Embed Size (px)
Citation preview

Morphological Diversity of Anatomical Strepsirrhinismand the Evolution of the Lemuriform Toothcomb
ROBERT J. ASHER*Doctoral Program in Anthropological Sciences, State University ofNew York, Stony Brook, New York 11794
KEY WORDS rostrum; vomeronasal organ; grooming; primates
ABSTRACT The hypothesis that the vomeronasal organ has an impor-tant functional relationship with, and led to the evolution of, the prosimiantoothcomb has not been well tested. This paper examines the diversity ofanatomical strepsirrhinism across several mammalian taxa to determine iffossil and living strepsirrhine primates exhibit any derived characters thatmay highlight the functional link between the vomeronasal organ and thetoothcomb, and to examine the potential importance of anatomical strepsir-rhinism to toothcomb origins. Results indicate that extant gregarious lemuri-forms are derived in having a relatively wide interincisal gap, providing anunobstructed line of communication between the vomeronasal organ andanterior rostral structures such as the toothcomb. This finding is consistentwith the proposal that anatomical strepsirrhinism is functionally related touse of the toothcomb in grooming. However, the importance of the vomerona-sal organ to toothcomb origins is less clear. If the morphology of adapiformsand non-gregarious lemuriforms is representative of the morphology of basallemuriforms, then it can be inferred that early lemuriforms did not possessthe wide-gap autapomorphy; hence, anatomical evidence discussed herecannot be used to rule out non-social hypotheses of toothcomb origins. Am JPhys Anthropol 105:355–367, 1998. r 1998 Wiley-Liss, Inc.
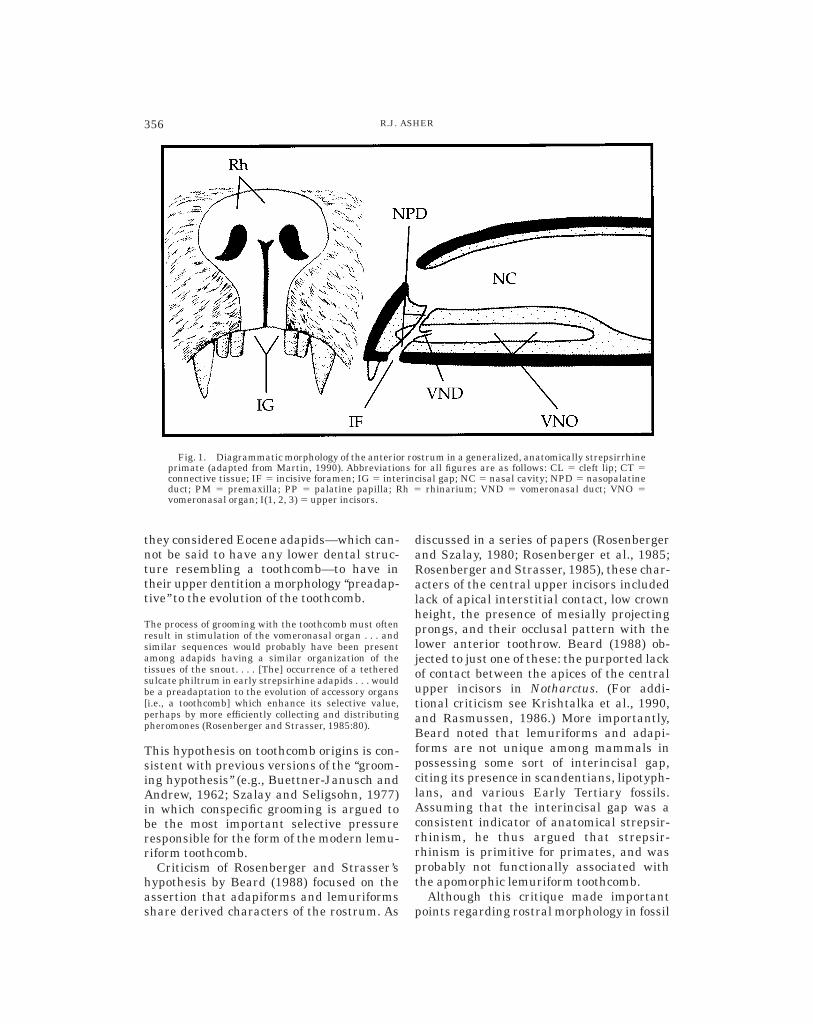
The classification of primates into thesuborders Haplorhini and Strepsirrhini (Po-cock, 1918) is nominally based on the mor-phology of the nasal region. The word ‘‘strep-sirrhine’’ literally refers to the ‘‘twisted’’appearance of the external nares (Hofer,1976; I follow Jenkins, 1987, for spelling of‘‘strepsirrhine’’ and lemuriform familialnames). However, as used by most workers(e.g., Maier, 1991), anatomical strepsir-rhinism indicates the continuity of the up-per lip with the vomeronasal organ (alsoknown as Jacobson’s organ; see Fig. 1).
The morphology of the anterior rostrum instrepsirrhine primates led Rosenberger andStrasser (1985) to propose a functional linkbetween the vomeronasal organ and thetoothcomb of lemuriform primates. Based onresearch that supports the role of the vomero-
nasal organ in sociosexual communication(Estes, 1972; Verberne, 1976), and on theconnection between the rhinarium and vom-eronasal organ via the gap between thecentral upper incisors (Hofer, 1977), theseresearchers hypothesized that the lemuri-form toothcomb evolved as a sensory exten-sion of the vomeronasal organ, serving as atool for pheromone collection.
Rosenberger and Strasser (1985) furtherargued that the upper anterior dentition ofEocene adapid primates closely resemblesthat of modern lemuriforms and would havepermitted the same continuity between theupper lip and vomeronasal organ. Hence,
*Correspondence to: Robert Asher, Department of Anthropol-ogy, State University of New York, Stony Brook, NY 11794-4364.Fax: (516) 444-3947. E-mail: [email protected]
Received 9 September 1996; accepted 22 November 1997.
AMERICAN JOURNAL OF PHYSICAL ANTHROPOLOGY 105:355–367 (1998)
r 1998 WILEY-LISS, INC.
Article ID # 920@xyserv1/disk8/CLS_liss/GRP_phan/JOB_phan105-3/DIV_920z16 dawn
they considered Eocene adapids—which can-not be said to have any lower dental struc-ture resembling a toothcomb—to have intheir upper dentition a morphology ‘‘preadap-tive’’ to the evolution of the toothcomb.
The process of grooming with the toothcomb must oftenresult in stimulation of the vomeronasal organ . . . andsimilar sequences would probably have been presentamong adapids having a similar organization of thetissues of the snout. . . . [The] occurrence of a tetheredsulcate philtrum in early strepsirhine adapids . . . wouldbe a preadaptation to the evolution of accessory organs[i.e., a toothcomb] which enhance its selective value,perhaps by more efficiently collecting and distributingpheromones (Rosenberger and Strasser, 1985:80).
