Embed Size (px)
Citation preview

ARCHIVES OF BIOCHEMISTRY AND BIOPHYSICS 162, 6-11 (1974)
Modification of Sarcoplasmic Reticulum Adenosine Triphosphatase
by Adenosine Triphosphate and Magnesium
DOUGLAS J. HORGAN
CAIRO Division of Food Research, Meat Research Laboratory, Cannon Hill, Queensland, Australia 4i70
Received August 2, 1973
Lineweaver-Burk plots of Cat+-activated adenosine triphosphatase from rabbit muscle sarcoplasmic reticulum have been determined for a wide range of substrate concentrations. The plots measured at constant Mg2f concentrations are normally nonlinear, but approach linearity either as the sarcoplasmic reticulum ages, or when small quantities of Triton-X100 are added. Titration with N-ethylmaleimide has the same effect on the activity of the ATPase measured either at high or low sub- strate concentrations. Lineweaver-Burk plots measured under conditions where the Mg2+ concentration is varied so as to be always equal to the ATP concentration are linear. These results have been interpreted as evidence that the adenosine triphos- phatase has a single active site which uses MgATP as its substrate and which can be modified by free Mgz+.
The sarcoplasmic reticulum of skeletal muscle contains two adenosine triphos- phatases (ATPases). These are the so-called “basal” ATPase which is a Mg2+-dependent ATPase active in the complete absence of calcium ions, and the “extra” ATPase which is responsible for the enhanced ac- tivity shown when micromolar concentra- tions of calcium are added to the “basal” assay system (1). Because of its involve- ment in calcium uptake by the sarcoplasmic reticulum (l-4), the “extra” ATPase has been studied in greater detail than the “basal” ATPase.
Previous kinetic studies (5, 6) of the extra ATPase have shown that the Lineweaver- Burk plot (7) of l/V versus l/[ATP] at con- stant Mg2+ concentration is nonlinear. K, values have been assigned to the high and low substrate concentration portions of these plots. On this basis, Inesi et al. (5) postulated the existence of two active sites with Km values of 10 P&I and >l mM. Ya- mamoto and Tonomura found Km values of 0.7 PM and 50 pM, but concluded that the activities shown at high and low ATP con-
centrations were due to only one active site (6). The highest substrate concentration used in these studies, however, was only 1 mM. This may be significant, because as Raison (8) has pointed out, early studies of the temperature dependence of sarcoplasmic reticulum extra ATPase, using 1 mM ATP as substrate, yielded linear Arrhenius plots (6, 9). Charnock (lo), however, using 5 mM ATP found a discontinuity at 10°C.
In the present study the reaction rate was measured over the range from 5 mM to 1 PM ATP. The effects of aging the sarco- plasmic reticulum preparation, adding Tri- ton-X100, varying the Mg2+ to ATP ratio, and titrating with N-ethylmaleimide (11) were studied. The results of these studies are discussed in terms of the existence of one or two active sites.
METHODS
Preparations of sarcoplasmic reticulum were made from the psoas muscle of New Zealand White rabbits essentially by the method of Martinosi, Donley. and Halpin (12). Dithiothreitol (13) was present at all stages of the preparation (1 maa) and in the final suspending medium (5 mM). It was
