Embed Size (px)
Citation preview

Modulation of bcl-xL in Tumor Cells RegulatesAngiogenesis through CXCL8 Expression
Simona Giorgini,1 Daniela Trisciuoglio,1 Chiara Gabellini,1 Marianna Desideri,1
Laura Castellini,1 Cristina Colarossi,2 Uwe Zangemeister-Wittke,3
Gabriella Zupi,1 and Donatella Del Bufalo1
1Experimental Chemotherapy Laboratory, Regina Elena Cancer Institute, and 2Pathology Unit,Sant’ Andrea Hospital, University ‘‘La Sapienza,’’ Rome, Italy; and 3Division ofMedical Oncology, University Hospital Zurich, Zurich, Switzerland
AbstractIn this paper, we investigated whether bcl-xL can
be involved in the modulation of the angiogenic
phenotype of human tumor cells. Using the ADF human
glioblastoma and the M14 melanoma lines, and their
derivative bcl-xL–overexpressing clones, we showed
that the conditioned medium of bcl-xL transfectants
increased in vitro endothelial cell functions, such
as proliferation and morphogenesis, and in vivo
vessel formation in Matrigel plugs, compared with
the conditioned medium of control cells. Moreover,
the overexpression of bcl-xL induced an increased
expression of the proangiogenic interleukin-8 (CXCL8),
both at the protein and mRNA levels, and an enhanced
CXCL8 promoter activity. The role of CXCL8 on
bcl-xL–induced angiogenesis was validated using
CXCL8-neutralizing antibodies, whereas
down-regulation of bcl-xL through antisense
oligonucleotide or RNA interference strategies
confirmed the involvement of bcl-xL on CXCL8
expression. Transient overexpression of bcl-xL led to
extend this observation to other tumor cell lines with
different origin, such as colon and prostate carcinoma.
In conclusion, our results showed that CXCL8
modulation by bcl-xL regulates tumor angiogenesis,
and they point to elucidate an additional function of
bcl-xL protein. (Mol Cancer Res 2007;5(8):761–71)
IntroductionAngiogenesis is essential for the growth of both primary and
metastatic tumors (1), and the understanding of the genetic
alterations involved in angiogenic dysregulation could lead to
the development of antiangiogenic therapies. Angiogenesis is
activated by different factors, including hypoxia, oncogenes,
and antioncogenes (2). We have previously shown that bcl-2
overexpression in several human tumor histotypes synergizes
with hypoxia to increase angiogenesis through vascular
endothelial growth factor (VEGF) expression and hypoxia
inducible factor-1 transcriptional activity (3-7). Moreover,
treatment of melanoma cells with bcl-2/bcl-xL antisense
oligonucleotides reduces angiogenic activity (8). The ability
of bcl-2 to induce angiogenesis has also been shown in prostate
carcinoma and microvascular endothelial cells (9, 10).
The present study aims to explore the ability of bcl-xL, an
antiapoptotic member of the bcl-2 family, to modulate tumor
angiogenesis. bcl-xL is a prosurvival multidomain protein that,
in addition to promoting cell survival (11), has been implicated
in the regulation of cell cycle (12), in the modulation of cell
differentiation (13), and in the resolution of inflammation (14).
A pivotal role for bcl-xL in breast cancer metastatization (15)
and in anaplastic lymphoma kinase–mediated oncogenicity
(16) has also been shown. In the present study, we analyzed the
functional effect of the modulation of bcl-xL in the angiogenic
process of glioblastoma and melanoma.
In this regard, glioblastoma is a particular, suitable model for
our study in which vascular changes accompany the advance-
ment of this neoplasia (17), and the expression of bcl-xL is
elevated when compared with normal glial or astrocytic cells
(18, 19). Recently, the induction of bcl-xL in human
glioblastoma cells has been found to confer infiltrative growth
in vivo and reduced survival of tumor-bearing mice (20).
Moreover, the relevance of bcl-xL in the resistance of
glioblastoma to therapy has been widely shown (21, 22).
Furthermore, the role of bcl-xL as an antiapoptotic
molecule has been well established also in various melanoma
models (23, 24), and the general consensus in the literature is
that the bcl-xL protein level seems to increase with melanoma
progression (24-26).
In this paper, we show that bcl-xL modulation regulates
CXCL8 expression in several tumor cell lines with different
histotypes and angiogenesis of melanoma and glioblastoma.
Resultsbcl-xL Overexpression in the ADF Glioblastoma and M14Melanoma Cell Lines Enhances In vivo Angiogenesis
The human ADF glioblastoma and M14 melanoma cell lines
were stably transfected with a bcl-xL expression vector or with
the vector carrying the neomycin-resistant gene (control clone).
About 5-fold increase of bcl-xL protein expression was
Received 2/15/07; revised 4/18/07; accepted 5/9/07.Grant support: Ministero della Salute (D. Del Bufalo) and Italian Association forCancer Research (D. Del Bufalo). Dr. D. Trisciuoglio and Dr. C. Gabellini arerecipients of a fellowship from the Italian Foundation for Cancer Research(FIRC).The costs of publication of this article were defrayed in part by the payment ofpage charges. This article must therefore be hereby marked advertisement inaccordance with 18 U.S.C. Section 1734 solely to indicate this fact.Requests for reprints: Donatella Del Bufalo, Experimental ChemotherapyLaboratory, Regina Elena Cancer Institute, Via delle Messi d’Oro 156, 00158Rome, Italy. Phone: 39-6-52662537; Fax: 39-6-52662592. E-mail: [email protected] D 2007 American Association for Cancer Research.doi:10.1158/1541-7786.MCR-07-0088
Mol Cancer Res 2007;5(8). August 2007 761
Research. on November 18, 2018. © 2007 American Association for Cancermcr.aacrjournals.org Downloaded from

observed in bcl-xL–overexpressing clones (AXL42, AXL74)
when compared with the parental glioblastoma cells (ADF) or
the control clone (AN8; Fig. 1A). As reported in Fig. 1A, an
undetectable level of bcl-xL protein was observed in M14
parental and control (Mneo) cells under our experimental
condition, whereas both bcl-xL transfectants (MXL90, MXL12)
expressed a consistent level of bcl-xL protein.
To examine whether bcl-xL plays a role in the angiogenesis
of glioblastoma and melanoma, in vivo vessel formation in
Matrigel was examined (Fig. 1B-E). Matrigel plugs containing
the conditioned medium (CM) of control or bcl-xL–over-
expressing clones were injected in mice, and the degree of
vascularization into Matrigel plugs was evaluated. As
evidenced by macroscopic analysis, CM of the glioblastoma
(AN8) and melanoma (Mneo, M14) control cells induced only a
slight angiogenic response compared with the negative control
(Fig. 1B and D). In contrast, Matrigel plugs containing CM of
bcl-xL transfectants from glioblastoma (AXL42, AXL74;
Fig. 1B) or melanoma (MXL90, MXL12; Fig. 1D) showed a
higher angiogenic response, as well as the co-option of
surrounding vessels into the plugs, when compared with the
response induced by CM from the control clones. Quantifica-
tion of the hemoglobin content in the Matrigel plugs evidenced
an induction of about 4- to 5-fold for glioblastoma (AXL42,
P = 0.039; AXL74, P = 0.046; Fig. 1C) and of about 7-fold for
melanoma (MXL12, P = 0.027; MXL90, P = 0.042; Fig. 1E)
at the level of hemoglobin in the Matrigel plugs containing
CM from bcl-xL transfectants than the ones containing CM of
control cells (Fig. 1C and E). Similar results were obtained
when in vivo Matrigel assay was done using cell suspensions
instead of CM (data not shown). Histologic examination of
vessels into Matrigel plugs confirmed the results of quantitative
analysis of hemoglobin (Fig. 2). Matrigel plugs containing CM
from bcl-xL–overexpressing cells revealed an increased
capillary vessel formation, as compared with Matrigel contain-
ing CM from parental cells: the number of vessels counted was
about 2.7 for glioblastoma parental cells and increases to 14.6
in bcl-xL transfectants (P < 0.001); similarly, the number of
vessels increases from 3.2 in melanoma parental cells to 21
after bcl-xL overexpression (P < 0.001).
