Embed Size (px)
Citation preview

Mc
C
Fb
C
(ptmobcI�dpv
onrl
p
0d
Seminars in Anesthesia, Perioperative Medicine and Pain (2007) 26, 28-34
olecular insights into �2 adrenergic receptor function:linical implications
arl Hurt, MD, PhD,a and Timothy Angelotti, MD, PhDb
rom the aDepartment of Molecular and Cellular Physiology, andDepartment of Anesthesia and Division of Critical Care Medicine, Stanford University Medical Center, Stanford,
alifornia.Efferent signaling from the sympathetic nervous system integrates myriad vital functions. Modulationof sympathetic neuron signaling occurs in part by feedback inhibition of neurotransmitter release,mediated largely via �2a and/or �2c adrenergic receptors (�2a&c ARs). This review will describe anoverview of our current understanding of �2a&c AR biology, clinical implications of these findings,and the possible clinical importance of genetic variants of �2a&c ARs.© 2007 Elsevier Inc. All rights reserved.
KEYWORDS:Adrenergic receptor;Sympathetic nervoussystem;Neurotransmitterrelease;Dexmedetomidine;Sympathetic ganglionneuron
oa
S
D
insmpdmPpn
Abnormal function of the sympathetic nervous systemSNS; dysautonomia) can be found in many aspects oferioperative medicine, from pre-operative evaluation ofhe patient, intraoperative changes in vital signs, to abnor-al SNS function in the intensive care unit (ICU). Release
f norepinephrine from sympathetic neurons is modulatedy �2a and/or �2c adrenergic receptors (�2a&c ARs) lo-ated at nerve terminals (ie, presynaptic autoreceptors).1-4
n addition, naturally occurring variant genetic forms of2a&c ARs have also been identified and linked to disor-ers of the SNS, such as congestive heart failure and chronicain, although the abnormal physiological function of theseariant forms has not been completely characterized.5-7
�2a&c AR subtypes, and their variant forms, may haveverlapping but unique physiological roles in neuronal sig-aling, although it is not clear as to why two similar auto-eceptors have been maintained in the SNS throughout evo-ution. It is plausible that their differential function may
Address reprint requests and correspondence: Dr. Tim Angelotti, De-artment of Anesthesia, H3580, Stanford University, Stanford, CA 94305.
aE-mail: [email protected].
277-0326/$ -see front matter © 2007 Elsevier Inc. All rights reserved.oi:10.1053/j.sane.2006.11.004
ccur at two molecular levels, such as cellular localizationnd differential modulation of neurotransmitter release.
NS overview
ysregulation and disease
The autonomic nervous system (ANS) operates as anntegrative structure, connecting afferent and efferent sig-aling of multiple vital organs with the central nervousystem. Modulation of the ANS can arise from variousechanisms, including drugs and acute or chronic patho-
hysiological conditions. Primary neurodegenerative disor-ers that involve the ANS include pure autonomic failure,ultiple system atrophy (“Shy-Drager Syndrome”), andarkinson’s Disease, which can manifest as orthostatic hy-otension, bradycardia, and other symptoms of dysauto-omic function.8,9
Secondary dysfunction of the ANS can also occur, as in
utonomic dysreflexia following spinal cord injury, diabetic
nanedwih
dscaacpo
S
a(plrcat
(“
lmNDnoi
�
H
smIsasahAaweATwa
�
np�m�tctKAtncdim
laev
Ftt��M3l
29Hurt and Angelotti �2 Adrenergic Receptor Function
europathy, and certain cancers.10-12 Compensatory alter-tions in sympathetic nervous system function (ie, increasedorepinephrine release) are seen in congestive heart failure,ventually leading to worsening of the disease.13 Acuteisease states can also involve dysfunction of the ANS. Asell, the decrease in heart rate variability seen in critically
ll patients may relate to dysregulation of normal autonomiceart reflexes.14,15
In the perioperative setting, management of sympatheticysfunction can be the major challenge facing an anesthe-iologist. Alterations in blood pressure and heart rate areommon occurrences, and when they occur in the setting ofpatient with cardiovascular co-morbidities, effective man-
gement can be difficult. Although inhalational anestheticsan modulate these functions, direct and indirect actingharmacological agents active on these end organ systemsften are better choices for management.
ympathetic neuron–neuroeffector model
A well-described example of SNS–neuroeffector inter-ction is sympathetic neurons and cardiac myocytesFigure 1).16 Norepinephrine (NE) released from the sym-athetic neuron activates various � adrenergic receptors,eading to changes in cardiac performance. In addition, theelease of NE can activate presynaptic �2a&c ARs to de-rease further release of NE from nerve terminals. Similardrenergic autoreceptors serve as negative feedback loopso regulate SNS activation at most neuroeffector sites.
