Embed Size (px)
Citation preview

JOURNAL OF PATHOLOGY, VOL. 178: 369-378 (1996)
REVIEW ARTICLE
MOLECULAR PATHOBIOLOGY AND IMMUNOLOGY OF MALIGNANT MESOTHELIOMA
H. BIELEFELDT-OHMANN", A. G. JARNICKI~ AND D. R. FITZPATRICK~
*Centre for Molecular Biotechnology, School of Life Science, Queensland University of Technology, Brisbane, Australia; ?Institute f o r Child Health Research, Subiaco, Western Australia; $Transplantation Biology Unit, Queensland Institute of Medical Research,
Herston, Queensland, Australia
INTRODUCTION
The mesothelium, a mesodermally derived single-layer epithelium, lines the pleural, pericardial, and peritoneal cavities, where it functions as a non-adhesive surface for the internal organs and as a selective barrier that regu- lates the transport of molecules and cells between the circulation and the body cavities. The mesothelium is supported by a layer of connective tissue consisting of collagen and elastin fibres interspersed with fibroblasts, mononuclear phagocytes, lymphocytes, capillaries, and lymphatics. The mesothelial cells are the progenitor cells of mesothelioma, a malignant neoplasm often associated with past exposure to asbestos; this accounts for approximately 50 per cent of all cases, the remainder being of unknown aetiology.'-3 Malignant meso- thelioma presents two particularly important problems, its intractable character and its difficult differential diag- nosis. The latter problem is now being tackled by a multi-method approach involving conventional histo- pathology, electron microscopy, and immunocytochem- istry with the application of a range of monoclonal antibodies to cellular antigen^.^ Progress in solving the former problem has, however, been relatively slow. Conventional approaches to the treatment of meso- thelioma have been largely unsuccessful, and newer immunotherapeutic a proaches have yet to have any impact on prognosis.'This may be partly explained by the lack of comprehensive knowledge of the pathogen- esis and immunobiology of mesothelioma which would assist the rational design of new therapeutic strategies.
The main focus of this review will be to discuss recent advances in our understanding of the role of endogenous cytokines and tumour infiltrating leukocytes in the pathophysiology and immunology of malignant meso- thelioma, based on studies in human patients and in animal models. The relevance of these findings for an improvement of the design of therapeutic strategies will also be discussed.
Addressee for correspondence: Dr H. Bielefeldt-Ohmann, Centre for Molecular Biotechnology, School of Life Science, Queensland University of Technology, GPO Box 2434, 2 George Street, Brisbane, Queensland 4001, Australia.
CCC 0022-341 7/96/040369-10 1996 by John Wiley & Sons, Ltd.
PATHOGENESIS OF MALIGNANT MESOTHELIOMA
In the current consensus model of the pathogenesis of mesothelioma, it is suggested that neoplastic growth is a result of two concurrent, perhaps mutually reinforcing processes: firstly, genetic changes in the mesothelial cells, whereby their responsiveness to autocrine and paracrine growth signals is altered, and, secondly, a chronic inflammatory response. Either or both may be induced by mineral fibres of the appropriate shape and chemical composition or by other carcinogenic/pro-inflammatory agents.&' To this model may be added a third process, immuno-evasion' (Fig. l), as discussed later.
Cytogenetic changes in known and putative tumour suppressor genes have been reported in a proportion of mesotheliomas,8,'0 although it is possible that even the relatively low frequencies reported have been overesti- mated due to extensive inferences from results obtained with established mesothelioma cell lines rather than primary tumours. Furthermore, the tumour sup- pressor genes most commonly altered in other human malignancies, including p53 and Rb, are only rarely mutated in malignant mesothelioma.' Activation of cellular oncogenes in mesothelioma development has received some attention, but again the most common oncogenes, e.g., rus, are rarely mutated." Heintz et ~ 1 . ' ~ showed that asbestos can induce c-fos and c-jun mRNA in mesothelial cells in a dose-dependent manner and proposed that by chronic stimulation of cell prolifera- tion, fixation of genetic damage caused by asbestos itself or other agents may eventually culminate in the selection of neoplastic cells. This may be reinforced by the selec- tive resistance of mesothelioma cells to direct mineral fibre t0x i~ i ty . l~
The inflammatory component of the hypothesized multi-stage cell carcinogenesis is even more elusive than the role played by chromosomal damagekhanges. Normal mesothelial cells have been shown to produce spontaneously a range of cytokines and growth factors in vitro, with up-regulation of these and additional molecular species, including interleukin-8 (IL-S), macro- phage inflammatory protein-1 (MIP-l), IL-lu//?, IL-6, and macrophage-, granulocyte-, and granulocyte- macrophage-colony stimulating factors (M-, GP-, and GM-CSF, respectively), following in vitro exposure to
Accepted 14 July 1995

370 H. BIELEFELDT-OHMANN ET AL.
1 Phagocytosis ~ Translocation of Fibres ~ by Macrophages 1
I Chemoattraction
& Stimulation 1 - ______-
Mesothelial Injury & ~ Chromosomal Changes ROS, NO, TGF-B, 1 -
I PDGF, IGF, etc. I Increased Responsiveness
Angiogenesis to Growth Factors Fibrosis (ECM) Growth Stimulation
~~~
__-
m c k e d Growth of Transformed Cells
I
& Deactivation 1 _____
Anti-Tumour / ~ Autocrine Cytokine I Production & ,
Effector Functions b I Immune Evasion 1 ._.____ - ~ ~~
Fig. I-A model for the pathogenesis of mesothelioma: after translocation of fibres from the lung to a serosal cavity, macrophages make abortive attempts to phagocytose the long fibres and in the process release reactive oxygen species (ROS), nitric oxide (NO), and a range of cytokines, notably TGFP. This cytokine will attract more macrophages to the site, as well as stimulate angiogenesis and the formation of extracellular matrix (ECM). The ROS, NO, and pcrhaps asbestos fibres cause direct mesothelial injury, as well as DNA injury. Chronic stimulation of cell proliferation (TGFP, PDGF, bFGF, etc.) leads to the fixation of the genetic damage. The inflammatory milieu will attract lymphocytes, which, however, soon become deactivated and induction of anti-tumour immune responses is therefore aborted. Collectively, these processes lead to unchecked growth of transformed cells and clinical malignant mesothelioma
mineral fibres or macrophage-derived c tokines [e.g., tumour necrosis factor-a (TNFa), IL-1].6, Macrophages specifically bind asbestos fibres in vitro,14 followed by phagocytosis and induction of cytokine release, including insulin-dependent growth factor-1 (IGF-I), transforming growth factor$ (TGFB, platelet- derived rowth factor (PDGF), TNFa, and interleukin- 1 (IL-l).1F16 In vivo studies in various animal species,
Y in which large doses of asbestos were administered intrapleurally, intraperitoneally, intratracheally, or by inhalation, demonstrated acute inflammatory reac- tions with focal mesothelioma cell death, regeneration and angiogenesis, haemorrha e and accumulation of macrophages and fibroblasts. ''"I However, so far there have been no reports of in situ localization of any of the above cytokines following experimental or 'natural'

PATHOBIOLOGY OF MALIGNANT MESOTHELIOMA 37 1
asbestos exposure. It is important to emphasize that cells may respond differently in vitro when taken out of context with extracellular matrix and fluid, and surrounding cell types.22 Likewise, the large doses of asbestos fibres used in both in vitro and in vivo studies, frequently given as a single bolus, may not reflect the natural history of asbestos-induced mesothelioma.