6 Copyright @ 1974 by Academic Press, Inc. All rights of reproduction in any form reserved.
SARCOPLASMIC RETICULUM ADENOSINE TRIPHOSPHATASE
0 10 20 30 40
[bii] 6 M-‘)
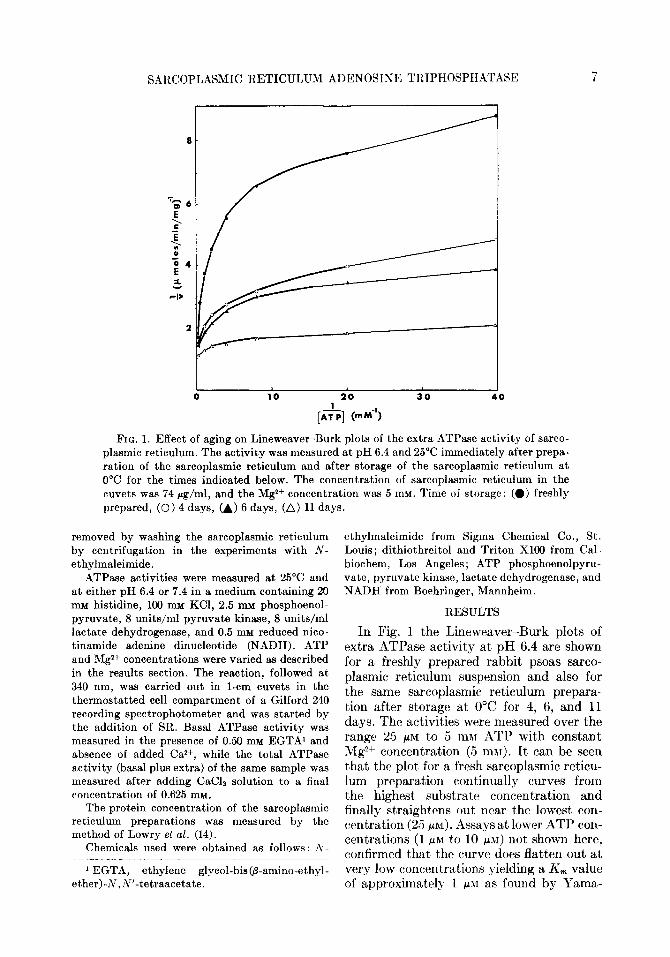
FIG. 1. Effect of aging on Lineweaver-Burk plots of the extra ATPase activity of sarco- plasmic reticulum. The activity was measured at pH 6.4 and 25°C immediately after prepa- ration of the sarcoplasmic reticulum and after storage of the sarcoplasmic reticulum at 0°C for the times indicated below. The concentration of sarcoplasmic reticulum in the cuvets was 74 rg/ml, and the Mg2+ concentration was 5 mM. Time of storage: (0) freshly prepared, (0) 4 days, (A) 6 days, (A) 11 days.
removed by washing the sarcoplasmic reticulum by centrifugation in the experiments with N- ethylmaleimide.
ATPase activities were measured at 25°C and at either pH 6.4 or 7.4 in a medium containing 20 mu histidine, 100 mu KCl, 2.5 mu phosphoenol- pyruvate, 8 units/ml pyruvate kinase, 8 units/ml lactate dehydrogenase, and 0.5 mM reduced nico- tinamide adenine dinucleotide (NADH). ATP and Mg2+ concentrations were varied as described in the results section. The reaction, followed at 340 nm, was carried out in l-cm cuvets in the thermostatted cell compartment of a Gilford 240 recording spectrophotometer and was started by the addition of SR. Basal ATPase activity was measured in the presence of 0.50 mu EGTA’ and absence of added Ca2+, while the total ATPase activity (basal plus extra) of the same sample was measured after adding CaClz solution to a final concentration of 0.625 mM.
The protein concentration of the sarcoplasmic reticulum preparations was measured by the method of Lowry et al. (14).
Chemicals used were obtained as follows: A’-
1 EGTA, ethylene glycol-bis@amino-ethyl- ether)-N,N’-tetraacetate.
ethylmaleimide from Sigma Chemical Co., St. Louis; dithiothreitol and Triton Xl00 from Cal- biochem, Los Angeles; ATP phosphoenolpyru- vate, pyruvate kinase, lactate dehydrogenase, and NADH from Boehringer, Mannheim.
