Embed Size (px)
Citation preview

Vol. 32, No. 2INFECTION AND IMMUNITY, May 1981, p. 563-5700019-9567/81/050563-08$02.00/0
Monoclonal Antibodies to Stage-Specific, Species-Specific, andCross-Reactive Antigens of the Rodent Malarial Parasite,
Plasmodium yoeliiDIANE W. TAYLOR,* K. JIN KIM, PHILLIP A. MUNOZ, CHARLES B. EVANS, AND
RICHARD ASOFSKYLaboratory ofMicrobial Immunity, National Institute ofAllergy and Infectious Diseases, National
Institutes ofHealth, Bethesda, Maryland 20205
Eighteen hybridoma cell lines were used to study species-specific, stage-specific,and serological cross-reactive antigens of the rodent malarial parasite, Plasmo-dium yoelii. Specificity and location of plasmodial antigens were determined byindirect fluorescent-antibody analysis. Results showed that a minimum of 12distinct plasmodial antigens could be distinguished by the 18 hybridomas. Anti-gens were found on the surface or within the cytoplasm of the parasite, but noton the surface of erythrocytes from infected animals. The majority (11 of 12) ofantigens were present in all erythrocytic stages of the parasite, but one was stage-specific for merozoites. Additional studies showed that 6 of 18 of the monoclonalantibodies identified species-specific antigens, 2 of 18 recognized antigens confinedto related rodent malarial parasites (Plasmodium berghei, Plasmodium vinckei,and Plasmodium chabaudi), whereas 8 of 18 detected cross-reactive antigenscommon to rodent, primate (Plasmodium knowlesi, Plasmodium falciparum),and avian (Plasmodium gallinaceum) malarias.
During malaria infection antiplasmodial anti-bodies (Ab's) are produced, but the number,location, and characteristics of many parasiteantigens (Ag's) which induce these responseshave not been identified (3, 7, 18, 20, 21, 24). Inthe past it has been difficult to identify andisolate individual plasmodial antigens and eval-uate their roles in production or suppression ofimmunity. The use of hybridoma technologynow makes it possible to produce large amountsof Ab against single antigenic determinants ofmalarial parasites. Monoclonal Ab's can be usedto identify and ultimately to purify malarialAg's.
It is well established that mice infected withnonlethal (strain 17XNL) Plasmodium yoeliidevelop protective immunity during primary in-fection, but mice infected with the lethal (17XL)substrain routinely die (27). Unfortunately, theimmune mechanism(s), including the Ag(s) in-volved in initiating protective immunity duringprimary nonlethal malaria, is unknown. SinceAb plays a role in immune protection (11) andsince antiplasmodial Ab's appear in the serumof infected mice before peak parasitemias, inves-tigations of plasmodial Ag's which induce hu-moral responses during the early stages of infec-tion are of interest. Because Ab's produced be-fore peak parasitemia bind to circulating para-sites and are removed from the serum, conven-tional serological and immunochemical tech-niques are not applicable. Therefore, the most
feasible approach was to produce hybridomasby using spleen cells from mice infected 4 to 6days previously with lethal or nonlethal malaria.Eighteen antiplasmodial hybridoma cell lineswere produced by using lymphocytes from in-fected animals. Then monoclonal Ab's were usedto partially characterize P. yoelii Ag's.
MATERIALS AND METHODSParasites. The 17XL and 17XNL substrains of P.
yoelii were used in this study (26).Production of monoclonal Ab's. BALB/c mice
(Cumberland View Farms, Clinton, Tenn.) wereprimed to malarial Ag's either by infection with 106 P.yoelii 17XL parasites 4 days before fusion (group 1) orby infection with 104 P. yoelii 17XNL parasites 4 or 6days before fusion (groups 2 and 3). The mean para-sitemia in the peripheral blood of the three groups ofanimals at the time of fusion was 17.5, <0.1, and 6.1%,respectively. Spleen cells from malaria-infected micewere fused at a 10:1 ratio with the P3-X63-NS/1 (NS1)cell line by the procedure of Margulies et al. (19).Culture supernatants collected from microtiter wells14 days after fusion were screened for antiplasmodialactivity by radioimmunoassay (15) or the indirect-fluorescent Ab (IFA) technique described below (16)or both. Antiplasmodial hybrids were cloned twice,using a thymocyte feeder layer (2), and ascites fluidwas produced in pristane (2,3,6,10-tetramethyl penta-decane; Aldrich Chemical Co., Milwaukee, Wis.)-primed mice.Hybridoma nomenclature. The hybrid cell lines
used in this study have been identified by: (i) anArabic numeral to designate the number of days after
563
on August 19, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

564 TAYLOR ET AL.
infection the fusion was performed; (ii) the strain of P.yoelii used, i.e., 17X lethal (L) or 17X nonlethal (NL);and (iii) a number to designate the individual hybridcell line. Thus, hybrid 6NL-3 is the third hybridomaproduced by fusion of NS1 and spleen cells fromBALB/c mice experimentally infected 6 days previ-ously with the 17XNL strain of P. yoelii.