As feedback regulators of neurotransmitter (NT) releaseeg, epinephrine or norepinephrine), �2a&c ARs act as
igure 1 Schematic of ARs at sympathetic neuron–neuroeffec-or site. �2a&c ARs shown localized to an axonal terminal, wherehey function as autoreceptors to modulate norepinephrine release.
ARs are shown at the neuroeffector cell membrane (ie, Cardiac).2a&c ARs have also been localized to somatodendritic sites.odified from Figure 1 in Hajjar and MacRae: N Engl J Med
47:1196-1199, 2002.16 (Color version of figure is available on-ine.)
brakes” on the SNS, thus playing key roles in the modu- a
ation of sympathetic neurotransmission. For example, ad-inistration of clonidine, an �2 agonist, leads to decreasedT release and thus decreased heart rate and blood pressure.evelopment of �2a&c AR-specific agonists and antago-ists may lead to new therapeutic options for the treatmentf a variety of disorders of dysregulated sympathetic activ-ty.
2 AR pharmacology and physiology
istorical perspective
Current adrenergic receptor classifications have evolvedince the late 1940s, from physiologic responses to phar-acologic binding profiles with relatively selective ligands.
n 1949, Ahlquist proposed that physiological effects ofynthetic adrenaline were mediated by an activating (� AR)nd an inhibiting (� AR) receptor. This concept was sub-equently modified due to the finding that the � adrenergicgonist isoproterenol had both activating (eg, increasedeart rate) and inhibiting (eg, bronchial relaxation) effects.s more physiologic responses were found for adrenergic
gents, a classification system based on anatomic locationas attempted. This scheme proposed that the �1 ARs were
xcitatory and located in postsynaptic tissue, whereas �2Rs were inhibitory and located in presynaptic neurons.his classification was subsequently found to be lackingith the development of ligands that showed that “presyn-
ptic” �2 ARs were found outside synaptic locations.
2 AR molecular biology
With the development of molecular cloning tech-iques,17 the �2 AR family has been subdivided into fourharmacologically distinct subtypes: �2a, �2b, �2c, and2d. However, molecular cloning has revealed that mostammalian species possess only three genes encoding �2b,2c, and either �2a or �2d ARs. This third gene is now
ermed �2a/d (termed �2a for this review). �2a&c ARsouple to pertussis toxin-sensitive Gi/Go signaling pathwayso inhibit cAMP production as well as to regulate Ca2� and
� channel signaling.18 Even following the cloning of �2R subtypes and their expression in heterologous cell sys-
ems, no highly specific and sensitive pharmacologic ago-ist for any �2 AR subtypes has been identified. Molecularloning of murine �2a&c AR genes revealed structuraleterminants of differential agonist and antagonist bind-ng19 and led to the development of gene knockout (KO)odels for in vivo and in vitro physiological research.20,21
These various studies with KO mice have aided in de-ineating the different roles that �2a&c ARs play in centralnd peripheral SNS function mediated by endogenous cat-cholamines and other drugs.22 For example, agonist acti-ation of centrally located �2a ARs leads to decreased SNS
ctivation and thus decreased blood pressure. However, �2c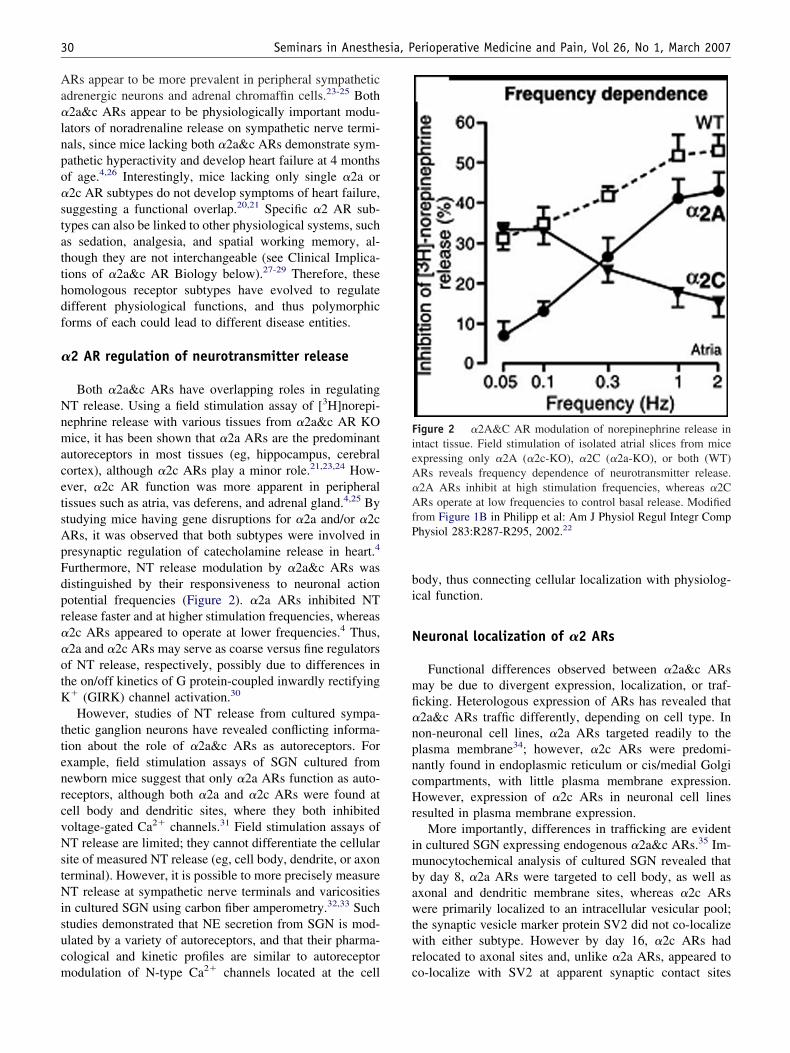
