Embed Size (px)
Citation preview

Molecular Cell, Vol. 5, 671–682, April, 2000, Copyright 2000 by Cell Press
MEX-5 and MEX-6 Functionto Establish Soma/Germline Asymmetryin Early C. elegans Embryos
sperm entry appears to define the posterior pole of theC. elegans embryo. After fertilization, the proteins PAR-1and PAR-2 localize to the posterior cortex of the embryo(Guo and Kemphues, 1995; Boyd et al., 1996) and PAR-3,PKC-3, and PAR-6 localize to the anterior cortex
Charlotte M. Schubert,*† Rueyling Lin,*†‖Corry J. de Vries,‡ Ronald H. A. Plasterk,‡and James R. Priess*†§
*Zoology DepartmentUniversity of Washington
(Etemad-Moghadam et al., 1995; Tabuse et al., 1998;Seattle, Washington 98109Hung and Kemphues, 1999). We refer here to these pro-†Department of Basic Sciencesteins collectively as PAR proteins. The PAR proteinsFred Hutchinson Cancer Research Center andare interdependent, as a mutation in any of several parHoward Hughes Medical Institutegenes can disrupt the localization of all PAR proteinsSeattle, Washington 98109(Etemad-Moghadam et al., 1995; Boyd et al., 1996; Ta-‡Division of Molecular Biologybuse et al., 1998; Hung and Kemphues, 1999). MutationsThe Netherlands Cancer Institutein the par genes also cause marked defects in many, or1066 CX Amsterdamall, of the subsequent asymmetries observed betweenwild-type blastomeres after cell division. These includedifferences in blastomere size, cleavage rate, and in theSummaryexpression of proteins that specify cell fate (reviewedin Rose and Kemphues, 1998). Although PAR asymmetryAn asymmetrical network of cortically localized PARclearly is linked to blastomere differences, the natureproteins forms shortly after fertilization of the C. ele-of this linkage has remained mysterious. Some PARgans egg. This network is required for subsequentproteins contain structural domains that are found inasymmetries in the expression patterns of several pro-diverse cell signaling molecules; PAR-1 and PKC-3, forteins that are encoded by nonlocalized, maternally ex-example, both have putative kinase domains (Guo andpressed mRNAs. We provide evidence that two nearlyKemphues, 1995; Tabuse et al., 1998). The PAR-2 pro-identical genes, mex-5 and mex-6, link PAR asymmetrytein contains a motif called a RING finger (Boyd et al.,to those subsequent protein asymmetries. MEX-5 is a1996), and recent studies have shown that RING fingersnovel, cytoplasmic protein that is localized throughmay function to regulate degradation by adding ubiqui-PAR activities to the anterior pole of the 1-cell stagetin to proteins (Freed et al., 1999; Joazeiro et al., 1999;embryo. MEX-5 localization is reciprocal to that of aLorick et al., 1999; Seol et al., 1999; Waterman et al.,group of posterior-localized proteins called germline1999). However, specific biochemical functions of mostproteins. Ectopic expression of MEX-5 is sufficient toPAR proteins have not been determined, nor have anyinhibit the expression of germline proteins, suggestingtargets of the PAR proteins been identified.that MEX-5 functions to inhibit anterior expression of
One approach toward elucidating the linkage betweenthe germline proteins.
the PAR proteins and the differences between earlyblastomeres is to work upstream from specific proteins
Introduction that show a PAR-dependent distribution. There are threegeneral, PAR-dependent patterns of protein distribition
Embryonic blastomeres become committed to distinct that have been described in the early blastomeres (seefates within the first few cell cycles after fertilization of Figure 1 for diagram). After the first division, anteriorthe C. elegans egg. This rapid diversification occurs proteins are present only in the anterior blastomere;because the egg contains a pool of maternally provided these proteins persist, or continue to be expressed, inmRNAs that encode determinative factors, and the early all the descendants of the anterior blastomere for theblastomeres have markedly different potentials for ex- next few cell cycles (Evans et al., 1994). Posterior pro-pressing these factors. This results in the asymmetric teins are localized to the posterior blastomere after the
first division and are present in all the early descendantsexpression patterns of transcriptional factors that ap-of the posterior blastomere (Bowerman et al., 1993;pear to determine blastomere fates directly, and compo-Hunter and Kenyon, 1996). A third group of proteins isnents of NOTCH-like and WNT/WINGLESS-like signalinglocalized to only a single branch of the descendants ofpathways that allow position-specific cell interactionsthe posterior blastomere (Mello et al., 1996; Guedes and(for general review, see Bowerman and Shelton, 1999).Priess, 1997; Tenenhaus et al., 1998; Tabara et al., 1999).Little is known about the molecular differences be-Blastomeres in this branch eventually form the germlinetween blastomeres that result in asymmetric protein ex-and so are called germline blastomeres to distinguishpression from these maternal mRNAs. However, a criti-them from all other blastomeres that produce only so-cal first step in this process occurs during the 1-cellmatic cell types. We refer here to the proteins that arestage, shortly after fertilization of the egg. The point oflocalized to the germline blastomeres as germline pro-teins.§ To whom correspondence should be addressed (e-mail: jpriess@
Almost all of the mRNAs encoding anterior, posterior,fred.fhcrc.org).and germline proteins are distributed uniformly through-‖ Present address: Department of Molecular Biology and Oncology,out the early embryo (Evans et al., 1994; Seydoux andUniversity of Texas Southwestern Medical Center, Dallas, Texas
75395. Fire, 1994; Guedes and Priess, 1997; Tenenhaus et al.,

Molecular Cell672
embryonic lethal mutants, only par-1 mutants werefound to completely abolish PIE-1 asymmetry (Tenen-haus et al., 1998). Although there are several possibleexplanations for these results, they are consistent withthe view that there may be only a few factors that linkPAR asymmetry to protein localization. In the presentstudy, we provide evidence that two closely related pro-teins, MEX-5 and MEX-6, are part of this linkage.
Results
The posterior protein SKN-1 is a transcription factorrequired for the development of muscles in the pharynx,among other cell types. Mutations in any of several pargenes, or in a gene called mex-1, result in misexpressionof SKN-1 in anterior blastomeres; such mutants haveabnormally large numbers of muscles (see Bowermanand Shelton, 1999). We performed a genetic screen formaternal effect lethal mutants with this muscle excess(mex) phenotype (see Experimental Procedures). In ad-Figure 1. Protein Expression Patterns in Early Embryosdition to alleles of the par and mex-1 genes, we identifiedA lineage diagram of the first few embryonic divisions is shown onan allele, zu199, of a novel gene we call mex-5. Embryosthe left. Lineage branches expressing anterior proteins are shown
in red, posterior proteins in purple, and germline proteins in green. from homozygous mex-5(zu199) adults (hereafter re-The successive blastomeres in the germline branch are named P0 ferred to as mex-5 embryos) produce approximately the(the 1-cell embryo), P1, P2, P3, and P4. Schematic drawings of the wild-type number of cells but do not undergo body mor-early blastomeres are shown on the right; sister blastomeres are phogenesis and die without hatching (Figure 2C). Theindicated by dashes. Each germline blastomere is green, and its
mex-5 embryos contain abnormally large numbers ofsomatic sister is light green. For comparison with later figures, themuscles toward their anterior poles (Figure 2D; comparegermline and somatic sisters are indicated by arrowheads. At thewith wild-type embryo in Figure 2B). In cell isolationbeginning of the 4-cell stage, one of the two anterior (white) blasto-
meres is forced toward the posterior by the surrounding eggshell. experiments on 2-cell stage mex-5 embryos, we foundThis movement results in the two anterior and two posterior blasto- that the anterior blastomere produced muscles inappro-meres adopting the rhombohedral configuration shown. priately (14/14 cases). In contrast, the anterior blasto-
mere from wild-type embryos, or from skn-1(zu67);mex-1998). Therefore, protein asymmetry must result from 5(zu199) embryos, did not produce muscles in similarprotein movement, or from differences in either protein experiments (0/11 and 0/20, respectively). Consistentstability or mRNA translation. There is suggestive evi- with these results, we found that 2-cell stage mex-5dence that localization of the germline proteins PIE-1, embryos misexpress the posterior transcription factorMEX-1, and POS-1 may, at least in part, involve protein SKN-1 at high levels in the anterior blastomere (Figuredegradation. As a germline blastomere divides, high lev- 2H; compare with wild-type in Figure 2G). In this defect,els of the germline proteins are inherited by the germline mex-5 embryos resemble the embryos from par mutantsdaughter, and either low (PIE-1; MEX-1) or high (POS-1) or mex-1 mutants (Bowerman et al., 1993, 1997). How-levels are inherited by the somatic daughter (Mello et ever, mex-5 embryos have normal early cleavage planesal., 1996; Guedes and Priess, 1997; Tenenhaus et al., and cell cycles, distinguishing them from par embryos1998; Tabara et al., 1999). These proteins persist at high (data not shown). In addition, germ cells were visible inlevels in the germline daughter during the cell cycle but almost all mex-5 embryos (15/17 embryos; Figure 2C,rapidly disappear from the somatic daughter (Mello et white arrows), while par embryos and mex-1 embryosal., 1996; Guedes and Priess, 1997; Tenenhaus et al., do not produce germ cells (see Bowerman and Shelton,1998; Tabara et al., 1999). 1999). Thus, the mex-5 gene appears to be a novel com-
The localization of the anterior protein GLP-1 is due ponent of the pathways that establish asymmetrical pat-in part to a difference between the anterior and posterior terns of protein expression in the early embryo.blastomeres in their ability to translate the glp-1 mRNA(Evans et al., 1994). A reporter construct containing only MEX-5 Shares a CCCH Motif with Germline Proteinsthe 39 untranslated region (UTR) from the glp-1 mRNA We cloned the mex-5 gene as described in Experimental
Procedures. In brief, mex-5(zu199) maps to a region onis expressed exclusively in anterior blastomeres. Dele-tion of a small region within the 39UTR results in ectopic chromosome V near the rescuing cosmid W02A2; RNA-
mediated inhibition of the predicted gene W02A2.7expression in posterior blastomeres. These results sug-gest that posterior blastomeres contain a trans-acting causes wild-type adults to produce inviable embryos
that are indistinguishable from mex-5 embryos; theinhibitor of translation. Thus, it is possible that PARproteins could modify factors involved in either transla- W02A2.7 gene in mex-5(zu199) animals contains a non-
sense mutation that would truncate the predicted pro-tion or protein degradation.A previous genetic screen to identify regulators of tein product (Figure 3A); and a monoclonal antibody
generated against a W02A2.7 fusion protein stains wild-GLP-1 localization yielded only alleles of known pargenes, and a candidate for a new par gene (Crittenden type embryos (see below), but not mex-5(zu199) em-
bryos. We henceforth name W02A2.7 the gene mex-5,et al., 1997). Similarly, in a survey of previously identified
MEX-5, MEX-6, and Soma/Germline Asymmetry673
POS-1 each contain two finger domains but do not ap-pear to have other similarity to MEX-5 (Figure 3B).
