Embed Size (px)
Citation preview

Models of Circadian Rhythms
Daniel Forger

~24-hour Clocks time important events



Circadian Clocks important for
• Bipolar Disorder• FASPS (and other Sleep Disorders)• Alzheimer’s Disease (institutionalization)• Leukemia (chemotherapy most effective at
night, direct link to cell cycle and cancer)• Cholesterol (4-fold increase in biosynthesis at
night controlled by liver pacemakers)• The Blind (can’t synchronize)• Schedules/Missions/Shiftwork

movies

Tyson-Hong-Thron Novak model of circadian rhythms in
Drosophila
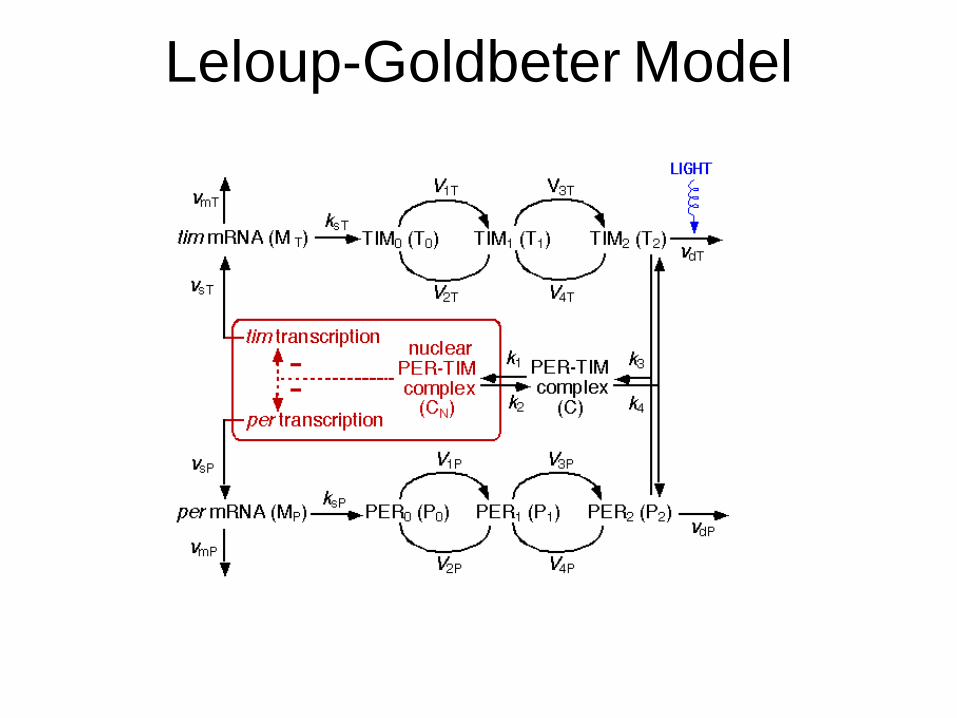
Leloup-Goldbeter Model


Equations Transcription Translation Degradation Phosphorylation Binding Nuclear TransportL = Light LeveldGRv/dt = binRv*RvnRvn *(1 - GRv) - unbinRv*GRvdMnRo/dt = trRo*(1 - G)*(1 - GRv)3 - tmc*MnRodMcRo/dt = tmc*MnRo - umR*McRodMnRt/dt = trRt*(1 - G) - tmc*MnRtdMcRt/dt = tmc*MnRt - umR*McRtdMnPo/dt = trPo*(1 - G)5 + L - tmc*MnPodMcPo/dt = tmc*MnPo - umPo*McPodMnPt/dt = trPt*(1 - G)5 + L - tmc*MnPtdMcPt/dt = tmc*MnPt - umPt*McPtdMnRv/dt = trRv*(1 - G)3 - tmc*MnRvdMcRv/dt = tmc*MnRv - umRv*McRvdRv/dt = tlrv*McRv - 2*arv*Rv*Rv + 2*drv*RvRv - nl*Rv + ne*Rvn - uRv*RvdRvn/dt = - 2*Nf*arv*Rvn*Rvn + 2*drv*RvnRvn + nl*Rv - ne*Rvn - uRv*RvndRvRv/dt = arv*Rv*Rv - drv*RvRv - nl*RvRv + ne*RvnRvn - 2*uRv*RvRvdRvnRvn/dt = Nf*arv*Rvn*Rvn - drv*RvnRvn + nl*RvRv - ne*RvnRvn - 2*uRv*RvnRvndPo/dt = tlp*McPo - ac*Po*C + dc*PoC - upu*PodPt/dt = tlp*McPt - ac*Pt*C + dc*PtC - upu*PodPoC/dt = ac*Po*C - dc*PoC - hoo*PoCdPtC/dt = ac*Pt*C - dc*PtC - hot*PtCdPopC/dt = hoo*PoC + ac*Pop*C - dc*PopC - up*PopC - ht*PopC- nl*PopC + ne*PonpCn - ar*PopC*Ro + dr*PopCRo - ar*PopC*Rt + dr*PopCRtdPtpC/dt = hot*PtC + ac*Ptp*C - dc*PtpC - up*PtpC - ht*PtpC- nl*PtpC + ne*PtnpCn - ar*PtpC*Ro + dr*PtpCRo - ar*PtpC*Rt + dr*PtpCRtdPop/dt = - ac*Pop*C+ dc*PopC - up*Pop - ar*Pop*Ro + dr*PopRo - ar*Pop*Rt + dr*PopRt - nl*Pop + ne*PonpdPtp/dt = - ac*Ptp*C+ dc*PtpC - up*Ptp - ar*Ptp*Ro + dr*PtpRo - ar*Ptp*Rt + dr*PtpRt - nl*Ptp + ne*PtnpdPoppC/dt = ht*PopC - up*PoppC+ ac*Popp*C- dc*PoppC + ne*PonppCn - ar*PoppC*Ro + dr*PoppCRo - ar*PoppC*Rt + dr*PoppCRtdPtppC/dt = ht*PtpC - up*PtppC+ ac*Ptpp*C- dc*PtppC + ne*PtnppCn - ar*PtppC*Ro + dr*PtppCRo - ar*PtppC*Rt + dr*PtppCRtdPopRo/dt = ar*Pop*Ro - dr*PopRo - ac*PopRo*C + dc*PopCRo - nl*PopRo + ne*PonpRondPtpRo/dt = ar*Ptp*Ro - dr*PtpRo - ac*PtpRo*C+ dc*PtpCRo - nl*PtpRo + ne*PtnpRondPopRt/dt = ar*Pop*Rt - dr*PopRt - ac*PopRt*C+ dc*PopCRt - nl*PopRt + ne*PonpRtndPtpRt/dt = ar*Ptp*Rt - dr*PtpRt - ac*PtpRt*C + dc*PtpCRt - nl*PtpRt + ne*PtnpRtndPoppRo/dt = ar*Popp*Ro - dr*PoppRo - ac*PoppRo*C + dc*PoppCRo + ne*PonppRondPoppRt/dt = ar*Popp*Rt - dr*PoppRt - ac*PoppRt*C + dc*PoppCRt + ne*PonppRtndPtppRo/dt = ar*Ptpp*Ro - dr*PtppRo - ac*PtppRo*C + dc*PtppCRo + ne*PtnppRondPtppRt/dt = ar*Ptpp*Rt - dr*PtppRt - ac*PtppRt*C + dc*PtppCRt + ne*PtnppRtndPopp/dt = - ac*Popp*C + dc*PoppC + ne*Ponpp - ar*Popp*Ro + dr*PoppRo –ar*Popp*Rt + dr*PoppRt - up*PoppdPtpp/dt = - ac*Ptpp*C+ dc*PtppC+ ne*Ptnpp - ar*Ptpp*Ro + dr*PtppRo - ar*Ptpp*Rt + dr*PtppRt - up*PtppdPopCRo/dt = ar*PopC*Ro - dr*PopCRo + ac*PopRo*C- dc*PopCRo - nl*PopCRo + ne*PonpCnRon - ht*PopCRodPtpCRo/dt = ar*PtpC*Ro - dr*PtpCRo + ac*PtpRo*C - dc*PtpCRo - nl*PtpCRo + ne*PtnpCnRon - ht*PtpCRodPopCRt/dt = ar*PopC*Rt - dr*PopCRt + ac*PopRt*C- dc*PopCRt - nl*PopCRt + ne*PonpCnRtn - ht*PopCRtdPtpCRt/dt = ar*PtpC*Rt - dr*PtpCRt + ac*PtpRt*C - dc*PtpCRt - nl*PtpCRt + ne*PtnpCnRtn - ht*PtpCRtdPoppCRo/dt = ar*PoppC*Ro - dr*PoppCRo + ac*PoppRo*C- dc*PoppCRo + ne*PonppCnRon + ht*PopCRo