The preclinical phases of malignant mesothelioma in humans are essentially unknown and currently un- diagnosable. Cases of benign asbestos-induced pleural effusion or ascites formation have been described, but these did not appear to precede malignant meso- t h e l i ~ m a . ~ ~ Pleural fibrotic plaques have received much attention and although a recent rigorous study would suggest that patients with pleural plaques have a higher risk of developing malignant mesothelioma, it was still an extremely small proportion of patients who did The fact that mesothelioma does develop in individuals with only brief and long-since-discontinued exposure suggests either that asbestos fibres continue to provoke a response, chromosomal and inflam- matory, throughout the long latency period, or that asbestos fibres have the ability to initiate a subse- quently self-sustaining inflammation. The pathogenesis of asbestos-unrelated mesothelioma also remains completely unknown, perhaps with the exception of the one suspected unifying component: chronic inf lammati~n.~,~’
PATHOLOGY: NEOPLASTIC AND PARANEOPLASTIC
The most common site of origin of mesothelioma is the pleura, with peritoneal mesothelioma second in frequency, although it may be added that the frequent diagnosis of the latter in certain geographical areas is now being questioned.26 Mesotheliomas are classified histologically as either epithelial, sarcomatous, or mixed/ b i p h a ~ i c , ~ ~ but the epithelial type especially can be difficult to distinguish from adenocarcinoma. Reactive, hyperplastic mesothelium may also, if entrapped within scar tissue, resemble biphasic mesothelioma. Table I summarizes some of the key features of the pathology of mesothelioma in humans and compares these with the pathological features of experimental mesothelioma in two rodent specie^.'^^^-^^ Comprehensive reviews of the pathology of malignant mesothelioma in humans have appeared recently.27T31 Unfortunately, missing from most histopathological descriptions of malignant meso- thelioma is an analysis of the degree of vascularization, fibrosis, necrosis, or leucocyte infiltration. This is regret- table since such data, as will be discussed below, might assist in the understanding of the relative involvement of various growth factors and cytokines in the patho- biology of mes~thelioma,~ as well as providing possible prognostic i n f ~ r m a t i o n . ’ ~ , ~ ~
Studies in a murine model of mesothelioma demon- strated that specific cytokines produced and/or induced by mesothelioma cells may be intimately involved in most of the athology and pathophysiology of this malignancy. Table I1 summarizes the functional 9 , B
characteristics, including pathological effects, of key cytokines/growth factors known or surmised to play a role in me~othelioma.”~ In mice inoculated intra- peritoneally with asbestos or syngeneic TGFP-producing mesothelioma cells (‘Pattern 1’ in Table I), there was nodular growth of mixed-type tumours with a substan- tial fibrotic stromal component. Vascularization was moderate and necrosis widespread.’ Typically, an inflammatory response was lacking from the necrotic foci, despite massive macrophage infiltration in the tumours. Both the fibrosis and the macrophage infiltra- tion may be TGFP-ir~duced,~~ while the tumour necrosis could, at least in part, be caused by excessively high levels of TNFa in the ascitic and interstitial fluids. TNFa was also considered the likely inducer of acute coagulation-necrosis of liver parenchyma and terminal hypovolaemic shock.’ The source of TNFa is not known, but tumour infiltrating macrophages (TIM) and macrophages accumulating in the ascitic fluid (see below) are likely candidates, stimulated by high levels of bioactive TGFP in the local mic ro -en~ i ronmen t .~~’~~
In mice inoculated with mesothelioma cells which produce high levels of IL-6 but not TGFP, the atho- logical features differed (‘Pattern 2’ in Table I). The tumours tended to grow as a large central mass in the mesentery, often invading and obstructing the intestines and other organs. The turnours contained only minimal fibrotic stroma, but appeared well vascularized by thin- walled vessels with a tendency to rupture, associated with haematoma formation. Tumour necrosis was most likely caused by the subsequent ischaemia. The paraneo- plastic pathology in these animals was striking; an early feature was hepatocyte vacuolation, a sign of metabolic disturbance, followed by necrobiosis. Later, the animals developed diarrhoea and severe cachexia; TNFa levels, however, remained low or undetectable throughout the clinical course.
Paraneoplastic syndromes including weight loss, fever, and thrombocytosis are among the more common presenting signs in mesothelioma patients; hepato- megaly is occasionally seen. IL-6 mRNA expression and protein production have been demonstrated by human and murine mesothelioma cells in vivo and in vitro and IL-6-producing mesotheliomas have been linked to thrombocytosis, fever, acute phase reaction, cachexia, and systemic amyloidosis in human patient^.^^-^' Treat- ment of mice carrying IL-6-producing mesotheliomas with anti-IL-6 antibodies curtailed the development of cachexia and liver pathology and extended the survival time without significantly inhibiting tumour develop- ment,30 suggesting that IL-6 plays no direct role in tumour growth per se. The result in the murine meso- thelioma model corroborates the association of IL-6, rather than TNFa, with cachexia in other cancer mod- e l ~ ~ ~ , ~ ~ and emphasizes the potential importance of paraneoplastic events in the a gressive tumourigenesis of malignant mes~thelioma.’~~~’This is referred to again in the concluding remarks on immunotherapy.
The predominant growth pattern of mesotheliomas is by invasion of tissues in the pleural or abdominal cavities, by transdiaphragmatic extension and, more rarely, by lymph node or distant haematogenous
F O

Tabl
e IL
Com
para
tive
pat
holo
gy o
f m
alig
nant
mes
othe
liom
a in
hum
ans
and
rode
nts
Mou
se’.’
’
Hum
an2’
%”
Gro
ss
Smal
l nod
ules
, pla
ques
to
diff
use
shee
t-lik
e th
icke
ning
of
the
pleu
ra
or m
esen
tery
, ofte
n w
ith e
ncas
emen
t of
org
ans,
and
usu
ally
ass
ocia
ted
with
eff
usio
n
Ligh
t mic
rosc
opy
Epith
elia
l mes
othe
liom
a Tu
bulo
papi
llary
or
tubu
lar
form
atio
ns li
ned
by c
uboi
dal o
r el
onga
ted
cells
Sarc
omat
ous
mes
othe
liom
a El
onga
ted
or s
pind
le-s
hape
d ce
lls
with
con
side
rabl
e pl
eom
orph
ism
Bip
hasi
clm
ixed
mes
othe
liom
a M
ixtu
re o
f ep
ithel
ial
and
sarc
omat
ous
grow
th p
atte
rns
in
varia
ble
ratio
s
Ultr
astr
uctu
re
Epith
elia
l com
pone
nt:
Long
mic
rovi
lli, a
bund
ant
inte
rmed
iate
fila
men
ts,
desm
osom
es, a
nd ju
nctio
nal
com
plex
es p
rese
nt.