MEX-5 and MEX-6 Have Overlapping FunctionsDatabase searches with the mex-5 sequence showedthat the C. elegans genome contains a predicted genesequence, AH6.5, that could encode a protein with veryhigh similarity to MEX-5; we have named this proteinMEX-6. MEX-6 is about 70% identical and 85% similarto MEX-5 in amino acid sequence (Figure 3A). The mex-6and mex-5 genes contain similarly placed introns andshare extensive nucleic acid identity in 59 and 39 untrans-lated regions and in intron sequences (data not shown).Thus, mex-5 and mex-6 are nearly identical genes thatare likely to have arisen from a relatively recent geneduplication.
We used the technique of double-stranded RNA-medi-ated inhibition (RNAi) to assay potential functions ofMEX-5 and MEX-6. When wild-type larvae were treatedwith mex-5 dsRNA, the resulting adults produced em-bryos (called mex-5(RNAi) embryos) that were inviableand appeared indistinguishable from mex-5(zu199) em-bryos by light microscopy (0% embryonic viability, n 583). In contrast, adults treated with mex-6 dsRNA pro-duced viable embryos that hatched and grew into fertileadults (100% embryonic viability, n 5 277). We identifieda deletion in the mex-6 gene by a PCR-based screeningmethod (Jansen et al., 1997). The mex-6(pk440) allele
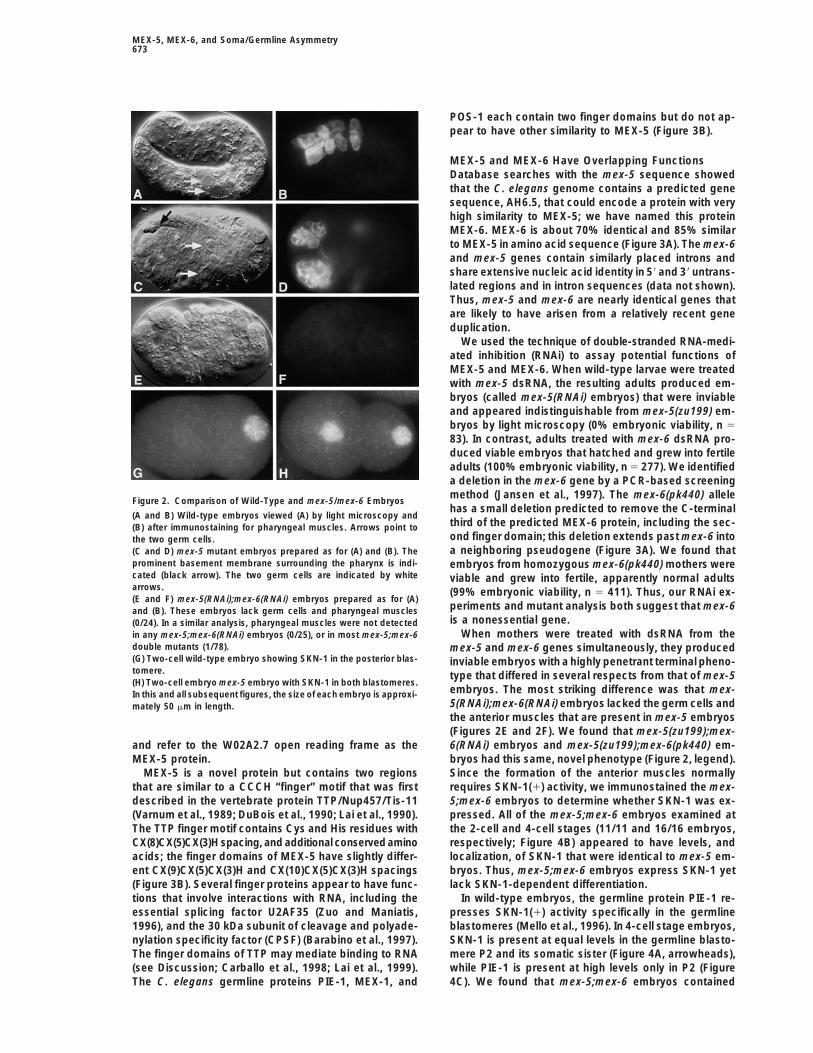
Figure 2. Comparison of Wild-Type and mex-5/mex-6 Embryoshas a small deletion predicted to remove the C-terminal
(A and B) Wild-type embryos viewed (A) by light microscopy andthird of the predicted MEX-6 protein, including the sec-(B) after immunostaining for pharyngeal muscles. Arrows point toond finger domain; this deletion extends past mex-6 intothe two germ cells.a neighboring pseudogene (Figure 3A). We found that(C and D) mex-5 mutant embryos prepared as for (A) and (B). The
prominent basement membrane surrounding the pharynx is indi- embryos from homozygous mex-6(pk440) mothers werecated (black arrow). The two germ cells are indicated by white viable and grew into fertile, apparently normal adultsarrows. (99% embryonic viability, n 5 411). Thus, our RNAi ex-(E and F) mex-5(RNAi);mex-6(RNAi) embryos prepared as for (A)
periments and mutant analysis both suggest that mex-6and (B). These embryos lack germ cells and pharyngeal musclesis a nonessential gene.(0/24). In a similar analysis, pharyngeal muscles were not detected
When mothers were treated with dsRNA from thein any mex-5;mex-6(RNAi) embryos (0/25), or in most mex-5;mex-6double mutants (1/78). mex-5 and mex-6 genes simultaneously, they produced(G) Two-cell wild-type embryo showing SKN-1 in the posterior blas- inviable embryos with a highly penetrant terminal pheno-tomere. type that differed in several respects from that of mex-5(H) Two-cell embryo mex-5 embryo with SKN-1 in both blastomeres.
embryos. The most striking difference was that mex-In this and all subsequent figures, the size of each embryo is approxi-5(RNAi);mex-6(RNAi) embryos lacked the germ cells andmately 50 mm in length.the anterior muscles that are present in mex-5 embryos(Figures 2E and 2F). We found that mex-5(zu199);mex-6(RNAi) embryos and mex-5(zu199);mex-6(pk440) em-and refer to the W02A2.7 open reading frame as the
MEX-5 protein. bryos had this same, novel phenotype (Figure 2, legend).Since the formation of the anterior muscles normallyMEX-5 is a novel protein but contains two regions
that are similar to a CCCH “finger” motif that was first requires SKN-1(1) activity, we immunostained the mex-5;mex-6 embryos to determine whether SKN-1 was ex-described in the vertebrate protein TTP/Nup457/Tis-11
(Varnum et al., 1989; DuBois et al., 1990; Lai et al., 1990). pressed. All of the mex-5;mex-6 embryos examined atthe 2-cell and 4-cell stages (11/11 and 16/16 embryos,The TTP finger motif contains Cys and His residues with
CX(8)CX(5)CX(3)H spacing, and additional conservedamino respectively; Figure 4B) appeared to have levels, andlocalization, of SKN-1 that were identical to mex-5 em-acids; the finger domains of MEX-5 have slightly differ-
ent CX(9)CX(5)CX(3)H and CX(10)CX(5)CX(3)H spacings bryos. Thus, mex-5;mex-6 embryos express SKN-1 yetlack SKN-1-dependent differentiation.(Figure 3B). Several finger proteins appear to have func-
tions that involve interactions with RNA, including the In wild-type embryos, the germline protein PIE-1 re-presses SKN-1(1) activity specifically in the germlineessential splicing factor U2AF35 (Zuo and Maniatis,
1996), and the 30 kDa subunit of cleavage and polyade- blastomeres (Mello et al., 1996). In 4-cell stage embryos,SKN-1 is present at equal levels in the germline blasto-nylation specificity factor (CPSF) (Barabino et al., 1997).
The finger domains of TTP may mediate binding to RNA mere P2 and its somatic sister (Figure 4A, arrowheads),while PIE-1 is present at high levels only in P2 (Figure(see Discussion; Carballo et al., 1998; Lai et al., 1999).
