plant mitochondrial mutations and male sterility.Annu. Rev. Genet. 25, 461–486
31 Oudot-Le Secq, M.P. et al. (2001) The complete sequence of a brown
Review TRENDS in Genetics Vol.19 No.12 December 2003 715
http://tigs.trends.com

algal mitochondrial genome, the ectocarpale Pylaiella littoralis (L.)Kjellm. J. Mol. Evol. 53, 80–88
32 Bullerwell, C.E. et al. (2000) A novel motif for identifying rps3 homologsin fungal mitochondrial genomes. Trends Biochem. Sci. 25, 363–365
33 Heinonen, T.Y. et al. (1987) Rearranged coding segments, separated bya transfer RNA gene, specify the two parts of a discontinuous largesubunit ribosomal RNA in Tetrahymena pyriformis mitochondria.J. Biol. Chem. 262, 2879–2887
34 Schnare, M. et al. (1986) A discontinuous subunit ribosomal RNA inTetrahymena pyriformis mitochondria. J. Biol. Chem. 261, 5187–5193
35 Boer, P.H. and Gray, M.W. (1988) Scrambled ribosomal RNA genepieces in Chlamydomonas reinhardtii mitochondrial DNA. Cell 55,399–411
36 Gillespie, D.E. et al. (1999) The fragmented mitochondrial ribosomalRNAs of Plasmodium falciparum have short A tails. Nucleic Acids Res.27, 2416–2422
37 Edqvist, J. et al. (2000) Expression of mitochondrial protein-codinggenes in Tetrahymena pyriformis. J. Mol. Biol. 297, 381–393
38 Nedelcu, A.M. et al. (2000) The complete mitochondrial DNA sequence ofScenedesmus obliquus reflects an intermediate stage in the evolution ofthe green algal mitochondrial genome. Genome Res. 10, 819–831
39 Perez-Martinez, X. et al. (2001) Subunit II of cytochrome c oxidase inchlamydomonad algae is a heterodimer encoded by two independentnuclear genes. J. Biol. Chem. 276, 11302–11309
40 Adams, K.L. et al. (2001) Mitochondrial gene transfer in pieces: fissionof the ribosomal protein gene rpl2 and partial or complete genetransfer to the nucleus. Mol. Biol. Evol. 18, 2289–2297
41 Gray, M.W. et al. (1998) Genome structure and gene content in protistmitochondrial DNAs. Nucleic Acids Res. 26, 865–878
42 Seif, E.R. et al. (2003) Mitochondrial RNase P RNAs in ascomycetefungi: lineage-specific variations in RNA secondary structure. RNA 9,1073–1083
43 Bullerwell, C.E. et al. (2003) Evolution of monoblepharidalean fungibased on complete mitochondrial genome sequences. Nucleic AcidsRes. 31, 1614–1623
44 Kennell, J.C. and Cohen, S.M. In Handbook of Fungal Biotechnology(Arora, D., ed.), Marcel Dekker (in press)
45 Schardl, C.L. et al. (1985) Mitochondrial DNA rearrangements associ-ated with fertile revertants of S-type male-sterile maize. Cell 43,361–368
46 Chesnick, J.M. et al. (2000) The mitochondrial genome of thestramenopile alga Chrysodidymus synuroideus. Complete sequence,gene content and genome organization. Nucleic Acids Res. 28,2512–2518
47 Forget, L. et al. (2002) Hyaloraphidium curvatum: a linear mitochon-drial genome, tRNA editing, and an evolutionary link to lower fungi.Mol. Biol. Evol. 19, 310–319
48 Bullerwell, C.E. et al. (2003) A comparison of three fission yeastmitochondrial genomes. Nucleic Acids Res. 31, 759–768
49 Marienfeld, J. et al. (1999) The mitochondrial genome of Arabidopsis iscomposed of both native and immigrant information. Trends Plant Sci.4, 495–502
50 Koulintchenko, M. et al. (2003) Plant mitochondria actively import DNAvia the permeability transition pore complex. EMBO J. 22, 1245–1254
51 Adams, K.L. et al. (2000) Repeated, recent and diverse transfers of amitochondrial gene to the nucleus in flowering plants. Nature 408,354–357
52 Adams, K.L. et al. (2001) Multiple losses and transfers to the nucleus oftwo mitochondrial succinate dehydrogenase genes during angiospermevolution. Genetics 158, 1289–1300
53 Adams, K.L. et al. (2002) Punctuated evolution of mitochondrial genecontent: high and variable rates of mitochondrial gene loss andtransfer to the nucleus during angiosperm evolution. Proc. Natl. Acad.Sci. U. S. A. 99, 9905–9912
54 Chomyn, A. and Attardi, G. (2003) MtDNA mutations in aging andapoptosis. Biochem. Biophys. Res. Commun. 304, 519–529
55 Wei, Y.H. and Lee, H.C. (2002) Oxidative stress, mitochondrial DNAmutation, and impairment of antioxidant enzymes in aging. Exp. Biol.Med. (Maywood) 227, 671–682
56 Lonergan, K.M. and Gray, M.W. (1994) The ribosomal RNA gene regionin Acanthamoeba castellanii mitochondrial DNA. A case of evolution-ary transfer of introns between mitochondria and plastids? J. Mol.Biol. 239, 476–499