Equations Cont. Transcription Translation Degradation Phosphorylation Binding Nuclear Transport
dPtppCRo/dt = ar*PtppC*Ro - dr*PtppCRo + ac*PtppRo*C - dc*PtppCRo + ne*PtnppCnRon + ht*PtpCRodPoppCRt/dt = ar*PoppC*Rt- dr*PoppCRt+ ac*PoppRt*C - dc*PoppCRt+ ne*PonppCnRtn + ht*PopCRtdPtppCRt/dt = ar*PtppC*Rt- dr*PtppCRt+ ac*PtppRt*C - dc*PtppCRt+ ne*PtnppCnRtn + ht*PtpCRtdRo/dt = - ar*Ro*Pop - ar*Ro*Popp - ar*Ro*PopC - ar*Ro*PoppC + dr*PopRo + dr*PoppRo + dr*PopCRo + dr*PoppCRo - ar*Ro*Ptp - ar*Ro*Ptpp - ar*Ro*PtpC - ar*Ro*PtppC +
dr*PtpRo + dr*PtppRo + dr*PtpCRo + dr*PtppCRo + tlr*McRo - uro*RodRt/dt = - ar*Rt*Pop - ar*Rt*Popp - ar*Rt*PopC - ar*Rt*PoppC + dr*PopRt+ dr*PoppRt+ dr*PopCRt+ dr*PoppCRt- ar*Rt*Ptp - ar*Rt*Ptpp - ar*Rt*PtpC - ar*Rt*PtppC +
dr*PtpRt+ dr*PtppRt+ dr*PtpCRt+ dr*PtppCRt+ tlr*McRt - urt*RtdPonpCn/dt = ac*Nf*Ponp*Cn - dc*PonpCn - ht*PonpCn + nl*PopC - ne*PonpCn - ar*Nf*PonpCn*Ron + dr*PonpCnRon - ar*Nf*PonpCn*Rtn + dr*PonpCnRtn - upn*PonpCndPtnpCn/dt = ac*Nf*Ptnp*Cn - dc*PtnpCn - ht*PtnpCn + nl*PtpC - ne*PtnpCn - ar*Nf*PtnpCn*Ron + dr*PtnpCnRon - ar*Nf*PtnpCn*Rtn + dr*PtnpCnRtn - upn*PtnpCndPonp/dt = - ac*Nf*Ponp*Cn + dc*PonpCn - ar*Nf*Ponp*Ron + dr*PonpRon - ar*Nf*Ponp*Rtn + dr*PonpRtn + nl*Pop - ne*Ponp - upn*PonpdPtnp/dt = - ac*Nf*Ptnp*Cn + dc*PtnpCn - ar*Nf*Ptnp*Ron + dr*PtnpRon - ar*Nf*Ptnp*Rtn + dr*PtnpRtn + nl*Ptp - ne*Ptnp - upn*PtnpdPonppCn/dt = ht*PonpCn + ac*Nf*Ponpp*Cn - dc*PonppCn - ne*PonppCn - ar*Nf*PonppCn*Ron + dr*PonppCnRon – ar*Nf*PonppCn*Rtn + dr*PonppCnRtn - upn*PonppCndPtnppCn/dt = ht*PtnpCn + ac*Nf*Ptnpp*Cn - dc*PtnppCn - ne*PtnppCn - ar*Nf*PtnppCn*Ron + dr*PtnppCnRon - ar*Nf*PtnppCn*Rtn + dr*PtnppCnRtn - upn*PtnppCndPonpRon/dt = ar*Nf*Ponp*Ron - dr*PonpRon - ac*Nf*PonpRon*Cn + dc*PonpCnRon + nl*PopRo - ne*PonpRondPtnpRon/dt = ar*Nf*Ptnp*Ron - dr*PtnpRon - ac*Nf*PtnpRon*Cn + dc*PtnpCnRon + nl*PtpRo - ne*PtnpRondPonpRtn/dt = ar*Nf*Ponp*Rtn - dr*PonpRtn - ac*Nf*PonpRtn*Cn + dc*PonpCnRtn + nl*PopRt- ne*PonpRtndPtnpRtn/dt = ar*Nf*Ptnp*Rtn - dr*PtnpRtn - ac*Nf*PtnpRtn*Cn + dc*PtnpCnRtn + nl*PtpRt- ne*PtnpRtndPonppRon/dt = ar*Nf*Ponpp*Ron - dr*PonppRon - ac*Nf*PonppRon*Cn + dc*PonppCnRon - ne*PonppRondPtnppRon/dt = ar*Nf*Ptnpp*Ron - dr*PtnppRon - ac*Nf*PtnppRon*Cn + dc*PtnppCnRon - ne*PtnppRondPonppRtn/dt = ar*Nf*Ponpp*Rtn - dr*PonppRtn - ac*Nf*PonppRtn*Cn + dc*PonppCnRtn - ne*PonppRtndPtnppRtn/dt = ar*Nf*Ptnpp*Rtn - dr*PtnppRtn - ac*Nf*PtnppRtn*Cn + dc*PtnppCnRtn - ne*PtnppRtndPonpp/dt = - ac*Nf*Ponpp*Cn + dc*PonppCn - ne*Ponpp - ar*Nf*Ponpp*Ron + dr*PonppRon - ar*Nf*Ponpp*Rtn + dr*PonppRtn - upn*PonppdPtnpp/dt = - ac*Nf*Ptnpp*Cn + dc*PtnppCn - ne*Ptnpp - ar*Nf*Ptnpp*Ron + dr*PtnppRon - ar*Nf*Ptnpp*Rtn + dr*PtnppRtn - upn*PtnppdPonpCnRon/dt = ar*Nf*PonpCn*Ron - dr*PonpCnRon + ac*Nf*PonpRon*Cn - dc*PonpCnRon + nl*PopCRo - ne*PonpCnRon - ht*PonpCnRondPtnpCnRon/dt = ar*Nf*PtnpCn*Ron - dr*PtnpCnRon + ac*Nf*PtnpRon*Cn - dc*PtnpCnRon + nl*PtpCRo - ne*PtnpCnRon - ht*PtnpCnRondPonpCnRtn/dt = ar*Nf*PonpCn*Rtn - dr*PonpCnRtn + ac*Nf*PonpRtn*Cn - dc*PonpCnRtn + nl*PopCRt- ne*PonpCnRtn - ht*PonpCnRtndPtnpCnRtn/dt = ar*Nf*PtnpCn*Rtn - dr*PtnpCnRtn + ac*Nf*PtnpRtn*Cn - dc*PtnpCnRtn + nl*PtpCRt- ne*PtnpCnRtn - ht*PtnpCnRtndPonppCnRon/dt = ar*Nf*PonppCn*Ron - dr*PonppCnRon + ac*Nf*PonppRon*Cn - dc*PonppCnRon - ne*PonppCnRon + ht*PonpCnRondPtnppCnRon/dt = ar*Nf*PtnppCn*Ron - dr*PtnppCnRon + ac*Nf*PtnppRon*Cn - dc*PtnppCnRon - ne*PtnppCnRon + ht*PtnpCnRondPonppCnRtn/dt = ar*Nf*PonppCn*Rtn - dr*PonppCnRtn + ac*Nf*PonppRtn*Cn - dc*PonppCnRtn - ne*PonppCnRtn + ht*PonpCnRtndPtnppCnRtn/dt = ar*Nf*PtnppCn*Rtn - dr*PtnppCnRtn + ac*Nf*PtnppRtn*Cn - dc*PtnppCnRtn - ne*PtnppCnRtn + ht*PtnpCnRtndRon/dt = - ar*Nf*Ron*Ponp - ar*Nf*Ron*Ponpp - ar*Nf*Ron*PonpCn - ar*Nf*Ron*PonppCn + dr*PonpRon + dr*PonppRon + dr*PonpCnRon + dr*PonppCnRon -
ar*Nf*Ron*Ptnp - ar*Nf*Ron*Ptnpp - ar*Nf*Ron*PtnpCn - ar*Nf*Ron*PtnppCn + dr*PtnpRon + dr*PtnppRon + dr*PtnpCnRon + dr*PtnppCnRon - uro*RondRtn/dt = - ar*Nf*Rtn*Ponp - ar*Nf*Rtn*Ponpp - ar*Nf*Rtn*PonpCn - ar*Nf*Rtn*PonppCn + dr*PonpRtn + dr*PonppRtn + dr*PonpCnRtn + dr*PonppCnRtn - ar*Nf*Rtn*Ptnp -
ar*Nf*Rtn*Ptnpp - ar*Nf*Rtn*PtnpCn - ar*Nf*Rtn*PtnppCn + dr*PtnpRtn + dr*PtnppRtn + dr*PtnpCnRtn + dr*PtnppCnRtn - urt*RtndCn/dt = - ac*Nf*Cn*Ponp - ac*Nf*Cn*Ponpp - ac*Nf*Cn*PonpRon - ac*Nf*Cn*PonppRon + dc*PonpCn + dc*PonppCn + dc*PonpCnRon + dc*PonppCnRon - ac*Nf*Cn*Ptnp -
ac*Nf*Cn*Ptnpp - ac*Nf*Cn*PtnpRon - ac*Nf*Cn*PtnppRon + dc*PtnpCn + dc*PtnppCn + dc*PtnpCnRon + dc*PtnppCnRon - ac*Nf*Cn*PonpRtn - ac*Nf*Cn*PonppRtn + dc*PonpCnRtn + dc*PonppCnRtn - ac*Nf*Cn*PtnpRtn - ac*Nf*Cn*PtnppRtn + dc*PtnpCnRtn + dc*PtnppCnRtn + upn*PonpCn + upn*PonppCn + upn*PtnpCn + upn*PtnppCn
RelationsC = Ct - (Ron + PonpRon + PonppRon + PonpCnRon + PonppCnRon + PtnpRon + PtnppRon + PtnpCnRon + PtnppCnRon + Rtn + PonpRtn + PonppRtn + PonpCnRtn +
PonppCnRtn + PtnpRtn + PtnppRtn + PtnpCnRtn + PtnppCnRtn)Rn = (Ron + PonpRon + PonppRon + PonpCnRon + PonppCnRon + PtnpRon + PtnppRon + PtnpCnRon + PtnppCnRon + Rtn + PonpRtn + PonppRtn + PonpCnRtn +
PonppCnRtn + PtnpRtn + PtnppRtn + PtnpCnRtn + PtnppCnRtn