Intr
acel
lula
r lu
min
a co
mm
on
Sarc
omat
ous
com
pone
nt:
Mic
rovi
lli u
sual
ly a
bsen
t Fi
lam
ents
pre
sent
in s
ome
cells
. D
esm
osom
es u
sual
ly a
bsen
t. M
ay
cont
ain
peri
pher
al a
ctin
fil
amen
ts
Rat28
.2Y
Patte
rn 1
Pa
ttern
2
Gro
ss
Asc
ites
(ofte
n w
ith b
lood
).
Wid
espr
ead,
non
-inva
sive
neo
plas
tic
impl
ants
ove
r pe
riton
eal
surf
aces
. N
o m
etas
tase
s
Ligh
t m
icro
scop
y Ty
pe 1
: rou
nded
epi
thel
ial c
ells
ove
r ce
ntra
l sp
indl
e ce
lls a
nd fi
brot
ic
tissu
e
Type
2: p
apill
ary
with
cel
ls o
f ro
unde
d ep
ithel
ial t
ype
grow
ing
in
solid
col
umns
Type
3: v
esic
ular
gro
wth
line
d by
fla
ttene
d ep
ithel
ial c
ells
. Min
imal
co
nnec
tive
tissu
e el
emen
t
(All
thre
e hi
stol
ogic
al t
ypes
may
be
pres
ent i
n la
rge
tum
ours
)
Ultr
astr
uctu
re
Type
1: c
ells
on
surf
ace
of t
umou
rs
only
loos
ely
conn
ecte
d, w
ith f
ew
desm
osom
es,
and
poss
esse
d lo
ng
mic
rovi
lli. S
pind
le c
ells
in l
oose
‘n
etw
ork’
bet
wee
n re
ticul
in f
ibre
s
Type
2: c
ells
in c
olum
ns fi
rmly
at
tach
ed b
y de
smos
omes
. Cel
ls o
n tu
mou
r su
rfac
e w
ith l
ong
mic
rovi
lli
Type
3: c
ell m
orph
olog
y va
riabl
e Fe
w d
esm
osom
es
Gro
ss
Blo
ody
asci
tes.
N
odul
ar tu
mou
r gr
owth
thr
ough
out
mes
ente
ry a
nd ‘
flesh
y’ pl
aque
-like
tu
mou
r gr
owth
on
diap
hrag
m.
Live
r nec
rose
s.
Thym
ic a
nd s
plen
ic a
trop
hy
Ligh
t mic
rosc
opy
Tum
our
biph
asic
, pre
dom
inan
tly
sarc
omat
ous,
with
ext
ensi
ve fi
bros
is,
low
to m
oder
ate
vasc
ular
izat
ion,
and
w
ides
prea
d ne
cros
is (
inde
pend
ent o
f tu
mou
r si
ze)
Live
r: fo
cal t
o di
ffus
e co
agul
ativ
e ne
cros
is
Thym
us:
cort
ical
and
med
ulla
ry
lym
phoc
yte
depl
etio
n
Ultr
astr
uctu
re
Not
don
e*
Blo
ody
asci
tes.
Si
ngle
larg
e tu
mou
r m
ass
cent
rally
in
mes
ente
ry,
ofte
n w
ith h
aem
atom
as
and
inva
sive
gro
wth
. Li
ver
enla
rged
, dis
colo
ured
. Th
ymic
and
lym
ph n
ode
atro
phy
Tum
our
biph
asic
with
dom
inan
t ep
ithel
ial c
ompo
nent
, m
inim
al
vasc
ular
ized
. Nec
rosi
s va
riab
le
Live
r: he
pato
cyte
vac
uola
tion
and
necr
osis
Thym
us: l
ymph
ocyt
e de
plet
ion
univ
ersa
l z
conn
ectiv
e tis
sue
form
atio
n, w
ell
3 2 r
m F 5 z z h
Y
b r-
Ultr
astr
uctu
re
Not
don
e
*A d
escr
iptio
n of
the
ultr
astr
uctu
re o
f so
me
mur
ine
mes
othe
liom
as c
an b
e fo
und
in r
ef. 2
8

PATHOBIOLOGY OF MALIGNANT MESOTHELIOMA 373
Table 11-Growth factors and cytokines with known or purported roles in the pathobiology of malignant mesothelioma
Cytokine Physiological
function Pathological
effects
IL-1
IL-2
IL-6
IL-10
TNFa
M- and GM-CSF
TGFB,, -82, $83
PDGF-A, -B
bFGF
T-cell activation Induction of 11-1, TNFa, GM-CSF synthesis Induction of acute phase response (APR)
Immunomodulation
Immunomodulation Induction of APR Angiogenesis
Immunomodulation
Immunomodulation Tissue repair and remodelling Induction of IL-1, IL-6, CSFs synthesis
Macrophage differentiation and activation
Immunomodulation Tissue repair and remodelling IL-6 induction
Tissue repair Growth factor for multiple dissimilar cell types ECM synthesis
Tissue repair and remodelling ECM synthesis (e.g., HA)
Pyrogenic action Fibrosis
Vascular leakage
Cachexia Thrombocytosis Am yloidosis Angiogenesis Pyrogenic action
Suppression of macrophage anti-tumour activity
Tissue necrosis Vascular leakage and hypovolaemic shock Cachexia
Immunosuppression ECM formation and fibrosis Angiogenesis Anchorage-independent growth of mesothelioma cells Cachexia
Fibrosis Tumour invasion Immunomodulation Autocrine mesothelioma growth
Stimulation of HA secretion by mesothelial cells Autocrine tumour growth Fibrosis Angiogenesis
metastases. The latter may, however, be more common than clinical assessments would suggest, as judged by post-mortem findings.42 Hyaluronan (HA), a ubiquitous component of the extracelluar matrix, is likely to play a major role in these aspects of mesothelioma patho- biology.43 Human mesothelioma cells produce factors, probably including PDGF and bFGF, which stimulate the production of HA by mesothelial cells and fibro- b l a s t ~ . ~ ~ Mesothelioma cells, but not normal mesothelial cells, express HA re~eptors.4~ TGFP appears to play a role in tumour cell mobility by simultaneously inducing the expression of at least one of the HA rece tors, receptor for HA-mediated motility (RHAMM)?' and
the secretion of HA.47 The expression of HA receptors, in particular CD44, has not yet been explored for its prognostic value in the context of mesothelioma, such as has been the case in other human cancers,43 where the prognosis may be dramatically better for patients with tumours lacking expression of CD44 variants. It is not known how such findings can be made compatible with the known function of CD44 isoforms as 'homing' receptors for lymphocytes48349 and the generally positive correlation between tumour infiltrating lymphocyte (TIL) numbers and a good prognosis in many human tum~urs ,~ ' including me~othe l ioma,~~ which is referred to later. This is in need of clarification.

