Embed Size (px)
Citation preview

Sadiqa Firdous
2017
Department of Biotechnology
Pakistan Institute of Engineering and Applied Sciences
Nilore, Islamabad, Pakistan
Molecular Characterization of Glyphosate
Degrading and/or Resistant Bacterial
Strains


Thesis Approval Form
Student’s Name: Sadiqa Firdous Department: Biotechnology
Registration Number: 10-7-1-066-2011 Date of Registration: 25-05-2010
Thesis Title: Molecular Characterization of Glyphosate Degrading and/or Resistant Bacterial
Strains
RECOMMENDATION (if any) by:
General comments (attach additional sheet if required)
When the final thesis defense of the student has been concluded and all other requirements have been
met, I
a. Do Recommend that the candidate be certified to the faculty for the degree of
Doctor of Philosophy
b. Do Recommend that the candidate be certified to the faculty for the degree of
Doctor of Philosophy subject to the minor correction in the thesis.
c. Do Recommend that the candidate should reappear in the oral defense
d. Do NOTRecommend that the candidate be certified to the faculty for the degree
of Doctor of Philosophy
Examiners Signatures
1. Internal Examiner (Name & Affiliation):
2. Internal Examiner (Name & Affiliation):
3. Internal Examiner (Name & Affiliation):
4. Supervisor (Name & Affiliation):
5. Head of Department (Name):
6. Dean (Research) :
Approved by:
Head of the Department (Name) :___________________________ Signatures/Date ______________
Distribution:
1. Original to be placed in Student‟s Personal file in the office of Dean (Research)
2. Copy to be included in the thesis prior to final submission.

Thesis Submission Approval
This is to certify that the work contained in this thesis entitled Molecular
Characterization of Glyphosate Degrading and/or Resistant Bacterial Strains,
was carried out by Sadiqa Firdous, and in my opinion, it is fully adequate, in scope
and quality, for the degree of Ph.D. Furthermore, it is hereby approved for submission
for review and thesis defense.
Supervisor: _____________________
Name: Dr. Samina Iqbal
Date: 21 June, 2017
Place: NIBGE, Faisalabad.
Head, Department of Biotechnology: ___________________
Name: Dr. Shahid Mansoor
Date: 21 June, 2017
Place: NIBGE, Faisalabad.

Molecular Characterization of Glyphosate
Degrading and/or Resistant Bacterial
Strains
Sadiqa Firdous
Submitted in partial fulfillment of the requirements
for the degree of Ph.D.
2017
Department of Biotechnology
Pakistan Institute of Engineering and Applied Sciences
Nilore, Islamabad, Pakistan

ii
Dedicated to My beloved Parents and Sisters Without whom none of my success
would have been possible

iii
Declaration of Originality
I hereby declare that the work contained in this thesis and the intellectual content of
this thesis are the product of my own work. This thesis has not been previously
published in any form nor does it contain any verbatim of the published resources
which could be treated as infringement of the international copyright law. I also
declare that I do understand the terms „copyright‟ and „plagiarism,‟ and that in case of
any copyright violation or plagiarism found in this work, I will be held fully
responsible of the consequences of any such violation.
__________________
(Sadiqa Firdous)
21 June, 2017
NIBGE, Faisalabad.

iv
Copyrights Statement
The entire contents of this thesis entitled Molecular Characterization of Glyphosate
Degrading and/or Resistant Bacterial Strains by Sadiqa Firdous are an intellectual
property of Pakistan Institute of Engineering & Applied Sciences (PIEAS). No
portion of the thesis should be reproduced without obtaining explicit permission from
PIEAS.

v
Table of Contents
Dedication ...................................................................................................................... ii
Declaration of Originality .............................................................................................. iii
Copyrights Statement ..................................................................................................... iv
List of Figures ................................................................................................................ ix
List of Tables.................................................................................................................. xi
Abstract .........................................................................................................................xii
List of Publications ....................................................................................................... xiv
List of Abbreviations ..................................................................................................... xv
Acknowledgements ..................................................................................................... xvii
1. Introduction .............................................................................................................. 1
1.1 Xenobiotics in Environment .............................................................................. 1
1.2 Pesticides .......................................................................................................... 1
1.3 Classification of Pesticides ................................................................................ 2
1.4 Organophosphonate Pesticides........................................................................... 3
1.5 Herbicides ......................................................................................................... 5
1.5.1 Group 1: Acetyl CoA Carboxylase (ACCase) Inhibitors (Lipid
Biosynthesis Inhibitors) ................................................................................ 5
1.5.2 Group 2: Acetolactate Synthase (ALS) or Acetohydroxy Acid Synthase
(AHAS) Inhibitors ........................................................................................ 6
1.5.3 Group 3: Mitosis Inhibitors (Root Growth Inhibitors) ................................... 6
1.5.4 Group 4: Plant Growth Regulators (Synthetic Auxins) .................................. 6
1.5.5 Groups 5, 6, 7: Photosynthesis inhibitors (Photosystem II (PSII)
Inhibitors) ..................................................................................................... 7
1.5.6 Groups 8 and 15: Shoot Growth Inhibitors .................................................... 7
1.5.7 Group 9: Aromatic Amino Acid Inhibitors .................................................... 8
1.5.8 Group 10: Glutamine Synthesis Inhibitors ..................................................... 8
1.5.9 Groups 12, 13, and 27: Pigment Synthesis Inhibitors ..................................... 8
1.5.10 Group 14: Protoporphyrinogen Oxidase (PPO) Inhibitors.............................. 9
1.5.11 Group 22: Photosynthesis Inhibitors (Photosystem I (PSI) Inhibitors) ........... 9
1.6 Glyphosate ........................................................................................................ 9
1.6.1 Mode of Action of Glyphosate .................................................................... 10
1.6.2 Uptake and Translocation of Glyphosate ..................................................... 12
1.6.3 Fate of Glyphosate in Soil ........................................................................... 12

vi
1.6.4 Fate of Glyphosate in Water ........................................................................ 13
1.6.5 Global Use of Glyphosate ........................................................................... 14
1.6.6 Environmental Impact and Toxicity of Glyphosate ...................................... 15
1.7 Biodegradation ................................................................................................ 18
1.7.1 Biodegradation of Glyphosate ..................................................................... 19
2. Materials and Methods ........................................................................................... 21
2.1 Chemicals Used in This Study ......................................................................... 21
2.2 Bacterial Strains Used in This Study ................................................................ 21
2.3 Soil Sample Collection .................................................................................... 23
2.4 Growth Media ................................................................................................. 23
2.5 Maintenance and Preservation of the Isolated Bacterial Strains ........................ 23
2.6 Equipments Used in Current Study .................................................................. 23
2.7 Enrichment of Glyphosate Tolerant Bacterial Strains ....................................... 24
2.8 Isolation of Glyphosate Tolerant Bacterial Strains ........................................... 24
2.9 Glyphosate Tolerance of Bacterial Isolates ...................................................... 24
2.10 Morphological Characterization of the Bacterial Isolates ................................. 25
2.10.1 Morphological Characterization .................................................................. 25
2.10.2 Gram Staining Method ................................................................................ 25
2.11 Molecular Characterization of Isolated Bacterial Strains .................................. 26
2.11.1 DNA Isolation ............................................................................................ 26
2.11.2 Amplification of 16S rRNA Gene from Isolated Bacterial Strains ............... 27
2.11.3 Agarose Gel Electrophoresis ....................................................................... 27
2.11.4 Ligation and Cloning of 16S rRNA Gene .................................................... 27
2.11.5 Sequencing of 16S rRNA Gene and Bacterial Identification ........................ 27
2.12 Bacterial Diversity Assessment through PCR-DGGE ...................................... 28
2.12.1 DGGE Gel Casting and Running ................................................................. 28
2.12.2 DGGE Gel Staining .................................................................................... 29
2.12.3 Excision and Re-amplification of DGGE PCR Fragments ........................... 29
2.13 Inoculum Preparation of Bacterial Strains ........................................................ 29
2.14 Glyphosate Degradation Studies ...................................................................... 30
2.15 Detection of Glyphosate .................................................................................. 30
2.15.1 Detection Wavelength of Glyphosate .......................................................... 30
2.15.2 Derivatization of Glyphosate ....................................................................... 30
2.15.3 HPLC Conditions for Glyphosate Residual Analysis ................................... 31
2.16 Study of Potential Genes in Glyphosate Tolerant Bacterial Strains ................... 31
2.16.1 Amplification of Glyphosate Resistant and Degrading Genes ...................... 31

vii
2.16.2 Agarose Gel Electrophoresis, Cloning and Sequencing of the Amplified
Genes .......................................................................................................... 32
3. Isolation and Characterization of Glyphosate Resistant Bacterial Strains ................ 34
3.1 Introduction ..................................................................................................... 34
3.1.1 Glyphosate Resistance ................................................................................ 35
3.1.2 5-Enolpyruvylshikimate-3-Phosphate Synthase (EPSPS, EC 2.5.1.26) ........ 35
3.1.3 Classification of EPSP Synthase ................................................................. 37
3.1.4 History and Engineering of Glyphosate Resistant EPSPS ............................ 37
3.1.5 Glyphosate Detoxification........................................................................... 39
3.2 Materials and Methods .................................................................................... 39
3.2.1 Enrichment, Isolation and Screening of Glyphosate Tolerant Bacterial
Strains......................................................................................................... 39
3.2.2 Morphological and Taxonomic Analysis of Glyphosate Tolerant Bacterial
Isolates ....................................................................................................... 40
3.2.3 Amplification and Sequencing of aroAO.intermedium Sq20 Gene ......................... 40
3.2.4 Tolerance/ Degradation of Glyphosate by Sq20........................................... 40
3.2.5 HPLC Analysis of Glyphosate .................................................................... 41
3.2.6 Detection of Metabolites Produced During Glyphosate Degradation ........... 41
3.2.7 Bioinformatic Analysis of aroAO.intermedium Sq20 ............................................. 41
3.3 Results............................................................................................................. 43
3.3.1 Isolation and Identification of Glyphosate Tolerant Bacterial Strains .......... 43
3.3.2 Morphological and Taxonomic Characterization of Glyphosate Tolerant
Bacterial Strains .......................................................................................... 43
3.3.3 Glyphosate Resistance and Degradation by Ochrobactrum Intermedium
Sq20 ........................................................................................................... 51
3.3.4 Detection of Glyphosate Degradation Products ........................................... 52
3.3.5 Bioinformatic Analysis of aroAO.intermedium Sq20 ............................................. 52
3.4 Discussion ....................................................................................................... 64
4. Isolation and Characterization of Glyphosate Degrading Bacterial Strains .............. 69
4.1 Introduction ..................................................................................................... 69
4.1.1 Glyphosate Biodegradation ......................................................................... 70
4.1.2 Pathways of Glyphosate Biodegradation ..................................................... 74
4.1.3 Response Surface Methodology .................................................................. 76
4.2 Materials and Methods .................................................................................... 77
4.2.1 Enrichment and Isolation of Glyphosate Degrading Bacterial Strains .......... 77
4.2.2 Selection of Competent Glyphosate Degrading Bacterial Strain .................. 77
4.2.3 Identification and Characterization of Glyphosate Degrading Bacterial
Strains......................................................................................................... 78

viii
4.2.4 Denaturing Gradient Gel Electrophoresis (DGGE) Analysis of Glyphosate
Contaminated Soil ...................................................................................... 78
4.2.5 Biodegradation of Glyphosate by Isolated Bacterial Strains ........................ 78
4.2.6 HPLC Analysis of Glyphosate Residues ..................................................... 79
4.2.7 Optimization of Culture Conditions Using Response Surface
Methodology (RSM) ................................................................................... 79
4.2.8 Detection of Genes Conferring Glyphosate Degradation ............................. 80
4.3 Results............................................................................................................. 81
4.3.1 Selection and Identification of Glyphosate Degrading Bacterial Strain ........ 81
4.3.2 Identification and Characterization of Bacterial Isolates .............................. 83
4.3.3 DGGE Analysis of Enrichment Cultures ..................................................... 91
4.3.4 Optimization of Parameters for Glyphosate Degradation Using RSM .......... 95
4.3.5 Response Surface Plots for Glyphosate Degradation ................................... 98
4.3.6 Identification of Glyphosate Degrading Genes from Isolated Bacterial
Strains....................................................................................................... 103
4.4 Discussion ..................................................................................................... 105
5. Discussion ............................................................................................................ 109
5.1 Aim of Thesis ................................................................................................ 109
5.2 Major Findings .............................................................................................. 111
5.2.1 Ochrobactrum intermedium Sq20 ............................................................. 111
5.2.2 Comamonas odontotermitis P2 .................................................................. 111
5.2.3 Glyphosate Degrading Genes .................................................................... 112
5.3 Explicit Future Recommendations ................................................................. 112
6. References ............................................................................................................ 114
Appendices .................................................................................................................. 135

ix
List of Figures
Figure 1-1 General chemical structure of organophosphonate pesticides. ............. 4
Figure 1-2 Schematic illustration of mode of action of glyphosate. .................... 11
Figure 1-3 Accumulation and distribution of glyphosate in environment. ........... 16
Figure 1-4 Pathways of glyphosate biodegradation. ........................................... 20
Figure 3-1 Inhibition of penultimate step of shikimic acid pathway by glyphosate.
......................................................................................................... 36
Figure 3-2 Growth of the isolated bacterial strains in MSM. .............................. 44
Figure 3-3 Phylogenetic analysis of isolated bacterial strain Sq20. ..................... 48
Figure 3-4 Phylogenetic analysis of isolated bacterial strain Sq11. ..................... 49
Figure 3-5 Phylogenetic analysis of isolated bacterial strain Sq14. ..................... 50
Figure 3-6 Phylogenetic analysis of isolated bacterial strain Sq16. ..................... 51
Figure 3-7 Glyphosate degradation by Ochrobactrum intermedium Sq20 at 500
mg/L initial concentration, 37 °C and pH 7 in minimal salt media
(MSM). ............................................................................................ 53
Figure 3-8 Phylogenetic analysis of EPSPS O.intermedium Sq20. ................................ 54
Figure 3-9 Multiple sequence alignment of EPSPSO.intermedium Sq20. ...................... 55
Figure 3-10 Phylogenetic analysis of Ochrobactrum intermedium strain Sq20 with
Class I and class II enzymes. ............................................................ 56
Figure 3-11 Secondary structure of EPSPSO.intermedium Sq20. .................................... 59
Figure 3-12 Secondary structure (SS) and disorder prediction of EPSPSO.intermedium
Sq20 through Phyre2 representing three states α-helix, β-strand and coil.
......................................................................................................... 60
Figure 3-13 Ramachandran plot (created by PROCHECK) showing 91.8% of
amino acid residues in core region (red colour). ................................ 61
Figure 3-14 Structure validation of EPSPSO.intermedium Sq20. .................................... 62
Figure 3-15 3-D structure analysis of class II EPSPS(s). ...................................... 63
Figure 4-1 Utilization of glyphosate (500 mg/L) by isolated bacterial strains. .... 82
Figure 4-2 Degradation of glyphosate by bacterial isolates. ................................ 83
Figure 4-3 Phylogenetic tree of Pseudomonas straminea P1. ............................. 87
Figure 4-4 Neighbor joining tree illustrating the phylogenetic relationship of P2
strain. ............................................................................................... 88
Figure 4-5 Phylogenetic analysis of Ochrobactrum anthropi P3 using Mega 6
based on 16S rRNA sequence analysis. ............................................. 89
Figure 4-6 Phylogenetic tree based on homologous sequences of the
Achromobacter spanius P4. .............................................................. 90

x
Figure 4-7 Dendrogram exhibiting genetic relationships of P5 isolate. ............... 91
Figure 4-8 (a, b) 16S rDNA denaturing gradient gel electrophoresis (DGGE)
analysis of bacterial community in enrichment cultures of glyphosate
contaminated soils. ........................................................................... 93
Figure 4-9 RSM analysis of glyphosate degradation......................................... 102
Figure 4-10 GOX amplification from Comamonas odontotermitis P2. ............... 103
Figure 4-11 Phylogenetic analysis of Comamonas odontotermitis P2 GOX. ....... 104
Figure 4-12 Amplification of phnJ gene from glyphosate degrading bacterial
isolates. .......................................................................................... 104
Figure 4-13 Blast analysis of phnJ gene amplified Comamonas odontotermitis P2.
....................................................................................................... 105

xi
List of Tables
Table 2.1 Glyphosate tolerant/degrading bacterial strains isolated in current
study................................................................................................. 22
Table 2.2 Sequences of primers designed by aligning the already reported
glyphosate resistant and degrading genes. ......................................... 33
Table 3.1 Morphological characteristics of glyphosate resistant bacterial strains.
......................................................................................................... 45
Table 3.2 Characterization of the glyphosate resistant bacterial strains isolated in
current study from glyphosate contaminated soil .............................. 47
Table 3.3 Amino acid composition of EPSPSO.intermedium Sq20 representing amino
acid number as well as percentage composition. ............................... 57
Table 3.4 Physico-chemical characteristics of EPSPSO.intermedium Sq20 computed by
Expasy‟s Protparam software. ........................................................... 58
Table 3.5 Comparative analysis of Swiss Model and Geno 3D computed models
of EPSPSO.intermedium Sq20 done by Ramachandran plot calculations. .... 67
Table 4.1 Glyphosate degrading bacterial strains with type of metabolism. ...... 72
Table 4.2 Experimental ranges and coded levels of independent variables. ....... 80
Table 4.3 Characterization of the glyphosate degrading bacterial strains isolated
in current study from glyphosate contaminated soil .......................... 84
Table 4.4 Morphological characteristics of glyphosate degrading bacterial
strains. .............................................................................................. 85
Table 4.5 Sequence analysis of DGGE bands obtained from glyphosate enriched
culture .............................................................................................. 94
Table 4.6 Predicted and experimental values of glyphosate degradation by CCD
matrix ............................................................................................... 96
Table 4.7 Analysis of variance (ANOVA) for the glyphosate degradation
response (%) ..................................................................................... 97
Table 4.8 Regression analysis and model coefficients of variables for glyphosate
degradation response (%).................................................................. 98

xii
Abstract
Glyphosate is an important organophosphonate herbicide used to eliminate grasses
and herbaceous plants in many vegetation management situations. It is one of the
most widely used herbicides owing to its non selective and post emergent properties.
Glyphosate is involved in blockage of an enzyme, 5-enolpyruvylshikimate-3-
phosphate synthase (EPSPS), entailed in catalysis of an essential step in the
biosynthesis of aromatic amino acids. The objective of the current study was to isolate
glyphosate tolerant bacterial strains and to characterize the gene(s) encoding
glyphosate resistance in these bacteria. A glyphosate tolerant bacterium,
Ochrobactrum intermedium Sq20 was isolated from glyphosate contaminated
indigenous soil, capable of utilizing glyphosate as sole carbon and energy source. A
1353 bp open reading frame (ORF) representing aroAO.intermedium Sq20 was amplified
from Sq20 which showed 97% homology with aroA genes from other Ochrobactrum
spp. Sequence analysis revealed that EPSPSO.intermedium Sq20 belongs to class II EPSPS.
In silico analysis was used for identification and characterization of EPSPS gene
through physicochemical properties. Methodical optimization and validation of
protein structure helped to build a reliable protein model of EPSPSO.intermedium Sq20
which will provide strong basis for functional analysis of EPSPSO.intermedium Sq20. The
results indicated that cloning and characterization of EPSPSO.intermedium Sq20 will further
help to understand its role at molecular level and its plausible usage for production of
glyphosate resistant transgenic crops.
The extensive use of glyphosate is detrimental for flora and fauna regarding
current studies about its toxic effects, and most appropriate strategy to remove it from
environment is bioremediation. As a step to address this problem, a novel bacterial
strain Comamonas odontotermitis P2 capable to utilize glyphosate as carbon and
phosphorus source was isolated and characterized. The glyphosate degradation
potential of C. odontotermitis P2 was optimized using response surface methodology
under various culture conditions. The strain P2 was proficient to degrade 1.5 g/L
glyphosate completely within 104 h. Moreover, GOX (glyphosate oxidoreductase) and
phnJ (C-P lyase) genes were identified from C. odontotermitis P2 signifying the

xiii
degradation potential through AMPA and sarcosine metabolic pathways. These results
demonstrate the potential of C. odontotermitis P2 for efficient degradation of
glyphosate which can be exploited for remediation of glyphosate.

xiv
List of Publications
Journal Publications
S. Firdous, S. Iqbal, S. Anwar, and H. Jabeen, “Identification and analysis of
5-enolpyruvylshikimate-3-phosphate synthase (EPSPS) gene from glyphosate
resistant Ochrobactrum intermedium Sq20,” Pest Manag Sci,
DOI:10.1002/ps.4624. 2017.
S. Firdous, S. Iqbal, and S. Anwar, “Optimization and modeling of
glyphosate biodegradation by a novel Comamonas odontotermitis P2 through
response surface methodology,” Pedosphere, Manuscript ID:
pedos201610546.R1, 2016. (In Press)
H. Jabeen, S. Iqbal, F. Ahmad, M. Afzal, and S. Firdous, “Enhanced
remediation of chlorpyrifos by ryegrass (Lolium multiflorum) and a
chlorpyrifos degrading bacterial endophyte Mezorhizobium sp. HN3,” Int. J.
Phytoremediation, vol. 1, no. 2, pp.126-133, 2016.

xv
List of Abbreviations
2,4D 2,4-Dichlorophenoxyacetic acid
2-AEP 2-Amino ethylphosphonic acid
AARI Ayub agricultural research institute
ACCase Acetyl CoA carboxylase
AHAS Acetohydroxy acid synthase
ALS Acetolactate synthase
AMPA Aminomethyl phosphonic acid
ANOVA Analysis of variance
ATPase Adenosine triphosphate
BHC Benzene hexachloride
BLAST Basic local alignment search tool
CCRD Central composite rotatable design
CFU Colony forming units
ChE Cholinesterase
CNBF 4-Chloro-3,5-dinitrobenzotrifluoride
CTAB Cetyl tri-methyl ammonium bromide
CYP Cytochrome P450
DAHP 3-Deoxy-D-arabinoheptulosonate-7-phosphate synthase
DCPA Dimethyl-2,3,5,6-tetrachloroterephthalate
DDT Dichlorodiphenyltrichloroethane
DENs Phenylpyrazolin
DF Degrees of freedom
DGGE Denaturing gradient gel electrophoresis
DHQase 3-Dehydroquinate synthase
DIMs Cyclohexanedione
E4P Phosphoenol pyruvate and erythrose-4-phosphate
EPSP 5-Enolpyruvylshikimate-3-phosphate synthase
EPSPS 5-Enolpyruvyl-shikimate-3-phosphate synthase
FOPs Aryloxyphenoxypropionate

xvi
GO Glycine oxidase
GOX Glyphosate oxidoreductase
GRAVY Grand average hydropathy
HPLC High performance liquid chromatography
HPPD 4-Hydroxyphenyl pyruvate dioxygenase
HRAC Herbicide resistance action committee
IAA Indole acetic acid
IARC International agency for research on cancer
LB Luria Bertani
MCL Maximum contaminant limit
MEGA Molecular evolutionary genetics analysis
MS Mean square
MSM Mineral salts medium
NADPH2 Nicotinamide adenine dinucleotide hydrogen phosphate
NHL Non-Hodgkin lymphoma
OCs Organochlorines
OPs Organophosphates
ORF Open reading frame
PEP Phosphoenolpyruvate
POEA Polyethoxylated tallow amine
POPs Persistent organic pollutants
PSI Photosystem I
PSII Photosystem II
RSM Response surface methodology
S3P Shikimate 3-phosphate
SAM s-Adenosyl-l-methionine
SOPMA Self optimized prediction method with alignment
SS Sum of squares
USGS U.S. geological survey
WHO World health organization

xvii
Acknowledgements
In the inception, all commendation and admiration is for the Almighty ALLAH,
whose blessings have been the main reason for the completion of my study, and His
Holy Prophet Mohammad (S.A.W.W.), who is forever torch of guidance and
knowledge for humanity as a whole.
It is my greatest pleasure to avail this opportunity to extend my gratitude to
Dr. Shahid Mansoor (DCS, SI), Director NIBGE for providing and facilitating a
competitive environment for research.
My cordial gratitude goes to my supervisor, Dr. Samina Iqbal (DCS), who
proficiently guided me during my PhD research work. She has been there to support
me in all circumstances throughout my PhD. She has always been passionate to create
an inimitable piece of research and her vital contribution to this thesis is beyond
description. In a nutshell, I would have never been able to complete my work without
her concerned involvement.
I am also indebted to Dr. Sajjad Mirza (DCS, HOD, SEBD) for his
encouragement and motivation. My humble thanks also goes out to Dr. Michael
Kertesz, my foreign supervisor for providing me a chance to utilize my research skills
and facilitating me with recourses at his lab during tenure of IRSIP fellowship
(International Research Support Initiative Program, HEC) at University of Sydney,
Australia.
I am strongly obliged to Samina Anwar (SS) for helping, encouraging and
guiding me throughout the research work. Her valuable help in experimental work
and sincere guidance was an inspiration. Without her help, I would have never been
able to conduct my research and reach its goal.
My amenable thanks go to my senior lab fellow and friend Dr. Hina Jabeen
for never refusing me whenever I approached her for seeking guidance. Her co-
operation genuinely helped to complete the journey of research.

xviii
I am highly thankful to my lab fellows Masooma Hammad, Fiaz Ahmed,
Salma Mughal, Sidra Gulnar, Muhammad Asif Nadeem and Abdul Qadeer
Wahla for their kind cooperation during my PhD research. The cooperation of my lab
fellows genuinely helped to complete the journey of research.
It fills me with immense gratitude to convey my thanks to my friends Mariam
Masood, Saima Majeed, Nida Iram, Maryam Zafar for always being there and
helping me at several places during the pace of my research.
I am highly indebted to my parents and sisters for their support, patience and
guidance. My words can never justify their role in this venture. I can never ever be
able to thank my parents Mr. and Mrs. Zafar Iqbal enough for all the love and
affection they generously poured on me; for all the tears they shed in front of ALLAH
to bless me with His innumerable blessings and for all the hardships they suffered to
comfort me. I wholeheartedly thank my sisters Amina, Ayesha and Quratulain for
their love, support and encouragement throughout my educational career.
Sadiqa Firdous

1
1. Introduction
1.1 Xenobiotics in Environment
Our environment encloses a wide-ranging variety of naturally occurring and synthetic
(biogenic or anthropogenic) chemical compounds [1]. Xenobiotic is a combination of
two Greek words xenos meaning foreign or eccentric and bios meaning life referring
to the substances foreign to the biosphere usually turn out due to industrial activities.
Some xenobiotic compounds are degraded by microorganisms (weak xenobiotics)
whereas some persist for longer time in environment and hardly degradable
(recalcitrant xenobiotics). Chief sources involved in the introduction of xenobiotic
compounds into the environment are pharmaceutical and chemical industries, pulp
and paper bleaching industries, agriculture and forestry (fertilizers, pesticides and
herbicides).
Xenobiotics were developed for the well-being of human beings in the last
century but their escalated use is disrupting the biosphere resulting in everlasting
contamination. Chemical industries are synthesizing 1,000 new chemicals every year
and about 60,000 to 95,000 chemicals are currently in commercial use subsequently
releasing more than one billion pounds in air and water. The environmentalists are
trying to raise the awareness about toxicity of these chemicals therefore numerous
persistent organic pollutants (POPs) have been banned around the world [2].
1.2 Pesticides
Pesticides are most commonly used amongst all the xenobiotics due to their
multipurpose applications in agriculture, forestry and domestic areas. A pesticide is
defined as any compound used for preventing, destroying, repelling or mitigating
any pest, covering a broad range of insecticides, fungicides, herbicides, rodenticides,
molluscicides, nematicides, plant growth regulators and others. Pesticides particularly
refer to chemical substances that modify biological processes of living organisms
reckoned to be pests (insects, mould, fungi, weeds or noxious plants) [3].

___ 1. Introduction
2
Pesticides are extensively used in different areas of agriculture to avoid yield
losses and low quality products. Crops are damaged by 9,000 species of insects and
mites, 50,000 species of plant pathogens and 8,000 species of weeds approximately.
Crop loss is minimized to 35% to 42% with use of pesticides [4]. The worldwide
consumption of pesticides was found to cost around 5.2 billion pounds in 2007 with
herbicides contributing major portion of total cost as compared to other classes of
pesticides. Agricultural countries rely on pesticides conspicuously to secure and
enhance crop yields and consequently strengthen economy. Pesticides are not newly
invented but deliberately used thousand years ago by Sumerians, Greeks, and Romans
who used a variety of compounds such as sulphur, mercury, arsenic, copper and plant
extracts to kill pests. But the results were not encouraging due to insufficient
knowledge about their chemistry and applications. Introduction of DDT
(dichlorodiphenyltrichloroethane), BHC (benzene hexachloride), aldrin, dieldrin,
endrin and 2, 4-D (2, 4-dichlorophenoxyacetic acid) augmented the use of pesticides
after World War II. These newly introduced chemicals were found to be efficient,
user-friendly and economical [5].
1.3 Classification of Pesticides
Classification of pesticides is done on the basis of their physical properties, chemical
structure, target organism and mode of action. Target organism based classification of
pesticides includes insecticides (kill the insect pests of crops, flies, mosquitoes and
insect vectors for human diseases), herbicides (alleviate the superfluous plants),
fungicides (kill fungus), avicides (kill bird pests) and acaricides (diminish tick and
mites). Although pesticide classification involves different grounds but chemical
structure based classification is more favoured by the scientists because it establishes
a correlation among structure, activity, toxicity and degradation mechanisms of
pesticide groups. Foremost pesticides according to chemical composition are
organophosphates (OPs), organochlorines (OCs), carbamates and pyrethroids.
Organochlorides (OCs) were used effectively to control different diseases such
as malaria and typhus, banned after 1960s in developed countries due to its
toxicological effects but still used in developing countries. Organochloride pesticides
are recalcitrant, hardly degradable and pretense chronic health effects including

___ 1. Introduction
3
cancer, neurological and teratogenic effects [6-9]. Most popular OC pesticides are
DDT, eldrin and endosulfan.
Other pesticides introduced in different decades as organophosphate (OPs) in
1960s, carbamates in 1970s, pyrethroids in 1980s and herbicides and fungicides in
1970s-1980s facilitated in controlling pests and boosting agricultural output.
Organophosphorus pesticides are esters of phosphoric acid and have adverse effects
on the nervous system of pests and human beings, influencing their reproductive
system as well [10, 11]. They inhibit the activity of cholinesterase (ChE) enzyme
accountable for the nerve impulse in living organisms [12]. Generally used OP
pesticides include chlorpyrifos, diazinon, profenofos, parathion, malathion and
triazophos.
Carbamate pesticides contain a wide variety of compounds including carbaryl,
carbofuran and aldicarb posing low toxicity to living organisms [13]. Although, they
inhibit the activity of enzyme acetylcholinesterase but inhibition is less severe and
reversible. Accumulation of acetylcholine due to inhibition of its hydrolysis reaction
causes various symptoms, such as sweating, lacrimation, hypersalivation and
convulsion of extremities [14].
Pyrethroids are classified as lipophilic esters containing an alcohol and an acid
moiety. Pyrethroid pesticides block neuronal activity by displaying high affinity and
binding to Na+ channels. Thus prolonged channel opening result in complete
depolarization of the cell membrane thus blocking neuronal activity. Even though less
toxic and persistent than other groups of insecticides but still poses harmful effects.
1.4 Organophosphonate Pesticides
Organophosphonates embodies a group of organic compounds distinguished by direct
carbon to phosphorus (C-P) covalent bond. These compounds are resistant to thermal
decomposition, chemical hydrolysis, enzymatic degradation and photolysis due to the
presence of stable C-P bond. Although there is no significant difference between
strength of C-P bond energy and other bond energies but former has higher activation
energy [15]. These compounds are extensively used as pesticides, lubricant additives,
flame retardants, plasticizers, corrosion inhibitors, drugs antibiotics, adhesives,
chelating agents etc. Some of the most discernible organophosphonates include

___ 1. Introduction
4
expansively used herbicides such as glyphosate and phosphinothricin [16]; insectides
such as ethylphosphonate and phenylphosphonate derivatives; flame retardant such as
Phyrol 76, an oligomer of vinylphosphonate-methylphosphonate; corrosion inhibitors
such as polyaminopolyphosphonic acids [17]; Aminotri (methylenephosphonic and
hydroxyethylidenediphosphonic acids) as chelative agents to household detergents
[18], bisphosphonates for the treatment of bone mineralization disorders;
alaphosphaline and phosphonomycin (biphosphonates) as antibiotics and cyclic esters
of aromatic bisphosphonates as polymer additives [19, 20].
In 1944, Pikl synthesized first organophosphonate compound
aminomethylphosphonic acid but later on Kosolapoff and Chavane produced diverse
amino and amino-substituted phosphonic acids [21]. First biogenic
organophosphonate 2-amino ethylphosphonic acid (2-AEP) was isolated by
Horiguchi in 1959 from acid hydrolysates of ether ethanol fraction of Protozoa [22].
Later on 2-AEP was also found in Tetrahymena pyriformis and Anthopleura
elegantissima (sea anemone) in the form of phosphonolipids.
Regarding the chemical structure, Organophosphonates are distinguished by
the presence of C-P bond which substitute one of the four C-O-P bonds commonly
found in most esters [23]. They contain C-PO(OH)2 or C-PO(OR)2 groups where R is
alkyl or aryl group, therefore R1 is directly bonded to the P atom while R2 is linked
either to an oxygen or sulphur atom (Figure 1-1).
Figure 1-1 General chemical structure of organophosphonate pesticides.

___ 1. Introduction
5
1.5 Herbicides
Since the existence of human race, its decisive enterprise has been the plant
production for food and fiber. Humans have battled to control weeds threatening
crop survival and productivity since the dawn of agriculture. Crops are greatly
affected by weeds every year therefore herbicides are extensively used for weed
management in agriculture sector [24]. They are used in industry, forestry and urban
areas as well. Herbicides are organophosphonate compounds that hinder or disrupt
the normal growth and development of plants resulting in their obliteration.
Contemporary agriculture practices greatly depend on herbicides for cost effective
weed control and soaring crop yield. Herbicides are classified in numerous ways
depending on their application method (soil, foliar, broadcast, spot or band),
application time (pre-plant incorporation, pre-emergent or post emergent), chemical
composition (organic or inorganic), general symptoms in plants (contact or
systemic), period of soil persistence (persistent, residual or long residual), response
among plants (nonselective or selective), formulation (liquid or dry) and mode of
action. Although different approaches are applied for herbicide classification but
classification hinged on herbicidal mode of action is relatively better and imperative
due to its significance in weed resistance management, exploring toxicological
problems and improving herbicide application techniques [25]. About 20 different
target sites for herbicides are reported [26] but herbicides with only 6 modes of
action embodies approximately 75% of herbicide sales [27].
Herbicides are classified into different groups on the basis of site of action. A
classification system was published by the International Herbicide Resistance Action
Committee (HRAC) based on letters for each group [28]. Classification was updated
by Mallory-Smith and Retzinger [29] afterwards and some herbicides listed in the
Weed Science Society of America 2002 Herbicide Handbook [30] were also added.
The modes of action of herbicides are discussed below:
1.5.1 Group 1: Acetyl CoA Carboxylase (ACCase) Inhibitors (Lipid
Biosynthesis Inhibitors)
Herbicides of this group obstruct the synthesis of fatty acids by suppressing acetyl
coenzyme A carboxylase (ACCase) enzyme activity. This blockage hinders the
production of phospholipids involved in synthesis of lipid bilayer, which plays an

___ 1. Introduction
6
important role in cell structure and function [31-33]. Aryloxyphenoxypropionate
(FOPs), cyclohexanedione (DIMs), and phenylpyrazolin (DENs) are ACCase
enzymes used for eliminating grass during the cultivation of broadleaf crop varieties
or crop rotation. Certain grasses and broadleaf crop varieties have natural resistance
to these herbicides due to their strong ACCase system [34].
1.5.2 Group 2: Acetolactate Synthase (ALS) or Acetohydroxy Acid
Synthase (AHAS) Inhibitors
This group includes imidazolinones, pyrimidinylthiobenzoates,
sulfonylaminocarbonyltriazolinones, sulfonylureas, and triazolopyrimidines which
hinders the activity of acetolactate synthase (ALS) also known as acetohydroxy acid
synthase (AHAS) involved in the formation of 2-acetolactate or 2-aceto-2-
hydroxybutyrate [35]. Therefore inhibition of AHAS results in obstruction of
branched-chain amino acids (leucine, isoleucine, and valine) synthesis [36] and this
amino acid deficiency halts the synthesis of proteins ultimately causing plant death.
1.5.3 Group 3: Mitosis Inhibitors (Root Growth Inhibitors)
They are used as preemergent or preplant herbicides in vegetables and ornamental
plants and their mode of action involves the inhibition of cell division. This group
includes benzamide, benzoic acid [dimethyl-2, 3, 5, 6-tetrachloroterephthalate
(DCPA)], dinitroaniline, phosphoramidate, and pyridine herbicides which binds to
major microtubule protein tubulin. This herbicide-protein complex hinders the
polymerization of microtubules during assembly but remains impervious during
depolymerization [37], resulting in loss of structure and function of the
microtubules. Therefore spindle apparatus is not formed and cell death occurs due to
inhibition of cell wall formation.
1.5.4 Group 4: Plant Growth Regulators (Synthetic Auxins)
Benzoic acid, phenoxycarboxylic acid, pyridine carboxylic acid and quinoline
carboxylic acid of this group halts the activity of indole acetic acid (IAA) [38]. They
are used to remove broadleaf weeds during the cultivation of corn, wheat, and
sorghum. Specific binding site involving activity of IAA and synthetic auxins is still
unknown however these herbicides disrupt the plasticity of cell wall and nucleic acid
metabolism. They activate the adenosine triphosphate (ATPase) proton pump which
results in increased enzymatic activity of cell wall [39]. Activity of IAA is mimicked

___ 1. Introduction
7
by these regulators and RNA, DNA, and protein biosynthesis is increased leading to
continuous vascular growth causing cell bursts and ultimate cell and plant death.
1.5.5 Groups 5, 6, 7: Photosynthesis inhibitors (Photosystem II
(PSII) Inhibitors)
These herbicides inhibit the Photosystem II (PSII), photosynthetic pathway. Group 5
exemplifies triazine, triazinone, phenylcarbamates, pyridazinones, and uracils.
Nitriles, benzothiadiazinones, and phenylpyridazines are included in Group 6.
Group 7 represents phenyl urea and amides. All groups of PSII inhibitors exhibit
different binding patterns among them except a few similarities. They binds with
QB binding site of D1 protein of photosystem II complex located in chloroplast
thylakoid membranes. This binding interrupts transport of electrons from QA to QB
and blocks the CO2 fixation, ATP generation and nicotinamide adenine dinucleotide
hydrogen phosphate (NADPH2) production essential for plant growth and
development [40, 41]. Failure of QA reoxidation leads to the formation of triplet
state chlorophyll which converts ground state oxygen to singlet oxygen. This leads
to the peroxidation of lipids with a release of lipid radical. Whole mechanism
results in the loss of chlorophyll and carotenoids from the cell membranes and
exposure of cell contents to ruthless environmental conditions ultimately causing
plant death [42].
1.5.6 Groups 8 and 15: Shoot Growth Inhibitors
Herbicides of both groups are applied in the soil prior to the emergence of grass and
broadleaf weeds to remove them effectively. The herbicides included in Group 8 are
phosphorodithioates and thiocarbamates which disturb the lipid synthesis
mechanism and halt the biosynthesis of lipids, fatty acids, proteins, isoprenoids,
flavonoids, and gibberellins [43]. Group 15 herbicides include chloroacetamide,
acetamide, oxyacetamide, and tetrazolinone which act on a very long chain fatty
acid present in the cell membrane [44]. They make a complex with acetyl COA and
some sulfhydryl-containing molecules via thiocarbamate sulfoxides and inhibit the
long chain fatty acids during the seedling shoot growth stage of the plant and
distress the weeds preemergence.