RESULTS
In Fig. 1 the Lineweaver-Burk plots of extra ATPase activity at pH 6.4 are shown for a freshly prepared rabbit psoas sarco- plasmic reticulum suspension and also for the same sarcoplasmic reticulum prepara- tion after storage at 0°C for 4, 6, and 11 days. The activities were measured over the range 25 PM to 5 mM ATP with constant Mg2+ concentration (5 m&r). It can be seen that the plot for a fresh sarcoplasmic reticu- lum preparation continually curves from the highest substrate concentration and finally straightens out near the lowest con- centration (25 pnr). Assays at lower ATP con- centrations (1 PM to 10 PM) not shown here, confirmed that the curve does flatten out at very low concentrations yielding a K, value of approximately 1 ~JZ as found by Yama-
DOUGLAS J. HORGAN
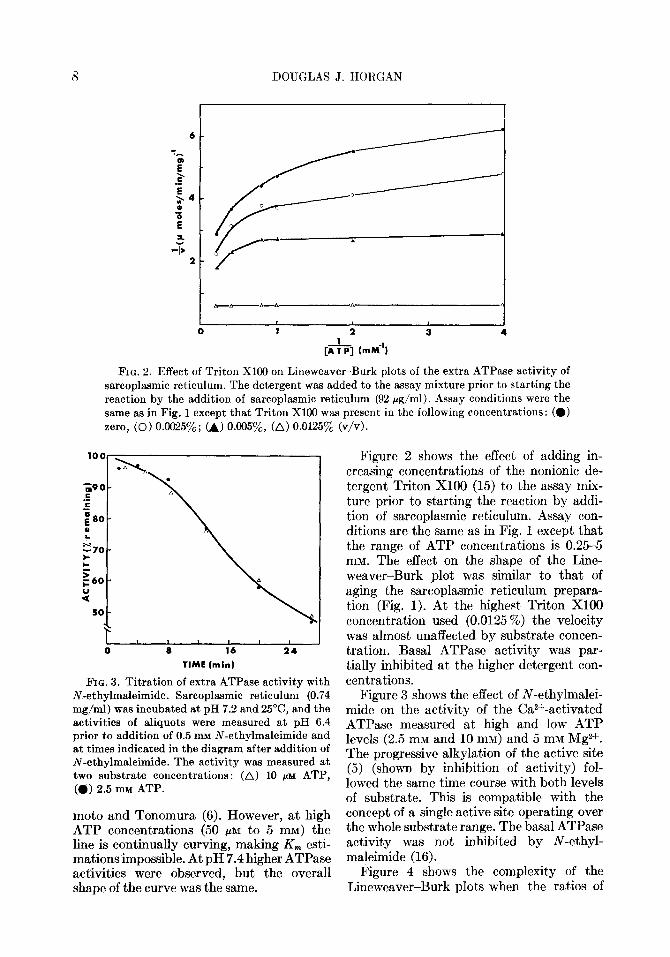
FIG. 2. Effect of Triton Xl00 on Lineweaver-Burk plots of the extra ATPase activity of sarcoplasmic reticulum. The detergent was added to the assay mixture prior to starting the reaction by the addition of sarcoplasmic reticulum (92 rg/ml). Assay conditions were the same as in Fig. 1 except that Triton Xl00 was present in the following concentrations: (0) zero, (0) 0.0025%; (A) 0.00597,, (A) 0.0125% (v/v).
-,oo- .c c ;ao- u L
;70-
I- 5 =60-
Y
50 -
% L , I I I I 0 8 16 24
TIME (min)
FIG. 3. Titration of extra ATPase activity with N-ethylmaleimide. Sarcoplasmic reticulum (0.74 mg/ml) was incubated at pH 7.2 and 25”C, and the activities of aliquots were measured at pH 6.4 prior to addition of 0.5 mu N-ethylmaleimide and at ,times indicated in the diagram after addition of N-ethylmaleimide. The activity was measured at two substrate concentrations: (A) 10 PM ATP, (a) 2.5 mu ATP.
moto and Tonomura (6). However, at high ATP concentrations (50 PM to 5 mM) the line is continually curving, making K,,, esti- mations impossible. At pH 7.4 higher ATPase activities were observed, but the overall shape of the curve was the same.
Figure 2 shows the effect of adding in- creasing concentrations of the nonionic de- tergent Triton Xl00 (15) to the assay mix- ture prior to starting the reaction by addi- t’ion of sarcoplasmic reticulum. Assay con- ditions are the same as in Fig. 1 except that the range of ATP concentrations is 0.25-5 mM. The effect on the shape of the Line- weaver-Burk plot was similar to that of aging the sarcoplasmic reticulum prepara- tion (Fig. 1). At the highest Triton Xl00 concentration used (0.0125 %) the velocity was almost unaffected by substrate concen- tration. Basal ATPase activity was par- tially inhibited at. the higher detergent con- centrations.
Figure 3 shows the effect of N-ethylmalei- mide on the activity of the Ca2+-activated ATPase measured at high and low ATP levels (2.5 mM and 10 mM) and 5 mM Mg2+. The progressive alkylation of the active site (5) (shown by inhibition of activity) fol- lowed the same time course with both levels of substrate. This is compatible with the concept of a single active site operating over the whole substrate range. The basal ATPase activity was not, inhibited by N-ethyl- maleimide (16).