bcl-xL Overexpression in the ADF Glioblastoma and M14Melanoma Cell Lines Enhances CXCL8 Protein andmRNA Expression
To determine whether the enhanced neovascularization
induced by bcl-xL overexpression was influenced by the
FIGURE 1. bcl-xL overex-pression in ADF glioblastomaand M14 melanoma cel lsincreases in vivo angiogenesis.A. Western blot analysis of bcl-xL protein in the ADF glioblasto-ma line and its derivative control(AN8 ) and bcl-xL–overexpress-ing (AXL42, AXL74) clones andin the M14 melanoma line and itsderivative control (Mneo ) andbcl-xL – overexpressing clones(MXL12, MXL90 ). Western blotanalysis representative of twoindependent experiments withsimilar results are shown.B–E. In vivo vessel formationassessed after injection of micewith Matrigel containing CM fromthe glioblastoma (B and C) andfrom the melanoma (D and E)lines. B and D. Pictures of threerepresentative Matrigel plugs outof eight done with superimpos-able results. In the negativecontrol (NEG ), the CM isreplaced with SFM. C and E.Measurement of hemoglobincontent in each Matrigel plugexpressed as absorbance (OD )per gram of Matrigel plug.P values are reported in the text.
Giorgini et al.
Mol Cancer Res 2007;5(8). August 2007
762
Research. on November 18, 2018. © 2007 American Association for Cancermcr.aacrjournals.org Downloaded from

synthesis of endogenous mediators of angiogenesis, we
evaluated the expression of several angiogenic factors through
a human angiogenesis protein array (Fig. 3). A schematic
representation of the proangiogenic factors that can be detected
by the use of the array was reported in Fig. 3C. An increased
secretion of CXCL8 was observed in both glioblastoma (about
1.5-fold induction; Fig. 3A) and melanoma (from 1.5- to 3-fold
induction; Fig. 3B) lines after bcl-xL overexpression. To
confirm the results obtained with the protein array, the level
of CXCL8 protein in the CM of bcl-xL transfectants and control
cells was measured (Figs. 4 and 5). A significant increase in the
amounts of secreted CXCL8 was observed when bcl-xL–
overexpressing glioblastoma cells were compared with control
cells: about 3-fold increase in CXCL8 protein secretion was
evident after both 24 h (AXL42, P = 0.031; AXL74, P = 0.029)
and 48 h (AXL42 and AXL74, P = 0.009) of cell culture
(Fig. 4A). Figure 4B shows Northern blot analysis of CXCL8
mRNA expression in control (AN8) cells and AXL42 bcl-xL
transfectant: the AN8 control cells expressed a slightly
detectable level of CXCL8 transcript, whereas CXCL8 mRNA
was clearly evident in the bcl-xL transfectant exposed to the
same experimental condition. The effect of bcl-xL on the
expression of other chemokines and angiogenic factors that
are involved in the growth of glioblastoma was also evaluated
(Fig. 4C). The expression of transforming growth factor-h1(TGF-h1) and VEGF proteins that were not detectable either in
parental and bcl-xL–overexpressing clones using the protein
array was evaluated by ELISA assay. As reported in Fig. 4C, no
difference in the levels of both proteins was observed when
comparing CM from bcl-xL–overexpressing cells to control
clones. We also analyzed by ELISA the secretion of interleukin-
1h (IL-1h) protein, an angiogenic factor, and TGF-h2, a
molecule recently shown to be regulated by bcl-xL over-
expression in glioma cells (20). The antibodies for these two
proteins were not included in the protein membrane. As
reported in Fig. 4C, TGF-h2 protein was found to be equally
secreted in parental and bcl-xL–overexpressing cells, whereas
IL-1h was not detectable both in control and bcl-xL trans-
fectants at the experimental condition used. Because from the
protein array bcl-xL seems to modulate tissue inhibitor of
metalloproteinase 1 (TIMP1) and TIMP2 in glioblastoma cells,
we also tested the expression of these two proteins by Western
blot analysis: no differences in TIMP1 and TIMP2 expression
were observed between parental and bcl-xL–overexpressing
cells (data not shown).
Regarding the effect of bcl-xL overexpression on the
expression of angiogenic factors by melanoma cells, whereas
no significant differences in VEGF secretion (Fig. 5C) or
expression (Fig. 5D) were observed between control cells and
bcl-xL–overexpressing clones, a significant difference in
CXCL8 secretion was evidenced by ELISA in time course
experiments (Fig. 5A). This difference was already evident
12 h after cell culture: CXCL8 was detected at a low level
in parental cells (100 F 5.3 pg/106 cells), whereas about
FIGURE 2. bcl-xL overex-pression in ADF glioblastomaand M14 melanoma cellsincreases in vivo vessel for-mation in a Matrigel assay.Representative pictures of his-tologic analysis in Matrigelplug sections. Matrigel plugscontaining CM from bcl-xL–overexpressing cells (AXL42and AXL74 glioblastoma;MXL12 and MXL90 melano-ma) reveal a marked capillaryvessel proliferation, as com-pared with Matrigel plugs con-taining CM from control cells(AN8 glioblastoma and Mneomelanoma). In the negativeand positive controls, the CMis replaced with SFM or an-giogenic factors (8), respec-tively (Masson’s trichromestaining; original magnifica-tion, 10�).
Regulation of Angiogenesis by bcl-xL
Mol Cancer Res 2007;5(8). August 2007
763
Research. on November 18, 2018. © 2007 American Association for Cancermcr.aacrjournals.org Downloaded from

1,000 F 47.7 pg/106 cells of CXCL8 were secreted by bcl-xL–
overexpressing cells (MXL12, P = 0.041; MXL90, P = 0.036).
This difference was maintained until 72 h of cell culture
(MXL12, P = 0.035; MXL90, P = 0.029). Western blot analysis
of CM confirmed the higher CXCL8 protein secretion in bcl-xL
transfectants compared with the control cells (Fig. 5B). As
reported for glioblastoma, no modulation of TGF-h2 was
observed after forced expression of bcl-xL in melanoma cells
(Fig. 5C), whereas TGF-h1 and IL-1h were not detectable both
in control and bcl-xL transfectants.