This hypothesis on toothcomb origins is con-sistent with previous versions of the ‘‘groom-ing hypothesis’’ (e.g., Buettner-Janusch andAndrew, 1962; Szalay and Seligsohn, 1977)in which conspecific grooming is argued tobe the most important selective pressureresponsible for the form of the modern lemu-riform toothcomb.
Criticism of Rosenberger and Strasser’shypothesis by Beard (1988) focused on theassertion that adapiforms and lemuriformsshare derived characters of the rostrum. As
discussed in a series of papers (Rosenbergerand Szalay, 1980; Rosenberger et al., 1985;Rosenberger and Strasser, 1985), these char-acters of the central upper incisors includedlack of apical interstitial contact, low crownheight, the presence of mesially projectingprongs, and their occlusal pattern with thelower anterior toothrow. Beard (1988) ob-jected to just one of these: the purported lackof contact between the apices of the centralupper incisors in Notharctus. (For addi-tional criticism see Krishtalka et al., 1990,and Rasmussen, 1986.) More importantly,Beard noted that lemuriforms and adapi-forms are not unique among mammals inpossessing some sort of interincisal gap,citing its presence in scandentians, lipotyph-lans, and various Early Tertiary fossils.Assuming that the interincisal gap was aconsistent indicator of anatomical strepsir-rhinism, he thus argued that strepsir-rhinism is primitive for primates, and wasprobably not functionally associated withthe apomorphic lemuriform toothcomb.
Although this critique made importantpoints regarding rostral morphology in fossil
Fig. 1. Diagrammatic morphology of the anterior rostrum in a generalized, anatomically strepsirrhineprimate (adapted from Martin, 1990). Abbreviations for all figures are as follows: CL 5 cleft lip; CT 5connective tissue; IF 5 incisive foramen; IG 5 interincisal gap; NC 5 nasal cavity; NPD 5 nasopalatineduct; PM 5 premaxilla; PP 5 palatine papilla; Rh 5 rhinarium; VND 5 vomeronasal duct; VNO 5vomeronasal organ; I(1, 2, 3) 5 upper incisors.
356 R.J. ASHER
Article ID # 920@xyserv1/disk8/CLS_liss/GRP_phan/JOB_phan105-3/DIV_920z16 dawn

primates, Beard’s response to Rosenbergerand Strasser (1985) did not adequately exam-ine their proposal that the vomeronasalorgan is relevant to the function of thetoothcomb. Beard (1988) asserted that thepossession of a space between the centralupper incisors in various eutherian ordersindicates that the rostrum in lemuriforms isplesiomorphic. However, in addition to sup-porting the point that many groups of mam-mals have some kind of gap between theircentral upper incisors, the data collected—but not quantified—by Beard (1988) on rela-tive gap size in various eutherians also showconsiderable differences among those thatare anatomically strepsirrhine. These differ-ences, and their potential relevance forRosenberger and Strasser’s theory of tooth-comb evolution, have yet to be explored.
It is therefore my intent in this paper toreexamine Rosenberger and Strasser’s pro-posal that the vomeronasal organ in pri-mates is relevant to the function and evolu-tion of the toothcomb. In order to accomplishthis, I will ask two questions related to thepolarity of the rostrum in toothcomb prosim-ians: 1) is anatomical strepsirrhinism homo-geneous in primates and other mammals;and 2) are there data supporting Rosen-berger and Strasser’s proposal that the ros-trum in lemuriforms and adapiforms is auta-pomorphic? If the answers to these questionsare, respectively, yes and no, then Beard’s(1988) criticism of Rosenberger and Stras-ser’s hypothesis is justified. Otherwise, theirproposal may be relevant to toothcomb evo-lution at some level.
MATERIALS AND METHODS
In order to assess the first question listedabove, the palatal region of Eulemur fulvus(Primates), Tupaia minor (Scandentia), Bla-rina brevicauda (Lipotyphla), Felis catus(Carnivora), and Ovis aries (Artiodactyla)was dissected and dry skulls of genera in allof the preceding ordinal taxa were exam-ined. Histologically prepared cross sectionsof Blarina were also used to document ros-tral morphology in that taxon. Soft-tissuespecimens examined in this study were ob-tained from the Division of Laboratory Ani-mal Resources (DLAR) and Anatomical Sci-ences (USBA) at the State University of
New York, Stony Brook, except for a speci-men of Tupaia minor, which is in the collec-tions of the Department of Mammalogy,American Museum of Natural History(AMNH), New York.
In order to address the second questionlisted above, I focused on the means bywhich the vomeronasal organ communicateswith the external environment in primates,i.e., the upper interincisal gap (Hofer, 1977).In outlining their hypothesis, Rosenbergerand Strasser (1985) emphasized that thegap between the upper incisors allows foreffective communication between the tooth-comb and vomeronasal organ. Thus, if Rosen-berger and Strasser’s proposed associationbetween grooming and the vomeronasal or-gan is correct, then one might predict thatlemuriforms that use the toothcomb mostoften for grooming would have a relativelywider channel between their central upperincisors than other mammals. That is, if oneassumes that gregarious lemuriforms (e.g.,Eulemur), due to their complex social struc-ture and relatively frequent contact withconspecifics (Richard, 1987), allogroom morefrequently than non-gregarious mammals(e.g., Loris), then one would expect the gre-garious forms to possess a wider interincisalgap than other mammals.