Tau mutation
• First discovered mutation in mammals– Subject of the widest study (many science papers)
• Missense mutation in CKIe which decreases kinase activity
• Similar mutations in Drosophila• Similar mutations in man (FASPS, CKId-T44A)
cause sleep disorders• Phenotype:
– 22 hour period in heterozygotes– 20 hour period in homozygotes
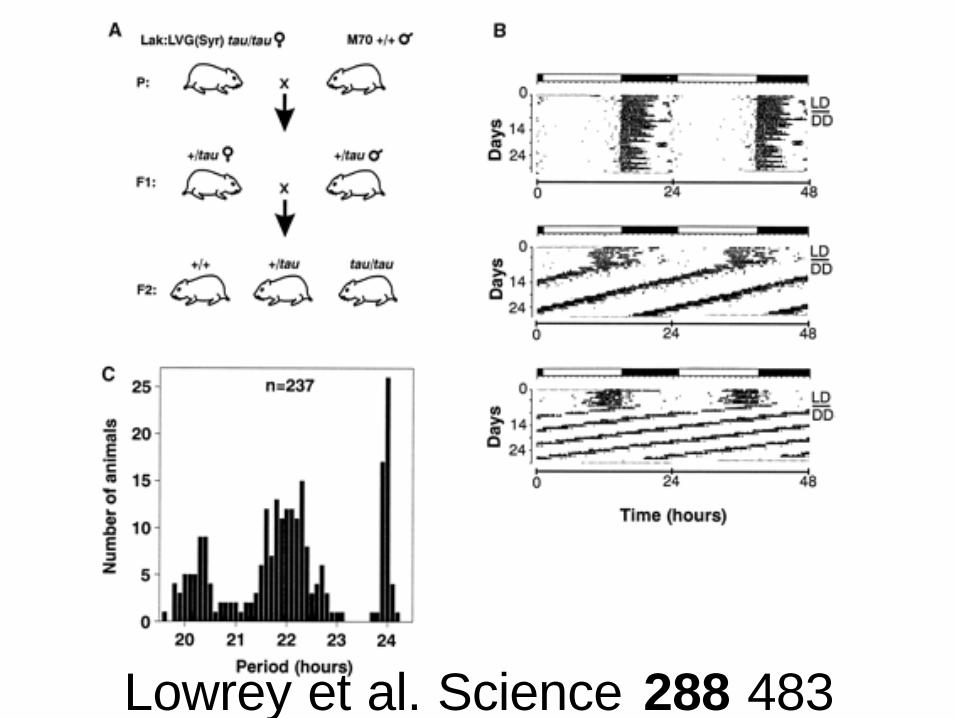
Lowrey et al. Science 288 483
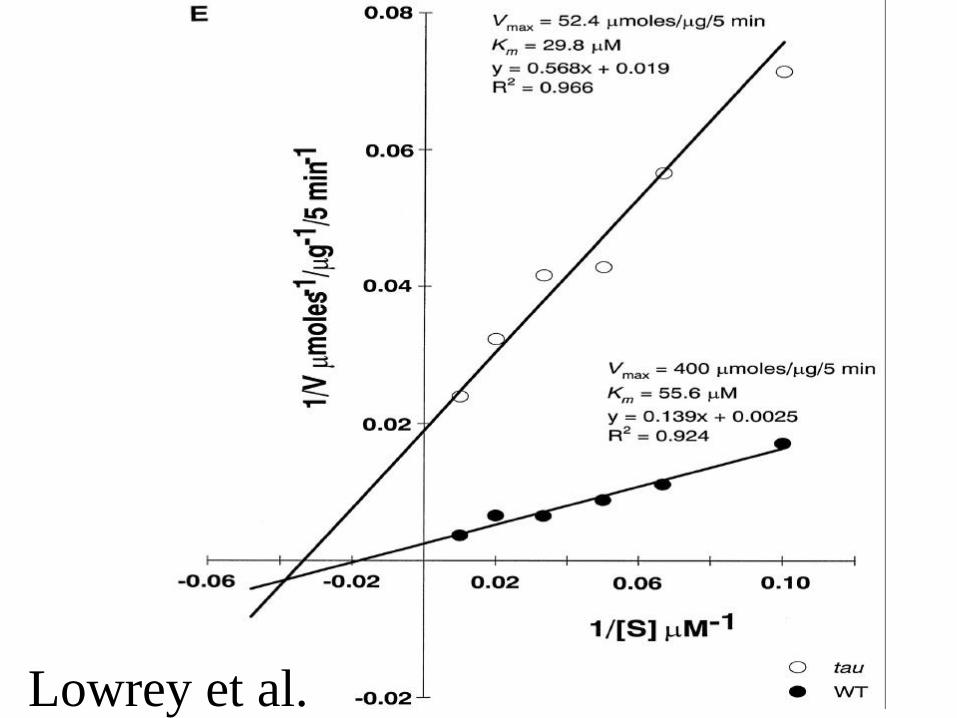
Lowrey et al.