Aa�lnpo�statthdf
�
NnmacetsApFdpr��otK
ttenrcvNstNisucm
bi
N
mfi�npncHr
imbawtwr
FieA�AfP
30 Seminars in Anesthesia, Perioperative Medicine and Pain, Vol 26, No 1, March 2007
Rs appear to be more prevalent in peripheral sympatheticdrenergic neurons and adrenal chromaffin cells.23-25 Both2a&c ARs appear to be physiologically important modu-
ators of noradrenaline release on sympathetic nerve termi-als, since mice lacking both �2a&c ARs demonstrate sym-athetic hyperactivity and develop heart failure at 4 monthsf age.4,26 Interestingly, mice lacking only single �2a or2c AR subtypes do not develop symptoms of heart failure,uggesting a functional overlap.20,21 Specific �2 AR sub-ypes can also be linked to other physiological systems, suchs sedation, analgesia, and spatial working memory, al-hough they are not interchangeable (see Clinical Implica-ions of �2a&c AR Biology below).27-29 Therefore, theseomologous receptor subtypes have evolved to regulateifferent physiological functions, and thus polymorphicorms of each could lead to different disease entities.
2 AR regulation of neurotransmitter release
Both �2a&c ARs have overlapping roles in regulatingT release. Using a field stimulation assay of [3H]norepi-ephrine release with various tissues from �2a&c AR KOice, it has been shown that �2a ARs are the predominant
utoreceptors in most tissues (eg, hippocampus, cerebralortex), although �2c ARs play a minor role.21,23,24 How-ver, �2c AR function was more apparent in peripheralissues such as atria, vas deferens, and adrenal gland.4,25 Bytudying mice having gene disruptions for �2a and/or �2cRs, it was observed that both subtypes were involved inresynaptic regulation of catecholamine release in heart.4
urthermore, NT release modulation by �2a&c ARs wasistinguished by their responsiveness to neuronal actionotential frequencies (Figure 2). �2a ARs inhibited NTelease faster and at higher stimulation frequencies, whereas2c ARs appeared to operate at lower frequencies.4 Thus,2a and �2c ARs may serve as coarse versus fine regulatorsf NT release, respectively, possibly due to differences inhe on/off kinetics of G protein-coupled inwardly rectifying
� (GIRK) channel activation.30
However, studies of NT release from cultured sympa-hetic ganglion neurons have revealed conflicting informa-ion about the role of �2a&c ARs as autoreceptors. Forxample, field stimulation assays of SGN cultured fromewborn mice suggest that only �2a ARs function as auto-eceptors, although both �2a and �2c ARs were found atell body and dendritic sites, where they both inhibitedoltage-gated Ca2� channels.31 Field stimulation assays ofT release are limited; they cannot differentiate the cellular
ite of measured NT release (eg, cell body, dendrite, or axonerminal). However, it is possible to more precisely measureT release at sympathetic nerve terminals and varicosities
n cultured SGN using carbon fiber amperometry.32,33 Suchtudies demonstrated that NE secretion from SGN is mod-lated by a variety of autoreceptors, and that their pharma-ological and kinetic profiles are similar to autoreceptor
odulation of N-type Ca2� channels located at the cell cody, thus connecting cellular localization with physiolog-cal function.