To test these predictions, I measured theraw interincisal gap (i.e., the shortest dis-tance in millimeters between the alveoli ofthe central upper incisors) across strepsir-rhine mammals listed in Table 1. This mea-surement was scaled using two indices: thewidth of the palate at M1 (i.e., the shortestdistance in millimeters between the lateralmargins of the mesiobuccal cusps of the twoupper M1s), and the estimated area of thefirst upper molar (i.e., the product of maxi-mum mesiodistal and buccolingual dimen-sions). Raw measurements for all individualspecimens are reported in Table 1. Measure-ments for extant taxa were obtained fromcrania housed in the Departments of Mam-malogy at the AMNH and the Museum ofComparative Zoology (MCZ), Harvard Uni-versity. Measurements for fossil taxa wereobtained from casts in the USBA collectionand from Beard (1988). Statistical compari-sons were facilitated using the JMP statisti-cal program for the Macintosh (Sall et al.,
357STREPSIRRHINISM AND TOOTHCOMB EVOLUTION
Article ID # 920@xyserv1/disk8/CLS_liss/GRP_phan/JOB_phan105-3/DIV_920z16 dawn
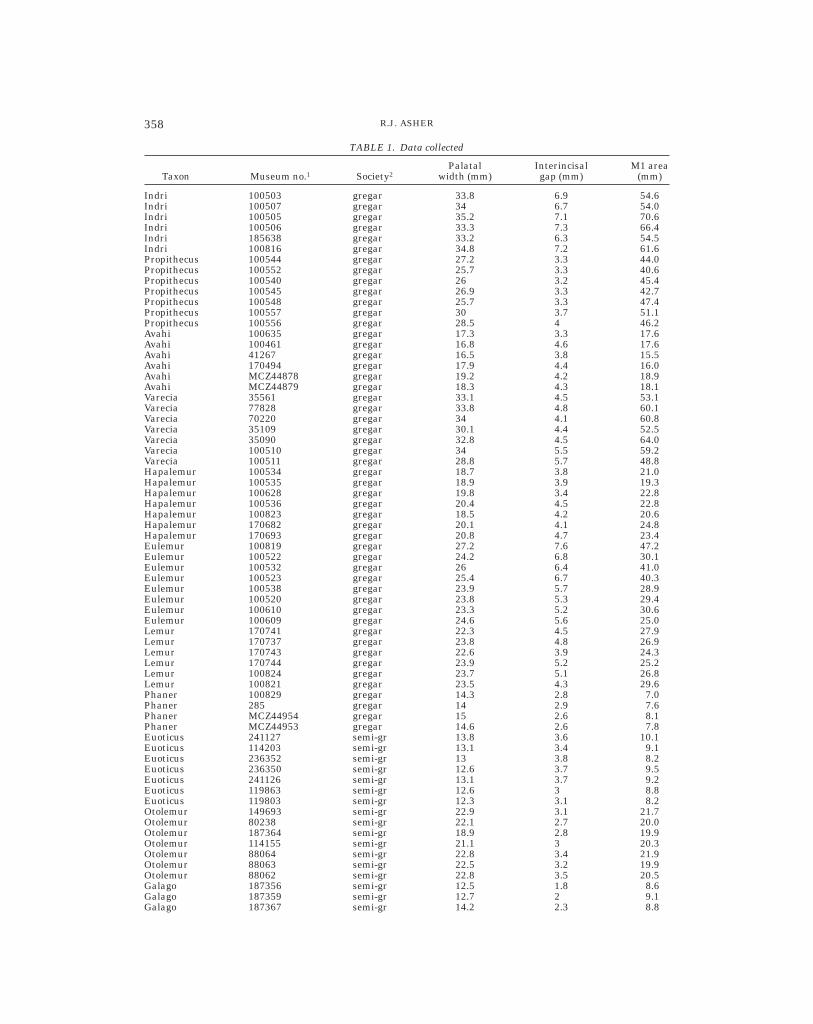
TABLE 1. Data collected
Taxon Museum no.1 Society2Palatal
width (mm)Interincisalgap (mm)
M1 area(mm)
Indri 100503 gregar 33.8 6.9 54.6Indri 100507 gregar 34 6.7 54.0Indri 100505 gregar 35.2 7.1 70.6Indri 100506 gregar 33.3 7.3 66.4Indri 185638 gregar 33.2 6.3 54.5Indri 100816 gregar 34.8 7.2 61.6Propithecus 100544 gregar 27.2 3.3 44.0Propithecus 100552 gregar 25.7 3.3 40.6Propithecus 100540 gregar 26 3.2 45.4Propithecus 100545 gregar 26.9 3.3 42.7Propithecus 100548 gregar 25.7 3.3 47.4Propithecus 100557 gregar 30 3.7 51.1Propithecus 100556 gregar 28.5 4 46.2Avahi 100635 gregar 17.3 3.3 17.6Avahi 100461 gregar 16.8 4.6 17.6Avahi 41267 gregar 16.5 3.8 15.5Avahi 170494 gregar 17.9 4.4 16.0Avahi MCZ44878 gregar 19.2 4.2 18.9Avahi MCZ44879 gregar 18.3 4.3 18.1Varecia 35561 gregar 33.1 4.5 53.1Varecia 77828 gregar 33.8 4.8 60.1Varecia 70220 gregar 34 4.1 60.8Varecia 35109 gregar 30.1 4.4 52.5Varecia 35090 gregar 32.8 4.5 64.0Varecia 100510 gregar 34 5.5 59.2Varecia 100511 gregar 28.8 5.7 48.8Hapalemur 100534 gregar 18.7 3.8 21.0Hapalemur 100535 gregar 18.9 3.9 19.3Hapalemur 100628 gregar 19.8 3.4 22.8Hapalemur 100536 gregar 20.4 4.5 22.8Hapalemur 100823 gregar 18.5 4.2 20.6Hapalemur 170682 gregar 20.1 4.1 24.8Hapalemur 170693 gregar 20.8 4.7 23.4Eulemur 100819 gregar 27.2 7.6 47.2Eulemur 100522 gregar 24.2 6.8 30.1Eulemur 100532 gregar 26 6.4 41.0Eulemur 100523 gregar 25.4 6.7 40.3Eulemur 100538 gregar 23.9 5.7 28.9Eulemur 100520 gregar 23.8 5.3 29.4Eulemur 100610 gregar 23.3 5.2 30.6Eulemur 100609 gregar 24.6 5.6 25.0Lemur 170741 gregar 22.3 4.5 27.9Lemur 170737 gregar 23.8 4.8 26.9Lemur 170743 gregar 22.6 3.9 24.3Lemur 170744 gregar 23.9 5.2 25.2Lemur 100824 gregar 23.7 5.1 26.8Lemur 100821 gregar 23.5 4.3 29.6Phaner 100829 gregar 14.3 2.8 7.0Phaner 285 gregar 14 2.9 7.6Phaner MCZ44954 gregar 15 2.6 8.1Phaner MCZ44953 gregar 14.6 2.6 7.8Euoticus 241127 semi-gr 13.8 3.6 10.1Euoticus 114203 semi-gr 13.1 3.4 9.1Euoticus 236352 semi-gr 13 3.8 8.2Euoticus 236350 semi-gr 12.6 3.7 9.5Euoticus 241126 semi-gr 13.1 3.7 9.2Euoticus 119863 semi-gr 12.6 3 8.8Euoticus 119803 semi-gr 12.3 3.1 8.2Otolemur 149693 semi-gr 22.9 3.1 21.7Otolemur 80238 semi-gr 22.1 2.7 20.0Otolemur 187364 semi-gr 18.9 2.8 19.9Otolemur 114155 semi-gr 21.1 3 20.3Otolemur 88064 semi-gr 22.8 3.4 21.9Otolemur 88063 semi-gr 22.5 3.2 19.9Otolemur 88062 semi-gr 22.8 3.5 20.5Galago 187356 semi-gr 12.5 1.8 8.6Galago 187359 semi-gr 12.7 2 9.1Galago 187367 semi-gr 14.2 2.3 8.8