Xu et al. Nature 434, 640
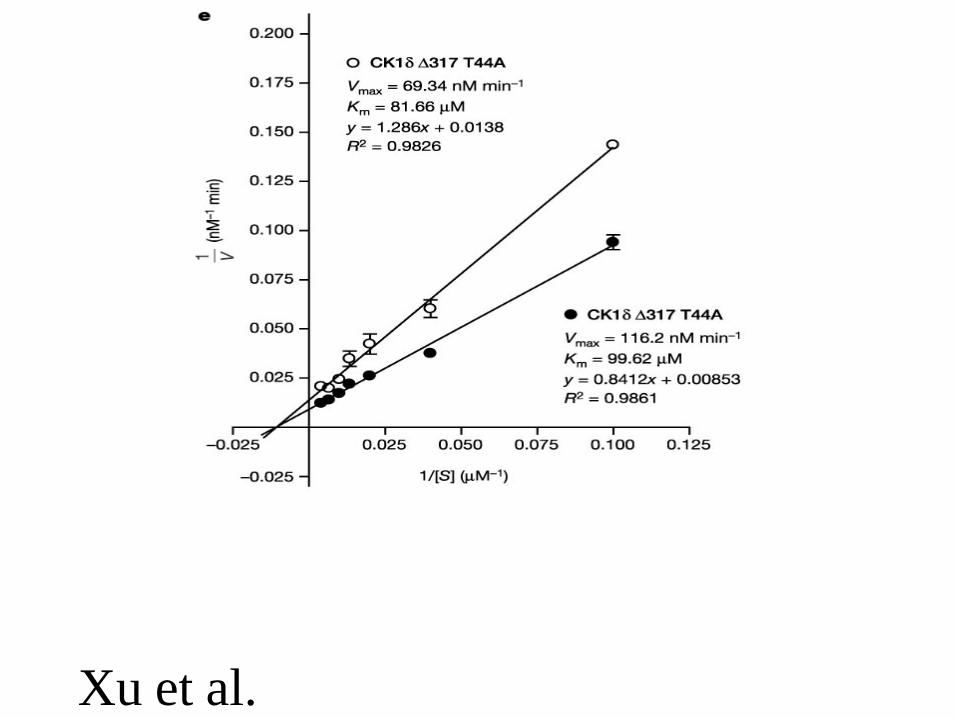
Xu et al.

Tau mutant predictions• WT period 24.2991• Half primary phosphorylation rate of PER1
– 24.43• Half secondary phosphorylation rate of PER1
– 26.73• Half primary phosphorylation rate of PER2
– 24.33• Half binding rate to kinases
– 26.62• Decreased BMAL activation
– 24.42

80 100 120 140Percent Rate
24.25
24.3
24.35
24.4
Period
80 100 120 140Percent Rate
23.9
24.124.224.3
24.424.5
Period
80 100 120 140Percent Rate
24.5
25
25.5
Period
80 100 120 140Percent Rate
24
25
26
Period
80 100 120 140Percent Rate
24.2
24.3
24.4
Period
80 100 120 140Percent Rate
24.27
24.28
24.29
24.31
24.32
24.33Period
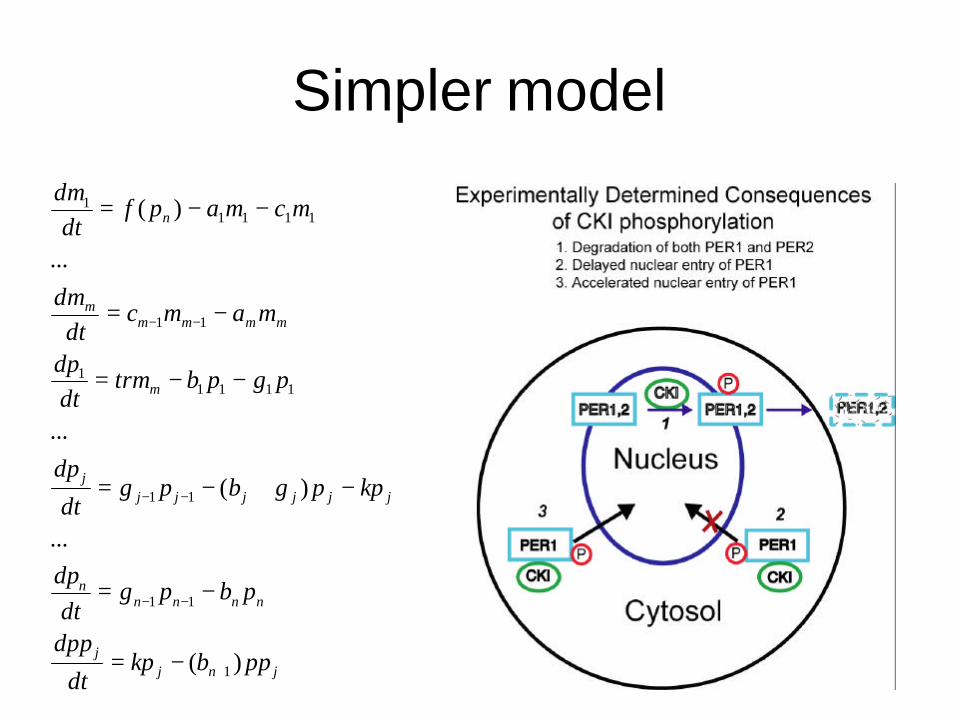
Simpler model
jnjj
nnnnn
jjjjjjj
m
mmmmm
n
ppbkpdt
dpp
pbpgdt
dp
kppgbpgdt
dp
pgpbtrmdtdp
mamcdt
dm
mcmapfdt
dm
)(
...
)(
...
...
)(
1
11
11
11111
11
11111
+
−−
−−
−−
−=
−=
−+−=
−−=
−=
−−=

Period Estimate
...
)(...
)(...
11
11
jjjjjj
jjjjjj
PkgbPgiwP
McaMciwM
++−=
+−=
−−
−−
...(iw + a j + c j )...(iw + b j + g j + k)...F(pn )
Pn
c1...cm−1trg1...gn−1
=1
Decreasing gj, k or bj (because of a lower phosphorylation rate) increases the phase of (iw + bj+gj+k). To balance this, wmust decrease and the period lengthens.

Nothing in the Model can explain tau!
(get help from friends)
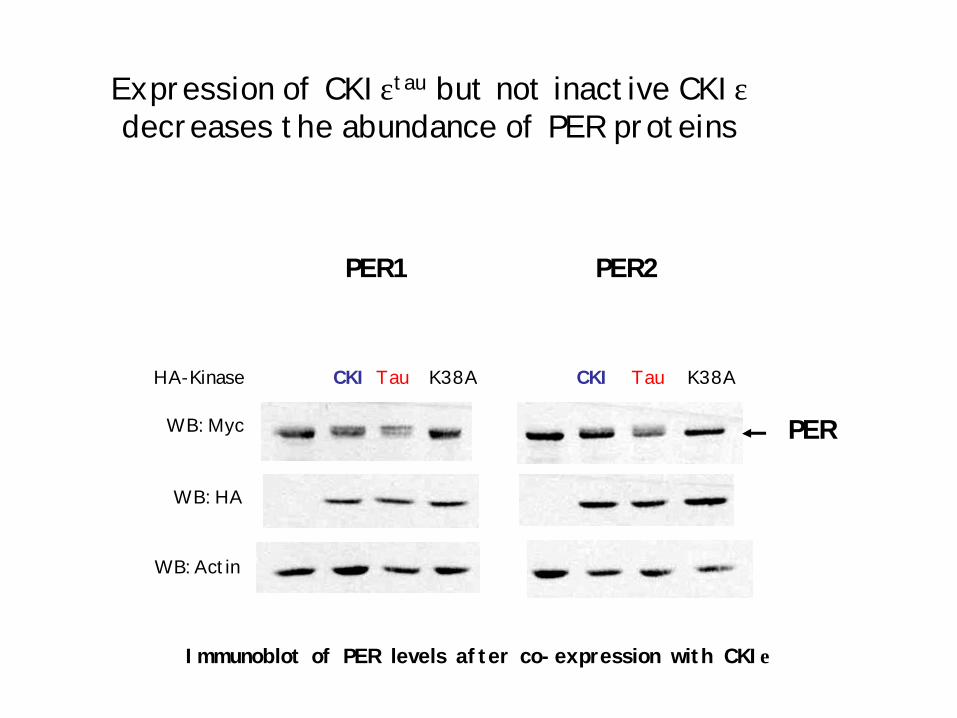
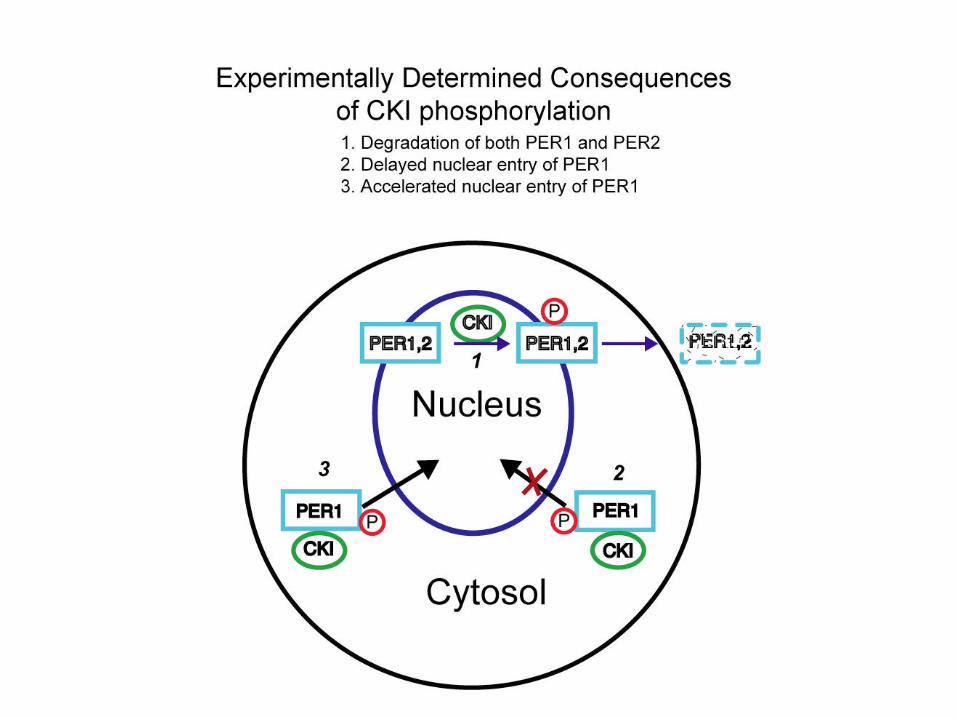
Expression of CKIεtau but not inactive CKIεdecreases the abundance of PER proteins
CKI Tau K38A CKI Tau K38A
WB: Myc
WB: Actin
WB: HA
HA-Kinase
PER1 PER2
Immunoblot of PER levels after co-expression with CKIε
PER