Purification of antimalarial hybridoma Ab's.Partially purified monoclonal Ab's were prepared byprecipitation from pooled ascites fluid with ammoniumsulfate at half saturation. After dialysis against phos-phate-buffered saline (PBS), the immunoglobulin con-centration was determined spectrophotometrically(optical density, 280 nm), using the extinction coeffi-cient for mouse immunoglobulin, E?l% = 1.2. Ab'swere diluted to 1 mg/ml in PBS and used at thatconcentration unless otherwise specified. Serumpooled from BALB/c mice which had recovered from17XNL infections and subsequent 17XL challenge wastreated as described above, adjusted to 1 mg/ml, andused as a source of positive control Ab. To confirmthat Ab's present in partially purified ascites fluidswere responsible for the results obtained, selectedmonoclonal Ab's of the immunoglobulin G (IgG) sub-class were isolated by protein A Sepharose CL-4B(Pharmacia, Uppsala, Sweden) column chromatogra-phy (10).
Isotype analysis. The isotypes of immunoglobu-lins secreted by in vitro maintained cell lines weredetermined by polyethylene glycol Ouchterlony anal-ysis (8), using culture supernatants and affinity-puri-fied class-specific reagents (goat antimouse IgM, IgG1,IgG2a, IgG2b, IgG3, and IgA) (22). In addition, par-tially purified ascites fluids and purified monoclonalAb's were analyzed with the above heavy chain-spe-cific reagents by immunoelectrophoresis to insure thepresence of a single paraprotein.IFA staining. The IFA procedure described by
Kuvin et al. was used (16). In brief, smears of periph-eral blood from malaria-infected animals were dehe-moglobinized, treated with 20 Ml of partially purifiedascites fluids (1 mg/ml) for 20 min at room tempera-ture, washed for 15 min in three changes of PBS, andthen counterstained for 20 min with fluorescein iso-thiocyanate-conjugated goat anti-mouse immunoglob-ulin (GaMIg-a polyvalent reagent containing 3 mg ofAb per ml, with a fluorescein-to-protein ratio of 7).Smears were washed for an additional 15 min in PBS,mounted in 50% glycerol in barbital buffer (pH 8.6),and examined by epifluorescence microscopy (LeitzOrtholux microscope, Leitz, Wetzlar, Germany). Pho-tographs were taken using an Orthomat II camera andKodak Tri/X film.
Blood smears of Plasmodium spp. and Babesiamicroti were examined to determine whether the Agrecognized by each monoclonal Ab was species specificor serologically cross-reactive. The species of Plas-modium used included P. yoelii (17XL and 17XNLstrains), Plasmodium berghei (NYU-2 strain), Plas-modium vinckei vinckei-62 (kindly supplied by R. L.Beaudoin, Naval Medical Research Institute, Be-thesda, Md.), Plasmodium chabaudi (provided by J.F. Finerty, Laboratory of Microbial Immunity, Na-tional Institute of Allergy and Infectious Diseases)Plasmodium knowlesi (H strain grown in rhesus mon-
keys and supplied by R. W. Gwadz, Laboratory ofParasitic Diseases, National Institute of Allergy andInfectious Diseases), Plasmodium falciparum Campstrain (produced by continuous in vitro cultivation inhuman type 0 erythrocytes by D. Haynes, WalterReed Army Medical Institute, Washington, D.C.), andPlasmodium gallinaceum grown in New HampshireRed chickens (graciously donated by R. Carter, Lab-oratory of Parasitic Diseases, National Institute ofAllergy and Infectious Diseases). The King-67 strainof B. microti, routinely maintained in our lab, also wasused. Smears of P. yoelii sporozoites (kindly suppliedby R. L. Beaudoin, Naval Medical Research Institute,Bethesda, Md.) were treated as described above.
In each IFA assay, a blood smear was stained withGiemsa stain to verify that all erythrocytic stages werepresent. Smears of all parasites were prepared andexamined at least twice. In synchronous malarial in-fections, blood was drawn several times during thecycle. However, blood smears containing B. microtidid not contain many late-stage parasites.
Partially purified ascites fluids were tested for anti-Toxoplasma gondii (RH strain) activity by MiltonLunde (Laboratory of Parasitic Diseases, NationalInstitute of Allergy and Infectious Diseases), using thetechniques of passive hemagglutination (13) or theSabin-Feldman dye test (23) or both.
