Embed Size (px)
Citation preview

Molecular and Structural Basis for the Roles of Hepatitis C VirusPolymerase NS5B Amino Acids 15, 223, and 321 in Viral Replicationand Drug Resistance
Angela M. Lam,a Thomas E. Edwards,b Ralph T. Mosley,a Eisuke Murakami,a,c Shalini Bansal,a Christopher Lugo,a Haiying Bao,a
Michael J. Otto,a Michael J. Sofia,a Phillip A. Furmana
Pharmasset, Inc., Princeton, New Jersey, USAa; Beryllium, Bainbridge Island, Washington, USAb; Gilead Sciences, Foster City, California, USAc
Resistance to the 2=-F-2=-C-methylguanosine monophosphate nucleotide hepatitis C virus (HCV) inhibitors PSI-352938 andPSI-353661 was associated with a combination of amino acid changes (changes of S to G at position 15 [S15G], C223H, andV321I) within the genotype 2a nonstructural protein 5B (NS5B), an RNA-dependent RNA polymerase. To understand the role ofthese residues in viral replication, we examined the effects of single and multiple point mutations on replication fitness and inhi-bition by a series of nucleotide analog inhibitors. An acidic residue at position 15 reduced replicon fitness, consistent with itsproximity to the RNA template. A change of the residue at position 223 to an acidic or large residue reduced replicon fitness, con-sistent with its proposed proximity to the incoming nucleoside triphosphate (NTP). A change of the residue at position 321 to acharged residue was not tolerated, consistent with its position within a hydrophobic cavity. This triple resistance mutation wasspecific to both genotype 2a virus and 2=-F-2=-C-methylguanosine inhibitors. A crystal structure of the NS5B S15G/C223H/V321Imutant of the JFH-1 isolate exhibited rearrangement to a conformation potentially consistent with short primer-template RNAbinding, which could suggest a mechanism of resistance accomplished through a change in the NS5B conformation, which wasbetter tolerated by genotype 2a virus than by viruses of other genotypes.
Hepatitis C virus (HCV) is a single-stranded positive-senseRNA virus and the cause of hepatitis C. Nonstructural pro-
tein 5B (NS5B) is the RNA-dependent RNA polymerase (RdRp)responsible for replication of the viral genome (1) and adopts aright-hand structure with palm, finger, and thumb domains com-mon to nucleotide polymerases (2–4). In addition, HCV NS5Bcontains an extension of the finger region called the fingertips, aswell as a �-hairpin loop or �-flap insertion into the thumb do-main that protrudes deep into the active site and appears to be ahallmark of Flaviviridae RdRps (5). It has been suggested that asignificant structural rearrangement of the thumb domain, in-cluding relocation of the �-flap and a C-terminal-membrane-an-choring linker from the active-site cavity, must take place to ac-commodate a growing primer-template RNA chain (3, 5, 6).Indeed, such a rearrangement was observed in an elongation-phase binary complex of an NS5B �-flap deletion mutant withprimer-template RNA (7). A crystal structure of an HCV NS5Bternary assembly is not yet available, and thus, insights into nucle-oside triphosphate (NTP) binding and incorporation have beenderived from biochemical studies and homology modeling.
NS5B is critical for viral replication, exhibits important differ-ences from cellular polymerases, and has become a major targetfor antiviral drug development. Chain-terminating nucleotide an-alog inhibitors are promising drugs against HCV because theytarget the conserved active site and exhibit pan-genotype activityand a high barrier of resistance (8). One such compound, sofos-buvir (GS-7977) (Fig. 1), was recently approved by the FDA. Be-cause clinical resistance is a major concern when treating viralinfections, understanding what mutations might arise and conferviral resistance is important in the development of any antiviralagent. We recently reported that the 2=-F-2=-C-methylguanosine(2=-F-2=-C-MeGTP) analogs PSI-352938 and PSI-353661 selectedfor a number of amino acid changes within the finger and palm
domains (changes of S to G at position 15 [S15G], C223H, andV321I) that, in combination, conferred resistance to this series ofinhibitors (9). Here, we examine the importance of these residuesin viral replication using mutational analysis coupled with assaysof replicon fitness and inhibition by nucleotide analog inhibitors,as well as a crystal structure of the NS5B S15G/C223H/V321I mu-tant of the JFH-1 isolate that we obtained. Our combined cell-based and structural results provide a better understanding of theroles of residues 15, 223, and 321 in HCV replication.
MATERIALS AND METHODSCompounds. All nucleoside/-tide analog inhibitors (Fig. 1) were synthe-sized at Pharmasset and shown to be more than �95 to 99% pure byproton nuclear magnetic resonance, mass spectrometry, and high-perfor-mance liquid chromatography analyses.
Replicons and cells. All plasmid sequences were confirmed by se-quencing. Plasmid DNA containing the genotype 1a (GT 1a) replicon ofisolate H77 (NCBI reference NC_004102.1) with adaptive amino acidchanges P1496L and S2204I and the J6/JFH-1 isolate-derived GT 2a(JFH-1 GT 2a) were licensed from Apath. The JFH-1 GT 2a repliconcontains the first 19 amino acids and 3=-nontranslated region (NTR) fromthe J6 strain (GenBank accession no. AF177036) and the 5= NTR andNS3-to-NS5B region from the JFH-1 strain (GenBank accession no.AB047639). The JFH-1 replicon open reading frame also contains a Re-
Received 9 July 2014 Returned for modification 7 August 2014Accepted 29 August 2014
Published ahead of print 2 September 2014
Address correspondence to Angela M. Lam, [email protected], orThomas E. Edwards, [email protected].
Copyright © 2014, American Society for Microbiology. All Rights Reserved.
doi:10.1128/AAC.03847-14
November 2014 Volume 58 Number 11 Antimicrobial Agents and Chemotherapy p. 6861– 6869 aac.asm.org 6861
on June 27, 2018 by guesthttp://aac.asm
.org/D
ownloaded from
nilla luciferase reporter gene upstream from the neomycin phosphotrans-ferase gene (Neo). We generated a GT 1a replicon with a Renilla luciferasereporter gene in between the 5=NTR and Neo by cloning and introducedan adaptive change (NS3 K583E) by site-directed mutagenesis to enhanceits replication fitness. The Lunet cell line and the Con1 isolate-derived GT1b ET replicon (GenBank accession no. AJ238799.1) with the firefly lucif-erase reporter gene and adaptive changes E1202G, T1280I, and K1846Twere provided by R. Bartenschlager (University of Heidelberg). The plas-mid containing the J6 GT 2a NS5B sequence was synthesized at IDT. GT3a NS5B was cloned from a human serum sample (EMBL ENA accessionno. LN555582). J6 and GT 3a NS5B were each cloned into a Con1 GT 1breplicon that encodes a Renilla luciferase reporter gene to generate thechimeric replicons GT 1b/J6 GT 2a NS5B and GT 1b/GT 3a NS5B.