___ 1. Introduction
8
1.5.7 Group 9: Aromatic Amino Acid Inhibitors
This group of herbicides inhibits the synthesis of aromatic amino acids by disturbing
the shikimate pathway (aromatic amino acid synthesis pathway) [45]. They are non
specific herbicides which kills every plant they come in contact with therefore they
are suggested to be used in glyphosate resistant crops as cotton, canola, corn and
soybean. This group includes glyphosate which is available as ammonium salts,
diammonium salts, dimethylammonium salts, isopropylamine and potassium salts.
Due to their broad spectrum nature, they are most commonly used in the world in
agriculture and forestry.
1.5.8 Group 10: Glutamine Synthesis Inhibitors
Glufosinate and bialophos (phosphinic acids) of this group disrupts the nitrogen
metabolism in plants. Their site of action is glutamine synthase which converts
ammonia and glutamate to glutamine [46]. Glutamine synthase re-assimilate the
ammonia generated during respiration and these herbicides impair its activity. They
degrade the proteins leading to the accumulation of ammonia resulting in lower pH
in the cell. This leads to the shutting down of the PSI and PSII systems eventually
causing the uncoupling of photophosphorylation [47]. These herbicides are effectual
for managing weeds and other unwanted plants due to the presence of glutamine
synthase in chloroplast and cytoplasm. They can also be used as postemergent
herbicides in glufosinate resistant crops.
1.5.9 Groups 12, 13, and 27: Pigment Synthesis Inhibitors
These herbicides destroy the photosynthesis pigment in plants, the chlorophyll. They
are also recognized as carotenoid biosynthesis inhibitors or bleaches as they remove
the green color of the plant tissues, destroying the cells and ultimately plant death.
They also inhibit the catalysis of 4-hydroxyphenyl pyruvate dioxygenase (HPPD)
enzyme therefore also known as HPPD inhibitors.
Group 12 includes amides, anilidex, furanones, phenoxybutan-amides,
pyridiazinones, and pyridines which inhibits the activity of phytoene desaturase
enzyme and interrupt the carotenoid biosynthetic pathway [48]. Group 13 represents
the chemical family of Isoxazolidinone which disrupts the diterpene synthesis.
Group 27 includes Isoxazole which inhibits HPPD enzyme. The application of these

___ 1. Introduction
9
groups of herbicides reduces the carotenoid levels in cells and increases the number
of unbound lipid radicals. Lipid peroxidation caused by these lipid radicals leads to
the impaired function of chlorophyll and membrane lipids. The cell contents are
destroyed due to membrane leakage causing plant death.
1.5.10 Group 14: Protoporphyrinogen Oxidase (PPO) Inhibitors
These herbicides are also called cell membrane disrupters as they disrupt the cell
membrane. They inhibit the PPO enzyme which is involved in the biosynthesis of
chlorophyll and heme. The herbicides included in this group are diphenylether, aryl
triazolinone, N-phenylphthalimides, oxadiazoles, oxazolidinediones,
phenylpyrazoles, pyrimidindiones and thiadiazoles. Inhibition of PPO eventually
leads to lipid peroxidation, cell disintegration and ultimately plant death [49]. Group
14 chemicals are used preemergent as well as postemergent herbicides.
1.5.11 Group 22: Photosynthesis Inhibitors (Photosystem I (PSI)
Inhibitors)
They are also known as cell membrane disrupters as they penetrate through plant
foliage and disrupt the lipid bilayer of cell resulting in cell membrane disruption.
Bipyridilium chemical family represents this group which produces herbicide
radicals by accepting electrons from PSI. Therefore they are also called PSI electron
diverters. These herbicides interact with molecular oxygen and produce superoxide
radicals further converted to hydrogen peroxide and hydroxyl radicals in the
presence of superoxide dismutase enzyme [50]. These radicals interrupt the
unsaturated fatty acids, chlorophyll, lipids, and proteins in the cell membrane. The
cell membrane disruption leads to the cell cytoplasm leakage and ultimately death of
plant. Photosynthesis inhibitors are nonselective in nature therefore applied
generally before crop harvest.
1.6 Glyphosate
Glyphosate [N-(phosphonomethyl) glycine] is a systemic, broad spectrum, non-
selective and postemergent herbicide. It is recurrently used in agricultural, industrial,
silvicultural amenity and domestic areas to control weeds [51, 52]. It is found in the
form of white crystalline powder with a chemical formula of C3H8NO5P and
molecular weight of 169.07 g/mol. The solubility of glyphosate in water is about 1.01

___ 1. Introduction
10
g/100mL at 20 °C. Its melting temperature is 184.5 °C and boiling temperature is 187
°C. Glyphosate was formerly used as plant growth promoter to increase production of
sucrose in cane but later on introduced as herbicide by John E Franz of Monsanto in
1970 [53]. It is most commonly used herbicide applied to control annual and perennial
weeds. Glyphosate is usually formulated in the form of its isopropylamine salt and is
the active ingredient of more than 50 formulations sold under various trade names
such as Roundup Ultra®, Roundup Pro®, Accord
®, Honcho
®, Pondmaster
® and
Protocol® etc [54]. In agricultural areas glyphosate is generally applied in fields
before sowing, between the rows in row crops and around perennial crops. In non-
agricultural areas, it is used along irrigation channels, roadsides and around recreation
areas. Glyphosate use has significantly increased with the introduction of glyphosate
tolerant crops [55, 56].
1.6.1 Mode of Action of Glyphosate
Mode of action of glyphosate involves the inhibition of 5-enolpyruvoyl-shikimate-3-
phosphate synthase (EPSPS, E.C. 2.5.1.19). EPSPS is a plant enzyme that catalyzes
the formation of EPSP from phosphoenolpyruvate (PEP) and shikimate 3-phosphate
(S3P) (Figure 1-2).
___ 1. Introduction
11
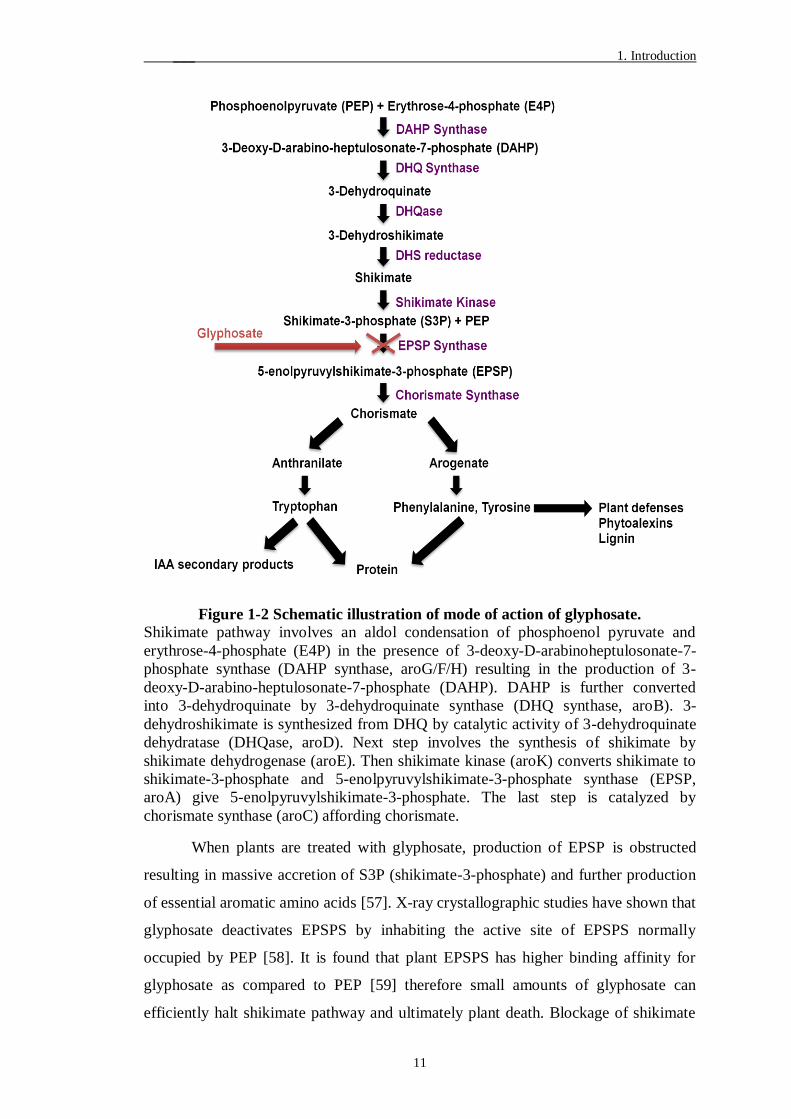
Figure 1-2 Schematic illustration of mode of action of glyphosate.
Shikimate pathway involves an aldol condensation of phosphoenol pyruvate and
erythrose-4-phosphate (E4P) in the presence of 3-deoxy-D-arabinoheptulosonate-7-
phosphate synthase (DAHP synthase, aroG/F/H) resulting in the production of 3-
deoxy-D-arabino-heptulosonate-7-phosphate (DAHP). DAHP is further converted
into 3-dehydroquinate by 3-dehydroquinate synthase (DHQ synthase, aroB). 3-
dehydroshikimate is synthesized from DHQ by catalytic activity of 3-dehydroquinate
dehydratase (DHQase, aroD). Next step involves the synthesis of shikimate by
shikimate dehydrogenase (aroE). Then shikimate kinase (aroK) converts shikimate to
shikimate-3-phosphate and 5-enolpyruvylshikimate-3-phosphate synthase (EPSP,
aroA) give 5-enolpyruvylshikimate-3-phosphate. The last step is catalyzed by
chorismate synthase (aroC) affording chorismate.
When plants are treated with glyphosate, production of EPSP is obstructed
resulting in massive accretion of S3P (shikimate-3-phosphate) and further production
of essential aromatic amino acids [57]. X-ray crystallographic studies have shown that
glyphosate deactivates EPSPS by inhabiting the active site of EPSPS normally
occupied by PEP [58]. It is found that plant EPSPS has higher binding affinity for
glyphosate as compared to PEP [59] therefore small amounts of glyphosate can
efficiently halt shikimate pathway and ultimately plant death. Blockage of shikimate

___ 1. Introduction
12
pathway prevents the production of chorismate which is an important intermediate
involved in the synthesis of essential aromatic amino acids (phenylalanine, tyrosine
and tryptophan). These aromatic amino acids are further implicated in the production
of several secondary plant products such as anthocyanins, lignin, growth promoters,
growth inhibitors, phenolics and protein production as well. Aromatic amino acids
and their secondary products molecules resulting from shikimate pathway contribute
approximately 35% of the total plant dry weight [53]. After the application of
glyphosate, injury symptoms appear as wilting, chlorosis and finally death of plants
within two weeks [54]. Glyphosate also affects bacteria and fungi due to the presence
of shikimate pathway.
1.6.2 Uptake and Translocation of Glyphosate
Glyphosate due to its non-selective nature is used to control all types of weeds. It
systemically moves through the leaves and shoots up to the roots after foliar
application. Franz et al. has reported that glyphosate translocates through the phloem,
accumulates in the meristematic regions of the plants and inhibits shikimate pathway
[53]. Shikimate pathway is the source of about 20% of all fixed carbon in plants and
its inhibition interrupts this carbon flow ensuing in the accumulation of huge amounts
of shikimate and eventually plant death [60]. Uptake rate of glyphosate differ between
species due to its susceptibility in different species and environmental factors such as
temperature, light intensity, wind speed and humidity etc. Glyphosate is usually
applied in formulation with some surfactants to aid in tissue penetration and avoid
droplet formation on waxy cuticle of leaves [61].
1.6.3 Fate of Glyphosate in Soil
The soil is the final destination of glyphosate translocation after its application in
forestry and agriculture. Glyphosate is a small molecule with three polar functional
groups (carboxyl, amino and phosphonate groups) which binds readily with soil
particles [62]. The persistence and transportation of glyphosate in soil depends on
different factors such as composition of soil, climate conditions, microbial activity
and agricultural management [62, 63]. Glyphosate competes with inorganic phosphate
for binding sites due to the presence of same phosphonic acid moiety which may
influence its sorption and mobility in phosphate rich soil [64]. Phosphate has ability to
outcompete glyphosate for sorption depending on soil properties and environmental

___ 1. Introduction
13
conditions. Glyphosate sorption strongly depends on soil pH and declines with
decrease in pH while sorption of phosphate is not affected by soil pH [65]. Moreover
different glyphosate degradation pathways (AMPA pathway, sarcosine pathway) are
employed by soil microbes therefore presence or absence of glyphosate and phosphate
in soil creates selection pressure on soil biota [66, 67].
Although glyphosate binds tightly with soil particles but it is hard to
generalize the glyphosate sorption due to different soil types. It is found that
glyphosate binds efficiently with soils containing less inorganic phosphate, high Al3+
and Fe3+
concentrations, broad surface area and high pH. Binding pattern of
glyphosate with clay and organic matter depends upon the type of cations and as a
polyprotic acid it has high affinity for trivalent cations such as Al3+
and Fe3+
[68].
Glyphosate sorption is found to increase with increase in surface area and soil pH
[69]. Average half life of glyphosate in soil is reported between 2-197 days [70] and
therefore its typical field half life is suggested of 47 days [30].
1.6.4 Fate of Glyphosate in Water
Glyphosate is believed to have low probability of leaching due to its high soil sorption
property but it has been detected in surface waters of irrigation channel near
application site in agricultural fields [71]. The binding capacity of glyphosate in soil
depending on soil, climatic and spray conditions might contaminate groundwater
through leaching [63, 65, 72]. Glyphosate has an estimated half life of 7-14 days in
surface water [70]. Glyphosate leaching has been reported in gravelly materials with
less retention capacity [73]. Moreover unnecessary irrigation or rainfall
instantaneously after application, washing and cleaning the tanks of the fumigation
machines in streams and adjacent water can also result in contamination of aquatic
systems [74, 75].
About 3% of the total applied glyphosate has been found in aquatic
environments [76]. France has declared the distribution and leaching of glyphosate in
water bodies as an environmental risk [77]. The long term study of pesticides
including glyphosate, conducted by Danish government revealed that concentration of
glyphosate (4.7 µg/L) in water drainage waterways near glyphosate treated sites was
five times higher than maximum contaminant limit (MCL) for glyphosate (0.1 µg/L)
in European Union conducts long-term monitor of a variety of pesticides, including

___ 1. Introduction
14
glyphosate [78]. Battaglin et al. found glyphosate (0.1-8.7 µg/L) in 36% of tested
water samples collected from 51 streams of Midwestern United States during growing
season in 2002 [79]. The presence of glyphosate in runoff from fields with different
crops and tillage practices was examined by Shipitalo and Owens. They found
maximum concentration of glyphosate (887 µg/L higher than US MCL of 700 µg/L)
during rainfall [80]. Moreover, glyphosate and AMPA are found among the
recurrently reported pesticides detected in water pollution monitoring [81]. Therefore
glyphosate leaches from sites of application to nearby water ways resulting in aquatic
toxicity.
1.6.5 Global Use of Glyphosate
Atrazine and metolachlor were most profoundly used global herbicides in 1970s. In
1995 about 60 million kg of these herbicides was applied on maize crop in U.S. [82].
51.3 million kg (113 million pounds) of glyphosate was applied by farmers globally in
1995. Global use of glyphosate in agricultural sector heightened with the introduction
of glyphosate tolerant crops. Glyphosate tolerant soybean, maize, and cotton varieties
were approved for planting in the U.S. in 1996 whereas glyphosate resistant corn was
introduced in 1998. These roundup ready crops contribute about 93% of soybeans,
82% of cotton, and 85% of corn to planted crops. Genetically engineered glyphosate
tolerant alfalfa and sugar beets were approved in 2005 and commercialized in 2008.
This breakthrough led to 14.6 fold increase in glyphosate use from 51 million kg (113
million pounds) in 1995 to 747 million kg (1.65 billion pounds) in 2014 encasing
about 1.4 billion hectares of cropland worldwide. Non agricultural use of glyphosate
has also boosted globally up to five folds from 16 million kg in 1995 to 79 million kg
in 2014. Total global use of glyphosate in agricultural and non-agricultural sectors
ascended more than 12 folds from about 67 million kg in 1995 to 826 million kg in
2014. This trend of glyphosate use was found adequate to treat 22 and 30% of
globally cultivated cropland. No such widely used herbicide has been reported in
history so far [82].
Various factors are involved in the elevated use of glyphosate after its
commercialization in 1974. Introduction of glyphosate tolerant crops has mushroomed
the use of glyphosate not only by increasing treatment area but also one time
application rate per hectare. In soybean sector of U.S. the glyphosate application

___ 1. Introduction
15
number mounted from 1.1 per crop year in 1996 to 1.52 in 2014 whereas single rate of
application increased from 0.7 kg/hectare (0.63 pound/acre) to 1.1 kg/hectare (0.98
pound/acre) simultaneously [82]. Emergence of less sensitive and glyphosate resistant
weeds has elevated the use of glyphosate on glyphosate tolerant crops [83]. Other
factors involved in the increased use of glyphosate include steady expansion in the
number of crops registered for use on glyphosate product labels, short half life in soil,
low leaching and transport potential in rhizosphere, absence of shikimate pathway in
animals and humans hence low toxicity, the adoption of no-till and conservation
tillage systems, the declining price per pound of active ingredient, new application
method and timing options [84-87].
1.6.6 Environmental Impact and Toxicity of Glyphosate
Excessive and continuous use of glyphosate is drastically effecting environment.
Once applied on crops, glyphosate enters in ground water due to its water soluble
properties. U.S. Geological Survey (USGS) found glyphosate content in air and water
samples during two growing seasons in states of Iowa and Mississippi and its higher
levels were observed during rainfall than for any other previously monitored
pesticide. Glyphosate is also found to turn down the pollinator habitats. It removes
milkweeds from crop fields which are key source of food for butterflies. Glyphosate
resistant crops grown in close proximity of organic crops increase the risk of genetic
cross-contamination, as pollens from glyphosate resistant crops have the potential to
drift to non resistant crops and produce offspring. Glyphosate is converted into more
toxic and persistent metabolite aminomethyl phosphonic acid (AMPA) which is found
persistent in soil and also effects human health (Figure 1-3).

___ 1. Introduction
16
Figure 1-3 Accumulation and distribution of glyphosate in environment.
Glyphosate sprayed on plants may directly enter the soil or absorbed by plants
through foliage and transported systemically to roots through phloem. From roots it
enters in soil and adsorbed by soil or degraded by microbes to
aminomethylphosphonic acid (AMPA). Glyphosate is also found in different plant
parts and taken up by herbivores. Glyphosate may be taken up by non-target plants
affecting their growth and resistance against pathogens. Glyphosate fate in soil
depends on soil type and presence of sorption surfaces. Glyphosate residues may
become part of animal feed through manure or end up to human consumption.
Fertilizers added to soil and glyphosate residues may enter aquatic system through
erosion or leaching and causes hardening of water. This hard water disturbs aquatic
life as well as human beings through drinking water. Glyphosate residues in soil may
enhance virulence of disease borne organism and disturb soil microflora. Glyphosate
consumed by human beings through inhalation or dermal absorption during spray may
adversely affect human health.
Glyphosate herbicides contain glyphosate as an active ingredient along with
certain additives (mostly surfactants). Richard et al. reported that sole glyphosate is
not as much toxic to humans as glyphosate in the form of its formulations [88].
Glyphosate herbicides were reckoned to be the reason of for pregnancy problems in
some agricultural workers [89]. Glyphosate was found to affect endocrine and human
placental JEG3 cells at concentrations lower than those used for agricultural purposes
[88]. Watts showed that glyphosate can change the DNA of sister chromatids in
human lymphocytes. In glyphosate formulations, different surfactants are used to
enhance glyphosate solubility and diffusion in plants [90]. Polyethoxylated tallow

___ 1. Introduction
17
amine (POEA) is a surfactant commonly used in glyphosate herbicides. It is found to
have toxic effects on human peripheral blood mononuclear cells (56.4 µg/mL
concentration at lab scale) [91, 92]. Moreover 0.5 and 10 µg/mL of glyphosate
concentration has been found to be toxic for human liver Hep G2 cells and also
disrupts the endocrine system of human beings [93]. Kimmel et al. reviewed a 2002
finding describing the role of glyphosate in cardiovascular defects during pregnancy
and found that glyphosate exposure does not pose potential risks for cardiovascular
defects during pregnancy [94]. Furthermore glyphosate and its formulations were
found to pose no genotoxic effects on human beings under normal conditions of
humans or environmental exposures [95]. Glyphosate is found to disturb gut
microflora by killing beneficial microbes and enhancing growth of pathogenic
bacteria in animals [96]. It is also reported to bind with iron and cobalt resulting in
suppression of cytochrome P450 (CYP) enzymes [97]. Schinasi and Leon reported
that there is a momentous relationship between non-Hodgkin lymphoma (NHL) and
occupational exposure of glyphosate [98]. In March 2015 the International Agency for
Research on Cancer (IARC) of World Health Organization (WHO) has placed
glyphosate in category 2A and classified it as possibly carcinogenic to humans on the
basis of epidemiological, animal and in vitro studies [99].
Glyphosate was claimed to be non toxic for terrestrial and aquatic animals by
Monsanto [100]. But different studies proved that it is toxic to non target organisms.
Carcinogenic effects on non human mammals include renal tubule carcinoma and
haemangiosarcoma incidence in male mice and prevalence of pancreatic islet-cell
adenoma in male rats [101]. Glyphosate is found to perturb aquatic communities as
compared to 2,4-D. Biodiversity of more than 25 animal species and algae of aquatic
environment were found to decrease up to 22% due to glyphosate exposure [102].
Soil ecosystem upholds different biological and biochemical processes and
soil microbes play an important role in accomplishment of these processes
(xenobiotics degradation, transformation and release of nutrients from complex
organic compounds). Agricultural practices used to enhance crop yield are seriously
affecting the quality and health of soil [103]. Although different parameters can be
employed to study these effects on soil but microbes are found to be more proficient
indicators [104]. Glyphosate toxicity to soil biota varies among different species and
organisms. Life span of some soil invertebrates such as springtail, Onychiurus

___ 1. Introduction
18
quadriocellatus and beneficial predatory mite, Amblyseius fallacies was found to
decrease due to toxic effects of glyphosate [105]. Glyphosate also pose risk to
beneficial insects and earthworms in soil [106, 107]. Repeated use of glyphosate (1
µg/g) also reduces beneficial microorganisms from soil such as saprophytic,
mycorrhizal fungi and nitrogen-fixing bacteria [108]. Glyphosate formulations
stimulate the growth of fungal pathogens causing diseases in plants.
Taking into account all aforementioned ramifications of intemperate use of
glyphosate on environment, there is a dire need to identify effective methods for its
detoxification. Various conventional methods such as photolysis, thermal
decomposition, incineration, chemical degradation, dispersion, diffusion and
volatilization, landfill, sedimentation, adsorption etc are available to remove
contaminants from the environment. But the limitations of these methods involve
formation of toxic products [109] as well as their costly, lengthy and environment
unfriendly nature [110]. In case of glyphosate, it is highly resistant to these non
biological methods due to the presence of stable C-P linkage [111]. Therefore use of
microorganisms able to degrade phosphonate compounds seems to be the appropriate
way out of this problem.
1.7 Biodegradation
Biodegradation is a natural attenuation process which involves utilization of
pesticides by microorganisms such as bacteria, fungi etc. as energy source via
conversion of pesticides into less toxic compounds. Soil is reported to hold more than
one hundred million bacteria (5000-7000 different species) and more than ten
thousand fungal colonies [112]. Removal of pollutants by using microorganisms is
safer and economical as compared to physic-chemical processes [113].
Biodegradation competence rely on different factors such as reactivity of pesticides in
the environment; condition of rhizosphere including soil type, precipitation, existence
of degrading microorganism, temperature, nutrient availability and pH of field; time
and method of pesticide application in fields [114].
The microorganisms required for biodegradation must have high survival rate
in the environment, pathogenicity and toxicity deficient, high herbicide degradation
efficiency irrespective of external environmental conditions and herbicide
mineralization capability without accumulation of toxic metabolites [115]. But these

___ 1. Introduction
19
requirements are not fulfilled concurrently. However such types of bacterial strains
are required to remove herbicides from contaminated sites in order to avoid their
leaching into ground water bodies. Moreover bioremediation procedures can be used
to remove herbicides in case of their leakage at production, storage and application
sites.
1.7.1 Biodegradation of Glyphosate
When glyphosate comes in contact with soil, it is inactivated after strongly binding
with soil particles. However soil microorganisms use it as energy source (C, P and N)
after degradation. Rate of biodegradation of glyphosate depends upon its persistence
in soil and diversity of soil microbes. Glyphosate biodegradation through soil
microbes is mostly carried out by two metabolic pathways [67] (Figure 1-4). First
pathway involves the cleavage of C-N bond of glyphosate by glyphosate
oxidoreductase (GOX) enzyme resulting in the production of aminomethylphosphonic
acid (AMPA) and glyoxylate. Glyphosate oxidoreductase is a flavoprotein which
utilizes FAD as a cofactor and this cofactor is reduced by glyphosate at its active site.
Oxygen is used as a cofactor under aerobic conditions while phenazine methosulfate
and ubiquinone are used as electron acceptors under anaerobic conditions. Glyphosate
oxidoreductase enzyme has been used for production of glyphosate resistant crops
[116]. Glyoxylate is a glyphosate degradation product and endogenous metabolite of
plants as well also engaged in different metabolic pathways [117]. AMPA is further
converted to methylamine through C-P lyase activity which is eventually converted to
formaldehyde by methylamine dehydrogenase enzyme [118]. Formaldehyde further
reacts with water or hydroxyl radicals and converted to methanol. Therefore
glyphosate biodegradation ultimately yields carbon dioxide, phosphate, ammonia and
methanol [119].

___ 1. Introduction
20
C-P lyase Glyphosate
oxidoreductase
Sarcosine Phosphate Aminomethyl
Phosphonic
acid
Glyoxylate
Sarcosine
oxidase
Glycine Formaldehyde Formaldehyde Phosphate
Phosphonatase
C-P lyase pathway AMPA pathway
Glyphosate
Figure 1-4 Pathways of glyphosate biodegradation.
One pathway involves the conversion of glyphosate into sarcosine by the action of
microbial C-P lyase gene. Sarcosine is further converted to glycine and formaldehyde.
Other pathway of glyphosate biodegradation encompasses the cleavage of C-N bond
resulting in production of aminomethyl phosphonic acid (AMPA). AMPA is further
degraded to phosphonoformaldehyde and formaldehyde by action of aminotransferase
and phosphonatase respectively.
Second pathway involves the degradation of glyphosate to sarcosine (N-
methylglycine) through C-P lyase enzyme activity [120]. This reaction is thought to
be a radical based redox dependent dephosphorylation, involving the formation of a
phosphonyl radical [121]. The activity of phn genes is regulated by Pho regulon and
their expression is synchronized by exogenous phosphate. Therefore activity of C-P
lyase gene is stimulated under phosphate starvation and microorganisms utilize
phosphonates as an alternative phosphorus source. Sarcosine is further converted to
glycine and formaldehyde by sarcosine oxidase activity. Formaldehyde enters the
tetrahydrofolate directed pathway of single carbon transfers and glycine is
metabolized to carbon dioxide and ammonia [65].

21
2. Materials and Methods
2.1 Chemicals Used in This Study
Technical (95%) and analytical (99.5%) grade standards of glyphosate were
purchased from Pak China Chemicals, Lahore, Pakistan and Chem Service (West
Chester PA) respectively. 4-chloro 3,5 dinitrobenzo trifluoride (99%) , boric Acid
(99.5%), potassium dihydrogen phosphate (99%), HCl (32%), and sodium tetraborate
decahydrate (99%) were procured from Sigma-Aldrich USA. Acetonitrile of HPLC
grade was purchased from Merck Germany. Rest of the chemicals utilized in bacterial
growth media preparation was obtained from Merck. Enzymes, PCR and cloning kits
used in molecular analysis were acquired from Fermentas.
2.2 Bacterial Strains Used in This Study
Glyphosate tolerant/degrading bacterial strains were isolated from glyphosate
contaminated soil samples collected from Pakistan and Australia (Table 2.1).
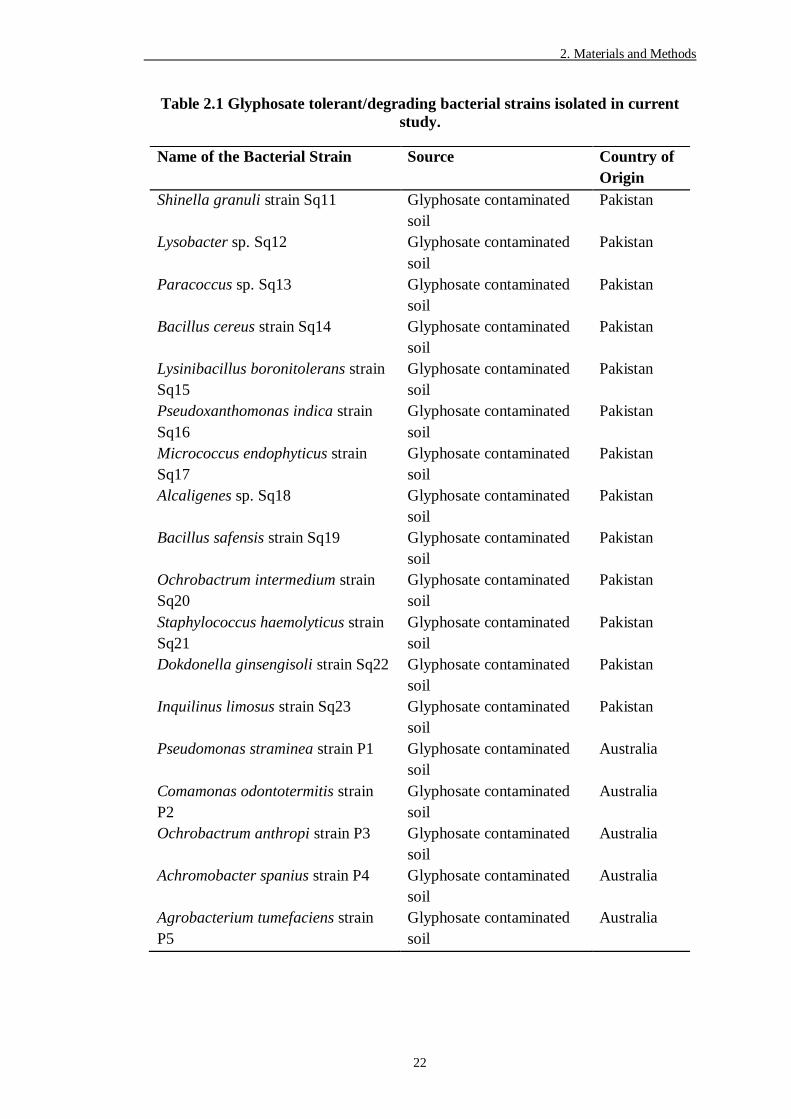
2. Materials and Methods
22
Table 2.1 Glyphosate tolerant/degrading bacterial strains isolated in current
study.
Name of the Bacterial Strain Source Country of
Origin
Shinella granuli strain Sq11 Glyphosate contaminated
soil
Pakistan
Lysobacter sp. Sq12 Glyphosate contaminated
soil
Pakistan
Paracoccus sp. Sq13 Glyphosate contaminated
soil
Pakistan
Bacillus cereus strain Sq14 Glyphosate contaminated
soil
Pakistan
Lysinibacillus boronitolerans strain
Sq15
Glyphosate contaminated
soil
Pakistan
Pseudoxanthomonas indica strain
Sq16
Glyphosate contaminated
soil
Pakistan
Micrococcus endophyticus strain
Sq17
Glyphosate contaminated
soil
Pakistan
Alcaligenes sp. Sq18 Glyphosate contaminated
soil
Pakistan
Bacillus safensis strain Sq19 Glyphosate contaminated
soil
Pakistan
Ochrobactrum intermedium strain
Sq20
Glyphosate contaminated
soil
Pakistan
Staphylococcus haemolyticus strain
Sq21
Glyphosate contaminated
soil
Pakistan
Dokdonella ginsengisoli strain Sq22 Glyphosate contaminated
soil
Pakistan
Inquilinus limosus strain Sq23 Glyphosate contaminated
soil
Pakistan
Pseudomonas straminea strain P1 Glyphosate contaminated
soil
Australia
Comamonas odontotermitis strain
P2
Glyphosate contaminated
soil
Australia
Ochrobactrum anthropi strain P3 Glyphosate contaminated
soil
Australia
Achromobacter spanius strain P4 Glyphosate contaminated
soil
Australia
Agrobacterium tumefaciens strain
P5
Glyphosate contaminated
soil
Australia

2. Materials and Methods
23
2.3 Soil Sample Collection
Soil samples were collected from fields of Ayub Agricultural Research Institute
(AARI) Faisalabad, Pakistan and Plant Breeding Institute, University of Sydney at
Cobbitty, NSW Australia. The collected soil samples had a long history of exposure
to widely used herbicide glyphosate consequently making this soil a crucial source for
isolation of glyphosate tolerant/degrading bacterial strains.
2.4 Growth Media
Growth media used in this study were Luria Bertani (LB), Minimal Salt Medium
(MSM), Enrichment medium (C and P free medium), Minimal medium (P free
medium). The compositions of all growth media are given in appendices.
2.5 Maintenance and Preservation of the Isolated Bacterial
Strains
Glyphosate tolerant bacterial isolates were preserved as glycerol stocks and agar
slants. Glycerol stocks (50% v/v) were prepared by adding bacterial cultures grown in
LB broth in eppendorfs containing 50% glycerol under aseptic conditions. The stocks
were kept at -80 °C for months. Agar slants or stab stocks were prepared from LB
agar, inoculated with bacterial strains, incubated for 16 h at ambient temperature and
preserved at 4 °C.
2.6 Equipments Used in Current Study
The majority of the facilities and equipments used in current study were available at
NIBGE. These included Spectrophotometer; High Performance Liquid
Chromatography (HPLC) Varian Pro Star 325, UV VIS detector; Eppendorf
centrifuge machines (models 5424 and 5816); Biorad Thermocycler;
Stereomicroscope; Rotary shaker; Light microscope. DIONEX High Performance
Liquid Chromatography (HPLC) and NanoDrop 2000c spectrophotometer was
availed at University of Sydney, Australia. DNA sequencing was carried out using
commercial sequencing facility of Macrogen, South Korea, until otherwise
mentioned.

2. Materials and Methods
24
2.7 Enrichment of Glyphosate Tolerant Bacterial Strains
Soil samples were collected from the fields of Ayub Agriculture Research Institute
Faisalabad having prior history of extensive exposure to glyphosate. Samples were
collected at 10-15 cm depth from three different areas of same field, mixed together,
transferred to sterile plastic bags and stored at 4 °C. The soil was air dried and sieved
using a 2 mm mesh prior to its use. Enrichment culture technique was employed for
isolation of glyphosate tolerant bacteria under the selective pressure of herbicide
[122]. Soil (5 g) was added in 250 mL Erlenmeyer flask containing a mixture of 50
mL of mineral salts medium (MSM) and 1000 mg/L glyphosate and the flasks were
incubated on a rotary shaker at 100 rpm for one week at 37 °C. Same procedure was
repeated by taking 1 mL of supernatant from enrichment culture and transferred to
fresh MSM containing 1000 mg/L glyphosate and incubated under same conditions.
This process was repeated for four weeks.
2.8 Isolation of Glyphosate Tolerant Bacterial Strains
The enrichment culture obtained after four weeks was further used for isolation of
glyphosate tolerant strains. Enrichment cultures were serially diluted (10-1
to 10-7
) in
saline solution (0.9%) and selected dilutions 10-5
, 10-6
and 10-7
of each culture were
used to spread (100 µL) in triplicates on solid LB agar plates containing 100 mg/L
glyphosate. The plates were incubated at 37 °C for 2 days and colonies with different
morphologies were isolated and sub-cultured on nutrient agar plates and bacterial
colonies with distinct morphology were observed.
2.9 Glyphosate Tolerance of Bacterial Isolates
The growth and glyphosate tolerance of bacterial isolates was monitored in liquid
MSM supplemented with 100 mg/L glyphosate as carbon and energy source. To
evaluate the growth of isolated strains, they were added individually in sterile MSM
containing 500 mg/L glyphosate with 2% inoculum in 250 mL Erlenmeyer flasks
along with respective negative controls. The flasks were placed in a shaker incubator
at 37 °C and 100 rpm, and after 72 hours, their absorption was measured by the
spectrophotometer at 590 nm wavelength. The growth of isolates was measured after
5 days of incubation to select the best isolates in the presence of glyphosate.
Turbidometric method was employed for monitoring the growth of inoculated

2. Materials and Methods
25
bacterial strains [123]. Cell dry mass of the bacterial cultures having an OD (optical
density at 600 nm) of 1.0 was calculated and was used as standard for calculating cell
dry mass of all the culture samples.
After five days of incubation, bacterial cultures were harvested and herbicide
residues were derivatized and analyzed by quantifying its residual concentrations on
HPLC. Preeminent tolerant bacterial strains were characterized and employed for
further degradation studies.
2.10 Morphological Characterization of the Bacterial
Isolates
2.10.1 Morphological Characterization
Morphological characterization of bacterial isolates was carried out by studying their
morphological characteristics (size, margins, shape and surface texture) and
microscopic features (pigmentation, motility and gram stain) as per the standard
procedures [124].
2.10.2 Gram Staining Method
The concentrations and compositions of all reagents used in Gram‟s method of
staining e.g., crystal violet solution (staining reagent), iodine solution (mordant) and
safranin solution (counter stain) are illustrated in Appendix F.
Following procedure was used for gram staining of bacterial isolates:
Bacterial strains were streaked on LB agar plates and incubated at 37 C overnight.
Single colony of each bacterial strain was selected in sterile conditions and mixed
with a drop of saline (0.9%) to make a thin smear on a glass slide.
The slide was air dried, fixed through heat and flood with primary crystal violet
staining reagent for 1 minute.
The smear was washed with gentle and indirect stream of distilled water for 2
seconds.
Then slide was flooded with iodine solution for 1 minute.
The slide was exposed to distilled water for 2 seconds and decolorizing agent (70%
ethanol) was applied for 30 seconds.

2. Materials and Methods
26
The glass slide was again washed with distilled water for 2 seconds and counter
stain, safranin staining reagent was applied for 1 minute.
Glass slide was washed with gentle and indirect stream of distilled water until
effluent seemed colourless and then dried with absorbent paper.
The stained bacterial isolates were observed under oil immersion using a Bright
field microscope.
2.11 Molecular Characterization of Isolated Bacterial
Strains
2.11.1 DNA Isolation
CTAB (Cetyl Tri-methyl Ammonium Bromide) Method reported by Mateen [125]
was used for DNA isolation of bacterial isolates and procedure is described as
follows:
Bacterial isolates were inoculated in MSM containing glucose at 37 C overnight
and harvested by centrifugation at 8000 rpm for 10 minutes.
Cell pellet was resuspended in 5 mL T.E buffer and lysozyme (20 mg) was added
to this solution.
The resulting suspension was incubated at 37 C for 5 minutes in water bath.
After that 500 µL of 10% SDS, 25 µL proteinase K (25 mg/mL) and 3 µL RNAase
were added to the suspension.
It was thoroughly mixed and again incubated at 37 C for 10 min in water bath.
Then 0.9 mL 5 M NaCl and 0.75 mL NaCl/CTAB were added thoroughly mixed
and incubated at 65 C for 20 min.
The protein contents of the bacterial suspension were extracted (twice) with an
equal volume of phenol-chloroform-isoamyl alcohol (25:24:1) and centrifuged at
8000 rpm and 4 °C for 10 min in order to separate the organic and aqueous phase
(containing DNA).
Aqueous phase was separated in fresh falcon tube and 0.6 volume of isopropanol
was added and thoroughly mixed until a white DNA thread appeared in the
solution. The solution containing DNA was stored at -20 °C overnight.
The solution containing DNA was centrifuged at 8000 rpm for 5 min next day and
supernatant was discarded.

2. Materials and Methods
27
DNA pellet was washed with 70% ethanol two times and dried at room
temperature.
Dried DNA pellet was resuspended in 50 µL T.E and saved at -20 °C until used.
2.11.2 Amplification of 16S rRNA Gene from Isolated Bacterial
Strains
16S rRNA gene analysis was carried out by amplifying this gene from all bacterial
isolates using forward primer FD1, 5-AGAGTTTGATCCTGGCTCAG-3 (E. coli
bases 8-27) and reverse primer RP1, 5-ACGGHTACC TTGTTACGACTT-3 (E. coli
bases 1507-1492) [126]. PCR reactions were performed in 50 µL reaction volumes
containing 25 µL of 2X Dreamtaq PCR master mix, 2 µL of each of the primers (10
µM), 2 µL of bacterial DNA and 19 µL of sterile distilled water. Thermal cycler
(Biorad) was used for amplification and thermocycling conditions consisted of a
denaturation step at 94 °C for 10 min, 30 amplification cycles at 94 °C for 30 s, 55 °C
for 25 s and 72 °C for 90 s and a final extension at 72 °C for 10 min. QIAQuick spin
column (QIAGEN) kit was used for purification of PCR product.
2.11.3 Agarose Gel Electrophoresis
The amplified PCR products were examined on 1% agarose gel stained with ethidium
bromide (100 μg/mL). PCR products were mixed with 6X loading dye (bromophenol
blue), loaded on the agraose gel and electrophoresed in 0.5X TAE buffer at 80-100
volts for about 30 to 45 minutes. Then gel was visualized under UV transilluminator
for analyzing the PCR products.
2.11.4 Ligation and Cloning of 16S rRNA Gene
PCR products of 16S rRNA gene were ligated into TA cloning vector obtained from
PCR Product cloning Kit of Fermentas) and the ligation mixtures were kept at 16 °C
for 16 h. The competent cells of E. coli were used for transformation of ligation
mixtures following heat shock method next day. Mini prep kit (Fermentas) was used
to isolate plasmids from E. coli and restricted with EcoRI (restriction enzyme,
Fermentas) to authenticate the insert size.
2.11.5 Sequencing of 16S rRNA Gene and Bacterial Identification
The cloned PCR products were sequenced with M13 primers and resulting sequences
were submitted to BLAST software (version 2.2.12) for homology studies. By using

2. Materials and Methods
28
Molecular Evolutionary Genetics Analysis (MEGA) software, version 6.0 was used
for multiple alignments and distance matrix analyses and consensus neighbor joining
tree construction.
2.12 Bacterial Diversity Assessment through PCR-DGGE
For analysis of bacterial diversity PCR-DGGE was carried out as described by
Cunliffe and Kertesz [127]. The primers used for DGGE PCR were GC-341F (with
GC clamp at 5')
(CGCCCGCCGCGCGCGGCGGGCGGGGCGGGGGCACGGGGGGCCTACGGG
AGGCAGCAG) and 518R (ATTACCGCGGCTGCTGG) [128]. The PCR profile
includes the following steps.
1. 95 °C for 5 min
2. 95 °C for 30 sec
3. 60 °C for 30 sec (touchdown -1 °C/cycle)
4. 68 °C for 30 sec (10 cycles from step 2)
5. 95 °C for 30 sec
6. 50 °C for 30 sec
7. 68 °C for 30 sec (20 cycles from step 5)
8. 68 °C for 5 min
2.12.1 DGGE Gel Casting and Running
After the preparation and mixing of 0% denaturant solution and 100% denaturant
solution, the gel was casted immediately as it starts to polymerize due to the presence
of APS.
Glass plates (large & small) along with spacers was washed with 70% ethanol,
fixed with sandwich clamps and consigned on the casting stand.
Gel solution of both concentrations was filled in two 50 mL syringes and they
were fixed to gradient maker. It was kept in mind that there should be no air
bubbles in the syringes and the tubes.
A syringe needle was connected with the syringe tubes and fixed at the middle of
the gel chamber. The hand of gradient maker was constantly rotated during gel
casting.

2. Materials and Methods
29
The comb was washed with 70% ethanol and placed it into the gel chamber while
avoiding air bubbles. The gel was left to solidify for two hours.
After two hours, the comb was removed from gel and wells were washed with
deionized water.
The core was placed in the DGGE tank filled with 6 L of 1X TAE buffer and
circulation was started to warm the buffer up to 60 °C for 30 minutes.
After attaining the acquired temperature, core was removed from tank and gel was
attached to it.
The core was placed in tank and gel was washed with 1X TAE buffer.
The samples were loaded in the gel and tank was filled with 1X TAE buffer up to
the mark. The gel was run at 63 V for 16 hours.
2.12.2 DGGE Gel Staining
The gel was removed from core and placed on plastic tray.
20-40 mL of the SYBR Gold staining solution (light sensitive) was poured onto
the gel until it covered the whole gel. The tray was placed in dark room or covered
with aluminium foil and left for 30 minutes in the dark.
The gel was rinsed with deionized water for several times after 30 minutes and
deionized water was added to the tray. Left it for 10 minutes in the dark and then
visualized under transilluminator.
2.12.3 Excision and Re-amplification of DGGE PCR Fragments
Selected DGGE bands were excised from the gel and soaked in MilliQ H2O for 24
hours at 4 °C. The supernatant was used for re-amplification with same DGGE
primers without GC clamp. The PCR products were purified and sequenced from
Australian Genome Research Facility Ltd. DGGE sequences obtained were compared
with DNA database using NCBI BLAST and submitted in Genbank.
2.13 Inoculum Preparation of Bacterial Strains
The glyphosate tolerant bacterial strains were inoculated in MSM medium containing
500 mg/L of glyphosate in Erlenmeyer flasks at 37 °C on a rotary shaker for two days.
Then the grown bacterial culture was centrifuged at 4600×g for 8 to 10 minutes and
used as inoculum. The cell pellet was washed with saline solution (0.9%) and

2. Materials and Methods
30
resuspended to acquire an optical density (OD at 600 nm) of 0.8. The dilution plate
count method was used to determine colony forming units (CFU/mL). The suspension
(2% v/v) obtained was further used as inoculum in biodegradation experiments until
otherwise depicted.
2.14 Glyphosate Degradation Studies
Glyphosate degradation studies were done using 250 mL Erlenmeyer flasks
containing 100 mL of MSM enriched with 500 mg/L glyphosate and 2% inoculum of
degrading strains under different culture conditions as described in respective
sections. The flasks were incubated in rotary shaker of 100 rpm at 37 °C (or at other
temperatures according to the respective experiment conditions) for one week. For all
the treatments, uninoculated flasks were used as control and all the experiments were
executed in triplicates. Samples from flasks were collected from time to time for
investigating the growth rates of bacterial strains and residues of glyphosate.
2.15 Detection of Glyphosate
2.15.1 Detection Wavelength of Glyphosate
Although the chromatographic conditions were followed as described by Qian et al.
[129] but for further corroboration, optimum wavelength (λmax) of glyphosate
derivative was determined spectrophotometrically. λmax was measured by recording
wavelength of glyphosate derivative at maximum UV absorption and also estimated
using HPLC and comparing the absorbance of the derivative at about three different
wavelengths.
2.15.2 Derivatization of Glyphosate
Detection of glyphosate is difficult due to its low molecular weight, low volatility,
high water solubility and lack of chromophores or fluorophores that could facilitate its
detection therefore glyphosate was derivatized with 4-chloro-3, 5-
dinitrobenzotrifluoride (CNBF) and quantified by HPLC.
The bacterial cultures harvested after specific intervals were centrifuged for 10
minutes at 12,000 rpm and supernatant was used for derivatization. The derivatization
procedure was performed as described by Qian et al. [129]. Supernatant (200 µL)
containing glyphosate was added to a vial containing a mixture of 300 µL of H3BO3-

2. Materials and Methods
31
Na2B4O7 buffer (mixture of 0.2 mol/L H3BO3 solution with 0.05 mol/L Na2B4O7,
pH: 9.5) and 100 µL of CNBF methanol solution (2.5 mmol/L). Then the entire
solution was diluted to 1 mL with ddH2O and incubated at 60 °C for 30 min. HCl (2
M, 10 µL) was added to knock down the reaction. The resulting solutions were
filtered through 0.45 µm nylon filters and injected in the chromatographic system.
Each sample was analyzed in triplicate and all the assays were consummated at
ambient temperature.
2.15.3 HPLC Conditions for Glyphosate Residual Analysis
For HPLC analysis, Kromasil ODS C18 column was used and pre-equilibrated with
the mobile phase for 30 min. The mobile phases used in the analysis were phosphate
buffer (50 mM, pH: 2.5 with phosphoric acid) (eluent A) and acetonitrile:water (1:1,
v/v, eluent B). All the solvents were filtered through a 0.45 µm membrane filter and
the program was set for a linear gradient starting from 20% of eluent B to reach its
100% at 30 min. The detector used was Photodiode array detector and the sample
injection volume was 20 µL at flow rate 1 mL/min. The detection wavelength was
360 nm. Retention time of glyphosate was calculated by running its standards
solutions [122]. Unit method was employed to measure the concentrations of
glyphosate in inoculated and non-inoculated control samples.
2.16 Study of Potential Genes in Glyphosate Tolerant
Bacterial Strains
2.16.1 Amplification of Glyphosate Resistant and Degrading Genes
Degenerate primers were designed and synthesized to amplify the potential genes
involved in glyphosate degradation/resistance from glyphosate tolerant bacterial
isolates. Primers were designed by retrieving already reported sequences of 5-
Enolpyruvylshikimate-3-phosphate synthase (EPSPS) gene, glyphosate
oxidoreductase gene (GOX), C-P lyase encoding gene (PhnJ) from NCBI GenBank.
The sequences were aligned through ClustalW software and conserved regions of the
sequences were utilized for primer designing. Furthermore degenerate bases were
added in case of mismatches between aligned gene sequences. The parameters of
primers were validated by using OligoDT software. The sequences of the designed
primers are given in Table 2.2.