358 R.J. ASHER
Article ID # 920@xyserv1/disk8/CLS_liss/GRP_phan/JOB_phan105-3/DIV_920z16 dawn
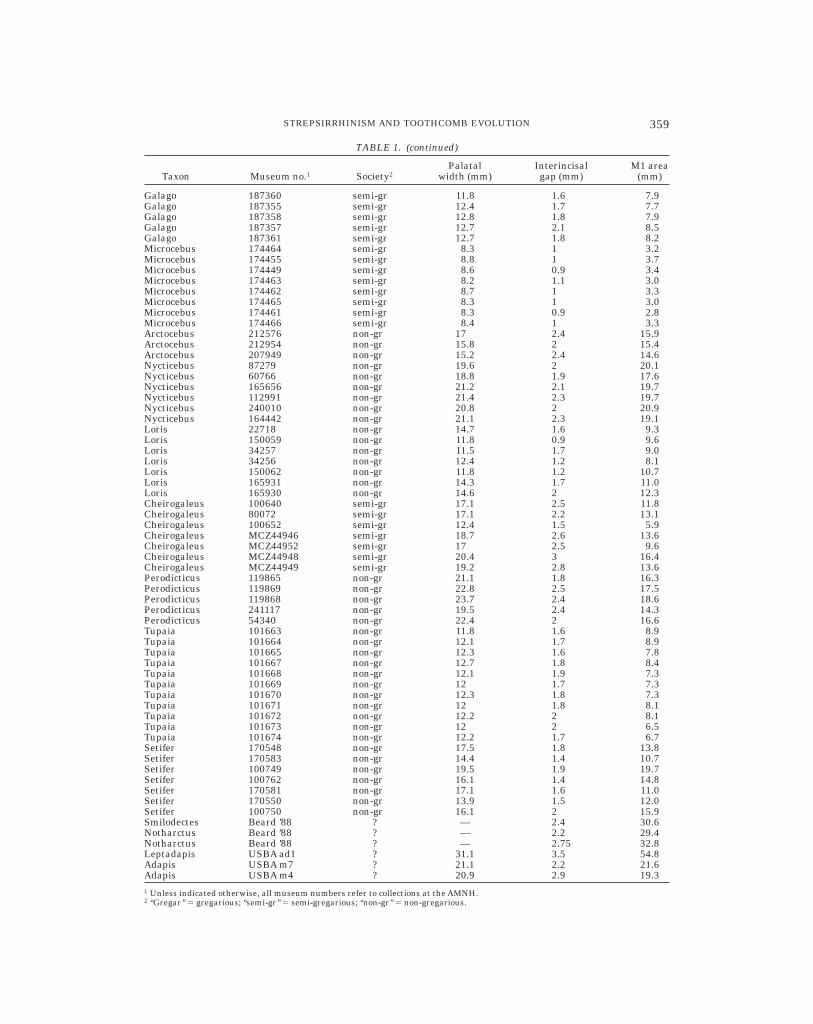
TABLE 1. (continued)
Taxon Museum no.1 Society2Palatal
width (mm)Interincisalgap (mm)
M1 area(mm)
Galago 187360 semi-gr 11.8 1.6 7.9Galago 187355 semi-gr 12.4 1.7 7.7Galago 187358 semi-gr 12.8 1.8 7.9Galago 187357 semi-gr 12.7 2.1 8.5Galago 187361 semi-gr 12.7 1.8 8.2Microcebus 174464 semi-gr 8.3 1 3.2Microcebus 174455 semi-gr 8.8 1 3.7Microcebus 174449 semi-gr 8.6 0.9 3.4Microcebus 174463 semi-gr 8.2 1.1 3.0Microcebus 174462 semi-gr 8.7 1 3.3Microcebus 174465 semi-gr 8.3 1 3.0Microcebus 174461 semi-gr 8.3 0.9 2.8Microcebus 174466 semi-gr 8.4 1 3.3Arctocebus 212576 non-gr 17 2.4 15.9Arctocebus 212954 non-gr 15.8 2 15.4Arctocebus 207949 non-gr 15.2 2.4 14.6Nycticebus 87279 non-gr 19.6 2 20.1Nycticebus 60766 non-gr 18.8 1.9 17.6Nycticebus 165656 non-gr 21.2 2.1 19.7Nycticebus 112991 non-gr 21.4 2.3 19.7Nycticebus 240010 non-gr 20.8 2 20.9Nycticebus 164442 non-gr 21.1 2.3 19.1Loris 22718 non-gr 14.7 1.6 9.3Loris 150059 non-gr 11.8 0.9 9.6Loris 34257 non-gr 11.5 1.7 9.0Loris 34256 non-gr 12.4 1.2 8.1Loris 150062 non-gr 11.8 1.2 10.7Loris 165931 non-gr 14.3 1.7 11.0Loris 165930 non-gr 14.6 2 12.3Cheirogaleus 100640 semi-gr 17.1 2.5 11.8Cheirogaleus 80072 semi-gr 17.1 2.2 13.1Cheirogaleus 100652 semi-gr 12.4 1.5 5.9Cheirogaleus MCZ44946 semi-gr 18.7 2.6 13.6Cheirogaleus MCZ44952 semi-gr 17 2.5 9.6Cheirogaleus MCZ44948 semi-gr 20.4 3 16.4Cheirogaleus MCZ44949 semi-gr 19.2 2.8 13.6Perodicticus 119865 non-gr 21.1 1.8 16.3Perodicticus 119869 non-gr 22.8 2.5 17.5Perodicticus 119868 non-gr 23.7 2.4 18.6Perodicticus 241117 non-gr 19.5 2.4 14.3Perodicticus 54340 non-gr 22.4 2 16.6Tupaia 101663 non-gr 11.8 1.6 8.9Tupaia 101664 non-gr 12.1 1.7 8.9Tupaia 101665 non-gr 12.3 1.6 7.8Tupaia 101667 non-gr 12.7 1.8 8.4Tupaia 101668 non-gr 12.1 1.9 7.3Tupaia 101669 non-gr 12 1.7 7.3Tupaia 101670 non-gr 12.3 1.8 7.3Tupaia 101671 non-gr 12 1.8 8.1Tupaia 101672 non-gr 12.2 2 8.1Tupaia 101673 non-gr 12 2 6.5Tupaia 101674 non-gr 12.2 1.7 6.7Setifer 170548 non-gr 17.5 1.8 13.8Setifer 170583 non-gr 14.4 1.4 10.7Setifer 100749 non-gr 19.5 1.9 19.7Setifer 100762 non-gr 16.1 1.4 14.8Setifer 170581 non-gr 17.1 1.6 11.0Setifer 170550 non-gr 13.9 1.5 12.0Setifer 100750 non-gr 16.1 2 15.9Smilodectes Beard ’88 ? — 2.4 30.6Notharctus Beard ’88 ? — 2.2 29.4Notharctus Beard ’88 ? — 2.75 32.8Leptadapis USBA ad1 ? 31.1 3.5 54.8Adapis USBA m7 ? 21.1 2.2 21.6Adapis USBA m4 ? 20.9 2.9 19.31 Unless indicated otherwise, all museum numbers refer to collections at the AMNH.2 ‘‘Gregar’’ 5 gregarious; ‘‘semi-gr’’ 5 semi-gregarious; ‘‘non-gr’’ 5 non-gregarious.