374 H. BIELEFELDT-OHMANN ET AL.
TUMOUR-INFILTRATING LYMPHOCYTES AND MACROPHAGES
Another striking difference between the two types of murine mesothelioma tumours described above was the phenotypic profile of the TIL and TIM. IL-6-positive/ TGFB-negative tumours contained fewer TIM, but more TIL than TGF/3-positive/IL-6-negative t u m ~ u r s . ~ , ~ ~ Furthermore, the CD4:CD8 ratio amongst TIL was approximately 1 in IL-6-producing tumours, but greater than 4 in TGFP-producing tumours. This occurred despite similarly uniform MHC class I expression on all tumour cells and no, or low, expression of MHC class I1 antigen on tumour cells and TIM, respectively, in both types of tumours,’ suggesting that it is not tumour antigens and their presentation (or lack of) which govern lymphocyte infiltration and TIL phenotype. Rather, cytokines in the local tumour milieu, expression of ‘homing’ receptors, or other factors may determine these differences.
Only a few studies of TIL in solid human mesothelio- mas have been reported”,” and no attempts have apparently yet been made to phenotype cells beyond the distinction of T and B cells, despite the availability of reagents and the widespread application of such studies in other human cancers. It is therefore not possible at present to assess the relevance of the findings in the murine model for human disease. However, considering the potential for using TIL markers in the monitoring of immunotherapy responses in mesothelioma, as demon- strated in preclinical investigation^,^^,^^ such studies seem warranted. It should, however, be kept in mind that the studies in the murine model had the exceptional advantage of being conducted with tumour cells with distinct cytokine profile^.^',^'
Likewise, only limited information is available regard- ing the inflammatory cells in pleural and ascitic effusions in human patients. Hoogsteden et d3 and Gusman rt uI. ’~ described a marked influx of CDS-positive/CD3- positive T lymphocytes in pleural effusions, with an increased CD4:CD8 ratio compared with peripheral blood. The phenotype of pleural effusion macrophages suggested influx of monocytes from the peripheral blood.”.54 Although possible implications of these find- ings on anti-tumour responses were discussed, these reports contained no assessment of cellular functions. De Pangher Manzini ct uI.’~ listed the cell composition in pleural fluid as a prognostic factor; there was no mention, however, of leukocytes. From studies in healthy individuals there is evidence to suggest that the peritoneal cavity contains a non-random steady-state population of lymphocytes and macro phage^'^ and that any changes therefore have the potential to be of diagnostic/prognostic value, as has been the case for the assessment of cells in broncho-alveolar lavage fluid in various lung diseases.57 This contention is also sup- ported by the findings of Monti e t ~ l . , ~ ~ who reported on an assessment of macrophages and CD8-positive cell activation in pleural IL-6-containing mesothelioma effu- sions following interferon-y (IFNy) therapy. The results obtained were indicative of macrophage and selective T-cell activation, although at this stage the results
cannot be definitively related to clinical status or prog- nosis. The study does, however, suggest a worthwhile avenue for monitoring mesothelioma patients, including those undergoing immunotherapy.
In the murine mesothelioma model, it was shown that during growth of tumours in the peritoneal cavity of syngeneic mice, the total number of leukocytes initially increased, followed by a slight decrease by the time that the animals showed severe clinical signs. Macrophages constituted the majority (>70 per cent) of cells through- out the tumour growth period.9330 However, MHC class I1 antigen expression on the peritoneal macrophages decreased progressively, while their ‘spontaneous’ cytokine (IL-6, IL-1, TNFn) production increased,”.”” suggesting a tumour-induced phenotypic change of the population. Although the mechanism at this stage remains speculative, it is known that TGFP can induce such changes in vitro5’ and that other cytokines with macrophage modulatory activities, including M-CSF and IL-10, are also present in the ascitic fluid of mesothelioma-bearing
IMMUNOLOGY OF MALIGNANT MESOTHELIOMA
A variety of immune effector cells and functions, including cytotoxic T cells, macrophages, and natural killer (NK) and lymphokine-activated killer (LAK) cells, are known to mediate anti-tumour effects in vitro. Although it could be surmised that the long latency period of mesothelioma might be due to efficient immune-mediated surveillance mechanisms, with malig- nant growth arising due to a final ‘breakdown’ in these reactions, the known inhibitory effects of asbestos fibres on many anti-cancer immune defences6() challenge this explanation. More importantly, as described above, TGFP, one of the most potent endogenous immune modulatory molecules, is present at high levels in the micro-environment of transformed mesothelium and many human and murine mesothelioma cell lines are among the most active TGFP producers de~cribed.~” High levels of bioactive TGFP, and TGFP, have been reported in pleural effusions from mesothelioma patient^.^' TGFB appears to be a very potent immuno- suppressant: it inhibits the priming of CD8-positive MHC class I-restricted T lymphocytes, down-regulates both constitutive and IFNy-induced expression of M HC class I1 antigen on lymphoid and non-lymphoid cells, and suppresses nitric oxide (NO) and TNFa production by macro phage^.'^^^'
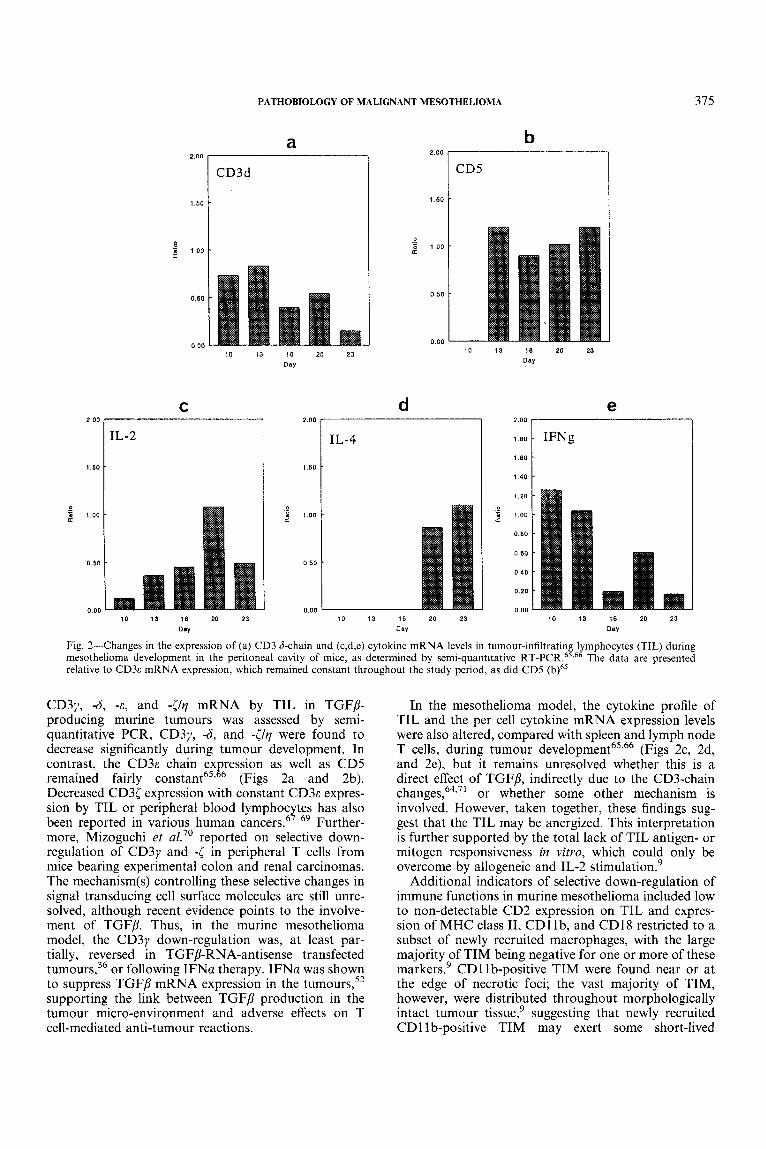
Early studies of TIL in human mesothelioma patients suggested a positive correlation between TIL numbers and survival time.33 In a murine model of mesothelioma, not only were TIL numbers found to be low in TGFP- producing tumours,’ as described above, but there was a selective down-regulation of chains of the T-cell receptor (TCR)-associated signalling molecule CD3. The CD3 complex consists of six transmembrane protein subunits, y , 6, E , [, rl, and cc), which are involved in the stabilization of the surface expression of the TCR and the transduc- tion of T-cell activation signals.62p64 When expression of
PATHOBIOLOGY OF MALIGNANT MESOTHELIOMA
1.50
a
IL-2
.