2. Materials and Methods
32
PCR reactions were carried out in reaction volumes of 25 µL containing
Taq DNA polymerase (0.5 µL, 2.5 U/µL), 10X PCR reaction buffer (2.5 µL),
forward and reverse primers (0.75 µL each, 10 µM), bacterial genomic DNA (1
µL) and sterile PCR water (19.5 µL). The PCR protocol was: denaturation at 94 C
for 1 min, annealing at 52-65 C (on the basis of Tm of primer pairs) for 1 min,
extension at 72 C for 90 s, all steps were repeated for 35 cycles and a final
extension at 72 °C for 10 min.
2.16.2 Agarose Gel Electrophoresis, Cloning and Sequencing of the
Amplified Genes
The PCR products were analyzed on agarose gel (1%) on the basis of their size in
base pairs. Same protocol was followed as described in Section 2.11.3 for 16S rRNA
gene.
The purification of amplified PCR products was done by using PCR
purification kit (Fermentas), ligated to PTZ vector and transformed in E. coli DH5
through heat shock method. Plasmids were isolated from transformants by mini prep
kit (Fermentas). Isolated plasmids were digested with suitable restriction enzymes to
confirm the amplicon product size. The plasmids were sequenced and then analyzed
using the BLASTN search program of the GenBank database in National Center for
Biotechnology Information, NCBI.
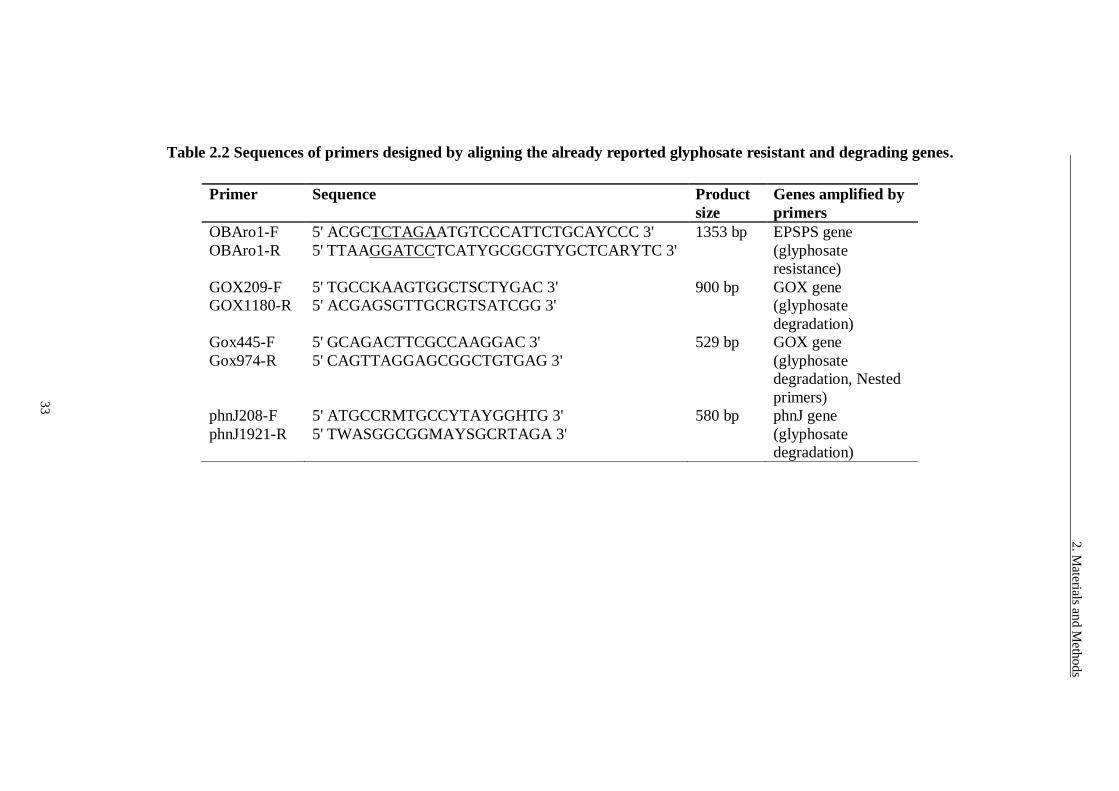
33
2. M
aterials and M
etho
ds
Table 2.2 Sequences of primers designed by aligning the already reported glyphosate resistant and degrading genes.
Primer Sequence Product
size
Genes amplified by
primers
OBAro1-F
OBAro1-R
5' ACGCTCTAGAATGTCCCATTCTGCAYCCC 3'
5' TTAAGGATCCTCATYGCGCGTYGCTCARYTC 3'
1353 bp EPSPS gene
(glyphosate
resistance)
GOX209-F
GOX1180-R
5' TGCCKAAGTGGCTSCTYGAC 3'
5' ACGAGSGTTGCRGTSATCGG 3'
900 bp GOX gene
(glyphosate
degradation)
Gox445-F
Gox974-R
5' GCAGACTTCGCCAAGGAC 3'
5' CAGTTAGGAGCGGCTGTGAG 3'
529 bp
GOX gene
(glyphosate
degradation, Nested
primers)
phnJ208-F
phnJ1921-R
5' ATGCCRMTGCCYTAYGGHTG 3'
5' TWASGGCGGMAYSGCRTAGA 3'
580 bp phnJ gene
(glyphosate
degradation)

34
3. Isolation and Characterization of
Glyphosate Resistant Bacterial Strains
3.1 Introduction
Glyphosate is a glycine derivative and belongs to the group of amino acid inhibitor
herbicides. It is distinguished from organophosphorus compounds by the presence of
a stable covalent carbon to phosphorus (C-P) bond that is resistant against chemical
hydrolysis, thermal decomposition and photolysis [121]. Since 1970, it has been
widely used as weedicide and crop dessicant in the form of various formulations [53,
130]. Glyphosate exhibits its herbicidal activity by halting the penultimate step of
aromatic amino acid synthesis pathway (shikimate pathway) [131]. Phenylalanine,
tyrosine and tryptophan synthesized through this pathway are further involved in
protein synthesis as well as secondary metabolite production (phenolics, lignins,
tannins, and other phenylpropanoids). This pathway is present in bacteria, algae,
fungi, apicomplexan parasites and plants but absent in animals which attributes to the
effective use of glyphosate as herbicide. Glyphosate acts in systemic manner as it
permeates through the surface of plant leaves to the whole plant by dint of phloem
and amasses in meristematic cells [132]. Inhibition of shikimate pathway leads to the
obstruction of physiological processes inside the plant resulting in chlorosis, growth
stunting and consequent plant death [87]. Shikimate pathway works in cytosol of
fungi and bacteria whereas in case of plants it is also found in plastid organelles.
Microorganisms use more than 90% of their metabolic energy for protein synthesis
whereas plants use more energy on secondary metabolite production. About 30%
carbon fixed by plants flows through shikimate pathway at normal growth conditions.
The shikimate pathway is indispensable for plants as well as for pathogenic
microorganisms as Streptomyces pneumonia strains [133], Bordetella bronchiseptica
[134], Mycobacteria [135] , Thermotoga gondii [136] and Plasmodium falciparum
[137] which makes it a potential antimicrobial target.

3. Glyphosate Resistant Bacterial Strains
35
3.1.1 Glyphosate Resistance
The introduction of glyphosate resistant transgenic soybean in 1996 led to the colossal
use of glyphosate. Since glyphosate can be used as post emergent herbicide in case of
glyphosate resistant crops therefore they have been promptly adopted. More than 90%
of total soybean, >75% of total cotton and >70% of total maize cultivated in USA was
glyphosate resistant in 2010 [138]. Powles claimed that glyphosate is one in a century
herbicide encompassing the same importance in food production as possessed by
penicillin for combating diseases [139]. The efficiency of glyphosate to remove vast
variety of weeds, low price and less toxicity contributed to its extensive use [56, 140].
Additionally, cessation of glyphosate patent in 2000 escorted different manufacturing
companies to immense production of glyphosate resulting in lower price and
substantial adoption.
The possible mechanisms involved in glyphosate resistance are: (a) Evolution
of resistance due to selection pressure from extreme use of glyphosate without
modifying mode of action of glyphosate, (b) target site resistance through substitution
of amino acids which influence glyphosate interactions with target enzyme, (c)
metabolism of glyphosate either by conjugation or degradation, (d) non target
resistance through changes at molecular or physiological level. Target site resistance
and glyphosate detoxification has been found thriving for introduction of glyphosate
resistance in crops. EPSPS acquired from A. tumefaciens CP4 or double mutated
EPSPS (TIPS) insensitive to glyphosate have been effectively used for commercial
production of glyphosate resistant crops. The speculative shortcoming of this strategy
is that glyphosate pile up in growing points of plants thus impeding reproductive
growth and lowering crop yield [141]. The metabolic detoxification of glyphosate
through plant enzymes or transgene encoded enzymes may help in elimination of
glyphosate residues and robust glyphosate resistance.
3.1.2 5-Enolpyruvylshikimate-3-Phosphate Synthase (EPSPS, EC
2.5.1.26)
The unearthing of EPSPS as glyphosate target enzyme impelled scientists to study its
structure, function and catalytic mechanisms in detail [131]. EPSPS catalyzes the
reaction between phosphoenolpyruvate (PEP) and shikimate-3-phosphate (S3P)
resulting in production of enolpyruvyl shikimate-3-phosphate (EPSP) and inorganic

3. Glyphosate Resistant Bacterial Strains
36
phosphate [142]. This reaction is sixth step of aromatic amino acid synthesis pathway,
shikimate pathway. Previously, only MurA bacterial enzyme (involved in bacterial
cell wall synthesis) has been reported to catalyze similar reaction. Glyphosate acts as
a competitive inhibitor of EPSPS binding adjoining to S3P in the PEP binding site
configuring a stable non-covalent ternary complex of EPSPS-S3P-glyphosate [143,
144]. The inhibition of EPSPS by glyphosate is uncompetitive with respect to S3P
[143] (Figure 3-1). Consequently, the shikimate pathway is hindered by either
reduced amounts of EPSPS or lacking of EPSPS functionality resulting in reduced
synthesis of aromatic amino acids and their derivatives [145].
Figure 3-1 Inhibition of penultimate step of shikimic acid pathway by
glyphosate.
Glyphosate hampers the conversion of S3P and PEP to EPSP resulting in scarcity of
aromatic amino acids [146].
First X-ray crystallography studies of EPSPS of E. coli demonstrated that it
has two globular domains each having three similar folding units tied to each other via
a two stranded hinge region [147]. Further studies showed that reaction of EPSPS
pursue induced fit mechanism, in which both globular domains move towards each
other in a screw like manner after S3P binding [58]. This changeover generates a
restrained and highly charged surroundings in the vicinity of hydroxyl group of S3P
where PEP or glyphosate bind. One more high resolution study revealed that
glyphosate and PEP share the identical binding site with similar binding interactions
[148]. Similar structural properties were also reported in EPSPS of S. pneumoniae
[149] and Agrobacterium sp. [150].

3. Glyphosate Resistant Bacterial Strains
37
3.1.3 Classification of EPSP Synthase
EPSPS has been categorized into four distinctive classes: Class I [151], Class II [152],
Class III [153] and Class IV [154] depending on its sensitivity towards glyphosate
concentrations. Naturally glyphosate sensitive Class I EPSP synthases are found in
some bacteria such as Escherichia coli and Salmonella enterica serovar
Typhimurium, Pseudomonas putida and all green plants [155, 156]. They have been
expansively characterized for enzyme active site and kinetics [58]. Catalytic activity
of such enzymes is interrupted at low (micromolar) glyphosate concentrations. Class I
EPSP synthases have a conserved amino acid motif (LFLGNAGTAMRPL) inside the
PEP binding pocket [157-159]. In contrast, catalytic activity of class II EPSP
synthases found in some bacteria including Pseudomonas sp. PG2982, Agrobacterium
tumefaciens CP4, Achromobacter sp. strain LBAA, Bacillus subtilis, Ochrobactrum
anthropi, Streptococcus pneumonia, Staphylococcus aureus is retained even at high
glyphosate concentrations [160]. They also have high affinity for the substrate PEP
[161] and hence are significantly helpful in developing glyphosate tolerant crops; e.g.,
soybeans, maize, cotton, etc. Glyphosate tolerant crops thus facilitate effective weed
control through post emergent herbicide application on such crops. Class II EPSPS do
not cross react with the polyclonal antibodies against Class I EPSPS and have <50%
amino acid similarity to class I enzymes [162]. Following five conserved amino acid
motifs GDKX, SAQXK, RXHXE, NXTR and RPMXR (X represents non conserved
amino acids) have been reported to contribute in glyphosate resistance in class II
EPSPS [163].
3.1.4 History and Engineering of Glyphosate Resistant EPSPS
High specificity of glyphosate towards EPSPS has exceedingly augmented its
application as herbicide. Even MurA instead of encompassing similarities with
EPSPS is not considerably hindered by glyphosate. Therefore it is not just an analog
of PEP but imitate intermediate state of PEP. Many glyphosate analogs have been
synthesized but no one showed significant inhibition of EPSPS due to changes in
chemical structure [53]. Scientists have strived for identification of EPSPS resistant to
glyphosate for development of glyphosate resistant crops since 1980s [152, 158].
Various glyphosate resistant EPSPS were identified through microbial screening, site
directed mutagenesis and selective evolution [149, 155, 158, 160, 164-167].
Glyphosate and PEP shares same binding site therefore improved glyphosate

3. Glyphosate Resistant Bacterial Strains
38
resistance most commonly results in reduced affinity of EPSPS towards PEP leading
to significant fitness cost.
Single Site Mutation
The first single mutations, P101S and G96A, conferring resistance to glyphosate were
reported in EPSPS from S. typhimurium [168] and K. Pneumonia [169] respectively.
The EPSPS from E. coli harbouring single mutation of G96A has been found highly
resistant to glyphosate owing to the overhanging of methyl group in binding site. But
this mutation has severely reduced affinity of EPSPS for PEP and its catalytic activity
[166]. In contradiction to G96A mutation, P101S is positioned away (about 9 Å) from
glyphosate binding site and its substitution results in slight changes in active site.
Therefore it exhibits low glyphosate resistance while sustaining high catalytic activity
and low fitness cost as compared to active site mutations. Naturally glyphosate
resistant plants have been found to contain single mutation consistently at site
analogous to Pro101 of EPSPS from E. coli [159, 170-173].
Multi Site Mutation
More than one mutation accompanying favourable characteristics were reported in
EPSPS from P. hybrid (G101A/G137D and G101A/P158S) [174], E. coli
(G96A/A183T) [175, 176] and Z. mays (T102I/P106S) [175, 177, 178]. The double
mutant (T102I/P106S analogous to T97I/P101S in E.coli), designated as TIPS EPSPS,
with favourable characteristics has been used for production of glyphosate resistant
maize (commercial field corn). The only glyphosate resistant class I EPSPS reported
so far is TIPS EPSPS from E. coli and has high affinity for PEP. X-ray
crystallography analysis revealed that double mutation in TIPS EPSPS moves G96
towards glyphosate whereas I97 reallocates away from binding site consequently
aiding in PEP utilization [157]. Agrobacterium sp. strain CP4 has been reported to
yield an efficient glyphosate resistant EPSPS designated as CP4 EPSPS. It has also
been utilized for production of glyphosate resistant crops (Roundup Ready, NK603)
[152]. The CP4 EPSPS belongs to the class II EPSPS and its catalytic activity
depends on monovalent cations (K+
and NH+4
). High resistance against glyphosate is
ascribed to the presence of A100 and L105 residues instead of G96 and P101 residues
found in plants and E. coli. Although binding of PEP is not affected by A100 but
glyphosate prefer to bind in high energy and non inhibitory conformation. By

3. Glyphosate Resistant Bacterial Strains
39
replacing A100 with glycine, the sensitivity of glyphosate can be restored partially
[138].
3.1.5 Glyphosate Detoxification
Another approach used for introduction of glyphosate resistance is detoxification of
glyphosate compound. Two different pathways are employed by soil microbes for
metabolism of glyphosate. First pathway is C-P lyase pathway which involves
cleavage of the C-P bond present in glyphosate leading to the production of sarcosine
and inorganic phosphate. This pathway is catalyzed by C-P lyase. Second pathway is
AMPA pathway which involves the oxidative cleavage of C-N bond leading to the
production of aminomethylphosphonic acid (AMPA) and glyoxylate. This pathway is
catalyzed by glyphosate oxidoreductase (GOX). None of these pathways has been
found to transpire in higher plants to a considerable level. C-P lyase pathway needs
phn operon which consists of 14 genes rendering its regulation difficult in transgenic
plants. AMPA pathway has been found most predominant among soil microbes for
glyphosate degradation. Another enzyme, Glycine oxidase (GO) from B. subtilis has
been reported to utilize glyphosate yielding AMPA and glyoxylate [138].
The current study was premeditated to isolate and characterize glyphosate
tolerant strains from indigenous soil and screen for the presence of EPSPS encoding
gene(s). A glyphosate resistant strain Ochrobactrum intermedium Sq20 was isolated
from the soil obtained from agricultural fields where glyphosate had been frequently
applied. Moreover, in this strain, aroAO.intermedium Sq20 gene was sequenced and
EPSPSO.intermedium Sq20 was characterized thereafter by means of bioinformatics tools.
3.2 Materials and Methods
3.2.1 Enrichment, Isolation and Screening of Glyphosate Tolerant
Bacterial Strains
Soil was collected from the fields with a previous history of extensive glyphosate
exposure. Enrichment culture technique was employed for isolation of glyphosate
tolerant bacterial strains (chapter 2, section 2.7, 2.8) and 13 isolates were obtained.
The growth of isolated bacterial strains at high glyphosate concentrations (up to 2
g/L) in LB was monitored by using a turbidometric method [123]. Glyphosate
tolerance by these isolates was also checked on MSM agar plates supplemented with

3. Glyphosate Resistant Bacterial Strains
40
2 g/L glyphosate as the sole carbon and energy source. Simultaneously, residual
glyphosate in MSM cultures containing glyphosate (500 mg/L) as sole
carbon/phosphorus source was measured to find the potential of the isolated bacteria
for glyphosate degradation. A glyphosate tolerant bacterial strain Sq20 was thereafter
selected for further studies.
3.2.2 Morphological and Taxonomic Analysis of Glyphosate
Tolerant Bacterial Isolates
The standard morphological and taxonomic methods were used for characterization of
glyphosate tolerant bacterial strains (chapter 2 Section 2.10).
3.2.3 Amplification and Sequencing of aroAO.intermedium Sq20 Gene
Degenerate primers OBAro1For 5′-ACGCTCTAGAATGTCCCATTCTGCAYCCC-
3′ (Y=C/T) and OBAro1Rev 5′-TTAAGGATCCTCATYGCGCGTYGCTCARYTC-
3′ (R=A/G) were designed through alignment of already known aroA gene sequences
of Ochrobactrum spp (GenBank accession No. GU992200, CP000758,
ACQA01000001). Using these primers, full length aroAO.intermedium Sq20 gene was
amplified whereby PCR conditions were, denaturation at 94 °C for 10 min, 30
amplification cycles at 94 °C for 1 min, 59 °C for 1 min and 72 °C for 90 s and a final
extension at 72 °C for 10 min. The PCR product (1.353 Kb) was purified through
PCR purification kit (Fermentas) and cloned in a pTZ57R/T vector under T7 promoter
by using specific restriction sites (XbaI and BamHI). Then plasmids were isolated
from clones using mini prep kit (Fermentas), and digested with XbaI and BamHI
restriction enzymes to confirm the insert size. The plasmids were sent to commercial
sequencing facility of Macrogen, South Korea for sequencing. The sequences
obtained were analyzed through BLASTN program of the GenBank database in NCBI
and submitted in Genbank.
3.2.4 Tolerance/ Degradation of Glyphosate by Sq20
Biodegradation studies of glyphosate were performed by incubating MSM augmented
with Sq20 inoculum (2%), glucose (1%) and 500 mg/L glyphosate for 5 days along
with negative and positive controls. Samples were collected at 8 h intervals for
analyzing the growth and biodegradation through BioTek Microplate Reader and
HPLC respectively. The bacterial cultures were centrifuged for 10 min at 12,000 rpm
and the supernatant was used for residual glyphosate analysis.

3. Glyphosate Resistant Bacterial Strains
41
Kinetics of glyphosate degradation was determined by plotting ln[Ct/C0]
against time (h). A linear trend showed that rate of this degradation process is
following first order kinetics model. Degradation rate constant (k, 1/h) and half-life
(t1/2, h) were determined using Equations (3-1) and (3-2) respectively [179].
Ct = C0 × e-kt
(3-1)
t1/2 = ln(2)/k (3-2)
Where Ct and C0 are the concentrations of glyphosate (mg/L) at time t and at
time zero respectively, k is first order rate constant and t is the degradation period (h).
3.2.5 HPLC Analysis of Glyphosate
Detection of glyphosate is difficult due to lack of chromophores or fluorophores in
this molecule that could facilitate its detection therefore glyphosate was derivatized
with 4-chloro-3,5-dinitrobenzotrifluoride (CNBF) [129] and quantified by HPLC. A
Kromasil ODS C18 column was used and pre-equilibrated with the mobile phase i.e.,
phosphate buffer (50 mM, pH 2.5) (eluent A) and acetonitrile: water (1:1, v/v, eluent
B). A modified method of Qian et al. [129] was employed for more sensitivity by
adjusting a linear gradient starting from 20% of eluent B to reach its 100% at 30 min.
Photodiode array UV/VIS detector was used for detection and the sample injection
volume was 20 µL at 1 mL/min flow rate and detection wavelength at 360 nm. After a
35 minute complete run, three peaks of glyphosate-CNBF complex, CNBF and
CNBF-OH at 15.05, 24.7 and 25.4 minutes were detected, respectively.
3.2.6 Detection of Metabolites Produced During Glyphosate
Degradation
The metabolites (AMPA, sarcosine, and glycine) produced during glyphosate
degradation were analyzed using Silufol UV254 plates by thin-layer chromatography.
O. intermedium Sq20 was incubated with 500 mg/L glyphosate for 24 h and liquid
culture was used for detection of glyphosate degradation products. The plates were
developed with ninhydrin reagent (0.5%) in acetone [180].
3.2.7 Bioinformatic Analysis of aroAO.intermedium Sq20
Bioinformatic analysis of the aroAO.intermedium Sq20 gene sequence was performed by
using different tools. Nucleotide and deduced amino acid sequences of aroAO.intermedium

3. Glyphosate Resistant Bacterial Strains
42
Sq20 gene were employed for sequence similarity searches using the Basic Local
Alignment Search Tool (BLAST) program. The multiple sequence alignment of
aroAO.intermedium Sq20 gene and EPSPS protein sequences with those of other species was
carried out by Clustal W with default parameters and phylogenetic trees that were
built using MEGA version 6 [181] by neighbor joining method.
Physicochemical Characterization
Physicochemical characterization of EPSPSO.intermedium Sq20 was carried out through
ProtParam server of Expasy [182]. Different physical and chemical parameters such
as theoretical isoelectric point (pI), molecular weight, total number of positive and
negative residues, extinction coefficient, instability index, aliphatic index and grand
average hydropathy (GRAVY) of EPSPSO.intermedium Sq20 were enumerated.
Secondary Structure Prediction
Secondary structure and disorder prediction of EPSPSO.intermedium Sq20 was carried out
by using Phyre2 (Protein Homology/AnalogY Recognition Engine). It modelled about
98% of the protein (440 residues) with 100% confidence and 98% query coverage by
the single highest scoring template [183]. Secondary structural features of
EPSPSO.intermedium Sq20 protein sequence were calculated with the help of Self
Optimized Prediction Method with Alignment (SOPMA) software [184] by applying
default parameters (window width: 17, similarity threshold: 8 and number of states:
4).
Homology Modeling and Quality Assessment
Three dimensional structure of EPSPSO.intermedium Sq20 was determined by using two
homology modeling softwares, Geno3D [185] and SWISS-MODEL [186]. The
modeled 3D structures of EPSPSO.intermedium Sq20 obtained from both software tools
were authenticated by PROCHECK [187] software by generating Ramachandran plot
of amino acid sequences in most favoured and disallowed regions [188]. The ProSA-
web server was further used to calculate energy profile and to validate the protein
structure in terms of Z score demonstrating overall quality of model and computing
the total energy deviation [189].

3. Glyphosate Resistant Bacterial Strains
43
3.3 Results
3.3.1 Isolation and Identification of Glyphosate Tolerant Bacterial
Strains
Through repeated sub-culturing in MSM containing 500 mg/L glyphosate as sole
carbon and energy source, 13 bacterial strains were isolated from glyphosate
contaminated soil. To choose the best one, growth of isolates was evaluated in MSM
containing 500 mg/L glyphosate with and without added glucose after 72 hours
incubation. The bacterial isolates Sq13, Sq17, Sq20 and Sq21 showed significant
growth in the presence of glucose whereas Sq20 isolate proclaimed the highest growth
rate in the presence of glucose (Figure 3-2a, 3-2b). Therefore Sq20 was found
proficient at glyphosate utilization and consequently used in further studies.
3.3.2 Morphological and Taxonomic Characterization of Glyphosate
Tolerant Bacterial Strains
Morphological analysis revealed that all isolated bacterial strains have distinct cell
morphologies and pigmentation. Microbiological analysis showed that Sq13, Sq17,
Sq21, Sq22 and Sq23 were non-motile whereas other 8 bacterial strains were motile.
Moreover, gram staining results demonstrated high incidence of gram negative
bacteria among isolations and only Sq14, Sq15, Sq17, Sq19 and Sq21 bacterial strains
were found gram positive (Table 3.1). Bacterial strains can be identified and classified
into different taxonomic groups on the basis of their cultural characteristics. The 16S
rRNA gene analysis revealed the taxonomy of isolated bacterial strains and each
isolate exhibited maximum identity with different genera. The closest sequence
homologs of bacterial isolates, their codes, accession numbers, maximum identity and
accession numbers of hits are illustrated in Table 3.2. The 16S rRNA gene sequence
analysis of Sq20 demonstrated that Sq20 has 99% identity with Ochrobactrum
intermedium type strain CNS-275 and was clustered in a well-supported branch with
various Ochrobactrum spp. type strains (Figure 3-3). Phylogenetic analysis of
bacterial isolates Sq11, Sq14 and Sq16 carried out through Neighbor joining method
showed their maximum identity with Shinella sp., Bacillus sp. and
Pseudoxanthomonas sp. respectively. The phylogenetic trees of these bacterial strains
are represented in Figures 3-4, 3-5 and 3-6.
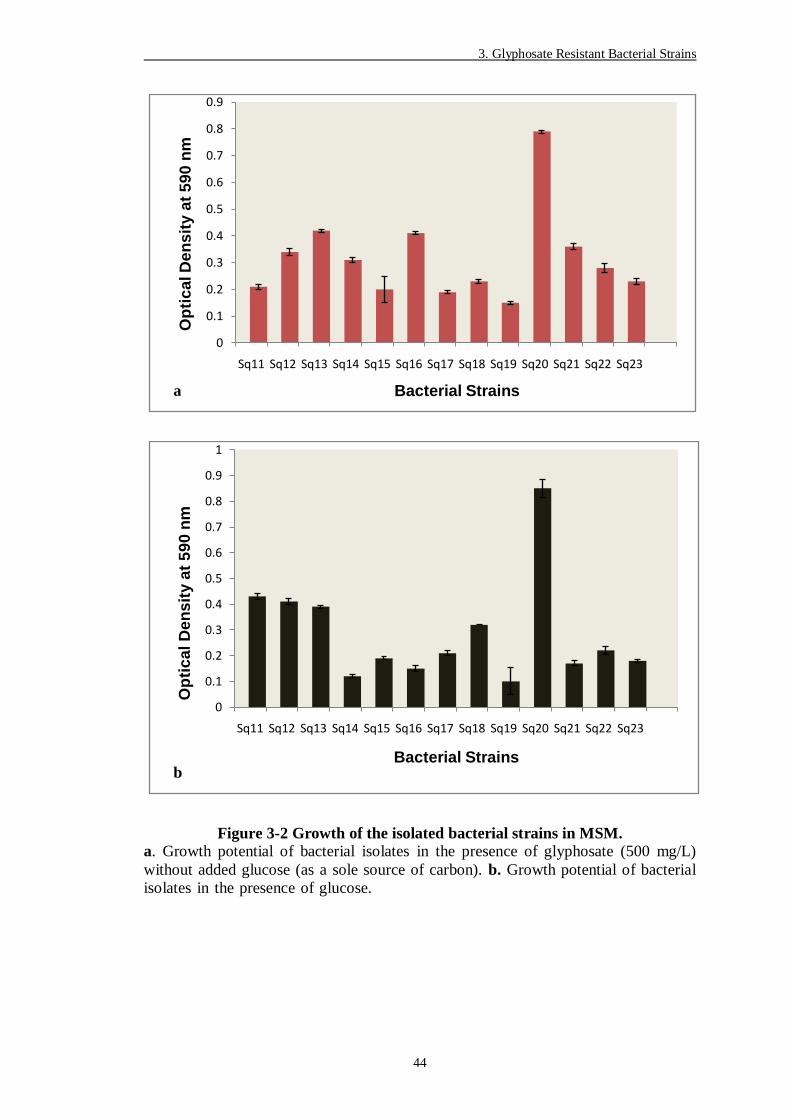
3. Glyphosate Resistant Bacterial Strains
44
Figure 3-2 Growth of the isolated bacterial strains in MSM.
a. Growth potential of bacterial isolates in the presence of glyphosate (500 mg/L)
without added glucose (as a sole source of carbon). b. Growth potential of bacterial
isolates in the presence of glucose.jjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjj
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
Sq11 Sq12 Sq13 Sq14 Sq15 Sq16 Sq17 Sq18 Sq19 Sq20 Sq21 Sq22 Sq23
Bacterial Strains
Op
tic
al D
en
sit
y a
t 5
90
nm
0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0.8
0.9
1
Sq11 Sq12 Sq13 Sq14 Sq15 Sq16 Sq17 Sq18 Sq19 Sq20 Sq21 Sq22 Sq23
Bacterial Strains
Op
tic
al D
en
sit
y a
t 5
90
nm
a
b
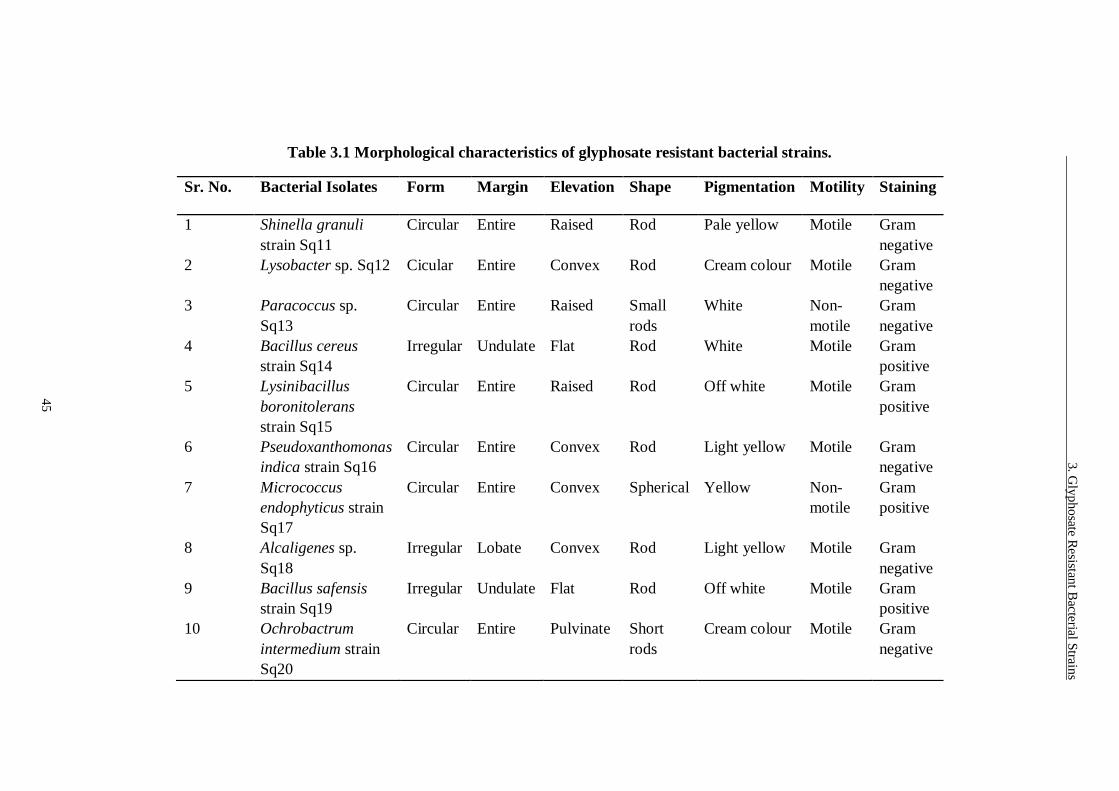
45
3. G
lyp
hosate R
esistant B
acterial Strain
s
Table 3.1 Morphological characteristics of glyphosate resistant bacterial strains.
Sr. No. Bacterial Isolates Form Margin Elevation Shape Pigmentation Motility Staining
1 Shinella granuli
strain Sq11
Circular Entire Raised Rod Pale yellow Motile Gram
negative
2 Lysobacter sp. Sq12 Cicular Entire Convex Rod Cream colour Motile Gram
negative
3 Paracoccus sp.
Sq13
Circular Entire Raised Small
rods
White Non-
motile
Gram
negative
4 Bacillus cereus
strain Sq14
Irregular Undulate Flat Rod White Motile Gram
positive
5 Lysinibacillus
boronitolerans
strain Sq15
Circular Entire Raised Rod Off white Motile Gram
positive
6 Pseudoxanthomonas
indica strain Sq16
Circular Entire Convex Rod Light yellow Motile Gram
negative
7 Micrococcus
endophyticus strain
Sq17
Circular Entire Convex Spherical Yellow Non-
motile
Gram
positive
8 Alcaligenes sp.
Sq18
Irregular Lobate Convex Rod Light yellow Motile Gram
negative
9 Bacillus safensis
strain Sq19
Irregular Undulate Flat Rod Off white Motile Gram
positive
10 Ochrobactrum
intermedium strain
Sq20
Circular Entire Pulvinate Short
rods
Cream colour Motile Gram
negative
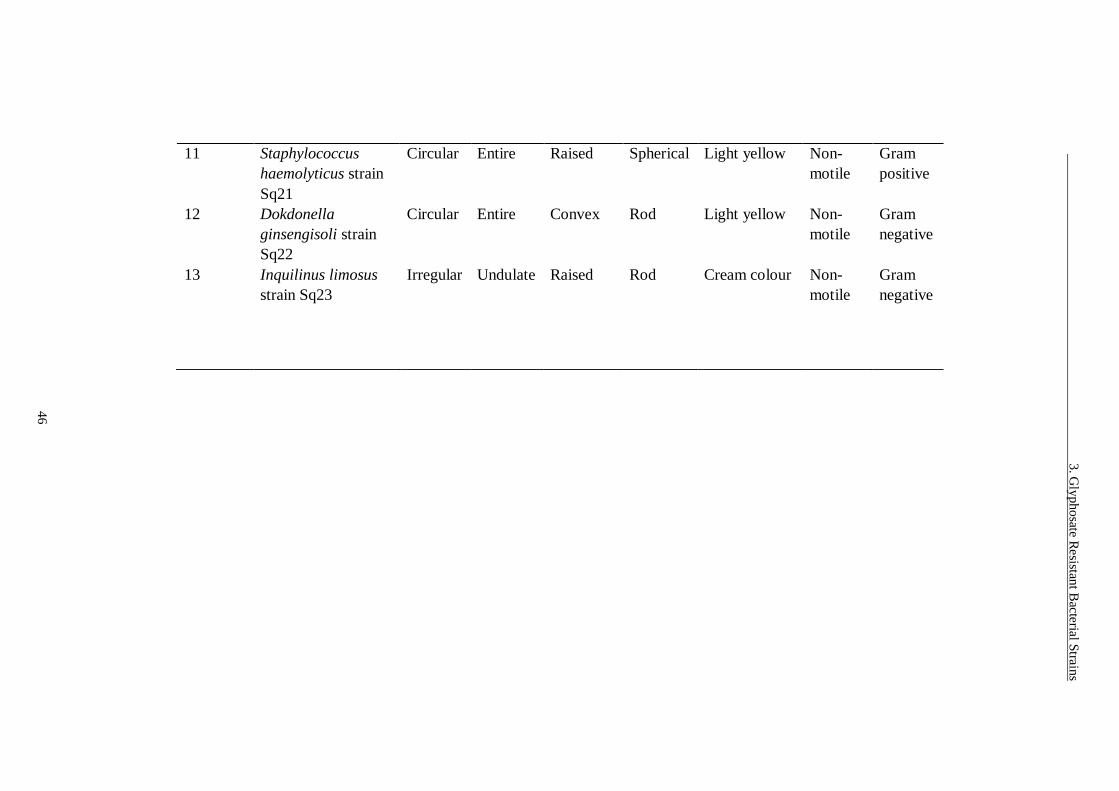
46
3. G
lypho
sate Resistan
t Bacterial S
trains
11 Staphylococcus
haemolyticus strain
Sq21
Circular Entire Raised Spherical Light yellow Non-
motile
Gram
positive
12 Dokdonella
ginsengisoli strain
Sq22
Circular Entire Convex Rod Light yellow Non-
motile
Gram
negative
13 Inquilinus limosus
strain Sq23
Irregular Undulate Raised Rod Cream colour Non-
motile
Gram
negative

3. Glyphosate Resistant Bacterial Strains
47
3
Table 3.2 Characterization of the glyphosate resistant bacterial strains isolated
in current study from glyphosate contaminated soil.
Sr.
No.
Isolates
Codes
Closest Sequence
Homologs of Bacterial
Isolates
Accession
No.
Accession
No. of Hits
Identity
(%)
1 Sq11 Shinella granuli KX255009 NR_041239.1 99%
2 Sq12 Lysobacter sp. KX255010 JN585697.1 99%
3 Sq13 Paracoccus sp. KC195786 KM603073.1 99%
4 Sq14 Bacillus cereus KX255011 HQ143564.1 99%
5 Sq15 Lysinibacillus
boronitolerans
KX255012 NR_114207.1 98%
6 Sq16 Pseudoxanthomonas indica KX255013 JQ659945 99%
7 Sq17 Micrococcus endophyticus KX255014 NR_044365.1 99%
8 Sq18 Alcaligenes sp. KX255015 KP876586.1 99%
9 Sq19 Bacillus safensis KX255016 NR_113945.1 99%
10 Sq20 Ochrobactrum
intermedium
KC195785 NR_113812.1 99%
11 Sq21 Staphylococcus
haemolyticus
KX255017 NR_113345.1 99%
12 Sq22 Dokdonella ginsengisoli KX255018 KF378756.1 99%
13 Sq23 Inquilinus limosus KX255019 NR_029046.1 99%
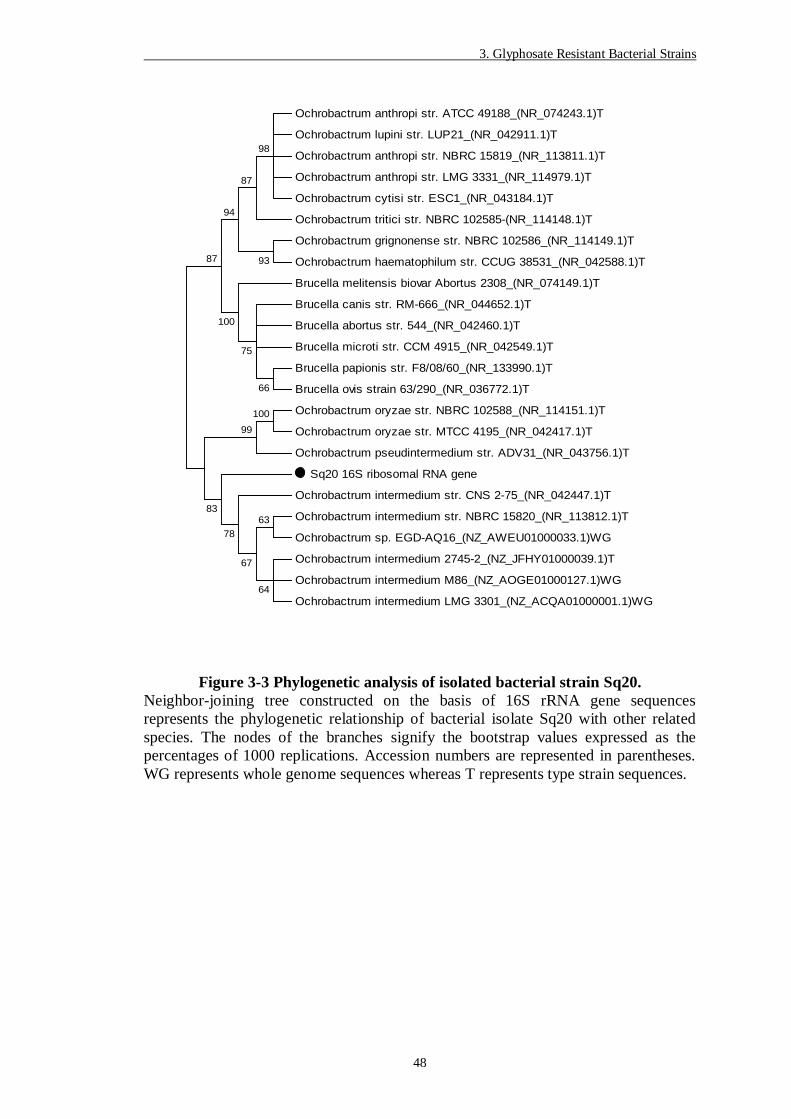
3. Glyphosate Resistant Bacterial Strains
48
Figure 3-3 Phylogenetic analysis of isolated bacterial strain Sq20.
Neighbor-joining tree constructed on the basis of 16S rRNA gene sequences
represents the phylogenetic relationship of bacterial isolate Sq20 with other related
species. The nodes of the branches signify the bootstrap values expressed as the
percentages of 1000 replications. Accession numbers are represented in parentheses.
WG represents whole genome sequences whereas T represents type strain sequences.
Ochrobactrum anthropi str. ATCC 49188_(NR_074243.1)T
Ochrobactrum lupini str. LUP21_(NR_042911.1)T
Ochrobactrum anthropi str. NBRC 15819_(NR_113811.1)T
Ochrobactrum anthropi str. LMG 3331_(NR_114979.1)T
Ochrobactrum cytisi str. ESC1_(NR_043184.1)T
Ochrobactrum tritici str. NBRC 102585-(NR_114148.1)T
Ochrobactrum grignonense str. NBRC 102586_(NR_114149.1)T
Ochrobactrum haematophilum str. CCUG 38531_(NR_042588.1)T
Brucella melitensis biovar Abortus 2308_(NR_074149.1)T
Brucella canis str. RM-666_(NR_044652.1)T
Brucella abortus str. 544_(NR_042460.1)T
Brucella microti str. CCM 4915_(NR_042549.1)T
Brucella papionis str. F8/08/60_(NR_133990.1)T
Brucella ovis strain 63/290_(NR_036772.1)T
Ochrobactrum oryzae str. NBRC 102588_(NR_114151.1)T
Ochrobactrum oryzae str. MTCC 4195_(NR_042417.1)T
Ochrobactrum pseudintermedium str. ADV31_(NR_043756.1)T
Sq20 16S ribosomal RNA gene
Ochrobactrum intermedium str. CNS 2-75_(NR_042447.1)T
Ochrobactrum intermedium str. NBRC 15820_(NR_113812.1)T
Ochrobactrum sp. EGD-AQ16_(NZ_AWEU01000033.1)WG
Ochrobactrum intermedium 2745-2_(NZ_JFHY01000039.1)T
Ochrobactrum intermedium M86_(NZ_AOGE01000127.1)WG
Ochrobactrum intermedium LMG 3301_(NZ_ACQA01000001.1)WG
66
75
100
100
99
98
64
63
67
78
83
87
87
94
93
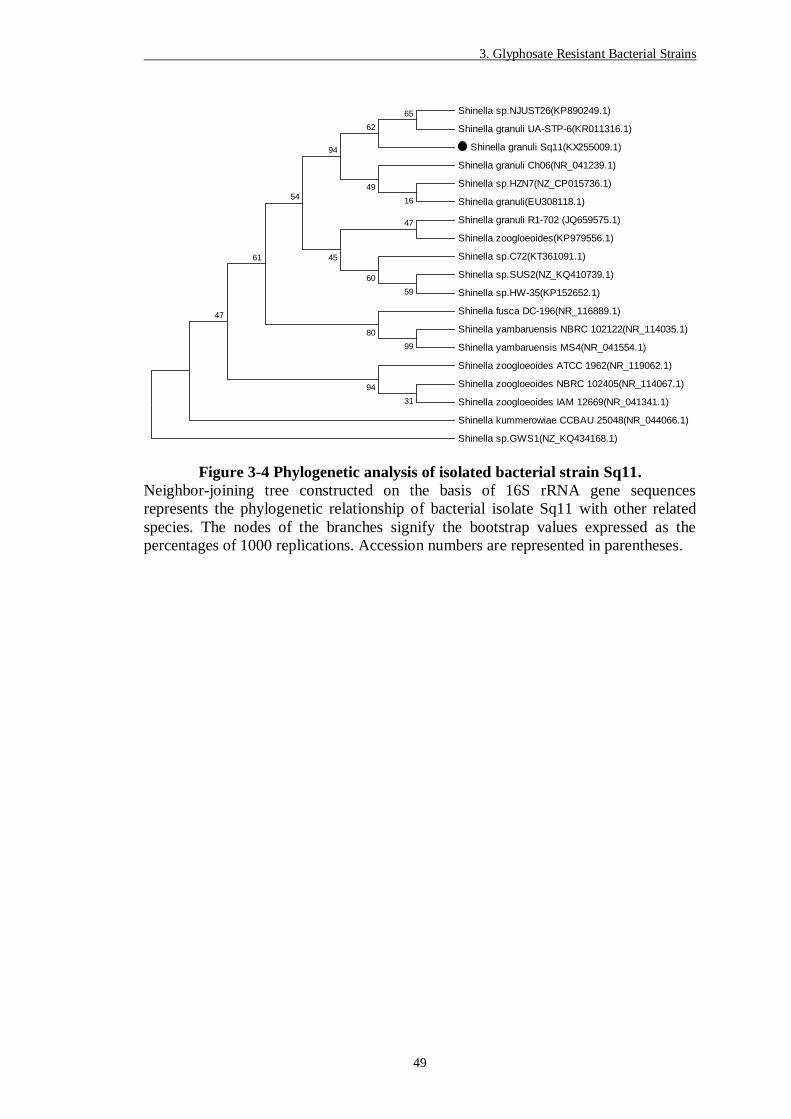
3. Glyphosate Resistant Bacterial Strains
49
Figure 3-4 Phylogenetic analysis of isolated bacterial strain Sq11.
Neighbor-joining tree constructed on the basis of 16S rRNA gene sequences
represents the phylogenetic relationship of bacterial isolate Sq11 with other related
species. The nodes of the branches signify the bootstrap values expressed as the
percentages of 1000 replications. Accession numbers are represented in parentheses.
Shinella sp.NJUST26(KP890249.1)
Shinella granuli UA-STP-6(KR011316.1)
Shinella granuli Sq11(KX255009.1)
Shinella granuli Ch06(NR_041239.1)
Shinella sp.HZN7(NZ_CP015736.1)
Shinella granuli(EU308118.1)
Shinella granuli R1-702 (JQ659575.1)
Shinella zoogloeoides(KP979556.1)
Shinella sp.C72(KT361091.1)
Shinella sp.SUS2(NZ_KQ410739.1)
Shinella sp.HW-35(KP152652.1)
Shinella fusca DC-196(NR_116889.1)
Shinella yambaruensis NBRC 102122(NR_114035.1)
Shinella yambaruensis MS4(NR_041554.1)
Shinella zoogloeoides ATCC 1962(NR_119062.1)
Shinella zoogloeoides NBRC 102405(NR_114067.1)
Shinella zoogloeoides IAM 12669(NR_041341.1)
Shinella kummerowiae CCBAU 25048(NR_044066.1)
Shinella sp.GWS1(NZ_KQ434168.1)
99
80
65
62
59
60
47
45
31
94
47
61
54
94
16
49

3. Glyphosate Resistant Bacterial Strains
50
Figure 3-5 Phylogenetic analysis of isolated bacterial strain Sq14.
Neighbor-joining tree constructed on the basis of 16S rRNA gene sequences
represents the phylogenetic relationship of bacterial isolate Sq14 with other related
species. The nodes of the branches signify the bootstrap values expressed as the
percentages of 1000 replications. Accession numbers are represented in parentheses.
Bacillus cereus Sq14(KX255011.1)
Bacillus cereus 03BB108(CP009641.1)
Bacillus cereus DZ4(HQ143564.1)
Bacillus cereus BFE 5392(GU250443.1)
Bacillus cereus 165PP(KM349191.1)
Bacillus thuringiensis HD1011(CP009335.1)
Bacillus sp.A52(KF479557.1)
Bacillus anthracis Vollum 1B(CP009328.1)
Bacillus cereus 03BB102(CP009318.1)
Bacillus cereus 9_julio(KM349186.1)
Bacillus cereus XX2010(JX993816.1)
Bacillus cereus L12(KU551157.1)
Bacillus sp.190Cu-As(KM349198.1)
Bacillus cereus ATCC 14579(NR_114582.1)
Bacillus cereus IAM 12605(NR_115526.1)
Bacillus cereus ATCC 14579(NR_074540.1)
Bacillus cereus CCM 2010(NR_115714.1)
Bacillus cereus JCM 2152(NR_113266.1)
Bacillus cereus NBRC 15305(NR_112630.1)
Bacillus pseudomycoides NBRC 101232(NR_113991.1)
Bacillus thuringiensis ATCC 10792(NR_114581.1)
Bacillus thuringiensis NBRC 101235(NR_112780.1)
Bacillus mycoides 273(NR_036880.1)
Bacillus cereus S2-8(CP009605.1)
Bacillus thuringiensis HD682(CP009720.1)100
84
59
53