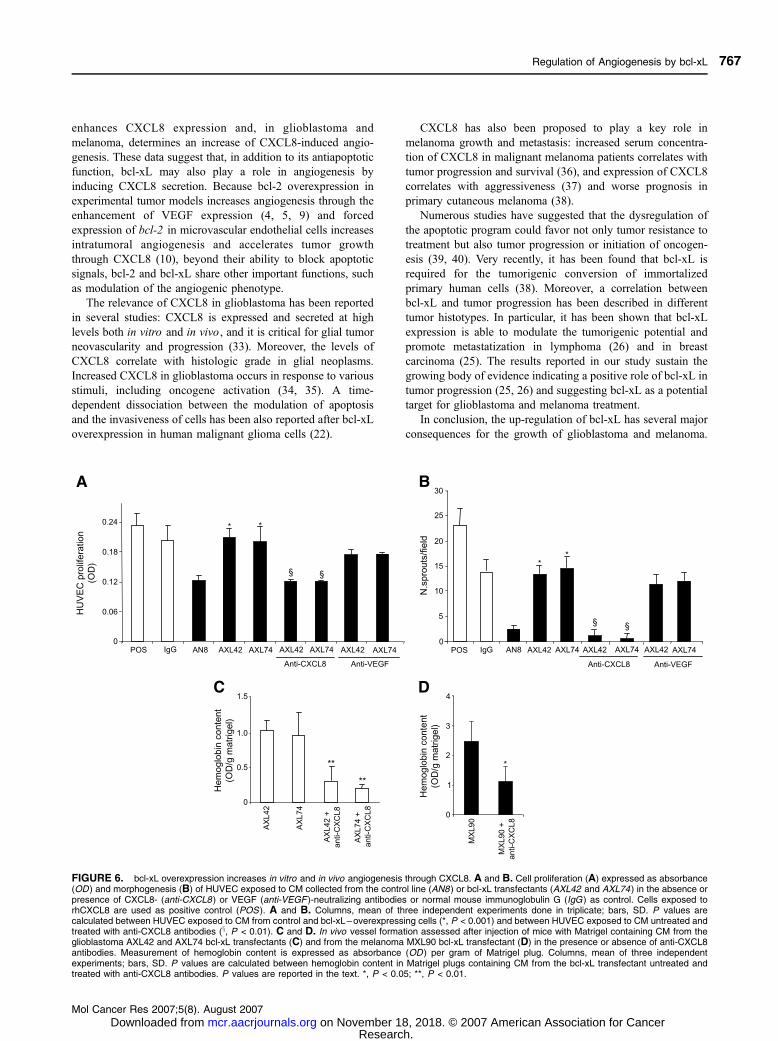
bcl-xL Overexpression Increases In vitro and In vivoAngiogenesis through CXCL8
Several evidences indicate that CXCL8 is an angiogenic
factor (27, 28). Thus, we analyzed whether the increase in
CXCL8 secretion induced by bcl-xL was responsible for the
enhancement of angiogenesis we previously observed in bcl-
xL–overexpressing cells (Fig. 1). In particular, to validate the
role of CXCL8 on bcl-xL–induced angiogenesis, we evaluated
in vitro endothelial cell functions, such as proliferation and
morphogenesis, in the presence of CM from different
glioblastoma lines containing CXCL8-neutralizing antibodies
or not. Figure 6A shows the proliferation of human umbilical
vascular endothelial cells (HUVEC) in the presence of CM
obtained after 48 h of cell culture. The CM from the bcl-xL–
overexpressing clones induced about a 2-fold increase of
HUVEC proliferation (AXL42 and AXL74, P = 0.001) when
compared with CM from the AN8 control clone. Addition of
CXCL8-neutralizing antibodies to the CM from bcl-xL trans-
fectants reduced HUVEC proliferation by about 40% (AXL42,
P = 0.005; AXL74, P = 0.001) to the level observed with CM
from control cells. VEGF-neutralizing antibodies did not induce
a significant inhibition of cell proliferation.
Next, we examined HUVEC capillary sprouting in the
presence of CM from the different glioblastoma lines. As
reported in Fig. 6B, HUVEC seeded on Matrigel in the
presence of CM from bcl-xL transfectants developed capillary-
like sprouts throughout the Matrigel surface; on the contrary,
endothelial cells exposed to CM from the control cells showed
only a reduced spread. In particular, 3 F 2 sprouts per field
were counted for HUVEC exposed to CM from the AN8
clone, whereas 13 F 5 and 15 F 3 sprouts per field were
counted for HUVEC exposed to CM from AXL42 (P =
0.007) and AXL74 (P = 0.005), respectively. Moreover,
whereas VEGF-neutralizing antibodies added to CM from
bcl-xL transfectants did not induce a significant decrease of
HUVEC morphogenesis, addition of CXCL8-neutralizing
antibodies reduced the number of spreads in HUVEC exposed
to CM from bcl-xL transfectants by about 80% (AXL42 and
AXL74, P = 0.001; Fig. 6B).
FIGURE 3. bcl-xL overex-pression in ADF glioblastomaand M14 melanoma cellsincreases CXCL8 expression.Image of membranes from aprotein array. Membranes areprobed with CM from the (A)glioblastoma control (AN8 )and bcl-xL – overexpressing(AXL42 and AXL74) clonesand (B) melanoma control(Mneo ) and bcl-xL–overex-p r e s s i n g (MXL12 andMXL90 ) clones. In the nega-tive control (NEG ), the CM isreplaced with an appropriatemock buffer according to ar-ray protocol. The intensity ofprotein signals for each mem-brane was compared with therelative positive signals bydensitometric analysis. C.Schematic representation ofproangiogenic factors thatcan be detected by the useof the membrane. TheCXCL8, TGF-1h, TIMP1,TIMP2, and VEGF proteinsignals (two spots) are indi-cated by red, blue, yellow,violet, and green rectangles,respectively, in each image.
Giorgini et al.
Mol Cancer Res 2007;5(8). August 2007
764
Research. on November 18, 2018. © 2007 American Association for Cancermcr.aacrjournals.org Downloaded from

In addition, the angiogenic response in Matrigel plugs in the
presence of CM from both melanoma and glioblastoma cells
overexpressing bcl-xL was markedly reduced by adding
specific neutralizing antibodies directed to CXCL8. The
hemoglobin content in the Matrigel plugs containing CM from
the bcl-xL transfectant was reduced by 70% (AXL42, P =
0.009; AXL74, P = 0.008) and 55% (P = 0.038) when using
CXCL8-neutralizing antibodies in glioblastoma (Fig. 6C) and
melanoma (Fig. 6D), respectively.
bcl-xL Overexpression Induces CXCL8 Secretion andPromoter Activity in Several Glioblastoma Cell Lines
We next evaluated the effect of bcl-xL overexpression on
several human glioblastoma cell lines. Figure 7A shows
Western blot analysis of bcl-xL protein expression in LI,
T98G, U373MG, CRS-A2, and U138 glioblastoma lines:
bcl-xL was expressed in all the lines analyzed. Moreover,
in those lines expressing high levels of bcl-xL, such as LI,
U373MG, and T98G, CXCL8 secretion was found to be
higher (respectively, 3,348 F 225, 2,005 F 1, and 1,763 F104 pg of CXCL8) than CRS-A2 and U138, expressing
undetectable levels of CXCL8 and low levels of bcl-xL. Then,
we evaluated whether transient overexpression of bcl-xL was
able to increase CXCL8 expression in the ADF and in other
glioblastoma lines (U138, CRS-A2, U373MG, and T98G). As
reported in Fig. 7B, an increase in CXCL8 expression of about
1.7-fold (U373MG, P = 0.042; T98G, P = 0.036), 2.7-fold
(ADF, P = 0.029; CRS-A2, P = 0.026), and 3.9-fold (U138,
P = 0.008) was observed after bcl-xL overexpression, thus
confirming the effect of bcl-xL on CXCL8 secretion in ADF
and extending this result to four other glioblastoma lines.