315
1 5 0
b
IL-4
.
2.00
1.50
2 2 1.00
0.50
0.00
C
CD3d
10 13 1 8 20 23
Day
d
2 00
1 5 0
1 0 0
0 50
0 00
CD5
10
2.00 , 2.00 , 1
I 1.00 ii 1.00
0.50 0.50
0.00 0.00 10 13 1 6 20 23
DW
10 13 15 20 23
O W
13 16 20 23
Day
2.00
1 .a0
1 8 0
1 4 0
1 2 0
I 1.00
0.80
0.80
0 . 4 0
0.20
0.00
e
IFNg
1 0 13 1 6 20 23
Day
Fig. 2-Changes in the expression of (a) CD3 &chain and (c,d,e) cytokine mRNA levels in tumour-infiltrating lymphocytes (TIL) during mesothelioma development in the peritoneal cavity of mice, as determined by semi-quantitative RT-PCR.65,66 The data are presented relative to CD3c mRNA expression, which remained constant throughout the study period, as did CD5 (b)65
CD3y, -6, -E , and -[/?I mRNA by TIL in TGFP- producing murine tumours was assessed by semi- quantitative PCR, CD3y, -6, and -[/?I were found to decrease significantly during tumour development. In contrast, the C D ~ E chain ex ression as well as CD5
Decreased CD3[ expression with constant C D ~ E expres- sion by TIL or peripheral blood lymphoc tes has also been reported in various human cancers!’-69 Further- more, Mizoguchi et a1.” reported on selective down- regulation of CD3y and -[ in peripheral T cells from mice bearing experimental colon and renal carcinomas. The mechanism(s) controlling these selective changes in signal transducing cell surface molecules are still unre- solved, although recent evidence points to the involve- ment of TGFP. Thus, in the murine mesothelioma model, the CD3y down-regulation was, at least par- tially, reversed in TGFP-RNA-antisense transfected turn our^,^^ or following IFNa therapy. IFNa was shown to suppress TGFP mRNA expression in the turn our^,^^ supporting the link between TGFP production in the tumour micro-environment and adverse effects on T cell-mediated anti-tumour reactions.
remained fairly constant65’ P (Figs 2a and 2b).
In the mesothelioma model, the cytokine profile of TIL and the per cell cytokine mRNA expression levels were also altered, compared with spleen and lymph node T cells, during tumour d e v e l ~ p m e n t ~ ~ ~ ~ ~ (Figs 2c, 2d, and 2e), but it remains unresolved whether this is a direct effect of TGFP, indirectly due to the CD3-chain change^,^^,^' or whether some other mechanism is involved. However, taken together, these findings sug- gest that the TIL may be anergized. This interpretation is further supported by the total lack of TIL antigen- or mitogen responsiveness in vitro, which could only be overcome by allogeneic and IL-2 stimulation.’
Additional indicators of selective down-regulation of immune functions in murine mesothelioma included low to non-detectable CD2 expression on TIL and expres- sion of MHC class 11, CD1 Ib, and CD18 restricted to a subset of newly recruited macrophages, with the large majority of TIM being negative for one or more of these markers.’ CD1 Ib-positive TIM were found near or at the edge of necrotic foci; the vast majority of TIM, however, were distributed throughout morphologically intact tumour tissue,’ suggesting that newly recruited CD11 b-positive TIM may exert some short-lived

376 H. BIELEFELDT-OHMANN ET AL.
c - -
I .< ch emoatt ract ion
TGF-I3 - (MQ, stimulation \,
\ J
- _ a\ ,MM\,
J. Growth Stimulation TGF-I3
TNF-a IL-I 11-6
Tumour T Cell Deactivation
Systemic Pathology \
Liver degenerationlnecrosis
I CD3 i CD2 1 Cytokines
Vascular LeakagdHaemorrhage Ascites Production Shock, Death
Fig. 3-A model for the role of TGFP in the immunohiology of malignant mesothelioma. This model does not take into account tumour-produced IL-6, which alone or in combination with TGFP has also been shown to play a rolc in the pathophysiology of malignant mesothelioina
cytotoxic anti-tumour activity, but soon after arrival become deactivated or even switched to a mode of tumour promotion via the production of PDGF, TGFa, TGFP, etc6 (Fig. 3) . TGFP may play a role both in the recruitment of macrophages to the tumour and in the subsequent down-regulation of their anti-tumour cytotoxic ~ a p a c i t y . ~ ~ . ~ ~ , ~ ~ , ’ ~
Very little is known about the systemic immune reactivit in humans or animals during the latency period.6X Mice challenged intraperitoneally with either TGFP- or IL-&producing mesothelioma cells show pro- gressive thymic atrophy and down-regulation of all lymphocyte-markers, andlor loss of marker-positive spleen cells was These changes were ac- companied by changes in mitogen-induced proliferation and cytokine production, characterized by an initial increase followed by a return to control levels or a depressed response. I t is notable that these changes preceded detectable tumour growth by 10-12 days, and might be due to either cytokine-induced changes in lymphocyte trafficking and cell activities, chronic expo- sure to tumour antigens at inappropriate high levels, or corticosteroid- or cytokine-induced apoptosis of acti- vated lymphocyte^.^,^^ Since these mechanisms are not mutually exclusive, they may also be invoked alone or in
combinations to explain the varied results found in studies of systemic immune reactivity in human mesothelioma patient^.^',^'
Another notable result of the investigations in the murine mesothelioma model was the total absence of detectable tumour-specific T-cell responses. Other studies have shown that the murine mesothelioma cell lines employed are immunogenic and constitutively express MHC class I antigen^,"'^ as do their human counterpart^.^^ However, down-regulation of MHC class I1 antigen on peritoneal, spleen, and lymph node macrophages and of CD3 and CD2 on lymphoc tes may have curtailed tumour-specific T-cell This may be further aggravated by the lack of B7 (CD70, CD86), integrins, and perha s other accessory molecules on the mesothelioma cells.’ Additional direct inhibitory effects may be exerted by TGFPs8 (Fig. 3 ) and perhaps other, as yet unidentified, factors.