The C. elegans germline proteins PIE-1, MEX-1, and 4C). We found that mex-5;mex-6 embryos contained

Molecular Cell674
Figure 3. MEX-5 and MEX-6
(A) Alignment of the predicted MEX-5 andMEX-6 proteins. The CCCH finger domainsare highlighted in gray. A point mutation in themex-5(zu199) allele introduces a stop codon(UAA) that is predicted to truncate the proteinbefore the first finger (arrowhead). The 59
breakpoint of the mex-6(pk440) deficiency isindicated with an arrow. Amino acid identitiesbetween MEX-5 and MEX-6 are indicated bydouble dots, similarities with single dots.(B) Alignment of the CCCH fingers. The topand bottom panels compare the first and sec-ond CCCH fingers of MEX-5 and MEX-6 withthe corresponding fingers in PIE-1, POS-1,MEX-1 (C. elegans), and TTP (mouse). Aminoacids shared with MEX-5 are highlighted ingray.
ectopic PIE-1: between the 1-cell stage and the 44-cell that the germline proteins MEX-1 and POS-1 were ex-pressed inappropriately in all blastomeres in mex-stages, PIE-1 was distributed uniformly in all blasto-
meres (Figure 4D). We constructed and examined pie- 5;mex-6 mutants (Figures 4F and 4H, respectively). Inwild-type embryos, cytoplasmic granules called P gran-1(RNAi);mex-5(RNAi);mex-6(RNAi) embryos and found
that these embryos produced cell types that normally ules are segregated asymmetrically to each germlineblastomere; in mex-5;mex-6 embryos, these granulesrequire SKN-1(1) function. For example, while mex-
5(RNAi);mex-6(RNAi) embryos lack intestinal cells (0/24 were present in all blastomeres (data not shown). Thetranscription factor PAL-1 is a posterior, nuclear proteinembryos), the pie-1(RNAi);mex-5(RNAi);mex-6(RNAi)
embryos contained numerous intestinal cells (54/54 em- in wild-type embryos (Figure 4I) but was present in allnuclei in mex-5;mex-6 embryos (Figure 4J). In wild-typebryos). Thus, PIE-1 is mislocalized in mex-5;mex-6 mu-
tants and appears to repress the activity of SKN-1. embryos, anterior expression of PAL-1 appears to beprevented, at least in part, by high levels of the MEX-3We immunostained mex-5;mex-6 embryos to examine
other proteins that, like SKN-1 and PIE-1, are asymmetri- protein in anterior blastomeres (Figure 4K; Draper et al.,1996; Hunter and Kenyon, 1996). We found that MEX-3cally localized in wild-type embryos (Figure 4). We found
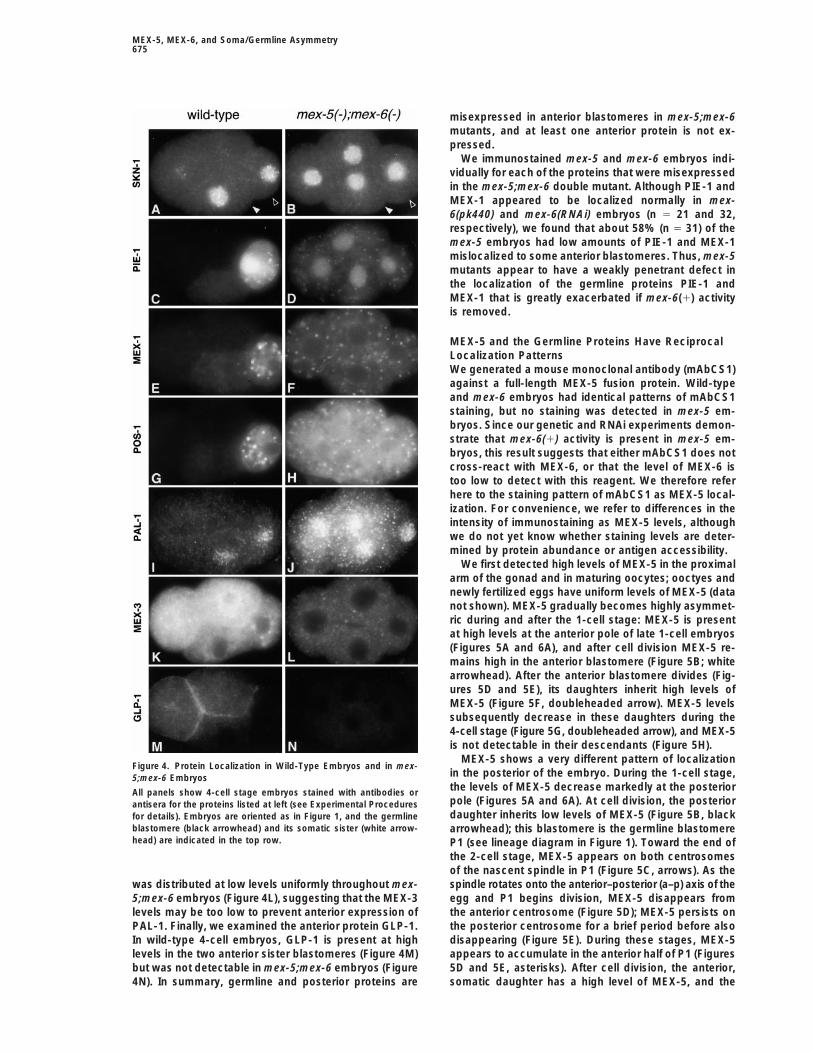
MEX-5, MEX-6, and Soma/Germline Asymmetry675
misexpressed in anterior blastomeres in mex-5;mex-6mutants, and at least one anterior protein is not ex-pressed.
We immunostained mex-5 and mex-6 embryos indi-vidually for each of the proteins that were misexpressedin the mex-5;mex-6 double mutant. Although PIE-1 andMEX-1 appeared to be localized normally in mex-6(pk440) and mex-6(RNAi) embryos (n 5 21 and 32,respectively), we found that about 58% (n 5 31) of themex-5 embryos had low amounts of PIE-1 and MEX-1mislocalized to some anterior blastomeres. Thus, mex-5mutants appear to have a weakly penetrant defect inthe localization of the germline proteins PIE-1 andMEX-1 that is greatly exacerbated if mex-6(1) activityis removed.
MEX-5 and the Germline Proteins Have ReciprocalLocalization PatternsWe generated a mouse monoclonal antibody (mAbCS1)against a full-length MEX-5 fusion protein. Wild-typeand mex-6 embryos had identical patterns of mAbCS1staining, but no staining was detected in mex-5 em-bryos. Since our genetic and RNAi experiments demon-strate that mex-6(1) activity is present in mex-5 em-bryos, this result suggests that either mAbCS1 does notcross-react with MEX-6, or that the level of MEX-6 istoo low to detect with this reagent. We therefore referhere to the staining pattern of mAbCS1 as MEX-5 local-ization. For convenience, we refer to differences in theintensity of immunostaining as MEX-5 levels, althoughwe do not yet know whether staining levels are deter-mined by protein abundance or antigen accessibility.
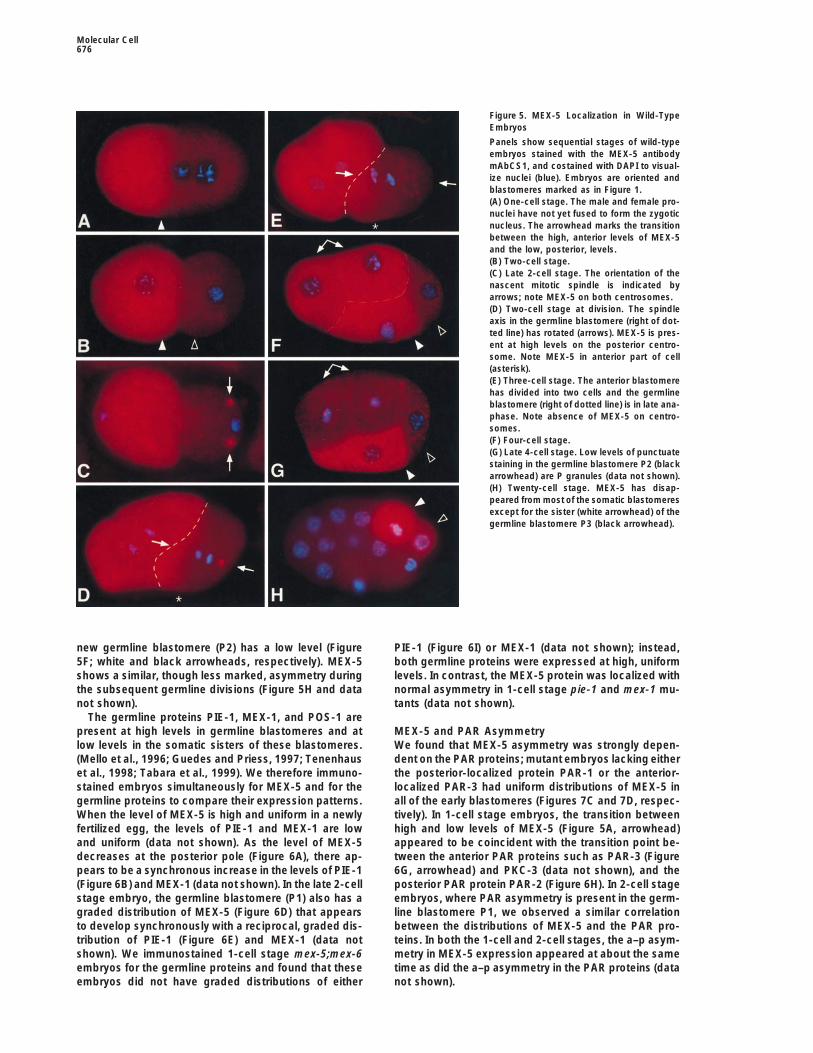
We first detected high levels of MEX-5 in the proximalarm of the gonad and in maturing oocytes; ooctyes andnewly fertilized eggs have uniform levels of MEX-5 (datanot shown). MEX-5 gradually becomes highly asymmet-ric during and after the 1-cell stage: MEX-5 is presentat high levels at the anterior pole of late 1-cell embryos(Figures 5A and 6A), and after cell division MEX-5 re-mains high in the anterior blastomere (Figure 5B; whitearrowhead). After the anterior blastomere divides (Fig-ures 5D and 5E), its daughters inherit high levels ofMEX-5 (Figure 5F, doubleheaded arrow). MEX-5 levelssubsequently decrease in these daughters during the4-cell stage (Figure 5G, doubleheaded arrow), and MEX-5is not detectable in their descendants (Figure 5H).