Mutagenesis and HCV replicon transient-transfection assays. Mu-tations were introduced into the appropriate replicon plasmids usingQuikChange II site-directed mutagenesis (Agilent) and primers (IDT).Replicon RNA was generated by in vitro transcription from linearizedplasmids (digested with ScaI for Con1 GT 1b and GT 1b/J6 NS5B, HpaIfor H77 GT 1a, and XbaI for JFH-1 GT 2a) using the RiboMAX large-scaleRNA T7 kit (Promega). Replicon RNA (10 �g) was electroporated intoLunet cells as described previously (10) to evaluate replication efficiencyand susceptibility to inhibition by nucleoside/-tide analogs. Transient-transfection assays were performed by transfecting cells and seeding themat a density of 8,000 cells/well. Cells were incubated with serially dilutedcompounds for 4 days prior to the luciferase assay. Replication fitness wasdetermined by normalizing the luciferase expression at 96 h with that at 4h and then dividing the normalized level of replicon mutant luciferaseexpression by that of the wild type. HCV replicon inhibition was deter-mined by measuring the levels of luminescence expressed via the firefly orRenilla luciferase reporter gene using Bright-Glo or Renilla-Glo, respec-tively (Promega). The concentrations at which 50% inhibition wasachieved (50% effective concentration [EC50]) were determined usingGraphPad Prism.
Expression and purification of JFH-1 NS5B. JFH-1 NS5B �21 wascloned from the plasmid DNA described above by amplification usingPlatinum PCR supermix high fidelity (HF) (Invitrogen), with forwardprimer 5=-GACGCTCATGAGCTCCATGTCATACTCCTGGACCGGGGCTC and reverse primer 5=-GTGCTCGAGGCGGGGTCGGGCGCGC,into the pET28a(�) expression vector (EMD) containing a C-terminalHis6 tag. Site-directed mutagenesis was performed using QuikChange II(Agilent) to obtain the plasmid encoding the S15G/C223H/V321I muta-tions. The plasmid was transformed into Rosetta DE3 E. coli competentcells, and the cells were grown in LB medium at 37°C to an optical density
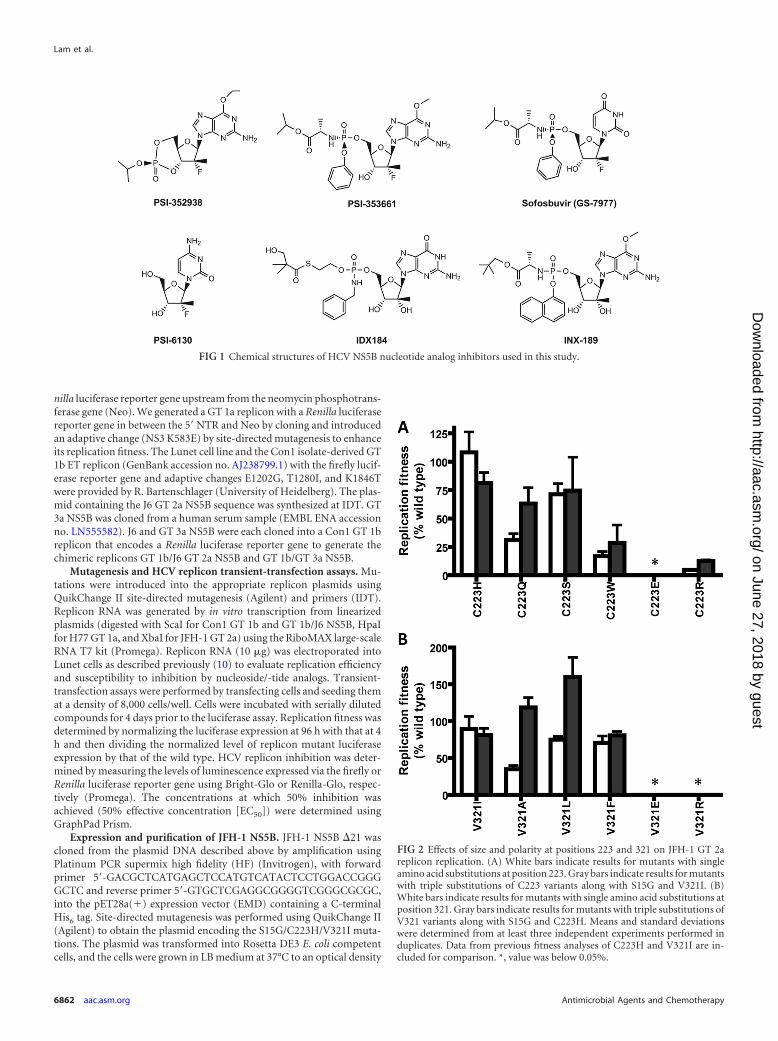
FIG 1 Chemical structures of HCV NS5B nucleotide analog inhibitors used in this study.
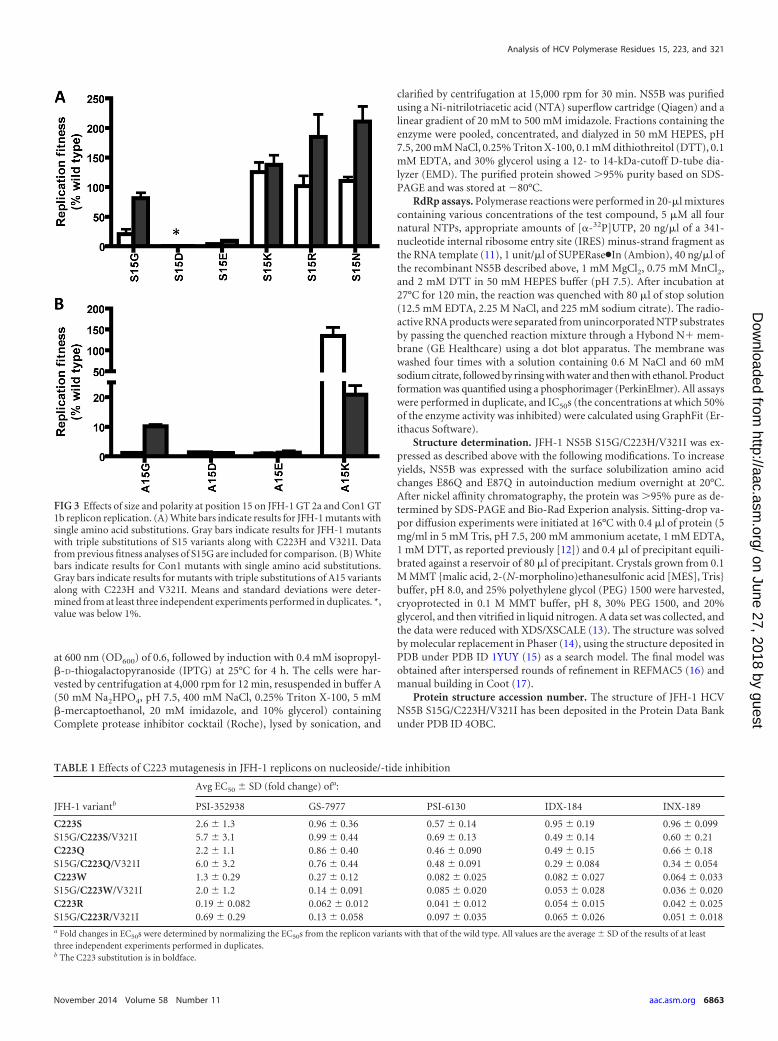
FIG 2 Effects of size and polarity at positions 223 and 321 on JFH-1 GT 2areplicon replication. (A) White bars indicate results for mutants with singleamino acid substitutions at position 223. Gray bars indicate results for mutantswith triple substitutions of C223 variants along with S15G and V321I. (B)White bars indicate results for mutants with single amino acid substitutions atposition 321. Gray bars indicate results for mutants with triple substitutions ofV321 variants along with S15G and C223H. Means and standard deviationswere determined from at least three independent experiments performed induplicates. Data from previous fitness analyses of C223H and V321I are in-cluded for comparison. *, value was below 0.05%.
Lam et al.