Because gene transcription is one major point of control at
which expression of CXCL8 is regulated (29), we next
determined whether the increase in CXCL8 secretion induced
by bcl-xL overexpression was due to a transcriptional up-
regulation of the corresponding gene (Fig. 5). To this end,
luciferase reporter construct containing wild-type CXCL8
proximal promoter was cotransfected with the control vector
PEQ-176 into control cells (AN8 and Mneo) and bcl-xL
transfectants from glioblastoma (AXL42 and AXL74,
Fig. 7C). As shown in Fig. 7C, basal activation of CXCL8
promoter region was up-regulated of about 2-fold in the
glioblastoma (AXL42, P = 0.025; AXL74, P = 0.031) bcl-
xL–overexpressing clones when compared with control
clones. U138 glioblastoma line was also used to confirm the
ability of bcl-xL to regulate CXCL8 promoter activity. As
shown in Fig. 7D, the transient expression of increasing
amounts of bcl-xL expression vector (from 0.25 to 0.75 Ag) inthis line induced a significant increase of CXCL8 promoter
activity (from about 1.7-fold to 4.5-fold, depending on the
amount of bcl-xL vector used; 0.25 Ag, P = 0.005; 0.5 and
0.75 Ag, P = 0.001) compared with the basal promoter activity.
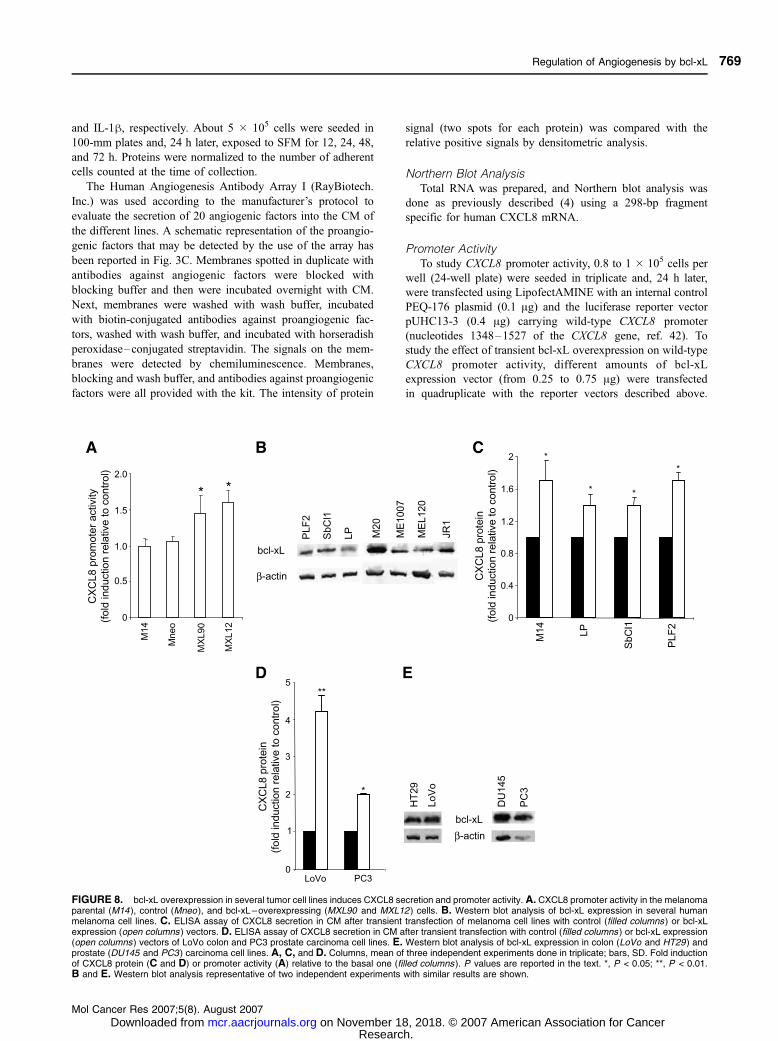
bcl-xL Overexpression Induces CXCL8 Secretion inSome Tumor Cell Lines of Different Origin
CXCL8 promoter activity after bcl-xL overexpression was
also investigated in melanoma cells. As shown in Fig. 8A, basal
activation of CXCL8 promoter region was up-regulated by
about 1.5-fold in melanoma (MXL90, P = 0.037; MXL12, P =
0.038) bcl-xL–overexpressing clones when compared with
control clones. bcl-xL protein was also found to be expressed in
several human melanoma lines, although different expression
levels were observed (Fig. 8B). Moreover, transient over-
expression of bcl-xL was also able to increase CXCL8
expression (from 1.4- to 1.7-fold) in the M14 (P = 0.042),
LP (P = 0.041), SbCl1 (P = 0.038), and PLF2 (P = 0.033)
melanoma lines (Fig. 8C). An increase in CXCL8 secretion
after transient overexpression of bcl-xL was also shown in
human colon (LoVo, 4.2-fold induction, P = 0.007) and prostate
(PC3, 2.0-fold induction, P = 0.036) carcinoma cells (Fig. 8D),
two tumor histotypes with a high expression level of bcl-xL
protein (30, 31), thus confirming the capability of bcl-xL to
modulate CXCL8 expression to tumor cells with different
origins. Despite a variability in bcl-xL protein expression from
line to line, Western blot analysis of bcl-xL protein expression
shows high levels of bcl-xL protein in two colon and two
prostate carcinoma lines (Fig. 8E).
Down-Regulation of bcl-xL Expression Reduces CXCL8Expression
To confirm the role of bcl-xL in the regulation of CXCL8
expression, bcl-xL protein expression was down-regulated
through antisense oligonucleotide (AS) or RNA interference
FIGURE 4. bcl-xL overexpression in ADF glioblastoma cells increasesCXCL8 expression. A. Expression of CXCL8 protein evaluated by ELISAin CM of the control cells (ADF, AN8 ) and bcl-xL–overexpressing clones(AXL42 and AXL74 ) after 24 (open columns ) and 48 h (filled columns ) ofcell culture. B. Northern blot analysis of CXCL8 gene expression in theAN8 and AXL42 clones. 28s and 18s rRNA are internal control for mRNAquality and loading. Northern blot analysis representative of twoindependent experiments with similar results is shown. C. Expression ofVEGF, TGF-h1, and TGF-h2 proteins evaluated by ELISA in CM fromADF, AN8, AXL42, and AXL74 cells after 48 h of cell culture. A and C.Columns, mean of three independent experiments done in triplicates; bars,SD. P values, calculated between control and bcl-xL–overexpressingcells, are reported in the text. *, P < 0.05; **, P < 0.01.
Regulation of Angiogenesis by bcl-xL
Mol Cancer Res 2007;5(8). August 2007
765
Research. on November 18, 2018. © 2007 American Association for Cancermcr.aacrjournals.org Downloaded from

strategies. Western blot analyses (Fig. 9A and C) showed that
treatment with AS specific for bcl-xL (32) decreased bcl-xL
expression both in the AXL74 bcl-xL transfectant from
glioblastoma (about 60% inhibition, Fig. 9A) and MXL12
bcl-xL transfectant from melanoma (about 40% inhibition,
Fig. 9C). Down-regulation of CXCL8 protein expression was
also induced by bcl-xL AS treatment: ELISA assays evidenced
the decrease of CXCL8 protein secretion after AS treatment
both in AXL74 (about 70% inhibition, P = 0.009, Fig. 9B) and
MXL12 (about 40% inhibition, P = 0.007, Fig. 9D) bcl-xL–
overexpressing clones. The control sequence (SC) did not affect
bcl-xL or CXCL8 expression (Fig. 9A-D). Similar results were
obtained using AXL42 glioblastoma and MXL90 melanoma
transfectants (data not shown). On the contrary, the decrease of
bcl-xL expression by AS exposure did not significantly modify
the level of secreted VEGF in both histotypes (data not shown).