Taken together, the data available so far on the cytokine- and immunobiology of malignant mesothe- lioma suggest that a range of cytokines, but in particular TGFP, with potential adverse effects on immune effector functions are produced during the development of mesothelioma. This profoundly affects, both locally and systemically, the development of anti-tumour effector
2 ”

PATHOBIOLOGY OF MALIGNANT MESOTHELIOMA 377
cells, including helper and cytotoxic T-lymphocyte ac- tivity and macrophage-mediated anti-tumour functions, thereby contributing to the escape of mesothelioma cells from immune-mediated elimination.9336
IMPLICATIONS OF PATHOBIOLOGY AND IMMUNOBIOLOGY FOR IMMUNOTHERAPY
If the above assumptions are correct, it would imply that immunotherapy, in order to be successful, must not only aim to target the local cytokine-mediated pathways promoting tumour cell growth, but perhaps just as importantly, aim to inhibit the cytokine-mediated path- ways which exert a negative effect on the development and expression of anti-tumour immune responses. How- ever, in order to achieve these goals, it may be necessary not only to refine the selection and combination of therapeutic agents, but also to commence treatment earlier than is currently the routine in clinical trial^.^ In recent studies of IFNa therapy in a murine meso- thelioma model, it was clearly shown that only if treat- ment was commenced early in tuniour development (a stage probably comparable to ‘mesothelioma in situ’ or with only minimal effusion and little solid tumour detectable by imaging) could a significant therapeutic effect be obtained, in this case by inhibition of TGFP production by tumour cells.52
Considering the pivotal role of TGFP in the patho- and immunobiology of mesothelioma, this molecule should be considered a prime target for therapy. Anti- sense technology, although a useful research tool,36 is not yet readily amenable to achievable therapeutic approaches. However, recombinant soluble TGFP receptors, decorin, and the TGFP latency-associated peptide (LAP) might be useful as antagonists, alone, or in combination with other therapeutic agents.
As mentioned above, paraneoplastic syndromes may contribute significantly to the weakened constitution of mesothelioma patients and intensify the side-effects of chemo- and immuno-therapeutic treatments. The recent findings of a role for IL-6, and perhaps TNFa, IL- 1, and other c y t o k i n e ~ , ~ . ’ . ~ . ~ ~ . ~ ~ 39 in the pathophysiology of mesothelioma opens up the avenue for new approaches to palliative treatment: in its own right to alleviate pain and discomfort and, perhaps more importantly, as adjuvant therapy for conventional and new immuno- therapeutic treatments. Cytokine antagonists, soluble cytokine receptors, cytokine synthesis inhibitors, and anti-cytokine antibodies are now being employed in the treatment of sepsis, persistent viral infections, and autoimmune diseases, and thus should be considered available adjuncts for mesothelioma treatment.
Mesothelioma may be a relatively rare and so far intractable cancer, but new developments in cancer biology and medicine offer fresh hope for patients with this malignancy. Further progress will be aided signifi- cantly by more emphasis on the assessment of various local and systemic pathological features in addition to those required for a correct diagnosis and treatment, in- cluding paraneoplastic phenomena and immunological parameters.
1.
2.
3.
4.
5.
6.
7.
8.
9.
10.
1 1 .
12.
13.
14.
15.
16.
17.
18.
19.
20.
21.
22. 23.
24.
25.
26.
27.
28.
29.
30.
REFERENCES
Pelnar PV. Further evidence of non-asbestos-related mesothelioma. Scund J Work Eneiron Health 1988; 1 4 141-144. Saebe A, Elgjo K , Lassen J. Could development of malignant mesothelioma by induced by Yersiniu enrerocolitrcu infection? Med Hypoth 1993; 4 0 275-277. Cicala C, Pompetti F, Carbone M. SV40 induces mesotheliomas in hamsters. Am J Puthul 1993; 142 152kI533 Bedrossian CW, Bonsib S, Moran C. Differential diagnosis between meso- thelioma and adenocarcinoma: a multimodal approach based on ultrastruc- ture and immunocytochemistry. Semin Diugn Pathoi 1992; 9 124140. Fitzpatrick DR, Manning LS, Musk AW, Robinson BWS, Bielefeldt- Ohmann H. Potential for cytokine therapy of malignant mesothelioma. Cuncer Treutment Rev 1995, 21: 273-288. Genvin BI. Mesothelial carcinogenesis. Possible avenues of growth factor promotion. In: Jaurand MC, Bignon J, eds. The Mesothelial Cell and Mesothelioma. New York: Marcel Dekker, 1994; 223-243. Fitzpatrick DR, Peroni DJ, Bielefeldt-Ohmann H. The role of growth factors and cytokines in the tumorigenesis and immunobiology of malignant mesothelioma. Am J Respir Cell Mul Biol 1995; 1 2 435460. Jaurand MC, Barrett JC. Neoplastic transformation of mesothelial cells. In: Jaurand MC, Bignot J, eds. The Mesothelial Cell and Mesothelioma. New York: Marcel Dekker, 1994; 207-223. Bielefeldt-Ohmann H, Marzo A, Fitzpatrick DR, et ul. lmmunobiology of murine malignant mesothelioma (MM): characterisation of tumour infil- trating leukocytes and cytokine production in a murine model of MM. Cunrer Immunol Imniunuther 1994; 39 347-359. Walker C , Everitt J , Barrett JC. Possible cellular and molecular mechanisms for asbestos carcinogenicity. Am J Ind Med 1992; 21: 253-273. Cheng JQ, Jhanwar SC, Klein WM, er a/. p16 alterations and deletion mapping of 9p21Lp22 in malignant mesothelioma. Cuncer Res 1994; 5 4 5547-5551. Heintz NH, Janssen YM, Mossman BT. Persistent induction of c$Os and c-jun expression by asbestos. Proc Nut/ AcudSci USA 1993; Y O 3299-3303. Grabrielson EW, Van der Meeren A, Reddel RR, Reddel H, Gerwin B1, Harris CC. Human mesothelioma cells and asbestos-exposed mesothelial cells are selectively resistant to amosite toxicity: a possible mechanism for tumor promotion by asbestos. Curcinogenesis 1992; 1 3 1359-1 363. Resnick D, Freedman NJ, Xu S, Krieger M. Secreted extracellular domains of macrophage scavenger receptors from elongated trimers which specifi- cally bind crocidolite asbestos. J B i d Chem 1993; 