359STREPSIRRHINISM AND TOOTHCOMB EVOLUTION
Article ID # 920@xyserv1/disk8/CLS_liss/GRP_phan/JOB_phan105-3/DIV_920z16 dawn

1994). Unless stated otherwise, statementsof statistical difference are based on thenonparametric Kruskal-Wallis test (see So-kal and Rohlf, 1995).
Generalizations about social behavior ingroups of mammals are difficult to make.Nevertheless, for the purposes of the pres-ent study three different categories wererecognized: First, taxa that habitually spendactive time in the presence of one or moreconspecifics were designated ‘‘gregarious;’’this category includes lemurids and indrids(Richard, 1987), plus the cheirogaleid Phaner(Charles-Dominique, 1978). Second, taxathat often forage alone but typically formsleeping groups containing multiple adultswere designated ‘‘semi-gregarious;’’ this cat-egory includes galagonids, Cheirogaleus, andMicrocebus (Bearder, 1987; Charles-Domi-nique, 1977). Third, taxa that forage aloneand do not typically form sleeping groupscontaining more than mother and offspringwere designated ‘‘non-gregarious;’’ this cat-egory includes lorids, Tupaia, and Setifer(Bearder, 1987; Charles-Dominique, 1977;Eisenberg and Gould, 1970; Nowak, 1991).
RESULTSIs anatomical strepsirrhinism
homogeneous across mammals?
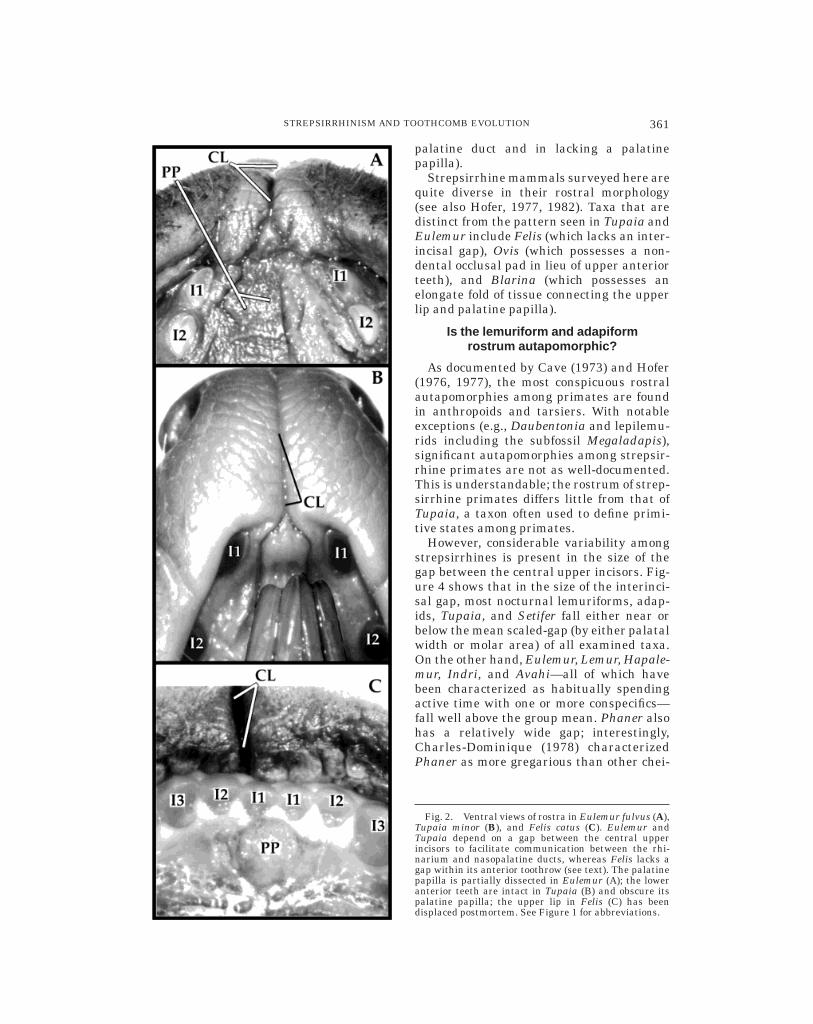
In Eulemur fulvus and Tupaia minor, thepath between the nasopalatine ducts andupper lip is folded to provide greater surfacearea, permitting material to flow freely be-tween the oral cavity and cleft upper lip(Figs. 2A, 2B). In both taxa, the upperinterincisal gap clearly facilitates the connec-tion between the vomeronasal organ andrhinarium. Microcebus murinus has a simi-lar morphology (Schilling, 1970); in thistaxon, the anterior lumina of the pairedvomeronasal organ are connected to thenasopalatine canals via short vomeronasalducts. The nasopalatine canals, in turn,connect the nasal and oral cavities, ven-trally opening up into ducts on either side ofthe palatine papilla. In cross section, thepalatine papilla is an inverted mushroom-shaped structure (Schilling, 1970) that al-lows Eulemur, Microcebus, and Tupaia ameans of closing the nasopalatine canals bypressing up against the papilla with thetongue. A stereotyped behavior involving the
tongue and lip (known as ‘‘flehmen’’) ob-served in Lemur catta probably augmentsthe transport of substances from the ante-rior rostrum to the vomeronasal organ (Bai-ley, 1978).
The domestic cat has a cleft upper lip,moist rhinarium, patent nasopalatine ca-nals, palatine papilla (Fig. 2C) and a well-developed vomeronasal organ (Verberne,1976). By virtue of these characters, it isstrepsirrhine. Nevertheless, cats have atightly packed row of incisors, allowing forno gaps within the upper anterior toothrow.Examination of dry skulls of canid, mus-telid, procyonid, ursid, viverrid, and othercarnivoran taxa also reveals the absence of agap in the upper anterior toothrow. Theoccurrence of flehmen in Felis (Verberne,1976) probably provides a method of trans-porting substances from the upper lip to thenasopalatine canals, circumventing the ante-rior dentition.