6862 aac.asm.org Antimicrobial Agents and Chemotherapy
on June 27, 2018 by guesthttp://aac.asm
.org/D
ownloaded from
at 600 nm (OD600) of 0.6, followed by induction with 0.4 mM isopropyl-�-D-thiogalactopyranoside (IPTG) at 25°C for 4 h. The cells were har-vested by centrifugation at 4,000 rpm for 12 min, resuspended in buffer A(50 mM Na2HPO4, pH 7.5, 400 mM NaCl, 0.25% Triton X-100, 5 mM�-mercaptoethanol, 20 mM imidazole, and 10% glycerol) containingComplete protease inhibitor cocktail (Roche), lysed by sonication, and
clarified by centrifugation at 15,000 rpm for 30 min. NS5B was purifiedusing a Ni-nitrilotriacetic acid (NTA) superflow cartridge (Qiagen) and alinear gradient of 20 mM to 500 mM imidazole. Fractions containing theenzyme were pooled, concentrated, and dialyzed in 50 mM HEPES, pH7.5, 200 mM NaCl, 0.25% Triton X-100, 0.1 mM dithiothreitol (DTT), 0.1mM EDTA, and 30% glycerol using a 12- to 14-kDa-cutoff D-tube dia-lyzer (EMD). The purified protein showed �95% purity based on SDS-PAGE and was stored at �80°C.
RdRp assays. Polymerase reactions were performed in 20-�l mixturescontaining various concentrations of the test compound, 5 �M all fournatural NTPs, appropriate amounts of [�-32P]UTP, 20 ng/�l of a 341-nucleotide internal ribosome entry site (IRES) minus-strand fragment asthe RNA template (11), 1 unit/�l of SUPERase●In (Ambion), 40 ng/�l ofthe recombinant NS5B described above, 1 mM MgCl2, 0.75 mM MnCl2,and 2 mM DTT in 50 mM HEPES buffer (pH 7.5). After incubation at27°C for 120 min, the reaction was quenched with 80 �l of stop solution(12.5 mM EDTA, 2.25 M NaCl, and 225 mM sodium citrate). The radio-active RNA products were separated from unincorporated NTP substratesby passing the quenched reaction mixture through a Hybond N� mem-brane (GE Healthcare) using a dot blot apparatus. The membrane waswashed four times with a solution containing 0.6 M NaCl and 60 mMsodium citrate, followed by rinsing with water and then with ethanol. Productformation was quantified using a phosphorimager (PerkinElmer). All assayswere performed in duplicate, and IC50s (the concentrations at which 50%of the enzyme activity was inhibited) were calculated using GraphFit (Er-ithacus Software).
Structure determination. JFH-1 NS5B S15G/C223H/V321I was ex-pressed as described above with the following modifications. To increaseyields, NS5B was expressed with the surface solubilization amino acidchanges E86Q and E87Q in autoinduction medium overnight at 20°C.After nickel affinity chromatography, the protein was �95% pure as de-termined by SDS-PAGE and Bio-Rad Experion analysis. Sitting-drop va-por diffusion experiments were initiated at 16°C with 0.4 �l of protein (5mg/ml in 5 mM Tris, pH 7.5, 200 mM ammonium acetate, 1 mM EDTA,1 mM DTT, as reported previously [12]) and 0.4 �l of precipitant equili-brated against a reservoir of 80 �l of precipitant. Crystals grown from 0.1M MMT {malic acid, 2-(N-morpholino)ethanesulfonic acid [MES], Tris}buffer, pH 8.0, and 25% polyethylene glycol (PEG) 1500 were harvested,cryoprotected in 0.1 M MMT buffer, pH 8, 30% PEG 1500, and 20%glycerol, and then vitrified in liquid nitrogen. A data set was collected, andthe data were reduced with XDS/XSCALE (13). The structure was solvedby molecular replacement in Phaser (14), using the structure deposited inPDB under PDB ID 1YUY (15) as a search model. The final model wasobtained after interspersed rounds of refinement in REFMAC5 (16) andmanual building in Coot (17).
Protein structure accession number. The structure of JFH-1 HCVNS5B S15G/C223H/V321I has been deposited in the Protein Data Bankunder PDB ID 4OBC.
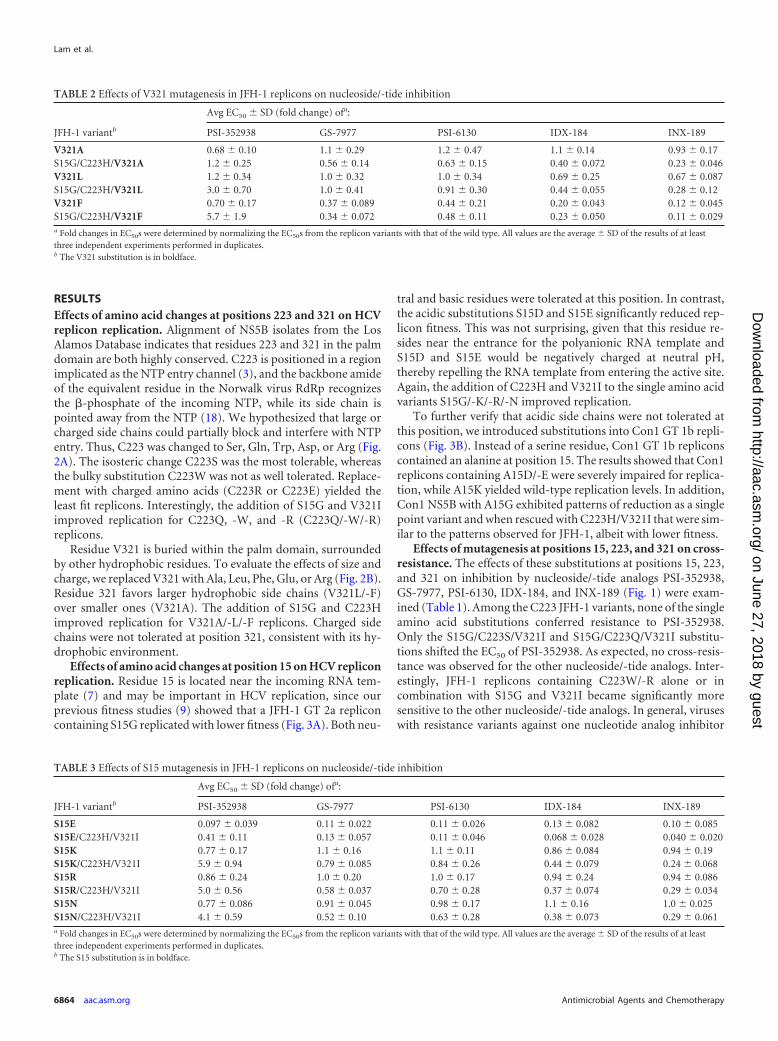
FIG 3 Effects of size and polarity at position 15 on JFH-1 GT 2a and Con1 GT1b replicon replication. (A) White bars indicate results for JFH-1 mutants withsingle amino acid substitutions. Gray bars indicate results for JFH-1 mutantswith triple substitutions of S15 variants along with C223H and V321I. Datafrom previous fitness analyses of S15G are included for comparison. (B) Whitebars indicate results for Con1 mutants with single amino acid substitutions.Gray bars indicate results for mutants with triple substitutions of A15 variantsalong with C223H and V321I. Means and standard deviations were deter-mined from at least three independent experiments performed in duplicates. *,value was below 1%.