57 Turmel, M. et al. (1995) Evolutionary transfer of ORF-containinggroup I introns between different subcellular compartments (chlor-oplast and mitochondrion). Mol. Biol. Evol. 12, 533–545
58 Burger, G. et al. (1999) Complete sequence of the mitochondrial DNA ofthe red alga Porphyra purpurea. Cyanobacterial introns and sharedancestry of red and green algae. Plant Cell 11, 1675–1694
59 Lang, B.F. (1984) The mitochondrial genome of the fission yeastSchizosaccharomyces pombe: highly homologous introns are insertedat the same position of the otherwise less conserved cox1 genes inSchizosaccharomyces pombe and Aspergillus nidulans. EMBO J. 3,2129–2136
60 Cho, Y. et al. (1998) Explosive invasion of plant mitochondria by agroup I intron. Proc. Natl. Acad. Sci. U. S. A. 95, 14244–14249
61 Cho, Y. and Palmer, J.D. (1999) Multiple acquisitions via horizontaltransfer of a group I intron in the mitochondrial cox1 gene duringevolution of the Araceae family. Mol. Biol. Evol. 16, 1155–1165
62 Paquin, B. et al. (1994) Interspecific transfer of mitochondrial genes infungi and creation of a homologous hybrid gene. Proc. Natl. Acad. Sci.U. S. A. 91, 11807–11810
63 Bergthorsson, U. et al. (2003) Widespread horizontal transfer ofmitochondrial genes in flowering plants. Nature 424, 197–201
64 Hanson, M.R. and Folkerts, O. (1992) Structure and function of thehigher plant mitochondrial genome. Int. J. Cytol. 141, 129–172
65 Anderson, S. et al. (1981) Sequence and organization of the humanmitochondrial genome. Nature 290, 457–465
66 Lander, E.S. et al. (2001) Initial sequencing and analysis of the humangenome. Nature 409, 860–921
67 Venter, J.C. et al. (2001) The sequence of the human genome. Science291, 1304–1351
68 Foury, F. et al. (1998) The complete sequence of the mitochondrialgenome of Saccharomyces cerevisiae. FEBS Lett. 440, 325–331
69 Giaever, G. et al. (2002) Functional profiling of the Saccharomycescerevisiae genome. Nature 418, 387–391
70 Elmerot, C. et al. (2002) The mitochondrial genome of the pufferfish,Fugu rubripes, and ordinal teleostean relationships. Gene 295,163–172
71 Aparicio, S. et al. (2002) Whole-genome shotgun assembly and analysisof the genome of Fugu rubripes. Science 297, 1301–1310
72 Thorsness, P.E. and Fox, T.D. (1990) Escape of DNA from mitochondriato the nucleus in Saccharomyces cerevisiae. Nature 346, 376–379
73 Woischnik, M. and Moraes, C.T. (2002) Pattern of organization ofhuman mitochondrial pseudogenes in the nuclear genome. GenomeRes. 12, 885–893
74 Wolff, G. et al. (1994) Complete sequence of the mitochondrial DNA ofthe chlorophyte alga Prototheca wickerhamii. Gene content andgenome organization. J. Mol. Biol. 237, 75–86
75 Kuck, U. et al. (2000) DNA sequence analysis of the completemitochondrial genome of the green alga Scenedesmu obliquus:evidence for UAG being a leucine and UCA being a non-sense codon.Gene 253, 13–18
76 Gray, M.W. and Boer, P.H. (1988) Organization and expression of algal(Chlamydomonas reinhardtii) mitochondrial DNA. Philos. Trans.R. Soc. Lond. B Biol. Sci. 319, 135–147
77 Tessier, L.H. et al. (1997) The cox1 gene from Euglena gracilis: a protistmitochondrial gene without introns and genetic code modifications.Curr. Genet. 31, 208–213
78 Covello, P.S. and Gray, M.W. (1992) Silent mitochondrial and activenuclear genes for subunit 2 of cytochrome c oxidase (cox2) in soybean:evidence for RNA-mediated gene transfer. EMBO J. 11, 3815–3820
79 Gabaldon, T. and Huynen, M.A. (2003) Reconstruction of the proto-mitochondrial metabolism. Science 301, 609
80 Rasmusson, A.G. et al. (1999) Homologues of yeast and bacterialrotenone-insensitive NADH dehydrogenases in higher eukaryotes:two enzymes are present in potato mitochondria. Plant J. 20, 79–87
81 Baldauf, S.L. (2003) The deep roots of eukaryotes. Science 300,1703–1706
82 Cavalier-Smith, T. (1983) A 6-kingdom classification and a unifiedphylogeny. In Endocytobiology II (Schwemmler, W. and Schenk,H.E.A., eds), pp. 1027–1034, De Gruyter
83 Martin, W. and Muller, M. (1998) The hydrogen hypothesis for the firsteukaryote. Nature 392, 37–41
84 Cermakian, N. et al. (1997) On the evolution of the single-subunit RNApolymerases. J. Mol. Evol. 45, 671–681
Review TRENDS in Genetics Vol.19 No.12 December 2003716
http://tigs.trends.com