euronal localization of �2 ARs
Functional differences observed between �2a&c ARsay be due to divergent expression, localization, or traf-cking. Heterologous expression of ARs has revealed that2a&c ARs traffic differently, depending on cell type. Inon-neuronal cell lines, �2a ARs targeted readily to thelasma membrane34; however, �2c ARs were predomi-antly found in endoplasmic reticulum or cis/medial Golgiompartments, with little plasma membrane expression.owever, expression of �2c ARs in neuronal cell lines
esulted in plasma membrane expression.More importantly, differences in trafficking are evident
n cultured SGN expressing endogenous �2a&c ARs.35 Im-unocytochemical analysis of cultured SGN revealed that
y day 8, �2a ARs were targeted to cell body, as well asxonal and dendritic membrane sites, whereas �2c ARsere primarily localized to an intracellular vesicular pool;
he synaptic vesicle marker protein SV2 did not co-localizeith either subtype. However by day 16, �2c ARs had
elocated to axonal sites and, unlike �2a ARs, appeared to
igure 2 �2A&C AR modulation of norepinephrine release inntact tissue. Field stimulation of isolated atrial slices from micexpressing only �2A (�2c-KO), �2C (�2a-KO), or both (WT)Rs reveals frequency dependence of neurotransmitter release.2A ARs inhibit at high stimulation frequencies, whereas �2CRs operate at low frequencies to control basal release. Modified
rom Figure 1B in Philipp et al: Am J Physiol Regul Integr Comphysiol 283:R287-R295, 2002.22
o-localize with SV2 at apparent synaptic contact sites

(ttc
snsotftanfA
C
�
AtahpApA�h
hretdktep
�
tyvhsfs�cfoake
sD�i
Ffca harma
31Hurt and Angelotti �2 Adrenergic Receptor Function
Figure 3). The differences in SGN targeting observed be-ween these two autoreceptors correlated with their abilityo modulate neuronal function, as measured by regulation ofalcium ([Ca2�]i) influx.
Differential �2a&c AR trafficking seen in SGN wasimilar in context to that observed in neuronal versus non-euronal cell lines, respectively. Namely, that a neuronal-pecific effect is necessary for proper membrane expressionf �2c ARs, and it appears that this occurs during matura-ion of SGN cultures. This result may account for the dif-erences seen in earlier SGN cultures when compared withissue studies. It is possible that �2c AR localization atpparent synaptic sites partially explains their responsive-ess to low frequency neuron stimulation and higher affinityor NE4; they reside closer to the site of NT release than �2aRs.
linical implications of �2a&c AR biology
2a&c ARs and hemodynamic control
Blood pressure homeostasis is regulated in part by �2Rs located within the central and peripheral nervous sys-
ems, as well as end organs. Non-�2 AR subtype selectivectivation by agonist leads to a biphasic response, a briefypertensive period followed by a prolonged hypotensivehase. The initial hypertensive phase is mediated by �2bRs located on vascular smooth muscle, whereas the hy-otensive phase is mediated by centrally located �2aRs.21,36 The latter response is the basis for the search for2a AR subtype selective agents as possible new anti-