268: 3538-3545. Noble PW, Henson PM, Riches DW. Insulin-like growth factor-I (IGF-I) mRNA expression in bone marrow derived macrophages is stimulated by chrysotile asbestos and bleomycin. A potential marker for a reparative macrophage phenotype. Chest 1991; 9 9 79s-80s. Bonner JC, Brody AR. Asbestos-induced alveolar injury. Evidence for macrophage-derived PDGF as a mediator of the fibrogenic response. Chest 1991; 9 9 54S-55S. Davis JMG, Addison J, Bolton RE, Donaldson K, Jones AD, Smith T. The pathogenicity of long versus short fibre samples of amosite asbestos administered to rats by inhalation and intraperitoneal injection. Br J E.xp Puthol 1986; 67: 415430. Moalli PA, MacDonald JL, Goodglick LA, Kane AB. Acute injury and regeneration of the mesothelium in response to asbestos fibers. Am I Pathoi 1987; 128: 426445. Monchaux G, Bignon J, Jaurdnd MC, et a/. Mesotheliomas in rats following inoculation with acid-leached chrysotile asbestos and other mineral fibers. Curcinogenesis 1991; 2 229-236. Brdnchaud RM, MacDonald JL, Kane AB. Induction of angiogenesis by intraperitoneal injections of asbestos fibers. FASEB J 1989; 3 174771752, Begin R, Rola-Pleszczynski M, Masse S, et ul. Asbestos-induced lung injury in the sheep model: the initial alveolitis. Environ Res 1983; 3 0 195-210. Nathan C, Sporn M. Cytokines in context. J Cell Biol 1991; 113 981-986. Robinson BWS, Musk AW. Benign asbestos pleural effusion: diagnosis and course. Thurux 1981; 3 6 896-900. Hillerdal G. Pleural plaques and risk for bronchial carcinoma and meso- thelioma. A prospective study. Chest 1994; 105 144--150. Browne K. Asbestos related malignancy and the Cairns hypothesis. Br J Ind Med 1991; 48: 73-76. Soosay GN, Griffiths M, Papadaki L, Happerfield L, Borrow L. The differential diagnosis of epithelial-type mesothelioma from adenocarcinoma and reactive mesothelial proliferation. J Puthol 1991; 163 299-305. Henderson DW, Shilkin KB, Whitaker D, ef d. The pathology of malignant mesothelioma, including immunohistology and ultrastructure. In: Henderson DW, Shilkin KB, Langois SL, Whitaker D, eds. Malignant Mesothelionia. New York: Hemisphere, 1992; 69-1 39. Davis JMG. Histogenesis and fine structure of peritoneal tumors in animals by injections of asbestos. J Null Cuncer lnsr 52: 1823-1837. Davis JMG. The histopathology and ultrastructure of pleural mesothelio- mas produced in the rat by injections of crocidolite asbestos. Br J Exp Puthol 1979; 6 0 642-652. Bielefeldt-Ohmann H, Marro AL, Himbech RP, Jarnicki AG, Robinson BWS, Fitzpatrick DR. Interleukin-6 involvement in mesothelioma

378 H. BIELEFELDT-OHMANN ET AL.
pathobiology: inhibition by interferon-u immunotherapy. Cancer fmmunol lmnninothcv 1995; 4 0 241-250.
31. Roggli VL, Sanfilippo F. Shelburne JD. Mesothelioma. In: Roggli VL, Greenberg SD. Pratt PC, eds. Pathology of Asbestos-Associated Diseases. Boston: Little, Brown & Co. 1992; 109-164.
32. Symington FW, Symington BE, Liu PY, Viguet H, Santhanam U, Sehgdl PB. The relationship of serum IL-6 levels to acute graft-versus-host and hepatorenal discase after human bone marrow transplantation. Trun.~plan- turiun 1992; 54 457462.
33. Leigh RA, Webster I. Lymphocytic infiltration of pleural mesothelioma and its significance for survival. S Afr Med .I 1982; 61: 1007--1009.
34. Wahl SM, Hunt DA, Wakefield LM, e t a / . Transforming growth factor type / j induces monocyte chemotaxis and growth factor production. Proc Nut/ Acud Sci USA 1987: 84 5788-5792.
35. Wiseman DM, Polverini PJ, Kamp DW, Leibovich SJ. Transforming growth factor-beta (TGFP) is chemotactic for human monocytes and induces their expression of angiogenic activity. Biochem Bir~phph~..~ Res Corn-
36. Fitqatrick DR, Biclefeldt-Ohmann H, Himbcck RP, Jarnicki AG, Marzo AL. Robinson BWS. Transforming growth factor-beta: antisense RNA- mediated inhibition arects anchorage-independent growth, tumorigenicity and tumour-infiltrating T-cells in malignant mesothelioma. Griwlh Furfurs 1994; 11: 2944.
37. Higashihara M, Suiiaga S, Tange T, Oohashi H, Kurokdwa K, Increased cccretion of interleukin-6 i n malignant mesothelioina cells from a patient with marked thronibocytosis. Curicrr 1992; 7U: 2105-2108.
38. Motoyama T, Honma T, Walanabe H, Honma S, Kumanishi T, Abe S. Interleukin-6-producing malignant mesothelioma. Virrhoiv.~ Arch B /Cell Puthid] 1993; 64: 367-~372.
39. Monti G. Jaurand MC, Monnet I, ef a/. Intrapleural production of interleukin 6 during mesothelioma and its modulation by y-interferon treatment. Cuncer Res 1994; 54 44194423.
40. Strassniann G, Fong M, Kenney JS, Jacob CO. Evidence for the involve- ment of interleukin-6 in cxpcrimcntal cancer cachexia. J Clin f n w s t 1992; 89 1681 1684.
41. Oliff A. The role of tumor necrosis factor (cachectin) in cachexia. Cell 1988; 54: 141 143.
42. Wronski M, Burt M. Cerebral metastases in pleural mesothelioma: case report and review of the literature. J Neuro-Oncol 1993; 17: 21-26.
43. Sherman I-, Sleeman J, Herrlich P, Ponta H. Hyaluronate receptors: key players in growth, differentiation, migration and tumor progression. Curr Opinion (‘dl Biol 1994; 6 726-733.
44. Asplund T, Versnel MA, Laurent TC, Heldin P. Human mesothelioma cells produce Factors that stimulate the production of hyaluronan by mesothelial cells and fibroblasts. Cuncer Res 1993; 53 388 392.
45. Asplund T, Heldin P. Hyaluronan receptors are expressed on human malignant niesothelionia cells but not on normal mesothelial cells. Cuncer Red 1994; 54 45164523.
46. Hardwick C, Hoare K. Owens R, c’f N I . Molecular cloning of a novel hyaluronan receptor that mediates tumor cell motility. J Cell B i d 1992; 117: 1343- 1350.
47. Samuel SK. Hurta RAR, Spearman MA, Wright JA, Turley EA, Greenberg AH. TGF-PI stimulation of cell locomotion utilizes the hyaluronan receptor RIfAMM and hyaluronan. J Cdl Biol 1993; 123: 749 758.
48 Stanienkovic I , Aruffo A, Amiot A, Seed B. The hematopoietic and epithelial forms of CD44 are distinct polypeptides with different adhesion potentials for hyaluronate-bearing cells. EMBO J 1991; 10 343 348.