TABLE 1 Effects of C223 mutagenesis in JFH-1 replicons on nucleoside/-tide inhibition
JFH-1 variantb
Avg EC50 SD (fold change) ofa:
PSI-352938 GS-7977 PSI-6130 IDX-184 INX-189
C223S 2.6 1.3 0.96 0.36 0.57 0.14 0.95 0.19 0.96 0.099S15G/C223S/V321I 5.7 3.1 0.99 0.44 0.69 0.13 0.49 0.14 0.60 0.21C223Q 2.2 1.1 0.86 0.40 0.46 0.090 0.49 0.15 0.66 0.18S15G/C223Q/V321I 6.0 3.2 0.76 0.44 0.48 0.091 0.29 0.084 0.34 0.054C223W 1.3 0.29 0.27 0.12 0.082 0.025 0.082 0.027 0.064 0.033S15G/C223W/V321I 2.0 1.2 0.14 0.091 0.085 0.020 0.053 0.028 0.036 0.020C223R 0.19 0.082 0.062 0.012 0.041 0.012 0.054 0.015 0.042 0.025S15G/C223R/V321I 0.69 0.29 0.13 0.058 0.097 0.035 0.065 0.026 0.051 0.018a Fold changes in EC50s were determined by normalizing the EC50s from the replicon variants with that of the wild type. All values are the average SD of the results of at leastthree independent experiments performed in duplicates.b The C223 substitution is in boldface.
Analysis of HCV Polymerase Residues 15, 223, and 321
November 2014 Volume 58 Number 11 aac.asm.org 6863
on June 27, 2018 by guesthttp://aac.asm
.org/D
ownloaded from
RESULTSEffects of amino acid changes at positions 223 and 321 on HCVreplicon replication. Alignment of NS5B isolates from the LosAlamos Database indicates that residues 223 and 321 in the palmdomain are both highly conserved. C223 is positioned in a regionimplicated as the NTP entry channel (3), and the backbone amideof the equivalent residue in the Norwalk virus RdRp recognizesthe �-phosphate of the incoming NTP, while its side chain ispointed away from the NTP (18). We hypothesized that large orcharged side chains could partially block and interfere with NTPentry. Thus, C223 was changed to Ser, Gln, Trp, Asp, or Arg (Fig.2A). The isosteric change C223S was the most tolerable, whereasthe bulky substitution C223W was not as well tolerated. Replace-ment with charged amino acids (C223R or C223E) yielded theleast fit replicons. Interestingly, the addition of S15G and V321Iimproved replication for C223Q, -W, and -R (C223Q/-W/-R)replicons.
Residue V321 is buried within the palm domain, surroundedby other hydrophobic residues. To evaluate the effects of size andcharge, we replaced V321 with Ala, Leu, Phe, Glu, or Arg (Fig. 2B).Residue 321 favors larger hydrophobic side chains (V321L/-F)over smaller ones (V321A). The addition of S15G and C223Himproved replication for V321A/-L/-F replicons. Charged sidechains were not tolerated at position 321, consistent with its hy-drophobic environment.
Effects of amino acid changes at position 15 on HCV repliconreplication. Residue 15 is located near the incoming RNA tem-plate (7) and may be important in HCV replication, since ourprevious fitness studies (9) showed that a JFH-1 GT 2a repliconcontaining S15G replicated with lower fitness (Fig. 3A). Both neu-
tral and basic residues were tolerated at this position. In contrast,the acidic substitutions S15D and S15E significantly reduced rep-licon fitness. This was not surprising, given that this residue re-sides near the entrance for the polyanionic RNA template andS15D and S15E would be negatively charged at neutral pH,thereby repelling the RNA template from entering the active site.Again, the addition of C223H and V321I to the single amino acidvariants S15G/-K/-R/-N improved replication.
To further verify that acidic side chains were not tolerated atthis position, we introduced substitutions into Con1 GT 1b repli-cons (Fig. 3B). Instead of a serine residue, Con1 GT 1b repliconscontained an alanine at position 15. The results showed that Con1replicons containing A15D/-E were severely impaired for replica-tion, while A15K yielded wild-type replication levels. In addition,Con1 NS5B with A15G exhibited patterns of reduction as a singlepoint variant and when rescued with C223H/V321I that were sim-ilar to the patterns observed for JFH-1, albeit with lower fitness.
Effects of mutagenesis at positions 15, 223, and 321 on cross-resistance. The effects of these substitutions at positions 15, 223,and 321 on inhibition by nucleoside/-tide analogs PSI-352938,GS-7977, PSI-6130, IDX-184, and INX-189 (Fig. 1) were exam-ined (Table 1). Among the C223 JFH-1 variants, none of the singleamino acid substitutions conferred resistance to PSI-352938.Only the S15G/C223S/V321I and S15G/C223Q/V321I substitu-tions shifted the EC50 of PSI-352938. As expected, no cross-resis-tance was observed for the other nucleoside/-tide analogs. Inter-estingly, JFH-1 replicons containing C223W/-R alone or incombination with S15G and V321I became significantly moresensitive to the other nucleoside/-tide analogs. In general, viruseswith resistance variants against one nucleotide analog inhibitor
TABLE 2 Effects of V321 mutagenesis in JFH-1 replicons on nucleoside/-tide inhibition
JFH-1 variantb
Avg EC50 SD (fold change) ofa:
PSI-352938 GS-7977 PSI-6130 IDX-184 INX-189
V321A 0.68 0.10 1.1 0.29 1.2 0.47 1.1 0.14 0.93 0.17S15G/C223H/V321A 1.2 0.25 0.56 0.14 0.63 0.15 0.40 0.072 0.23 0.046V321L 1.2 0.34 1.0 0.32 1.0 0.34 0.69 0.25 0.67 0.087S15G/C223H/V321L 3.0 0.70 1.0 0.41 0.91 0.30 0.44 0.055 0.28 0.12V321F 0.70 0.17 0.37 0.089 0.44 0.21 0.20 0.043 0.12 0.045S15G/C223H/V321F 5.7 1.9 0.34 0.072 0.48 0.11 0.23 0.050 0.11 0.029a Fold changes in EC50s were determined by normalizing the EC50s from the replicon variants with that of the wild type. All values are the average SD of the results of at leastthree independent experiments performed in duplicates.b The V321 substitution is in boldface.
TABLE 3 Effects of S15 mutagenesis in JFH-1 replicons on nucleoside/-tide inhibition
JFH-1 variantb
Avg EC50 SD (fold change) ofa:
PSI-352938 GS-7977 PSI-6130 IDX-184 INX-189
S15E 0.097 0.039 0.11 0.022 0.11 0.026 0.13 0.082 0.10 0.085S15E/C223H/V321I 0.41 0.11 0.13 0.057 0.11 0.046 0.068 0.028 0.040 0.020S15K 0.77 0.17 1.1 0.16 1.1 0.11 0.86 0.084 0.94 0.19S15K/C223H/V321I 5.9 0.94 0.79 0.085 0.84 0.26 0.44 0.079 0.24 0.068S15R 0.86 0.24 1.0 0.20 1.0 0.17 0.94 0.24 0.94 0.086S15R/C223H/V321I 5.0 0.56 0.58 0.037 0.70 0.28 0.37 0.074 0.29 0.034S15N 0.77 0.086 0.91 0.045 0.98 0.17 1.1 0.16 1.0 0.025S15N/C223H/V321I 4.1 0.59 0.52 0.10 0.63 0.28 0.38 0.073 0.29 0.061a Fold changes in EC50s were determined by normalizing the EC50s from the replicon variants with that of the wild type. All values are the average SD of the results of at leastthree independent experiments performed in duplicates.b The S15 substitution is in boldface.
Lam et al.