igure 3 Co-localization of �2A&C ARs and SV2 in Cultured Sor �2a (A) and �2c (D) ARs and the synaptic vesicle protein So-localization of �2a ARs and SV2 (C), but the presence of co-lrrows (F). Modified from Figures 6 and 7 in Brum et al: Neurop
ypertensive agents. Unfortunately, it appears that the anti- m
ypertensive effect of �2a AR activation cannot be sepa-ated from its sedative effects. Although �2c ARs arexpressed exclusively in adrenal chromaffin cells, no sub-ype-specific drugs exist for them, and the exact role suchrugs would play in blood pressure regulation is notnown.25 However, it has been suggested that abnormalrafficking of �2c ARs at cold temperatures may partlyxplain the peripheral vasoconstriction seen in Raynaud’shenomenon.37
2 AR regulation of analgesia and sedation
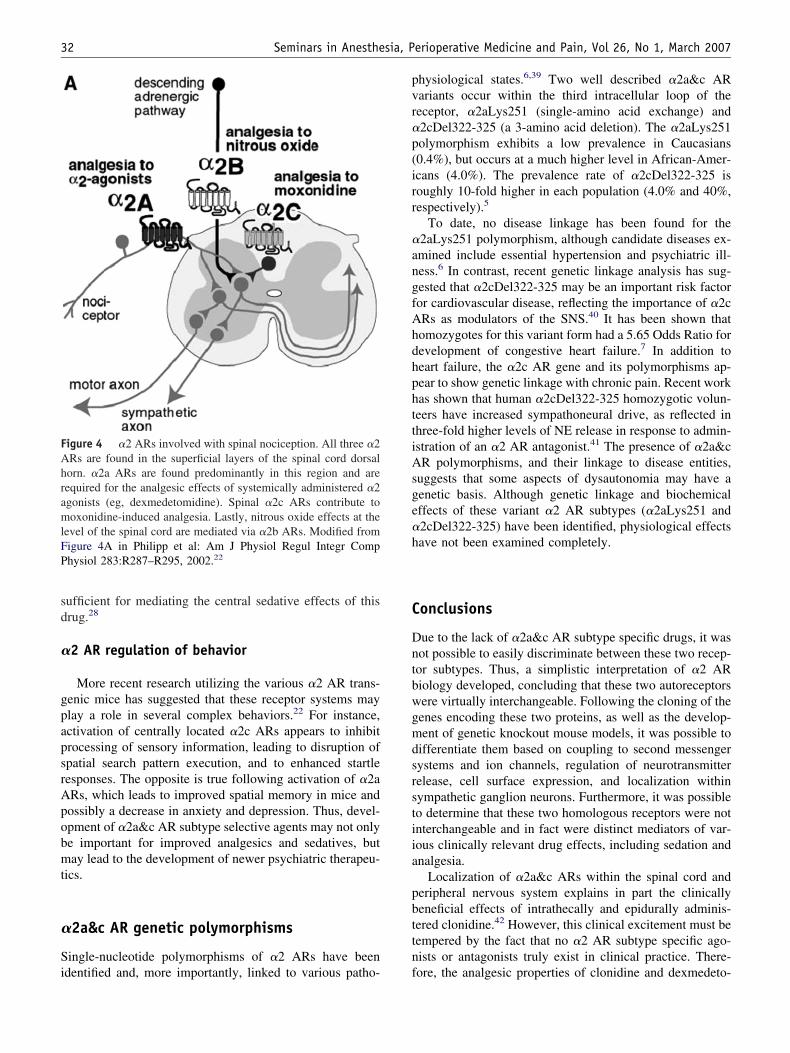
�2 AR agonists have been proven to be potent analgesicshat can potentiate the effects of opioids. In addition, anal-sis of various �2 AR KO mouse models has revealed thatarious other anesthetic agents also possess sedative andypnotic effects that are due to activation of various �2 ARubtypes (Figure 4). For instance, �2a ARs are necessaryor the antinociception effects of various �2 AR agonists,uch as clonidine and dexmedetomidine.27 However, mixed2 AR/imidazoline receptor agonists, such as moxonidine,ause spinal antinociception that requires spinal �2c ARsor their effect.38 Lastly, the potent inhaled analgesic nitrousxide requires spinal �2b ARs to mediate a supraspinalnalgesic pathway via the dorsal horn of the spinal cord. Innockout mice lacking �2b ARs, this descending noradren-rgic pathway has been abolished.
Dexmedetomidine is commonly used in ICUs as a potentedative, and with increasing frequency in operating rooms.exmedetomidine is approximately 1600x more potent at2 over �1 ARs (compared to 200x for clonidine); however,
t is not subtype selective. Analysis of various �2 AR KO
ixteen-day-old SGN cultures were labeled with antibodies specific, E). Insets denoted by boxes were merged. Note the absence ofd �2c ARs and SV2 immunostaining (yellow), denoted by whitecology 51:397-413, 2006.35
GN. SV2 (Bocalize
ice revealed that the �2a AR subtype is necessary and
sd
�
gpapsrApobmt
�
Si
pvr�p(irr
�angfAhdhphttiAsge�h
C
Dntbwgmdsrstiia
pbttn
FAhramlFP
32 Seminars in Anesthesia, Perioperative Medicine and Pain, Vol 26, No 1, March 2007
ufficient for mediating the central sedative effects of thisrug.28
2 AR regulation of behavior
More recent research utilizing the various �2 AR trans-enic mice has suggested that these receptor systems maylay a role in several complex behaviors.22 For instance,ctivation of centrally located �2c ARs appears to inhibitrocessing of sensory information, leading to disruption ofpatial search pattern execution, and to enhanced startleesponses. The opposite is true following activation of �2aRs, which leads to improved spatial memory in mice andossibly a decrease in anxiety and depression. Thus, devel-pment of �2a&c AR subtype selective agents may not onlye important for improved analgesics and sedatives, butay lead to the development of newer psychiatric therapeu-
ics.