MEX-5 shows a very different pattern of localizationFigure 4. Protein Localization in Wild-Type Embryos and in mex-
in the posterior of the embryo. During the 1-cell stage,5;mex-6 Embryosthe levels of MEX-5 decrease markedly at the posteriorAll panels show 4-cell stage embryos stained with antibodies orpole (Figures 5A and 6A). At cell division, the posteriorantisera for the proteins listed at left (see Experimental Proceduresdaughter inherits low levels of MEX-5 (Figure 5B, blackfor details). Embryos are oriented as in Figure 1, and the germline
blastomere (black arrowhead) and its somatic sister (white arrow- arrowhead); this blastomere is the germline blastomerehead) are indicated in the top row. P1 (see lineage diagram in Figure 1). Toward the end of
the 2-cell stage, MEX-5 appears on both centrosomesof the nascent spindle in P1 (Figure 5C, arrows). As thespindle rotates onto the anterior–posterior (a–p) axis of thewas distributed at low levels uniformly throughout mex-
5;mex-6 embryos (Figure 4L), suggesting that the MEX-3 egg and P1 begins division, MEX-5 disappears fromthe anterior centrosome (Figure 5D); MEX-5 persists onlevels may be too low to prevent anterior expression of
PAL-1. Finally, we examined the anterior protein GLP-1. the posterior centrosome for a brief period before alsodisappearing (Figure 5E). During these stages, MEX-5In wild-type 4-cell embryos, GLP-1 is present at high
levels in the two anterior sister blastomeres (Figure 4M) appears to accumulate in the anterior half of P1 (Figures5D and 5E, asterisks). After cell division, the anterior,but was not detectable in mex-5;mex-6 embryos (Figure
4N). In summary, germline and posterior proteins are somatic daughter has a high level of MEX-5, and the
Molecular Cell676
Figure 5. MEX-5 Localization in Wild-TypeEmbryos
Panels show sequential stages of wild-typeembryos stained with the MEX-5 antibodymAbCS1, and costained with DAPI to visual-ize nuclei (blue). Embryos are oriented andblastomeres marked as in Figure 1.(A) One-cell stage. The male and female pro-nuclei have not yet fused to form the zygoticnucleus. The arrowhead marks the transitionbetween the high, anterior levels of MEX-5and the low, posterior, levels.(B) Two-cell stage.(C) Late 2-cell stage. The orientation of thenascent mitotic spindle is indicated byarrows; note MEX-5 on both centrosomes.(D) Two-cell stage at division. The spindleaxis in the germline blastomere (right of dot-ted line) has rotated (arrows). MEX-5 is pres-ent at high levels on the posterior centro-some. Note MEX-5 in anterior part of cell(asterisk).(E) Three-cell stage. The anterior blastomerehas divided into two cells and the germlineblastomere (right of dotted line) is in late ana-phase. Note absence of MEX-5 on centro-somes.(F) Four-cell stage.(G) Late 4-cell stage. Low levels of punctuatestaining in the germline blastomere P2 (blackarrowhead) are P granules (data not shown).(H) Twenty-cell stage. MEX-5 has disap-peared from most of the somatic blastomeresexcept for the sister (white arrowhead) of thegermline blastomere P3 (black arrowhead).
new germline blastomere (P2) has a low level (Figure PIE-1 (Figure 6I) or MEX-1 (data not shown); instead,both germline proteins were expressed at high, uniform5F; white and black arrowheads, respectively). MEX-5
shows a similar, though less marked, asymmetry during levels. In contrast, the MEX-5 protein was localized withnormal asymmetry in 1-cell stage pie-1 and mex-1 mu-the subsequent germline divisions (Figure 5H and data
not shown). tants (data not shown).The germline proteins PIE-1, MEX-1, and POS-1 are
present at high levels in germline blastomeres and at MEX-5 and PAR AsymmetryWe found that MEX-5 asymmetry was strongly depen-low levels in the somatic sisters of these blastomeres.
(Mello et al., 1996; Guedes and Priess, 1997; Tenenhaus dent on the PAR proteins; mutant embryos lacking eitherthe posterior-localized protein PAR-1 or the anterior-et al., 1998; Tabara et al., 1999). We therefore immuno-
stained embryos simultaneously for MEX-5 and for the localized PAR-3 had uniform distributions of MEX-5 inall of the early blastomeres (Figures 7C and 7D, respec-germline proteins to compare their expression patterns.
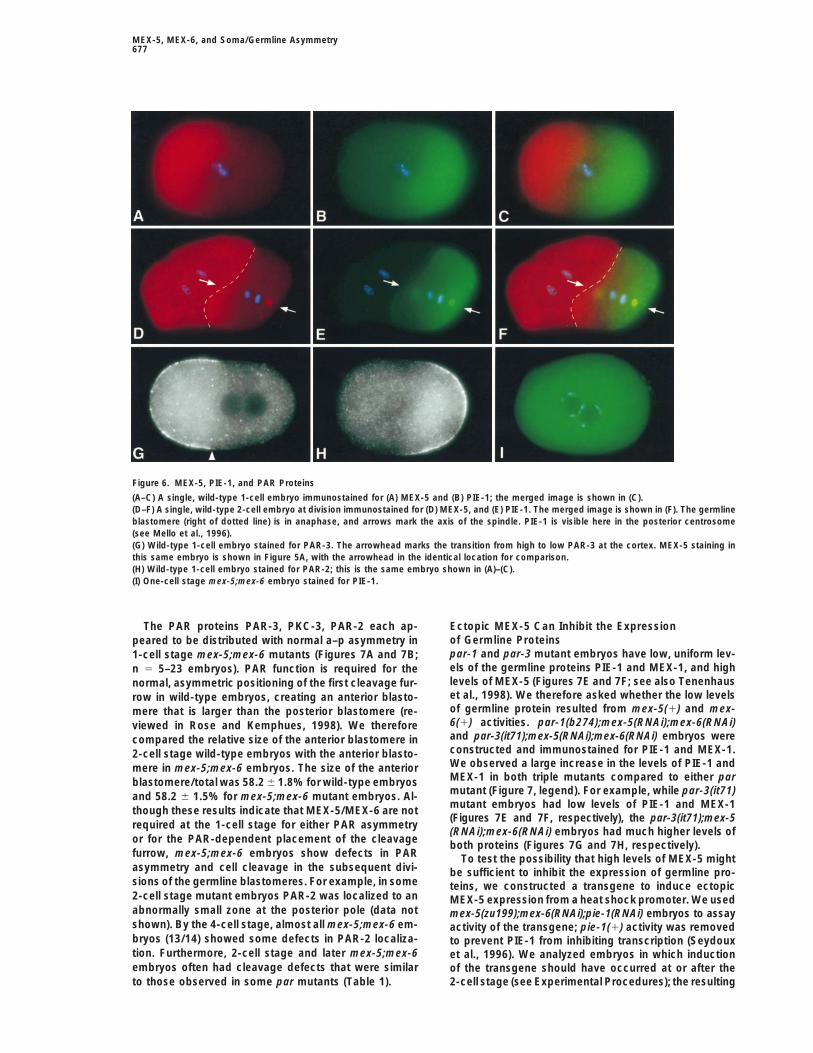
When the level of MEX-5 is high and uniform in a newly tively). In 1-cell stage embryos, the transition betweenhigh and low levels of MEX-5 (Figure 5A, arrowhead)fertilized egg, the levels of PIE-1 and MEX-1 are low
and uniform (data not shown). As the level of MEX-5 appeared to be coincident with the transition point be-tween the anterior PAR proteins such as PAR-3 (Figuredecreases at the posterior pole (Figure 6A), there ap-
pears to be a synchronous increase in the levels of PIE-1 6G, arrowhead) and PKC-3 (data not shown), and theposterior PAR protein PAR-2 (Figure 6H). In 2-cell stage(Figure 6B) and MEX-1 (data not shown). In the late 2-cell
stage embryo, the germline blastomere (P1) also has a embryos, where PAR asymmetry is present in the germ-line blastomere P1, we observed a similar correlationgraded distribution of MEX-5 (Figure 6D) that appears
to develop synchronously with a reciprocal, graded dis- between the distributions of MEX-5 and the PAR pro-teins. In both the 1-cell and 2-cell stages, the a–p asym-tribution of PIE-1 (Figure 6E) and MEX-1 (data not
shown). We immunostained 1-cell stage mex-5;mex-6 metry in MEX-5 expression appeared at about the sametime as did the a–p asymmetry in the PAR proteins (dataembryos for the germline proteins and found that these
embryos did not have graded distributions of either not shown).
MEX-5, MEX-6, and Soma/Germline Asymmetry677
Figure 6. MEX-5, PIE-1, and PAR Proteins
(A–C) A single, wild-type 1-cell embryo immunostained for (A) MEX-5 and (B) PIE-1; the merged image is shown in (C).(D–F) A single, wild-type 2-cell embryo at division immunostained for (D) MEX-5, and (E) PIE-1. The merged image is shown in (F). The germlineblastomere (right of dotted line) is in anaphase, and arrows mark the axis of the spindle. PIE-1 is visible here in the posterior centrosome(see Mello et al., 1996).(G) Wild-type 1-cell embryo stained for PAR-3. The arrowhead marks the transition from high to low PAR-3 at the cortex. MEX-5 staining inthis same embryo is shown in Figure 5A, with the arrowhead in the identical location for comparison.(H) Wild-type 1-cell embryo stained for PAR-2; this is the same embryo shown in (A)–(C).(I) One-cell stage mex-5;mex-6 embryo stained for PIE-1.