Preparation of free parasites for differentiat-ing internal and membrane-associated Ag's. TheIFA procedure described above, using smears contain-ing erythrocytic-stage parasites, does not distinguishAg's located on the outer membranes of parasites frominternal plasmodial cytoplasmic Ag's. To determinewhether monoclonal Ab's could bind to Ag's on thesurface of freed P. yoelii parasites, parasites werereleased from erythrocytes by the following methodbefore staining. Blood cells from P. yoelii 17XL-in-fected BALB/c mice were washed three times withPBS, passed through a CF 11 Whatman cellulosecolumn to remove leukocytes (12), centrifuged, andswollen in the presence of 0.65% sodium chloride.Weakened erythrocytes were lysed by passage througha French pressure cell at 200 lb/in2, which resulted inthe release of intact parasites. Parasites were carefullywashed free of erythrocyte ghosts by differential cen-trifugation. Suspensions of intact parasites weretreated with 50 p1 of monoclonal Ab for 30 min on ice,washed, and then incubated with 10M1 of fluorescein-ated GaMIg for 30 min.Assaying for neoantigens on the surface of
infected erythrocytes. The technique of flow micro-fluorometry was employed for the detection of neoan-tigens on the surface of erythrocytes from P. yoelii-infected mice. A 1% suspension of erythrocytes wasprepared with washed erythrocytes from normal andmalaria-infected mice. Next, 100 pI-samples of theabove erythrocyte suspensions were incubated with100 pl of partially purified monoclonal Ab at roomtemperature for 30 min. Cells were washed twice withPBS and then incubated with a fluoresceinated F(ab')2fragment of affinity-purified goat anti-mouse kappalight chain immunoglobulin for 30 min at room tem-perature. Cells were washed and suspended in PBS.
Flow microfluorometry was performed with a FACSII (Becton, Dickinson & Co., Sunnyvale, Calif.). Light
INFECT. IMMUN.
on August 19, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

MALARIAL ANTIGENS 565
scatter windows were set to exclude small parasitesspontaneously released during sample processing. Inall samples, 50,000 cells were analyzed. Data were
collected in 1,000-channel linear distribution, stored,and processed in a PDP 11/40 computer (DigitalEquipment Corporation, Maynard, Mass.). A sampleof the cells also was examined by conventional epiflu-orescence microscopy.
RESULTS
Hybridomas. Eighteen anti-P. yoelii hybrid-oma cell lines were successfully established andcloned. Ten of the hybrids were produced byusing spleen cells from mice infected for 4 dayswith the lethal substrain of P. yoelii. Thesehybrids secreted IgM and IgG2a Ab's (Table 1).Four IgM-secreting hybridomas were producedby using spleen cells from mice infected with thenonlethal substrain of P. yoelii for 4 days. Fu-sions with cells from animals with 6-day nonle-thal infections resulted in six hybrids which se-
creted IgM, IgG2a, and IgG3 Ab's. In this study,antiplasmodial Ab's of the IgGl and IgG2b sub-classes were not produced. Monoclonal Ab'sfrom all 18 hybrids showed antiplasmodial activ-ity by both radioimmunoassay and IFA analysis.Patterns of immunofluorescence pro-
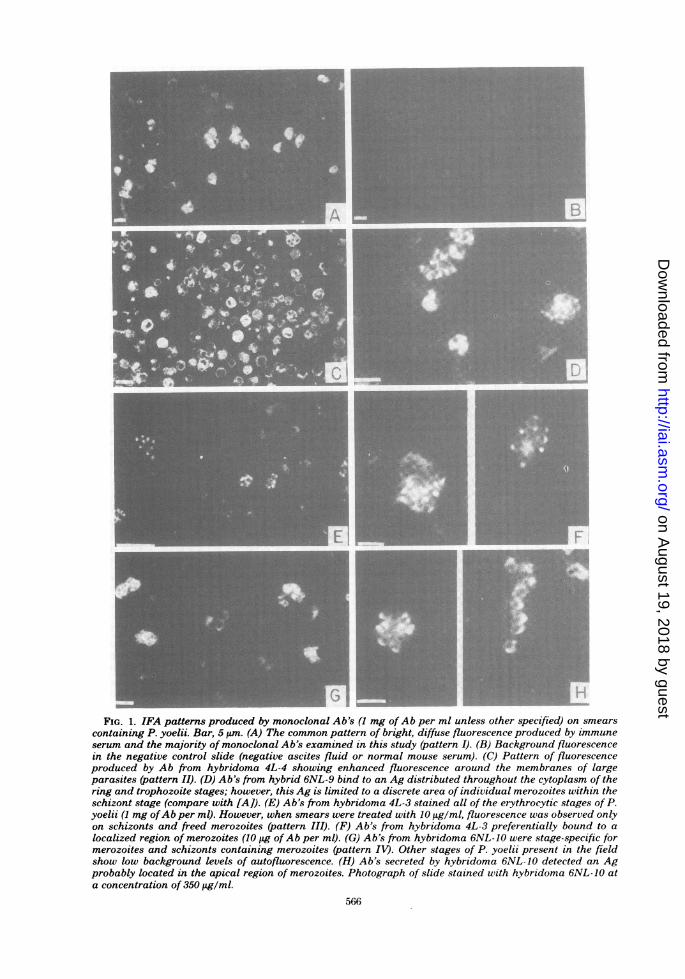
duced by monoclonal Ab's. Examination ofsmears of P. yoelii (strains 17XNL and 17XL)stained by IFA revealed that the 18 monoclonalAb's produced at least four distinct patterns ofimmunofluorescence.The majority of monoclonal Ab's produced a
fluorescence pattern similar to that commonlyobserved with immune serum (pattern I); i.e., allerythrocytic stages of the parasite showedbright, diffuse fluorescence (Fig. 1A). This pat-tern of fluorescence resulted when smears were
stained with a variety ofAb concentrations (0.01to 10 mg/ml).The second pattem of fluorescence (pattern
II) was produced by hybridomas, 4L-1, 4L-4, and4L-6. These Ab's appeared to bind to all eryth-rocytic stages, but produced enhanced fluores-cence around the edge of large parasites (Fig.1C).A group of three hybrids (4L-3, 6NL-6, and
6NL-9) secreted Ab's which bound to all of theerythrocytic stages of P. yoelii when smears
were treated with Ab at 1 mg/ml. However,dilution of the Ab to 10 ,tg/ml resulted in char-acteristic staining pattern III, in which fluores-cence was confined to a portion of individualmerozoites within schizonts (Fig. 1D-F). Thus,it appears that all of the erythrocytic stages ofthe parasite contain low cytoplasmic levels ofthis Ag(s) but that the Ag(s) is in a higherconcentration in a specific region of the mero-
zoite.