We also evaluated whether bcl-xL down-regulation by bcl-xL
small interfering RNA (siRNA) in U373MG glioblastoma
cells (Fig. 9E) and MXL12 melanoma bcl-xL transfectant
(Fig. 9G) was able to affect CXCL8 expression: a reduction
of bcl-xL expression by bcl-xL siRNA in both histotypes
(Fig. 9E and G) was paralleled by a decrease of CXCL8
protein secretion of about 50% in glioblastoma (P = 0.008,
Fig. 9F) and of about 40% in melanoma (P = 0.016, Fig. 9H).
The control siRNA did not modify CXCL8 secretion (Fig. 9F
and H). These results confirm a relationship between bcl-xL
and CXCL8 expression.
DiscussionThe major finding of our study is that bcl-xL is able to
modulate CXCL8 expression in some human cancer cell lines.
In addition, bcl-xL–mediated-CXCL8 induction increases
angiogenesis in glioblastoma and melanoma cells. In particular,
stable bcl-xL overexpression in both tumor histotypes induced
CXCL8 protein secretion, mRNA expression, and promoter
activity. Moreover, CM from bcl-xL transfectants enhanced
in vitro proliferation and morphogenesis of endothelial cells
and in vivo vessel formation in a Matrigel assay. Enhancement
of CXCL8 expression induced by bcl-xL was dependent on
gene transcription: transient transfection experiments with wild-
type CXCL8 promoter reporter construct showed that bcl-xL
overexpression increased CXCL8 promoter activity in glioblas-
toma and melanoma lines. A decreased secretion of CXCL8
protein was observed after the down-regulation of bcl-xL
protein expression using bcl-xL –specific AS or siRNA, and the
use of CXCL8–specific neutralizing antibodies validated the
role of CXCL8 on bcl-xL–induced angiogenesis. Transient
transfection experiments confirmed the ability of bcl-xL to
increase CXCL8 protein expression not only in the glioblas-
toma and melanoma lines, but also in tumor cell lines of
different origin such as prostate and colon carcinoma, thus
indicating that the CXCL8 induction by bcl-xL is not a
phenomenon restricted to glioblastoma and melanoma, but it
could be extended to other kinds of tumors.
We excluded the possibility that bcl-xL could increase
angiogenesis in our models through the enhancement of other
important angiogenic factors or chemokines involved in the
angiogenic phenotype, such as VEGF, TGF-h1, IL-1h, TIMP1,
and TIMP2, because no modulation of these proteins was
observed after bcl-xL overexpression.
There are several novel aspects in our study. It is the first to
show that bcl-xL overexpression in several tumor histotypes
FIGURE 5. bcl-xL overex-pression in M14 melanoma cellsincreases CXCL8 protein expres-sion. CXCL8 and VEGF proteinexpression in M14 parental (o)and Mneo (.) control cells andMXL12 (5) and MXL90 (n)bcl-xL – overexpressing clones.ELISA evaluation in CM ofCXCL8 (A) after 12 to 72 h of cellculture and VEGF and TGF-h2(C) after 48 h of cell culture.A and C. Results represent themean F SD of three independentexperiments done in triplicates.P values are reported in the text.Western blot analysis of CXCL8in the CM (B) and VEGF in thecellular lysates (D). Red Ponceaustaining of filter and h-actin areused to check equal loading andtransfer of proteins. Westernblots representative of two inde-pendent experiments with similarresults are shown.
Giorgini et al.
Mol Cancer Res 2007;5(8). August 2007
766
Research. on November 18, 2018. © 2007 American Association for Cancermcr.aacrjournals.org Downloaded from
enhances CXCL8 expression and, in glioblastoma and
melanoma, determines an increase of CXCL8-induced angio-
genesis. These data suggest that, in addition to its antiapoptotic
function, bcl-xL may also play a role in angiogenesis by
inducing CXCL8 secretion. Because bcl-2 overexpression in
experimental tumor models increases angiogenesis through the
enhancement of VEGF expression (4, 5, 9) and forced
expression of bcl-2 in microvascular endothelial cells increases
intratumoral angiogenesis and accelerates tumor growth
through CXCL8 (10), beyond their ability to block apoptotic
signals, bcl-2 and bcl-xL share other important functions, such
as modulation of the angiogenic phenotype.
The relevance of CXCL8 in glioblastoma has been reported
in several studies: CXCL8 is expressed and secreted at high
levels both in vitro and in vivo , and it is critical for glial tumor
neovascularity and progression (33). Moreover, the levels of
CXCL8 correlate with histologic grade in glial neoplasms.
Increased CXCL8 in glioblastoma occurs in response to various
stimuli, including oncogene activation (34, 35). A time-
dependent dissociation between the modulation of apoptosis
and the invasiveness of cells has been also reported after bcl-xL
overexpression in human malignant glioma cells (22).
CXCL8 has also been proposed to play a key role in
melanoma growth and metastasis: increased serum concentra-
tion of CXCL8 in malignant melanoma patients correlates with
tumor progression and survival (36), and expression of CXCL8
correlates with aggressiveness (37) and worse prognosis in
primary cutaneous melanoma (38).
Numerous studies have suggested that the dysregulation of
the apoptotic program could favor not only tumor resistance to
treatment but also tumor progression or initiation of oncogen-
esis (39, 40). Very recently, it has been found that bcl-xL is
required for the tumorigenic conversion of immortalized
primary human cells (38). Moreover, a correlation between
bcl-xL and tumor progression has been described in different
tumor histotypes. In particular, it has been shown that bcl-xL
expression is able to modulate the tumorigenic potential and
promote metastatization in lymphoma (26) and in breast
carcinoma (25). The results reported in our study sustain the
growing body of evidence indicating a positive role of bcl-xL in
tumor progression (25, 26) and suggesting bcl-xL as a potential
target for glioblastoma and melanoma treatment.
In conclusion, the up-regulation of bcl-xL has several major
consequences for the growth of glioblastoma and melanoma.
FIGURE 6. bcl-xL overexpression increases in vitro and in vivo angiogenesis through CXCL8. A and B. Cell proliferation (A) expressed as absorbance(OD ) and morphogenesis (B) of HUVEC exposed to CM collected from the control line (AN8 ) or bcl-xL transfectants (AXL42 and AXL74) in the absence orpresence of CXCL8- (anti-CXCL8 ) or VEGF (anti-VEGF )-neutralizing antibodies or normal mouse immunoglobulin G (IgG ) as control. Cells exposed torhCXCL8 are used as positive control (POS ). A and B. Columns, mean of three independent experiments done in triplicate; bars, SD. P values arecalculated between HUVEC exposed to CM from control and bcl-xL–overexpressing cells (*, P < 0.001) and between HUVEC exposed to CM untreated andtreated with anti-CXCL8 antibodies (x, P < 0.01). C and D. In vivo vessel formation assessed after injection of mice with Matrigel containing CM from theglioblastoma AXL42 and AXL74 bcl-xL transfectants (C) and from the melanoma MXL90 bcl-xL transfectant (D) in the presence or absence of anti-CXCL8antibodies. Measurement of hemoglobin content is expressed as absorbance (OD ) per gram of Matrigel plug. Columns, mean of three independentexperiments; bars, SD. P values are calculated between hemoglobin content in Matrigel plugs containing CM from the bcl-xL transfectant untreated andtreated with anti-CXCL8 antibodies. P values are reported in the text. *, P < 0.05; **, P < 0.01.