Bovids lack upper anterior teeth alto-gether; instead, they possess a tough occlu-sal pad that can act with the lower dentitionto crop vegetation. Ovis (not figured) has acleft lip, well-developed vomeronasal organ,and a slight depression in the midline of theanterior palate that may serve as a conduitbetween the upper lip and the palatinepapilla. However, this depression provides arelatively longer and more indirect pathbetween the palatine papilla and upper lipthan the median interincisal gap seen inEulemur and Tupaia. Estes (1972) describedin detail flehmen in ungulates.
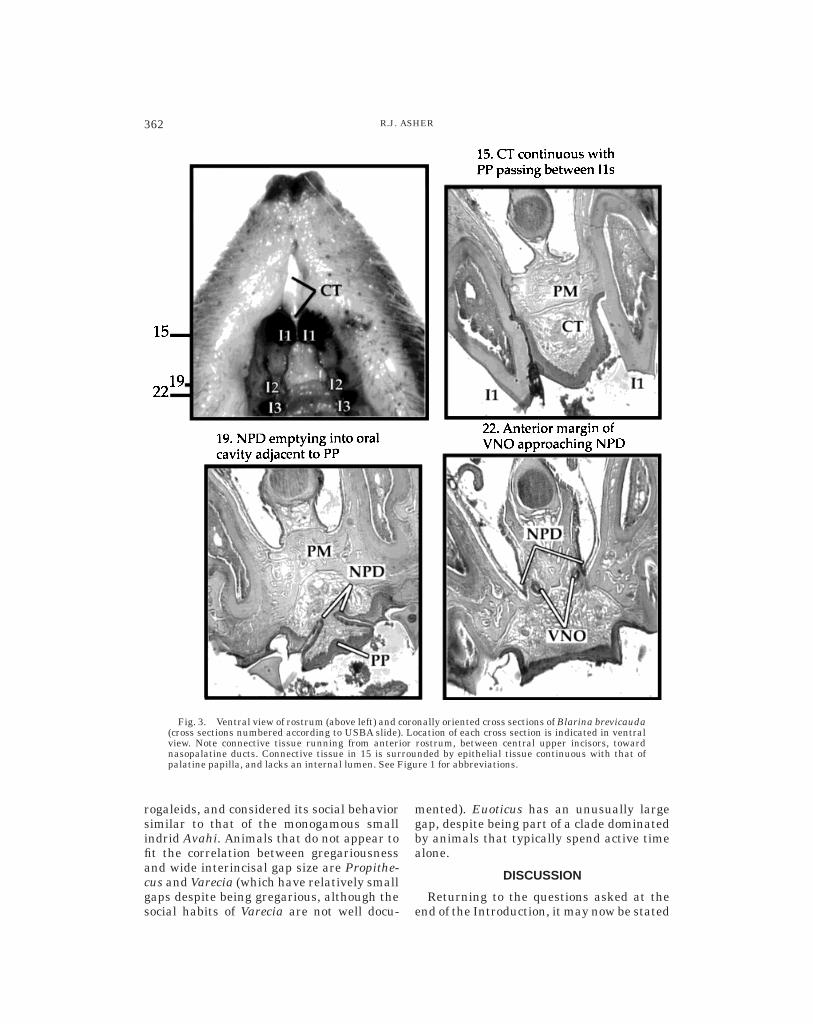
Blarina brevicauda has an elongate fold ofconnective tissue that runs between thecentral upper incisors, connecting the ante-rior rostrum and nasopalatine ducts (Fig. 3).Epithelial tissue continuous with that of thepalatine papilla surrounds this fold; no lu-men is evident within it. Figure 3 showsserial cross sections of the rostrum in Bla-rina, and demonstrates the connections be-tween the paired vomeronasal organ, thenasopalatine ducts, the palatine papilla, andthe fold of tissue between the central upperincisors. Certain tenrecine lipotyphlans ex-amined by Hofer (1982) lack this elongatefold of tissue, but are apomorphic in otherregards (e.g., in having an unpaired naso-
360 R.J. ASHER
Article ID # 920@xyserv1/disk8/CLS_liss/GRP_phan/JOB_phan105-3/DIV_920z16 dawn
palatine duct and in lacking a palatinepapilla).
Strepsirrhine mammals surveyed here arequite diverse in their rostral morphology(see also Hofer, 1977, 1982). Taxa that aredistinct from the pattern seen in Tupaia andEulemur include Felis (which lacks an inter-incisal gap), Ovis (which possesses a non-dental occlusal pad in lieu of upper anteriorteeth), and Blarina (which possesses anelongate fold of tissue connecting the upperlip and palatine papilla).
Is the lemuriform and adapiformrostrum autapomorphic?
As documented by Cave (1973) and Hofer(1976, 1977), the most conspicuous rostralautapomorphies among primates are foundin anthropoids and tarsiers. With notableexceptions (e.g., Daubentonia and lepilemu-rids including the subfossil Megaladapis),significant autapomorphies among strepsir-rhine primates are not as well-documented.This is understandable; the rostrum of strep-sirrhine primates differs little from that ofTupaia, a taxon often used to define primi-tive states among primates.
However, considerable variability amongstrepsirrhines is present in the size of thegap between the central upper incisors. Fig-ure 4 shows that in the size of the interinci-sal gap, most nocturnal lemuriforms, adap-ids, Tupaia, and Setifer fall either near orbelow the mean scaled-gap (by either palatalwidth or molar area) of all examined taxa.On the other hand, Eulemur, Lemur, Hapale-mur, Indri, and Avahi—all of which havebeen characterized as habitually spendingactive time with one or more conspecifics—fall well above the group mean. Phaner alsohas a relatively wide gap; interestingly,Charles-Dominique (1978) characterizedPhaner as more gregarious than other chei-
Fig. 2. Ventral views of rostra in Eulemur fulvus (A),Tupaia minor (B), and Felis catus (C). Eulemur andTupaia depend on a gap between the central upperincisors to facilitate communication between the rhi-narium and nasopalatine ducts, whereas Felis lacks agap within its anterior toothrow (see text). The palatinepapilla is partially dissected in Eulemur (A); the loweranterior teeth are intact in Tupaia (B) and obscure itspalatine papilla; the upper lip in Felis (C) has beendisplaced postmortem. See Figure 1 for abbreviations.
361STREPSIRRHINISM AND TOOTHCOMB EVOLUTION
Article ID # 920@xyserv1/disk8/CLS_liss/GRP_phan/JOB_phan105-3/DIV_920z16 dawn
rogaleids, and considered its social behaviorsimilar to that of the monogamous smallindrid Avahi. Animals that do not appear tofit the correlation between gregariousnessand wide interincisal gap size are Propithe-cus and Varecia (which have relatively smallgaps despite being gregarious, although thesocial habits of Varecia are not well docu-
mented). Euoticus has an unusually largegap, despite being part of a clade dominatedby animals that typically spend active timealone.