TABLE 1. Summary ofIFA staining patterns andlocation of P. yoelii antigens
Antigen located:
Hybridoma Isotype IFA On sur- WithinHybridoma Isot pe attemna face of para-freed sitepara- cyto-sites plasm
4L-1 IgG2a II - +4L-2 IgG2a I - +4L-3 IgM III - +4L-4 IgM II - +4L-5 IgM I - +4L-6 IgM II - +4L-7 IgG2a I +4L-8 IgG2a I - +
4NL-1 IgM I - +4NL-2 IgM I - +4NL-3 IgM I - +4NL-4 IgM I - +
6NL-5 IgM I - +6NL-6 IgM III - +6NL-7 IgG2a I - +6NL-8 IgG3 I - +6NL-9 IgM III - +6NL-10 IgG3 IV - +a For a description of patterns, see text.
Of the 18 hybrids, 1 produced Ab to a stage-specific Ag (pattern IV). Ab from hybrid 6NL-10 bound to merozoites within schizonts or tomerozoites released from schizonts, but not tothe other erythrocytic stages of P. yoelii (Fig.1G). At 1 mg of Ab per ml, fluorescence wasobserved throughout the cell, but upon dilution,fluorescence was noted only in a discrete area ofmerozoites (Fig. 1H).A single pattern of fluorescence was not re-
stricted to a single isotype (Table 1). For exam-ple, pattern I was produced by IgM-, IgG2a-,and IgG3-secreting hybrids. Thus, different pat-terms of fluorescence did not reflect a differencebetween IgM and IgG staining, but indicatedthat the monoclonal Abs were detecting differ-ent parasite Ag's.Differentiating Ag's recognized by the
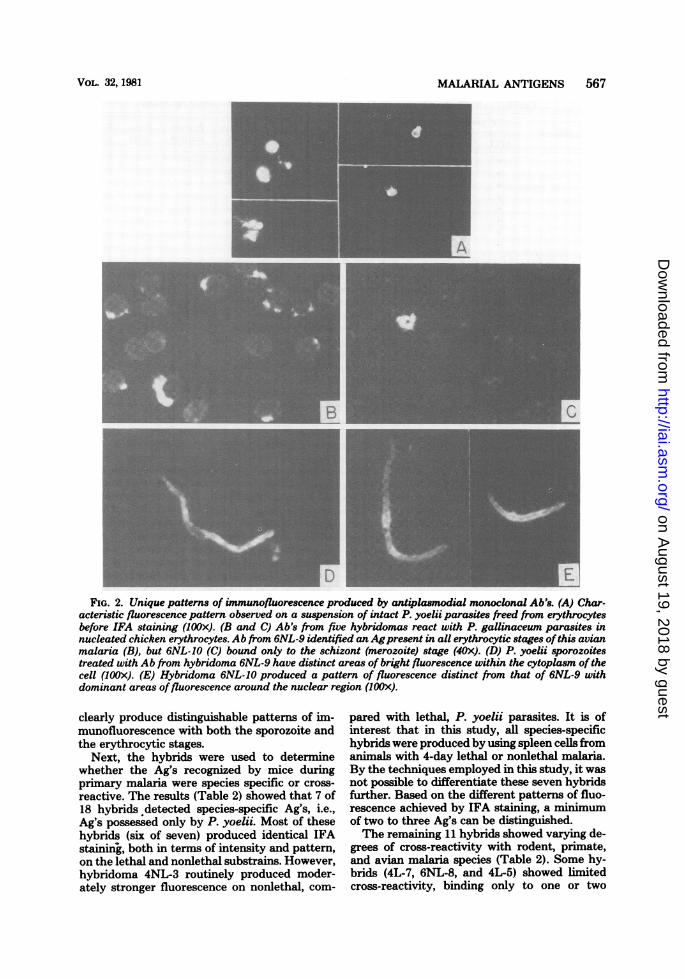
monoclonal Ab's. To characterize these Ag'sfurther, the monoclonal Ab's were first screenedby IFA analysis on P. yoelii sporozoites. Only 2of 18 hybrids detected Ag's common to the spo-rozoite and erythrocytic stages. Ab from hybrid6NL-9 produced a pattern ofpatchy fluorescencethroughout the cytoplasm of the sporozoite (Fig.2D), whereas bright fluorescence around thenuclear region and in a discrete spot posterior tothe nucleus was produced by Ab from hybrid6NL-10 (Fig. 2E). Hybrids 6NL-9 and 6NL-10apparently recognize different Ag's as they
VOL. 32, 1981
on August 19, 2018 by guest
http://iai.asm.org/
Dow
nloaded from
I
FIG. 1. IFA patterns produced by monoclonal Ab's (1 mg ofAb per ml unless other specified) on smearscontaining P. yoelii. Bar, 5 um. (A) The common pattern of bright, diffuse fluorescence produced by immuneserum and the majority of monoclonal Ab's examined in this study (pattern I). (B) Background fluorescencein the negative control slide (negative ascites fluid or normal mouse serum). (C) Pattern of fluorescenceproduced by Ab from hybridoma 4L-4 showing enhanced fluorescence around the membranes of largeparasites (pattern II). (D) Ab's from hybrid 6NL-9 bind to an Ag distributed throughout the cytoplasm of thering and trophozoite stages; however, this Ag is limited to a discrete area of individual merozoites within theschizont stage (compare with [A]). (E) Ab's from hybridoma 4L-3 stained all of the erythrocytic stages of P.yoelii (1 mg ofAb per ml). However, when smears were treated with 10 tiglml, fluorescence was observed onlyon schizonts and freed merozoites (pattern III). (F) Ab's from hybridoma 4L-3 preferentially bound to alocalized region of merozoites (10 jig of Ab per ml). (0) Ab's from hybridoma 6NL-10 were stage-specific formerozoites and schizonts containing merozoites (pattern IV). Other stages of P. yoelii present in the fieldshow low bachground levels of autofluorescence. (H) Ab's secreted by hybridoma 6NL- 10 detected an Agprobably located in the apical region of merozoites. Photograph of slide stained with hybridoma 6NL-10 ata concentration of 350 ig/ml.