6864 aac.asm.org Antimicrobial Agents and Chemotherapy
on June 27, 2018 by guesthttp://aac.asm
.org/D
ownloaded from

that exhibit loss of fitness may appear more susceptible to differ-ent nucleotide analog inhibitors than wild-type strains.
Among the V321 variants capable of replication (the variantwith a mutation to Asp failed to replicate), we observed that re-duction in PSI-352938 activity was dependent both on the com-bination with S15G and C223H and on the size of the side chain(Table 2). Essentially no change in EC50s was observed for thevariants with single point changes, and resistance was not ob-served for the other nucleoside/-tide analogs.
The effects of modifying residue 15 were evaluated using bothJFH-1 (Table 3) and Con1 (Table 4) replicons. JFH-1 S15 variantswith single amino acid substitutions remained susceptible to nu-cleoside/-tide analogs, and reduction of PSI-352938 activity wasonly observed in the presence of C223H and V321I together witha nonacidic amino acid at position 15 (Table 3). The only residue15 substitution that produced replication-competent replicons inCon1 was A15K as a single mutation or part of a triple mutation,and a more modest increase in the EC50 was observed for Con1(2.6-fold) (Table 4) than for JFH-1 (5.9-fold) (Table 3) with theA15K/C223H/V321I substitutions against PSI-352938 only.
Effects of the S15G/C223H/V321I mutations in GT 1a and in1b/2a and 1b/3a chimeras. JFH-1 replicons tolerated the S15G/C223H/V321I mutations much better than Con1 replicons con-taining these mutations (9). For this study, H77 GT 1a or chimericreplicons containing Con1 GT 1b as the backbone and either J6GT 2a or GT 3a NS5B were constructed as wild type or as theC223H, C223H/V321I or S15G/C223H/V321I variant and as-sayed for replication fitness (Table 5). H77 GT 1a replicons con-taining the C223H substitution showed reduced replication levels,which were further lowered by the addition of A15G and V321Imutations. The J6 GT 2a chimeric replicons tolerated the single,double, or triple mutation. Similar to GT 1 replicons, chimericCon1/GT 3a NS5B replicons with amino acid substitutions at sites15, 223, and/or 321 were severely impaired for replication. Dose-response studies using GT 1a and GT 1b/J6 NS5B replicon mu-
tants showed that the single C223H amino acid change did notsignificantly affect PSI-352938 activity, whereas double or triplemutants showed increased resistance (Table 5).
Biochemical studies of JFH-1 NS5B variants. The effects ofthe S15G/C223H/V321I triple variant on RNA synthesis and theapparent IC50s of PSI-352666 (active 5=-triphosphate of PSI-352938 and PSI-353661) and 2=-C-methylguanosine triphosphate(2=-C-MeGTP; the active form of IDX-184) were examined. Theactivity of the triple variant enzyme was approximately 50%higher than that of the wild-type enzyme, and both PSI-352666and 2=-C-MeGTP showed dose-dependent inhibition (Table 6).The variant enzyme demonstrated approximately 2-fold-reducedsensitivity to PSI-352666, whereas 2=-C-MeGTP was 2.5-foldmore active, demonstrating that 2=-F-2=-C-MeGTP was sensitiveto these amino acid changes and 2=-OH-2=-C-MeGTP was not.
JFH-1 S15G/C223H/V321I NS5B crystal structure. We ob-tained a crystal structure of JFH-1 S15G/C223H/V321I NS5B at2.5-Å resolution (Fig. 4A; Table 7). Each amino acid change wasclearly defined by the electron density. C223H adopted the thirdmost common rotamer conformation, while V321I adopted themost common rotamer conformation. Surprisingly, JFH-1 NS5BS15G/C223H/V321I exhibited a significantly different conforma-tion (Fig. 4B) than most other GT 2a crystal structures, such asthose of JFH-1 (PDB ID 3I5K [12]; C� root mean square deviation[RMSD], 1.80 Å, and PDB ID 2XXD [19]; C� RMSD, 1.76 Å), theGT 2a consensus sequence as apo (PDB ID 1YUY [15]; C� RMSD,1.58 Å) or in complex with a thumb site nonnucleoside inhibitor(PDB ID 1YVX [15]; C� RMSD, 1.10 Å), or the J6 isolate (PDB ID2XWH [19]; C� RMSD, 1.79 Å). Although the finger and palmdomains were quite similar, significant rearrangement occurredin the fingertip and thumb domains, including a partial disorder-ing of the C terminus and a refolding of the loop that spans theprimer grip helix and the primer buttress helix (residues W397 toN406) (Fig. 4B). All of these rearrangements were observed pre-viously in our structures of a �-hairpin loop deletion construct of
TABLE 4 Effects of A15 mutagenesis in Con1 replicons on nucleoside/-tide inhibition
Con1 variantb
Avg EC50 SD (fold change) ofa:
PSI-352938 GS-7977 PSI-6130 IDX-184 INX-189
A15K 1.1 0.33 1.3 0.39 1.3 0.21 1.1 0.21 1.1 0.46A15K/C223H/V321I 2.6 0.71 1.1 0.32 0.84 0.27 0.62 0.21 0.27 0.093a Fold changes in EC50s were determined by normalizing the EC50s from the replicon variants with that of the wild type. All values are the average SD of the results of at leastthree independent experiments performed in duplicates.b The A15 substitution is in boldface.
TABLE 5 Effects of changes in residues 15, 223, and 321 in NS5Bs from other isolates
Subgenomic repliconvariant
Avg SD of indicated value for varianta:
C223H C223H/V321I A-/S15Gb/C223H/V321I
Replication fitness(%)
PSI-352938 EC50
(fold change)Replicationfitness (%)
PSI-352938 EC50
(fold change)Replicationfitness (%)
PSI-352938 EC50
(fold change)
GT 1a 28.3 4.8 1.5 0.2 6.0 2.2 3.7 0.5 4.4 0.7 8.7 0.8GT 1b/J6 GT 2a 74.6 15.3 2.1 0.5 43.6 12.1 3.7 0.9 69.5 17.5 8.0 1.4GT 1b/GT 3a 8.5 4.4 ND 5.3 2.8 ND 5.5 1.3 NDa All values are the average SD of the results of at least three independent experiments performed in duplicates. Replication fitness values are the percentage of the value for thewild type. The EC50 values are the fold change relative to the value for the wild type. ND, not determined, because luciferase expression levels in GT 3a variants were similar tobackground values.b Residue 15 is an Ala in GT 1a and GT 3a and a Ser in J6 GT 2a.