DISCUSSION
Returning to the questions asked at theend of the Introduction, it may now be stated
Fig. 3. Ventral view of rostrum (above left) and coronally oriented cross sections of Blarina brevicauda(cross sections numbered according to USBA slide). Location of each cross section is indicated in ventralview. Note connective tissue running from anterior rostrum, between central upper incisors, towardnasopalatine ducts. Connective tissue in 15 is surrounded by epithelial tissue continuous with that ofpalatine papilla, and lacks an internal lumen. See Figure 1 for abbreviations.
362 R.J. ASHER
Article ID # 920@xyserv1/disk8/CLS_liss/GRP_phan/JOB_phan105-3/DIV_920z16 dawn
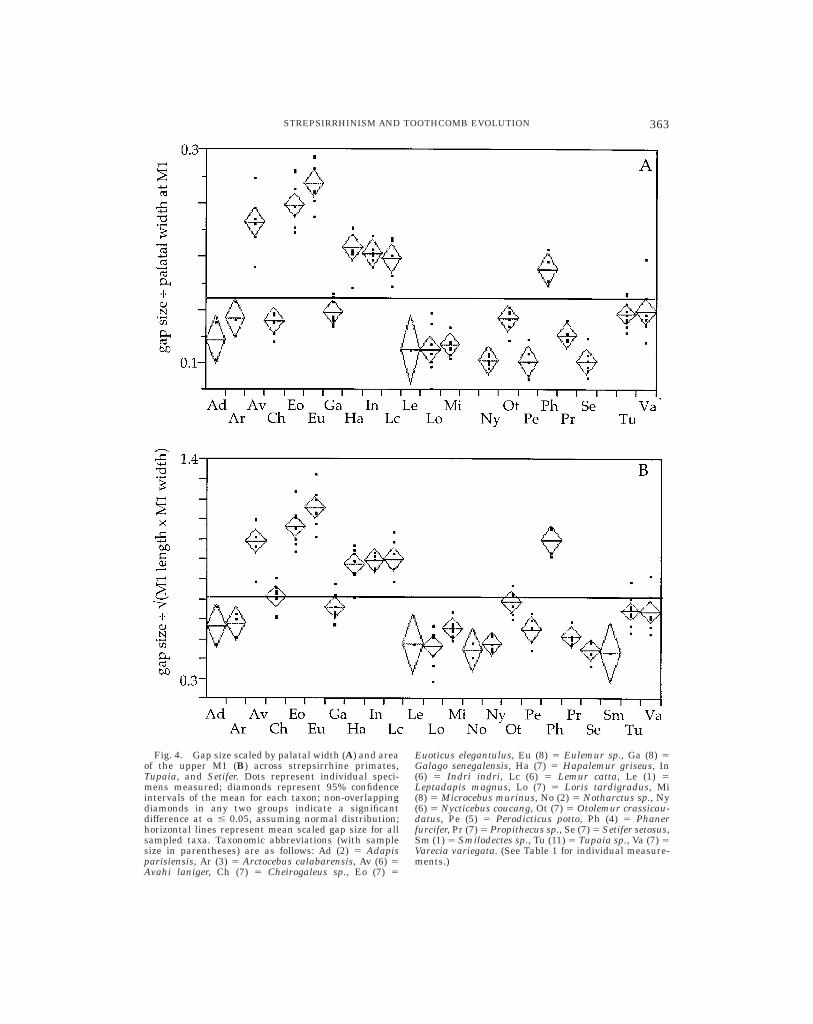
Fig. 4. Gap size scaled by palatal width (A) and areaof the upper M1 (B) across strepsirrhine primates,Tupaia, and Setifer. Dots represent individual speci-mens measured; diamonds represent 95% confidenceintervals of the mean for each taxon; non-overlappingdiamonds in any two groups indicate a significantdifference at a # 0.05, assuming normal distribution;horizontal lines represent mean scaled gap size for allsampled taxa. Taxonomic abbreviations (with samplesize in parentheses) are as follows: Ad (2) 5 Adapisparisiensis, Ar (3) 5 Arctocebus calabarensis, Av (6) 5Avahi laniger, Ch (7) 5 Cheirogaleus sp., Eo (7) 5
Euoticus elegantulus, Eu (8) 5 Eulemur sp., Ga (8) 5Galago senegalensis, Ha (7) 5 Hapalemur griseus, In(6) 5 Indri indri, Lc (6) 5 Lemur catta, Le (1) 5Leptadapis magnus, Lo (7) 5 Loris tardigradus, Mi(8) 5 Microcebus murinus, No (2) 5 Notharctus sp., Ny(6) 5 Nycticebus coucang, Ot (7) 5 Otolemur crassicau-datus, Pe (5) 5 Perodicticus potto, Ph (4) 5 Phanerfurcifer, Pr (7) 5 Propithecus sp., Se (7) 5 Setifer setosus,Sm (1) 5 Smilodectes sp., Tu (11) 5 Tupaia sp., Va (7) 5Varecia variegata. (See Table 1 for individual measure-ments.)
363STREPSIRRHINISM AND TOOTHCOMB EVOLUTION
Article ID # 920@xyserv1/disk8/CLS_liss/GRP_phan/JOB_phan105-3/DIV_920z16 dawn
that 1) no, anatomically strepsirrhine mam-mals do not display homogeneous rostralmorphologies, and 2) yes, data exist thatsupport Rosenberger and Strasser’s pro-posal that some lemuriforms (but not adapi-forms) have an autapomorphic rostrum. Theimplications of each of these answers for thepotential link between strepsirrhinism andtoothcomb evolution will be explored in turn.
Rostral diversity amongstrepsirrhine mammals
Outside of primates and tupaiids, theinterincisal gap is not a necessary correlateof anatomical strepsirrhinism. This findingis of particular concern to studies seeking toestablish the presence/absence of anatomi-cal strepsirrhinism in fossil taxa, which lackinformation on soft tissue structures. Fortu-nately, there are other osteological indica-tors: some lemurs (e.g., Varecia) possessbony prongs medial to the central upperincisors that may be related to the tetheredcondition of the upper lip.
A more consistent indicator may be re-lated to the morphology of the nasal fossa.With the possible exception of some aquaticmammals (e.g., Phoca), strepsirrhine mam-mals appear to be consistently macrosmatic.Across mammals as a whole, a relativelylarge and complex nasal fossa is bettercorrelated with strepsirrhinism than is theupper interincisal gap. All of the anatomi-cally strepsirrhine animals described here(i.e., Blarina, Eulemur, Felis, Ovis, andTupaia) possess a relatively large nasal fossadivided into respiratory and olfactory re-gions by the posterior transverse lamina(Cave, 1973); however, not all of these taxapossess a relatively large upper interincisalgap. Hence, studies on fossil taxa shouldincorporate internal nasal osteology in addi-tion to gaps in the anterior dentition to inferthe morphology of the soft tissues of therostrum (e.g., Moorman and Fleagle, 1980).