566
on August 19, 2018 by guest
http://iai.asm.org/
Dow
nloaded from
MALARIAL ANTIGENS 567
FIG. 2. Unique patterns of immunofluorescence produced by antiplasmodial monoclonal Ab's. (A) Char-acteristic fluorescence pattern observed on a suspension of intact P. yoelii parasites freed from erythrocytesbefore IFA staining (IOOx). (B and C) Ab's from five hybridomas react with P. gallinaceum parasites innucleated chicken erythrocytes. Ab from 6NL-9 identified an Agpresent in all erythrocytic stages ofthis avianmalaria (B), but 6NL-10 (C) bound only to the schizont (merozoite) stage (40x). (D) P. yoelii sporozoitestreated with Ab from hybridoma 6NL-9 have distinct areas of bright fluorescence within the cytoplasm of thecell (lOOx). (E) Hybridoma 6NL-10 produced a pattern of fluorescence distinct from that of 6NL-9 withdominant areas offluorescence around the nuclear region (1OOx).
clearly produce distinguishable patterns of im-munofluorescence with both the sporozoite andthe erythrocytic stages.
Next, the hybrids were used to determinewhether the Ag's recognized by mice duringprimary malaria were species specific or cross-reactive. The results (Table 2) showed that 7 of18 hybrids detected species-specific Ag's, i.e.,Ag's possessed only by P. yoelii. Most of thesehybrids (six of seven) produced identical IFAstaining, both in terms of intensity and pattern,on the lethal and nonlethal substrains. However,hybridoma 4NL-3 routinely produced moder-ately stronger fluorescence on nonlethal, com-
pared with lethal, P. yoelii parasites. It is ofinterest that in this study, all species-specifichybrids were produced by using spleen cells fromanimals with 4-day lethal or nonlethal malaria.By the techniques employed in this study, it wasnot possible to differentiate these seven hybridsfurther. Based on the different patterns of fluo-rescence achieved by IFA staining, a minimumof two to three Ag's can be distinguished.The remaining 11 hybrids showed varying de-
grees of cross-reactivity with rodent, primate,and avian malaria species (Table 2). Some hy-brids (4L-7, 6NL-8, and 4L-5) showed limitedcross-reactivity, binding only to one or two
VOL. 32, 1981
on August 19, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

TABLE 2. Detection of cross-reactive antigens among various species of Sporozoa
P. yoelii P. P. cha- P. P. P falcip- P. galli- B. mi- T. gon-17XL 17XNL berghei baudi vinckei lesi arum naceum croti dii
4L-1 + + _ _ _ _ _ _ _ _4L-2 + + _ _ _ _ _ _ _ _4L-3 + + + + + + + - - -4L-4 + + - - - - - - - -4L-5 + + - - - - - - + -4L-6 + + - _ _ _ _ _ _ _4L-7 + + - + + - - - - -4L-8 + + + + + +
4NL-1 + + - - - - - - - -4NL-2 + + + + + + + + + -4NL-3 ±a +4NL-4 + + _ _ _ _ _ _ _
6NL-5 + + + + + + + + + -6NL-6 + + + + + + + + + -6NL-7 + + + ± + + + - - -6NL-8 + + + - ± - _ _ _ _6NL-9 + + + + + + + + ± -6NL-10 + + + + + + + + _ _
Immune serum + + + + + + + + +a ±, Weak cross-reactivity.