Analysis of HCV Polymerase Residues 15, 223, and 321
November 2014 Volume 58 Number 11 aac.asm.org 6865
on June 27, 2018 by guesthttp://aac.asm
.org/D
ownloaded from
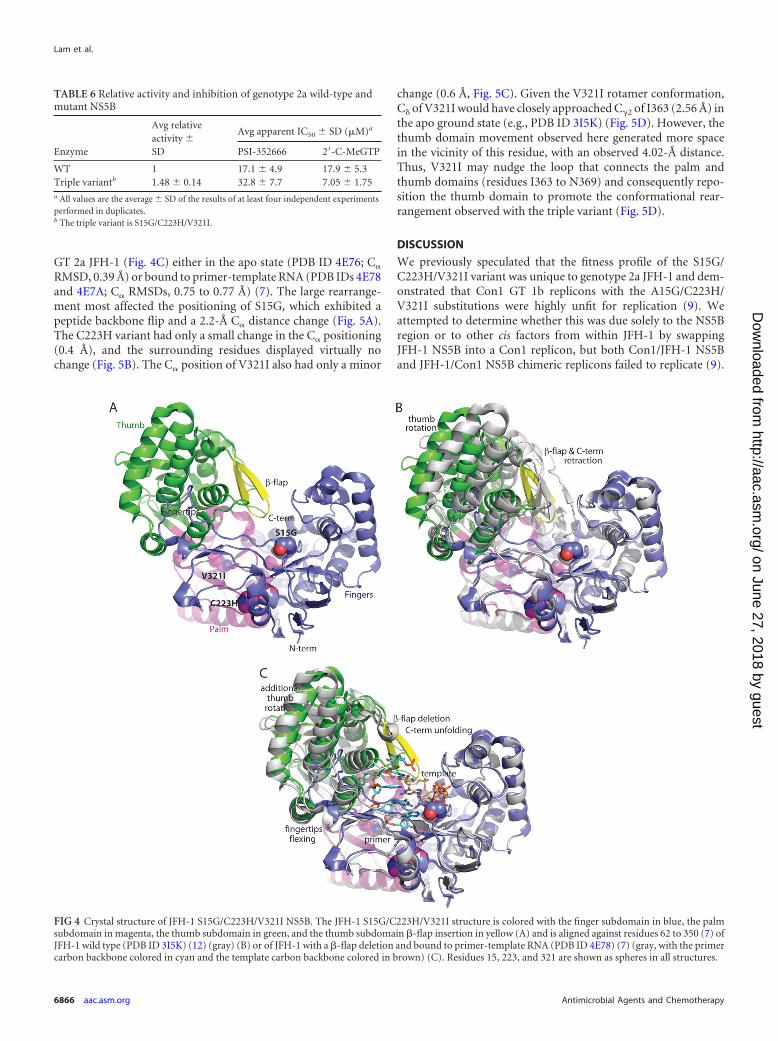
GT 2a JFH-1 (Fig. 4C) either in the apo state (PDB ID 4E76; C�
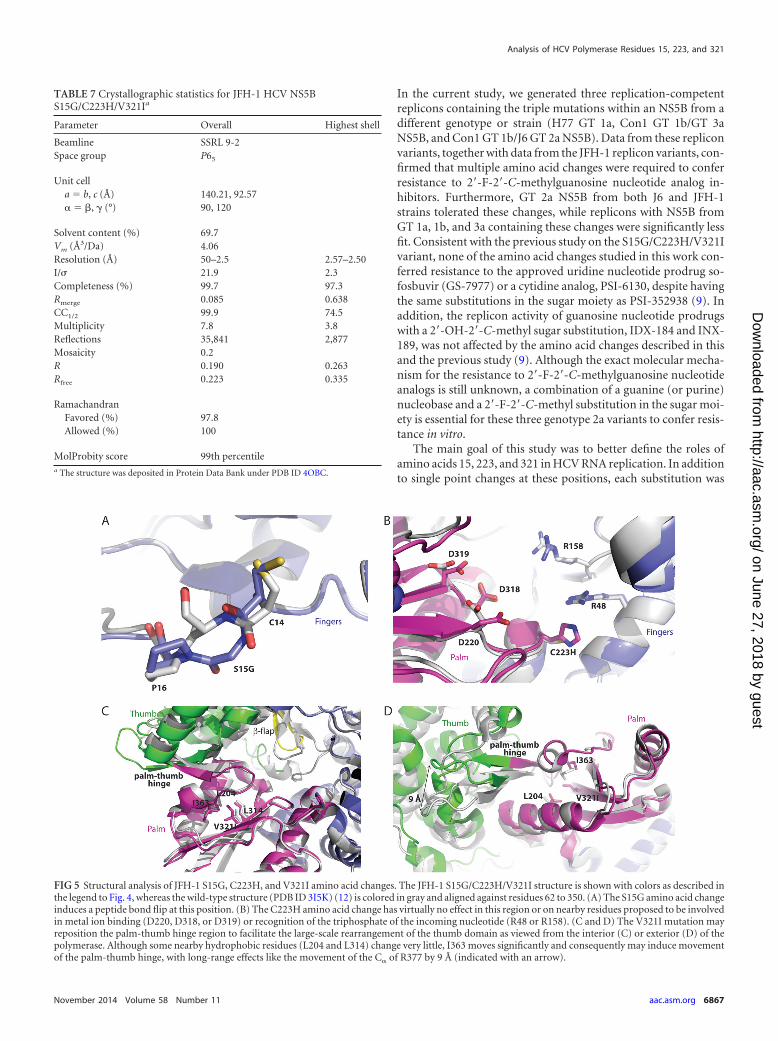
RMSD, 0.39 Å) or bound to primer-template RNA (PDB IDs 4E78and 4E7A; C� RMSDs, 0.75 to 0.77 Å) (7). The large rearrange-ment most affected the positioning of S15G, which exhibited apeptide backbone flip and a 2.2-Å C� distance change (Fig. 5A).The C223H variant had only a small change in the C� positioning(0.4 Å), and the surrounding residues displayed virtually nochange (Fig. 5B). The C� position of V321I also had only a minor
change (0.6 Å, Fig. 5C). Given the V321I rotamer conformation,C of V321I would have closely approached C�2 of I363 (2.56 Å) inthe apo ground state (e.g., PDB ID 3I5K) (Fig. 5D). However, thethumb domain movement observed here generated more spacein the vicinity of this residue, with an observed 4.02-Å distance.Thus, V321I may nudge the loop that connects the palm andthumb domains (residues I363 to N369) and consequently repo-sition the thumb domain to promote the conformational rear-rangement observed with the triple variant (Fig. 5D).
DISCUSSION
We previously speculated that the fitness profile of the S15G/C223H/V321I variant was unique to genotype 2a JFH-1 and dem-onstrated that Con1 GT 1b replicons with the A15G/C223H/V321I substitutions were highly unfit for replication (9). Weattempted to determine whether this was due solely to the NS5Bregion or to other cis factors from within JFH-1 by swappingJFH-1 NS5B into a Con1 replicon, but both Con1/JFH-1 NS5Band JFH-1/Con1 NS5B chimeric replicons failed to replicate (9).
TABLE 6 Relative activity and inhibition of genotype 2a wild-type andmutant NS5B
Enzyme
Avg relativeactivity SD
Avg apparent IC50 SD (�M)a
PSI-352666 2=-C-MeGTP
WT 1 17.1 4.9 17.9 5.3Triple variantb 1.48 0.14 32.8 7.7 7.05 1.75a All values are the average SD of the results of at least four independent experimentsperformed in duplicates.b The triple variant is S15G/C223H/V321I.
FIG 4 Crystal structure of JFH-1 S15G/C223H/V321I NS5B. The JFH-1 S15G/C223H/V321I structure is colored with the finger subdomain in blue, the palmsubdomain in magenta, the thumb subdomain in green, and the thumb subdomain �-flap insertion in yellow (A) and is aligned against residues 62 to 350 (7) ofJFH-1 wild type (PDB ID 3I5K) (12) (gray) (B) or of JFH-1 with a �-flap deletion and bound to primer-template RNA (PDB ID 4E78) (7) (gray, with the primercarbon backbone colored in cyan and the template carbon backbone colored in brown) (C). Residues 15, 223, and 321 are shown as spheres in all structures.
Lam et al.