Figure 4 shows the complexity of the Lineweaver-Burk plots when the ratios of
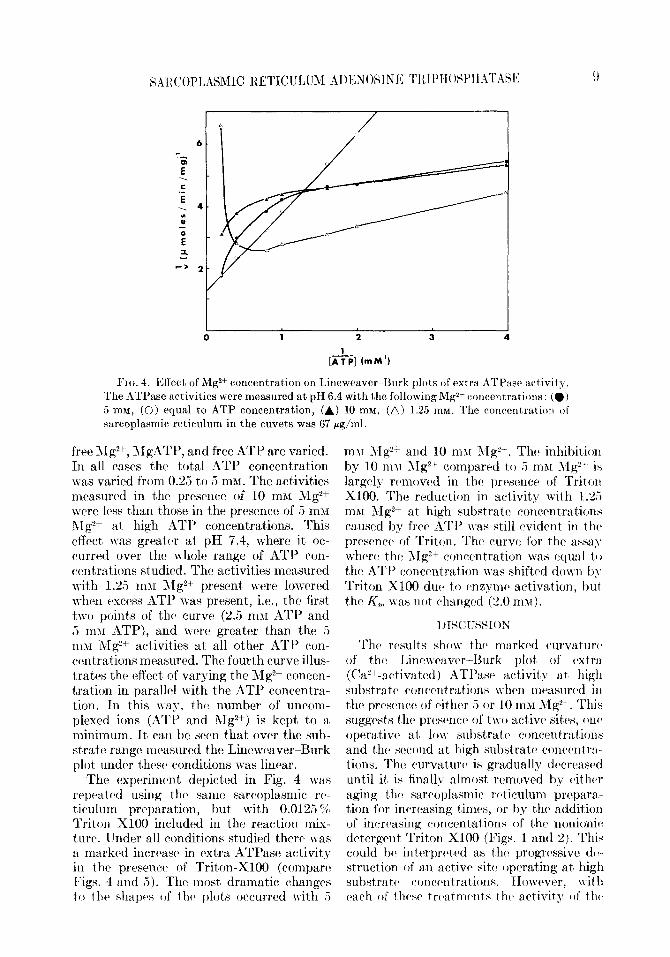
- [A:P] (mm’) FIG. 4. Effect of Mg2+ concentration on Lineweaver- Burk plots of extra ATPase activity.
The ATPase activities were measured at pH 6.4 with the following Mg2+ concentrations: (0) 5 mM, (oj equal to ATP concentration, (A) 10 mM, (A) 1.25 mu. The concentration of sarcoplasmic reticulum in the cuvets was 67 rgjml.
free 11g2+, AIgATP, and free ATP are varied. In all cases the tot,al ATE’ concent’ration was varied from 0.25 t,o 5 m;LI. The activities measuwd in t,he presence of 10 mar hIg2+ were less than those in the presence of 5 mnr i\Ig”+ at high ATl’ concentrations. This effects was grratw at pH 7.4, where it’ oc- curred over the whole range of ATP con- centrations studied. The activiDies measured with 1.25 rnlr RZg’+~ present. were lowered n-hen excess ATP was pwsenD, i.e., the first two points of th(l curve (2.5 rnX1 ATI’ and 5 m$I ATP), and ww greater than the 5 11111 i\Ig?+ activities at all other ATI’ corl- wntrations measured. The fourth curve illus- trates t#he effect of varying the Mg2+ concen- tration in paraM with the ATP concent’ra- tion. 111 t,his way, th(b number of uncom- plexed ions (ATI’ and hIg2+) is kept to a minimum. It can he swn that ovw the sub- strate range mcwured thr: Linewaver-Burk plot under t#hcsc conditions was linear.