3. Glyphosate Resistant Bacterial Strains
51
Figure 3-6 Phylogenetic analysis of isolated bacterial strain Sq16.
Neighbor-joining tree constructed on the basis of 16S rRNA gene sequences
represents the phylogenetic relationship of bacterial isolate Sq16 with other related
species. The nodes of the branches signify the bootstrap values expressed as the
percentages of 1000 replications. Accession numbers are represented in parentheses.
3.3.3 Glyphosate Resistance and Degradation by Ochrobactrum
Intermedium Sq20
To explore glyphosate resistance potential of strain Sq20, it was inoculated into LB
media containing 0.5-3.0 g/L glyphosate and growth was monitored until 24 h of
incubation. In this media, up to 2.5 g/L initial concentration of glyphosate, growth of
Ochrobactrum intermedium Sq20 was comparable to that of the control without
glyphosate indicating that the strain Sq20 is resistant to high glyphosate
concentration. Moreover, Ochrobactrum intermedium Sq20 was able to degrade
glyphosate at 500 mg/L initial concentration in MSM within three days at ambient
temperature and neutral pH. After 88h of incubation, 80% of the added glyphosate
was degraded and an increase in cell mass was observed (Figure 3-7a). Complete
degradation of glyphosate was attained after 104 h of incubation.
Experimental data of glyphosate degradation was interpreted statistically for
the calculation of first order reaction parameters and regression equation (Figure 3-
7b). Results showed rate constant (k) of 0.0464/h and half life (t1/2) of 14.9 h.
Regression coefficient (R2) value of 0.998 indicated a good fit of this model.
Pseudoxanthomonas indica Sq16(KX255013.1)
Pseudoxanthomonas indica C18(LN827737.1)
Pseudoxanthomonas sp.TMX-24(JN867356.1)
Pseudoxanthomonas sp.RD_MAAMIA_15(KU597505.1)
Pseudoxanthomonas indica S9-676(JQ660316.1)
Pseudoxanthomonas mexicana RA58(JN585700.1)
Pseudoxanthomonas indica R8-542(JQ659945.1)
Pseudoxanthomonas indica R4-389-1(JQ659701.1)
Pseudoxanthomonas mexicana L2(GQ480839.1)
Pseudoxanthomonas mexicana(EU119264.1)
Pseudoxanthomonas mexicana(AB246798.1)
Pseudoxanthomonas indica P15(NR_116019.1)
Pseudoxanthomonas indica R5-337(JQ659735.1)
Pseudoxanthomonas wuyuanensis XC21-2(NR_126229.1)
Pseudoxanthomonas jiangsuensis(NR_132712.1)
Pseudoxanthomonas japonensis NBRC 101033(NR_113972.1)
Pseudoxanthomonas japonensis 12-3(NR_024660.1)
Pseudoxanthomonas mexicana NBRC 101034(NR_113973.1)
Pseudoxanthomonas mexicana AMX 26B(NR_025105.1)
98
95
98
81
100
74
70
63
94
60

3. Glyphosate Resistant Bacterial Strains
52
3.3.4 Detection of Glyphosate Degradation Products
To study the route of glyphosate degradation followed by O. intermedium
Sq20, thin layer chromatography was used. The liquid culture of O. intermedium
Sq20 incubated with glyphosate was used for detection of major metabolites (AMPA,
glycine, and sarcosine). AMPA was not detected in the test samples confirming the
absence of glyphosate oxidoreductase (GOX) degradation pathway involving
breakdown of C-N bond. However, sarcosine (Rf = 0.53) and glycine (Rf = 0.65) were
detected by comparing their Rf values with those obtained for the standard substances,
affirming the breakdown of glyphosate through C-P lyase pathway. The control
samples containing O. intermedium Sq20 incubated with KH2PO4 showed no
sarcosine or glycine. These results indicate that O. intermedium Sq20 has C-P lyase
activity and cleaves the C-P bond of glyphosate resulting in the formation of sarcosine
which is further oxidized to glycine.
3.3.5 Bioinformatic Analysis of aroAO.intermedium Sq20
Sequencing of aroAO.intermedium insert indicated a 1353 bp open reading frame (ORF)
(Genbank Accession No. KU873038) with 63.8% GC contents encoding a peptide of
450 amino acids and was named aroAO.intermedium Sq20. The NCBI blast results and
phylogenetic analysis of aroAO.intermedium Sq20 revealed high homology with aroA genes
from other Ochrobactrum spp. Furthermore, BlastP and phylogenetic analysis of
translated EPSPSO.intermedium Sq20 sequence (Genbank Accession No. ANN25048)
revealed that this protein has 97-98% amino acid sequence identities to EPSPS
proteins from other Ochrobactrum spp. (Figure 3-8).
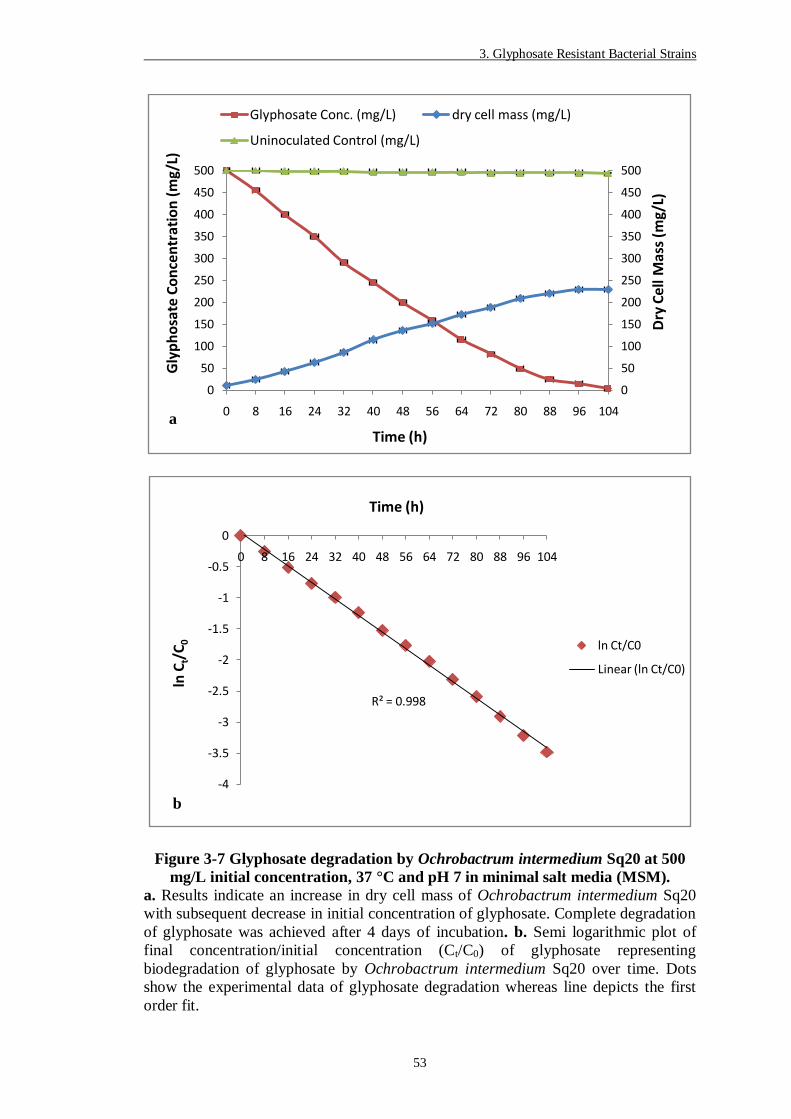
3. Glyphosate Resistant Bacterial Strains
53
Figure 3-7 Glyphosate degradation by Ochrobactrum intermedium Sq20 at 500
mg/L initial concentration, 37 °C and pH 7 in minimal salt media (MSM).
a. Results indicate an increase in dry cell mass of Ochrobactrum intermedium Sq20
with subsequent decrease in initial concentration of glyphosate. Complete degradation
of glyphosate was achieved after 4 days of incubation. b. Semi logarithmic plot of
final concentration/initial concentration (Ct/C0) of glyphosate representing
biodegradation of glyphosate by Ochrobactrum intermedium Sq20 over time. Dots
show the experimental data of glyphosate degradation whereas line depicts the first
order fit.
0
50
100
150
200
250
300
350
400
450
500
0
50
100
150
200
250
300
350
400
450
500
0 8 16 24 32 40 48 56 64 72 80 88 96 104
Glyphosate Conc. (mg/L) dry cell mass (mg/L)
Uninoculated Control (mg/L)
Time (h)
Dry
Cel
l Mas
s (m
g/L)
Gly
ph
osa
teC
on
cen
trat
ion
(m
g/L)
R² = 0.998
-4
-3.5
-3
-2.5
-2
-1.5
-1
-0.5
0
0 8 16 24 32 40 48 56 64 72 80 88 96 104
ln Ct/C0
Linear (ln Ct/C0)
Time (h)
lnC
t/C
0
a
b

3. Glyphosate Resistant Bacterial Strains
54
Figure 3-8 Phylogenetic analysis of EPSPS O.intermedium Sq20.
Neighbor-joining tree represents the phylogenetic relationship between
EPSPSO.intermedium Sq20 and EPSPS proteins of other related species. The percentage
values of the tree greater than 50 calculated from 1000 bootstrap resamples supporting
the topology are shown in phylogram. Accession numbers are indicated in
parentheses. The scale bar represents 0.01 substitutions per position.
It is reported that XLGNAGTAXRXL (X represents a variable amino acid
residue) is a highly conserved region of EPSPS(s) involved in the binding of substrate
PEP with EPSP synthases [190]. Clustal alignment showed that this region present in
the EPSPSO.intermedium Sq20 shows more sequence identity with that of the class II EPSPS
proteins than with class I analogs (Figure 3-9). Five other conserved regions GDKX,
SAQXK, RXHXE, NXTR and RPMXR reported in the class II EPSP synthases
including EPSPSA.tumefaciens CP4 and EPSPSO.anthropi were also found in EPSPSO.intermedium
Ochrobactrum sp. EGD-AQ16(WP_021587681.1)
Ochrobactrum intermedium(WP_006465745.1)
Ochrobactrum intermedium 229E(ERM02521.1)
Ochrobactrum intermedium(WP_025090682.1)
Ochrobactrum intermedium Sq20 EPSPS
Ochrobactrum anthropi(WP_043062739.1)
Ochrobactrum sp.(WP_029926097.1)
Ochrobactrum anthropi(WP_011982268.1)
Ochrobactrum anthropi(ADG03677.1)
Ochrobactrum anthropi(WP_010657837.1)
Brucella sp.BO2(WP_009364909.1)
Brucella inopinata(WP_008503805.1)
Brucella ceti str.Cudo(EEH15273.1)
Brucella microti(WP_012783809.1)
Brucella suis(WP_006206089.1)
Brucella sp.NVSL 07-0026(WP_008933634.1)
Brucella abortus NCTC 8038(EEW80290.1)
Brucella abortus str.2308(AEEP63548.1)
Brucella abortus 78/32(ENR88457.1)
Brucella abortus(WP_011265281.1)
Ochrobactrum rhizosphaerae(WP_024898290.1)
Ochrobactrum sp.CDB2(WP_007872980.1)
Pseudochrobactrum sp.(WP_022710242.1)
Phyllobacterium sp.(WP_027230834.1)
Phyllobacterium sp.YR531(WP_008127565.1)
Shinella sp.DD12(EYR84130.1)
Ensifer sp. USDA 6670(WP_029961663.1)
65
92
63
100
98
96
75
90
74
79
93
62
93
100
0.000.010.020.030.040.05

3. Glyphosate Resistant Bacterial Strains
55
Sq20. These regions have been revealed to impart glyphosate tolerance in class II EPSP
synthases. Moreover, phylogenetic analysis of EPSPSO.intermedium Sq20 protein with
already reported class I and II EPSPS(s) demonstrated that it belongs to class II
EPSPS (Figure 3-10).
Motif 1
Motif 2
Motif 3 Motif 4
Motif 5
Motif 6
Figure 3-9 Multiple sequence alignment of EPSPSO.intermedium Sq20.
With a typical class I enzyme EPSPSE.coli and typical class II enzymes
EPSPSA.tumefaciens CP4 and EPSPSO.anthropi, done by using ClustalW. The motif
important for interaction with PEP conserved in class I EPSPS enzymes is
boxed as motif 1 and the other five motifs involved in destabilizing glyphosate
binding in class II EPSPSs are boxed as motif 2, motif 3, motif 4, motif 5 and
motif 6.

3. Glyphosate Resistant Bacterial Strains
56
Figure 3-10 Phylogenetic analysis of Ochrobactrum intermedium strain Sq20 with
Class I and class II enzymes.
Phylogram bootstrapped with 1000 replicates constructed from Clustal W and
MEGA5 representing relationship of Sq20 EPSPS protein with reported class I and II
EPSPSs. Class I species are represented by (▲) and class II species are symbolized
by (■). Tree suggests that Ochrobactrum intermedium strain Sq20 (●) belongs to class
II EPSPS proteins.
Physicochemical Characterization
The amino acid composition and other physicochemical parameters of
EPSPSO.intermedium Sq20 calculated by ProtParam software are represented in Table 3.3
and Table 3.4 respectively. The amino acid content suggests that this protein contains
high concentration of polar (hydrophilic) amino acid residues (54.5%) as compared to
the nonpolar (hydrophobic) amino acid residues (45.5%) and hence can be described
as hydrophilic. Low extinction coefficient exhibited by EPSPSO.intermedium Sq20 indicates
low concentrations of these Tyr, Trp and Cys amino acids residues in the protein.
Generally, a protein with an instability index smaller than 40 is considered as stable.
On this account, EPSPSO.intermedium Sq20 is stable since its instability index is 32.06.
High aliphatic index further suggested that this protein is thermally stable. The Grand
Pseudomonas sp.(P0A2Y4.1)
Achromobacter sp.(P0A2Y5.1)
Ochrobactrum intermedium strain Sq20
Ochrobactrum anthropi(ADG03677.1)
A. tumefaciens strain CP4(Q9R4E4.2)
Bradyrhizobium diazoefficiens(NP_767378.1)
Dichelobacter nodosus(Q46550.1)
Staphylococcus aureus(Q05615.2)
Halothermothrix orenii(WP_012635978.1)
Pseudomonas fluorescens(ABM21481.1)
E.coli(P0A6D3.1)
Pantoea ananatis(WP_014593322.1)
Serratia odorifera(WP_004964814.1)
Zea mays(CAA44974.1)
Oryza sativa(AAL07437.1)
Arabidopsis thaliana(P05466.3)
Nicotiana tabacum(P23981.1)
87
100
100
100
93
60
100
59
100
100
100
100
0.00.10.20.30.40.50.6
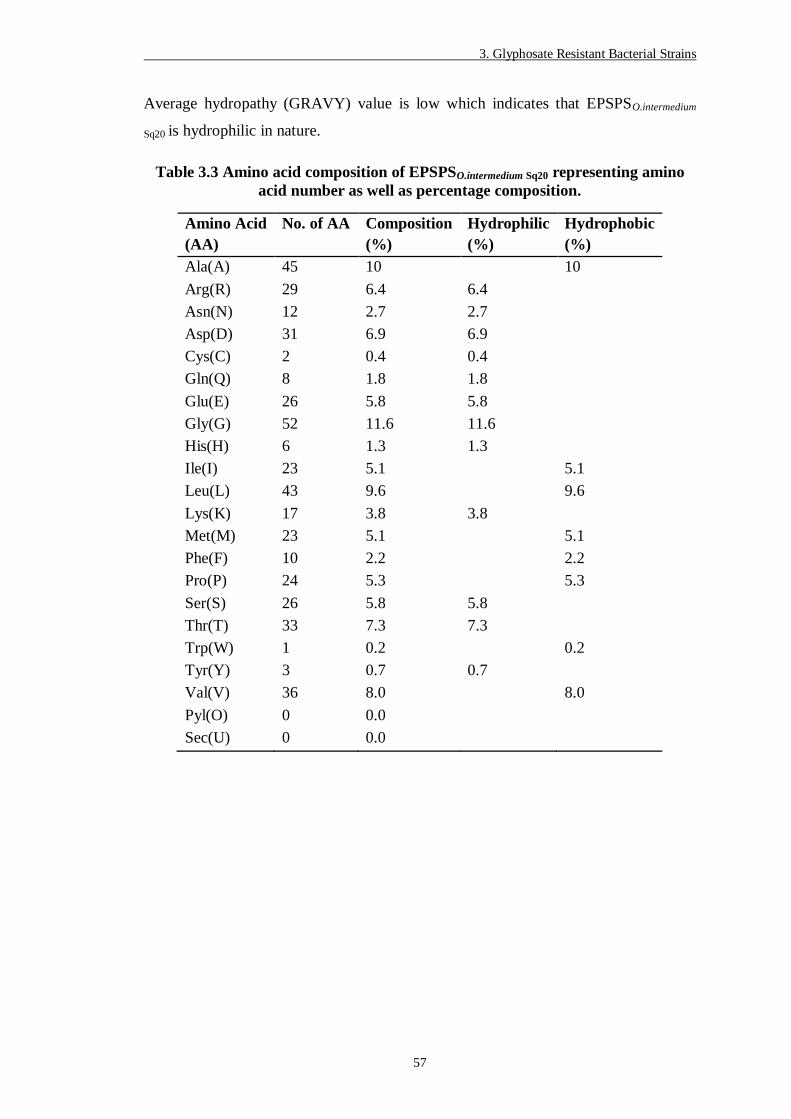
3. Glyphosate Resistant Bacterial Strains
57
Average hydropathy (GRAVY) value is low which indicates that EPSPSO.intermedium
Sq20 is hydrophilic in nature.
Table 3.3 Amino acid composition of EPSPSO.intermedium Sq20 representing amino
acid number as well as percentage composition.
Amino Acid
(AA)
No. of AA Composition
(%)
Hydrophilic
(%)
Hydrophobic
(%)
Ala(A) 45 10 10
Arg(R) 29 6.4 6.4
Asn(N) 12 2.7 2.7
Asp(D) 31 6.9 6.9
Cys(C) 2 0.4 0.4
Gln(Q) 8 1.8 1.8
Glu(E) 26 5.8 5.8
Gly(G) 52 11.6 11.6
His(H) 6 1.3 1.3
Ile(I) 23 5.1 5.1
Leu(L) 43 9.6 9.6
Lys(K) 17 3.8 3.8
Met(M) 23 5.1 5.1
Phe(F) 10 2.2 2.2
Pro(P) 24 5.3 5.3
Ser(S) 26 5.8 5.8
Thr(T) 33 7.3 7.3
Trp(W) 1 0.2 0.2
Tyr(Y) 3 0.7 0.7
Val(V) 36 8.0 8.0
Pyl(O) 0 0.0
Sec(U) 0 0.0

3. Glyphosate Resistant Bacterial Strains
58
Table 3.4 Physico-chemical characteristics of EPSPSO.intermedium Sq20 computed by
Expasy’s Protparam software.
No. Parameters Results
1 No. of amino acids 450
2 Molecular weight 47.3733 kDa
3 Isoelectrical point 5.21
4 Total number of negatively charged residues
(Asp + Glu)
57
5 Total number of positively charged residues
(Arg + Lys)
46
7 Chemical Formula C2048H3373N587O647S25
8 Extinction coefficient assuming all pairs of Cys
residues form cystines
10095 M-1
cm-1
9 Extinction coefficient assuming all Cys residues
are reduced
9970 M-1
cm-1
10 Instability index 32.06
11 Aliphatic index 90.4
12 Grand average of hydropathicity (GRAVY) -0.039

3. Glyphosate Resistant Bacterial Strains
59
Secondary Structure Prediction
The secondary structure of EPSPSO.intermedium Sq20 exhibits higher percentage of random
coils (33.56%) and alpha helices (30.44%) as compared to other secondary structure
elements (extended strands 24.44%, beta turns 11.56%). The positions of secondary
structure elements are presented in Figure 3-11. Disorder value (18%) showed that
EPSPSO.intermedium Sq20 has low incidence of disordered regions (Figure 3-12).
Figure 3-11 Secondary structure of EPSPSO.intermedium Sq20.
Secondary structure of EPSPSO.intermedium Sq20 protein indicating the positions of α
helices (h), extended strands (e), β turns (t) and random coils (c) computed by
SOPMA software. EPSPSO.intermedium Sq20 shows higher percentage of random coils as
compared to α helices, β turns and extended strands.
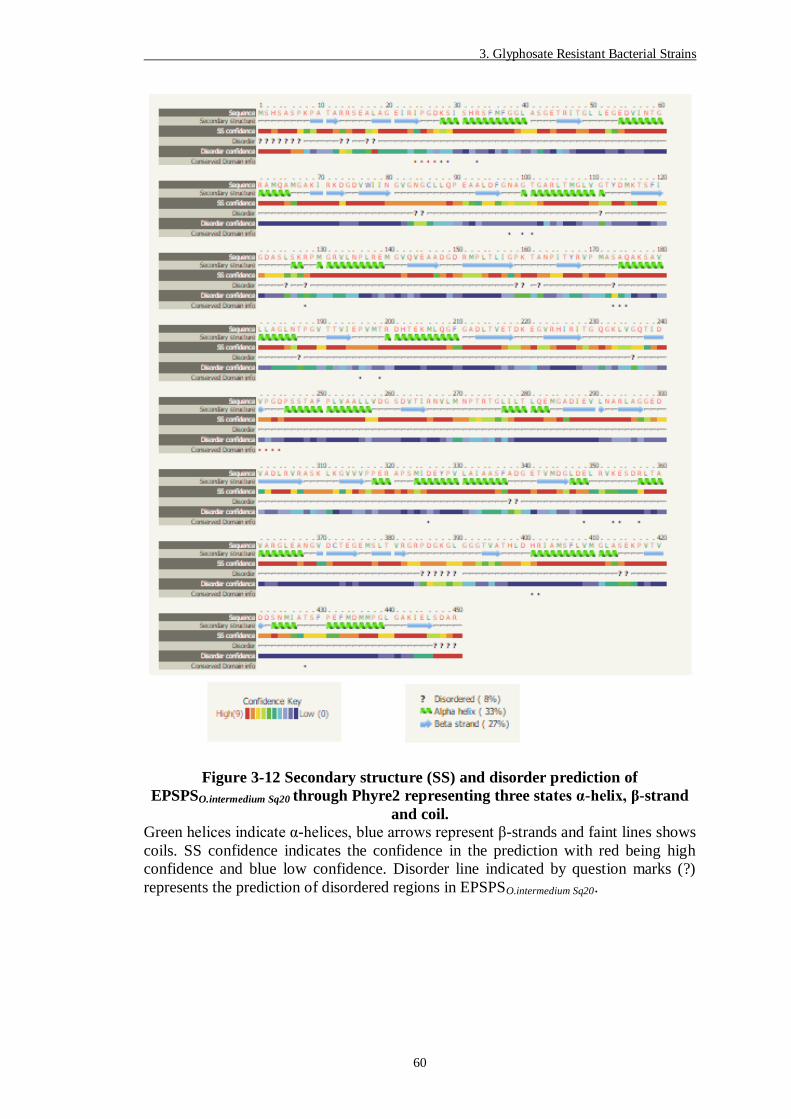
3. Glyphosate Resistant Bacterial Strains
60
Figure 3-12 Secondary structure (SS) and disorder prediction of
EPSPSO.intermedium Sq20 through Phyre2 representing three states α-helix, β-strand
and coil.
Green helices indicate α-helices, blue arrows represent β-strands and faint lines shows
coils. SS confidence indicates the confidence in the prediction with red being high
confidence and blue low confidence. Disorder line indicated by question marks (?)
represents the prediction of disordered regions in EPSPSO.intermedium Sq20.
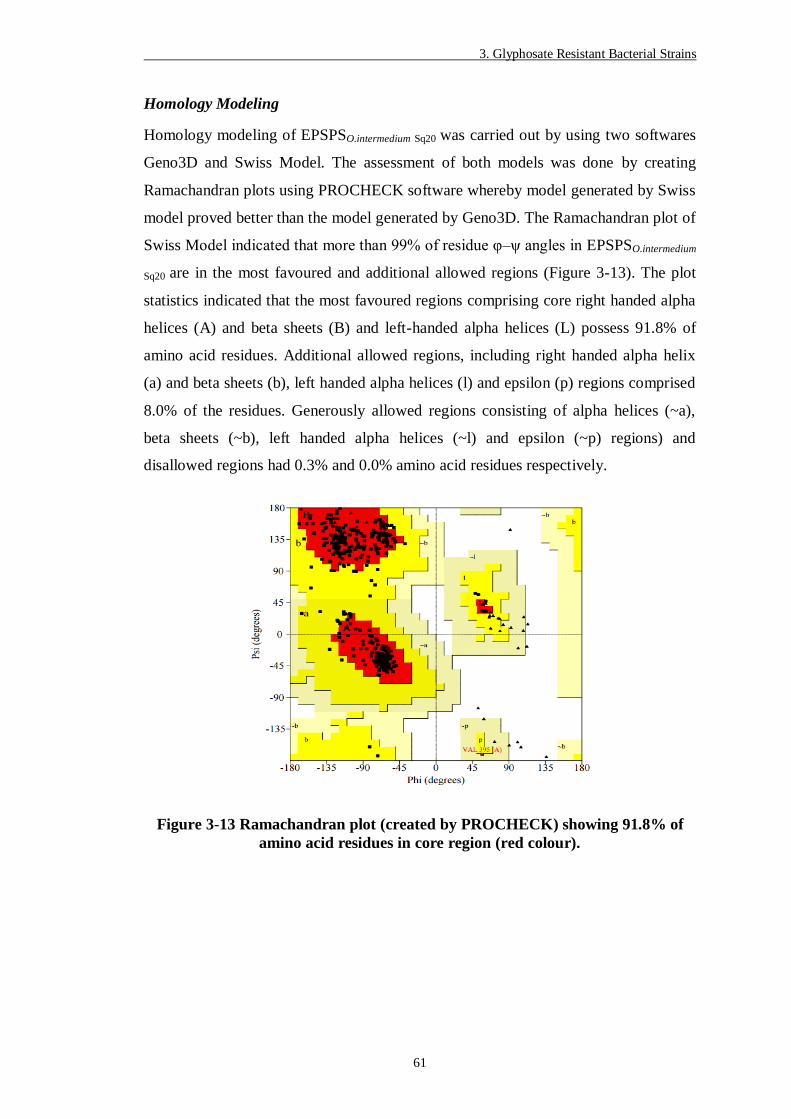
3. Glyphosate Resistant Bacterial Strains
61
Homology Modeling
Homology modeling of EPSPSO.intermedium Sq20 was carried out by using two softwares
Geno3D and Swiss Model. The assessment of both models was done by creating
Ramachandran plots using PROCHECK software whereby model generated by Swiss
model proved better than the model generated by Geno3D. The Ramachandran plot of
Swiss Model indicated that more than 99% of residue φ–ψ angles in EPSPSO.intermedium
Sq20 are in the most favoured and additional allowed regions (Figure 3-13). The plot
statistics indicated that the most favoured regions comprising core right handed alpha
helices (A) and beta sheets (B) and left-handed alpha helices (L) possess 91.8% of
amino acid residues. Additional allowed regions, including right handed alpha helix
(a) and beta sheets (b), left handed alpha helices (l) and epsilon (p) regions comprised
8.0% of the residues. Generously allowed regions consisting of alpha helices (~a),
beta sheets (~b), left handed alpha helices (~l) and epsilon (~p) regions) and
disallowed regions had 0.3% and 0.0% amino acid residues respectively.
Figure 3-13 Ramachandran plot (created by PROCHECK) showing 91.8% of
amino acid residues in core region (red colour).
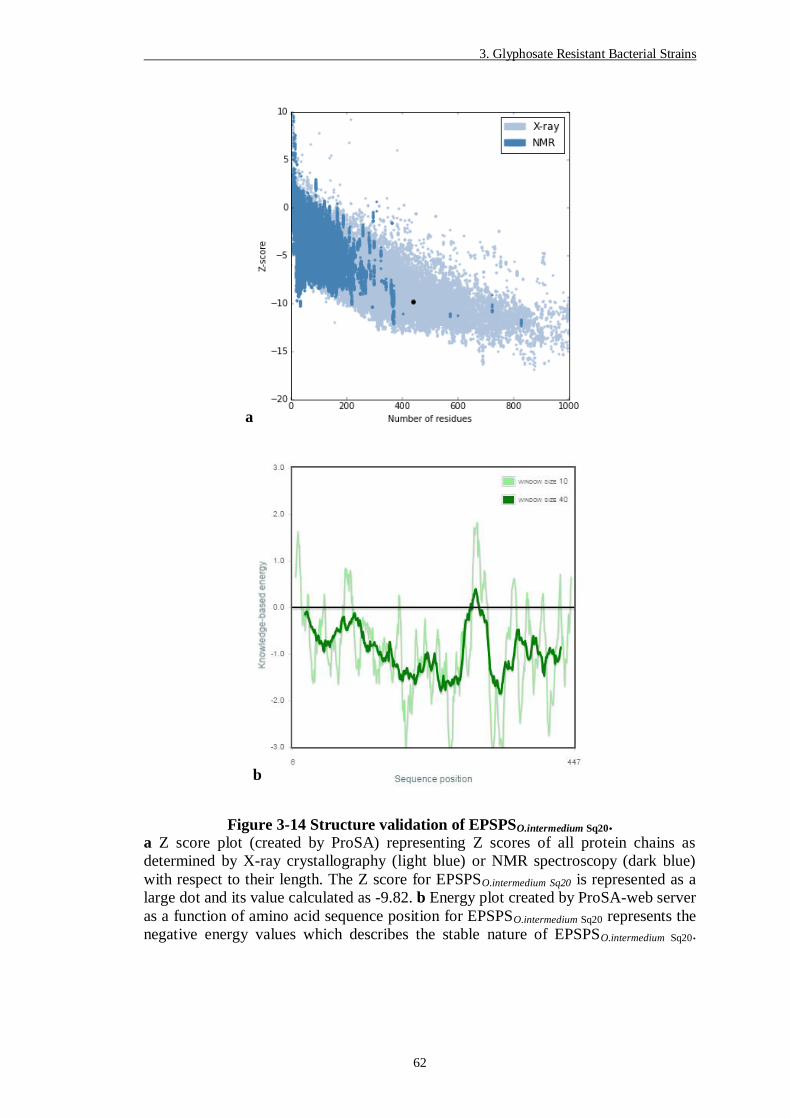
3. Glyphosate Resistant Bacterial Strains
62
Figure 3-14 Structure validation of EPSPSO.intermedium Sq20.
a Z score plot (created by ProSA) representing Z scores of all protein chains as
determined by X-ray crystallography (light blue) or NMR spectroscopy (dark blue)
with respect to their length. The Z score for EPSPSO.intermedium Sq20 is represented as a
large dot and its value calculated as -9.82. b Energy plot created by ProSA-web server
as a function of amino acid sequence position for EPSPSO.intermedium Sq20 represents the
negative energy values which describes the stable nature of EPSPSO.intermedium Sq20.
a
b
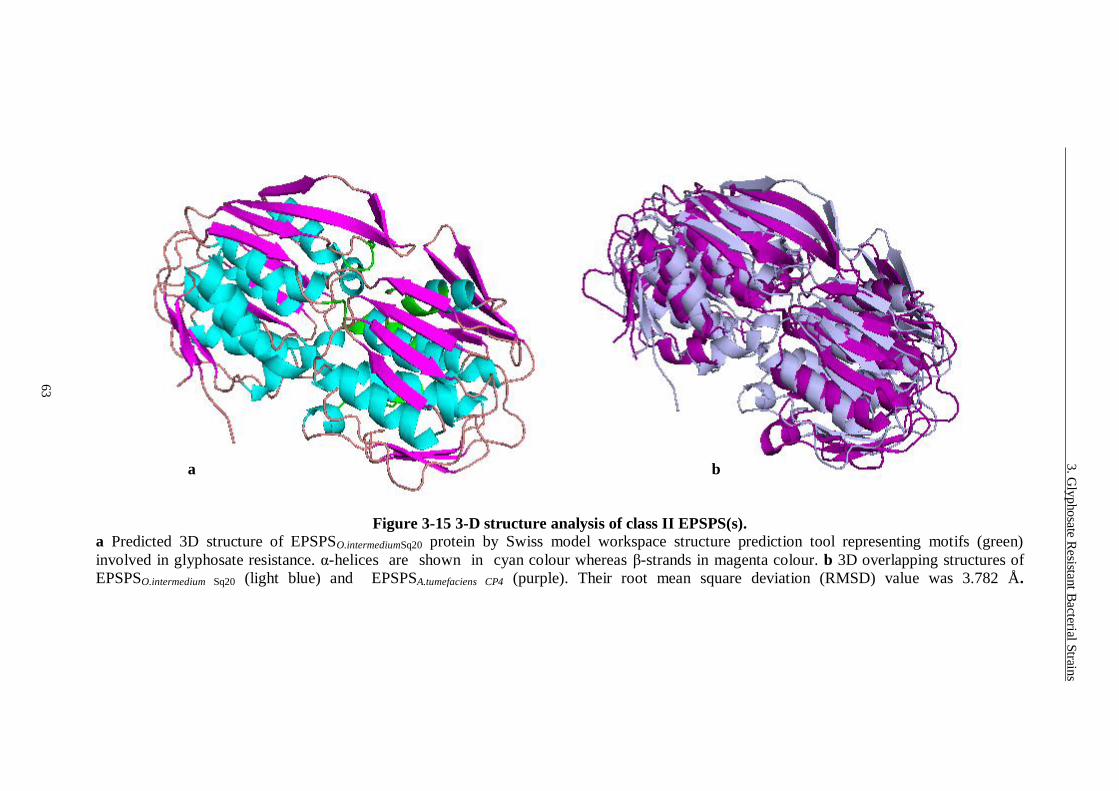
3. G
lyphosate R
esistant B
acterial Strain
s
63
Figure 3-15 3-D structure analysis of class II EPSPS(s).
a Predicted 3D structure of EPSPSO.intermediumSq20 protein by Swiss model workspace structure prediction tool representing motifs (green)
involved in glyphosate resistance. α-helices are shown in cyan colour whereas β-strands in magenta colour. b 3D overlapping structures of
EPSPSO.intermedium Sq20 (light blue) and EPSPSA.tumefaciens CP4 (purple). Their root mean square deviation (RMSD) value was 3.782 Å.
a b

3. Glyphosate Resistant Bacterial Strains
64
3
The Z score, a measure of model quality that predicts the total energy of the
structure [189] was predicted using the ProSA-web server. Z value of -9.82 was
obtained for EPSPSO.intermedium Sq20 protein model represents its good quality. It is
indicated with a dark black dot in the plot (Figure 3-14a). The ProSA-web server also
assesses model quality by plotting energies as a function of amino acid residues and
negative values of energy accounts for the stability of protein structure. In case of
EPSPSO.intermedium Sq20, most of the energy values of amino acid residues were found to
be in negative region further confirming that EPSPSO.intermedium Sq20 is a stable protein
(Figure 3-14b). Three dimensional structure of the EPSPSO.intermedium Sq20 was
generated by Swiss Model and the final model containing 6-448 amino acid residues
was visualized by PyMOL (Figure 3-15a). The EPSPSO.intermedium Sq20 model was
superimposed separately on the structure of EPSPSA.tumefaciens CP4 (Figure 3-15b) and
the results proclaimed that the allocation of α helices and β sheets in the
EPSPSO.intermedium Sq20 was similar to that of EPSPSA.tumefaciens CP4. In addition, all the
amino acid residues facilitating glyphosate binding in EPSPSA.tumefaciens CP4 were
present in the corresponding regions of EPSPSO.intermedium Sq20.
3.4 Discussion
The focal goal of our research was to isolate a bacterial strain from indigenous soil
competent to tolerate high glyphosate concentration, and thus characterize the gene(s)
involved. In this regard, a glyphosate resistant bacterial strain Ochrobactrum
intermedium Sq20 was isolated. Sq20 was found capable of degrading glyphosate as
well. The genus Ochrobactrum belongs to the family Brucellaceae and Ochrobactrum
intermedium is deemed as an emerging human environmental opportunistic pathogen
with mild virulence [191]. Ochrobactrum strains are studied as potential
bioremediation agents and plant growth-promoting rhizobacteria [192-196]. They are
involved in the degradation of organophosphorus pesticides such as parathion and
methylparathion [197], phenol [198], dimethylformamide (DMF, toxic organic
solvent) [199], petroleum waste [200] and chlorothalonil [201]. Ochrobactrum spp.
also plays an important role in removal of toxic metals such as chromium, cadmium
and copper from the environment [202]. In recent years, O. intermedium is of
particular interest due to its biocatalyst properties. The biosurfactant produced by
O. intermedium can potentially and successfully be used for the remediation of PAHs
contaminated soil in industrial areas [203]. These properties of O. intermedium have

3. Glyphosate Resistant Bacterial Strains
65
generated considerable interest for its use in bioremediation. Moreover,
O. intermedium Sq20 is indigenously isolated from glyphosate contaminated soil
therefore its degrading properties can be exploited for decontamination of glyphosate
at point sources.
Glyphosate biodegradation data by O. intermedium Sq20 showed that linear
regression equation can be used for its approximation. Furthermore, implication of
first order kinetics illustrated that time required for biodegradation of glyphosate
depends upon initial concentration of glyphosate. The rational of this research work
was to study glyphosate resistant genes therefore a glyphosate resistant gene (aroA)
was amplified from O. intermedium Sq20 in this context. Sequence analysis of this
putative aroA gene showed that it shares 97% sequence identity with other reported
Ochrobactrum spp. aroA genes. Moreover, EPSPSO.intermedium Sq20 belongs to class II
EPSP synthases and contains conserved domains identical to ones found in proteins
that impart natural glyphosate resistance in some bacteria. Particularly five already
reported motifs characteristic of class II EPSP synthases have been identified in
EPSPSO.intermedium Sq20 as: GDKX in which K28 residue is involved in PEP binding;
SAQXK in which S173 and Q175 residues facilitates S3P binding; RPMXR, RXHXE
and NXTR in which the positive charges of the guanidine groups of R128, R200 and
R274 destabilizes the glyphosate binding respectively [53].
Although researchers have discovered mutant EPSPS genes from different
resistant microbes [150, 161, 165], there is only one report of resistant EPSPS from
Ochrobactrum sp. having potential for tolerating 200 mM glyphosate [204]. In order
to comprehend molecular profile and physicochemical properties of EPSPSO.intermedium
Sq20 protein, Expasy‟s ProtParam tools were employed by measuring different
parameters. It is already reported that protein stability is predicted by computing its
instability index. Occurrence of certain dipeptides in stable proteins is different in
contrast with unstable ones. Weight values of instability are allocated to the proteins
on the basis of incidence of certain dipeptides. Proteins exhibiting an instability index
smaller than 40 are considered as stable, therefore the instability index of
EPSPSO.intermedium Sq20 protein calculated as 32.06 signifies the stable nature of EPSPS
protein. The extinction coefficient demonstrates about light absorbed by a protein
depending on concentrations of Cys, Trp and Tyr at a certain wavelength. It is also
helpful in the quantitative study of protein-protein and protein-ligand interactions in

3. Glyphosate Resistant Bacterial Strains
66
solution. Low extinction coefficient (10095, 9970) indicates less concentration of
Cys, Trp and Tyr in EPSPSO.intermedium Sq20 and therefore this protein cannot be
scrutinized by using UV spectral methods. Thermal stability of a protein is predicted
by its aliphatic index calculated on the basis of relative volume of that protein
occupied by its aliphatic side chains (alanine, valine, isoleucine, and leucine).
Aliphatic index is also considered as a positive factor in thermostability of globular
proteins and low thermal stability value of EPSPSO.intermedium Sq20 indicates flexible
structure of this protein [182]. High aliphatic index (90.4) of EPSPSO.intermedium Sq20
proposes that this protein has stable behaviour for a broad range of temperatures. The
Grand Average hydropathy (GRAVY) value for a protein is calculated as the sum of
hydropathy values of all the amino acids divided by the number of residues in the
protein sequence. High negative value of GRAVY (-0.039) demonstrates that
EPSPSO.intermedium Sq20 is hydrophilic in nature. Isoelectric point (pI) is described as the
pH upon which protein surface is covered with charge but has zero net electrical
charge. pI calculation is helpful in developing buffer systems for purification of
proteins by isoelectric focusing method [205]. The pI computed to be 5.21 indicates
EPSPSO.intermedium Sq20 as moderately acidic. Generally it is perceived that GRAVY
decreases with decrease in pI.
The secondary structure of EPSPSO.intermedium Sq20 protein revealed that random
coils were the most copious followed by alpha helix and other secondary structure
elements (extended strand and beta turns). Random coils are indispensable in the
context of protein flexibility and conformational changes as well as enzymatic
turnover [206]. Secondary structure and disorder prediction by using Phyre2 showed
that EPSPSO.intermedium Sq20 holds 8% disorder therefore indicating the absence of weak
regions in this protein. Phyre2 uses a folding simulation method to model regions of a
query with no detectable similarities to known structures and indicates the disordered
regions. The homology modeling of the protein was accomplished using two
programs Geno3D and Swiss Model. The results acquired from these programs are
compared in Table 3.5. In order to rationalize predicted models, Ramchandran map
calculations computed by using PROCHECK software were employed to create
Ramachandran plots illustrating the φ and ψ angles distribution in the models. The
results of Swiss Model showed that 91.8% of amino acid residues were found to be
localized in most favoured regions. Additional allowed regions hold 8.0% of amino

3. Glyphosate Resistant Bacterial Strains
67
acid residues. 0.2% of amino acid residues were found in generously allowed regions.
It is reported that a model having more than 90% residues in the most favoured
regions is expected as a good quality model so above results affirm the good quality
of EPSPSO.intermedium Sq20 predicted model. This comparison also suggests that the
model generated by Swiss model is better than the model generated by Geno3D.
Model reliability was further confirmed by Z score calculated by ProSA-web server
found to be -9.82. The black spot in optimum range represents that protein has valid
structure. Z score is found to depend on the protein length and negative Z scores are
excellent for a reliable model. Z score also embodies the overall quality of model and
calculates the total energy deviation of the protein structure with respect to energy
distribution acquired from random conformations. Local model quality was
determined by plotting energies as a function of amino acid sequence positions.
Positive values indicate the problematic or flawed parts of the input structure. Usually
a plot based on energies of single residues contains large fluctuations. Therefore, it is
not reliable for model evaluation. For that reason the plot is smoothed by plotting
average energies of 40 amino acid residues [189]. The calculated Z score and energy
plot revealed that Swiss Model of EPSPSO.intermedium Sq20 is reliable.
Table 3.5 Comparative analysis of Swiss Model and Geno 3D computed models
of EPSPSO.intermedium Sq20 done by Ramachandran plot calculations.
Software Parameters Value (%)
Swiss Model Residues in most favoured regions 91.8
Residues in additional allowed regions 8.0
Residues in generously allowed regions 0.3
Residues in disallowed regions 0.0
Geno 3D Residues in most favoured regions 63.9
Residues in additional allowed regions 24.9
Residues in generously allowed regions 4.9
Residues in disallowed regions 0.8

3. Glyphosate Resistant Bacterial Strains
68
This study demonstrates the isolation and identification of a glyphosate
resistant bacterial strain Ochrobactrum intermedium Sq20 that that can utilize
glyphosate as an energy source. Therefore the competence of this isolate to survive
and reproduce at high concentrations of the glyphosate in indigenous soil marks it as a
good candidate for the remediation of glyphosate contaminated environments. The
characterization of class II EPSPS encoded by the aroAO.intermedium Sq20 gene was
carried out and a computational 3-D model was predicted. 3-D model further
corroborated through various parameters of validation and authenticity was found
significant and reliable for investigating the molecular features that result in
glyphosate resistance.