Regulation of Angiogenesis by bcl-xL
Mol Cancer Res 2007;5(8). August 2007
767
Research. on November 18, 2018. © 2007 American Association for Cancermcr.aacrjournals.org Downloaded from

As shown by several papers, it inhibits apoptosis induced by
several stimuli, improving the ability of tumor cells to escape
from therapy (12, 13). Moreover, as reported in this study, bcl-
xL overexpression increases the expression of CXCL8, an
endogenous inducer of in vitro and in vivo angiogenesis (28).
These functions have a direct and significant influence on the
glioblastoma and melanoma progression and angiogenesis.
Thus, the blockade of bcl-xL protein expression by AS
treatment or chemical selective inhibitors could be used to
inhibit the angiogenic potential and improve the treatment of
these neoplasia.
Materials and MethodsCell Lines, Transfections, Oligonucleotide, and siRNATreatment
Human glioblastoma (ADF, CRS-A2, LI, T98G, U373MG,
and U138), melanoma (M14, PLF2, SbCl1, LP, M20, ME1007,
MEL120, and JR1), prostate carcinoma (PC3 and DU145), colon
carcinoma (LoVo and HT29), umbilical vein endothelial
(HUVEC) cell lines, two bcl-xL–overexpressing clones
(AXL42 and AXL74), and a control clone (AN8) derived from
the ADF line after stable transfection, two bcl-xL–over-
expressing clones (MXL90 and MXL12) and a control clone
(Mneo) derived from the M14 line after stable transfection were
used.
Stable and transient transfections with bcl-xL expression
vector (pcDNA3-bcl-xL), bcl-xL antisense oligonucleotides
(AS, 750 nmol/L, ref. 41) or bcl-xL siRNA (100 nmol/L, 5¶-GGAGAUGCAGGUAUUGGUGUU-3¶, Dharmacon Re-
search) were done using LipofectAMINE or lipofectin
(Invitrogen). After transient transfections, cells were exposed
to serum-free medium (SFM) for 24 h, and then adherent cells
and CM were differentially processed according to the analyses
to be done. Control experiments were done using scrambled
oligonucleotides (SC, 100 nmol/L, 5¶-CACGTCACGCGCG-
CACTATT-3¶, Dharmacon Research; ref. 41), siRNA against
unrelated mRNA, or empty expression vectors.
Western Blot AnalysisProteins from cellular lysates or CM were fractionated by
SDS-PAGE, transferred to a nitrocellulose filter, and subjected
to immunoblot assays. Antibodies against bcl-xL and TIMP2
(Santa Cruz Biotechnology), VEGF and HSP 72/73 (Calbio-
chem, Merck KGaA), CXCL8 (Biodesign International),
TIMP1 (Chemicon International), and h-actin (Sigma) were
used. Red Ponceau (Sigma) staining of filter and h-actin or HSP
72/73 protein amount were used to check equal loading and
transfer of proteins.
ELISA and Human Angiogenesis Antibody ArrayELISA assay (R&D Systems) was used to evaluate the level
of secreted CXCL8, VEGF, IL-1h, TGF-h1, and TGF-h2 in
CM. The sensitivity of the assay was 31.2 pg/mL for CXCL8
and TGF-h1, 15.6, 7.0, and 3.9 pg/mL for VEGF, TGF-h2,
FIGURE 7. bcl-xL overexpressionin several glioblastoma cell lines indu-ces CXCL8 secretion and promoteractivity. A. Western blot analysis ofbcl-xL expression in five human glio-blastoma cell lines. Western blot anal-ysis representative of two independentexperiments with similar results isshown. B. ELISA assay of CXCL8secretion in CM after transient trans-fection of glioblastoma cell lines withcontrol (filled columns ) or bcl-xL ex-pression (open columns ) vectors. C.CXCL8 promoter activity in the glio-blastoma parental (ADF ), control(AN8 ), and bcl-xL – overexpressing(AXL42 and AXL74 ) cells. D. CXCL8promoter activity in U138 glioblastomaline after transient transfection of in-creasing amounts (from 0.25 to 0.75 Ag)of bcl-xL expression vector (opencolumns ). B–D. Columns, mean ofthree independent experiments done intriplicate; bars, SD. Fold induction ofCXCL8 protein (B) or promoter activity(C and D) relative to the basal one(filled columns ). P values are reportedin the text. *, P < 0.05; **, P < 0.01.
Giorgini et al.
Mol Cancer Res 2007;5(8). August 2007
768
Research. on November 18, 2018. © 2007 American Association for Cancermcr.aacrjournals.org Downloaded from
and IL-1h, respectively. About 5 � 105 cells were seeded in
100-mm plates and, 24 h later, exposed to SFM for 12, 24, 48,
and 72 h. Proteins were normalized to the number of adherent
cells counted at the time of collection.
The Human Angiogenesis Antibody Array I (RayBiotech.
Inc.) was used according to the manufacturer’s protocol to
evaluate the secretion of 20 angiogenic factors into the CM of
the different lines. A schematic representation of the proangio-
genic factors that may be detected by the use of the array has
been reported in Fig. 3C. Membranes spotted in duplicate with
antibodies against angiogenic factors were blocked with
blocking buffer and then were incubated overnight with CM.
Next, membranes were washed with wash buffer, incubated
with biotin-conjugated antibodies against proangiogenic fac-
tors, washed with wash buffer, and incubated with horseradish
peroxidase–conjugated streptavidin. The signals on the mem-
branes were detected by chemiluminescence. Membranes,
blocking and wash buffer, and antibodies against proangiogenic
factors were all provided with the kit. The intensity of protein
signal (two spots for each protein) was compared with the
relative positive signals by densitometric analysis.
Northern Blot AnalysisTotal RNA was prepared, and Northern blot analysis was
done as previously described (4) using a 298-bp fragment
specific for human CXCL8 mRNA.
Promoter ActivityTo study CXCL8 promoter activity, 0.8 to 1 � 105 cells per
well (24-well plate) were seeded in triplicate and, 24 h later,
were transfected using LipofectAMINE with an internal control
PEQ-176 plasmid (0.1 Ag) and the luciferase reporter vector
pUHC13-3 (0.4 Ag) carrying wild-type CXCL8 promoter
(nucleotides 1348–1527 of the CXCL8 gene, ref. 42). To
study the effect of transient bcl-xL overexpression on wild-type
CXCL8 promoter activity, different amounts of bcl-xL
expression vector (from 0.25 to 0.75 Ag) were transfected
in quadruplicate with the reporter vectors described above.
FIGURE 8. bcl-xL overexpression in several tumor cell lines induces CXCL8 secretion and promoter activity. A. CXCL8 promoter activity in the melanomaparental (M14 ), control (Mneo ), and bcl-xL–overexpressing (MXL90 and MXL12 ) cells. B. Western blot analysis of bcl-xL expression in several humanmelanoma cell lines. C. ELISA assay of CXCL8 secretion in CM after transient transfection of melanoma cell lines with control (filled columns ) or bcl-xLexpression (open columns ) vectors. D. ELISA assay of CXCL8 secretion in CM after transient transfection with control (filled columns ) or bcl-xL expression(open columns) vectors of LoVo colon and PC3 prostate carcinoma cell lines. E. Western blot analysis of bcl-xL expression in colon (LoVo and HT29) andprostate (DU145 and PC3 ) carcinoma cell lines. A, C, and D. Columns, mean of three independent experiments done in triplicate; bars, SD. Fold inductionof CXCL8 protein (C and D) or promoter activity (A) relative to the basal one (filled columns ). P values are reported in the text. *, P < 0.05; **, P < 0.01.B and E. Western blot analysis representative of two independent experiments with similar results are shown.