The cJxperimc>nt depicted in Fig. 4 was rrpcatcd using thcl same sarcoplasmic r(‘- ticulum preparation, but wit,h 0.0125 ‘2, Triton Xl00 included in the reaction mix- t uw. Under all conditions studied there was a marked incwasc: in rlxtra ATPase activit,y in the prrsenw of Triton-X100 (cornpaw F’igs. 4 and 5). The most dramatic changw to the> shapw of thcl plots occurred with .5
mu hlg”+ and 10 m>r 11g2+. The inhibition by 10 m\l 1Ig*+ compawd to 5 mhl YIg2’ is largely wmowd in th(l prrscnce of Triton X100. The reduction in act,ivit,y with 1.25 ml1 ;\lgz+ at high substratcl cwlccntjrations causrd by frw ATl’ \vas still widcnt, in thcb prtwnw of Triton. The curve for the Assam wh(w thrl ;\Ig;“+ concentration was quaI t I J the ATP conwntration was shifted do\vn b> Triton Xl00 dw to cwymcb activation, hut the K, \vas Ilot changed (2.0 mu).
The wsults sholv t h(b marked curvat ur(s of th(a Lint,\\-ntvt~r-E3urk plot of cbxtru (Ca*+-ac*tivatcd) ATl’asc activity at high substrattx concentrations I\-hen measurcad ilr t’ho prcwnw of cithcr 5 or 10 m&I lVg2+. This suggests t,hc prcwnw of two actiw sitw, on(’ operatiw at, low substratcl ~on~(~ritr:ttio118 and th(l swond at high suhstratc> conwnt.ra- tions. The curvatuw is gradually dwwawcl until it is finally almost wmoved by (lit ti(sr aging thcb sarcoplasmic reticulum prcpara- tion for inclraaing times, or by thck addition of incwasing conccntations of thcl nonionic: drtcrgcnt Triton Xl00 (lcigs. 1 and 2). This could 1~~ intwpwtcd as th(j progwssivc dc~- structiori of an actiw site! operating at] high substratcx c,onc~t,lltratiorls. Howwcr, with clarh of thaw twatnwnts th(x activity 01’ the,
10 DOUGLAS J. HORGAN
0 1 2 3 4
[iii, (m m-‘1
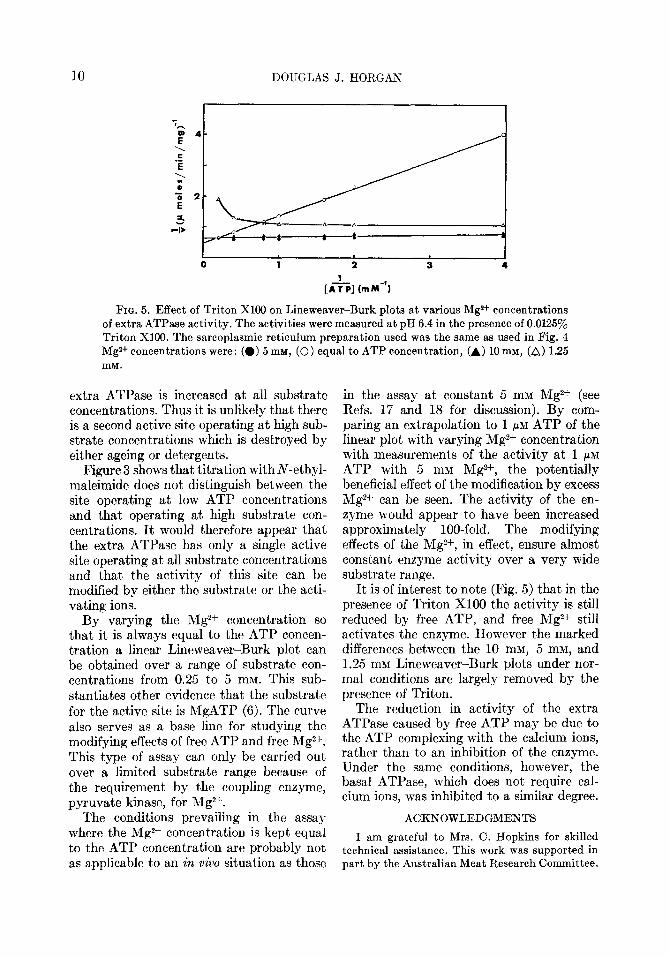
FIQ. 5. Effect of Triton Xl00 on Lineweaver-Burk plots at various Mg2+ concentrations of extra ATPase activity. The activities were measured at pH 6.4 in the presence of 0.01250/, Triton X100. The sarcoplasmic reticulum preparation used was the same as used in Fig. 4 Mg2+ concentrations were: (0) 5 rnr,r, (0) equal to ATP concentration, (A) 10 mM, (A) 1.25 NIM.
extra ATPase is increased at all substrate concentrations. Thus it is unlikely that there is a second active site operating at high sub- strate concentrations which is destroyed by either ageing or detergents.