2a&c AR genetic polymorphisms
ingle-nucleotide polymorphisms of �2 ARs have been
igure 4 �2 ARs involved with spinal nociception. All three �2Rs are found in the superficial layers of the spinal cord dorsalorn. �2a ARs are found predominantly in this region and areequired for the analgesic effects of systemically administered �2gonists (eg, dexmedetomidine). Spinal �2c ARs contribute tooxonidine-induced analgesia. Lastly, nitrous oxide effects at the
evel of the spinal cord are mediated via �2b ARs. Modified fromigure 4A in Philipp et al: Am J Physiol Regul Integr Comphysiol 283:R287–R295, 2002.22
dentified and, more importantly, linked to various patho- f
hysiological states.6,39 Two well described �2a&c ARariants occur within the third intracellular loop of theeceptor, �2aLys251 (single-amino acid exchange) and2cDel322-325 (a 3-amino acid deletion). The �2aLys251olymorphism exhibits a low prevalence in Caucasians0.4%), but occurs at a much higher level in African-Amer-cans (4.0%). The prevalence rate of �2cDel322-325 isoughly 10-fold higher in each population (4.0% and 40%,espectively).5
To date, no disease linkage has been found for the2aLys251 polymorphism, although candidate diseases ex-mined include essential hypertension and psychiatric ill-ess.6 In contrast, recent genetic linkage analysis has sug-ested that �2cDel322-325 may be an important risk factoror cardiovascular disease, reflecting the importance of �2cRs as modulators of the SNS.40 It has been shown thatomozygotes for this variant form had a 5.65 Odds Ratio forevelopment of congestive heart failure.7 In addition toeart failure, the �2c AR gene and its polymorphisms ap-ear to show genetic linkage with chronic pain. Recent workas shown that human �2cDel322-325 homozygotic volun-eers have increased sympathoneural drive, as reflected inhree-fold higher levels of NE release in response to admin-stration of an �2 AR antagonist.41 The presence of �2a&cR polymorphisms, and their linkage to disease entities,
uggests that some aspects of dysautonomia may have aenetic basis. Although genetic linkage and biochemicalffects of these variant �2 AR subtypes (�2aLys251 and2cDel322-325) have been identified, physiological effectsave not been examined completely.
onclusions
ue to the lack of �2a&c AR subtype specific drugs, it wasot possible to easily discriminate between these two recep-or subtypes. Thus, a simplistic interpretation of �2 ARiology developed, concluding that these two autoreceptorsere virtually interchangeable. Following the cloning of theenes encoding these two proteins, as well as the develop-ent of genetic knockout mouse models, it was possible to
ifferentiate them based on coupling to second messengerystems and ion channels, regulation of neurotransmitterelease, cell surface expression, and localization withinympathetic ganglion neurons. Furthermore, it was possibleo determine that these two homologous receptors were notnterchangeable and in fact were distinct mediators of var-ous clinically relevant drug effects, including sedation andnalgesia.
Localization of �2a&c ARs within the spinal cord anderipheral nervous system explains in part the clinicallyeneficial effects of intrathecally and epidurally adminis-ered clonidine.42 However, this clinical excitement must beempered by the fact that no �2 AR subtype specific ago-ists or antagonists truly exist in clinical practice. There-
ore, the analgesic properties of clonidine and dexmedeto-
msapnmactiet
R
1
11
1
1
1
1
1
1
1
2
2
2
2
2
2
2
2
2
2
3
3
3
3
3
3
3
3
3
3
4
33Hurt and Angelotti �2 Adrenergic Receptor Function
idine will always be accompanied by a dose-limitingedative side effect. Interestingly, various �2 AR agonistsre utilized in veterinary medicine to induce large animalsrior to induction of anesthesia, followed by reversal withon-subtype selective �2 AR antagonists.43 The develop-ent of paired �2a&c AR subtype selective agonists and
ntagonists would be beneficial for anesthetic and criticalare needs. Lastly, future translational research directedoward understanding the role of �2a&c AR polymorphismsn perioperative and intensive care medicine may help tolucidate the variability seen amongst patients with respecto autonomic nervous system function and dysfunction.