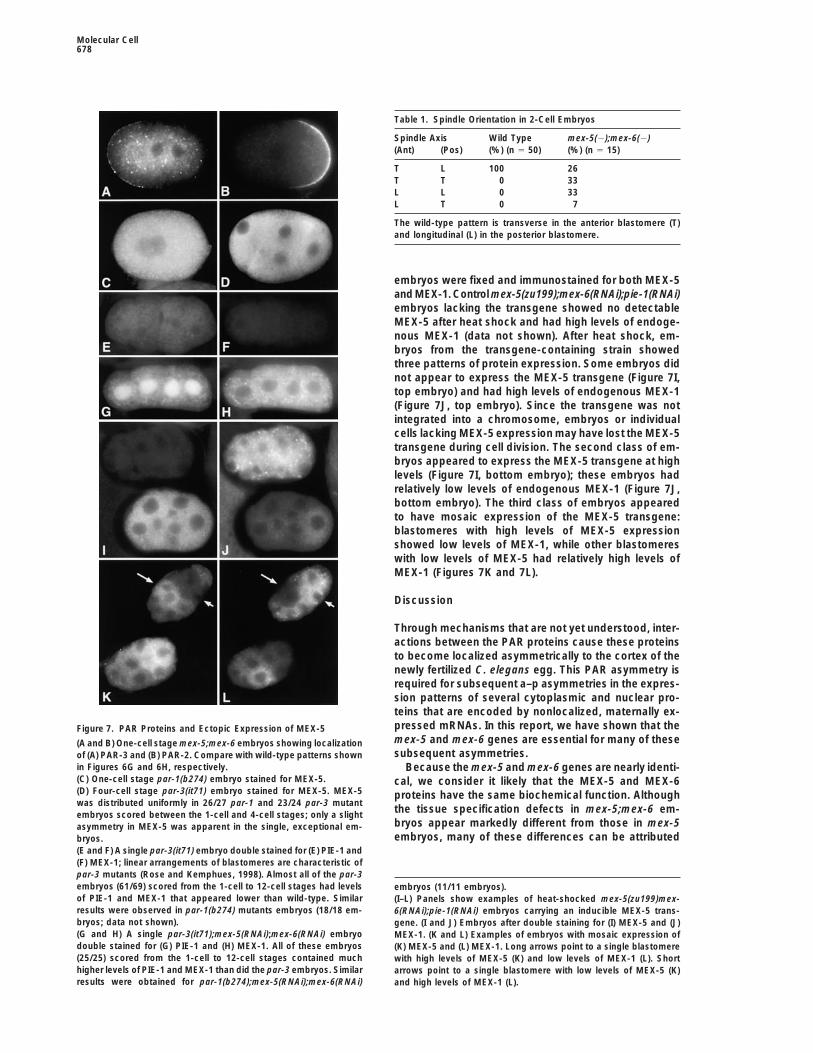
The PAR proteins PAR-3, PKC-3, PAR-2 each ap- Ectopic MEX-5 Can Inhibit the Expressionof Germline Proteinspeared to be distributed with normal a–p asymmetry inpar-1 and par-3 mutant embryos have low, uniform lev-1-cell stage mex-5;mex-6 mutants (Figures 7A and 7B;els of the germline proteins PIE-1 and MEX-1, and highn 5 5–23 embryos). PAR function is required for thelevels of MEX-5 (Figures 7E and 7F; see also Tenenhausnormal, asymmetric positioning of the first cleavage fur-et al., 1998). We therefore asked whether the low levelsrow in wild-type embryos, creating an anterior blasto-of germline protein resulted from mex-5(1) and mex-mere that is larger than the posterior blastomere (re-6(1) activities. par-1(b274);mex-5(RNAi);mex-6(RNAi)viewed in Rose and Kemphues, 1998). We thereforeand par-3(it71);mex-5(RNAi);mex-6(RNAi) embryos werecompared the relative size of the anterior blastomere inconstructed and immunostained for PIE-1 and MEX-1.2-cell stage wild-type embryos with the anterior blasto-We observed a large increase in the levels of PIE-1 andmere in mex-5;mex-6 embryos. The size of the anteriorMEX-1 in both triple mutants compared to either parblastomere/total was 58.2 6 1.8% for wild-type embryosmutant (Figure 7, legend). For example, while par-3(it71)and 58.2 6 1.5% for mex-5;mex-6 mutant embryos. Al-mutant embryos had low levels of PIE-1 and MEX-1though these results indicate that MEX-5/MEX-6 are not(Figures 7E and 7F, respectively), the par-3(it71);mex-5
required at the 1-cell stage for either PAR asymmetry(RNAi);mex-6(RNAi) embryos had much higher levels of
or for the PAR-dependent placement of the cleavage both proteins (Figures 7G and 7H, respectively).furrow, mex-5;mex-6 embryos show defects in PAR To test the possibility that high levels of MEX-5 mightasymmetry and cell cleavage in the subsequent divi- be sufficient to inhibit the expression of germline pro-sions of the germline blastomeres. For example, in some teins, we constructed a transgene to induce ectopic2-cell stage mutant embryos PAR-2 was localized to an MEX-5 expression from a heat shock promoter. We usedabnormally small zone at the posterior pole (data not mex-5(zu199);mex-6(RNAi);pie-1(RNAi) embryos to assayshown). By the 4-cell stage, almost all mex-5;mex-6 em- activity of the transgene; pie-1(1) activity was removedbryos (13/14) showed some defects in PAR-2 localiza- to prevent PIE-1 from inhibiting transcription (Seydouxtion. Furthermore, 2-cell stage and later mex-5;mex-6 et al., 1996). We analyzed embryos in which inductionembryos often had cleavage defects that were similar of the transgene should have occurred at or after the
2-cell stage (see Experimental Procedures); the resultingto those observed in some par mutants (Table 1).
Molecular Cell678
Table 1. Spindle Orientation in 2-Cell Embryos
Spindle Axis Wild Type mex-5(2);mex-6(2)(Ant) (Pos) (%) (n 5 50) (%) (n 5 15)
T L 100 26T T 0 33L L 0 33L T 0 7
The wild-type pattern is transverse in the anterior blastomere (T)and longitudinal (L) in the posterior blastomere.
embryos were fixed and immunostained for both MEX-5and MEX-1. Control mex-5(zu199);mex-6(RNAi);pie-1(RNAi)embryos lacking the transgene showed no detectableMEX-5 after heat shock and had high levels of endoge-nous MEX-1 (data not shown). After heat shock, em-bryos from the transgene-containing strain showedthree patterns of protein expression. Some embryos didnot appear to express the MEX-5 transgene (Figure 7I,top embryo) and had high levels of endogenous MEX-1(Figure 7J, top embryo). Since the transgene was notintegrated into a chromosome, embryos or individualcells lacking MEX-5 expression may have lost the MEX-5transgene during cell division. The second class of em-bryos appeared to express the MEX-5 transgene at highlevels (Figure 7I, bottom embryo); these embryos hadrelatively low levels of endogenous MEX-1 (Figure 7J,bottom embryo). The third class of embryos appearedto have mosaic expression of the MEX-5 transgene:blastomeres with high levels of MEX-5 expressionshowed low levels of MEX-1, while other blastomereswith low levels of MEX-5 had relatively high levels ofMEX-1 (Figures 7K and 7L).
Discussion
Through mechanisms that are not yet understood, inter-actions between the PAR proteins cause these proteinsto become localized asymmetrically to the cortex of thenewly fertilized C. elegans egg. This PAR asymmetry isrequired for subsequent a–p asymmetries in the expres-sion patterns of several cytoplasmic and nuclear pro-teins that are encoded by nonlocalized, maternally ex-pressed mRNAs. In this report, we have shown that theFigure 7. PAR Proteins and Ectopic Expression of MEX-5mex-5 and mex-6 genes are essential for many of these(A and B) One-cell stage mex-5;mex-6 embryos showing localizationsubsequent asymmetries.of (A) PAR-3 and (B) PAR-2. Compare with wild-type patterns shown
in Figures 6G and 6H, respectively. Because the mex-5 and mex-6 genes are nearly identi-(C) One-cell stage par-1(b274) embryo stained for MEX-5. cal, we consider it likely that the MEX-5 and MEX-6(D) Four-cell stage par-3(it71) embryo stained for MEX-5. MEX-5 proteins have the same biochemical function. Althoughwas distributed uniformly in 26/27 par-1 and 23/24 par-3 mutant
the tissue specification defects in mex-5;mex-6 em-embryos scored between the 1-cell and 4-cell stages; only a slightbryos appear markedly different from those in mex-5asymmetry in MEX-5 was apparent in the single, exceptional em-embryos, many of these differences can be attributedbryos.
(E and F) A single par-3(it71) embryo double stained for (E) PIE-1 and(F) MEX-1; linear arrangements of blastomeres are characteristic ofpar-3 mutants (Rose and Kemphues, 1998). Almost all of the par-3embryos (61/69) scored from the 1-cell to 12-cell stages had levels embryos (11/11 embryos).of PIE-1 and MEX-1 that appeared lower than wild-type. Similar (I–L) Panels show examples of heat-shocked mex-5(zu199)mex-results were observed in par-1(b274) mutants embryos (18/18 em- 6(RNAi);pie-1(RNAi) embryos carrying an inducible MEX-5 trans-bryos; data not shown). gene. (I and J) Embryos after double staining for (I) MEX-5 and (J)(G and H) A single par-3(it71);mex-5(RNAi);mex-6(RNAi) embryo MEX-1. (K and L) Examples of embryos with mosaic expression ofdouble stained for (G) PIE-1 and (H) MEX-1. All of these embryos (K) MEX-5 and (L) MEX-1. Long arrows point to a single blastomere(25/25) scored from the 1-cell to 12-cell stages contained much with high levels of MEX-5 (K) and low levels of MEX-1 (L). Shorthigher levels of PIE-1 and MEX-1 than did the par-3 embryos. Similar arrows point to a single blastomere with low levels of MEX-5 (K)results were obtained for par-1(b274);mex-5(RNAi);mex-6(RNAi) and high levels of MEX-1 (L).