Regulation of Angiogenesis by bcl-xL
Mol Cancer Res 2007;5(8). August 2007
769
Research. on November 18, 2018. © 2007 American Association for Cancermcr.aacrjournals.org Downloaded from

The luciferase activity of each sample was normalized to
h-galactosidase activity to calculate relative luciferase activity.
Densitometric AnalysesDeveloped films were acquired using GS-700 Imaging
Densitometer (Bio-Rad) and processed with Corel Photo Paint
7.0 to adjust image brightness and contrast. Densitometric
evaluation was done using Molecular Analyst Software (Bio-
Rad) and normalized with relative controls depending on the
analysis done.
In vitro Proliferation and Morphogenesis of HUVECExponentially growing HUVEC were seeded for prolifera-
tion (7 � 103 cells per 96-well microtiter plate) or morpho-
genesis (1.5 � 104 cells per 96-well microtiter plate containing
polymerized Matrigel) and incubated with CM obtained from
control cells or from bcl-xL transfectants, alone or in the
presence of specific human CXCL8- or VEGF-neutralizing
antibodies (anti-CXCL8 and anti-VEGF, 0.2 Ag/mL, R&D
Systems) or normal mouse immunoglobulin G (Pierce Biotech-
nology, Inc.) as control. Cells incubated in the presence of
rhCXCL8 (10 ng/mL, R&D Systems) were used as positive
control. Cell proliferation was evaluated after 48 h by a
colorimetric assay as previously described (5) and reported as
absorbance. In morphogenesis experiments, the number of
sprouts in six random microscopic fields was counted in
triplicate wells at 10� original magnification, using the inverted
phase contrast microscope Leitz Fluovert (Wetzlar).
In vivo Matrigel AssayIn vivo Matrigel assay and quantification of the hemoglobin
content in the Matrigel plugs were done as previously reported
(4) using cell suspensions (5 � 106 viable cells) or CM (10�concentrated) in the absence or presence of CXCL8-neutraliz-
ing antibodies (5 Ag/mL). Groups of eight mice were used for
each experimental point. All procedures involving animals and
their care were conducted in conformity with the institutional
guidelines, which are in compliance with national and
international laws (4). For histologic analysis, Matrigel plugs
were fixed with 10% formalin solution and paraffin-embedded.
Sections 5 Am thick were deparaffinized and stained with
Masson’s trichrome according to the manufacturer’s protocol
FIGURE 9. Down-regulation of bcl-xL expression in tumor cells reduces CXCL8 expression. A, C, E, and G. Western blot analyses of bcl-xL protein intotal extracts from AXL74 glioblastoma bcl-xL transfectant (A), U373MG glioblastoma cells (E), and MXL12 melanoma bcl-xL transfectant (C and G) aftertreatment with bcl-xL antisense (AS) or scrambled control (SC ) oligonucleotide (A and C) or after exposure to bcl-xL siRNA or control siRNA (E andG). HSP72/73 protein amounts are used to check equal loading and transfer of proteins. Representative Western blots out of two experiments done withsuperimposable results. B, D, F, and H. ELISA assays of CXCL8 protein in CM from AXL74 glioblastoma bcl-xL transfectant (B), U373MG glioblastomacells (F), and MXL12 melanoma bcl-xL transfectant (D and H) after treatment with bcl-xL AS or SC (B and D) or bcl-xL siRNA or control siRNA (F and H).Columns, mean of three independent experiments done in triplicate; bars, SD. P values are reported in the text. **, P < 0.01; *, P < 0.05.
Giorgini et al.
Mol Cancer Res 2007;5(8). August 2007
770
Research. on November 18, 2018. © 2007 American Association for Cancermcr.aacrjournals.org Downloaded from

(Bioptica Spa). In the negative and positive controls, the CM
was replaced with SFM or angiogenic factors, respectively.
Statistical AnalysisResults were expressed as the mean F SD from n
determinations. The significance of differences between the
means was determined with Student’s two-tailed t test, done
with PSP6 software. Values of P < 0.05 were accepted as
statistically significant.
AcknowledgmentsWe are grateful to Dr. Andrea Anichini (Istituto Nazionale Tumori, Milan, Italy)and Michael Kracht (Medical School Hannover, Institute of Pharmacology,Hannover, Germany) for providing us the ME1007 melanoma cell line andCXCL8 promoter reporter vector, respectively. We thank Dr. Adele Petricca forsecretarial assistance in the preparation of the manuscript.
References1. Folkman J, Watson K, Ingber D, Hanahan D. Induction of angiogenesis duringthe transition from hyperplasia to neoplasia. Nature 1989;339:58–61.
2. Mukhopadhyay D, Tsiokas L, Zhou, XM, Foster D, Brugge JS, Sukhatme VP.Hypoxic induction of human vascular endothelial growth factor expressionthrough c-Src activation. Nature 1995;375:577 –81.
3. Del Bufalo D, Biroccio A, Leonetti C, Zupi G. Bcl-2 overexpression enhancesthe metastatic potential of a human breast cancer line. FASEB J 1997;11:947– 53.
4. Biroccio A, Candiloro A, Mottolese M, et al. Bcl-2 overexpression andhypoxia synergistically act to modulate vascular endothelial growth factorexpression and in vivo angiogenesis in a breast carcinoma line. FASEB J 2000;14:652 –60.
5. Iervolino A, Trisciuoglio D, Ribatti D, et al. Bcl-2 overexpression in humanmelanoma cells increases angiogenesis through VEGF mRNA stabilization andHIF-1 –mediated transcriptional activity. FASEB J 2002;16:1453–5.
6. Trisciuoglio D, Desideri M, Ciuffreda L, et al. Bcl-2 overexpression inmelanoma cells increases tumor progression-associated properties and in vivo
tumor growth. J Cell Physiol 2005;205:414 –21.
7. Trisciuoglio D, Iervolino A, Zupi G, Del Bufalo D. Involvement of PI3K andMAPK signaling in bcl-2 – induced vascular endothelial growth factor expressionin melanoma cells. Mol Biol Cell 2005;16:4153–62.
8. Del Bufalo D, Trisciuoglio D, Scarsella M, Zangemeister-Wittke U, Zupi G.Treatment of melanoma cells with a bcl-2/bcl-xL antisense oligonucleotideinduces antiangiogenic activity. Oncogene 2003;22:8441–7.
9. Fernandez A, Udagawa T, Schwesinger C, et al. Angiogenic potential ofprostate carcinoma cells overexpressing bcl-2. J Natl Cancer Inst 2001;93:208– 13.
10. Karl E, Warner K, Zeitlin B, et al. Bcl-2 acts in a proangiogenic signalingpathway through nuclear factor-nB and CXC chemokines. Cancer Res 2005;65:5063–9.
11. Zhu H, Guo W, Zhang L, et al. Bcl-XL small interfering RNA suppresses theproliferation of 5-fluorouracil – resistant human colon cancer cells. Mol CancerTher 2005;4:451 –6.
12. Itoh T, Itoh A, Pleasure D. Bcl-2-related protein family gene expressionduring oligodendroglial differentiation. J Neurochem 2003;85:1500 –12.
13. Wakabayashi K, Saito H, Ebinuma H, et al. Bcl-2 related proteins aredramatically induced at the early stage of differentiation in human liver cancercells by a histone deacetylase inhibitor projecting an antiapoptotic role during thisperiod. Oncol Rep 2000;7:285– 8.