69
4. Isolation and Characterization of
Glyphosate Degrading Bacterial Strains
4.1 Introduction
Glyphosate is a broad spectrum herbicide differentiated by a direct carbon to
phosphorus (C-P) bond, thermally and chemically inert to non-biological degradation
processes [207, 208]. Glyphosate belongs to the amino acid herbicide family and its
mode of action involves inhibition of shikimic acid pathway enzyme, 5-
enolpyruvylshikimic acid-3-phosphate synthase (EPSPS), activity resulting in plant
death [209].
It has been used on crop and non crop areas in multifarious practices for many
years [65, 210]. Agricultural use of this herbicide increased drastically by the
introduction and commercial launch of genetically modified glyphosate resistant
crops [211]. Glyphosate is mostly used for weed management purposes in glyphosate
resistant crops in developed countries. Currently glyphosate resistant crops such as
maize, cotton, soybean, sugar beet, canola and wheat etc have covered more than 80%
of the planted area. The production of these crops has surpassed 500,000 tons per year
and is approximated to go beyond 10,00000 tons in next few years [67, 212].
Indiscriminate use of glyphosate in agriculture, horticulture and at amenity areas is
leading to accumulation of its residues in soil, food and water due to its mobility and
water solubility properties. Contamination of terrestrial and aquatic ecosystems has
detrimental effects on different biota and biogeochemical cycles as well. Weed
resistance is also an emanation of extensive use of glyphosate [213]. Glyphosate has
been reported to disturb the balance between beneficial and pathogenic soil micro
flora resulting in curtailed nutrient availability and disease severity owing to
weakened plant defense system [214]. The chelation property of glyphosate may
hinder the uptake and translocation of micronutrients within plant tissues through
formation of poorly soluble glyphosate metal complexes [215]. Glyphosate has been
reported to cause oxidative stress and genotoxicity in aquatic organisms [216].
Daphnia magna showed ~100% abortion rate of eggs and embryonic stages at 1.35

4. Glyphosate Degrading Bacterial Strains
70
mg a.i./L of Roundup [217]. Moreover, reproduction of soil dwelling earthworms was
found to reduce by 56% within three months after glyphosate application [218]. As
Acidobacteria plays an important role in biogeochemical processes, therefore their
low abundance in response to glyphosate exposure may hamper natural processes.
Furthermore, about 40% reduction of mycorrhization was observed in soils that had
been amended with the mycorrhizal fungi, Glomus mosseae after the application of
glyphosate [219].
Toxicology studies of glyphosate showed that glyphosate can change the DNA
of sister chromatids in human lymphocytes. Moreover occupational exposure of
glyphosate causes non-Hodgkin lymphoma (NHL) [98]. Glyphosate is reported to
have tumorigenic, hepatorenal and teratogenic effects which can be expounded on the
basis of oxidative stress and endocrine disruption resulting in metabolic alterations
[220]. International Agency for Research on Cancer (IARC) of World Health
Organization (WHO) has classified glyphosate as possibly carcinogenic (category 2A)
to humans on the basis of epidemiological, animal and in vitro studies [221].
Therefore its removal from environment is a priority and needs to be addressed
promptly. The easy, eco-friendly and cost effective way to decontaminate polluted
environment is bioremediation.
4.1.1 Glyphosate Biodegradation
Regarding glyphosate biodegradation certain potential glyphosate degrading micro
organisms have been isolated from organophosphonates contaminated soils [222].
Different glyphosate degrading bacteria and their metabolic products are given in
Table 4.1. Bacterial consortia were reported to involve in biodegradation of
glyphosate yielding AMPA [223, 224]. Pseudomonas sp. PG2982 was first bacterial
strain found proficient to utilize glyphosate as P source. This degradation mechanism
involved cleavage of C-P bond yielding sarcosine instead of AMPA [225]. Same
mechanism was reported in Alcaligenes sp. [118], Arthrobacter sp. [226],
Pseudomonas sp. GLC11 [227], Pseudomonas sp. 4ASW [66] and Achromobacter sp.
[228]. Certain bacterial strains were found capable of utilizing glyphosate as P source
while yielding AMPA such as Pseudomonas sp. SG-1 [229], Flavobacterium sp.
[223], Pseudomonas sp. LBr [230], Achromobacter sp. LW9 [231], O. anthropi
LBAA [116], O. anthropi GPK3 [228] and Ochrobactrum sp. GDOS [232]. The
number of bacterial strains capable of utilizing glyphosate other than P source is very

4. Glyphosate Degrading Bacterial Strains
71
limited. Arthrobacter sp. GLP-1 was found able to utilize glyphsate as N source
[233]. Streptomyces sp. StC [234] and Achromobacter sp. LW9 [231] were found to
use glyphosate as C and N source. The reasons may involve slow glyphosate
degradation in cytoplasm or inhibition of degrading enzymes by resultant P [77, 235].
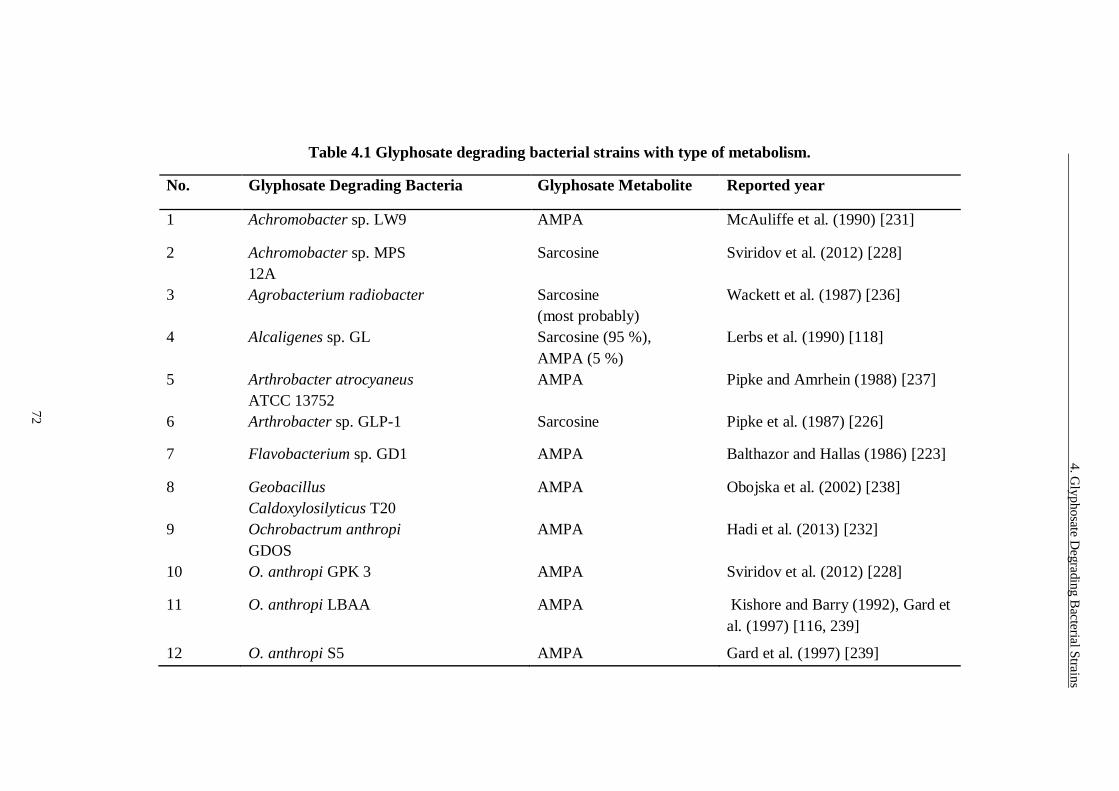
72
4. G
lyp
hosate D
egrad
ing B
acterial Strain
s
Table 4.1 Glyphosate degrading bacterial strains with type of metabolism.
No. Glyphosate Degrading Bacteria Glyphosate Metabolite Reported year
1 Achromobacter sp. LW9 AMPA McAuliffe et al. (1990) [231]
2 Achromobacter sp. MPS
12A
Sarcosine Sviridov et al. (2012) [228]
3 Agrobacterium radiobacter Sarcosine
(most probably)
Wackett et al. (1987) [236]
4 Alcaligenes sp. GL Sarcosine (95 %),
AMPA (5 %)
Lerbs et al. (1990) [118]
5 Arthrobacter atrocyaneus
ATCC 13752
AMPA Pipke and Amrhein (1988) [237]
6 Arthrobacter sp. GLP-1 Sarcosine Pipke et al. (1987) [226]
7 Flavobacterium sp. GD1 AMPA Balthazor and Hallas (1986) [223]
8 Geobacillus
Caldoxylosilyticus T20
AMPA Obojska et al. (2002) [238]
9 Ochrobactrum anthropi
GDOS
AMPA Hadi et al. (2013) [232]
10 O. anthropi GPK 3 AMPA Sviridov et al. (2012) [228]
11 O. anthropi LBAA AMPA Kishore and Barry (1992), Gard et
al. (1997) [116, 239]
12 O. anthropi S5 AMPA Gard et al. (1997) [239]
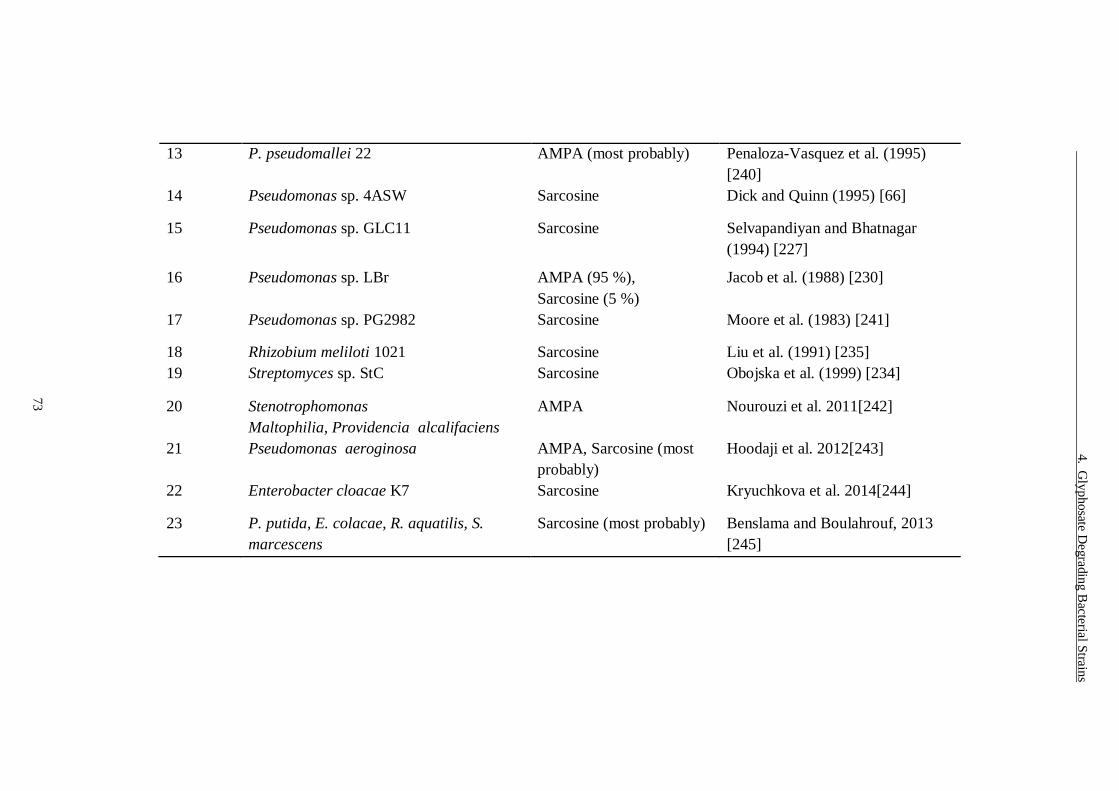
73
4. G
lypho
sate Deg
radin
g B
acterial Strain
s
13 P. pseudomallei 22 AMPA (most probably) Penaloza-Vasquez et al. (1995)
[240]
14 Pseudomonas sp. 4ASW Sarcosine Dick and Quinn (1995) [66]
15 Pseudomonas sp. GLC11 Sarcosine
Selvapandiyan and Bhatnagar
(1994) [227]
16 Pseudomonas sp. LBr AMPA (95 %),
Sarcosine (5 %)
Jacob et al. (1988) [230]
17 Pseudomonas sp. PG2982 Sarcosine Moore et al. (1983) [241]
18 Rhizobium meliloti 1021 Sarcosine Liu et al. (1991) [235]
19 Streptomyces sp. StC Sarcosine Obojska et al. (1999) [234]
20 Stenotrophomonas
Maltophilia, Providencia alcalifaciens
AMPA Nourouzi et al. 2011[242]
21 Pseudomonas aeroginosa AMPA, Sarcosine (most
probably)
Hoodaji et al. 2012[243]
22 Enterobacter cloacae K7 Sarcosine Kryuchkova et al. 2014[244]
23 P. putida, E. colacae, R. aquatilis, S.
marcescens
Sarcosine (most probably) Benslama and Boulahrouf, 2013
[245]

4. Glyphosate Degrading Bacterial Strains
74
4.1.2 Pathways of Glyphosate Biodegradation
Two major pathways of glyphosate biodegradation have been reported. One pathway
involves glyphosate utilization as C source resulting in aminomethylphosphonic acid
(AMPA) production involving glyphosate oxidoreductase (GOX) gene. Second
pathway utilizes glyphosate as P source with production of sarcosine and glycine via
C-P lyase [130].
AMPA Pathway of Glyphosate Biodegradation
This pathway involves glyphosate oxidoreductase (GOX) which cleaves C-N bond of
glyphosate yielding AMPA and glyoxylate in the presence of magnesium and flavin
adenine dinucleotide (FAD) [116]. Glyoxylate enters the Krebs cycle and metabolizes
to its intermediates. GOX gene was isolated from Ochrobactrum antrophi strain
LBAA previously known as Achromobacter sp. and transformed in maize and canola
to augment glyphosate resistance [116]. The glyphosate oxidoreductase (GOX) of
flavin monooxygenase superfamily has been recently purified from O. anthropi GPK
3. It shows maximum activity under alkaline conditions with relatively low affinity
for glyphosate and holds single noncovalently bound FAD molecule per subunit. The
activity of purified enzyme is influenced by oncotic and osmotic pressure with
relative low stability at normal conditions and is hindered by Cu2+
, Ag+, Mn
2+ and p-
chloromercurybenzoic acid. GOX has wide range of substrates including
iminodiacetic acid and phosphonomethyliminodiacetic acid and have ability to
transform glycine, sarcosine and D-alanine to some extent [228]. The thermostable
form of GOX is presumed to exist in thermophilic Geobacillus caldoxylosilyticus T20
isolated from central heating system [238]. C-P lyase pathway was exclusively
considered for glyphosate biodegradation before the discovery of phosphonatase
pathway in O. anthropi GPK 3 [228]. Phosphonatase pathway is involved in
metabolism of phosphonates and consists of two steps. First steps involves
transamination of 2-aminoethylphosphonate (2-AEP, a natural phosphonate ciliatine)
by 2-AEP pyruvate aminotransferase (EC 2.6.1.37, a 80 kDa homodimer with pH
optimum of 8.5) [246]. Second step involves hydrolysis of C-P bond of resulting
phosphonoacetaldehyde by phosphonoacetaldehyde hydrolase (EC 3.11.1.1, a 62 kDa
Mg2+
dependent homodimer with optimal pH of 7.5) [247]. Although these two
enzymes are highly substrate specific and no reports about their involvement in
glyphosate mineralization exist [248-250] however some reports showed likelihood of

4. Glyphosate Degrading Bacterial Strains
75
phosphonatase involvement in AMPA degradation [223, 229]. O. anthropi GPK 3
showed two distinctive activities of transaminase. First activity is related to ordinary
2-AEP pyruvate aminotransferase while the second one is related to AMPA
metabolism resulting in production of phosphonoformaldehyde. It is further
metabolized by phosphonatase to formaldehyde and inorganic phosphate [228].
Sarcosine Pathway of Glyphosate Biodegradation
C-P lyase pathway is most prevalent route used by microorganisms for phosphonates
utilization. C-P bond is cleaved by two types of enzymes, first one are substrate
specific hydrolases while second one are non specific C-P lyases. Glyphosate does not
have active C-P bond (destabilized by nearby oxygen of carboxyl, carbonyl, or epoxy
group) which is required by organophosphonate degrading hydrolases [249, 251].
Inactive C-P bond is highly resistant to enzymatic and chemical hydrolysis therefore
its cleavage needs a multienzyme complex activity such as C-P lyase capable of
converting phosphonates into inorganic phosphonate and corresponding metabolite.
C-P lyase studies in E. coli revealed that this complex have 14 gene which
collectively catalyzes the breakdown of phosphonates [252]. C-P lyases composition
in both gram negative and gram positive bacteria is assumed to be alike [249]. The
phn operon is a part of Pho regulon and induced by phosphorus starvation only. Of
this complex of 14 genes, phnCDE are considered to encode an ATP binding cassette
transporter, phnF acts as a repressor protein and phnNOP have regulatory or some
auxiliary roles [253, 254]. PhnGHIJKLM embodies the core components of the C-P
lyase pathway and mutation or deletion in any one of these genes components put an
end to C-P lyase activity [255]. phnJ is found to catalyze the main C-P cleavage and
this reaction involves the conversion of 5-phosphoribosyl-1-phosphonate to 5-
phosphoribosyl-1,2-cyclic phosphate in the presence of S-adenosyl-l-methionine
(SAM) [254]. PhnI deglycosylates GTP and ATP to ribose-5-triphosphate [256]. The
proposed pathway of phosphonate metabolism revealed that core component of C-P
lyase complex performs two major functions: PhnGHI facilitate the coupling of
phosphonate and ATP whereas PhnJ aids in cleavage of C-P bond.
Glyphosate mineralization through C-P lyase activity leads to the production
of sarcosine which is then metabolized into formaldehyde and glycine by sarcosine
oxidase. Formaldehyde is further oxidized to carbon dioxide and water whereas

4. Glyphosate Degrading Bacterial Strains
76
glycine enters in various biosynthesis processes as a substrate [225, 257]. Such type
of glyphosate degradation pathway is exhibited by those bacterial strains only which
have glyphosate as sole P source. But there are many easily available P sources in
natural environment affecting glyphosate utilization. The activity of C-P lyase is
down regulated by extracellular P which is commonly found in environment [66, 77,
237]. Certain bacterial strains such as mutant form of Arthrobacter sp. GLP-1/Nit
[233] and Alcaligenes sp. GL [118] metabolize glyphosate to sarcosine even in
presence of extracellular P.
4.1.3 Response Surface Methodology
Different parameters are involved in biodegradation process so their optimization is
required for acquisition of highest process efficiency [258]. Usually one factor at a
time approach is used for optimization studies but this method is tiresome and neglect
mutual interaction of parameters leading to misapprehension of results. Conversely,
statistical methods are well planned, economical and lessen the tendency of
ambiguous results [259]. Response Surface Methodology (RSM) is one of the
statistical designs used to optimize experimental procedures in a cost effective manner
by reducing their numerical noise. RSM is a multivariate statistical method that
determines the mutual effect of different variables on response of an experimental
design and has been used effectively for optimization of xenobiotics degradation. It is
very effective design which involves multiple factor experiments, investigate the
relationships among factors, unearth the best fit experimental conditions and predict
the response [260]. A response under the influence of different independent variables
can be optimized through careful designing of experiments. This leads to the model
with complete depiction of variables, their mutual interaction and effect on response.
RSM uses different experimental designs but central composite rotatable design
(CCRD) was used in current study. A comparison between CCRD and other RSM
designs has demonstrated that CCRD has better prediction than other designs because
it include a situation when all variables are in their boundary value simultaneously
[261].
The current work was aimed to study the biodegradation of glyphosate by
indigenously isolated bacterial strains and identification of potential genes involved in
degradation process. Therefore, an isolate found to be effective in glyphosate

4. Glyphosate Degrading Bacterial Strains
77
biodegradation, Comamonas odontotermitis P2, was then subjected to central
composite design (CCD) through RSM for optimization of different growth
parameters for biodegradation of glyphosate.
4.2 Materials and Methods
4.2.1 Enrichment and Isolation of Glyphosate Degrading Bacterial
Strains
Soil was collected from 3 different locations in an experimental field (-34.018°,
150.67°) at cobbitty site, Plant Breeding Institute, University of Sydney, NSW
Australia with a prior history of glyphosate spray. The field was again sprayed with
glyphosate, soil samples were collected at intervals of 4, 7 and 14 days after spraying
at a depth of 5-10 cm of A horizon and saved at 4 °C in sterile zip lock plastic bags.
These samples were dried, sieved and then used for isolation of glyphosate degrading
bacterial strains through enrichment culture technique. Soil samples (10 g each) were
added separately in 250 mL Erlenmeyer flasks containing 100 mL enrichment
medium (EM) (Appendix C) and 1 g/L glyphosate and incubated at 25 °C on rotary
shaker at 150 rpm. After 7 days, 5 mL culture was transferred to fresh EM with same
glyphosate concentration and this sub-culturing was repeated for four weeks. After
four weeks, serial dilutions of enrichment culture were plated on minimal medium
(MM) (Appendix D) agarose plates containing glyphosate and incubated at 30 °C.
Morphologically different bacterial strains were isolated and purified through
repeated streaking. Nutrient agar slants were prepared for preservation of stock
cultures and kept at 4 °C.
4.2.2 Selection of Competent Glyphosate Degrading Bacterial Strain
Growth and glyphosate degradation potential of the bacterial isolates were observed
in 250 mL Erlenmeyer flasks containing 100 mL phosphate free EM medium
supplemented with 500 mg/L glyphosate as sole source of carbon and phosphorus,
incubated in a rotary shaker at 150 rpm and 30 °C for 4 days. The absorption of these
cultures was measured by the spectrophotometer at 590 nm wavelengths. To
investigate utilization of glyphosate as sole carbon source EM medium was used,
whereas for glyphosate utilization as sole P source MM medium was employed
whereas other conditions remained same. All experiments were performed in
triplicates and uninoculated flasks were used as controls. The residual concentration

4. Glyphosate Degrading Bacterial Strains
78
of glyphosate was quantified to determine the degradation potential of all the bacterial
isolates.
4.2.3 Identification and Characterization of Glyphosate Degrading
Bacterial Strains
Genomic DNA of isolated bacterial strains was extracted and 16S rRNA gene analysis
was performed for their molecular characterization (chapter 2, section 2.9). The 16S
rRNA gene sequences of the isolated bacterial strains were submitted in GenBank.
Morphological characterization was carried out by inspecting form, margin and
elevation of isolates on nutrient agar through stereoscope. Shape, pigmentation and
motility were also studied [124]. Differentiation of isolates on the basis of their cell
wall composition was done through Gram staining (chapter 2, section 2.10.2).
Moreover, phylogenetic analysis was carried out as described in chapter 2, section
2.11.5.
4.2.4 Denaturing Gradient Gel Electrophoresis (DGGE) Analysis of
Glyphosate Contaminated Soil
DNA of enrichment cultures was extracted by using InvitrogenTM
genomic DNA
extraction kit according to the instructions provided with kit. The quality and quantity
of extracted DNA was measured by agarose gel electrophoresis and NanoDrop 2000c
spectrophotometer respectively and used for PCR amplification accordingly. PCR-
DGGE of partial eubacterial 16S rRNA gene fragments was carried out as described
in chapter 2 section 2.12. DGGE sequences were submitted in Genbank.
4.2.5 Biodegradation of Glyphosate by Isolated Bacterial Strains
Isolated bacterial strains (P1, P2, P3, P4 and P5) were grown in MM containing 500
mg/L of glyphosate, The cultures were harvested and centrifuged at 6000 rpm for 10
min. The cell pellets were washed with 0.9% saline solution and suspended in the
same solution to obtain an OD of 1. Colony forming units (CFU/mL) of the
suspensions were determined through dilution plate count method and used as
inoculum (2%) for glyphosate biodegradation experiments.
Glyphosate degradation potential of isolated bacterial strains as carbon and
phosphorus source was measured in Erlenmeyer flasks containing 50 mL MM (pH
7.0) supplemented with the 500 mg/L glyphosate. 2% inoculum was added to MM

4. Glyphosate Degrading Bacterial Strains
79
and flasks were incubated at 25 °C in a rotary shaker at 150 rpm. All experiments
were carried out in triplicates and uninoculated flasks were used as controls.
4.2.6 HPLC Analysis of Glyphosate Residues
For HPLC analysis, glyphosate was derivatized with 4-chloro-3,5-
dinitrobenzotrifluoride (CNBF) [129] and analyzed as described in chapter 2 section
2.15.
4.2.7 Optimization of Culture Conditions Using Response Surface
Methodology (RSM)
To optimize the combined effects of pH, temperature and inoculum size on
glyphosate degradation and determination of their optimum combinations, response
surface methodology (RSM) was used. Degradation of glyphosate (500 mg/L) was
monitored in MM as analyzed response after three days of incubation. A three
factor/five level central composite design (CCD) of 23 full factorial with 20 runs was
employed by using Design Expert software (trial version 10, Stat-Ease, Inc., MN,
USA).
Quadratic polynomial equation used to approximate the mathematical
relationship of the response (glyphosate % degradation) of these three variables is
given below:
Y= a0 + a1A + a2 B+ a3C + a12AB + a13AC +
a23BC + a11A2 + a22B
2 + a33C
2 (4-1)
Where Y is the predicted response value, a0 is the value of the fitted response
at the centre point of the design, a1, a2 and a3 are the linear coefficients; a12, a13 and a23
are the cross product coefficients; a11, a22 and a33 are the quadratic coefficients. Based
on the best results of the one-at-a-time method, the ranges and coded levels of three
variables viz. pH (A), temperature (B), and inoculum density (C) are listed in Table
4.2. The design matrix with three variables (pH, temperature, and inoculum
density)×five levels (−1.68, −1, 0, +1, +1.68) is presented in Table 4.2. All the
variables were taken at the coded values.
F test and R2 (correlative coefficient value) was employed to check statistical
significance and quality fit of model respectively. The general experimental setup of
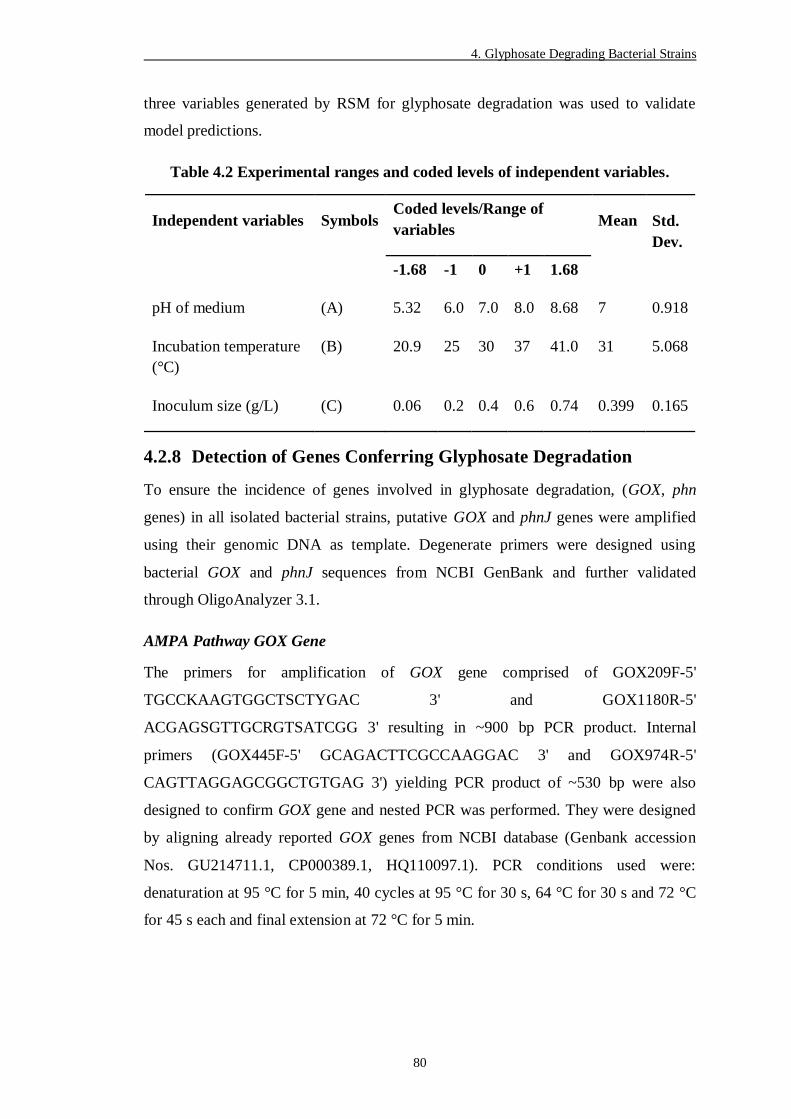
4. Glyphosate Degrading Bacterial Strains
80
three variables generated by RSM for glyphosate degradation was used to validate
model predictions.
Table 4.2 Experimental ranges and coded levels of independent variables.
Independent variables Symbols Coded levels/Range of
variables Mean Std.
Dev.
-1.68 -1 0 +1 1.68
pH of medium (A) 5.32 6.0 7.0 8.0 8.68 7 0.918
Incubation temperature
(°C)
(B) 20.9 25 30 37 41.0 31 5.068
Inoculum size (g/L) (C) 0.06 0.2 0.4 0.6 0.74 0.399 0.165
4.2.8 Detection of Genes Conferring Glyphosate Degradation
To ensure the incidence of genes involved in glyphosate degradation, (GOX, phn
genes) in all isolated bacterial strains, putative GOX and phnJ genes were amplified
using their genomic DNA as template. Degenerate primers were designed using
bacterial GOX and phnJ sequences from NCBI GenBank and further validated
through OligoAnalyzer 3.1.
AMPA Pathway GOX Gene
The primers for amplification of GOX gene comprised of GOX209F-5'
TGCCKAAGTGGCTSCTYGAC 3' and GOX1180R-5'
ACGAGSGTTGCRGTSATCGG 3' resulting in ~900 bp PCR product. Internal
primers (GOX445F-5' GCAGACTTCGCCAAGGAC 3' and GOX974R-5'
CAGTTAGGAGCGGCTGTGAG 3') yielding PCR product of ~530 bp were also
designed to confirm GOX gene and nested PCR was performed. They were designed
by aligning already reported GOX genes from NCBI database (Genbank accession
Nos. GU214711.1, CP000389.1, HQ110097.1). PCR conditions used were:
denaturation at 95 °C for 5 min, 40 cycles at 95 °C for 30 s, 64 °C for 30 s and 72 °C
for 45 s each and final extension at 72 °C for 5 min.

4. Glyphosate Degrading Bacterial Strains
81
Sarcosine Pathway PhnJ Gene
The primers designed for amplification of phnJ were phnJ208F-5'
ATGCCRMTGCCYTAYGGHTG 3' and phnJ1921R-5'
TWASGGCGGMAYSGCRTAGA 3' yielding a PCR product of ~600 bp. These
primers were designed through multiple sequence alignment of previously reported
phnJ genes (Genbank accession Nos. CP015005.1, CP006581.1, AP017605.1). PCR
conditions were: denaturation at 95 °C for 5 min, 35 amplification cycles at 95 °C for
1 min, 56 °C for 1 min and 72 °C for 1 min and final extension at 72 °C for 10 min.
All PCR products were sequenced from Australian Genome Research Facility Ltd.
and these partial gene sequences were submitted to Genbank.
4.3 Results
4.3.1 Selection and Identification of Glyphosate Degrading Bacterial
Strain
Through enrichment culture technique, five bacterial strains were isolated. All the five
bacterial isolates were found capable to utilize glyphosate as a sole C source (in EM
medium) and as sole P source (in MM medium) separately (Figure 4-1a, 4-1b).
Significantly higher growth and glyphosate utilization was observed by the isolate P2
in both cases. For this isolate, culture ODs of 0.29 and 0.24 were obtained
respectively when glyphosate was used as sole C and P sources as compared to ≤ 0.12
OD for all other isolates after 4 days of incubation. Moreover, these isolates were
capable to grow in EM medium (without KP) containing 500 mg/L glyphosate whilst
utilizing it as sole sources of phosphorus and carbon simultaneously. However, after 4
days of incubation, complete degradation of glyphosate was observed only by the
isolate P2 (Figure 4-2). Additionally, 100% glyphosate utilization was observed by P2
as compared to ≤ 50% utilization by the remaining four isolates.
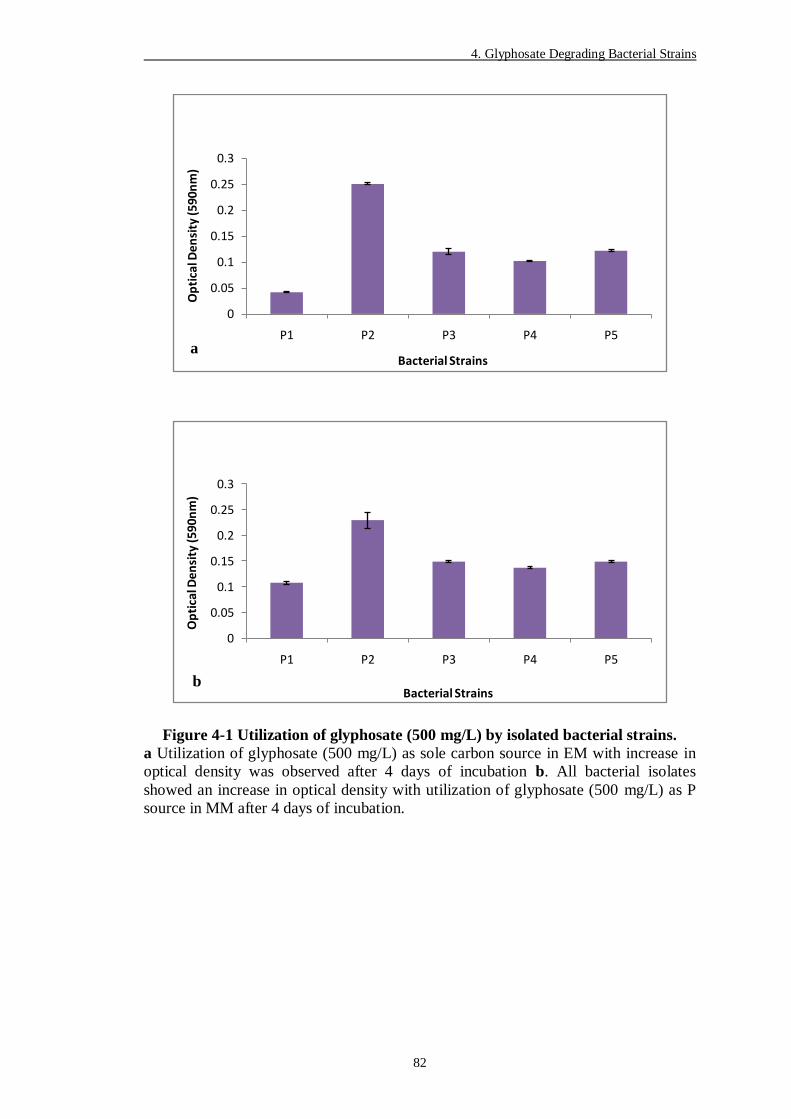
4. Glyphosate Degrading Bacterial Strains
82
Figure 4-1 Utilization of glyphosate (500 mg/L) by isolated bacterial strains.
a Utilization of glyphosate (500 mg/L) as sole carbon source in EM with increase in
optical density was observed after 4 days of incubation b. All bacterial isolates
showed an increase in optical density with utilization of glyphosate (500 mg/L) as P
source in MM after 4 days of incubation.
0
0.05
0.1
0.15
0.2
0.25
0.3
P1 P2 P3 P4 P5
Op
tica
l De
nsi
ty (
590n
m)
Bacterial Strains
0
0.05
0.1
0.15
0.2
0.25
0.3
P1 P2 P3 P4 P5
Op
tica
l Den
sity
(590
nm
)
Bacterial Strains
a
b
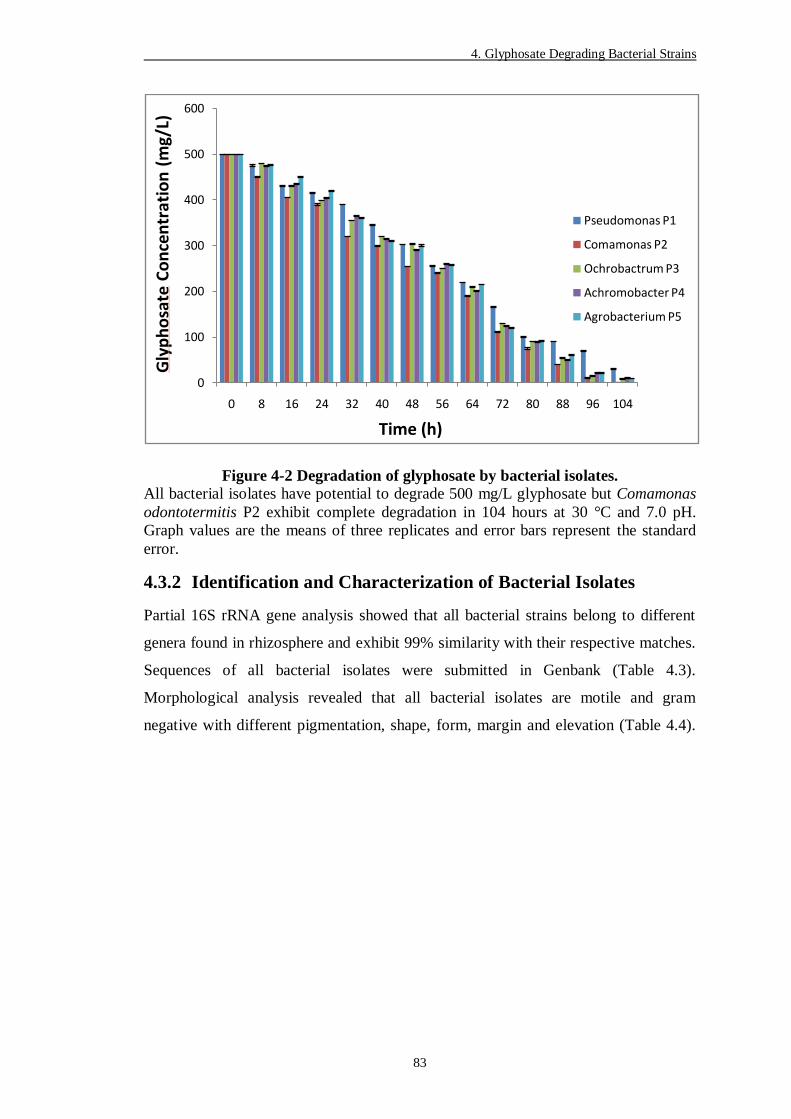
4. Glyphosate Degrading Bacterial Strains
83
Figure 4-2 Degradation of glyphosate by bacterial isolates.
All bacterial isolates have potential to degrade 500 mg/L glyphosate but Comamonas
odontotermitis P2 exhibit complete degradation in 104 hours at 30 °C and 7.0 pH.
Graph values are the means of three replicates and error bars represent the standard
error.
4.3.2 Identification and Characterization of Bacterial Isolates
Partial 16S rRNA gene analysis showed that all bacterial strains belong to different
genera found in rhizosphere and exhibit 99% similarity with their respective matches.
Sequences of all bacterial isolates were submitted in Genbank (Table 4.3).
Morphological analysis revealed that all bacterial isolates are motile and gram
negative with different pigmentation, shape, form, margin and elevation (Table 4.4).
0
100
200
300
400
500
600
0 8 16 24 32 40 48 56 64 72 80 88 96 104
Pseudomonas P1
Comamonas P2
Ochrobactrum P3
Achromobacter P4
Agrobacterium P5
Gly
ph
osa
teC
on
cen
trat
ion
(m
g/L)
Time (h)
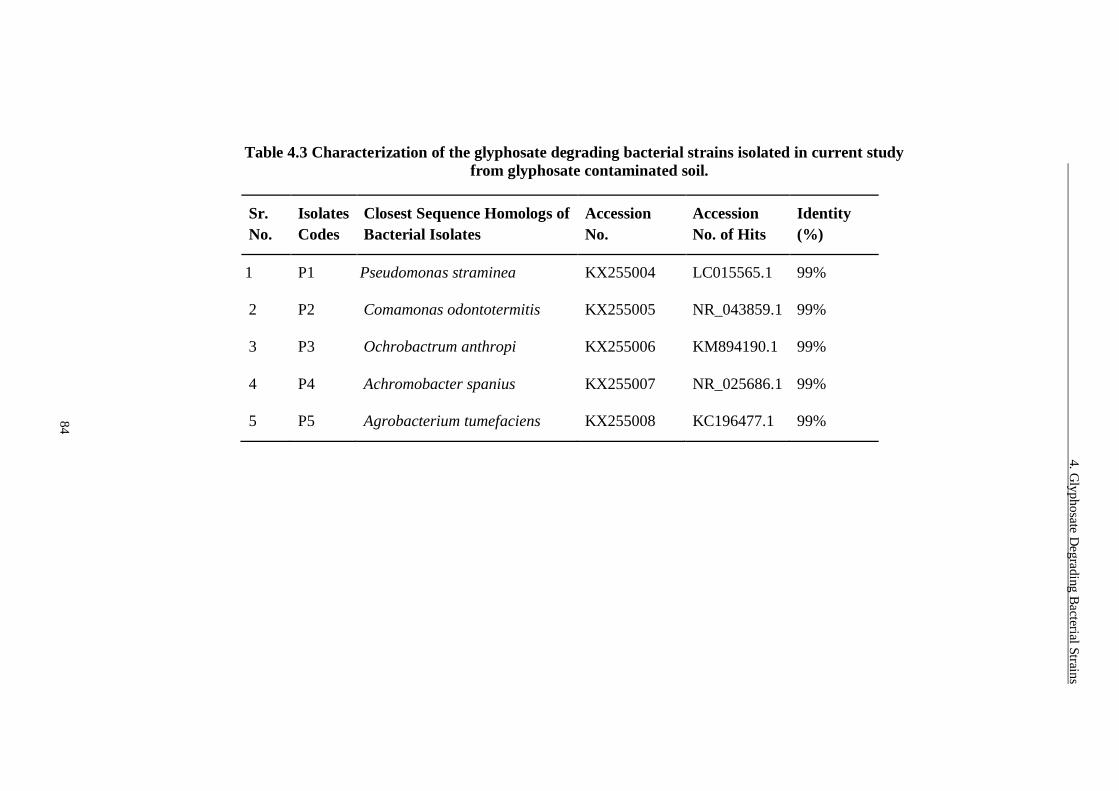
84
4. G
lyph
osate D
egrad
ing
Bacterial S
trains
Table 4.3 Characterization of the glyphosate degrading bacterial strains isolated in current study
from glyphosate contaminated soil.
Sr.
No.
Isolates
Codes
Closest Sequence Homologs of
Bacterial Isolates
Accession
No.
Accession
No. of Hits
Identity
(%)
1 P1 Pseudomonas straminea KX255004 LC015565.1 99%
2 P2 Comamonas odontotermitis KX255005 NR_043859.1 99%
3 P3 Ochrobactrum anthropi KX255006 KM894190.1 99%
4 P4 Achromobacter spanius KX255007 NR_025686.1 99%
5 P5 Agrobacterium tumefaciens KX255008 KC196477.1 99%
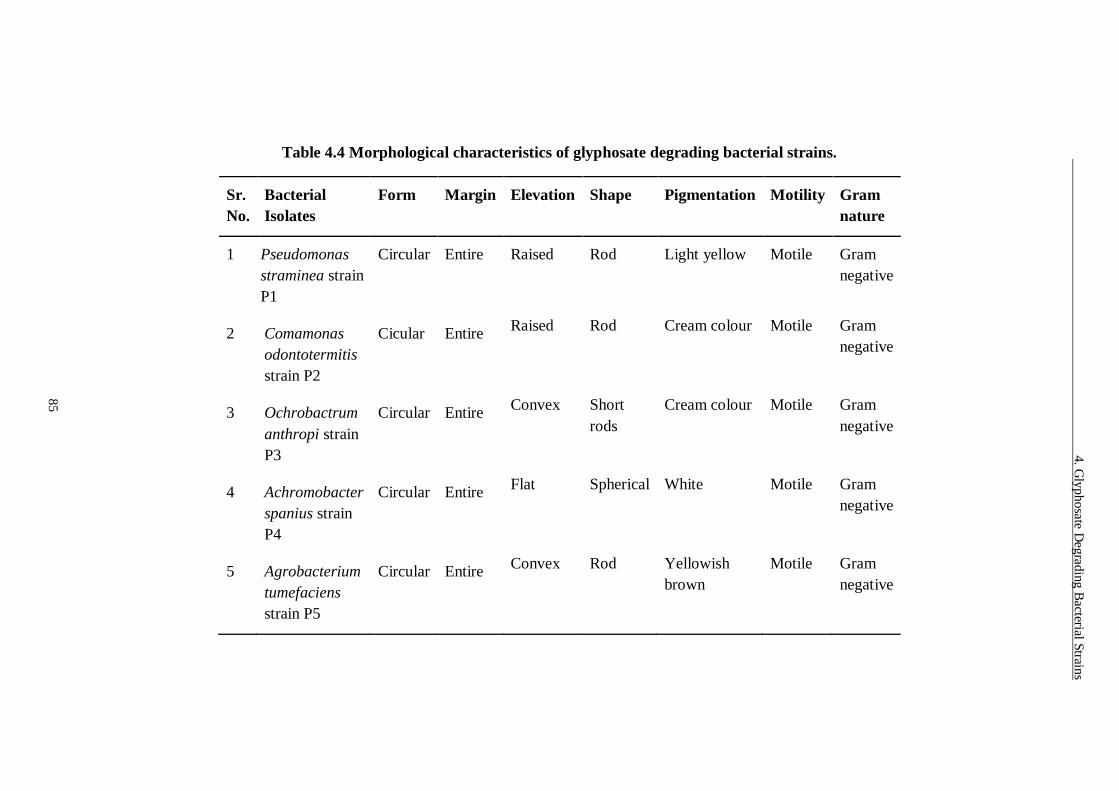
85
4. G
lyph
osate D
egrad
ing
Bacterial S
trains
Table 4.4 Morphological characteristics of glyphosate degrading bacterial strains.
Sr.
No.
Bacterial
Isolates
Form Margin Elevation Shape Pigmentation Motility Gram
nature
1 Pseudomonas
straminea strain
P1
Circular Entire Raised Rod Light yellow Motile Gram
negative
2 Comamonas
odontotermitis
strain P2
Cicular Entire Raised Rod Cream colour Motile Gram
negative
3 Ochrobactrum
anthropi strain
P3
Circular Entire Convex Short
rods
Cream colour Motile Gram
negative
4 Achromobacter
spanius strain
P4
Circular Entire Flat Spherical White Motile Gram
negative
5 Agrobacterium
tumefaciens
strain P5
Circular Entire Convex Rod Yellowish
brown
Motile Gram
negative

4. Glyphosate Degrading Bacterial Strains
86
Through partial 16S rRNA gene (900 bp) analysis using (BLASTn) of the
NCBI, sequences with maximum identity were retrieved for each isolate, aligned and
phylogenetic trees were constructed. P1 isolate showed 99% identity with
Pseudomonas straminea AF95 (LC015565.1) and clustered among type strains of
Pseudomonas straminea and other Pseudomonas spp. indicated in phlogenetic tree
(Figure 4-3). Isolate P2 was found in the lineage of genus Comamonas and showed
maximum sequence identity (99%) with C. odontotermitis strain Dant 3-8
(NR_043859.1). Homologous 16S rRNA gene sequences obtained from the NCBI
were aligned with P2 to generate a phylogenetic tree (Figure 4-4) showing that P2
falls in the same cluster with the type strains of C. odontotermitis and other
Comamonas sp. The approximate phylogenetic affiliations of P3 determined through
nucleotide BLAST analysis of 16S rRNA gene sequence exhibited 99% identity with
Ochrobactrum anthropi E46b (KM894190.1). Phylogenetic tree constructed by
Neighbor joining algorithm showed maximum similarity of P3 with nearest database
entries of other ochrobactrum spp (Figure 4-5). Similarly BLAST analysis of P4
showed maximum sequence similarity with Achromobacter spanius (NR_025686.1)
and phylogenetic analysis demonstrated distribution of this isolate among closely
related Achromobacter spp. (Figure 4-6). Isolate P5 illustrated 99% identity with
Agrobacterium tumefaciens A46 (KC196477.1) and dendrogram constructed through
MEGA 6 showed distribution of this isolate among Agrobacterium sp. on the basis of
sequence similarity (Figure 4-7).