49. Dougherty GJ, Lansdorp PM, Cooper DL, Humphries RK. Molecular cloning of CD44RI and CD44R2, two isoforms of the human CD44 lymphocyte ‘homing’ receptor expressed by hemopoietic cells. J E.I, Med 1991; 174 1-5.
50. Vose BM. Moore M. Human tumour-infiltrating lymphocytes. Semin Hrmutol 1985; 22: 27~40.
51. Henderson DW, Shilkin KB. Whitaker D, ef ul. Unusual histological types and anatomic sites of inesothelioma. In: Henderson DW, Shilkin KB, Langois SL, Whitaker D, eds. Malignant Mesothelioma. New York: Hemisphere, 1992; 140-166.
52. Bielefeldt-Ohmann H, Fitrpatrick DR, Marzo AL, Jarnicki AG, Musk AW, Robinson BWS. The potential for interferon-u therapy in mcsothelioma: assessment in a murine model. J Interferon Cytokine Res 1995; 15 213-223.
53. Hoogsteden HC, Versnel MA, Hop W, vander Kwast ThH, Hilvering C. Immunological characterization of leukocytes in pleural fluid in malignant mesothelioma. Eur Rr;pir R m 1993; 3: 30G32.
19x8; 157: 793-800.
54.
55.
56.
57.
58.
59.
60.
61.
62.
63.
64.
65.
66.
67.
68.
69.
70.
71.
72.
73.
74.
75.
76.
Guzman J: Bross KJ, Wiirtemberger G, Costabel U . lmmunocytology in malignant pleural mesothelioma. Chevt 1989: 9 5 590 595. De Pangher Manzini V, Brollo A, Franceschi S, De Matthaeis M, Talamini R, Bianchi C. Prognostic factors of malignant mesothelioma of the pleura. Cuncw 1993; 7 2 410 417. Kubicka U. Olszewski WL, Maldyk J, Wierzbicki 2, Orkiszewska A . Normal human immune peritoneal cells: phenotypic characteristics. Iinniimohiologj 1989; 180 80-92. Hunninghdke GW, Gadek JE, Kawanami 0, Ferrans VJ, Crystal RG. Inflammatory and immune processes in the human lung in health and disease. Evaluation by bronchoalveolar lavagc. Ani J Purhol 1979; 97: 149-206. Ruscetti FW, Palladino MA. Transforming growth factor-beta and the immune system. Prog Growth Factor Res 1991; 3 159-171. Alleva DG, Burger CJ, Elgert KD. Tumor-induced regulation of suppressor macrophage nitric oxide and TNF-u production Role of tumor-derived LL-10, TGF-/I and prostaglandin E,. .I h n ~ t i t n ~ i l 1994; 153: 1674-1686. Manning LS, Rose AH, Bowman RV, Christmas TI, Garlepp MJ, Robinson BWS. Immune function related to asbestos exposure and mesothelioma. and immunothcrapy of mesothclioma. In: Henderson DW, Shilkin KB, Langlois SLP. Whitaker D. eds. Malignant Mesothelioma. New York: Hemisphere, 1992; 278 291. Maeda J, Ueki N, Ohkawa T, et ul. Tr-ansfbrming growth factor-beta 1 (TGF-PI)- and @-like activitirs in malignant pleufiil effusions caused hy malignant mesothelioma or primary lung cancer. C ’ h Ekp lnrmunol 1994; 9 8 3 19-322. Alercon B, Berkhou B, Breitmeyer J, Terhorst C. Assembly of the human T cell receptor-CD3 complex takes place in the endoplasmic reticulum and involves intermediary complexes between CD3 7, 6 core and single T cell receptor a and P chains. J Biol Chrm 1988; 263 2953 -2961. Chetty R, Gatter K. CDB: structure, function, and role of immunostaining i n clinical practice. J Puthol 1994; 173: 303-307. Buferne M, Luton F, Letourneur F, et ul. Role of CD3B in surface expression of the TCR-CD3 coniplex and in activation for killing analyzed with CD3d’-negative cytotoxic T lymphocyte variant. J fmmunol 1992; 148: 657-664. Jarnicki AG, Fitzpatrick DR, Marzo A, Robinson B, Bielefeldt-Ohmann H. Surface antigen and cytokine profile of tumour infiltrating lymphocytes (TIL) in malignant mesothelioma. J Leuk B i d lYY3; 54 (Suppl): YO. Jarnicki A, FitLpatrick DR, Robinson BWS, Bielefeldt-Ohmann H. I n .r.itu characterization of lymphocytes infiltrating murine malignant mcso- thelioma: changes in the CD3-chain complex and cytokine expression during tumor development. (Submitted.) Fdrace F, Angevin E, Vanderplancke J, Escudier B, Triebel F. The decreased expression of CD3 [ chain, in cancer patients I S not reversed by 1L-2 administration. fn l J Canwr 1994; 59 752 755. Finke JH, Zea AH, Stanley J, pi ul. Loss of T-cell receptor zeta chain and p561ck in T-cells infiltrating human renal cell carcinoma. Cancer Res 1993: 53: 5613--5616. Nakagomi H, Petersson M, Magnusson I , rr ul. Decreased expression of the signal-transducing zeta chains in tumor-infiltrating T-cells and NK cells of patients with colorectal carcinoma. Ginrcr Reds 1993; 53 5620-5622. Mizoguchi H, O’Shea JJ, Longo DL, ef ul. Alterations in signal transduc- tion molecules in T lymphocytes from tumor-bearing mice. Science 1992: 258: 1795-1797. Weiss A, Imboden J, Hardy K, et ul. The role of the TYantigen receptor complex in T-cell activation. Annu Rev fmmunol 1986; 4 593-619. Lew F, Tsang P, Holland JF, Warner N, Selikoff U , Bekesi JG. High frequency of immune dysfunction in asbestos workers and in patients with malignant mesothelioma. J Clin fn~munol 1986; 6: 225-233. Manning LS, Davis MR, Bielefeldt-Ohmann H, el ul. Evaluation of immunogenicity of murine mesothelioma cells by immunization. Eur Re,.spir Rev 1993; 3: 234-237. Christmas TI, Manning LS, Davis MR, Robinson BWS, Garlepp MJ. HLA antigen expression and malignant mesothelioma. Am .I Rpspir Cc4 Mol Biol
Bierer BE, Sleckmaii BP, Ratnofsky SE, Burakoff SJ. Thc biological roles of CD2, CD4 and CDX in T-cell activation. Annu Rev Inununol 1989: 7: 579-599. Gregory CD, Murray RJ, Edwards CF, Rickinson AH. Downregulation of cell adhesion molecules LFA-3 and ICAM- I in Epstein Barr virus-positive Burkitt’s lymphoma underlies tumor cell escape from virus-specific T cell surveillance. J Exp Med 1988; 167: I8 I I -1 824.
1991; 5: 213 -220.





