6866 aac.asm.org Antimicrobial Agents and Chemotherapy
on June 27, 2018 by guesthttp://aac.asm
.org/D
ownloaded from
In the current study, we generated three replication-competentreplicons containing the triple mutations within an NS5B from adifferent genotype or strain (H77 GT 1a, Con1 GT 1b/GT 3aNS5B, and Con1 GT 1b/J6 GT 2a NS5B). Data from these repliconvariants, together with data from the JFH-1 replicon variants, con-firmed that multiple amino acid changes were required to conferresistance to 2=-F-2=-C-methylguanosine nucleotide analog in-hibitors. Furthermore, GT 2a NS5B from both J6 and JFH-1strains tolerated these changes, while replicons with NS5B fromGT 1a, 1b, and 3a containing these changes were significantly lessfit. Consistent with the previous study on the S15G/C223H/V321Ivariant, none of the amino acid changes studied in this work con-ferred resistance to the approved uridine nucleotide prodrug so-fosbuvir (GS-7977) or a cytidine analog, PSI-6130, despite havingthe same substitutions in the sugar moiety as PSI-352938 (9). Inaddition, the replicon activity of guanosine nucleotide prodrugswith a 2=-OH-2=-C-methyl sugar substitution, IDX-184 and INX-189, was not affected by the amino acid changes described in thisand the previous study (9). Although the exact molecular mecha-nism for the resistance to 2=-F-2=-C-methylguanosine nucleotideanalogs is still unknown, a combination of a guanine (or purine)nucleobase and a 2=-F-2=-C-methyl substitution in the sugar moi-ety is essential for these three genotype 2a variants to confer resis-tance in vitro.
The main goal of this study was to better define the roles ofamino acids 15, 223, and 321 in HCV RNA replication. In additionto single point changes at these positions, each substitution was
TABLE 7 Crystallographic statistics for JFH-1 HCV NS5BS15G/C223H/V321Ia
Parameter Overall Highest shell
Beamline SSRL 9-2Space group P65
Unit cella � b, c (Å) 140.21, 92.57� � �, � (°) 90, 120
Solvent content (%) 69.7Vm (Å3/Da) 4.06Resolution (Å) 50–2.5 2.57–2.50I/ 21.9 2.3Completeness (%) 99.7 97.3Rmerge 0.085 0.638CC1/2 99.9 74.5Multiplicity 7.8 3.8Reflections 35,841 2,877Mosaicity 0.2R 0.190 0.263Rfree 0.223 0.335
RamachandranFavored (%) 97.8Allowed (%) 100
MolProbity score 99th percentilea The structure was deposited in Protein Data Bank under PDB ID 4OBC.
FIG 5 Structural analysis of JFH-1 S15G, C223H, and V321I amino acid changes. The JFH-1 S15G/C223H/V321I structure is shown with colors as described inthe legend to Fig. 4, whereas the wild-type structure (PDB ID 3I5K) (12) is colored in gray and aligned against residues 62 to 350. (A) The S15G amino acid changeinduces a peptide bond flip at this position. (B) The C223H amino acid change has virtually no effect in this region or on nearby residues proposed to be involvedin metal ion binding (D220, D318, or D319) or recognition of the triphosphate of the incoming nucleotide (R48 or R158). (C and D) The V321I mutation mayreposition the palm-thumb hinge region to facilitate the large-scale rearrangement of the thumb domain as viewed from the interior (C) or exterior (D) of thepolymerase. Although some nearby hydrophobic residues (L204 and L314) change very little, I363 moves significantly and consequently may induce movementof the palm-thumb hinge, with long-range effects like the movement of the C� of R377 by 9 Å (indicated with an arrow).
Analysis of HCV Polymerase Residues 15, 223, and 321
November 2014 Volume 58 Number 11 aac.asm.org 6867
on June 27, 2018 by guesthttp://aac.asm
.org/D
ownloaded from

combined with the other two amino acid changes that were pre-viously determined to be essential in conferring resistance to 2=-F-2=-C-methylguanine nucleotides. Mutagenesis studies con-firmed the importance of each of these residues. Amino acid 15 islocated on the surface of the finger domain near the incomingtemplate (7) and is more polymorphic than positions 223 and 321.It is primarily an Ala in GT 1, 4, and 6, a Ser in GT 3 and 5, and aSer or Gly (�1:1 ratio) in GT 2 viruses. The reduced fitness ofreplicons with an acidic side chain could reflect disruption of in-teractions of the NS5B with the RNA template. Residue 223 mayinteract with the incoming NTP (3), and it could not tolerate achange to Glu or Arg. Furthermore, the bulky side chain of atryptophan reduced viral fitness and could block NTP entry orreduce the size of this channel. V321, located within the hydro-phobic interior of the palm domain, is also a highly conservedresidue, and its replacement with a smaller residue resulted in lossof fitness, while mutation to a larger hydrophobic residue wastolerated.
The crystal structure of JFH-1 NS5B S15G/C223H/V321I un-expectedly showed the same structural rearrangements as thoseobserved previously for an elongation-phase binary complex of a�-hairpin loop deletion polymerase bound to primer-templateRNA, in which the thumb domain rotates around the thumbhinge, accompanied by stretching of the strand that contains S15and flexing of the fingertip domains (7). In comparison with ear-lier structures (12, 15, 19), the �-hairpin loop retracted ca. 3 to 5 Åto generate more space in the active-site cavity in the vicinity of thecatalytic residues, which may allow entry of the 3= end of the viralRNA template and nucleotides for replication initiation; furtherstructure studies aimed at nucleic acid binding to this more openconformation are in progress. It is tempting to speculate that theS15G/C223H/V321I triple variant, and specifically the V321I mu-tation (Fig. 5C and D), may facilitate these rearrangements topromote a catalytically competent conformation for replicationinitiation. However, we cannot rule out the possibility that thecrystal form rather than these three amino acid changes is respon-sible. The current structure appears in the same crystal form asthat reported previously (7), which includes the surface solubili-zation modification E86Q that is involved in crystal lattice con-tacts. Interestingly, our biochemical de novo RNA synthesis assays(Table 6) showed that JFH-1 NS5B S15G/C223H/V321I was ap-proximately 50% more efficient at RNA synthesis than the wildtype. The two rate-limiting steps during de novo RNA synthesishave been identified as the synthesis of the first dinucleotide andthe transition from initiation to elongation, which requires rear-rangement of NS5B (6). Therefore, it is possible that the increasein our de novo RNA synthesis activity resulted from enhancementof a rate-limiting step by the triple-mutant enzyme with a confor-mation compatible with RNA binding (7), which is consistentwith the observation that JFH-1 is able to synthesize the first di-nucleotide and transition to elongation more efficiently thanother isolates (20). The molecular mechanism of how this confor-mation could confer resistance to 2=-F-2=-C-methylguanosinetriphosphates remains unclear. It can be speculated that bindingof the triphosphate analog, the initiation, the transition, and/orthe elongation steps or transitions between them could each beaffected as a result of the changes in NS5B conformation. Thesechanges may not be captured in the current static-state crystalstructure. In addition, because there was a discrepancy in the foldchanges in activity caused by the S15G/C223H/V321I mutation in
the replicon assay (10- to 15-fold) and the biochemical assay (2-fold), interactions with other HCV nonstructural proteins and/orhost factors in the replication complex may be involved in themechanism of resistance and such interactions may be affected bythe conformational changes. The site-directed mutagenesis stud-ies of each of the three amino acids as single or triple amino acidchanges supported the roles of these residues in regulating HCVreplication and the possibility that conformational selectionwithin the NS5B protein could also play a role in conferring resis-tance to 2=-F-2=-C-methylguanosine analogs.
ACKNOWLEDGMENTS
The authors are past employees of Pharmasset except for T.E.E., who is anemployee of Beryllium Discovery Corp. E.M. is a current employee ofGilead Sciences, which acquired Pharmasset.