0 3
Tau K38ACKI1.5 5 0 31.5 5 0 31.5 5 0 31.5 5CHX(hrs)
PER2
HA-CKIε
Actin
Tau CKI shortens PER2 half-life
MG

UbUbUbUb
Phosphorylation-dependent mPER2 proteolysis
26S Proteasome
Per2
P P
PP
+ +β−TrCP
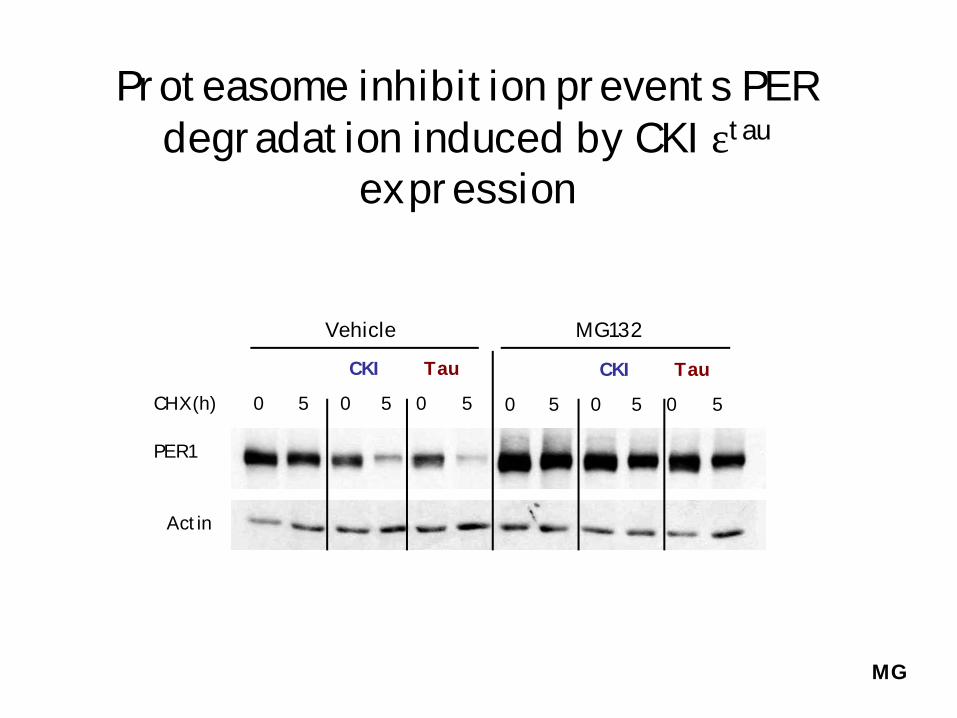
Proteasome inhibition prevents PER degradation induced by CKIεtau
expression
0 5 0 5 0 5
CKI Tau
CHX(h)
Vehicle
0 5 0 5 0 5
CKI Tau
MG132
PER1
Actin
MG
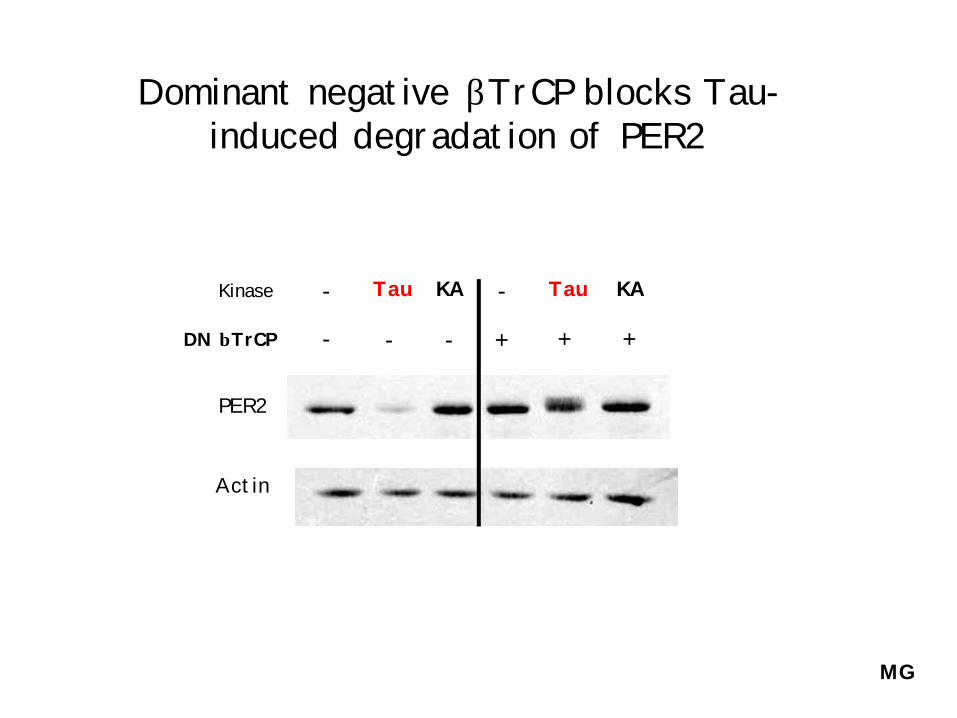
Dominant negative βTrCP blocks Tau-induced degradation of PER2
Kinase
DN βTrCP
Tau KA- Tau KA-
- - - + + +
PER2
Actin
MG

HA-Kinase Tau KA- CKI Tau KA- CKIWT-PER2 PER2(S477A/G479A)
PER2
Actin
Mutation of βTrCP binding site blocks Tau-induced degradation (but not phosphorylation) of PER2
NLSCKICLD Cry
450 755
PAS
MG