4. Glyphosate Degrading Bacterial Strains
87
Figure 4-3 Phylogenetic tree of Pseudomonas straminea P1.
Based on similarity of 16S rRNA gene sequences with closely related Pseudomonas
spp. Neigbor joining method was used to construct this dendrogram with 1000
iterations. Accession numbers of related sequences are given in brackets.
Pseudomonas sp.Acj 106(AB480754.1)
Pseudomonas fulva ATY63(HQ219982.1)
Pseudomonas putida F29(EF204245.1)
Pseudomonas fulva MRC41(KU672374.1)
Pseudomonas sp.Fa27(AY131221.1)
Pseudomonas parafulva AJ 2130(AB060133.1)
Pseudomonas argentinensis NWP3(KJ875601.1)
Pseudomonas straminea P1(KX255004.1)
Pseudomonas straminea AF95(LC015565.1)
Pseudomonas putida GG59(KJ850210.1)
Pseudomonas argentinensis HDDMM04(EU723826.1)
Pseudomonas sp.EK-I1(GU935266.1)
Pseudomonas sp.M549(AB461834.1)
Pseudomonas sp.M516(AB461802.1)
Pseudomonas straminea IAM 1587(AB060135.1)
Pseudomonas sp.Bg-8(HQ916748.1)
Pseudomonas straminea NBRC 16665(AB681101.1)
Pseudomonas sp.M130(AB461665.1)
Pseudomonas sp.M121(AB461657.1)
Pseudomonas straminea(D84023.1)
Pseudomonas fulva ALEB41(KF460530.1)
Pseudomonas sp.MG-2011-7-BJ(FR872475.1)

4. Glyphosate Degrading Bacterial Strains
88
Figure 4-4 Neighbor joining tree illustrating the phylogenetic relationship of P2
strain.
With other type strains based on the 16S rRNA gene sequences. Bootstrap values
expressed in percentages of 1000 replications are represented at the nodes of the
branches.
Comamonas odontotermitis(DQ453128.1)
Comamonas odontotermitis strain Dant 3-8(NR_043859.1)
Comamonas odontotermitis P2(KX255005.1)
Comamonas odontotermitis strain RJ40(KP701358.1)
Comamonas sp. NT2B(GU458274.1)
Comamonas sp. HM1(GU458261.1 )
Comamonas sp. JL4(JF740040.1)
Comamonas terrigena strain NBRC 13299(NR_113613.1)
Comamonas nitrativorans strain 2331(NR_025376.1)
Comamonas granuli NBRC 101663(NZ_BBJX01000025.1)
Comamonas serinivorans strain SP-35(NR_134010.1)
Comamonas testosteroni strain RCB266(KT260478.1)
Comamonas sp. KBB4(JX997984.1)
Comamonas thiooxydans strain S23(NR_115741.1)
Comamonas testosteroni strain KS 0043(NR_029161.1)
Comamonas testosteroni strain NBRC 14951(NR_113709.1)
Comamonas thiooxydans strain PHE2-6(NZ_LKFB01000041.1)
Comamonas testosteroni(NZ_BBQP01000035.1)
Comamonas sp. E6(NZ_BBXH01000054.1)
Comamonas testosteroni(NZ_JWJW01000032.1)
Comamonas kerstersii(NZ_LFYP01000018.1)
Comamonas testosteroni strain WDL7(NZ_LMXT01000002.1)
Comamonas thiooxydans(NZ_LIOM01000035.1)
Comamonas thiooxydans(NZ_BBVD01000034.1)
Comamonas testosteroni NBRC 14951(NZ_BBJZ01000028.1)
98
75
58
53
55
99
79
85
100
58
82
56

4. Glyphosate Degrading Bacterial Strains
89
Figure 4-5 Phylogenetic analysis of Ochrobactrum anthropi P3 using Mega 6
based on 16S rRNA sequence analysis.
The tree suggests that P3 isolate clades with other Ochrobactrum spp on the basis of
maximum sequence identity.
Ochrobactrum sp.G2(KR052826.1)
Ochrobactrum sp.CCBAU(EF377300.1)
Ochrobactrum anthropi LMG 3331(NR_114979.1)
Ochrobactrum sp.PP-2(KX549468.1)
Ochrobactrum anthropi NBRC 15819(NR_113811.1)
Ochrobactrum anthropi SH23C(KT337526.1)
Ochrobactrum sp.DX2(KC462882.1)
Ochrobactrum sp.n-9(KF010632.1)
Ochrobactrum sp.X-16(EU187496.1)
Ochrobactrum anthropi S4(KT380595.1)
Ochrobactrum anthropi TCC-1(KT820194.1)
Ochrobactrum sp.DS4(KJ817211.1)
Ochrobactrum anthropi ATCC 49188(NR_074243.1)
Ochrobactrum anthropi P3(KX255006.1)
Ochrobactrum anthropi E46b(KM894190.1)
Ochrobactrum anthropi XH02(KU999381.1)
Ochrobactrum sp.A9(KF958486.1)
Ochrobactrum anthropi S180b(KM894186.1)
Ochrobactrum anthropi W-7(EU187487.1)
Ochrobactrum sp.LC498(JN863520.1)
Ochrobactrum anthropi R058(KC252888.1)

4. Glyphosate Degrading Bacterial Strains
90
Figure 4-6 Phylogenetic tree based on homologous sequences of the
Achromobacter spanius P4.
The neighbor joining methods employed through MEGA 6. Topology of dendrogram
supported by bootstrap values of 1000 iterations is represented as percentage values
when >50. Accession numbers are shown in parentheses.
Achromobacter spanius 2P2A1(HF936998.1)
Achromobacter spanius 2P1F8(HF936970.1)
Achromobacter sp.G3Dc4(KF465936.1)
Achromobacter sp.G3Dc11(KF465937.1)
Achromobacter sp.5_24(HF954413.1)
Achromobacter sp.LMG 3441(HG324052.1)
Achromobacter sp. EA_R_2(KJ642225.1)
Achromobacter spanius DUCC3703(KP318450.1)
Achromobacter xylosoxidans C2(KP967463.1)
Achromobacter sp.TY3-4(KP410738.1)
Achromobacter sp.255B4_12AESBL(KU644204.1)
Achromobacter sp.OMC7(KP728242.1)
Achromobacter spanius CCUG 47062(NR_118402.1)
Achromobacter spanius P4(KX255007.1)
Achromobacter spanius LMG 5911(NR_025686.1)
Achromobacter spanius Nr_39(KT714140.1)
Achromobacter spanius B1(KP860309.1)
Achromobacter marplatensis CCUG 56371(NR_118400.1)
Achromobacter xylosoxidans LMG 1863(NR_118403.1)
Achromobacter denitrificans CCUG 407(NR_118398.1)99
70
63

4. Glyphosate Degrading Bacterial Strains
91
Figure 4-7 Dendrogram exhibiting genetic relationships of P5 isolate.
With closely related Agrobacterium spp. based on 16SrRNA gene sequences
submitted in NCBI Genbank. P5 showed 99% similarity with Agrobacterium
tumefaciens A46.
4.3.3 DGGE Analysis of Enrichment Cultures
DGGE analysis of glyphosate enrichment cultures was carried out to monitor the
persistence of bacterial community under glyphosate stress. DGGE profile analysis
revealed the selection of limited number of bacterial strains through enrichment steps.
The cultures enriched with glyphosate in the first week had higher number of bands as
compared to the fingerprints from later weeks signifying a selection for glyphosate
tolerant microorganisms. Two groups of DGGE bands were observed in gel. First
group of bands indicated the organisms that were present in soil at initial enrichment
step but became very weak or disappeared by the final enrichment steps. Second
group of bands were present in fingerprints of all enrichment steps. Those bands
represented dominant bacterial strains in glyphosate treated soil capable to utilize
glyphosate as C and P sources (Fig 4-8a, 4-8b). Sequence analysis showed their
affiliation with Pseudomonas sp. (DG1), Comamonas sp. (DG2), uncultured
phyllobacterium sp. (DG3), mesorhizobium sp. (DG4), uncultured chryseobacterium
Agrobacterium tumefaciens P5(KX255008.1)
Agrobacterium tumefaciens A46(KC196477.1)
Agrobacterium sp.AN16(KR051019.1)
Agrobacterium tumefaciens NFM2(KP410821.1)
Agrobacterium tumefaciens(KT825747.1)
Agrobacterium tumefaciens LM-1(KM884891.1)
Agrobacterium tumefaciens AF114(LC015594.1)
Agrobacterium tumefaciens SQ3-38-1(KM252932.1)
Agrobacterium sp.SAUBS3-4(KC243285.1)
Agrobacterium tumefaciens A14(KC196473.1)
Agrobacterium sp.AN17(KR051020.1)
Agrobacterium tumefaciens DD260(KR822276.1)
Agrobacterium tumefaciens MLS-1-10(KT997434.1)
Agrobacterium tumefaciens DSR11(JQ342861.1)
Agrobacterium tumefaciens NFM10(KP410823.1)
Agrobacterium tumefaciens DSS5(JQ342845.1)
Agrobacterium tumefaciens AF47(LC015593.1)
Agrobacterium tumefaciens NSBm.28(CFS1)(JF708879.1)
Agrobacterium tumefaciens(KP875541.1)
Agrobacterium sp.(KR081269.1)
Agrobacterium tumefaciens LM6-1(KM884893.1)

4. Glyphosate Degrading Bacterial Strains
92
sp. (DG5), ochrobactrum sp. (DG6), Uncultured bacteriodetes bacterium (DG7),
uncultured bacterium (DG8), uncultured gamma proteobacterium (DG9) uncultured
flavobacterium sp. (DG10), uncultured brucella sp. (DG11), uncultured
bradyrhizobium sp. (DG12). As indicated by sequencing of bands obtained from
DGGE gel P2 strain was quite persistent among the replicates of enrichment cultures
till the end of enrichment experiment (Table 4.5).
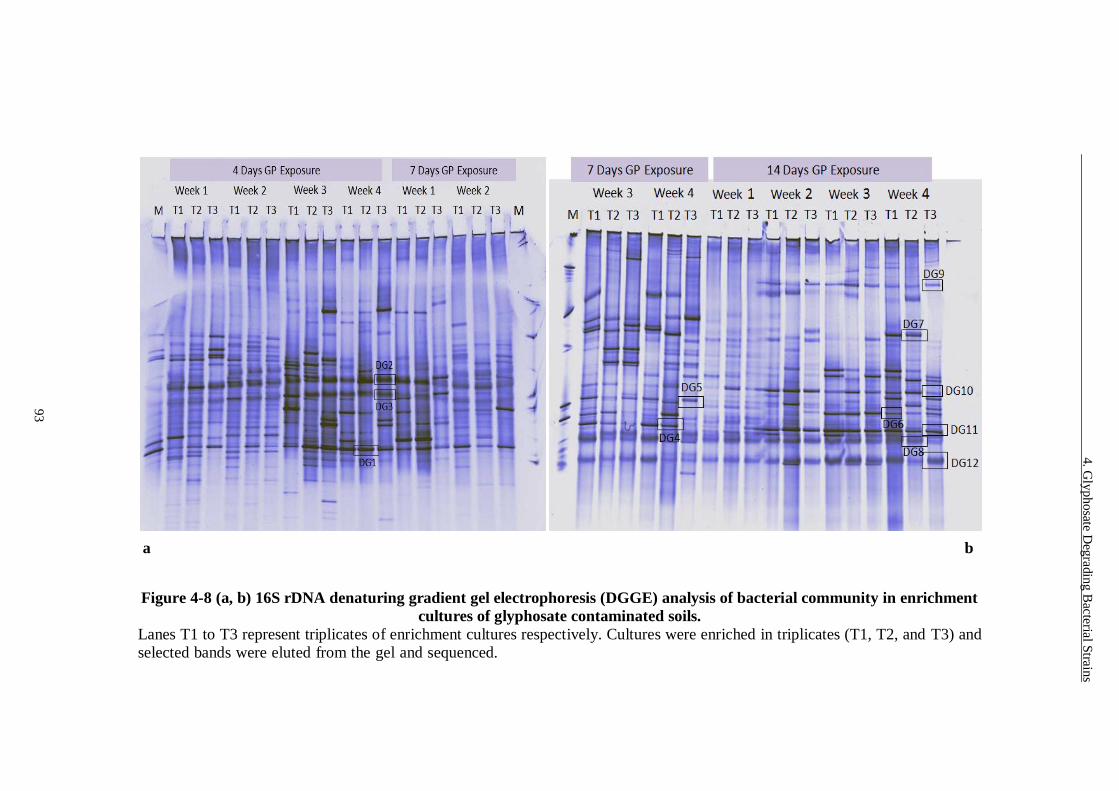
93
4. G
lypho
sate Deg
radin
g B
acterial Strain
s
Figure 4-8 (a, b) 16S rDNA denaturing gradient gel electrophoresis (DGGE) analysis of bacterial community in enrichment
cultures of glyphosate contaminated soils.
Lanes T1 to T3 represent triplicates of enrichment cultures respectively. Cultures were enriched in triplicates (T1, T2, and T3) and
selected bands were eluted from the gel and sequenced.jjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjj
a b
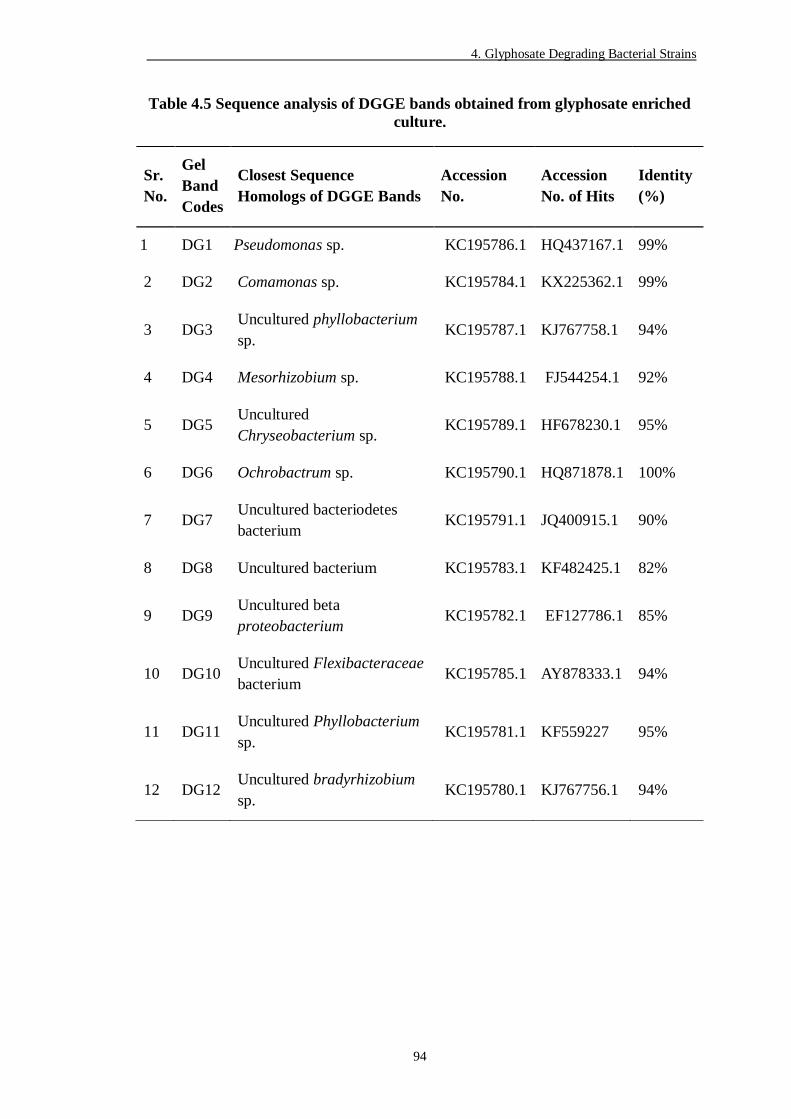
4. Glyphosate Degrading Bacterial Strains
94
Table 4.5 Sequence analysis of DGGE bands obtained from glyphosate enriched
culture.
Sr.
No.
Gel
Band
Codes
Closest Sequence
Homologs of DGGE Bands
Accession
No.
Accession
No. of Hits
Identity
(%)
1 DG1 Pseudomonas sp. KC195786.1 HQ437167.1 99%
2 DG2 Comamonas sp. KC195784.1 KX225362.1 99%
3 DG3 Uncultured phyllobacterium
sp. KC195787.1 KJ767758.1 94%
4 DG4 Mesorhizobium sp. KC195788.1 FJ544254.1 92%
5 DG5 Uncultured
Chryseobacterium sp. KC195789.1 HF678230.1 95%
6 DG6 Ochrobactrum sp. KC195790.1 HQ871878.1 100%
7 DG7 Uncultured bacteriodetes
bacterium KC195791.1 JQ400915.1 90%
8 DG8 Uncultured bacterium KC195783.1 KF482425.1 82%
9 DG9 Uncultured beta
proteobacterium KC195782.1 EF127786.1 85%
10 DG10 Uncultured Flexibacteraceae
bacterium KC195785.1 AY878333.1 94%
11 DG11 Uncultured Phyllobacterium
sp. KC195781.1 KF559227 95%
12 DG12 Uncultured bradyrhizobium
sp. KC195780.1 KJ767756.1 94%

4. Glyphosate Degrading Bacterial Strains
95
4.3.4 Optimization of Parameters for Glyphosate Degradation Using
RSM
Contrary to the conventional “single factor at a time experiments”, interaction and
concurrent effect of different factors; temperature, pH and initial inoculum size on
glyphosate degradation was studied using RSM. On the basis of central composite
design, 20 experimental setups were carried out consisting of 8 full factorial points, 6
central and 6 axial points located at the central and the extreme levels (Table 4.6).
Analysis of variance (ANOVA) values calculated by quadratic response surface
model are given in Table 4.7. The efficacy of model was demonstrated by
determination coefficient (R2
= 0.969) which corroborated that independent variables
ascribe 96.9% variability to glyphosate degradation. High value of the adjusted
determination coefficient (Adj. R2
= 0.9428) also showed the best fit of the model.
Furthermore high F-value (35.83) and low p-value showed that quadratic model was
significant. The non-significant value of lack of fit showed that degree of fitting is
fine. The p-value of individual variables exhibited that all factors significantly
influenced glyphosate degradation but effect of inoculum density was promising.

4. Glyphosate Degrading Bacterial Strains
96
Table 4.6 Predicted and experimental values of glyphosate degradation by CCD
matrix.
Standard
No.
Variables in coded levels Response
(glyphosate degradation %)
A B C Actual Predicted
1 -1 -1 -1 54.00 53.19
2 1 -1 -1 66.00 67.32
3 -1 1 -1 38.00 33.32
4 1 1 -1 68.00 65.94
5 -1 -1 1 88.00 89.83
6 1 -1 1 72.10 76.45
7 -1 1 1 72.00 70.45
8 1 1 1 75.00 75.58
9 -1.68 0 0 60.00 62.49
10 1.68 0 0 84.00 81.74
11 0 -1.68 0 76.00 71.81
12 0 1.68 0 50.00 54.52
13 0 0 -1.68 42.00 45.50
14 0 0 1.68 90.00 86.17
15 0 0 0 90.00 88.95
16 0 0 0 87.00 88.95
17 0 0 0 88.00 88.95
18 0 0 0 91.00 88.95
19 0 0 0 92.00 88.95
20 0 0 0 85.00 88.95
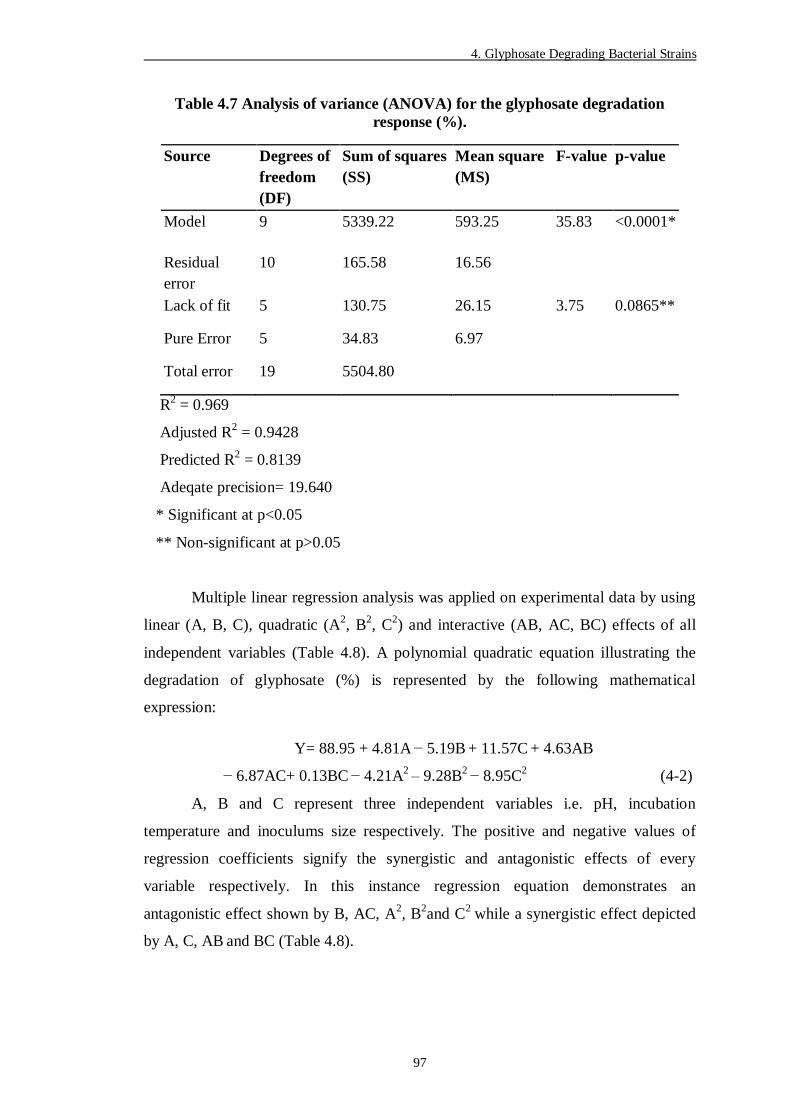
4. Glyphosate Degrading Bacterial Strains
97
Table 4.7 Analysis of variance (ANOVA) for the glyphosate degradation
response (%).
Source Degrees of
freedom
(DF)
Sum of squares
(SS)
Mean square
(MS)
F-value p-value
Model 9 5339.22 593.25 35.83 <0.0001*
Residual
error
10 165.58 16.56
Lack of fit 5 130.75 26.15 3.75 0.0865**
Pure Error 5 34.83 6.97
Total error 19 5504.80
R2 = 0.969
Adjusted R2 = 0.9428
Predicted R2 = 0.8139
Adeqate precision= 19.640
* Significant at p<0.05
** Non-significant at p>0.05
Multiple linear regression analysis was applied on experimental data by using
linear (A, B, C), quadratic (A2, B
2, C
2) and interactive (AB, AC, BC) effects of all
independent variables (Table 4.8). A polynomial quadratic equation illustrating the
degradation of glyphosate (%) is represented by the following mathematical
expression:
Y= 88.95 + 4.81A − 5.19B + 11.57C + 4.63AB
− 6.87AC+ 0.13BC − 4.21A2 – 9.28B
2 − 8.95C
2 (4-2)
A, B and C represent three independent variables i.e. pH, incubation
temperature and inoculums size respectively. The positive and negative values of
regression coefficients signify the synergistic and antagonistic effects of every
variable respectively. In this instance regression equation demonstrates an
antagonistic effect shown by B, AC, A2, B
2and C
2 while a synergistic effect depicted
by A, C, AB and BC (Table 4.8).
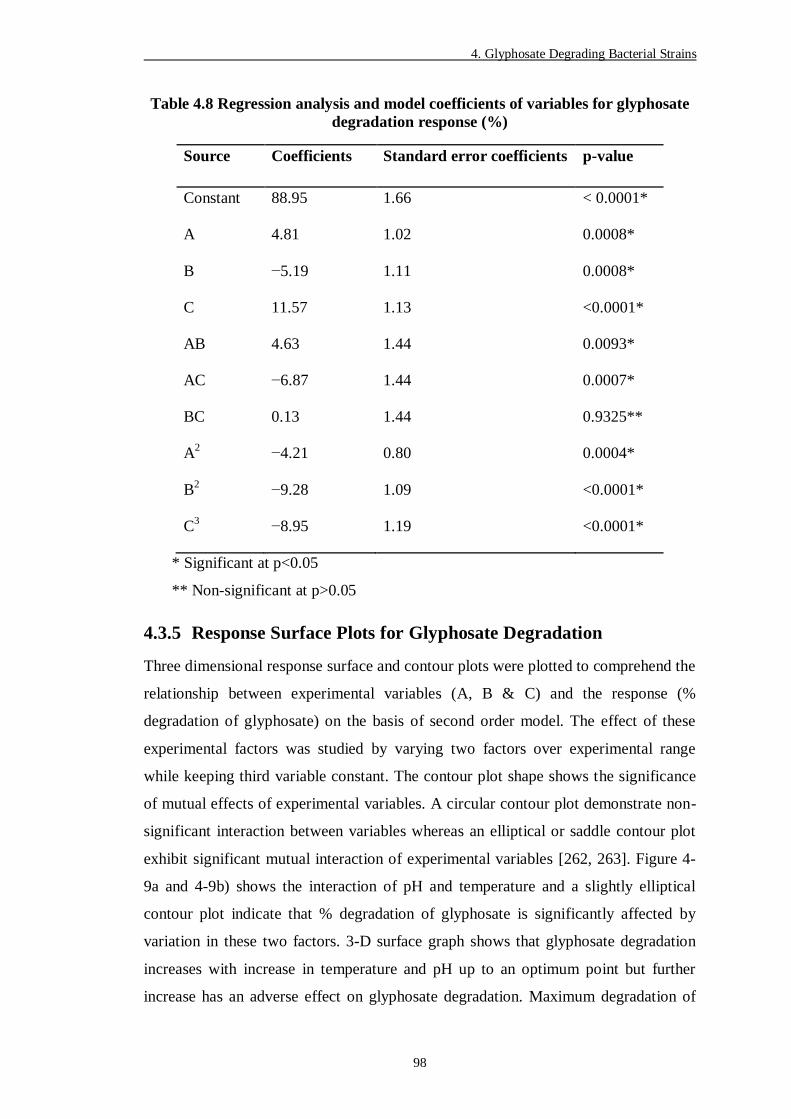
4. Glyphosate Degrading Bacterial Strains
98
Table 4.8 Regression analysis and model coefficients of variables for glyphosate
degradation response (%)
Source Coefficients Standard error coefficients p-value
Constant 88.95 1.66 < 0.0001*
A 4.81 1.02 0.0008*
B −5.19 1.11 0.0008*
C 11.57 1.13 <0.0001*
AB 4.63 1.44 0.0093*
AC −6.87 1.44 0.0007*
BC 0.13 1.44 0.9325**
A2 −4.21 0.80 0.0004*
B2 −9.28 1.09 <0.0001*
C3 −8.95 1.19 <0.0001*
* Significant at p<0.05
** Non-significant at p>0.05
4.3.5 Response Surface Plots for Glyphosate Degradation
Three dimensional response surface and contour plots were plotted to comprehend the
relationship between experimental variables (A, B & C) and the response (%
degradation of glyphosate) on the basis of second order model. The effect of these
experimental factors was studied by varying two factors over experimental range
while keeping third variable constant. The contour plot shape shows the significance
of mutual effects of experimental variables. A circular contour plot demonstrate non-
significant interaction between variables whereas an elliptical or saddle contour plot
exhibit significant mutual interaction of experimental variables [262, 263]. Figure 4-
9a and 4-9b) shows the interaction of pH and temperature and a slightly elliptical
contour plot indicate that % degradation of glyphosate is significantly affected by
variation in these two factors. 3-D surface graph shows that glyphosate degradation
increases with increase in temperature and pH up to an optimum point but further
increase has an adverse effect on glyphosate degradation. Maximum degradation of

4. Glyphosate Degrading Bacterial Strains
99
glyphosate was found at 31 °C. Figure 4-9c and 4-9d indicates that the interaction
between inoculum density and temperature is non-significant. Both high and low
values of inoculum size and temperature do not show effective glyphosate
degradation. An increase in temperature reduces glyphosate degradation. Moreover
glyphosate degradation increases with an increase in inoculum density and
temperature up to a certain limit. The elliptical contour plot shows that pH and
inoculum density significantly affect glyphosate degradation (Figure 4-9e and 4-9f).
Glyphosate degradation increases with increase in inoculum density up to an optimal
pH level.
The optimized % degradation of glyphosate predicted by software was found
to be 92.2% based on optimized values of variables i.e. temperature 29.9 °C, pH 7.4
and inoculum size 0.45 g/L after 3 days. Validity of the applied model was confirmed
by performing experiments with the optimized values predicted by RSM and 90%
degradation was attained. This experimental value is close to the predicted value of
glyphosate degradation (92.2%) signifying the sufficiency of the acquired model for
glyphosate degradation.
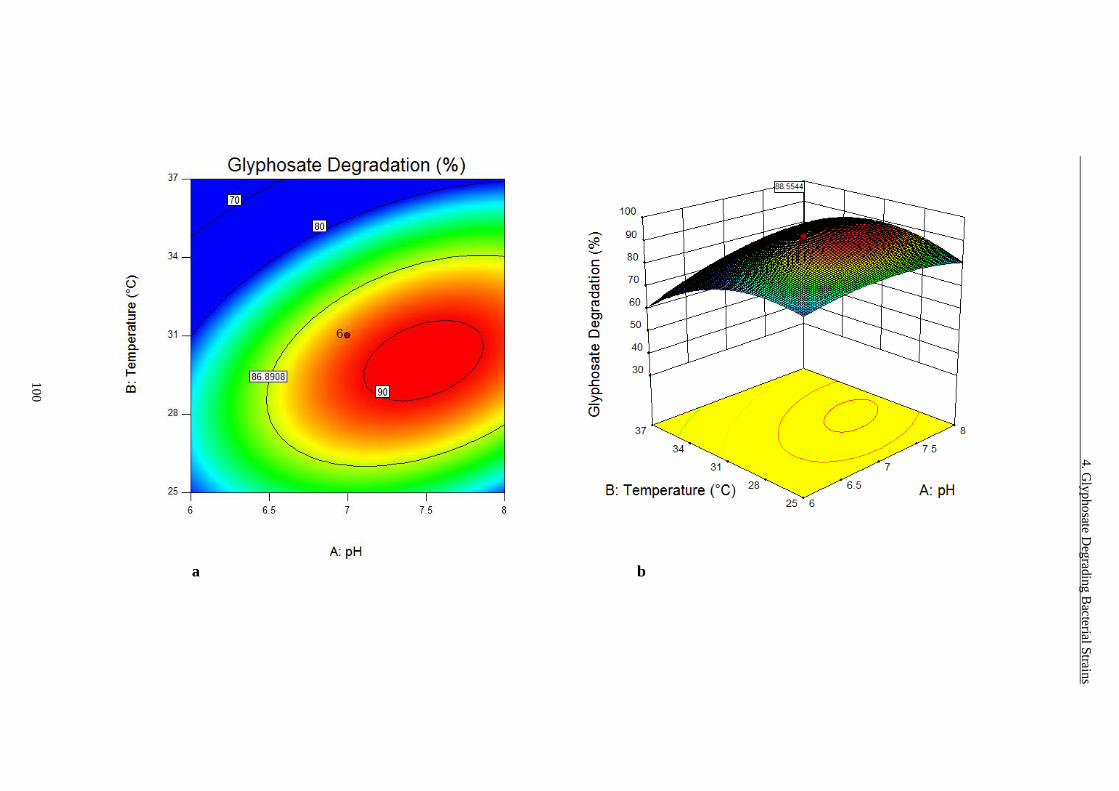
10
0
4. G
lypho
sate Deg
radin
g B
acterial Strain
s
a b
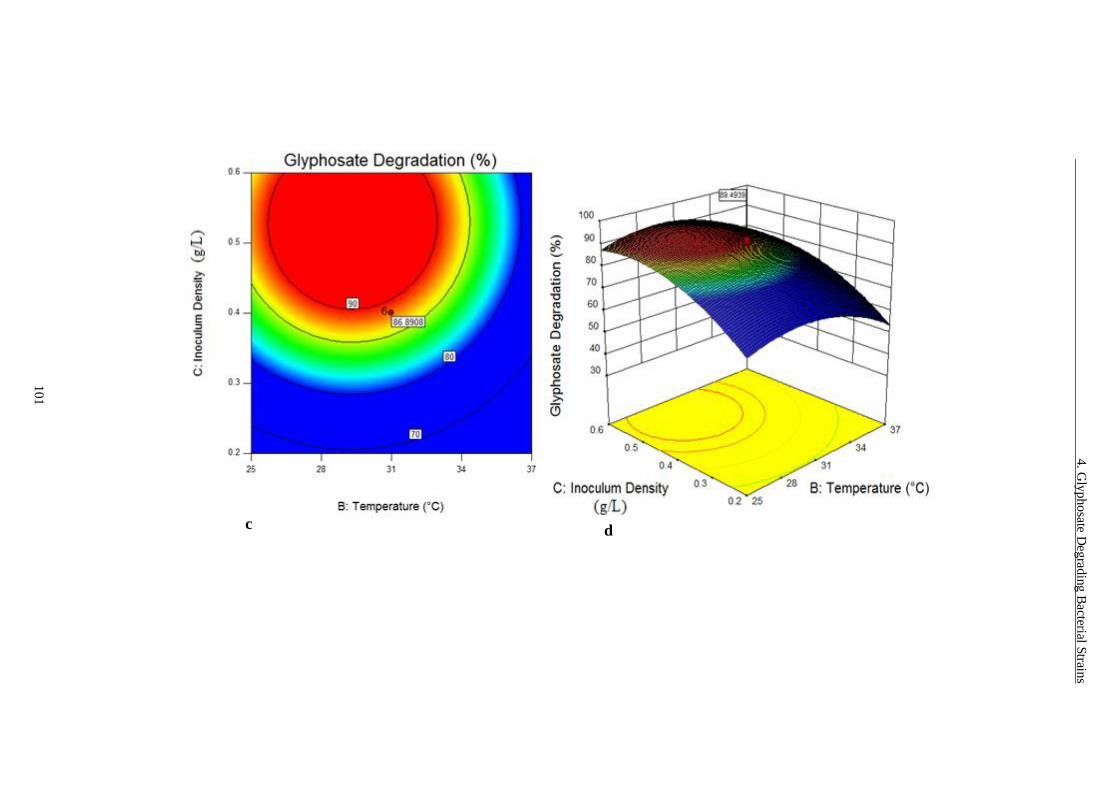
10
1
4. G
lyph
osate D
egrad
ing
Bacterial S
trains
c d
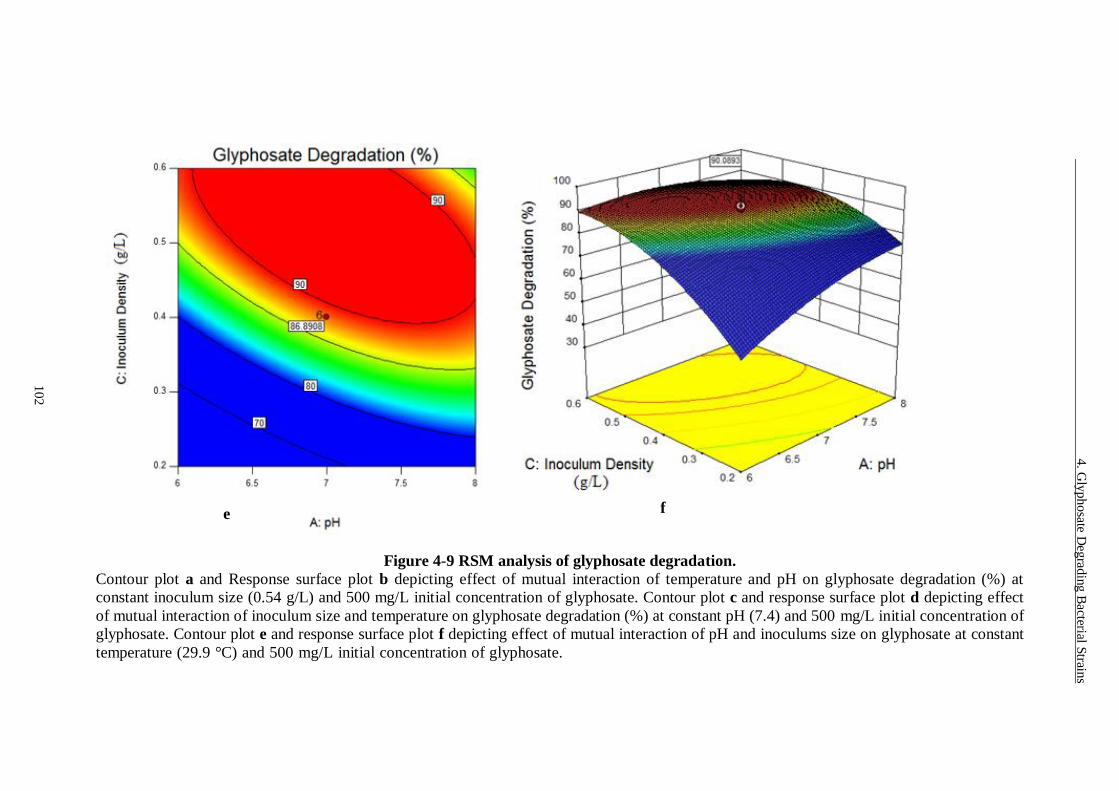
10
2
4. G
lyph
osate D
egrad
ing
Bacterial S
trains
Figure 4-9 RSM analysis of glyphosate degradation.
Contour plot a and Response surface plot b depicting effect of mutual interaction of temperature and pH on glyphosate degradation (%) at
constant inoculum size (0.54 g/L) and 500 mg/L initial concentration of glyphosate. Contour plot c and response surface plot d depicting effect
of mutual interaction of inoculum size and temperature on glyphosate degradation (%) at constant pH (7.4) and 500 mg/L initial concentration of
glyphosate. Contour plot e and response surface plot f depicting effect of mutual interaction of pH and inoculums size on glyphosate at constant
temperature (29.9 °C) and 500 mg/L initial concentration of glyphosate.jjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjjj
f e
e f
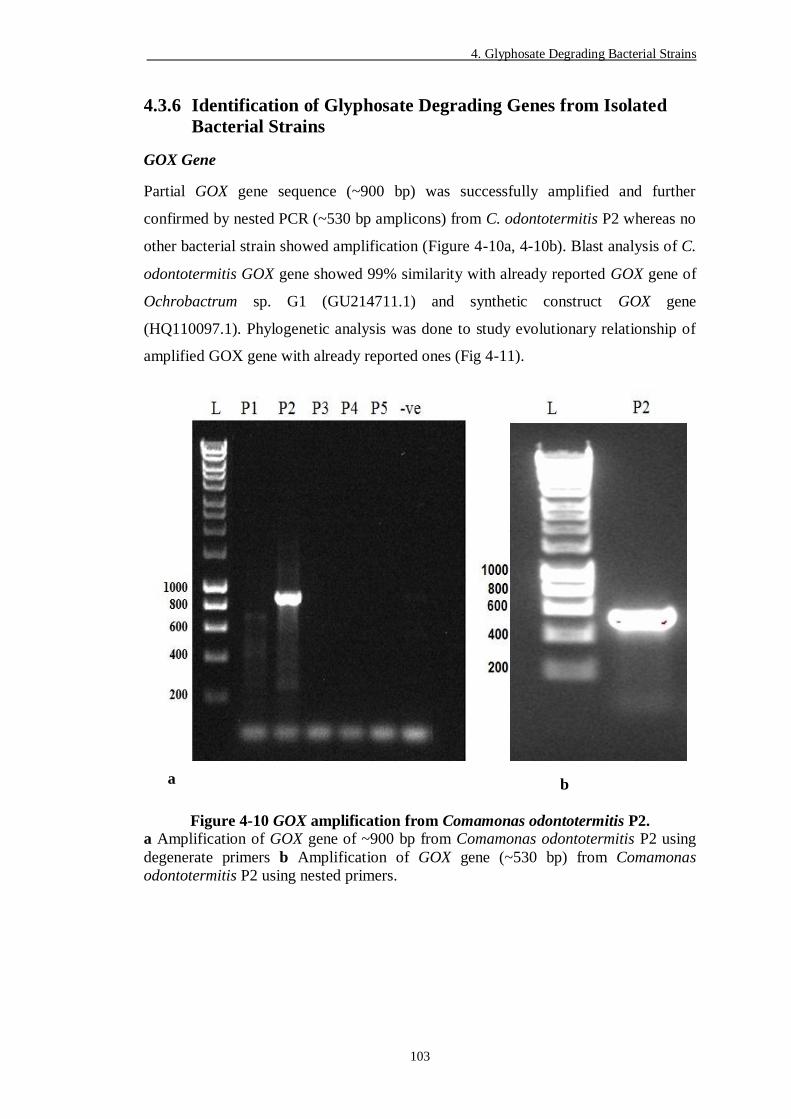
4. Glyphosate Degrading Bacterial Strains
103
4.3.6 Identification of Glyphosate Degrading Genes from Isolated
Bacterial Strains
GOX Gene
Partial GOX gene sequence (~900 bp) was successfully amplified and further
confirmed by nested PCR (~530 bp amplicons) from C. odontotermitis P2 whereas no
other bacterial strain showed amplification (Figure 4-10a, 4-10b). Blast analysis of C.
odontotermitis GOX gene showed 99% similarity with already reported GOX gene of
Ochrobactrum sp. G1 (GU214711.1) and synthetic construct GOX gene
(HQ110097.1). Phylogenetic analysis was done to study evolutionary relationship of
amplified GOX gene with already reported ones (Fig 4-11).
Figure 4-10 GOX amplification from Comamonas odontotermitis P2.
a Amplification of GOX gene of ~900 bp from Comamonas odontotermitis P2 using
degenerate primers b Amplification of GOX gene (~530 bp) from Comamonas
odontotermitis P2 using nested primers.
a b
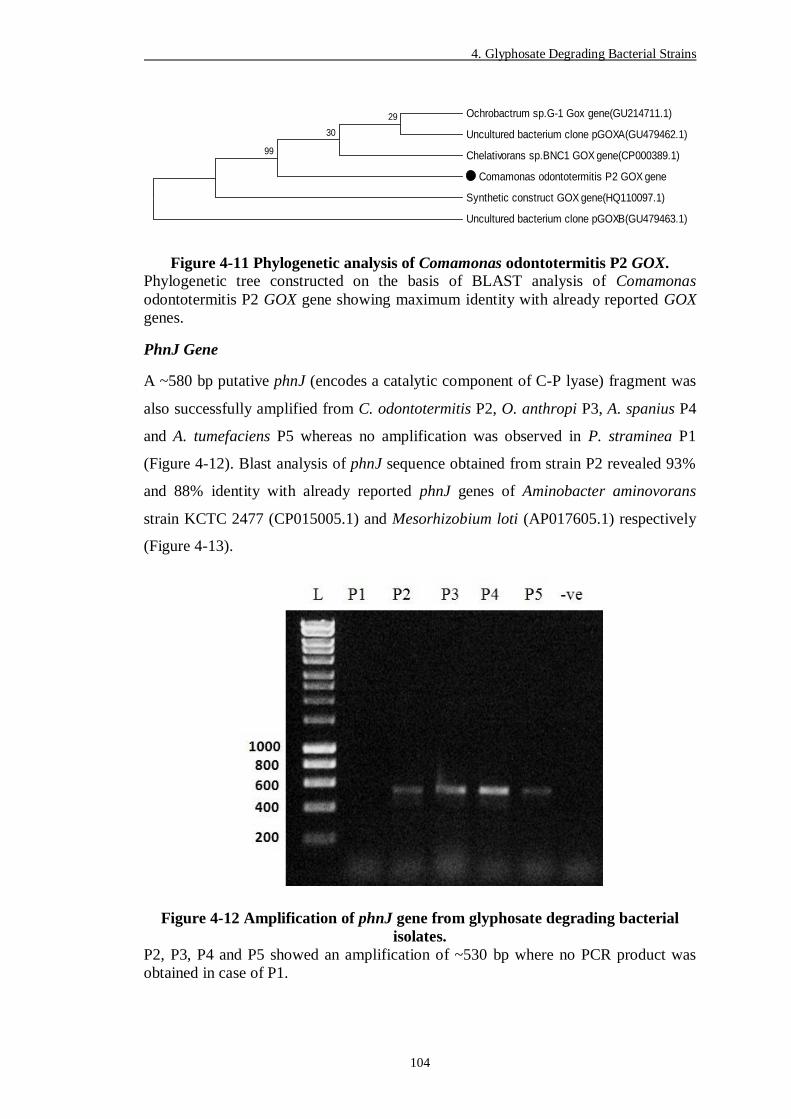
4. Glyphosate Degrading Bacterial Strains
104
Figure 4-11 Phylogenetic analysis of Comamonas odontotermitis P2 GOX.
Phylogenetic tree constructed on the basis of BLAST analysis of Comamonas
odontotermitis P2 GOX gene showing maximum identity with already reported GOX
genes.
PhnJ Gene
A ~580 bp putative phnJ (encodes a catalytic component of C-P lyase) fragment was
also successfully amplified from C. odontotermitis P2, O. anthropi P3, A. spanius P4
and A. tumefaciens P5 whereas no amplification was observed in P. straminea P1
(Figure 4-12). Blast analysis of phnJ sequence obtained from strain P2 revealed 93%
and 88% identity with already reported phnJ genes of Aminobacter aminovorans
strain KCTC 2477 (CP015005.1) and Mesorhizobium loti (AP017605.1) respectively
(Figure 4-13).
Figure 4-12 Amplification of phnJ gene from glyphosate degrading bacterial
isolates.
P2, P3, P4 and P5 showed an amplification of ~530 bp where no PCR product was
obtained in case of P1.
Ochrobactrum sp.G-1 Gox gene(GU214711.1)
Uncultured bacterium clone pGOXA(GU479462.1)
Chelativorans sp.BNC1 GOX gene(CP000389.1)
Comamonas odontotermitis P2 GOX gene
Synthetic construct GOX gene(HQ110097.1)
Uncultured bacterium clone pGOXB(GU479463.1)
99
29
30

4. Glyphosate Degrading Bacterial Strains
105
Figure 4-13 Blast analysis of phnJ gene amplified Comamonas odontotermitis P2.
Analysis revealed 90% identity with already reported phnJ genes. Phylogenetic tree
was constructed on the basis of PhnJ sequence identity.
4.4 Discussion
Herbicides have long been used for weed control in agriculture sector. Addition of
herbicides in soil is deterimental for soil biota thus their removal is essential to
prevent their accretion. Biodegradation is an eco-friendly and cost effective
technology to combat this problem. Generally biological ways of herbicide removal
are quite effective as compared to chemical ways especially in case of glyphosate
which has a strong C-P bond [264]. Biodegradation involves the conversion of
harmful substances into non toxic or less toxic substances. This methodology needs
the isolation of potential degrading microorganisms and their recruitment for
metabolizing toxic organic pollutants. Therefore bacterial strains acclimatized to high
concentrations of glyphosate were isolated and used for glyphosate biodegradation.
Different bacterial strains have been isolated and investigated for their glyphosate
degradation potential in this context [65]. Certain bacterial isolates have potential to
use glyphosate as C, P or N source [265]. Although certain Comamonas spp. has been
reported to degrade different recalcitrant pollutants [266-270] but there is no report
about C. odontotermitis as glyphosate degrader. C. odontotermitis P2 isolated in
current study has potential to utilize glyphosate as C and P source. Therefore this
finding is new addition to the list of glyphosate degrading bacteria that can be
exploited for decontaminating glyphosate polluted environment. Glyphosate
degradation potential of C. odontotermitis P2 may be attributed to its adaptation for
Sinorhizobium meliloti 2011 Phnj(CP004139.1)
Rhizobium meliloti phnj(M96263.1)
Ensifer adhaerens Casida A Phnj(CP015881.1)
Comamonas odontotermitis P2 Phnj gene
Mesorhizobium amorphae CCNWGS0123 Phnj(CP015318.1)
Mesorhizobium ciceri WSM1284 Phnj(CP015064.1)
Mesorhizobium huakuii 7653R Phnj(CP006581.1)
Mesorhizobium loti TONO Phnj(AP017605.1)
Aminobacter aminovorans KCTC 2477 Phnj(CP015005.1)
Mesorhizobium australicum WSM2073 Phnj(CP003358.1)
Agrobacterium sp. RAC06 Phnj(CP016499.1)
Sinorhizobium fredii HH103 Phnj(HE616899.1)
Sinorhizobium meliloti RMO17 Phnj(CP009146.1)
100
99
97
70
74
69
64
74
100
58

4. Glyphosate Degrading Bacterial Strains
106
glyphosate as energy source or genetic mutation leading to activation of enzymes
involved in degradation.
Bacterial cells require certain nutrients (C, N and P) for their growth and
metabolic activities and acclimatize themselves to the environment in case of
nutrients unavailability [121]. Therefore isolated bacteria were cultivated preliminary
in carbon and phosphate free media containing glyphosate to enhance their glyphosate
utilization efficiency. The adapted cells utilized glyphosate as carbon and phosphorus
source possibly due to the induction of glyphosate metabolic enzyme system. C.
odontotermitis P2 is found to utilize glyphosate as P source more efficiently rather
than as C source. Moneke et al. has also reported that Acetobacter sp. has better
ability to use glyphosate as P source than C source [271].
DGGE analysis revealed significant shifts in bacterial community structure
which may be attributed to the changed environment in laboratory conditions.
Bacterial strains showing glyphosate utilization under natural environmental
conditions might lose their activity due to change in required ratios of C, P or N.
Moreover, bands existing in the fingerprints of later enrichment cultures might be
consequence of constant selection pressure resulting in propagation of potential
glyphosate degrading bacteria. The bands present in initial enrichment steps but
disappeared in fingerprints of final enrichments steps may be due to their intolerance
to glyphosate, removal of bacteria that want to use soil particles as a solid substratum
to attach to or their replacement by bacteria with improved glyphosate degrading
abilities.
Certain growth parameters such as pH, inoculums size and temperature have
significant impact on biodegradation of organic pollutants [9, 272, 273].
Biodegradation process is regulated through variations in these parameters therefore
their optimization is crucial. Contrary to the conventional method involving single
factor at a time experiments, response surface methodology (RSM) was employed in
current study. Through RSM, more than one factor and their mutual interaction can be
studied simultaneously. The effect of three factors: temperature, pH and inoculums
density on glyphosate biodegradation was analyzed. Although C. odontotermitis P2
was found capable of degrading glyphosate at all experimental temperature, pH and
inoculums size but RSM provided set of conditions for maximum degradation. An

4. Glyphosate Degrading Bacterial Strains
107
increase in inoculum density enhances glyphosate degradation depending on pH and
temperature. Glyphosate degradation increased with increase in pH and inoculums
density which may be attributed to the activation of degrading enzymes at higher pH.
pH and temperature also affect glyphosate degradation at optimum inoculum density
however decreases at high temperature. RSM gave a set of optimized conditions to
achieve maximum glyphosate degradation (92.2%) based on optimized values of
variables i.e. temperature 29.9 °C, pH 7.4 and inoculum size 0.45 g/L within 3 days of
incubation.
C-P lyase encoding gene has been identified in Pseudomonas sp., Rhizobium
spp. and Streptomcyces sp. whereas GOX gene has been reported in Arthrobacter
atrocyaneus, Pseudomonas sp. and Ochrobactrum sp. [67]. Although C-P lyase
encoding genes have been identified in many bacterial strains but there are only few
reports on glyphosate oxidoreductase encoding GOX gene as found in Ochrobactrum
sp. G-1 (GenBank accession no. GU214711.1) and Chelativorans sp. BNC1
(CP000389.1) [116]. There is no data available in literature about GOX identification
in C. odontotermitis and a limited number of studies have addressed GOX activity by
which glyphosate is converted to AMPA and glyoxylic acid through C-N bond
cleavage within specific bacteria such as from industrial activated sludge that has
been exposed to glyphosate and byproducts of its production. The results of
glyphosate degradation showed that C. odontotermitis could utilize glyphosate as C
and P source. These findings are in agreement with previous reports which indicated
that C and P starvation results in glyphosate uptake as source of C and P leading to
the activation of glyphosate degradation pathways [121]. GOX involves cleavage of
C-N bond of glyphosate with formation of AMPA and provides available carbon to
bacteria whereas C-P lyase cleaves C-P bond of glyphosate and microorganisms can
therefore use it as alternative phosphorus source.
Current study shows the isolation and identification of C. odontotermitis P2 on
the basis its potential to metabolize glyphosate. This work has added a new species to
the list of already reported glyphosate degrading bacteria. Identification of potential
GOX and C-P lyase genes indicates the incidence of two glyphosate degradation
pathways in C. odontotermitis P2. This finding signifies that GOX and C-P lyase
pathways are potentially encoded by a single strain. Moreover, response surface
methodology (RSM) provides imperative information about optimization of various

4. Glyphosate Degrading Bacterial Strains
108
growth parameters and their mutual effects for maximum degradation of glyphosate.
DGGE analysis shows the dominance of strain C. odontotermitis P2 in enrichment
culture. Capability of C. odontotermitis P2 to degrade glyphosate under diverse
conditions demonstrates that it would be a good candidate for decontamination of
glyphosate affected environment.