We thank Todd C. Appleby for discussions on the manuscript.
REFERENCES1. Behrens SE, Tomei L, De Francesco R. 1996. Identification and proper-
ties of the RNA-dependent RNA polymerase of hepatitis C virus. EMBO J.15:12–22.
2. Ago H, Adachi T, Yoshida A, Yamamoto M, Habuka N, Yatsunami K,Miyano M. 1999. Crystal structure of the RNA-dependent RNA polymer-ase of hepatitis C virus. Structure 7:1417–1426. http://dx.doi.org/10.1016/S0969-2126(00)80031-3.
3. Bressanelli S, Tomei L, Roussel A, Incitti I, Vitale RL, Mathieu M, DeFrancesco R, Rey FA. 1999. Crystal structure of the RNA-dependent RNApolymerase of hepatitis C virus. Proc. Natl. Acad. Sci. U. S. A. 96:13034 –13039. http://dx.doi.org/10.1073/pnas.96.23.13034.
4. Lesburg CA, Cable MB, Ferrari E, Hong Z, Mannarino AF, Weber PC.1999. Crystal structure of the RNA-dependent RNA polymerase fromhepatitis C virus reveals a fully encircled active site. Nat. Struct. Biol.6:937–943. http://dx.doi.org/10.1038/13305.
5. Caillet-Saguy C, Lim SP, Shi PY, Lescar J, Bressanelli S. 2014. Poly-merases of hepatitis C viruses and flaviviruses: structural and mechanisticinsights and drug development. Antiviral Res. 105:8 –16. http://dx.doi.org/10.1016/j.antiviral.2014.02.006.
6. Harrus D, Ahmed-El-Sayed N, Simister PC, Miller S, Triconnet M,Hagedorn CH, Mahias K, Rey FA, Astier-Gin T, Bressanelli S. 2010.Further insights into the roles of GTP and the C terminus of the hepatitisC virus polymerase in the initiation of RNA synthesis. J. Biol. Chem.285:32906 –32918. http://dx.doi.org/10.1074/jbc.M110.151316.
7. Mosley RT, Edwards TE, Murakami E, Lam AM, Grice RL, Du J, SofiaMJ, Furman PA, Otto MJ. 2012. Structure of hepatitis C virus polymerasein complex with primer-template RNA. J. Virol. 86:6503– 6511. http://dx.doi.org/10.1128/JVI.00386-12.
8. Sofia MJ. 2013. Nucleotide prodrugs for the treatment of HCV infection.Adv. Pharmacol. 67:39 –73. http://dx.doi.org/10.1016/B978-0-12-405880-4.00002-0.
9. Lam AM, Espiritu C, Bansal S, Micolochick Steuer HM, Zennou V,Otto MJ, Furman PA. 2011. Hepatitis C virus nucleotide inhibitors PSI-352938 and PSI-353661 exhibit a novel mechanism of resistance requiringmultiple mutations within replicon RNA. J. Virol. 85:12334 –12342. http://dx.doi.org/10.1128/JVI.05639-11.
10. Lam AM, Frick DN. 2006. Hepatitis C virus subgenomic replicon re-quires an active NS3 RNA helicase. J. Virol. 80:404 – 411. http://dx.doi.org/10.1128/JVI.80.1.404-411.2006.
11. Reigadas S, Ventura M, Sarih-Cottin L, Castroviejo M, Litvak S, Astier-Gin T. 2001. HCV RNA-dependent RNA polymerase replicates in vitrothe 3= terminal region of the minus-strand viral RNA more efficiently thanthe 3= terminal region of the plus RNA. Eur. J. Biochem. 268:5857–5867.http://dx.doi.org/10.1046/j.0014-2956.2001.02532.x.
12. Simister P, Schmitt M, Geitmann M, Wicht O, Danielson UH, Klein R,Bressanelli S, Lohmann V. 2009. Structural and functional analysis ofhepatitis C virus strain JFH1 polymerase. J. Virol. 83:11926 –11939. http://dx.doi.org/10.1128/JVI.01008-09.
13. Kabsch W. 2010. XDS. Acta Crystallogr. D Biol. Crystallogr. 66(Pt 2):125–132. http://dx.doi.org/10.1107/S0907444909047337.
14. McCoy AJ, Grosse-Kunstleve RW, Adams PD, Winn MD, Storoni LC,
Lam et al.
6868 aac.asm.org Antimicrobial Agents and Chemotherapy
on June 27, 2018 by guesthttp://aac.asm
.org/D
ownloaded from

Read RJ. 2007. Phaser crystallographic software. J. Appl. Crystallogr. 40:658 – 674. http://dx.doi.org/10.1107/S0021889807021206.
15. Biswal BK, Cherney MM, Wang M, Chan L, Yannopoulos CG, Bilimo-ria D, Nicolas O, Bedard J, James MN. 2005. Crystal structures of theRNA-dependent RNA polymerase genotype 2a of hepatitis C virus revealtwo conformations and suggest mechanisms of inhibition by non-nucleoside inhibitors. J. Biol. Chem. 280:18202–18210. http://dx.doi.org/10.1074/jbc.M413410200.
16. Collaborative Computational Project N. 1994. The CCP4 suite: pro-grams for protein crystallography. Acta Crystallogr. D Biol. Crystallogr.50:760 –763. http://dx.doi.org/10.1107/S0907444994003112.
17. Emsley P, Cowtan K. 2004. Coot: model-building tools for moleculargraphics. Acta Crystallogr. D Biol. Crystallogr. 60(Pt 12 No. 1):2126 –2132. http://dx.doi.org/10.1107/S0907444904019158.
18. Zamyatkin DF, Parra F, Alonso JM, Harki DA, Peterson BR, Grochul-ski P, Ng KK. 2008. Structural insights into mechanisms of catalysis andinhibition in Norwalk virus polymerase. J. Biol. Chem. 283:7705–7712.http://dx.doi.org/10.1074/jbc.M709563200.
19. Schmitt M, Scrima N, Radujkovic D, Caillet-Saguy C, Simister PC,Friebe P, Wicht O, Klein R, Bartenschlager R, Lohmann V, BressanelliS. 2011. A comprehensive structure-function comparison of hepatitis Cvirus strain JFH1 and J6 polymerases reveals a key residue stimulatingreplication in cell culture across genotypes. J. Virol. 85:2565–2581. http://dx.doi.org/10.1128/JVI.02177-10.
20. Scrima N, Caillet-Saguy C, Ventura M, Harrus D, Astier-Gin T, Bres-sanelli S. 2012. Two crucial early steps in RNA synthesis by the hepatitis Cvirus polymerase involve a dual role of residue 405. J. Virol. 86:7107–7117.http://dx.doi.org/10.1128/JVI.00459-12.
Analysis of HCV Polymerase Residues 15, 223, and 321
November 2014 Volume 58 Number 11 aac.asm.org 6869
on June 27, 2018 by guesthttp://aac.asm
.org/D
ownloaded from



![CENTERITY SERVICE PACK FOR CLOUDERA€¦ · OOZIE [roles status] • CLOUDERA ROLES SOLR [roles status] • CLOUDERA ROLES SPARK [roles status] • CLOUDERA ROLES SQOOP [roles status]](https://img.pdfslide.net/doc/110x75/5fc0df6d43307a59a12ae0a7/centerity-service-pack-for-cloudera-oozie-roles-status-a-cloudera-roles-solr.jpg)

