The tau mutation in CKIεincreases activity in vivo
• Is this really increased kinase activity?• Is it specific to PER and circadian rhythm?
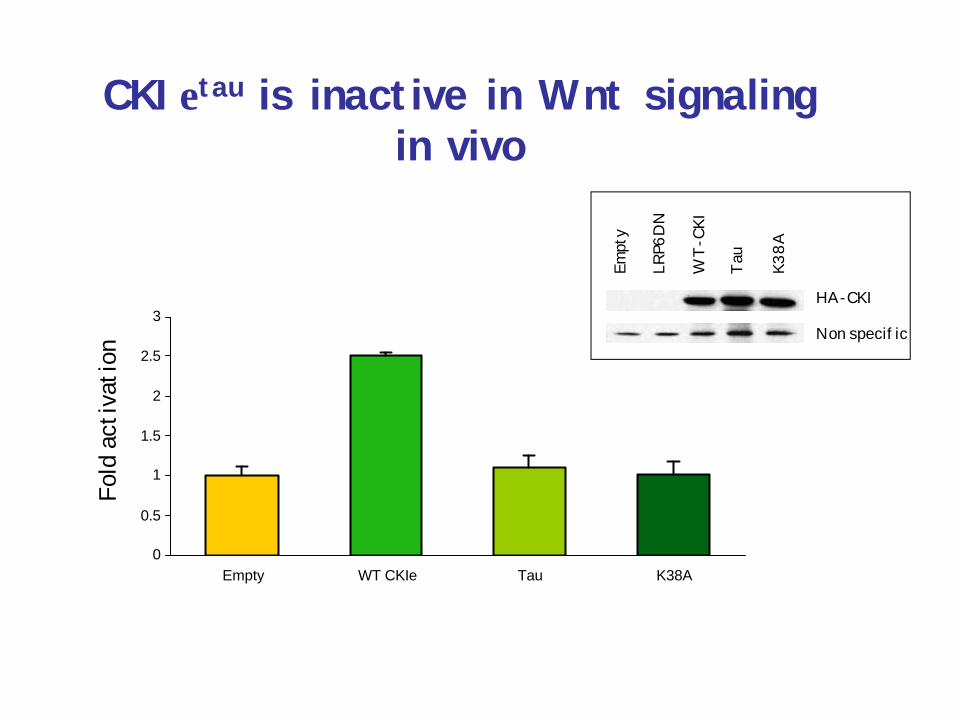
0
0.5
1
1.5
2
2.5
3
Empty WT CKIe Tau K38A
Fold
act
ivat
ion
CKIεtau is inactive in Wnt signaling in vivo
Empt
y
LRP6
DN
WT-
CKI
Tau
K38A
HA-CKI
Non specific

Phosphopeptide mapping of in vivometabolically labeled PER2
8
1
2a2b2c
3a3b 4a
4b
5
6
7
CKIεtau
1
2a2b
2c
3a
3b4a
4b
5
6
7
vector
700 cpm/TLC plate, 96 hr autoradiographic exposure EJE

Can Simulate these Directly(Gillespie’s SSA Method)
• Choose randomly among all possible reactions (weighted by their rates)
• Find the time until the next reaction:
• For speedup, do the above for reaction classes, and then choose individual reaction
• # of molecules given by experiments (100s to 1000s)• Measure period by several upcrossings (mean, 80% of
mean, 120% of mean etc.)
=
00
1ln
1ra
τ
PopCn ® Popn + Cn dc*PopCnPoppCn ® Poppn + Cn dc*PoppCnPtpCn ® Ptpn + Cn dc*PtpCnPtppCn ® Ptppn + Cn dc*PtppCnPopCRon ® PopRon + Cn dc*PopCRonPoppCRon ® PoppRon + Cn dc*PoppCRonPopCRtn ® PopRtn + Cn dc*PopCRtnPoppCRtn ® PoppRtn + Cn dc*PoppCRtnPtpCRon ® PtpRon + Cn dc*PtpCRonPtppCRon ® PtppRon + Cn dc*PtppCRonPtpCRtn ® PtpRtn + Cn dc*PtpCRtnPtppCRtn ® PtppRtn + Cn dc*PtppCRtnPoC ® PopC hoo*PoCPopC ® PoppC hto*PopCPtC ® PtpC hot*PtCPtpC ® PtppC ht*PtpCPopCn ® PoppCn hto*PopCnPtpCn ® PtppCn ht*PtpCnPopCRo ® PoppCRo hto*PopCRoPopCRt ® PoppCRt hto*PopCRtPtpCRo ® PtppCRo ht*PtpCRoPtpCRt ® PtppCRt ht*PtpCRtPopCRon ® PoppCRon hto*PopCRonPopCRtn ® PoppCRtn hto*PopCRtnPtpCRon ® PtppCRon ht*PtpCRonPtpCRtn ® PtppCRtn ht*PtpCRtnPop + Ro ® PopRo ar*Pop*RoPopp + Ro ® PoppRo ar*Popp*RoPop + Rt ® PopRt ar*Pop*RtPopp + Rt ® PoppRt ar*Popp*RtPtp + Ro ® PtpRo ar*Ptp*RoPtpp + Ro ® PtppRo ar*Ptpp*RoPtp + Rt ® PtpRt ar*Ptp*RtPtpp + Rt ® PtppRt ar*Ptpp*RtPopC + Ro ® PopCRo ar*PopC*RoPoppC + Ro ® PoppCRo ar*PoppC*RoPopC + Rt ® PopCRt ar*PopC*RtPoppC + Rt ® PoppCRt ar*PoppC*RtPtpC + Ro ® PtpCRo ar*PtpC*RoPtppC + Ro ® PtppCRo ar*PtppC*RoPtpC + Rt ® PtpCRt ar*PtpC*RtPtppC + Rt ® PtppCRt ar*PtppC*RtPopn + Ron ® PopRon ar*N*Popn*RonPoppn + Ron ® PoppRon ar*N*Poppn*RonPopn + Rtn ® PopRtn ar*N*Popn*RtnPoppn + Rtn ® PoppRtn ar*N*Poppn*RtnPtpn + Ron ® PtpRon ar*N*Ptpn*Ron
Reaction RatePo + C ® PoC ac*Po*CPop + C ® PopC ac*Pop*CPopp + C ® PoppC ac*Popp*CPt + C ® PtC ac*Pt*CPtp + C ® PtpC ac*Ptp*CPtpp + C ® PtppC ac*Ptpp*CPopRo + C ® PopCRo ac*PopRo*CPoppRo + C ->® PoppCRo ac*PoppRo*CPopRt + C ->® PopCRt ac*PopRt*CPoppRt + C ->® PoppCRt ac*PoppRt*CPtpRo + C ® PtpCRo ac*PtpRo*CPtppRo + C ® PtppCRo ac*PtppRo*CPtpRt + C ® PtpCRt ac*PtpRt*CPtppRt + C ® PtppCRt ac*PtppRt*CPopn + Cn ® PopCn ac*N*Popn*CnPoppn + Cn ® PoppCn ac*N*Poppn*CnPtpn + Cn ® PtpCn ac*N*Ptpn*CnPtppn + Cn ® PtppCn ac*N*Ptppn*CnPopRon + Cn ® PopCRon ac*N*PopRon*CnPoppRon + Cn ® PoppCRon ac*N*PoppRon*CPopRtn + Cn ® PopCRtn ac*N*PopRtn*CnPoppRtn + Cn ® PoppCRtn ac*N*PoppRtn*CnPtpRon + Cn ® PtpCRon ac*N*PtpRon*CnPtppRon + Cn ® PtppCRon ac*N*PtppRon*CnPtpRtn + Cn ®PtpCRtn ac*N*PtpRtn*CnPtppRtn + Cn ® PtppCRtn ac*N*PtppRtn*CnPoC ® Po + C dc*PoCPopC ® Pop + C dc*PopCPoppC ® Popp + C dc*PoppPtC ® Pt + C dc*PtCPtpC ® Ptp + C dc*PtpCPtppC ® Ptpp + C dc*PtppCPopCRo ® PopRo + C dc*PopCRoPoppCRo ® PoppRo + C dc*PoppCRoPopCRt ® PopRt + C dc*PopCRtPoppCRt ® PoppRt + C dc*PoppCRtPtpCRo ® PtpRo + C dc*PtpCRoPtppCRo ® PtppRo + C dc*PtppCRoPtpCRt ® PtpRt + C dc*PtpCRtPtppCRt ® PtppRt + C dc*PtppCRt