closely related rodent parasites. Three hybrids(4L-8, 4L-3, and 6NL-7) detected Ag's shared byrodent and primate malarias. However, five hy-brids demonstrated extensive reactivity by bind-ing to Ag(s) common to rodent and primatemalarias plus the avian parasite P. gallinaceum(Fig. 2B and C). Four of these hybrids alsoproduced Ab's that bound to Ag(s) of B. microti.Each of the 11 cross-reactive hybrids pro-
duced the same pattern of immunofluorescenceon various species of Plasmodium as that ini-tially described for P. yoelii. For example, hybrid6NL-10 was found to bind only to merozoitesbut not to the other erythrocytic stages of P.yoelii (pattern IV). Ab from hybrid 6NL-10 alsodetected an Ag restricted to merozoites of theother rodent, primate, and avian malarias tested(Fig. 20). Based on the results (Tables 1 and 2)plus data on sporozoite cross-reactivity, at least10 unique Ag's can be differentiated by the 11non-species-specific hybrids.
Neither immune serum nor the 18 monoclonalAb's showed serological cross-reactivity with T.gondii (Table 2).Location of P. yoelii Ag's. An attempt was
made to determine whether the Ag's recognizedby the monoclonal Ab's were (i) present on thesurface of erythrocytes from infected animals,(ii) on the surface of the parasite itself or itsassociated parasitophorous vacuolar membrane,(iii) contained within the cytoplasm of the par-
asite or whether a combination of these possi-bilities might be true. Thus, the 18 monoclonalAb's were surveyed by flow microfluorometryfor their ability to bind to the surface of eryth-rocytes from P. yoelii-infected animals. Al-though immune serum was observed to bind tothe surface of a population of erythrocytes frominfected animals (data not shown), none of themonoclonal Ab's examined possessed this char-acteristic. Similar results were obtained by con-ventional fluorescence microscopy.IFA staining of fixed smears containing ma-
larial parasites detects Ag's located on the sur-face and within the cytoplasm of parasites. Todetermine which, if any, of the 18 monoclonalAb's were directed against Ag's on the outermembrane surface of intact parasites, parasiteswere freed from erythrocytes and stained insuspension. With this technique, a typical pat-tern ofmembrane fluorescence was produced byusing immune serum (Fig. 2A). However, only 1of the 18 hybrid Ab's (4L-7) was determined torecognize Ag on the surface of intact parasites(Table 1). The remaining 17 hybrids, therefore,were assumed to bind to cytoplasmic Ags.
DISCUSSIONMonoclonal Ab's from 18 hybrid cell lines
were used to partially characterize Ag's of therodent malarial parasite, P. yoelii. By the tech-nique of indirect immunofluorescence, mono-
568 TAYLOR ET AL. INFECT. IMMUN.
on August 19, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

MALARIAL ANTIGENS 569
clonal Ab's were screened for their ability tobind to the erythrocytic and sporozoite stages ofP. yoelii and to the erythrocytic stages ofrelatedrodent, primate, and avian malarias. MonoclonalAb's also were tested for their ability to detectAg's on the surface of erythrocytes from infectedmice and on malarial parasites mechanically re-leased from erythrocytes. Based on results fromthis study, the 18 hybrids appeared to detect atleast 12 different antigenic determinants. Sev-eral of these Ag's were species specific, whereasothers showed varying degrees of serologicalcross-reactivity with other malarial parasites.Ag's were detected on the surface and within thecytoplasm of P. yoelii parasites, but not on thesurface of erythrocytes from infected animals.Two Ag's (6NL-9 + 6NL-10) were found to bepresent in both the erythrocytic and sporozoitestages of P. yoelii. Thus, BALB/c mice respondto a wide variety of parasite Ag's during theearly stages of P. yoelii infection.