MEX-5, MEX-6, and Soma/Germline Asymmetry679
simply to an increase in the penetrance of a single mex-5 proper cleavage asymmetry in the germline blastomeresP2 and P3 (Mello et al., 1992; Tabara et al., 1999), anddefect: the mislocalization of the germline protein PIE-1.
mex-6(pk440) embryos and mex-6(RNAi) embryos are both proteins are mislocalized in mex-5;mex-6 mutants.viable and develop into fertile adults, suggesting thatmex-6(1) function is not essential when mex-5(1) func- MEX-5/MEX-6 and Protein Localizationtion is present. In contrast, mex-5(zu155) embryos and in the Early Embryomex-5(RNAi) embryos are inviable, indicating that mex- Many of the examples of protein mislocalization we ob-6(1) activity does not compensate fully for the loss of serve in mex-5;mex-6 mutant embryos involve the ante-mex-5(1) activity. We do not yet know the expression rior misexpression of proteins that normally are ex-pattern of the MEX-6 protein, nor do we know whether pressed in posterior and germline blastomeres. Forartificially increasing the levels of MEX-6 would be suffi- example, the proteins SKN-1, PAL-1, MEX-1, PIE-1,cient to rescue the mex-5 mutant. If the expression pat- POS-1, and SKN-1 are each misexpressed in the anteriortern of MEX-6 coincides with that of MEX-5 in wild-type of mex-5;mex-6 mutant embryos. This result suggestsembryos, it may be that the levels of MEX-6 in mex-5 that in wild-type embryos, MEX-5/MEX-6 function tomutants are insufficient to perform all essential func- inhibit the anterior expression of these proteins. How-tions, such as those required for the proper localization ever, MEX-5/MEX-6 also plays a role in permitting theof SKN-1. expression of the anterior protein GLP-1, since this pro-
tein is not detectable in mex-5;mex-6 mutants. In wild-type embryos, translation of glp-1 mRNA appears to beMEX-5/MEX-6 Are Distinct from PAR Proteinsrepressed in posterior blastomeres (see Introduction).Blastomeres in mex-5;mex-6 embryos have several de-Thus, MEX-5/MEX-6 may prevent a posterior-localized,fects in protein expression that resemble defects de-trans-acting repressor of glp-1 mRNA from being ex-scribed for par mutants but do not have obvious defectspressed in the anterior of the embryo, consistent within PAR asymmetry at the 1-cell stage. While mutationsthe effect of MEX-5/MEX-6 on other posterior-localizedin any one of the par genes except par-4 alter the posi-proteins.tion of the first cleavage furrow (Rose and Kemphues,
1998), mex-5;mex-6 embryos have a normal, asymmetri-cally positioned furrow. These observations suggest MEX-5/MEX-6 and the Germline Proteins
In 2-cell stage wild-type embryos, the expression ofthat MEX-5/MEX-6 have functions that are distinct fromthose of the PAR proteins. In support of this view, we MEX-5 is reciprocal to that of all posterior proteins.
However, this is not the case at the 4-cell stage. Forhave shown that MEX-5/MEX-6 have effects on the ex-pression of the germline proteins PIE-1 and MEX-1 that example, SKN-1 is expressed at high, equal levels in
both of the posterior sister blastomeres at the 4-cellare independent of the asymmetrical network of PARproteins. First, our genetic experiments demonstrate stage: one sister lacks MEX-5 (the germline blastomere
P2), but the other sister contains high levels of MEX-5.that MEX-5/MEX-6 activities inhibit the expression ofboth PIE-1 and MEX-1 in mutant backgrounds lacking Since MEX-5/MEX-6 functions are required for the
proper localization of the posterior proteins, it is possi-either anterior PAR proteins (PAR-3), or posterior PARproteins (PAR-1). Second, heat shock induction of ble that MEX-5/MEX-6 functions before the 4-cell stage.
It is also possible that MEX-5/MEX-6 affects the localiza-MEX-5 at or after the 2-cell stage can inhibit the expres-sion of the MEX-1 protein in anterior blastomeres that do tion of some posterior proteins indirectly. For example,
MEX-3 appears to repress the anterior expression ofnot contain the asymmetrical network of PAR proteins.Although mex-5;mex-6 mutant embryos do not show PAL-1 in wild-type embryos (Draper et al., 1996; Hunter
and Kenyon, 1996), and MEX-3 is expressed at abnor-defects in the asymmetry of PAR proteins at the 1-cellstage, par mutant embryos show highly penetrant de- mally low levels in mex-5;mex-6 mutants.
Although the presence of MEX-5 does not directlyfects in MEX-5 protein localization. Thus, the PAR pro-teins may not be required for MEX-5/MEX-6 activity but correlate with the absence of all posterior proteins, it
correlates remarkably with the absence of the germlineare required for MEX-5, and possibly MEX-6, local-ization. proteins MEX-1, PIE-1, and POS-1: during the early
cleavages, MEX-5 is present at high levels in all regionsThe mex-5;mex-6 embryos often show PAR-likecleavage defects after the 1-cell stage. It is possible of the embryo that contain the mRNAs for the germline
proteins but that do not accumulate the germline pro-that these later defects result from the misexpressionof PAR, or non-PAR, proteins that regulate cell division. teins. The mRNAs for the germline proteins are exam-
ples of class II maternally provided mRNAs (SeydouxIn wild-type embryos, the asymmetrical network of PARproteins re-forms during each cell cycle of the asymmet- and Fire, 1994); these mRNAs are present in the oocyte
and persist at high levels in each successive germlinerically dividing germline blastomeres. PAR asymmetryin the germline lineage has not been studied as exten- blastomere. When a germline blastomere divides, its
somatic daughter and germline daughter inherit compa-sively as for the 1-cell stage and is complicated by thefact that the a–p axis of the germline blastomeres ap- rable levels of mRNA. However, after one or two addi-
tional cell cycles these mRNAs are degraded in the de-pears to invert during the third cell cycle (Schierenberg,1987). Nevertheless, these observations suggest that scendants of the somatic daughter. For example, mex-1
mRNA is present at high levels in the anterior, somaticsome factor(s) that regulates PAR asymmetry is re-stricted to the germline blastomeres and thus may be blastomere of a 2-cell embryo but disappears rapidly
from the daughters of the anterior blastomere duringmislocalized in mex-5;mex-6 mutants. For example, thegermline proteins MEX-1 and POS-1 are required for the 4-cell stage (Guedes and Priess, 1997), and MEX-5

Molecular Cell680
disappears from these same daughters late in the 4-cell positively regulate the translation of maternal mRNAsin the germline blastomere (Tabara et al., 1999). If so,stage. Thus, MEX-5 may not be necessary to prevent
expression of the germline proteins in anterior blasto- it will be interesting to determine how the molecularfunctions of MEX-1 and POS-1 compare with MEX-5meres once the germline mRNAs are targeted for degra-
dation. In contrast, MEX-5 remains at high levels in the and MEX-6.somatic sister of the germline blastomere throughoutthe 4-cell stage, and this somatic sister is the only blas- PAR Asymmetry and MEX-5/MEX-6tomere that retains high levels of the germline mRNAs Several studies have demonstrated that the asymmetri-without expressing the germline proteins. cal network of PAR proteins is required for a–p differ-
The most striking examples of MEX-5 and germline ences in the fates of the early embryonic blastomeresproteins having reciprocal asymmetry are the gradients in C. elegans (Kemphues et al., 1988; Bowerman et al.,that form during the 1-cell and 2-cell stages. mex-5(1) 1997). We propose that MEX-5/MEX-6 is a critical com-and mex-6(1) activities are required for the gradients ponent of the link between PAR proteins and cell fate.of PIE-1 and MEX-1; however, neither pie-1(1) nor mex- We have shown here that the asymmetric localization1(1) activities are required for the gradient of MEX-5. of MEX-5 closely matches the a–p boundary betweenWe have further shown that MEX-5 expression is recip- anterior and posterior PAR proteins, and that PAR func-rocal to that of the germline proteins when MEX-5 is tion is required for MEX-5 localization.expressed inappropriately in posterior blastomeres. In One intriguing aspect of MEX-5 localization is the as-our heat shock experiments, high levels of induced sociation with centrosomes. MEX-5 becomes associ-MEX-5 expression were correlated invariably with low ated with centrosomes only in the germline blastomeres,levels of endogenous MEX-1. Similarly, in various par which are the only blastomeres that reproduce themutants, MEX-5 is expressed at high levels in posterior asymmetrical network of PAR proteins during each ofblastomeres that lack, or have low levels of, germline the early cell cycles. In mutants defective for either ante-proteins, and MEX-5/MEX-6 function is required for rior or posterior PAR proteins, MEX-5 does not associatethose low levels. These several results suggest that with centrosomes, though MEX-5 is present at high lev-MEX-5/MEX-6 may directly inhibit the expression of at els. Thus, in wild-type embryos, PAR functions at theleast some of the germline proteins. posterior pole could cause MEX-5 to localize initially to