PopRo ® PopRon nl*PopRoPopRt ® PopRtn nl*PopRtPtpRo ® PtpRon nl*PtpRoPtpRt ® PtpRtn nl*PtpRtPopCRo ® PopCRon nl*PopCRoPopCRt ® PopCRtn nl*PopCRtPtpCRo ® PtpCRon nl*PtpCRoPtpCRt ® PtpCRtn nl*PtpCRtPopn ® Pop ne*PopnPtpn ® Ptp ne*PtpnPopCn ® PopC ne*PopCnPtpCn ® PtpC ne*PtpCnPopRon ® PopRo ne*PopRonPopRtn ® PopRt ne*PopRtnPtpRon ® PtpRo ne*PtpRonPtpRtn ® PtpRt ne*PtpRtnPopCRon ® PopCRo ne*PopCRonPopCRtn ® PopCRt ne*PopCRtnPtpCRon ® PtpCRo ne*PtpCRonPtpCRtn ® PtpCRt ne*PtpCRtnPoppn ® Popp ne*PoppnPtppn ® Ptpp ne*PtppnPoppCn ® PoppC ne*PoppCnPtppCn ® PtppC ne*PtppCnPoppRon ® PoppRo ne*PoppRonPoppRtn ® PoppRt ne*PoppRtnPtppRon ® PtppRo ne*PtppRonPtppRtn ® PtppRt ne*PtppRtnPoppCRon ® PoppCRo ne*PoppCRonPoppCRtn ® PoppCRt ne*PoppCRtnPtppCRon ® PtppCRo ne*PtppCRonPtppCRtn ® PtppCRt ne*PtppCRtnPop ® up*PopPopp ® up*PoppPtp ® up*PtpPtpp ® up*PtppPopC ® C up*PopCPoppC ® C up*PoppCPtpC ® C up*PtpCPtppC ® C up*PtppCPopn ® up*PopnPoppn ® up*PoppnPtpn ® up*PtpnPtppn ® up*PtppnPopCn ® Cn up*PopCnPoppCn ® Cn up*PoppCnPtpCn ® Cn up*PtpCnPtppCn ® Cn up*PtppCnRo ® uro*Ro
Rt ® urt*RtRon ® uro*RonRtn ® urt*Rtn® G (1.0-G)*bin*(Ron+Rtn+PopRon+PoppRon+
PtpRon+PtppRon+PopRtn+PoppRtn+PtpRtn+PtppRtn+PopCRon+PoppCRon+PtpCRon+PtppCRon+PopCRtn+PoppCRtn+PtpCRtn+PtppCRtn)
G ® unbin*G®MnPo trPo*(1.0-G)5
MnPo ® McPo tmc*MnPoMcPo ® umPo*McPo® Po tlpo*McPo® Pt tlpt*McPt® Ro tlro*McRo® Rt tlrt*McRtPo ® upu*PoPt ® upu*PtPoC ® C upu*PoCPtC ® C upu*PtC®MnRv trRv*(1.0-G)3
MnRv ® McRv tmc*MnRvMcRv ® umRv*McRv® Rv tlRv*McRvRv + Rv ® RvRv arv*Rv*RvRvRv ® Rv + Rv drv*RvRvRv ® Rvn nl*RvRvn ® Rv ne*RvnRvRv ® RvRvn nl*RvRvRvRvn ® RvRv ne*RvRvnRvn + Rvn ® RvRvn N*arv*Rvn*RvnRvRvn ® Rvn + Rvn drv*RvRvnRv ® uRv*RvRvn ® uRv*RvnRvRv ® 2.0*uRv*RvRvRvRvn ® 2.0*uRv*RvRvn®MnPt trPt*(1.0-G)5
MnPt ®McPt tmc*MnPtMcPt ® umPt*McPt®MnRo trRo*(1.0-G)*(1.0-GRv)3
MnRo ®McRo tmc*MnRoMcRo ® umR*McRo®MnRt trRt*(1.0-G)MnRt ®McRt tmc*MnRtMcRt ® umR*McRt® GRv binRv*(1.0-GRv)*RvRvnGRv ® unBinRv*GRv
Ptppn + Ron ® PtppRon ar*N*Ptppn*RonPtpn + Rtn ® PtpRtn ar*N*Ptpn*RtnPtppn + Rtn ® PtppRtn ar*N*Ptppn*RtnPopCn + Ron ® PopCRon ar*N*PopCn*RonPoppCn + Ron ® PoppCRon ar*N*PoppCn*RonPopCn + Rtn ® PopCRtn ar*N*PopCn*RtnPoppCn + Rtn ® PoppCRtn ar*N*PoppCn*RtnPtpCn + Ron ® PtpCRon ar*N*PtpCn*RonPtppCn + Ron ® PtppCRon ar*N*PtppCn*RonPtpCn + Rtn ® PtpCRtn ar*N*PtpCn*RtnPtppCn + Rtn ® PtppCRtn ar*N*PtppCn*RtnPopRo ® Pop + Ro dr*PopRoPoppRo ® Popp + Ro dr*PoppRoPopRt ® Pop + Rt dr*PopRtPoppRt ® Popp + Rt dr*PoppRtPtpRo ® Ptp + Ro dr*PtpRoPtppRo ® Ptpp + Ro dr*PtppRoPtpRt ® Ptp + Rt dr*PtpRtPtppRt ® Ptpp + Rt dr*PtppRtPopCRo ® PopC + Ro dr*PopCRoPoppCRo ® PoppC + Ro dr*PoppCRoPopCRt ® PopC + Rt dr*PopCRtPoppCRt ® PoppC + Rt dr*PoppCRtPtpCRo ® PtpC + Ro dr*PtpCRoPtppCRo ® PtppC + Ro dr*PtppCRoPtpCRt ® PtpC + Rt dr*PtpCRtPtppCRt ® PtppC + Rt dr*PtppCRtPopRon ® Popn + Ron dr*PopRonPoppRon ® Poppn + Ron dr*PoppRonPopRtn ® Popn + Rtn dr*PopRtnPoppRtn ® Poppn + Rtn dr*PoppRtnPtpRon ® Ptpn + Ron dr*PtpRonPtppRon ® Ptppn + Ron dr*PtppRonPtpRtn ® Ptpn + Rtn dr*PtpRtnPtppRtn ® Ptppn + Rtn dr*PtppRtnPopCRon ® PopCn + Ron dr*PopCRonPoppCRon ® PoppCn + Ron dr*PoppCRonPopCRtn ® PopCn + Rtn dr*PopCRtnPoppCRtn ® PoppCn + Rtn dr*PoppCRtnPtpCRon ® PtpCn + Ron dr*PtpCRonPtppCRon ® PtppCn + Ron dr*PtppCRonPtpCRtn ® PtpCn + Rtn dr*PtpCRtnPtppCRtn ® PtppCn + Rtn dr*PtppCRtnPop ® Popn nl*PopPtp ® Ptpn nl*PtpPopC ® PopCn nl*PopCPtpC ® PtpCn nl*PtpC

Stochastic Simulation of Mutations


1/n1/2 Behavior

What is Temperature Compensation?
Chemical reaction speed up as temperature increases• Circadian rhythms have an approximately constant
period regardless of external temperature• Not Temperature Independence
(Rhythms can entrain to temperature)
• Q10 = (Period at T)/(Period at T+10)(Q10 of circadian rhythms is typically 0.95-1.05,
Q10 is typically 2-4 in chemical reactions)
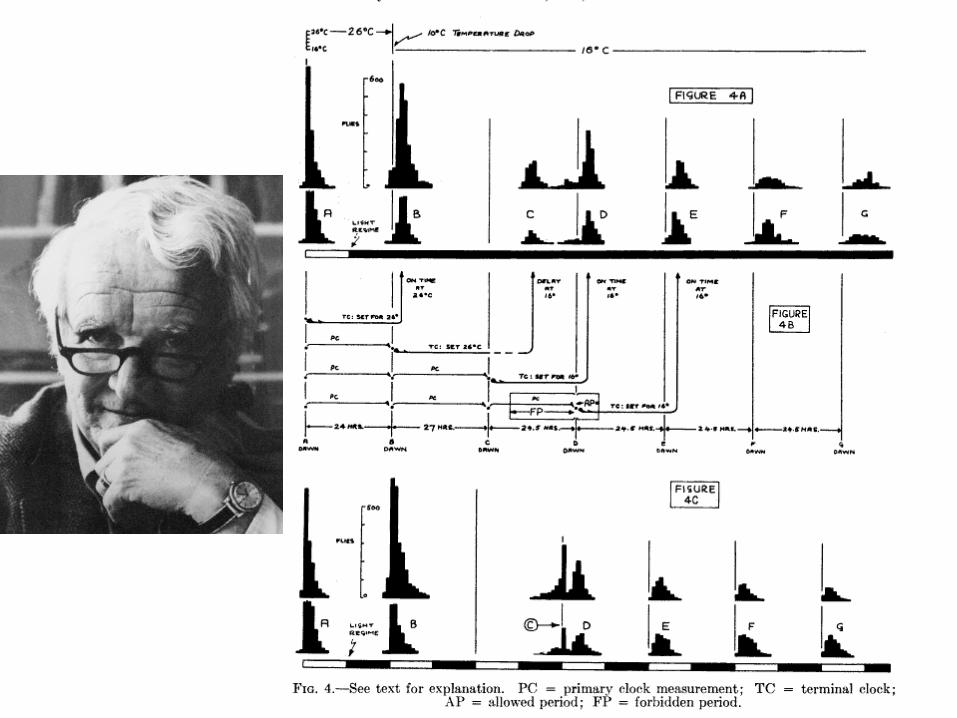
Pittendrigh

Hastings and Sweeney (1957)
• Some parts of the circadian clock increase period with increasing temperature (TCE)
• The rest decrease the period with increasing temperature
• Temperature compensation is achieved by balancing these processes.