Previous studies have demonstrated that atleast part of the protective immune response ofthe host is directed against stage-specific Ag's.This point has been well established recently intwo studies with monoclonal Ab's. Yoshida et al.used monoclonal Ab's to demonstrate that asingle Ag (Pb44) on the surface of P. bergheisporozoites is important in sporozoite immunity(28). Monoclonal Ab to this stage-specific Agneutralizes sporozoites in vitro and blocks spo-rozoite infectivity in vivo. In another study,Freeman et al. showed that monoclonal Ab to amerozoite-specific Ag of P. yoelii is capable ofpassively protecting mice from lethal P. yoeliimalaria (11). Although mice passively treatedwith monoclonal Ab developed parasitemias, thecourse of infection was similar to that observedin mice with nonlethal malaria.One stage-specific Ag (6NL-10) was identified
in our study. This merozoite-specific (Ag) wasnot species specific as monoclonal Ab 6NL-10bound to merozoites of all ofthe rodent, primate,and avian malarial parasites tested. Our prelim-inary results suggest that Ab from 6NL-10 is notprotective and that the Ag may be associatedwith an internal merozoite organelle. As tropho-zoites develop into merozoites, nuclear struc-tures, rhoptries, micronemes, and a surface coatare formed (1, 17, 20). Since Ab from 6NL-10 (i)did not bind to the surface of merozoites, (ii) wasrestricted primarily to a definite area of mero-zoites, and (iii) bound to two areas of sporozoites(which also possess rhoptries and similar acces-sory organelles), Ag 6NL-10 is probably associ-ated with structures involved in nuclear divisionor organelles important in cell penetration. How-ever, the observation thatBALB/c mice infected
with P. yoelii 17XNL for 6 days produce an Abwhich binds to a "universal" merozoite Ag is aninteresting finding that deserves further inves-tigation.One predicted use for monoclonal Ab's is for
taxonomic studies of malarial parasites. Duringthe evolutionary adaptation ofmalarial parasitesto various hosts, some Ag's have been retained,i.e., conserved, whereas others have been ac-quired and develop as species-species Ag's. Var-ious investigators have demonstrated serologicalcross-reactivity among the rodent malarial par-asites P. berghei, P. chabaudi, and P. vinckei(5, 6, 9), among primate malarias (4), and be-tween human and primate (25) and human andavian malarias (14). However, it has not beenpossible (i) to establish how many Ag's are in-volved in serological cross-reactivity, (ii) to de-termine whether common Ag's have taxonomicsignificance, or (iii) to determine whether cross-reactive determinants are located on homolo-gous molecules, i.e., molecules with equivalentbiological functions. Some of these questionsmay be approached by hybridoma technology.Our results suggest that serological cross-reac-tivity may be due to numerous antigenic deter-minants. For example, all of the rodent, primate,and avian malarial parasites we tested were ca-pable of binding Ab from three hybridomaswhich have different antigenic specificities([4NL-2 + 6NL-5], [6NL-6 + 6NL-9], and 6NL-10). One cannot conclude, however, that theobserved serological cross-reactivity denotes thepresence of homologous molecules in the aboveparasites. Ag's which bind monoclonal Ab's mustbe isolated from each species of malarial para-sites and biochemically analyzed before taxo-nomic interrelationships can be established.
In this investigation, serological cross-reactiv-ity studies were used to help differentiate theantigenic specificity of antimalarial hybrid celllines. Currently, monoclonal Ab's from thesehybridomas are being used to isolate and furthercharacterize Ag's which induce humoral immuneresponses in malaria-infected mice.
ACKNOWLEDGMENTSWe thank the following people for their contributions to
the project: T. Chused for flow microfluorometric analysis ofparasitized erythrocytes, Mary Leef for isolation of P. yoeliisporozoites, F. Rollwagen and G. D. Wallace for criticallyreviewing the manuscript, and V. Shaw and the NationalInstitute of Allergy and Infectious Diseases Editorial Officefor help in preparation of the manuscript.
LITERATURE CITED
1. Aikawa, M., and C. Sterling. 1974. Intracellular para-sitic protozoa. Academic Press, Inc., New York.
2. Andersson, J., A. Coutinho, W. Lernhardt, and F.Melcher. 1977. Clonal growth and maturation to im-
VOL. 32, 1981
on August 19, 2018 by guest
http://iai.asm.org/
Dow
nloaded from

570 TAYLOR ET AL.
munoglobulin secretion in vitro of every growth-induc-ible B lymphocyte. Cell 10:27-34.
3. Cohen, S. 1979. Immunity to malaria. Proc. R. Soc. Lon-don Ser. B 203:323-345.
4. Collins, W. E., J. C. Skinner, and E. G. Guinn. 1966.Antigenic variations in the plasmodia of lower pimatesas detected by immuno-fluorescence. Am. J. Trop. Med.Hyg. 15:483-485.
5. Cox, F. E. G. 1970. The specificity of immunoglobulin Gand immunoglobulin M in the fluorescent-antibody testfor malarial parasites in mice. Bull. W.H.O. 43:341-344.
6. Cox, F. E. G., and S. A. Turner. 1970. Antigenic rela-tionships between the malaria parasites and piroplasmsof mice as determined by the fluorescent-antibody tech-nique. Bull. W.H.O. 43:337-340.
7. Deans, J. A., E. D. Dennis, and S. Cohen. 1978. Anti-genic analysis of sequential erythrocytic stages of Plas-modium knowlesi. Parasitology 77:333-344.
8. Eby, W. C., B. S. Kim, S. Dray, G. 0. Young-Copper,and R. G. Mage. 1973. Detection of the e14 and el5rabbit allotypic specificities by immunodiffusion in PEGagar. Immunocheniistry 10:417-418.