both centrosomes. The subsequent 908 rotation of theControl of Germline Protein Levels centrosomes/nascent spindle complex toward the a–pSince the mRNAs for proteins like MEX-1 and PIE-1 are axis of the embryo would move the anterior-mostdistributed uniformly until the 4-cell stage, how might centrosome away from the posterior PAR proteins, pos-MEX-5/MEX-6 inhibit the expression of these proteins? sibly allowing MEX-5 to disassociate. Additional mecha-A mechanism for degrading germline proteins may exist nisms might then degrade MEX-5 remaining in the pos-in the somatic daughters of germline blastomeres, and terior centrosome at anaphase.therefore MEX-5/MEX-6 could be part of this process Recent studies have identified orthologs of PAR-1(see Introduction). We do not yet know whether degra- and PAR-3 in Drosophila (Kuchinke et al., 1998) and indation might also be involved in setting up the gradient mammals (Bohm et al., 1997; Izumi et al., 1998), whereof germline proteins that develops in germline blasto- they have asymmetrical, cortical distributions in polar-meres prior to division. As the gradients of PIE-1 and ized cells. For example, the PAR-3-related protein inMEX-1 form during the 1-cell stage, the levels of these Drosophila (Bazooka) is localized asymmetrically to theproteins remain low at the anterior pole while the levels cortex of neuroblasts and is required for the proper,increase toward the posterior pole. In principle, the gra- asymmetrical distribution of other neuroblast proteinsdient could be generated by coupling degradation to (Schober et al., 1999; Wodarz et al., 1999). In futurenonlocalized translation, or by restricting the translation studies, it will be interesting to determine whether MEX-of the germline mRNAs to the posterior pole. 5-related proteins have general roles in linking cortical
Finger motifs similar to those found in MEX-5 and asymmetries to cytoplasmic differences in protein ex-MEX-6 have been described in several proteins that presssion.appear to interact, directly or indirectly, with RNA. Inparticular, mouse TTP has been shown to bind tumor Experimental Proceduresnecrosis factor (TNF) mRNA and requires the finger do-
Nematode Strains and Allelesmains to do so (Carballo et al., 1998; Lai et al., 1999). IfThe Bristol strain N2 was used as the standard wild-type strain.the finger motifs function in general to bind RNA, anmex-5(zu199) was used in cis to unc-31(e169) or unc-30(e191). Inattractive model is that MEX-5 and MEX-6 might bindaddition, the following strains and genetic reagents were used: I,
to, and prevent the translation of, germline messages. dpy-5(e61); II, rol-1(e187); III, unc-32(e189),par-3(it71); Qc1 IV, skn-Like MEX-5 and MEX-6, the germline proteins PIE-1, 1(zu67), unc-5(e53), unc-26(e345), dpy-4(e1166), unc-24(e138),
unc-22(s7), let-324(s1727), dpy-20(e1282), nDf27, sDf66, nT1; V,MEX-1, and POS-1 each contain two copies of a relateddpy-11(e224), par-1(b274), rol-4(sc8); and X, lon-2(e678). C. elegansfinger motif. PIE-1 is a predominantly nuclear proteinculture, mutagenesis, and genetics were as described in Brenner,that is markedly different from MEX-1 and POS-1, and1974).that appears to repress transcription in the germline
blastomeres through an unknown mechanism (SeydouxIsolation of mex-5(zu199) and Mutant Characterization
et al., 1996). MEX-1 and POS-1 are cytoplasmic proteins; mex-5(zu199) was isolated in an F2 screen for maternal effect lethalalthough their biochemical functions are not known, mu- mutants as described by Priess et al. (1987) and mapped near unc-
30; map data is available from the C. elegans Genetic Center. Alltant analysis has suggested that these proteins might

MEX-5, MEX-6, and Soma/Germline Asymmetry681
self-progeny of homozygous mex-5(zu199) hermaphrodites arrested done on 8–16 cell stage embryos. Combined embryos analyzed(MEX-5 levels/MEX-1 levels) were: 7 high/low, 17 low/high, 11 mo-during embryogenesis (n 5 3412). All cross-progeny after male mat-
ing were indistinguishable from self-progeny (n 5 123). Development saic (n 5 35).of individual early blastomeres was assayed after killing all otherblastomeres with a laser microbeam (Bowerman et al., 1992; Mello Acknowledgmentset al., 1992). Size of the AB blastomere was determined with morpho-metric software in NIH Image on 12 wild-type and 13 mex-5;mex-6 We thank C. C. Mello for discussion and sharing unpublished resultsembryos. that expedited the cloning of mex-5; E. Wayner for assistance in
generating the monoclonal antibodies; C. Hunter, K. Kemphues, J.Molecular Analysis Kimble, and Y. Tabuse for providing reagents; and current and for-For mex-5 rescue, the cosmid W02A2 and marker rol-6 DNA were mer members of the Priess lab for advice and discussion. C. M. S.coinjected into the syncytial gonad of unc-31(e169)mex-5(zu199)/ was supported in part by a Public Health Services National ResearchnT1 adults following published procedures (Mello et al., 1991). The Service Award and the NIH. Some nematode strains used in thismex-5(zu199) and mex-6(pk440) alleles were sequenced using stan- work were provided by the Caenorhabditis elegans Genetic Center,dard protocols from PCR-amplified genomic DNA. mex-6(pk440) which is funded by the NIH. J. R. P. is supported by the HowardDNA is a deletion removing sequences 12737–15477 within the cos- Hughes Medical Institute.mid AH6 (GenBank).
Predicted structure of MEX-5 from genomic sequence (W02A2.7; Received January 19, 2000; revised March 21, 2000.GenBank) agreed with sequence analysis of the mex-5 cDNAyk328d9. The predicted MEX-6 protein (AH6.5; GenBank) contains References30 N-terminal amino acids not present in the 59 end sequences ofseveral mex-6 cDNAs (yk339c12, yk423a1, yk562a8, yk520g6). Barabino, S.M., Hubner, W., Jenny, A., Minvielle-Sebastia, L., and
Keller, W. (1997). The 30-kD subunit of mammalian cleavage anddsRNA Inhibition polyadenylation specificity factor and its yeast homolog are RNA-Regions of the mex-5 and mex-6 genes corresponding, respectively, binding zinc finger proteins. Genes Development 11, 1703–1716.to amino acids 153–186 and 156–190 in Figure 3 were amplified by Bohm, H., Brinkmann, V., Drab, M., Henske, A., and Kurzchalia, T.V.polymerase chain reaction (PCR) from phage yk45f12 and yk38b2, (1997). Mammalian homologues of C. elegans PAR-1 are asymmetri-respectively; oligonucleotides contained sites for T3 or T7 polymer- cally localized in epithelial cells and may influence their polarity.ase. pie-1 dsRNA was prepared from the plasmids pJP660 and Curr. Biol. 7, 603–606.pJP661 (Mello et al., 1996) after PCR amplification. Worms were
Bowerman, B., and Shelton, C.A. (1999). Cell polarity in the earlytreated with dsRNA either by injection (Powell-Coffman et al., 1996;Caenorhabditis elegans embryo. Curr. Opin. Genet. Dev. 9, 390–395.Rocheleau et al., 1997) or by soaking (Tabara et al., 1999).Bowerman, B., Eaton, B.A., and Priess, J.R. (1992). skn-1, a mater-nally expressed gene required to specify the fate of ventral blasto-Immunolocalizationmeres in the early C. elegans embryo. Cell 68, 1061–1075.Antibodies/antisera against the following proteins were used as de-Bowerman, B., Draper, B.W., Mello, C.C., and Priess, J.R. (1993).scribed: MEX-3 (Draper et al., 1996), POS-1 (Tabara et al., 1999),The maternal gene skn-1 encodes a protein that is distributed un-MEX-1 (Guedes and Priess, 1997), PIE-1 (Tenenhaus et al., 1998),equally in early C. elegans embryos. Cell 74, 443–452.PAL-1 (Hunter and Kenyon, 1996), SKN-1(Bowerman et al., 1993),
PAR-1 (Guo and Kemphues, 1995), PAR-2 (Boyd et al., 1996), PAR-3 Bowerman, B., Ingram, M.K., and Hunter, C.P. (1997). The maternal(Etemad-Moghadam et al., 1995), PKC-3 (Tabuse et al., 1998), P par genes and the segregation of cell fate specification activities ingranules (Strome and Wood, 1982). Pharyngeal muscles were early Caenorhabditis elegans embryos. Development 124, 3815–stained with mAb3NB12 (Mello et al., 1992). mAbCS1 was generated 3826.following published procedures (Wayner and Carter, 1987) in the Boyd, L., Guo, S., Levitan, D., Stinchcomb, D.T., and Kemphues,FHCRC Hybridoma Production Facility using bacterially expressed, K.J. (1996). PAR-2 is asymmetrically distributed and promotes asso-full-length, His-tagged MEX-5. Fixation and staining was as in Lin ciation of P granules and PAR-1 with the cortex in C. elegans em-et al. (1995). bryos. Development 122, 3075–3084.