Ruoff’s Theory• All biochemical oscillators have TCEs• Consider the period of the oscillator
• From kinetics and assumptionfor some I
• Approximate numerically (assume constant) and choose to achieve the balance
0=
∂∂
∂∂
= ∑ Tk
kP
dTdP i
i
0>∂∂
Tki 0<
∂∂
ikP
ikP
∂∂
Tki
∂∂
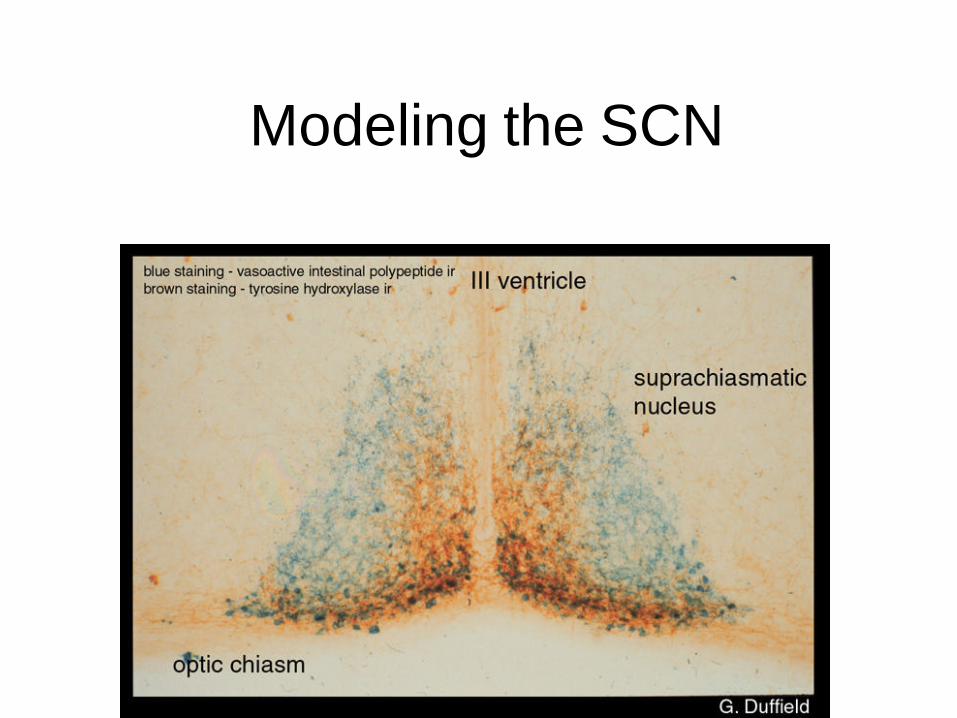
Modeling the SCN
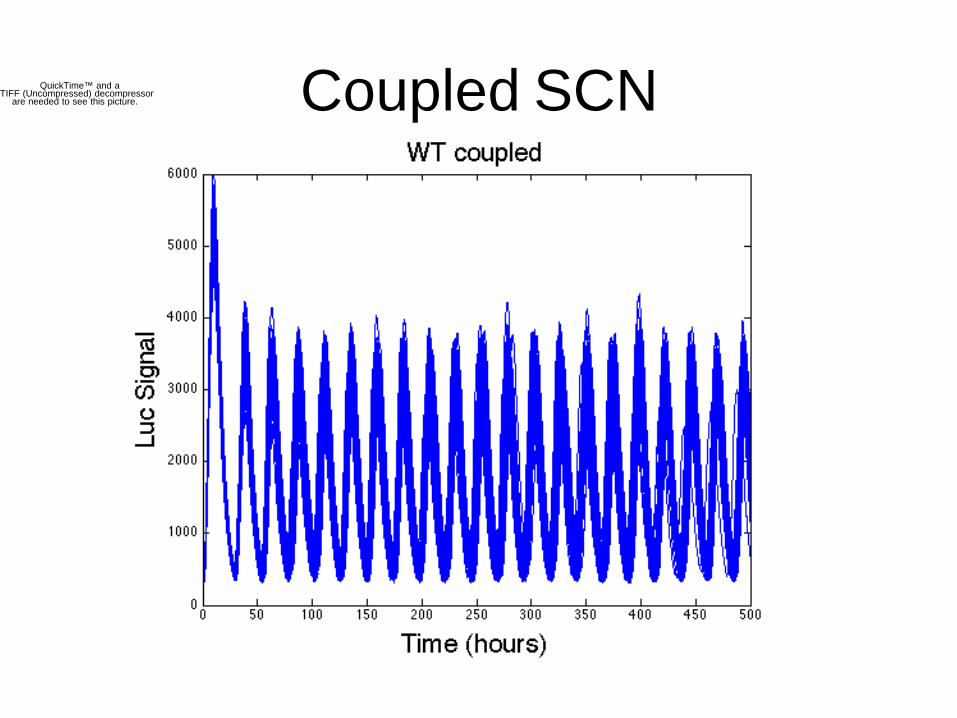
Coupled SCNQuickTime™ and aTIFF (Uncompressed) decompressor
are needed to see this picture.
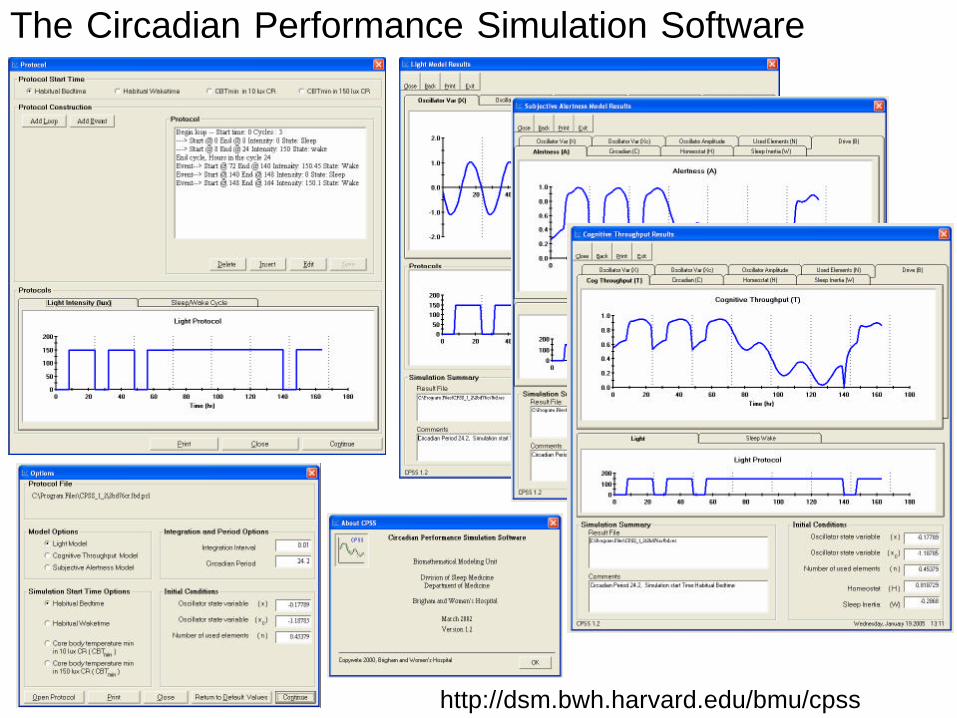
http://dsm.bwh.harvard.edu/bmu/cpss
The Circadian Performance Simulation Software





