9. El-Nahal, H. M. S. 1967. Serological cross-reaction be-tween rodent malaria parasites as determined by theindirect immunofluorescent technique. Bull. W.H.O.36:423-429.
10. Ey, P. L., S. J. Prowse, and C. R. Jenkin. 1978. Isolationof pure IgG,, IgG2., and IgG2b immunoglobulins frommouse serum using protein A-Sepharose. Immuno-chemistry 15:429-436.
11. Freeman, R. R., A. J. Trejdosiewicz, and G. A. M.Cross. 1980. Protective monoclonal antibodies recog-nising stage-specific merozoite antigens of a rodent ma-laria parasite. Nature (London) 284:366-368.
12. Howard, R. J., P. M. Smith, and G. F. Mitchell. 1978.Removal of leucocytes from red blood cells in Plasmo-dium berghei-infected mouse blood and purification ofschizont-infected cells. Ann. Trop. Med. Parasitol. 72:573-576.
13. Jacobs, L., and M. N. Lunde. 1957. A hemagglutinationtest for toxoplasmosis. J. Parasitol. 43:308-314.
14. Kielmann, A., and N. Weiss. 1968. Plasmodium galli-naceum as antigen in immunofluorescence antibodystudies. Acta Trop. 25:185-187.
15. Kim, K. J., D. W. Taylor, C. B. Evans, and R. Asofsky.1980. Radioimmunoassay for detecting antibodiesagainst murine malarial parasite antigens: monoclonalantibodies recognizing Plasmodium yoelii antigens. J.Immunol. 125:2565-2569.
16. Kuvin, S. F., J. E. Tobie, C. B. Evans, C. P. Coatney,and P. G. Contacos. 1962. Antibody production in
human malaria as determined by the fluorescence an-tibody technique. Science 135:1130-1131.
17. Langreth, S. G., J. B. Jensen, R. T. Reese, and W.Trager. 1978. Fine structure ofhuman malaria in vitro.J. Protozool. 25:443-452.
18. Langreth, S. G., and R. T. Reese. 1979. Antigenicity ofthe infected-erythrocyte and merozoite surfaces in Fal-ciparum malaria. J. Exp. Med. 150:1241-1254.
19. Margulies, D. H., W. Cieplinski, B. Dharmgrongar-tama, M. L. Gefter, S. L. Morrision, T. Kelly, andM. D. Scharff. 1977. Regulation of immunoglobulinexpression in mouse myeloma cells. Cold Spring HarborSymp. Quant. Biol. 41:781-791.
20. Miller, L. H., M. Aikawa, and J. F. Dvorak. 1975.Malaria (Plasmodium knowlesi) merozoites: immunityand the surface coat. J. Immunol. 114:1237-1242.
21. Mitchell, G. H., G. A. Butcher, and S. Cohen. 1974. Amerozoite vaccine effective against Plasmodium know-lesi malaria. Nature (London) 252:311-313.
22. Pierce, C. W., B. H. Johnson, H. E. Gershon, and R.Asofsky. 1971. Immune response in vitro. III. Devel-opment of primary yM, yG, and -yA plaque-forming cellresponses in mouse spleen cell cultures stimulated withheterologous erythrocytes. J. Exp. Med. 134:395-416.
23. Sabin, A. B., and H. A. Feldman. 1948. Dyes as micro-bial indicators of new immunity phenomenon affectinga protozoan parasite (Toxoplasma). Science 108:660.
24. Schmidt-Ullrich, R., D. F. H. Wallach, and J. Light-holder. 1979. Two Plasmodium knowlesi-specific an-tigens on the surface ofschizont-infected rhesus monkeyerythrocytes induce antibody production in immunehosts. J. Exp. Med. 150:86-99.
25. Tobie, J. E., S. F. Kuvin, P. G. Contacos, G. R. Coat-ney, and C. B. Evans. 1962. Fluorescent antibodystudies on cross reactions between human and simianmalaria in normal volunteers. Am. J. Trop. Med. Hyg.11:589-596.
26. Weinbaum, F. I., C. B. Evans, and R. E. Tigelaar.1976. An in vitro assay for T cell immunity to malariain mice. J. Immunol. 116:1280-1283.
27. Weinbaum, F. I., J. Weintraub, F. K. Nkrumah, C. B.Evans, R. E. Tigelaar, and Y. J. Rosenberg. 1978.Immunity to Plasmodium berghei yoelii in mice. II.Specific and nonspecific cellular and humoral responsesduring the course of infection. J. Immunol. 121:629-636.
28. Yoshida, N., R. S. Nussenzweig, P. Potocnjak, V.Nussenzweig, and M. Aikawa. 1980. Hybridoma pro-duces protective antibodies directed against the sporo-zoite stage of malaria parasite. Science 207:71-73.
INFECT. IMMUN.
on August 19, 2018 by guest
http://iai.asm.org/
Dow
nloaded from





