109
5. Discussion
5.1 Aim of Thesis
The decisive venture of human beings has been growing crops to fulfill food and fiber
requirements since their existence. Currently more than 6 billion people uphold major
crops worldwide and any intimidation to agricultural sector has detrimental upshots.
Crops are mainly affected by pests and herbs (weeds) every year. Different strategies
are used for pest and weed management to augment productivity and survival of
crops. Mechanical methods such as removal of weeds with hands or some primal tools
e.g., hoe were used for controlling weeds in ancient times. Nowadays different
herbicides have been developed and employed for this purpose thus making
noteworthy contribution to enhanced agricultural productivity. However, excessive
production and utilization of such xenobiotic compounds have engendered mayhem in
natural environment through contaminating air, water and soil resulting in commotion
of biogeochemical cycle and different biota.
Glyphosate is an active ingredient of most widely used herbicides such as
Roundup. Nonselective nature of glyphosate makes this compound ominous even for
desired plants. It performs its function by hampering protein synthesis through
inhibition of aromatic amino acid pathway. It has become indispensable in agriculture
and silviculture particularly after the preamble of glyphosate resistant crops. Since
biotechnology proffer economically beneficial approaches in the areas of industry,
agriculture, health and environment. Different bacterial strains isolated from
contaminated areas and bearing novel genes and proteins have been employed for
industrial, agricultural and environmental applications. Glyphosate resistant crops are
genetically engineered by using glyphosate resistant genes from such bacterial isolates
(Reviewed in chapter 1). Presently substantial attention has been given to the
identification of bacterial strains conferring high resistance to glyphosate. Keeping in
view the current scenario, glyphosate resistant bacterial strains were isolated and
assessed for their tolerance potential in this study (Reviewed in chapter 3).

5. Discussion
110
Toxicity of glyphosate has been misapprehended since its discovery owing to
the absence of shikimate pathway in animals and human beings. Duke and Powles,
2008 reported that glyphosate has low toxicity as compared to aspirin or sodium
chloride [56]. However later studies invalidated this customary dogma and divulged
toxic effects of glyphosate on living organisms. Glyphosate has detrimental effects on
soil biota as well as on human beings. Various diseases have been found mounting in
agricultural communities exposed to glyphosate such as Alzheimer's disease,
Parkinson's disease, autism, AD, PD, anxiety disorder, inflammatory bowel disease,
cholestasis, renal lithiasis, anxiety syndrome, osteoporosis, infertility and thyroid
dysfunction [274]. Consequently excessive use of glyphosate is leading to the
contamination of ecosystem and its repercussions are under consideration by scientific
community these days. The endeavor of glyphosate decontamination can be achieved
by employing microorganisms through a cost effective and environmentally safe
technology called bioremediation.
Bioremediation technology involves exploitation of microorganisms to
diminish, remove or transform toxic xenobiotics found in soil or water to less toxic
compounds. Microorganisms can endure variations in environmental conditions
therefore their metabolic competence is very important. Bioremediation upholds the
intensification of particular indigenous microbiota capable of executing desired
activities at contaminated sites [275]. Such type of microbiota can be established by
certain ways involving growth promotion through addition of terminal electron
acceptors and nutrients, and by controlling growth parameters [276, 277].
Microorganisms utilize the toxic compounds as nutrient or energy sources in
bioremediation process [276, 278]. Therefore this aptitude of the microorganism has
been used for the welfare of humanity, flora and fauna and also for tapering the stress
of toxic xenobiotic compounds on ecosystem. Hence, different bacterial strains
capable of degrading glyphosate were isolated in current study (Reviewed in chapter
4).
The bacterial isolates were scrutinized for their degradation potential
regarding glyphosate. Different growth parameters were optimized to attain maximum
degradation of glyphosate. The genes reported to involve in glyphosate degradation
were identified from isolates in order to make an attempt for elucidation of
biodegradation pathway. The major rationale of this study was to attain methodical

5. Discussion
111
insight of glyphosate utilization by bacteria to facilitate bioremediation of
contaminated water and soil.
5.2 Major Findings
5.2.1 Ochrobactrum intermedium Sq20
Screening for glyphosate tolerant bacterial strains led to the isolation of bacteria from
different genera. Among all the isolates Ochrobactrum intermedium Sq20 was found
capable to tolerate >2 g/L of glyphosate. Moreover, its degradation potential was also
assessed and Ochrobactrum intermedium Sq20 showed complete degradation of
glyphosate at 500 mg/L initial concentration as sole carbon and energy source within
4 days. Glyphosate resistance gene, aroA gene (designated as aroAO.intermedium Sq20) was
amplified from this indigenous isolate Sq20. An open reading frame (ORF) of 1353
bp representing aroAO.intermedium Sq20 was amplified from Sq20 which showed 97%
homology with aroA genes from other Ochrobactrum spp. The sequence and
phylogenetic analysis revealed that aroAO.intermedium Sq20 encodes a polypeptide of 450
amino acids designated as EPSPSO.intermedium Sq20 and belongs to class II EPSPS
enzymes. The glyphosate resistant microbes identified hitherto include Agrobacterium
sp. strain CP4, Achromobacter sp. strain LBAA, Ochrobactrum anthropi and
Pseudomonas sp. strain PG2982. The EPSPS enzymes of these bacteria are classified
as class II EPSP synthases on the basis of their catalytic efficiency in the presence of
glyphosate and their substantial sequence variation [149]. Physicochemical
characterization demonstrated the validation and authenticity of 3-D model of
EPSPSO.intermedium Sq20 and was found significant and reliable for investigating the
mechanisms underlying the glyphosate tolerance. Furthermore, it was surmised that
EPSPSO.intermedium Sq20 has auspicious aptitude for production of glyphosate resistant
transgenic crops.
5.2.2 Comamonas odontotermitis P2
While investigating for glyphosate degrading bacterial strains, five bacterial strains
capable to utilize glyphosate as carbon and phosphorus source under different
environmental conditions were isolated. One novel bacterial strain unearthed as
Comamonas odontotermitis P2 through biochemical features and 16S rRNA sequence
analysis exhibited significant glyphosate degrading activity. The strain P2 was found
proficient to degrade 1.5 g/L glyphosate completely within 104 h. Consequently this

5. Discussion
112
strain can be potentially exploited for decontamination of glyphosate polluted areas.
Comamonas sp. has been reported as metal resistant bacteria able to degrade different
aromatic compounds [238]. Growth parameters of strain P2 were optimized through
response surface methodology (RSM) for glyphosate degradation. RSM has been
found useful for elucidation of growth parameters as it allows to study more than one
parameters at a time. In present study, three parameters were studied with changing
two parameters while keeping one constant resulting in polynomial equation helpful
in prediction of glyphosate degradation (%). Three dimensional graphical
representation of RSM helped to develop an immediate understanding of the
biodegradation trend. Likewise this methodology has revolutionized the earlier trend
of lengthy and tiresome one factor at a time approach. Therefore RSM is a time
saving approach supporting the investigation of mutual interaction between multiple
factors and response.
5.2.3 Glyphosate Degrading Genes
With the intention of investigating fate of glyphosate during biodegradation process,
an attempt was made to identify potential genes involved in glyphosate degradation.
Therefore, primers were designed on the basis of formerly reported glyphosate
degrading genes and amplification was carried out under respective conditions. Two
imperative potentially encoding genes, glyphosate oxidoreductase (GOX) and phnJ
(C-P lyase encoding gene) genes were successfully amplified from Comamonas
odontotermitis P2. Sequencing of GOX and phnJ from C. odontotermitis P2 revealed
99 and 93% identity to already reported bacterial GOX and phnJ genes respectively.
Identification of these two genes in strain P2 correlates the findings with cleavage of
C-N and C-P bonds of glyphosate indicating its potential to degrade glyphosate
through GOX and C-P lyase metabolic pathways. This potential of Comamonas
odontotermitis P2 can be employed for efficient degradation of glyphosate which can
be exploited for reduction in soil pollution.
5.3 Explicit Future Recommendations
1. Functional characterization of EPSPSO.intermedium Sq20 from Ochrobactrum
intermedium Sq20 will help in understanding its role at molecular level and also
plausible usage for production of glyphosate resistant transgenic crops. In
addition, the competence of Ochrobactrum intermedium Sq20 to survive and

5. Discussion
113
reproduce at high concentrations of the glyphosate in indigenous soil will help in
the remediation of glyphosate contaminated environments.
2. In situ remediation of glyphosate can be done by using Comamonas
odontotermitis P2 at glyphosate exposed sites. This strategy will help to improve
soil quality and diminish health risks related to glyphosate accumulation in fields.
This technology will also help to improve quality standards and boost agricultural
export through curtailing pesticide residues in export products.
3. The identification of metabolites produced during glyphosate degradation through
LCMS or MS/MS analysis will help in prediction of metabolic pathways.
Furthermore, detection of unknown metabolites may escort to the prediction of
new metabolic pathways.
4. The protein expression analysis of glyphosate degrading genes amplified from
bacterial isolates from induced and un-induced cultures will help in establishing
enhanced bioremediation.

114
6. References
[1] T. Leisinger, R. Hutter, and A. M. Cook, "Microbial degradation of
xenobiotics and recalcitrant compounds," FEMS Symp, 1981.
[2] K. P. Shukla, N. K. Singh, and S. Sharma, "Bioremediation: developments,
current practices and perspectives," Genet. Eng. Biotechnol. J., vol. 3, pp. 1-
20, 2010.
[3] W. Zhang, F. Jiang, and J. Ou, "Global pesticide consumption and pollution:
with China as a focus," Proc. Intl. Acad. Ecol. Environ. Sci. U.S.A., vol. 1, p.
125, 2011.
[4] D. Pimentel, "Environmental and economic costs of the application of
pesticides primarily in the United States," Environ. Dev. Sustainability, vol. 7,
pp. 229-252, 2005.
[5] C. A. Damalas, "Understanding benefits and risks of pesticide use," Sci. Res.
Essays,, vol. 4, pp. 945-949, 2009.
[6] M. Dua, A. Singh, N. Sethunathan, and A. Johri, "Biotechnology and
bioremediation: successes and limitations," Appl. Microbiol. Biotechnol., vol.
59, pp. 143-152, 2002.
[7] G. R. Chaudhry and S. Chapalamadugu, "Biodegradation of halogenated
organic compounds," Microbiol. Rev., vol. 55, pp. 59-79, 1991.
[8] D. A. Vaccari, P. F. Strom, and J. E. Alleman, Environmental Biology for
Engineers and Scientists: John Wiley & Sons, 2005.
[9] M. Diez, "Biological aspects involved in the degradation of organic
pollutants," J. Soil Sci.Plant Nutr., vol. 10, pp. 244-267, 2010.
[10] C. Colosio, M. Tiramani, G. Brambilla, A. Colombi, and A. Moretto,
"Neurobehavioural effects of pesticides with special focus on
organophosphorus compounds: which is the real size of the problem?,"
Neurotoxicology, vol. 30, pp. 1155-1161, 2009.
[11] M. Jokanovic and M. Prostran, "Pyridinium oximes as cholinesterase
reactivators. Structure-activity relationship and efficacy in the treatment of
poisoning with organophosphorus compounds," Curr. Med. Chem., vol. 16,
pp. 2177-2188, 2009.
[12] S. Yair, B. Ofer, E. Arik, S. Shai, R. Yossi, D. Tzvika,and K. Amir,
"Organophosphate degrading microorganisms and enzymes as biocatalysts in
environmental and personal decontamination applications," Crit. Rev.
Biotechnol., vol. 28, pp. 265-275, 2008.
[13] N. L. Wolfe, R. G. Zepp, and D. F. Paris, "Use of structure-reactivity
relationships to estimate hydrolytic persistence of carbamate pesticides,"
Water Res., vol. 12, pp. 561-563, 1978.

6. References
115
[14] B. Eskenazi, A. Bradman, and R. Castorina, "Exposures of children to
organophosphate pesticides and their potential adverse health effects,"
Environ. Health Perspect., vol. 107, p. 409, 1999.
[15] N. G. Ternan, J. W. Mc Grath, G. Mc Mullan, and J. P. Quinn, "Review:
organophosphonates: occurrence, synthesis and biodegradation by
microorganisms," World J. Microbiol. Biotechnol., vol. 14, pp. 635-647, 1998.
[16] E. G. Jaworski, "Mode of action of N-phosphonomethylglycine. Inhibition of
aromatic amino acid biosynthsis," J. Agric. Food Chem., vol. 20, pp. 1195-
1198, 1972.
[17] R. L. Hilderbrand and T. Henderson, "The Effects of Phosphonates on Living
Systems," The Role of Phosphonates in Living Systems.ed Hilderbrand, R. L.
Boca Raton, Florida, USA. C.R.C. Press, Inc., pp. 139-170, 1983.
[18] T. Egli, "(An) aerobic breakdown of chelating agents used in household
detergents," Microbiol. Sci., vol. 5, pp. 36-41, 1988.
[19] R. J. Tyhach, R. Engel, and B. Tropp, "Metabolic fate of 3, 4-dihydroxybutyl-
1-phosphonate in Escherichia coli," J. Biol. Chem., vol. 251, pp. 6717-6723,
1976.
[20] H. Fleisch, "Bisphosphonates," Drugs, vol. 42, pp. 919-944, 1991.
[21] A. Lacoste, C. Dumora, and A. Cassaigne, "Cleavage of the carbon to
phosphorus bond of organophosphonates by bacterial systems," Biochemistry
(Life Sci. Adv.), vol. 8, pp. 97-111, 1989.
[22] M. Horiguchi and M. Kandatstu, "Isolation of 2-aminoethane phosphonic acid
from rumen protozoa," Nature, pp. 97-111, 1959.
[23] B. L. Wanner and W. W. Metcalf, "Molecular genetic studies of a 10.9-kb
operon in Escherichia coli for phosphonate uptake and biodegradation,"
FEMS Microbiol. Lett., vol. 100, pp. 133-139, 1992.
[24] S. B. Powles and Q. Yu, "Evolution in action: plants resistant to herbicides,"
Annu. Rev. Plant Biol., vol. 61, pp. 317-347, 2010.
[25] S. Vats, "Herbicides: History, Classification and Genetic Manipulation of
Plants for Herbicide Resistance,"Sustain. Agr. Rev., pp. 153-192, 2015.
[26] D. L. Shaner, "Herbicide safety relative to common targets in plants and
mammals," Pest Manag. Sci., vol. 60, pp. 17-24, 2004.
[27] H. Ohkawa, H. Miyagawa, and P. W. Lee, Pesticide Chemistry: Crop
Protection, Public Health, Environmental Safety, Wiley-VCH, Weinheim,
Germany, pp. 29-42, 2007.
[28] R. Schmidt, "HRAC classification of herbicides according to mode of action,"
in 1997 Brighton crop protection conference: weeds. Proceedings of an
international conference, Brighton, UK, 1997, pp. 1133-1140.
[29] C. A. Mallory-Smith and E. J. Retzinger Jr, "Revised Classification of
Herbicides by Site of Action for Weed Resistance Management Strategies 1,"
Weed Technol., vol. 17, pp. 605-619, 2003.
[30] W. K. Vencill, Herbicide Handbook, 8th edition. Weed Science Society of
America, 2002.

6. References
116
[31] T. Konishi and Y. Sasaki, "Compartmentalization of two forms of acetyl-CoA
carboxylase in plants and the origin of their tolerance toward herbicides,"
Proc. Natl. Acad. Sci. U.S.A., vol. 91, pp. 3598-3601, 1994.
[32] T. Nikolskaya, O. Zagnitko, G. Tevzadze, R. Haselkorn, and P. Gornicki,
"Herbicide sensitivity determinant of wheat plastid acetyl-CoA carboxylase is
located in a 400-amino acid fragment of the carboxyltransferase domain,"
Proc. Natl. Acad. Sci. U.S.A., vol. 96, pp. 14647-14651, 1999.
[33] X. Yang, I. A. Guschina, S. Hurst, S. Wood, M. Langford, T. Hawkes, and J.
L. Harwood, "The action of herbicides on fatty acid biosynthesis and
elongation in barley and cucumber," Pest Manag. Sci., vol. 66, pp. 794-800,
2010.
[34] O. Zagnitko, J. Jelenska, G. Tevzadze, R. Haselkorn, and P. Gornicki, "An
isoleucine/leucine residue in the carboxyltransferase domain of acetyl-CoA
carboxylase is critical for interaction with aryloxyphenoxypropionate and
cyclohexanedione inhibitors," Proc. Natl. Acad. Sci. U.S.A., vol. 98, pp. 6617-
6622, 2001.
[35] R. G. Duggleby and S. S. Pang, "Acetohydroxyacid synthase," J. Biochem.
Mol. Biol., vol. 33, pp. 1-36, 2000.
[36] C. E. Whitcomb, "An introduction to ALS-inhibiting herbicides," Toxicol. Ind.
Health, vol. 15, pp. 232-240, 1999.
[37] D. Wloga and J. Gaertig, "Post-translational modifications of microtubules," J.
Cell Sci., vol. 123, pp. 3447-3455, 2010.
[38] K. Grossmann, "Auxin herbicides: current status of mechanism and mode of
action," Pest Manag. Sci., vol. 66, pp. 113-120, 2010.
[39] N. P. Mira, M. C. Teixeira, and I. Sá-Correia, "Adaptive response and
tolerance to weak acids in Saccharomyces cerevisiae: a genome-wide view,"
OMICS: J. Integrative Biol., vol. 14, pp. 525-540, 2010.
[40] M. D Lambreva, D. Russo, F. Polticelli, V. Scognamiglio, A. Antonacci, V.
Zobnina, G. Campi, and G. Rea, "Structure/function/dynamics of photosystem
II plastoquinone binding sites," Curr. Protein Pept. Sci., vol. 15, pp. 285-295,
2014.
[41] T. Roach and A. Krieger-Liszkay, "Regulation of photosynthetic electron
transport and photoinhibition," Curr. Protein Pept. Sci., vol. 15, pp. 351-362,
2014.
[42] S. Santabarbara, "Limited sensitivity of pigment photo-oxidation in isolated
thylakoids to singlet excited state quenching in photosystem II antenna," Arch.
Biochem. Biophys., vol. 455, pp. 77-88, 2006.
[43] M. B. Colovic, D. Z. Krstic, T. D. Lazarevic-Pasti, A. M. Bondzic, and V. M.
Vasic, "Acetylcholinesterase inhibitors: pharmacology and toxicology," Curr.
Neuropharmacol., vol. 11, pp. 315-335, 2013.
[44] Y.-M. Qin, C.-Y. Hu, Y. Pang, A. J. Kastaniotis, J. K. Hiltunen, and Y.-X.
Zhu, "Saturated very-long-chain fatty acids promote cotton fiber and
Arabidopsis cell elongation by activating ethylene biosynthesis," Plant Cell,
vol. 19, pp. 3692-3704, 2007.

6. References
117
[45] H. Maeda and N. Dudareva, "The shikimate pathway and aromatic amino acid
biosynthesis in plants," Annu. Rev. Plant Biol., vol. 63, pp. 73-105, 2012.
[46] W. W. Metcalf and W. A. van der Donk, "Biosynthesis of phosphonic and
phosphinic acid natural products," Annu. Rev. Biochem., vol. 78, p. 65, 2009.
[47] J. J. van Rensen and W. J. Vredenberg, "Adaptation of photosystem II to high
and low light in wild-type and triazine-resistant Canola plants: analysis by a
fluorescence induction algorithm," Photosynth. Res., vol. 108, pp. 191-200,
2011.
[48] G. Qin, H. Gu, L. Ma, Y. Peng, X. W. Deng, Z. Chen, L. J. Qu, "Disruption of
phytoene desaturase gene results in albino and dwarf phenotypes in
Arabidopsis by impairing chlorophyll, carotenoid, and gibberellin
biosynthesis," Cell Res., vol. 17, pp. 471-482, 2007.
[49] C. D. Nwani, W. S. Lakra, N. S. Nagpure, R. Kumar, B. Kushwaha, and S. K.
Srivastava, "Toxicity of the herbicide atrazine: effects on lipid peroxidation
and activities of antioxidant enzymes in the freshwater fish Channa punctatus
(Bloch)," Int. J. Env. Res. Public Health, vol. 7, pp. 3298-3312, 2010.
[50] K. Oracz, H. El-Maarouf-Bouteau, I. Kranner, R. Bogatek, F. Corbineau, and
C. Bailly, "The mechanisms involved in seed dormancy alleviation by
hydrogen cyanide unravel the role of reactive oxygen species as key factors of
cellular signaling during germination," Plant Physiol., vol. 150, pp. 494-505,
2009.
[51] B. Walpola, S. Wanniarachchi, and J. Liyanage, "Responses of the soil
microbial biomass carbon to the herbicides propanil and glyphosate," J. Agric.
Sci, vol. 3, pp. 122-130, 2007.
[52] J. Lipok, H. Studnik, and S. Gruyaert, "The toxicity of Roundup® 360 SL
formulation and its main constituents: glyphosate and isopropylamine towards
non-target water photoautotrophs," Ecotoxicol. Environ. Saf., vol. 73, pp.
1681-1688, 2010.
[53] J. E. Franz, M. K. Mao, and J. A. Sikorski, Glyphosate's Molecular Mode of
Action in Glyphosate: A Unique Global Herbicide: American Chemical
Society, Washington, DC, pp. 521-642, 1997.
[54] Monsanto, "Backgrounder. History of Monsanto‟s Glyphosate Herbicides,"
Internet: www.monsanto.com/products/Documents/glyphosate-background-
materials/back_history.pdf. [Apr 15, 2014].
[55] S. B. Powles and C. Preston, "Evolved glyphosate resistance in plants:
biochemical and genetic basis of resistance 1," Weed Technol., vol. 20, pp.
282-289, 2006.
[56] S. O. Duke and S. B. Powles, "Glyphosate: a once‐in‐a‐century herbicide,"
Pest Manag. Sci., vol. 64, pp. 319-325, 2008.
[57] L. Artim, L. Schouten, B. V. Berg, and N. Carozzi, "Athenix." Internet:
http://www.fda.gov/downloads/Food/FoodScienceResearch/Biotechnology/Su
bmissions/UCM233624.pdf. [June 29, 2013].
[58] E. Schönbrunn, S. Eschenburg, W. A. Shuttleworth, J. V. Schloss, N.
Amrhein, J. N. Evans, and W. Kabsch, "Interaction of the herbicide glyphosate

6. References
118
with its target enzyme 5-enolpyruvylshikimate 3-phosphate synthase in atomic
detail," Proc. Natl. Acad. Sci. U.S.A., vol. 98, pp. 1376-1380, 2001.
[59] B. J. Vande Berg, P. E. Hammer, B. L. Chun, L. C. Schouten, B. Carr, R. Guo,
C. Peters, T. K. Hinson, V. Beilinson, A. Shekita, and R. Deter,
"Characterization and plant expression of a glyphosate‐tolerant
enolpyruvylshikimate phosphate synthase," Pest Manag. Sci., vol. 64, pp. 340-
345, 2008.
[60] J. A. Sikorski and K. J. Gruys, "Understanding glyphosate's molecular mode
of action with EPSP synthase: evidence favoring an allosteric inhibitor
model," Acc. Chem. Res., vol. 30, pp. 2-8, 1997.
[61] R. C. Kirkwood, R. Hetherington, T. L. Reynolds, and G. Marshall,
"Absorption, localisation, translocation and activity of glyphosate in
barnyardgrass (Echinochloa crus‐galli (L) Beauv): influence of herbicide and
surfactant concentration," Pest Manag. Sci., vol. 56, pp. 359-367, 2000.
[62] A. Gimsing, O. Borggaard, and M. Bang, "Influence of soil composition on
adsorption of glyphosate and phosphate by contrasting Danish surface soils,"
Eur. J. Soil Sci., vol. 55, pp. 183-191, 2004.
[63] P. Laitinen, K. Siimes, L. Eronen, S. Rämö, L. Welling, S. Oinonen, L.
Mattsoff, and M. Ruohonen‐Lehto, "Fate of the herbicides glyphosate,
glufosinate‐ammonium, phenmedipham, ethofumesate and metamitron in two
Finnish arable soils," Pest Manag. Sci., vol. 62, pp. 473-491, 2006.
[64] H. de Jonge, L. W. de Jonge, O. H. Jacobsen, T. Yamaguchi, and P. Moldrup,
"Glyphosate sorption in soils of different pH and phosphorus content," Soil
Sci., vol. 166, pp. 230-238, 2001.
[65] O. K. Borggaard and A. L. Gimsing, "Fate of glyphosate in soil and the
possibility of leaching to ground and surface waters: a review," Pest Manag.
Sci., vol. 64, pp. 441-456, 2008.
[66] R. E. Dick and J. P. Quinn, "Control of glyphosate uptake and metabolism in
Pseudomonas sp. 4ASW," FEMS Microbiol. Lett., vol. 134, pp. 177-182,
1995.
[67] S. O. Duke, "Glyphosate degradation in glyphosate-resistant and-susceptible
crops and weeds," J. Agric. Food Chem., vol. 59, pp. 5835-5841, 2010.
[68] A. L. Gimsing and O. K. Borggaard, "Phosphate and glyphosate adsorption by
hematite and ferrihydrite and comparison with other variable-charge
minerals," Clays Clay Miner., vol. 55, pp. 108-114, 2007.
[69] J. S. McConnell and L. R. Hossner, "pH-dependent adsorption isotherms of
glyphosate," J. Agric. Food Chem., vol. 33, pp. 1075-1078, 1985.
[70] J. P. Giesy, S. Dobson, and K. R. Solomon, "Ecotoxicological risk assessment
for Roundup® herbicide," in Rev. Environ. Contam. Toxicol., pp. 35-120,
2000.
[71] W. Edwards, G. Triplett, and R. Kramer, "A watershed study of glyphosate
transport in runoff," J. Environ. Qual., vol. 9, pp. 661-665, 1980.
[72] H. Vereecken, "Mobility and leaching of glyphosate: a review," Pest Manag.
Sci., vol. 61, pp. 1139-1151, 2005.

6. References
119
[73] L. Torstensson, E. Börjesson, and J. Stenström, "Efficacy and fate of
glyphosate on Swedish railway embankments," Pest Manag. Sci., vol. 61, pp.
881-886, 2005.
[74] J. Kjær, P. Olsen, M. Ullum, and R. Grant, "Leaching of glyphosate and
amino-methylphosphonic acid from Danish agricultural field sites," J.
Environ. Qual., vol. 34, pp. 608-620, 2005.
[75] L. Candela, J. Caballero, and D. Ronen, "Glyphosate transport through
weathered granite soils under irrigated and non-irrigated conditions-Barcelona,
Spain," Sci. Total Environ., vol. 408, pp. 2509-2516, 2010.
[76] R. Lajmanovich, M. Sandoval, and P. Peltzer, "Induction of mortality and
malformation in Scinax nasicus tadpoles exposed to glyphosate formulations,"
Bull. Environ. Contam. Toxicol., vol. 70, pp. 0612-0618, 2003.
[77] S. Bazot and T. Lebeau, "Simultaneous mineralization of glyphosate and
diuron by a consortium of three bacteria as free-and/or immobilized-cells
formulations," Appl. Microbiol. Biotechnol., vol. 77, pp. 1351-1358, 2008.
[78] J. Kjær, A. E. Rosenbom, W. Brüsch, R. K. Juhler, L. Gudmundsson, F.
Plauborg, R. Grant, and P. Olsen, "The Danish Pesticide Leaching Assessment
Programme: Monitoring Results May 1999-June 2010," Geological Survey of
Denmark and Greenland, Copenhagen, 2011.
[79] W. A. Battaglin, D. W. Kolpin, E. A. Scribner, K. M. Kuivila, and M. W.
Sandstrom, "Glyphosate, its transformation product AMPA, and other
herbicides in midwestern streams, 2002," J. Am. Water. Resour. As., vol. 41,
pp. 323-332, 2005.
[80] M. J. Shipitalo and L. B. Owens, "Comparative losses of glyphosate and
selected residual herbicides in surface runoff from conservation-tilled
watersheds planted with corn or soybean," J. Environ. Qual., vol. 40, pp.
1281-1289, 2011.
[81] E. A. Scribner, W. A. Battaglin, R. J. Gilliom, and M. T. Meyer,
"Concentrations of glyphosate, its degradation product,
aminomethylphosphonic acid, and glufosinate in ground- and surface-water,
rainfall, and soil samples collected in the United States, 2001-06," US
Geological Survey Scientific Investigations Report 2007-5122, p.111, 2007.
[82] C. M. Benbrook, "Trends in glyphosate herbicide use in the United States and
globally," Env. Sci. Eur., vol. 28, p. 1, 2016.
[83] S. B. Powles, "Evolved glyphosate‐resistant weeds around the world: lessons
to be learnt," Pest Manag. Sci., vol. 64, pp. 360-365, 2008.
[84] J. Kjær, V. Ernstsen, O. H. Jacobsen, N. Hansen, L. W. de Jonge, and P.
Olsen, "Transport modes and pathways of the strongly sorbing pesticides
glyphosate and pendimethalin through structured drained soils," Chemosphere,
vol. 84, pp. 471-479, 2011.
[85] R. H. Coupe, S. J. Kalkhoff, P. D. Capel, and C. Gregoire, "Fate and transport
of glyphosate and aminomethylphosphonic acid in surface waters of
agricultural basins," Pest Manag. Sci., vol. 68, pp. 16-30, 2012.

6. References
120
[86] A. A. Shehata, W. Schrödl, A. A. Aldin, H. M. Hafez, and M. Krüger, "The
effect of glyphosate on potential pathogens and beneficial members of poultry
microbiota in vitro," Curr. Microbiol., vol. 66, pp. 350-358, 2013.
[87] A. D. Baylis, "Why glyphosate is a global herbicide: strengths, weaknesses
and prospects," Pest Manag. Sci., vol. 56, pp. 299-308, 2000.
[88] S. Richard, S. Moslemi, H. Sipahutar, N. Benachour, and G.-E. Seralini,
"Differential effects of glyphosate and roundup on human placental cells and
aromatase," Environ. Health Perspect., pp. 716-720, 2005.
[89] D. A. Savitz, T. Arbuckle, D. Kaczor, and K. M. Curtis, "Male pesticide
exposure and pregnancy outcome," Am. J. Epidemiol., vol. 146, pp. 1025-
1036, 1997.
[90] M. Watts, "Glyphosate: Addendum 2012. Pesticide Action Network Asia and
the Pacific 2012." Internet:
http://library.ipamglobal.org/jspui/handle/ipamlibrary/789. [Oct 12, 2014].
[91] A. Martínez, I. Reyes, and N. Reyes, "Cytotoxicity of the herbicide glyphosate
in human peripheral blood mononuclear cells," Biomedica, vol. 27, pp. 594-
604, 2007.
[92] N. Benachour and G.-E. Séralini, "Glyphosate formulations induce apoptosis
and necrosis in human umbilical, embryonic, and placental cells," Chem. Res.
Toxicol., vol. 22, pp. 97-105, 2008.
[93] C. Gasnier, C. Dumont, N. Benachour, E. Clair, M.-C. Chagnon, and G. E.
Séralini, "Glyphosate-based herbicides are toxic and endocrine disruptors in
human cell lines," Toxicology, vol. 262, pp. 184-191, 2009.
[94] G. L. Kimmel, C. A. Kimmel, A. L. Williams, and J. M. DeSesso, "Evaluation
of developmental toxicity studies of glyphosate with attention to
cardiovascular development," Crit. Rev. Toxicol., vol. 43, pp. 79-95, 2013.
[95] L. D. Kier and D. J. Kirkland, "Review of genotoxicity studies of glyphosate
and glyphosate-based formulations," Crit. Rev. Toxicol., vol. 43, pp. 283-315,
2013.
[96] M. Kruger, W. Schrodl, J. Neuhaus, and A. A. Shehata, "Field investigations
of glyphosate in urine of Danish dairy cows," J. Environ. Anal. Toxicol., vol.
2013, 2013.
[97] A. Samsel and S. Seneff, "Glyphosate‟s suppression of cytochrome P450
enzymes and amino acid biosynthesis by the gut microbiome: Pathways to
modern diseases," Entropy, vol. 15, pp. 1416-1463, 2013.
[98] L. Schinasi and M. E. Leon, "Non-Hodgkin lymphoma and occupational
exposure to agricultural pesticide chemical groups and active ingredients: a
systematic review and meta-analysis," Int. J. Env. Res. Public Health, vol. 11,
pp. 4449-4527, 2014.
[99] International Agency for Research on Cancer, "Evaluation of five
organophosphate insecticides and herbicides," World Health Organization
(Ed.), Lyon, France, IARC Monographs, 2015.
[100] D. Buffin and T. Jewell, Health and environmental impacts of glyphosate: the
implications of increased use of glyphosate in association with genetically

6. References
121
modified crops: Friends of the earth, London, 2001.
(http://www.foe.co.uk/resource/reports/impacts_glyphosate.pdf).
[101] L. Fritschi, J. McLaughlin, C. Sergi, G. Calaf, F. Le Curieux, F. Forastiere, H.
Kromhout, P. Egeghy, G. D. Jahnke, C. W. Jameson, and M. T. Martin,
"Carcinogenicity of tetrachlorvinphos, parathion, malathion, diazinon, and
glyphosate," Red, vol. 114, 2015.
[102] M. A. Marin-Morales, B. de Campos Ventura-Camargo, and M. M. Hoshina,
"Toxicity of herbicides: impact on aquatic and soil biota and human health," in
Herbicides–Current Research and Case Studies in Use, ed. A. J. Price, and J.
A. Kelton, Rijeka, Croatia, pp. 399-443, 2013.
[103] Z. Filip, "International approach to assessing soil quality by ecologically-
related biological parameters," Agric., Ecosyst. Environ., vol. 88, pp. 169-174,
2002.
[104] L. Avidano, E. Gamalero, G. P. Cossa, and E. Carraro, "Characterization of
soil health in an Italian polluted site by using microorganisms as
bioindicators," Appl. Soil Ecol., vol. 30, pp. 21-33, 2005.
[105] S. Carlisle and J. Trevors, "Glyphosate in the environment," Water Air Soil
Pollut., vol. 39, pp. 409-420, 1988.
[106] S. Hassan, F. Bigler, H. Bogenschütz, E. Boller, J. Brun, J. Calis, P.
Chiverton, J. Coremans-Pelseneer, C. Duso, G. B. Lewis, and F. Mansour,
"Results of the fifth joint pesticide testing programme carried out by the
IOBC/WPRS-Working Group “Pesticides and Beneficial Organisms”,"
Entomophaga, vol. 36, pp. 55-67, 1991.
[107] J. Springett and R. Gray, "Effect of repeated low doses of biocides on the
earthworm Aporrectodea caliginosa in laboratory culture," Soil Biol.
Biochem., vol. 24, pp. 1739-1744, 1992.
[108] R. J. Kremer and N. E. Means, "Glyphosate and glyphosate-resistant crop
interactions with rhizosphere microorganisms," Eur. J. Agron., vol. 31, pp.
153-161, 2009.
[109] D. Paul, G. Pandey, J. Pandey, and R. K. Jain, "Accessing microbial diversity
for bioremediation and environmental restoration," Trends Biotechnol., vol.
23, pp. 135-142, 2005.
[110] R. K. Jain, M. Kapur, S. Labana, B. Lal, P. M. Sarma, D. Bhattacharya, and I.
S. Thakur, "Microbial diversity: Application of micro-organisms for the
biodegradation of xenobiotics," Curr. Sci., vol. 89, pp. 101-112, 2005.
[111] S. Le Borgne, D. Paniagua, and R. Vazquez-Duhalt, "Biodegradation of
organic pollutants by halophilic bacteria and archaea," J. Mol. Microbiol.
Biotechnol., vol. 15, pp. 74-92, 2008.
[112] D. L. Dindal, "Soil biology guide," Insecta: Coleóptera, vol. 5, p. 28, 1990.
[113] M. Vidali, "Bioremediation. an overview," Pure Appl. Chem., vol. 73, pp.
1163-1172, 2001.
[114] O. Tiryaki and C. Temur, "The fate of pesticide in the environment," J. Biol.
Environ. Sci., vol. 4, 2010.

6. References
122
[115] A. Sviridov, T. Shushkova, I. Ermakova, E. Ivanova, D. Epiktetov, and A.
Leontievsky, "Microbial degradation of glyphosate herbicides (Review),"
Appl. Biochem. Microbiol., vol. 51, pp. 188-195, 2015.
[116] G. F. Barry and G. M. Kishore, "Glyphosate tolerant plants," U. S. Patent No.
5,463,175, 1995.
[117] A. M. Rojano‐Delgado, J. Ruiz‐Jiménez, M. D. L. de Castro, and R. De Prado,
"Determination of glyphosate and its metabolites in plant material by
reversed‐polarity CE with indirect absorptiometric detection," Electrophoresis,
vol. 31, pp. 1423-1430, 2010.
[118] W. Lerbs, M. Stock, and B. Parthier, "Physiological aspects of glyphosate
degradation in Alcaligenes spec. strain GL," Arch. Microbiol., vol. 153, pp.
146-150, 1990.
[119] A. d. Araújo, R. Monteiro, and R. Abarkeli, "Effect of glyphosate on the
microbial activity of two Brazilian soils," Chemosphere, vol. 52, pp. 799-804,
2003.
[120] P. Kafarski, B. Lejczak, and G. Forlani, "Biodegradation of pesticides
containing carbon-to-phosphorus bond," ACS Symp. Ser., Vol. 777, pp. 145-
163, 2001.
[121] S. Kononova and M. Nesmeyanova, "Phosphonates and their degradation by
microorganisms," Biochemistry (Moscow), vol. 67, pp. 184-195, 2002.
[122] S. Anwar, F. Liaquat, Q. M. Khan, Z. M. Khalid, and S. Iqbal,
"Biodegradation of chlorpyrifos and its hydrolysis product 3, 5, 6-trichloro-2-
pyridinol by Bacillus pumilus strain C2A1," J. Hazard. Mater., vol. 168, pp.
400-405, 2009.
[123] K. Jyothi, K. S. Babu, N. Clara, and A. Kashyap, "Identification and isolation
of hydrocarbon degrading bacteria by molecular characterization," Helix, vol.
2, pp. 105-111, 2012.
[124] W. Koneman, D. Allen, M. Janda, C. Schreckenberger, and J. Winn, "Atlas
and textbook of diagonistic microbiology," 5th edition. Philadelphia, USA.
Lippincott Williams & Wilkins, 1997.
[125] A. Mateen, "Purification of the enzyme and cloning of the gene involved in
hydrolysis of carbofuran from Pseudomonas sp. 50432," PhD diss., University
of the Punjab, Lahore, 1998.
[126] K. H. Wilson, R. Blitchington, and R. Greene, "Amplification of bacterial 16S
ribosomal DNA with polymerase chain reaction," J. Clin. Microbiol., vol. 28,
pp. 1942-1946, 1990.
[127] M. Cunliffe and M. A. Kertesz, "Effect of Sphingobium yanoikuyae B1
inoculation on bacterial community dynamics and polycyclic aromatic
hydrocarbon degradation in aged and freshly PAH-contaminated soils,"
Environ. Pollut., vol. 144, pp. 228-237, 2006.
[128] G. Muyzer, E. C. De Waal, and A. G. Uitterlinden, "Profiling of complex
microbial populations by denaturing gradient gel electrophoresis analysis of
polymerase chain reaction-amplified genes coding for 16S rRNA," Appl.
Environ. Microbiol., vol. 59, pp. 695-700, 1993.