Figure 3 shows that titration with N-ethyl- maleimide does not distinguish between the site operating at low ATP concentrations and that operating at high substrate con- centrations. It would therefore appear that the extra ATPase has only a single active site operating at all substrate concentrations and that the activity of this site can be modified by either the substrate or the acti- vating ions.
By varying the Mg2+ concentration so that it is always equal to the ATP concen- tration a linear Lineweaver-Burk plot can be obtained over a range of substrate con- centrations from 0.25 to 5 mM. This sub- stantiates other evidence that the substrate for the active site is MgATP (6). The curve also serves as a base line for studying the modifying effects of free ATP and free Mg2+. This type of assay can only be carried out over a limited substrate range because of the requirement by the coupling enzyme, pyruvate kinase, for Mg2+.
The conditions prevailing in the assay where the Mg2+ concentration is kept equal to the ATP concentration are probably not as applicable to an in V&J situation as those
in the assay at constant 5 mM Mg2+ (see Refs. 17 and 18 for discussion). By com- paring an extrapolation to 1 PM ATP of the linear plot with varying Mg2+ concentration with measurement’s of the activity at 1 PM ATP with 5 m&r Mg2+, the potentially beneficial effect of the modification by excess Mg2+ can be seen. The activity of the en- zyme would appear to have been increased approximately loo-fold. The modifying effects of the Mg2+, in effect, ensure almost constant enzyme activity over a very wide substrate range.
It is of interest to note (Fig. 5) that in the presence of Triton Xl00 the activity is still reduced by free ATP, and free Mg2+ still activates the enzyme. However the marked differences between the 10 mM, 5 mM, and 1.25 rnM Lineweaver-Burk plots under nor- mal conditions are largely removed by the presence of Triton.
The reduction in activity of the extra ATPase caused by free ATP may be due to the ATP complexing with the calcium ions, rather than to an inhibition of the enzyme. Under the same conditions, however, the basal ATPase, which does not require cal- cium ions, was inhibited to a similar degree.
ACKNOWLEDGMENTS
I am grateful to Mrs. C. Hopkins for skilled technical assistance. This work was supported in part by the Australian Meat Research Committee.

SARCOPLASMIC RETICULUM ADENOSINE TRIPHOSPHATASE 11
REFERENCES 1. HASSELBACH, W., AND MAHINOSE, M. (1963)
Biochem. Z. 339, 94. 2. EBASHI, S. (1961) J. Biochem. 60,236. 3. HASSELBACH, W., AXD ~'V~AKINOSE, M. (1961)
Biochem. Z. 333, 518. 4. HASSELBACH, W., AND MAKINOSE, M. (1962)
Riochem. Biophys. Res. Commun. 7, 132. 5. INESI, G.. GOODMAN, J.J., AND WATANABE,~,
(1967) J. Biol. Chem. 242, 4637. 6. YAMAMOTO, T., AND TONOMURA, Y. (1967)
J. Biochem. 62, 558. 7. LISEWEAVER, H., AND BURK, D. F. (1934)
J. Amer. Chem. Sot. 56, 658. 8. RAISOS, J. K. (1972) Bioenergetics, 4, 285. 9. IXlrSI, c;., AND WATANABE, s. (1967) Arch.
Biopkys. 121, 665. 10. CII.IRNOCK, J. P., AND FRANKEL, D. (1971)
Abstr. 2nd ht. Congr. Muscle Diseases, Pertb, Abstr. 45.
11. PANET, It., AND SEIJNGER, Z. (1970) EUT. ,I. Biochem. 14, 440.
12. MARTOXOSI, ;1., L)ONLEY, J., A~YD H~LPIX, IL A. (1968) J. Biol. Chew 243, 61.
13. VAN DER KI,OUT, W. (1969) Science 164, 12!)4. 14. LOIVRI'. 0. H., I:osI.:DROUGH, N. J., FARR. A.
L., .4ND KAh-DALL, 1:. J. (1951) J. Bif)/. ('h('~2. 193,265.





