eferences
1. Rosin DL, Zeng D, Stornetta RL, et al: Immunohistochemical local-ization of alpha 2A-adrenergic receptors in catecholaminergic andother brainstem neurons in the rat. Neuroscience 56:139-155, 1993
2. Rosin DL, Talley EM, Lee A, et al: Distribution of alpha 2C-adren-ergic receptor-like immunoreactivity in the rat central nervous system.J Comp Neurol 372:135-165, 1996
3. Trendelenburg AU, Cox SL, Gaiser EG, et al: Noradrenaline releasefrom cultured mouse postganglionic sympathetic neurons: autorecep-tor-mediated modulation. J Neurochem 73:1439-1445, 1999
4. Hein L, Altman JD, Kobilka BK: Two functionally distinct alpha2-adrenergic receptors regulate sympathetic neurotransmission. Nature402:181-184, 1999
5. Small KM, Liggett SB: Identification and functional characterizationof alpha(2)-adrenoceptor polymorphisms. Trends Pharmacol Sci 22:471-477, 2001
6. Garland EM, Biaggioni I: Genetic polymorphisms of adrenergic re-ceptors. Clin Auton Res 11:67-78, 2001
7. Small KM, Wagoner LE, Levin AM, et al: Synergistic polymorphismsof beta1- and alpha2C-adrenergic receptors and the risk of congestiveheart failure. N Engl J Med 347:1135-1142, 2002
8. Mathias CJ, Polinsky RJ: Separating the primary autonomic failuresyndromes, multiple system atrophy, and pure autonomic failure fromParkinson’s disease. Adv Neurol 80:353-361, 1999
9. Chaudhuri KR: Autonomic dysfunction in movement disorders. CurrOpin Neurol 14:505-511, 2001
0. Mathias CJ: Disorders of the autonomic nervous system, in Bradley W,Daroff R, Fenichel G, et al (eds): Neurology in Clinical Practice (ed 3).Boston, MA, Butterworth-Heinemann, 2000, pp 2131-2165
1. Karlsson AK: Autonomic dysreflexia. Spinal Cord 37:383-391, 19992. Greene DA, Stevens MJ, Feldman EL: Diabetic neuropathy: scope of
the syndrome. Am J Med 107:2S-8S, 19993. Cohn JN: The management of chronic heart failure. N Engl J Med
335:490-498, 19964. Schmidt HB, Werdan K, Muller-Werdan U: Autonomic dysfunction in
the ICU patient. Curr Opin Crit Care 7:314-322, 20015. Gang Y, Malik M: Heart rate variability in critical care medicine. Curr
Opin Crit Care 8:371-375, 20026. Hajjar RJ, MacRae CA: Adrenergic-receptor polymorphisms and heart
failure. N Engl J Med 347:1196-1199, 20027. Bylund DB, Eikenberg DC, Hieble JP, et al: International Union of
Pharmacology nomenclature of adrenoceptors. Pharmacol Rev 46:121-136, 1994
8. MacDonald E, Kobilka BK, Scheinin M: Gene targeting–homing in onalpha 2-adrenoceptor-subtype function. Trends Pharmacol Sci 18:211-219, 1997
9. Link R, Daunt D, Barsh G, et al: Cloning of two mouse genes encodingalpha 2-adrenergic receptor subtypes and identification of a single aminoacid in the mouse alpha 2-C10 homolog responsible for an interspecies
variation in antagonist binding. Mol Pharmacol 42:16-27, 19920. Link RE, Stevens MS, Kulatunga M, et al: Targeted inactivation of thegene encoding the mouse alpha 2c-adrenoceptor homolog. Mol Phar-macol 48:48-55, 1995
1. Altman JD, Trendelenburg AU, MacMillan L, et al: Abnormal regu-lation of the sympathetic nervous system in alpha2A- adrenergic re-ceptor knockout mice. Mol Pharmacol 56:154-161, 1999
2. Philipp M, Brede M, Hein L: Physiological significance of alpha(2)-adrenergic receptor subtype diversity: one receptor is not enough.Am J Physiol Regul Integr Comp Physiol 283:R287-R295, 2002
3. Trendelenburg AU, Hein L, Gaiser EG, et al: Occurrence, pharmacol-ogy and function of presynaptic alpha2-autoreceptors in alpha2A/D-adrenoceptor-deficient mice. Naunyn Schmiedebergs Arch Pharmacol360:540-551, 1999
4. Trendelenburg AU, Klebroff W, Hein L, et al: A study of presynapticalpha2-autoreceptors in alpha2A/D-, alpha2B- and alpha2C-adreno-ceptor-deficient mice. Naunyn Schmiedebergs Arch Pharmacol 364:117-130, 2001
5. Brede M, Nagy G, Philipp M, et al: Differential control of adrenal andsympathetic catecholamine release by alpha 2-adrenoceptor subtypes.Mol Endocrinol 17:1640-1646, 2003