6. References
123
[129] K. Qian, T. Tang, T. Shi, F. Wang, J. Li, and Y. Cao, "Residue determination
of glyphosate in environmental water samples with high-performance liquid
chromatography and UV detection after derivatization with 4-chloro-3, 5-
dinitrobenzotrifluoride," Anal. Chim. Acta, vol. 635, pp. 222-226, 2009.
[130] I. T. Ermakova, N. I. Kiseleva, T. Shushkova, M. Zharikov, G. A. Zharikov,
and A. A. Leontievsky, "Bioremediation of glyphosate-contaminated soils,"
Appl. Microbiol. Biotechnol., vol. 88, pp. 585-594, 2010.
[131] H. Steinrücken and N. Amrhein, "The herbicide glyphosate is a potent
inhibitor of 5-enolpyruvylshikimic acid-3-phosphate synthase," Biochem.
Biophys. Res. Commun., vol. 94, pp. 1207-1212, 1980.
[132] L. Arnaud, F. Nurit, P. Ravanel, and M. Tissut, "Distribution of glyphosate
and of its target enzyme inside wheat plants," Pestic. Sci., vol. 40, pp. 217-
223, 1994.
[133] D. McDevitt, D. Payne, D. Holmes, and M. Rosenberg, "Novel targets for the
future development of antibacterial agents," J. Appl. Microbiol., vol. 92, 2002.
[134] J. D. McArthur, N. P. West, J. N. Cole, H. Jungnitz, C. A. Guzmán, J. Chin, P.
R. Lehrbach, S. P. Djordjevic, and M. J. Walker, "An aromatic amino acid
auxotrophic mutant of Bordetella bronchiseptica is attenuated and
immunogenic in a mouse model of infection," FEMS Microbiol. Lett., vol.
221, pp. 7-16, 2003.
[135] T. Parish and N. G. Stoker, "The common aromatic amino acid biosynthesis
pathway is essential in Mycobacterium tuberculosis," Microbiology, vol. 148,
pp. 3069-3077, 2002.
[136] F. Roberts, C. W. Roberts, J. J. Johnson, D. E. Kyle, T. Krell, J. R. Coggins,
G. H. Coombs, W. K. Milhous, S. Tzipori, D. J. Ferguson, and D. Chakrabarti,
"Evidence for the shikimate pathway in apicomplexan parasites," Nature, vol.
393, pp. 801-805, 1998.
[137] G. A. McConkey, "Targeting the shikimate pathway in the malaria parasite
Plasmodium falciparum," Antimicrob. Agents Chemother., vol. 43, pp. 175-
177, 1999.
[138] L. Pollegioni, E. Schonbrunn, and D. Siehl, "Molecular basis of glyphosate
resistance–different approaches through protein engineering," FEBS J., vol.
278, pp. 2753-2766, 2011.
[139] S. B. Powles, "Gene amplification delivers glyphosate-resistant weed
evolution," Proc. Natl. Acad. Sci. U.S.A., vol. 107, pp. 955-956, 2010.
[140] G. M. Dill, C. A. CaJacob, and S. R. Padgette, "Glyphosate‐resistant crops:
adoption, use and future considerations," Pest Manag. Sci., vol. 64, pp. 326-
331, 2008.
[141] W. A. Pline, J. W. Wilcut, S. O. Duke, K. L. Edmisten, and R. Wells,
"Tolerance and accumulation of shikimic acid in response to glyphosate
applications in glyphosate-resistant and nonglyphosate-resistant cotton
(Gossypium hirsutum L.)," J. Agric. Food Chem., vol. 50, pp. 506-512, 2002.

6. References
124
[142] J. Levin and D. Sprinson, "The enzymatic formation and isolation of 3-
enolpyruvylshikimate 5-phosphate," J. Biol. Chem., vol. 239, pp. 1142-1150,
1964.
[143] M. R. Boocock and J. R. Coggins, "Kinetics of 5-enolpyruvylshikimate-3-
phosphate synthase inhibition by glyphosate," FEBS Lett., vol. 154, pp. 127-
133, 1983.
[144] S. Castellino, G. C. Leo, R. D. Sammons, and J. A. Sikorski, "Phosphorus-31,
sup 15 N, and sup 13 C NMR of glyphosate: Comparison of pH titrations to
the herbicidal dead-end complex with 5-enolpyruvoylshikimate-3-phosphate
synthase," Biochemistry;(USA), vol. 28, 1989.
[145] W. Wang, H. Xia, X. Yang, T. Xu, H. J. Si, X. X. Cai, F. Wang, J. Su, A. A.
Snow and B. R. Lu, "A novel 5‐enolpyruvoylshikimate‐3‐phosphate (EPSP)
synthase transgene for glyphosate resistance stimulates growth and fecundity
in weedy rice (Oryza sativa) without herbicide," New Phytol., vol. 202, pp.
679-688, 2014.
[146] A. Oikawa, Y. Nakamura, T. Ogura, A. Kimura, H. Suzuki, N. Sakurai, Y.
Shinbo, D. Shibata, S. Kanaya, and D. Ohta, "Clarification of pathway-
specific inhibition by Fourier transform ion cyclotron resonance/mass
spectrometry-based metabolic phenotyping studies," Plant Physiol., vol. 142,
pp. 398-413, 2006.
[147] W. C. Stallings, S. S. Abdel-Meguid, L. W. Lim, H.-S. Shieh, H. E. Dayringer,
N. K. Leimgruber, R. A. Stegeman, K. S. Anderson, J. A. Sikorski, S. R.
Padgette, and G. M. Kishore, "Structure and topological symmetry of the
glyphosate target 5-enolpyruvylshikimate-3-phosphate synthase: a distinctive
protein fold," Proc. Natl. Acad. Sci. U.S.A., vol. 88, pp. 5046-5050, 1991.
[148] S. Eschenburg, W. Kabsch, M. L. Healy, and E. Schönbrunn, "A new view of
the mechanisms of UDP-N-acetylglucosamine enolpyruvyl transferase (MurA)
and 5-enolpyruvylshikimate-3-phosphate synthase (AroA) derived from X-ray
structures of their tetrahedral reaction intermediate states," J. Biol. Chem., vol.
278, pp. 49215-49222, 2003.
[149] H. Park, J. L. Hilsenbeck, H. J. Kim, W. A. Shuttleworth, Y. H. Park, J. N.
Evans, and C. Kang, "Structural studies of Streptococcus pneumoniae EPSP
synthase in unliganded state, tetrahedral intermediate‐bound state and
S3P‐GLP‐bound state," Mol. Microbiol., vol. 51, pp. 963-971, 2004.
[150] T. Funke, H. Han, M. L. Healy-Fried, M. Fischer, and E. Schönbrunn,
"Molecular basis for the herbicide resistance of Roundup Ready crops," Proc.
Natl. Acad. Sci. U.S.A., vol. 103, pp. 13010-13015, 2006.
[151] M. F. Alibhai, C. Cajacob, P. C. C. Feng, G. R. Heck, Y. Qi, S. Flasinski, W.
C. Stallings, "Glyphosate resistant class I 5-enolpyruvylshikimate-3-phosphate
synthase (epsps)," U. S. Patent No. 7 723 575, 2004.
[152] G. F. Barry, G. M. Kishore, S. R. Padgette, and W. C. Stallings, "Glyphosate-
tolerant 5-enolpyruvylshikimate-3-phosphate synthases," U. S. Patent No. 5
633 435, 1997.
[153] N. Carozzi, B. Carr, and P. E. Hammer, "Identification of a new class of epsp
synthases," U. S. Patent No. 7 700 842, 2010.

6. References
125
[154] J. M. Lira, R. M. CICCHILLO, and S. K. NAIR, "Novel class of glyphosate
resistance genes," U. S. Patent Application No. 15/152 305, 2013.
[155] W. Du, N. G. Wallis, M. J. Mazzulla, A. F. Chalker, L. Zhang, W. S. Liu, H.
Kallender, and D. J. Payne, "Characterization of Streptococcus pneumoniae
5‐enolpyruvylshikimate 3‐phosphate synthase and its activation by univalent
cations," Eur. J. Biochem., vol. 267, pp. 222-227, 2000.
[156] Y.-S. Tian, J. Xu, A.-S. Xiong, W. Zhao, X.-Y. Fu, R.-H. Peng, and Q. H.
Yao, "Improvement of glyphosate resistance through concurrent mutations in
three amino acids of the Ochrobactrum 5-enopyruvylshikimate-3-phosphate
synthase," Appl. Environ. Microbiol., vol. 77, pp. 8409-8414, 2011.
[157] T. Funke, Y. Yang, H. Han, M. Healy-Fried, S. Olesen, A. Becker, and E.
Schönbrunn, "Structural basis of glyphosate resistance resulting from the
double mutation Thr97→ Ile and Pro101→ Ser in 5-enolpyruvylshikimate-3-
phosphate synthase from Escherichia coli," J. Biol. Chem., vol. 284, pp. 9854-
9860, 2009.
[158] S. Padgette, D. B. Re, C. Gasser, D. A. Eichholtz, R. Frazier, C. M. Hironaka,
E. B. Levine, D. M. Shah, R. T. Fraley, and G. M. Kishore, "Site-directed
mutagenesis of a conserved region of the 5-enolpyruvylshikimate-3-phosphate
synthase active site," J. Biol. Chem., vol. 266, pp. 22364-22369, 1991.
[159] S. R. Baerson, D. J. Rodriguez, M. Tran, Y. Feng, N. A. Biest, and G. M. Dill,
"Glyphosate-resistant goosegrass. Identification of a mutation in the target
enzyme 5-enolpyruvylshikimate-3-phosphate synthase," Plant Physiol., vol.
129, pp. 1265-1275, 2002.
[160] M. He, Y.-F. Nie, and P. Xu, "A T42M substitution in bacterial 5-
enolpyruvylshikimate-3-phosphate synthase (EPSPS) generates enzymes with
increased resistance to glyphosate," Biosci., Biotechnol., Biochem., vol. 67, pp.
1405-1409, 2003.
[161] D. Jin, W. Lu, S. Ping, W. Zhang, J. Chen, B. Dun, R. Ma, Z. Zhao, J. Sha, L.
Li, and Z. Yang, "Identification of a new gene encoding EPSPS with high
glyphosate resistance from the metagenomic library," Curr. Microbiol., vol.
55, pp. 350-355, 2007.
[162] J. Fitzgibbon and H. Braymer, "Cloning of a gene from Pseudomonas sp.
strain PG2982 conferring increased glyphosate resistance," Appl. Environ.
Microbiol., vol. 56, pp. 3382-3388, 1990.
[163] L. Li, W. Lu, Y. Han, S. Ping, W. Zhang, M. Chen, Z. Zhao, Y. Yan, Y. Jiang,
and M. Lin, "A novel RPMXR motif among class II 5-enolpyruvylshikimate-
3-phosphate synthases is required for enzymatic activity and glyphosate
resistance," J. Biotechnol., vol. 144, pp. 330-336, 2009.
[164] P. Vaithanomsat and K. A. Brown, "Isolation and mutation of recombinant
EPSP synthase from pathogenic bacteria Pseudomonas aeruginosa," Process
Biochem., vol. 42, pp. 592-598, 2007.
[165] Y.-C. Sun, Y.-C. Chen, Z.-X. Tian, F.-M. Li, X.-Y. Wang, J. Zhang, Z. L.
Xiao, M. Lin, N. Gilmartin, D. N. Dowling, and Y. P. Wang, "Novel AroA
with high tolerance to glyphosate, encoded by a gene of Pseudomonas putida

6. References
126
4G-1 isolated from an extremely polluted environment in China," Appl.
Environ. Microbiol., vol. 71, pp. 4771-4776, 2005.
[166] S. Eschenburg, M. L. Healy, M. A. Priestman, G. H. Lushington, and E.
Schönbrunn, "How the mutation glycine96 to alanine confers glyphosate
insensitivity to 5-enolpyruvyl shikimate-3-phosphate synthase from
Escherichia coli," Planta, vol. 216, pp. 129-135, 2002.
[167] L. Wang, R. Peng, Y. Tian, J. Han, W. Zhao, B. Wang, M. Liu, and Q. Yao,
"Characterization of a class II 5-enopyruvylshikimate-3-phosphate synthase
with high tolerance to glyphosate from Sinorhizobium fredii," World J.
Microbiol. Biotechnol., vol. 30, pp. 2967-2973, 2014.
[168] L. Comai, L. C. Sen, and D. M. Stalker, "An altered aroA gene product
confers resistance to the herbicide glyphosate," Science, vol. 221, pp. 370-371,
1983.
[169] D. Sost and N. Amrhein, "Substitution of Gly-96 to Ala in the 5-
enolpyruvylshikimate 3-phosphate synthase of Klebsiella pneumoniae results
in a greatly reduced affinity for the herbicide glyphosate," Arch. Biochem.
Biophys., vol. 282, pp. 433-436, 1990.
[170] C. Ng, R. Wickneswari, S. Salmijah, Y. Teng, and B. Ismail, "Gene
polymorphisms in glyphosate‐resistant and‐susceptible biotypes of Eleusine
indica from Malaysia," Weed Res., vol. 43, pp. 108-115, 2003.
[171] A. Perez-Jones, K.-W. Park, N. Polge, J. Colquhoun, and C. A. Mallory-
Smith, "Investigating the mechanisms of glyphosate resistance in Lolium
multiflorum," Planta, vol. 226, pp. 395-404, 2007.
[172] A. Wakelin, D. Lorraine‐Colwill, and C. Preston, "Glyphosate resistance in
four different populations of Lolium rigidum is associated with reduced
translocation of glyphosate to meristematic zones," Weed Res., vol. 44, pp.
453-459, 2004.
[173] Q. Yu, A. Cairns, and S. Powles, "Glyphosate, paraquat and ACCase multiple
herbicide resistance evolved in a Lolium rigidum biotype," Planta, vol. 225,
pp. 499-513, 2007.
[174] S. Duke, "Herbicide Resistant Crops: Agricultural," Environmental,
Economic, Regulatory, and Technical Aspects, CRC Press, Boca Raton, FL,
420pp, 1996.
[175] D. A. Eichholtz, C. S. Gasser, and G. M. Kishore, "Modified gene encoding
glyphosate-tolerant 5-enolpruvyl-3-phosphoshikimate synthase," U. S. Patent
No. 6,225,114, 2001.
[176] D. Kahrizi, A. H. Salmanian, A. Afshari, A. Moieni, and A. Mousavi,
"Simultaneous substitution of Gly96 to Ala and Ala183 to Thr in 5-
enolpyruvylshikimate-3-phosphate synthase gene of E. coli (k12) and
transformation of rapeseed (Brassica napus L.) in order to make tolerance to
glyphosate," Plant Cell Rep., vol. 26, pp. 95-104, 2007.
[177] M. Lebrun, A. Sailland, G. Freyssinet, and E. DeGryse, "Mutated 5-
enolpyruvylshikimate-3-phosphate synthase, gene coding for said protein and
transformed plants containing said gene," U.S. Patent No. 6,566,587, 2003.

6. References
127
[178] M. Spencer, R. Mumm, and J. Gwyn, "Glyphosate resistant maize lines," U.S.
Patent No. 6,040,497, 2000.
[179] S. Beulke and C. D. Brown, "Evaluation of methods to derive pesticide
degradation parameters for regulatory modelling," Biol. Fertility Soils, vol. 33,
pp. 558-564, 2001.
[180] P. Sprankle, C. Sandberg, W. Meggitt, and D. Penner, "Separation of
glyphosate and possible metabolities by thin-layer chromatography," Weed
Sci., vol. 26, pp. 673-674, 1978.
[181] K. Tamura, G. Stecher, D. Peterson, A. Filipski, and S. Kumar, "MEGA6:
molecular evolutionary genetics analysis version 6.0," Mol. Biol. Evol., vol.
30, pp. 2725-2729, 2013.
[182] E. Gasteiger, C. Hoogland, A. Gattiker, S. e. Duvaud, M. R. Wilkins, R. D.
Appel, and A. Bairoch, Protein identification and analysis tools on the
ExPASy server, The Proteomics Protocols Handbook. Eds J. M. Walker,
Totowa, N.J., Humana Press, 2005.
[183] L. A. Kelley, S. Mezulis, C. M. Yates, M. N. Wass, and M. J. Sternberg, "The
Phyre2 web portal for protein modeling, prediction and analysis," Nat.
Protoc., vol. 10, pp. 845-858, 2015.
[184] C. Geourjon and G. Deleage, "SOPMA: significant improvements in protein
secondary structure prediction by consensus prediction from multiple
alignments," Computer applications in the biosciences: CABIOS, vol. 11, pp.
681-684, 1995.
[185] C. Combet, M. Jambon, G. Deleage, and C. Geourjon, "Geno3D: automatic
comparative molecular modelling of protein," Bioinformatics, vol. 18, pp.
213-214, 2002.
[186] K. Arnold, L. Bordoli, J. Kopp, and T. Schwede, "The SWISS-MODEL
workspace: a web-based environment for protein structure homology
modelling," Bioinformatics, vol. 22, pp. 195-201, 2006.
[187] R. A. Laskowski, J. A. C. Rullmann, M. W. MacArthur, R. Kaptein, and J. M.
Thornton, "AQUA and PROCHECK-NMR: programs for checking the quality
of protein structures solved by NMR," J. Biomol. NMR, vol. 8, pp. 477-486,
1996.
[188] S. A. Hollingsworth and P. A. Karplus, "A fresh look at the Ramachandran
plot and the occurrence of standard structures in proteins," Biomol. Concepts,
vol. 1, pp. 271-283, 2010.
[189] M. Wiederstein and M. J. Sippl, "ProSA-web: interactive web service for the
recognition of errors in three-dimensional structures of proteins," Nucleic
Acids Res., vol. 35, pp. W407-W410, 2007.
[190] S.-y. Yi, Y. Cui, Y. Zhao, Z.-d. Liu, Y.-j. Lin, and F. Zhou, "A Novel
Naturally Occurring Class I 5-Enolpyruvylshikimate-3-Phosphate Synthase
from Janibacter sp. Confers High Glyphosate Tolerance to Rice," Sci. Rep.,
vol. 6, 2016.
[191] F. Aujoulat, S. Romano-Bertrand, A. Masnou, H. Marchandin, and E. Jumas-
Bilak, "Niches, population structure and genome reduction in Ochrobactrum

6. References
128
intermedium: clues to technology-driven emergence of pathogens," PLoS One,
vol. 9, p. e83376, 2014.
[192] S. Pandey, P. K. Ghosh, S. Ghosh, T. K. De, and T. K. Maiti, "Role of heavy
metal resistant Ochrobactrum sp. and Bacillus spp. strains in bioremediation of
a rice cultivar and their PGPR like activities," J. Microbiol., vol. 51, p. 11,
2013.
[193] N. Wackerow-Kouzova, "Ochrobactrum intermedium ANKI, a nitrogen-fixing
bacterium able to decolorize azobenzene," Appl. Biochem. Microbiol., vol. 43,
p. 403, 2007.
[194] P. Waranusantigul, H. Lee, M. Kruatrachue, P. Pokethitiyook, and C.
Auesukaree, "Isolation and characterization of lead-tolerant Ochrobactrum
intermedium and its role in enhancing lead accumulation by Eucalyptus
camaldulensis," Chemosphere, vol. 85, pp. 584-590, 2011.
[195] B. Kavita and H. Keharia, "Reduction of hexavalent chromium by
Ochrobactrum intermedium BCR400 isolated from a chromium-contaminated
soil," 3 Biotech, vol. 2, pp. 79-87, 2012.
[196] Z. F. Boukhatem, O. Domergue, A. Bekki, C. Merabet, S. Sekkour, F.
Bouazza, R. Duponnois, P. De Lajudie, and A. Galiana, "Symbiotic
characterization and diversity of rhizobia associated with native and
introduced acacias in arid and semi-arid regions in Algeria," FEMS Microbiol.
Ecol., vol. 80, pp. 534-547, 2012.
[197] R. Zhang, Z. Cui, J. Jiang, J. He, X. Gu, and S. Li, "Diversity of
organophosphorus pesticide-degrading bacteria in a polluted soil and
conservation of their organophosphorus hydrolase genes," Can. J. Microbiol.,
vol. 51, pp. 337-343, 2005.
[198] W. S. El-Sayed, M. K. Ibrahim, M. Abu-Shady, F. El-Beih, N. Ohmura, H.
Saiki, and A. Ando,"Isolation and identification of a novel strain of the genus
Ochrobactrum with phenol-degrading activity," J. Biosci. Bioeng., vol. 96, pp.
310-312, 2003.
[199] Y. Veeranagouda, P. E. Paul, P. Gorla, D. Siddavattam, and T. Karegoudar,
"Complete mineralisation of dimethylformamide by Ochrobactrum sp.
DGVK1 isolated from the soil samples collected from the coalmine leftovers,"
Appl. Microbiol. Biotechnol., vol. 71, pp. 369-375, 2006.
[200] E. Katsivela, E. R. Moore, D. Maroukli, C. Strömpl, D. Pieper, and N.
Kalogerakis, "Bacterial community dynamics during in-situ bioremediation of
petroleum waste sludge in landfarming sites," Biodegradation, vol. 16, pp.
169-180, 2005.
[201] Y.-M. Kim, K. Park, G.-J. Joo, E.-M. Jeong, J.-E. Kim, and I.-K. Rhee,
"Glutathione-dependent biotransformation of the fungicide chlorothalonil," J.
Agric. Food Chem., vol. 52, pp. 4192-4196, 2004.
[202] G. Ozdemir, T. Ozturk, N. Ceyhan, R. Isler, and T. Cosar, "Heavy metal
biosorption by biomass of Ochrobactrum anthropi producing
exopolysaccharide in activated sludge," Bioresour. Technol., vol. 90, pp. 71-
74, 2003.

6. References
129
[203] S. Ferhat, R. Alouaoui, A. Badis, and N. Moulai-Mostefa, "Production and
characterization of biosurfactant by free and immobilized cells from
Ochrobactrum intermedium isolated from the soil of southern Algeria with a
view to environmental application," Biotechnol. Biotechnol. Equip., pp. 1-10,
2017.
[204] Y.-S. Tian, A.-S. Xiong, J. Xu, W. Zhao, F. Gao, X.-Y. Fu, H. Xu, J. L.
Zheng, R. H. Peng, and Q. H. Yao, "Isolation from Ochrobactrum anthropi of
a novel class II 5-enopyruvylshikimate-3-phosphate synthase with high
tolerance to glyphosate," Appl. Environ. Microbiol., vol. 76, pp. 6001-6005,
2010.
[205] K. Sivakumar and S. Balaji, "In silico characterization of antifreeze proteins
using computational tools and servers," J. Chem. Sci., vol. 119, pp. 571-579,
2007.
[206] A.-F. Miller, "Superoxide dismutases: active sites that save, but a protein that
kills," Curr. Opin. Chem. Biol., vol. 8, pp. 162-168, 2004.
[207] V. E. Hayes, N. G. Ternan, and G. McMullan, "Organophosphonate
metabolism by a moderately halophilic bacterial isolate," FEMS Microbiol.
Lett., vol. 186, pp. 171-175, 2000.
[208] B. K. Singh and A. Walker, "Microbial degradation of organophosphorus
compounds," FEMS Microbiol. Rev., vol. 30, pp. 428-471, 2006.
[209] A. L. Cerdeira and S. O. Duke, "The current status and environmental impacts
of glyphosate-resistant crops," J. Environ. Qual., vol. 35, pp. 1633-1658,
2006.
[210] V. Joseph P, S. Nader, R. Darren E, T. Francois J, L. Mark B, and S. Peter H,
"Glyphosate-resistant giant ragweed (Ambrosia trifida L.) in Ontario: dose
response and control with postemergence herbicides," Am. J. Plant Sci., vol.
2012, 2012.
[211] Y. Sharifi, A. A. Pourbabaei, A. Javadi, M. H. Abdolmohammad, M. Saffari,
and A. Morovvati, "Biodegradation of glyphosate herbicide by Salinicoccus
spp isolated from Qom Hoze-soltan lake, Iran," Environ. Health Eng. Manag.
J., vol. 2, pp. 31-36, 2015.
[212] G. M. Dill, R. D. Sammons, P. C. Feng, F. Kohn, K. Kretzmer, A.
Mehrsheikh, M. Bleeke, J. L. Honegger, D. Farmer, D. Wright, and E. A.
Haupfear, "Glyphosate: Discovery, Development, Applications, and
Properties," Glyphosate Resistance in Crops and Weeds: History,
Development, and Management, Hoboken, John Wiley and Sons, Inc., pp. 1-
33, 2010.
[213] T. Yamada, R. J. Kremer, P. R. d. C. e Castro, and B. W. Wood, "Glyphosate
interactions with physiology, nutrition, and diseases of plants: Threat to
agricultural sustainability?," Eur. J. Agron., vol. 31, pp. 111-113, 2009.
[214] G. Johal and D. Huber, "Glyphosate effects on diseases of plants," Eur. J.
Agron., vol. 31, pp. 144-152, 2009.
[215] S. Eker, L. Ozturk, A. Yazici, B. Erenoglu, V. Romheld, and I. Cakmak,
"Foliar-applied glyphosate substantially reduced uptake and transport of iron

6. References
130
and manganese in sunflower (Helianthus annuus L.) plants," J. Agric. Food
Chem., vol. 54, pp. 10019-10025, 2006.
[216] R. Annett, H. R. Habibi, and A. Hontela, "Impact of glyphosate and
glyphosate‐based herbicides on the freshwater environment," J. Appl. Toxicol.,
vol. 34, pp. 458-479, 2014.
[217] M. Cuhra, T. Traavik, and T. Bøhn, "Clone-and age-dependent toxicity of a
glyphosate commercial formulation and its active ingredient in Daphnia
magna," Ecotoxicology, vol. 22, pp. 251-262, 2013.
[218] M. Gaupp-Berghausen, M. Hofer, B. Rewald, and J. G. Zaller, "Glyphosate-
based herbicides reduce the activity and reproduction of earthworms and lead
to increased soil nutrient concentrations," Sci. Rep., vol. 5, 2015.
[219] J. G. Zaller, F. Heigl, L. Ruess, and A. Grabmaier, "Glyphosate herbicide
affects belowground interactions between earthworms and symbiotic
mycorrhizal fungi in a model ecosystem," Sci. Rep., vol. 4, 2014.
[220] R. Mesnage, N. Defarge, J. S. de Vendomois, and G. Seralini, "Potential toxic
effects of glyphosate and its commercial formulations below regulatory
limits," Food Chem. Toxicol., vol. 84, pp. 133-153, 2015.
[221] International Agency for Research on Cancer and World Health Organization,
"IARC Monographs Volume 112: Evaluation of Five Organophosphate
Insecticides and Herbicides," ed Lyon France, 2015.
[222] T. Shushkova, I. Ermakova, and A. Leontievsky, "Glyphosate bioavailability
in soil," Biodegradation, vol. 21, pp. 403-410, 2010.
[223] T. M. Balthazor and L. E. Hallas, "Glyphosate-degrading microorganisms
from industrial activated sludge," Appl. Environ. Microbiol., vol. 51, pp. 432-
434, 1986.
[224] L. Hallas, W. Adams, and M. Heitkamp, "Glyphosate degradation by
immobilized bacteria: field studies with industrial wastewater effluent," Appl.
Environ. Microbiol., vol. 58, pp. 1215-1219, 1992.
[225] D. L. Shinabarger and H. D. Braymer, "Glyphosate catabolism by
Pseudomonas sp. strain PG2982," J. Bacteriol., vol. 168, pp. 702-707, 1986.
[226] R. Pipke, N. Amrhein, G. S. Jacob, J. Schaefer, and G. M. Kishore,
"Metabolism of glyphosate in an Arthrobacter sp. GLP‐1," Europ. J.
Biochem., vol. 165, pp. 267-273, 1987.
[227] A. Selvapandiyan and R. K. Bhatnagar, "Isolation of a glyphosate-
metabolising Pseudomonas: detection, partial purification and localisation of
carbon-phosphorus lyase," Appl. Microbiol. Biotechnol., vol. 40, pp. 876-882,
1994.
[228] A. V. Sviridov, T. V. Shushkova, N. F. Zelenkova, N. G. Vinokurova, I. G.
Morgunov, I. T. Ermakova, and A. A. Leontievsky, "Distribution of
glyphosate and methylphosphonate catabolism systems in soil bacteria
Ochrobactrum anthropi and Achromobacter sp," Appl. Microbiol. Biotechnol.,
vol. 93, pp. 787-796, 2012.

6. References
131
[229] H. W. Talbot, L. M. Johnson, and D. M. Munnecke, "Glyphosate utilization by
Pseudomonas sp. and Alcaligenes sp. isolated from environmental sources,"
Curr. Microbiol., vol. 10, pp. 255-259, 1984.
[230] G. Jacob, J. Garbow, L. Hallas, N. Kimack, G. Kishore, and J. Schaefer,
"Metabolism of glyphosate in Pseudomonas sp. strain LBr," Appl. Environ.
Microbiol., vol. 54, pp. 2953-2958, 1988.
[231] K. S. McAuliffe, L. E. Hallas, and C. F. Kulpa, "Glyphosate degradation by
Agrobacterium radiobacter isolated from activated sludge," J. Ind. Microbiol.,
vol. 6, pp. 219-221, 1990.
[232] F. Hadi, A. Mousavi, K. A. Noghabi, H. G. Tabar, and A. H. Salmanian, "New
bacterial strain of the genus Ochrobactrum with glyphosate-degrading
activity," J. Environ. Sci. Health, Part B, vol. 48, pp. 208-213, 2013.
[233] R. Pipke and N. Amrhein, "Isolation and characterization of a mutant of
Arthrobacter sp. strain GLP-1 which utilizes the herbicide glyphosate as its
sole source of phosphorus and nitrogen," Appl. Environ. Microbiol., vol. 54,
pp. 2868-2870, 1988.
[234] A. Obojska, B. Lejczak, and M. Kubrak, "Degradation of phosphonates by
streptomycete isolates," Appl. Microbiol. Biotechnol., vol. 51, pp. 872-876,
1999.
[235] C.-M. Liu, P. McLean, C. Sookdeo, and F. Cannon, "Degradation of the
herbicide glyphosate by members of the family Rhizobiaceae," Appl. Environ.
Microbiol., vol. 57, pp. 1799-1804, 1991.
[236] L. P. Wackett, S. L. Shames, C. P. Venditti, and C. T. Walsh, "Bacterial
carbon-phosphorus lyase: products, rates, and regulation of phosphonic and
phosphinic acid metabolism," J. Bacteriol., vol. 169, pp. 710-717, 1987.
[237] R. Pipke and N. Amrhein, "Degradation of the phosphonate herbicide
glyphosate by Arthrobacter atrocyaneus ATCC 13752," Appl. Environ.
Microbiol., vol. 54, pp. 1293-1296, 1988.
[238] A. Obojska, N. G. Ternan, B. Lejczak, P. Kafarski, and G. McMullan,
"Organophosphonate utilization by the thermophile Geobacillus
caldoxylosilyticus T20," Appl. Environ. Microbiol., vol. 68, pp. 2081-2084,
2002.
[239] J. Gard, P. Feng, and W. Hutton, "Nuclear magnetic resonance tim ecourse
studies of glyphosate metabolism by microbial soil isolates," Xenobiotica, vol.
27, pp. 633-644, 1997.
[240] A. Peñaloza-Vazquez, G. L. Mena, L. Herrera-Estrella, and A. M. Bailey,
"Cloning and sequencing of the genes involved in glyphosate utilization by
Pseudomonas pseudomallei," Appl. Environ. Microbiol., vol. 61, pp. 538-543,
1995.
[241] J. K. Moore, H. D. Braymer, and A. D. Larson, "Isolation of a Pseudomonas
sp. which utilizes the phosphonate herbicide glyphosate," Appl. Environ.
Microbiol., vol. 46, pp. 316-320, 1983.

6. References
132
[242] M. M. Nourouzi, T. G. Chuah, T. S. Choong, and C. J. Lim, "Glyphosate
utilization as the source of carbon: isolation and identification of new
bacteria," J. Chem., vol. 8, pp. 1582-1587, 2011.
[243] M. Hoodaji, A. Tahmourespour, and M. Partoazar, "The Efficiency of
Glyphosate Biodegradation by Pseudomonas (aeruginosa)," Microbes in
Applied Research: Current Advances and Challenges: Malaga, Spain, 14-16
September 2011, p. 183, 2012.
[244] Y. V. Kryuchkova, G. L. Burygin, N. E. Gogoleva, Y. V. Gogolev, M. P.
Chernyshova, O. E. Makarov, E. E. Fedorov, and O. V. Turkovskaya,
"Isolation and characterization of a glyphosate-degrading rhizosphere strain,
Enterobacter cloacae K7," Microbiol. Res. vol. 169, pp. 99-105, 2014.
[245] O. Benslama and A. Boulahrouf, "Isolation and characterization of
glyphosate-degrading bacteria from different soils of Algeria," Afr. J.
Microbiol. Res., vol. 7, pp. 5587-5595, 2013.
[246] A. D. Kim, A. S. Baker, D. Dunaway-Mariano, W. Metcalf, B. Wanner, and
B. M. Martin, "The 2-aminoethylphosphonate-specific transaminase of the 2-
aminoethylphosphonate degradation pathway," J. Bacteriol., vol. 184, pp.
4134-4140, 2002.
[247] M. C. Morais, G. Zhang, W. Zhang, D. B. Olsen, D. Dunaway-Mariano, and
K. N. Allen, "X-ray crystallographic and site-directed mutagenesis analysis of
the mechanism of Schiff-base formation in phosphonoacetaldehyde hydrolase
catalysis," J. Biol. Chem., vol. 279, pp. 9353-9361, 2004.
[248] J. P. Quinn, A. N. Kulakova, N. A. Cooley, and J. W. McGrath, "New ways to
break an old bond: the bacterial carbon-phosphorus hydrolases and their role
in biogeochemical phosphorus cycling," Environ. Microbiol., vol. 9, pp. 2392-
2400, 2007.
[249] A. K. White and W. W. Metcalf, "Microbial metabolism of reduced
phosphorus compounds," Annu. Rev. Microbiol., vol. 61, pp. 379-400, 2007.
[250] J. W. McGrath, J. P. Chin, and J. P. Quinn, "Organophosphonates revealed:
new insights into the microbial metabolism of ancient molecules," Nat. Rev.
Microbiol., vol. 11, pp. 412-419, 2013.
[251] S. L. Shames, L. P. Wackett, M. S. LaBarge, R. L. Kuczkowski, and C. T.
Walsh, "Fragmentative and stereochemical isomerization probes for homolytic
carbon to phosphorus bond scission catalyzed by bacterial carbon-phosphorus
lyase," Bioorg. Chem., vol. 15, pp. 366-373, 1987.
[252] G. Yakovleva, S.-K. Kim, and B. Wanner, "Phosphate-independent expression
of the carbon-phosphorus lyase activity of Escherichia coli," Appl. Microbiol.
Biotechnol., vol. 49, pp. 573-578, 1998.
[253] B. Hove-Jensen, T. J. Rosenkrantz, D. L. Zechel, and M. Willemoës,
"Accumulation of intermediates of the carbon-phosphorus lyase pathway for
phosphonate degradation in phn mutants of Escherichia coli," J. Bacteriol.,
vol. 192, pp. 370-374, 2010.
[254] B. Hove-Jensen, F. R. McSorley, and D. L. Zechel, "Physiological role of
PhnP-specified phosphoribosyl cyclic phosphodiesterase in catabolism of

6. References
133
organophosphonic acids by the carbon-phosphorus lyase pathway," J. Am.
Chem. Soc., vol. 133, pp. 3617-3624, 2011.
[255] W. W. Metcalf and B. L. Wanner, "Mutational analysis of an Escherichia coli
fourteen-gene operon for phosphonate degradation, using TnphoA'elements,"
J. Bacteriol., vol. 175, pp. 3430-3442, 1993.
[256] S. S. Kamat, H. J. Williams, and F. M. Raushel, "Intermediates in the
transformation of phosphonates to phosphate by bacteria," Nature, vol. 480,
pp. 570-573, 2011.
[257] G. Kishore and G. S. Jacob, "Degradation of glyphosate by Pseudomonas sp.
PG2982 via a sarcosine intermediate," J. Biol. Chem., vol. 262, pp. 12164-
12168, 1987.
[258] D. Hamsaveni, S. Prapulla, and S. Divakar, "Response surface methodological
approach for the synthesis of isobutyl isobutyrate," Process Biochem., vol. 36,
pp. 1103-1109, 2001.
[259] Y. R. Abdel-Fattah, E. R. El-Helow, K. M. Ghanem, and W. A. Lotfy,
"Application of factorial designs for optimization of avicelase production by a
thermophilic Geobacillus isolate," Res. J. Microbiol., vol. 2, pp. 13-23, 2007.
[260] M. Seth and S. Chand, "Biosynthesis of tannase and hydrolysis of tannins to
gallic acid by Aspergillus awamori-optimisation of process parameters,"
Process Biochem., vol. 36, pp. 39-44, 2000.
[261] J. Zolgharnein, A. Shahmoradi, and J. B. Ghasemi, "Comparative study of
Box–Behnken, central composite, and Doehlert matrix for multivariate
optimization of Pb (II) adsorption onto Robinia tree leaves," J. Chemometrics,
vol. 27, pp. 12-20, 2013.
[262] Z.-W. Wang and X.-L. Liu, "Medium optimization for antifungal active
substances production from a newly isolated Paenibacillus sp. using response
surface methodology," Bioresour. Technol., vol. 99, pp. 8245-8251, 2008.
[263] J. Xu, G.-P. Sheng, H.-W. Luo, F. Fang, W.-W. Li, R. J. Zeng, Z. H. Tong,
and H. Q. Yu, "Evaluating the influence of process parameters on soluble
microbial products formation using response surface methodology coupled
with grey relational analysis," Water Res., vol. 45, pp. 674-680, 2011.
[264] E. Alexa, M. Hafner, M. Negrea, and A. Lazureanu, "HPLC and GC
determination of glyphosate and aminomethylphosphonic acid (AMPA) in
water samples," in 43rd Croatian and 3rd International Symposium on
Agriculture. Opatija, Croatia, pp. 100-105. 2008.
[265] A. Sviridov, T. Shushkova, I. Ermakova, E. Ivanova, and A. Leontievsky,
"Glyphosate: safety risks, biodegradation, and bioremediation," in Current
Environmental Issues and Challenges, ed: Springer, 2014, pp. 183-195.
[266] S. F. Nishino and J. C. Spain, "Oxidative athway for the biodegradation of
nitrobenzene by Comamonas sp. strain JS765," Appl. Environ. Microbiol., vol.
61, pp. 2308-2313, 1995.
[267] U. U. Jadhav, V. V. Dawkar, G. S. Ghodake, and S. P. Govindwar,
"Biodegradation of direct red 5B, a textile dye by newly isolated Comamonas
sp. UVS," J. Hazard. Mater., vol. 158, pp. 507-516, 2008.

6. References
134
[268] H. S. Bae, J. M. Lee, Y. B. Kim, and S.-T. Lee, "Biodegradation of the
mixtures of 4-chlorophenol and phenol by Comamonas testosteroni CPW301,"
Biodegradation, vol. 7, pp. 463-469, 1996.
[269] B. Zhao, X. Hua, F. Wang, W. Dong, Z. Li, Y. Yang, Z. Cui, and M. Wang,
"Biodegradation of propyzamide by Comamonas testosteroni W1 and cloning
of the propyzamide hydrolase gene camH," Bioresource Technol., vol. 179,
pp. 144-149, 2015.
[270] Y. Chen, L. Chai, Y. Zhu, Z. Yang, Y. Zheng, and H. Zhang, "Biodegradation
of kraft lignin by a bacterial strain Comamonas sp. B‐9 isolated from eroded
bamboo slips," J. Appl. Microbiol., vol. 112, pp. 900-906, 2012.
[271] A. Moneke, G. Okpala, and C. Anyanwu, "Biodegradation of glyphosate
herbicide in vitro using bacterial isolates from four rice fields," African J.
Biotechnol., vol. 9, pp. 4067-4074, 2010.
[272] N. Awasthi, R. Ahuja, and A. Kumar, "Factors influencing the degradation of
soil-applied endosulfan isomers," Soil Biol. Biochem., vol. 32, pp. 1697-1705,
2000.
[273] E. A. Wolski, S. E. Murialdo, and J. F. Gonzalez, "Effect of pH and inoculum
size on pentachlorophenol degradation by Pseudomonas sp.," Water SA, vol.
32, pp. 93-98, 2006.
[274] A. Samsel and S. Seneff, "Glyphosate, pathways to modern diseases III:
Manganese, neurological diseases, and associated pathologies," Surg. Neurol.
Int., vol. 6, p. 45, 2015.
[275] A. Kumar, B. Bisht, V. Joshi, and T. Dhewa, "Review on bioremediation of
polluted environment: a management tool," Int. J. Environ. Sci., vol. 1, p.
1079, 2011.
[276] A. Hess, B. Zarda, D. Hahn, A. Häner, D. Stax, P. Höhener, and J. Zeyer, "In
situ analysis of denitrifying toluene-and m-xylene-degrading bacteria in a
diesel fuel-contaminated laboratory aquifer column," Appl. Environ.
Microbiol., vol. 63, pp. 2136-2141, 1997.
[277] V. H. Smith, D. W. Graham, and D. D. Cleland, "Application of resource-ratio
theory to hydrocarbon biodegradation," Environ. Sci. Technol., vol. 32, pp.
3386-3395, 1998.
[278] C. Y. Tang, Q. S. Fu, C. S. Criddle, and J. O. Leckie, "Effect of flux
(transmembrane pressure) and membrane properties on fouling and rejection
of reverse osmosis and nanofiltration membranes treating perfluorooctane
sulfonate containing wastewater," Environ. Sci. Technol., vol. 41, pp. 2008-
2014, 2007.
135
Appendices
Appendix - A
Minimal Salt Media (MSM)
Chemical Concentration
Distilled water 1 L
Na2 H PO4 5.8 g/L
KH2PO4 3.0 g/L
NaCl 0.5 g/L
NH4Cl 1.0 g/L
MgSO4.7H2O 0.25 g/L
pH 6.8-7.00
Appendix - B
Lauria Bertani Medium
Chemical Concentration
Distilled water 1 L
Trypton 10 g/L
Yeast extract 5 g/L
NaCl 5 g/L
pH 7.2±0.2
Appendices
136
Appendix - C
Enrichment Media
Appendix - D
Minimal Media
Chemical Concentration
NH4Cl 20 mM
NaCl/KCl 20 mM
MgCl2 0.5 mM
KP 25 mM
Na2SO4 0.5 mM
Trace element
solutions
(100X)
10 mL
pH 7.0
Chemical Concentration
Tris base 50 mM
NH4Cl 20 mM
Nasuccinate 25 mM
NaCl/KCl 20 mM
KP 10 mM
Na2SO4 1 mM
Trace element
solutions
(100X)
10 mL
pH 7.0

Appendices
137
Appendix - E
100X Trace Element Solution
Appendix - F
Solutions for Gram’s reaction
Crystal violet solution
Chemical Concentration
Distilled water 400 mL
Crystal violet 10 g
Ammonium oxalate 4 g
Ethanol 100 mL
Iodine solution
Chemical Concentration
Distilled water 25 mL
Iodine 1 g
Potassium iodide 2 g
Ethanol 10 mL
Chemical Concentration
Na2-EDTA.2H2O 500 mg/L
FeCl2.4H2O 143 mg/L
ZnCl2 4.7 mg/L
MnCl2.4H2O 3.0 mg/L
H3BO3 30 mg/L
CoCl2.6H2O 20 mg/L
CuCl2.2H2O 1.0 mg/L
NiCl2.6H2O 2.0 mg/L
Na2MoO4.2H2O 3.0 mg/L
CaCl2.2H2O 100 mg/L

Appendices
138
Safranin solution
Chemical Concentration
Distilled water 100 mL
Safranin 2.5 g
Ethanol 10 mL
Appendix - G
Saline solution (0.9%)
Appendix - H
Phosphate Free Media
Chemical Concentration
Distilled water 100 mL
NaCl 0.9 g
Chemical Concentration
NH4Cl 20 mM
NaCl/KCl 20 mM
MgCl2 0.5 mM
Nasuccinate 25 mM
Na2SO4 0.5 mM
Trace element
solutions
(100X)
10 mL
pH 7.0
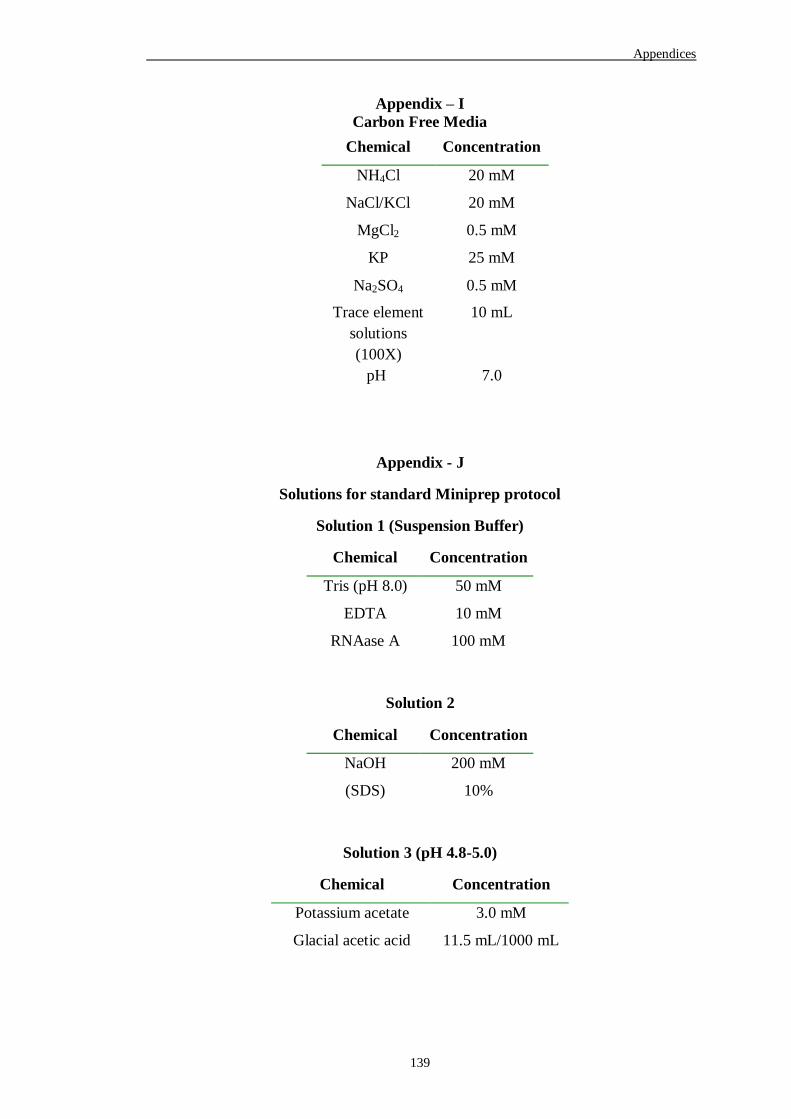
Appendices
139
Appendix – I
Carbon Free Media
Appendix - J
Solutions for standard Miniprep protocol
Solution 1 (Suspension Buffer)
Chemical Concentration
Tris (pH 8.0) 50 mM
EDTA 10 mM
RNAase A 100 mM
Solution 2
Chemical Concentration
NaOH 200 mM
(SDS) 10%
Solution 3 (pH 4.8-5.0)
Chemical Concentration
Potassium acetate 3.0 mM
Glacial acetic acid 11.5 mL/1000 mL
Chemical Concentration
NH4Cl 20 mM
NaCl/KCl 20 mM
MgCl2 0.5 mM
KP 25 mM
Na2SO4 0.5 mM
Trace element
solutions
(100X)
10 mL
pH 7.0





