14. Lakics V, Medvedev AE, Okada S, Vogel SN. Inhibition of LPS-inducedcytokines by Bcl-xL in a murine macrophage cell line. J Immunol 2000;165:2729–37.
15. Martin SS, Ridgeway AG, Pinkas J, et al. A cytoskeleton-based functionalgenetic screen identifies Bcl-xL as an enhancer of metastasis, but not primarytumor growth. Oncogene 2004;23:4641 –5.
16. Coluccia AM, Perego S, Cleris L, et al. Bcl-xL down-regulation suppressesthe tumorigenic potential of NPM/ALK in vitro and in vivo . Blood 2004;103:2787–94.
17. Brat DJ, Castellano-Sanchez A, Kaur B, Van Meir EG. Genetic and biologicprogression in astrocytomas and their relation to angiogenic dysregulation. AdvAnat Pathol 2002;91:24– 36.
18. Jiang Z, Zheng X, Rich KM. Down-regulation of Bcl-2 and Bcl-xLexpression with bispecific antisense treatment in glioblastoma cell lines inducecell death. J Neurochem 2003;84:273–81.
19. Nagane M, Levitzki A, Gazit A, Cavenee WK, Huang HJ. Drug resistance ofhuman glioblastoma cells conferred by a tumor-specific mutant epidermal growthfactor receptor through modulation of Bcl-XL and caspase-3 – like proteases. ProcNatl Acad Sci U S A 1998;95:5724 –9.
20. Weiler M, Bahr O, Hohlweg U, et al. BCL-xL: time-dependent dissociationbetween modulation of apoptosis and invasiveness in human malignant gliomacells. Cell Death Differ 2006;13:1156 –69.
21. Krajewski S, Krajewska M, Ehrmann J, et al. Immunohistochemical analysisof Bcl-2, Bcl-X, Mcl-1, and Bax in tumors of central and peripheral nervoussystem origin. Am J Pathol 1997;150:805– 14.
22. Martin S, Toquet C, Oliver L, et al. Expression of bcl-2, bax and bcl-xL inhuman gliomas: a re-appraisal. J Neurooncol 2001;52:129– 39.
23. Olie RA, Hafner C, Kuttel R, et al. Bcl-2 and bcl-xL antisenseoligonucleotides induce apoptosis in melanoma cells of different clinical stages.J Invest Dermatol 2002;118:505– 12.
24. Leiter U, Schmid RM, Kaskel P, Peter RU, Krahn G. Antiapoptotic bcl-2 andbcl-xL in advanced malignant melanoma. Arch Dermatol Res 2000;292:225 –32.
25. Bush JA, Li G. The role of Bcl-2 family members in the progression ofcutaneous melanoma. Clin Exp Metastasis 2003;20:531– 9.
26. Zhang H, Rosdahl I. Bcl-xL and bcl-2 proteins in melanoma progression andUVB-induced apoptosis. Int J Oncol 2006;28:661–6.
27. Koch AE, Polverini PJ, Kunkel SL, et al. Interleukin-8 as a macrophage-derived mediator of angiogenesis. Science 1992;258:1798 –801.
28. Li A, Dubey S, Varney ML, Dave BJ, Singh RK. IL-8 directly enhancedendothelial cell survival, proliferation, and matrix metalloproteinases productionand regulated angiogenesis. J Immunol 2003;170:3369–76.
29. Mukaida N, Mahe Y, Matsushima K. Cooperative interaction of nuclearfactor-nB and cis-regulatory enhancer binding protein-like factor bindingelements in activating the interleukin-8 gene by pro-inflammatory cytokines.J Biol Chem 1990;265:21128 –33.
30. Castilla C, Congregado B, Chinchon D, et al. Bcl-xL is overexpressed inhormone-resistant prostate cancer and promotes survival of LNCaP cells viainteraction with proapoptotic Bak. Endocrinology 2006;147:4960–7.
31. Sulkowski S, Kanczuga-Koda L, Koda M, et al. M Insulin-like growth factor-I receptor correlates with connexin 26 and Bcl-xL expression in human colorectalcancer. Ann N Y Acad Sci 2006;1090:265–75.
32. Del Bufalo D, Trisciuoglio D, Scarsella M, Zangemeister-Wittke U, Zupi G.Lonidamine causes inhibition of angiogenesis-related endothelial cell functions.Neoplasia 2004;6:513 –22.
33. Brat DJ, Bellail AC, Van Meir EG. The role of interleukin-8 and its receptorsin gliomagenesis and tumoral angiogenesis. Neuro-oncol 2005;7:122 –33.
34. Sparmann A, Bar-Sagi D. Ras-induced interleukin-8 expression plays acritical role in tumor growth and angiogenesis. Cancer Cell 2004;6:447 –58.
35. Garkavtsev I, Kozin SV, Chernova O, et al. The candidate tumour suppressorprotein ING4 regulates brain tumour growth and angiogenesis. Nature 2004;428:328 –32.
36. Ugurel S, Rappl G, Tilgen W, Reinhold U. Increased serum concentration ofangiogenic factors in malignant melanoma patients correlates with tumorprogression and survival. J Clin Oncol 2001;19:577– 83.
37. Varney ML, Johansson SL, Singh RK. Distinct expression of CXCL8 and itsreceptors CXCR1 and CXCR2 and their association with vessel density andaggressiveness in malignant melanoma. Am J Clin Pathol 2006;125:209 –16.
38. Nurnberg W, Tobias D, Otto F, Henz BM, Schadendorf D. Expression ofinterleukin-8 detected by in situ hybridization correlates with worse prognosis inprimary cutaneous melanoma. J Pathol 1999;189:546 –51.
39. Reed JC. Dysregulation of apoptosis in cancer. J Clin Oncol 1999;17:2941–53.
40. Kim SH, Nakagawa H, Navaraj A, et al. Tumorigenic conversion of primaryhuman esophageal epithelial cells using oncogene combinations in the absence ofexogenous Ras. Cancer Res 2006;66:10415–24.
41. Zangemeister-Wittke U, Leech SH, Olie RA, et al. A novel bispecificantisense oligonucleotide inhibiting both bcl-2 and bcl-xL expression efficientlyinduces apoptosis in tumor cells. Clin Cancer Res 2000;6:2547–55.
42. Holtmann H, Winzen R, Holland P, et al. Induction of interleukin-8 synthesisintegrates effects on transcription and mRNA degradation from at least threedifferent cytokine- or stress-activated signal transduction pathways. Mol Cell Biol1999;19:6742–53.
Regulation of Angiogenesis by bcl-xL
Mol Cancer Res 2007;5(8). August 2007
771
Research. on November 18, 2018. © 2007 American Association for Cancermcr.aacrjournals.org Downloaded from

2007;5:761-771. Mol Cancer Res Simona Giorgini, Daniela Trisciuoglio, Chiara Gabellini, et al. through CXCL8 ExpressionModulation of bcl-xL in Tumor Cells Regulates Angiogenesis
Updated version
http://mcr.aacrjournals.org/content/5/8/761
Access the most recent version of this article at:
Cited articles
http://mcr.aacrjournals.org/content/5/8/761.full#ref-list-1
This article cites 42 articles, 14 of which you can access for free at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. (CCC)Click on "Request Permissions" which will take you to the Copyright Clearance Center's
.http://mcr.aacrjournals.org/content/5/8/761To request permission to re-use all or part of this article, use this link
Research. on November 18, 2018. © 2007 American Association for Cancermcr.aacrjournals.org Downloaded from