PAR and germline protein staining staining was scored at theBrenner, S. (1974). The genetics of Caenorhabditis elegans. Genetics1-cell stage embryos after the pronuclei had decondensed and be-77, 71–94.gun migration. Figure 4 is based on analysis of between 16–40 wild-Carballo, E., Lai, W.S., and Blackshear, P.J. (1998). Feedback inhibi-type embryos and 11–58 mex-5;mex-6 mutant embryos. For thetion of macrophage tumor necrosis factor-alpha production byexperiments shown in Figures 4 and 7, wild-type and mutant em-tristetraprolin. Science 281, 1001–1005.bryos were placed on the same slide for immunostaining and were
photographed using identical exposure times. Crittenden, S.L., Rudel, D., Binder, J., Evans, T.C., and Kimble, J.(1997). Genes required for GLP-1 asymmetry in the early Caenorhab-ditis elegans embryo. Dev. Biol. 181, 36–46.Heat Shock Experiments
Full-length MEX-5 was cloned into heat shock vectors pPD49.78 Draper, B.W., Mello, C.C., Bowerman, B., Hardin, J., and Priess, J.R.and pPD49.83 (Seydoux et al., 1996) and coinjected with rol-6 DNA (1996). MEX-3 is a KH domain protein that regulates blastomereinto unc-30(e191)mex-5(zu199)/nT1 hermaphrodites (Mello et al., identity in early C. elegans embryos. Cell 87, 205–216.1991). Stage-selected, calibration embryos and gravid hermaphro- DuBois, R.N., McLane, M.W., Ryder, K., Lau, L.F., and Nathans,dites containing embryos at multiple stages were subjected to one D. (1990). A growth factor-inducible nuclear protein with a novelof two protocols with equivalent results. (1) One hour heat shock cysteine/histidine repetititve sequence. J. Biol. Chem. 265, 19185–348C was followed by a 1.5 hr recovery period at room temperature. 19191.One-cell stage calibration embryos had not progressed beyond the
Etemad-Moghadam, B., Guo, S., and Kemphues, K.J. (1995). Asym-4-cell stage before fixation (6/6 embryos), and most 2-cell stagemetrically distributed PAR-3 protein contributes to cell polarity andembryos did not progress beyond the 12-cell stage (4/5 embryos).spindle alignment in early C. elegans embryos. Cell 83, 743–752.Thus, nonstaged, fixed embryos with 8–16 cells must have beenEvans, T.C., Crittenden, S.L., Kodoyianni, V., and Kimble, J. (1994).past the 1-cell stage before induction of MEX-5. (2) Three hour heatTranslational control of maternal glp-1 mRNA establishes an asym-shock at 308C was terminated without a recovery period. One-cellmetry in the C. elegans embryo. Cell 77, 183–194.stage calibration embryos progressed to about the 24-cell stage
before fixation, and no MEX-5 could be detected in control embryos Freed, E., Lacey, K.R., Huie, P., Lyapina, S.A., Deshaies, R.J.,Stearns, T., and Jackson, P.K. (1999). Components of an SCF ubiqui-fixed after the first 2 hr of heat shock. Thus, nonstaged, fixed em-
bryos with 8–16 cells are unlikely to have had detectable levels of tin ligase localize to the centrosome and regulate the centrosomeduplication cycle. Genes Dev. 13, 2242–2257.MEX-5 before the 1–2 cell stage. After either protocol, analysis was

Molecular Cell682
Guedes, S., and Priess, J.R. (1997). The C. elegans MEX-1 protein Schierenberg, E. (1987). Reversal of cellular polarity and early cell-cell interaction in the embryos of Caenorhabditis elegans. Dev. Biol.is present in germline blastomeres and is a P granule component.122, 452–463.Development 124, 731–739.
Schober, M., Schaefer, M., and Knoblich, J.A. (1999). Bazooka re-Guo, S., and Kemphues, K. (1995). par-1, a gene required for estab-cruits Inscuteable to orient asymmetric cell divisions in Drosophilalishing polarity in C. elegans embryos, encodes a putative Ser/Thrneuroblasts. Nature 402, 548–551.kinase that is asymmetrically distributed. Cell 81, 611–620.
Seol, J.H., Feldman, R.M., Zachariae, W., Shevchenko, A., Correll,Hung, T.J., and Kemphues, K.J. (1999). PAR-6 is a conserved PDZC.C., Lyapina, S., Chi, Y., Galova, M., Claypool, J., Sandmeyer, S.,domain-containing protein that colocalizes with PAR-3 in Caeno-Nasmyth, K., and Deshaies, R.J. (1999). Cdc53/cullin and the essen-rhabditis elegans embryos. Development 126, 127–135.tial Hrt1 RING-H2 subunit of SCF define a ubiquitin ligase moduleHunter, C.P., and Kenyon, C. (1996). Spatial and temporal controlsthat activates the E2 enzyme Cdc34. Genes Dev. 13, 1614–1626.target pal-1 blastomere-specification activity to a single blastomereSeydoux, G., and Fire, A. (1994). Soma-germline asymmetry in thelineage in C. elegans embryos. Cell 87, 217–226.distributions of embryonic RNAs in Caenorhabditis elegans. Devel-Izumi, Y., Hirose, T., Tamai, Y., Hirai, S., Nagashima, Y., Fujimoto,opment 120, 2823–2834.
T., Tabuse, Y., Kemphues, K.J., and Ohno, S. (1998). An atypicalSeydoux, G., Mello, C.C., Pettitt, J., Wood, W.B., Priess, J.R., andPKC directly associates and colocalizes at the epithelial tight junc-Fire, A. (1996). Repression of gene expression in the embryoniction with ASIP, a mammalian homologue of Caenorhabditis elegansgerm lineage of C. elegans. Nature 382, 713–716.polarity protein PAR-3. J. Cell Biol. 143, 95–106.Strome, S., and Wood, W.B. (1982). Immunofluorescence visualiza-Jansen, G., Hazendonk, E., Thijssen, K.L., and Plasterk, R.H. (1997).tion of germ-line-specific cytoplasmic granules in embryos, larvae,Reverse genetics by chemical mutagenesis in Caenorhabditis ele-and adults of Caenorhabditis elegans. Proc. Natl. Acad. Sci. USAgans. Nat. Genet. 17, 119–121.79, 1558–1562.
Joazeiro, C.A., Wing, S.S., Huang, H., Leverson, J.D., Hunter, T.,Tabara, H., Hill, R., Mello, C., Priess, J., and Kohara, Y. (1999). pos-1and Liu, Y.C. (1999). The tyrosine kinase negative regulator c-Cblencodes a cytoplasmic zinc-finger protein essential for germlineas a RING-type, E2-dependent ubiquitin-protein ligase. Science 286,specification in C. elegans. Development 126, 1–11.
309–312.Tabuse, Y., Izumi, Y., Piano, F., Kemphues, K.J., Miwa, J., and Ohno,
Kemphues, K.J., Priess, J.R., Morton, D.G., and Cheng, N.S. (1988).S. (1998). Atypical protein kinase C cooperates with PAR-3 to estab-
Identification of genes required for cytoplasmic localization in earlylish embryonic polarity in Caenorhabditis elegans. Development
C. elegans embryos. Cell 52, 311–320.125, 3607–3614.
Kuchinke, U., Grawe, F., and Knust, E. (1998). Control of spindle Tenenhaus, C., Schubert, C., and Seydoux, G. (1998). Genetic re-orientation in Drosophila by the Par-3-related PDZ-domain protein quirements for PIE-1 localization and inhibition of gene expressionBazooka. Curr. Biol. 8, 1357–1365. in the embryonic germ lineage of Caenorhabditis elegans. Dev. Biol.Lai, W.S., Stumpo, D.J., and Blackshear, P.J. (1990). Rapid insulin- 200, 212–224.stimulated accumulation of an mRNA encoding a proline-rich region. Varnum, B.C., Lim, R.W., Sukhatme, V.P., and Herschman, H.R.J. Biol. Chem. 265, 16556–16563. (1989). Nucleotide sequence of a cDNA encoding TIS11, a messageLai, W.S., Carballo, E., Strum, J.R., Kennington, E.A., Phillips, R.S., induced in Swiss 3T3 cells by the tumor promoter tetradecanoyland Blackshear, P.J. (1999). Evidence that tristetraprolin binds to phorbol acetate. Oncogene 4, 119–120.AU-rich elements and promotes the deadenylation and destabiliza- Waterman, H., Levkowitz, G., Alroy, I., and Yarden, Y. (1999). Thetion of tumor necrosis factor alpha mRNA. Mol. Cell. Biol. 19, 4311– RING finger of c-Cbl mediates desensitization of the epidermal4323. growth factor receptor. J. Biol. Chem. 274, 22151–22154.Lin, R., Thompson, S., and Priess, J.R. (1995). pop-1 encodes an Wayner, E.A., and Carter, W.G. (1987). Identification of multiple cellHMG box protein required for the specification of a mesoderm pre- adhesion receptors for collagen and fibronectin in human fibrosar-cursor in early C. elegans embryos. Cell 83, 599–609. coma cells possessing unique alpha and common beta subunits. J.
Cell Biol. 105, 1873–1884.Lorick, K.L., Jensen, J.P., Fang, S., Ong, A.M., Hatakeyama, S., andWeissman, A.M. (1999). RING fingers mediate ubiquitin-conjugating Wodarz, A., Ramrath, A., Kuchinke, U., and Knust, E. (1999). Bazookaenzyme (E2)-dependent ubiquitination. Proc. Natl. Acad. Sci. USA provides an apical cue for Inscuteable localization in Drosophila96, 11364–11369. neuroblasts. Nature 402, 544–547.
Mello, C.C., Kramer, J.M., Stinchcomb, D., and Ambros, V. (1991). Zuo, P., and Maniatis, T. (1996). The splicing factor U2AF35 mediatesEfficient gene transfer in C. elegans: extrachromosomal mainte- critical protein-protein interactions in constitutive and enhancer-
dependent splicing. Genes Dev. 10, 1356–1368.nance and integration of transforming sequences. EMBO J. 10,3959–3970.
Mello, C.C., Draper, B.W., Krause, M., Weintraub, H., and Priess,J.R. (1992). The pie-1 and mex-1 genes and maternal control ofblastomere identity in early C. elegans embryos. Cell 70, 163–176.
Mello, C.C., Schubert, C., Draper, B., Zhang, W., Lobel, R., andPriess, J.R. (1996). The PIE-1 protein and germline specification inC. elegans embryos. Nature 382, 710–712.
Powell-Coffman, J.A., Knight, J., and Wood, W.B. (1996). Onset ofC. elegans gastrulation is blocked by inhibition of embryonic tran-scription with an RNA polymerase antisense RNA. Dev. Biol. 178,472–483.
Priess, J.R., Schnabel, H., and Schnabel, R. (1987). The glp-1 locusand cellular interactions in early C. elegans embryos. Cell 51,601–611.
Rocheleau, C.E., Downs, W.D., Lin, R., Wittmann, C., Bei, Y., Cha,Y.H., Ali, M., Priess, J.R., and Mello, C.C. (1997). Wnt signaling andan APC-related gene specify endoderm in early C. elegans embryos.Cell 90, 707–716.
Rose, L.S., and Kemphues, K.J. (1998). Early patterning of the C.elegans embryo. Annu. Rev. Genet. 32, 521–545.