6. Brum PC, Kosek J, Patterson A, et al: Abnormal cardiac functionassociated with sympathetic nervous system hyperactivity in mice.Am J Physiol Heart Circ Physiol 283:H1838-H1845, 2002
7. Stone LS, MacMillan LB, Kitto KF, et al: The alpha2a adrenergicreceptor subtype mediates spinal analgesia evoked by alpha2 agonistsand is necessary for spinal adrenergic-opioid synergy. J Neurosci17:7157-7165, 1997
8. Lakhlani PP, MacMillan LB, Guo TZ, et al: Substitution of a mutantalpha2a-adrenergic receptor via “hit and run” gene targeting revealsthe role of this subtype in sedative, analgesic, and anesthetic-sparingresponses in vivo. Proc Natl Acad Sci U S A 94:9950-9955, 1997
9. Scheinin M, Sallinen J, Haapalinna A: Evaluation of the alpha2C-adrenoceptor as a neuropsychiatric drug target studies in transgenicmouse models. Life Sci 68:2277-2285, 2001
0. Bunemann M, Bucheler MM, Philipp M, et al: Activation and deac-tivation kinetics of alpha 2A- and alpha 2C-adrenergic receptor-acti-vated G protein-activated inwardly rectifying K� channel currents.J Biol Chem 276:47512-47517, 2001
1. Trendelenburg AU, Norenberg W, Hein L, et al: Alpha2-adrenoceptor-mediated inhibition of cultured sympathetic neurons: changes inalpha2A/D-adrenoceptor-deficient mice. Naunyn Schmiedebergs ArchPharmacol 363:110-119, 2001
2. Koh DS, Hille B: Modulation by neurotransmitters of catecholaminesecretion from sympathetic ganglion neurons detected by amperom-etry. Proc Natl Acad Sci U S A 94:1506-1511, 1997
3. Travis ER, Wightman RM: Spatio-temporal resolution of exocytosis fromindividual cells. Annu Rev Biophys Biomol Struct 27:77-103, 1998
4. Daunt DA, Hurt C, Hein L, et al: Subtype-specific intracellular traffickingof alpha2-adrenergic receptors. Mol Pharmacol 51:711-720, 1997
5. Brum PC, Hurt CM, Shcherbakova OG, et al: Differential targeting andfunction of alpha(2A) and alpha(2C) adrenergic receptor subtypes incultured sympathetic neurons. Neuropharmacology 51:397-413, 2006
6. Link RE, Desai K, Hein L, et al: Cardiovascular regulation in micelacking alpha2-adrenergic receptor subtypes b and c. Science 273:803-805, 1996
7. Jeyaraj SC, Chotani MA, Mitra S, et al: Cooling evokes redistributionof alpha2C-adrenoceptors from Golgi to plasma membrane in trans-fected human embryonic kidney 293 cells. Mol Pharmacol 60:1195-1200, 2001
8. Fairbanks CA, Stone LS, Kitto KF, et al: alpha(2C)-Adrenergic recep-tors mediate spinal analgesia and adrenergic-opioid synergy. J Phar-macol Exp Ther 300:282-290, 2002
9. Zaugg M, Schaub MC: Genetic modulation of adrenergic activity inthe heart and vasculature: implications for perioperative medicine.Anesthesiology 102:429-446, 2005
0. Small KM, Mialet-Perez J, Seman CA, et al: Polymorphisms of car-
diac presynaptic alpha2C adrenergic receptors: Diverse intragenic
4
4
4
34 Seminars in Anesthesia, Perioperative Medicine and Pain, Vol 26, No 1, March 2007
variability with haplotype-specific functional effects. Proc Natl AcadSci U S A 101:13020-13025, 2004
1. Neumeister A, Charney D, Belfer I, et al: Sympathoneural andadrenomedullary functional effects of alpha2C-adrenoreceptor genepolymorphism in healthy humans. Pharmacogenetics 15:143-150,
20052. Eisenach JC, De Kock M, Klimscha W: alpha(2)-adrenergic agonistsfor regional anesthesia. A clinical review of clonidine (1984-1995).Anesthesiology 85:655-674, 1996
3. Sinclair MD: A review of the physiological effects of alpha2-agonistsrelated to the clinical use of medetomidine in small animal practice.
The Canadian veterinary journal 44:885-897, 2003













![FOSTER, MICHAEL D., M.S. Computational Study of RTI ......adrenergic receptor ( β2-AR) [6-8], β1-adrenergic receptor ( β1-AR) [9], adenosine A2A receptor [10] and most recently](https://img.pdfslide.net/doc/110x75/607b302caf43ed024c5d3e7b/foster-michael-d-ms-computational-study-of-rti-adrenergic-receptor.jpg)