Gregarious lemuriforms arerostrally autapomorphic
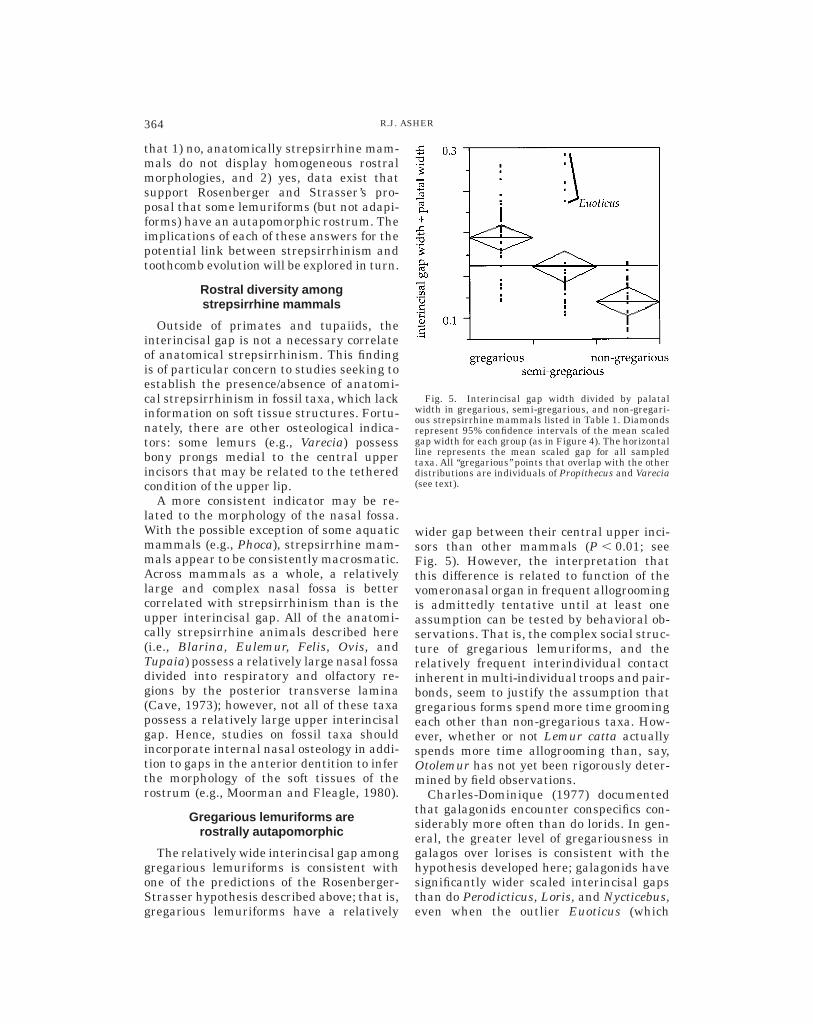
The relatively wide interincisal gap amonggregarious lemuriforms is consistent withone of the predictions of the Rosenberger-Strasser hypothesis described above; that is,gregarious lemuriforms have a relatively
wider gap between their central upper inci-sors than other mammals (P , 0.01; seeFig. 5). However, the interpretation thatthis difference is related to function of thevomeronasal organ in frequent allogroomingis admittedly tentative until at least oneassumption can be tested by behavioral ob-servations. That is, the complex social struc-ture of gregarious lemuriforms, and therelatively frequent interindividual contactinherent in multi-individual troops and pair-bonds, seem to justify the assumption thatgregarious forms spend more time groomingeach other than non-gregarious taxa. How-ever, whether or not Lemur catta actuallyspends more time allogrooming than, say,Otolemur has not yet been rigorously deter-mined by field observations.
Charles-Dominique (1977) documentedthat galagonids encounter conspecifics con-siderably more often than do lorids. In gen-eral, the greater level of gregariousness ingalagos over lorises is consistent with thehypothesis developed here; galagonids havesignificantly wider scaled interincisal gapsthan do Perodicticus, Loris, and Nycticebus,even when the outlier Euoticus (which
Fig. 5. Interincisal gap width divided by palatalwidth in gregarious, semi-gregarious, and non-gregari-ous strepsirrhine mammals listed in Table 1. Diamondsrepresent 95% confidence intervals of the mean scaledgap width for each group (as in Figure 4). The horizontalline represents the mean scaled gap for all sampledtaxa. All ‘‘gregarious’’ points that overlap with the otherdistributions are individuals of Propithecus and Varecia(see text).
364 R.J. ASHER
Article ID # 920@xyserv1/disk8/CLS_liss/GRP_phan/JOB_phan105-3/DIV_920z16 dawn

greatly increases mean galagonid gap size)is removed (P , 0.01; see Fig. 4). The loridArctocebus calabarensis provides a less-remarkable outlier; the small sample of thistaxon examined here does not have a signifi-cantly smaller interincisal gap than that ofGalago or Otolemur. There is also consider-able overlap between most galagos and treeshrews.
Dietary factors impinging on the morphol-ogy of the rostrum are clearly relevant tothis study, and may also explain some of theobserved variation. The fact that Propithe-cus has well-developed upper anterior inci-sors and uses them as foraging tools (Szalayand Seligsohn, 1977) is undoubtedly relatedto the morphology of its rostrum; the samemay be said of the anterior dentition inVarecia, and in the gumnivorous taxa Phanerand Euoticus. If diet alone is cited to explainthe gap size in these taxa, then it does nothave a consistent effect, as Propithecus andVarecia have unusually narrow gaps forindrids and lemurids, and Phaner and Euoti-cus have unusually large gaps for cheiroga-leids and galagonids (respectively). It islikely that both dietary and social factorshad some influence on the morphology of theanterior rostrum in all of these animals; butit is difficult to determine the relative impor-tance of these factors. In any event, moredata on the behavior of certain nocturnalprosimians (e.g., Euoticus and Phaner), andon the extent to which they can be classifiedas ‘‘gregarious,’’ are desirable to test whetherthe size of the interincisal gap in theseanimals could be related to frequent groom-ing.
Despite some exceptions, there remainsan intriguing correlation between the widthof the upper interincisal gap and socialpatterns among lemuriforms. Five of sevenlemurids and indrids that spend active timewith one or more adult conspecifics (Eule-mur, Hapalemur, Lemur, Avahi, and Indri),and one cheirogaleid (Phaner) classified as‘‘pre-gregarious’’ by Charles-Dominique(1978), have significantly wider scaled inter-incisal gaps than all lorids, cheirogaleids,non-primates, and all but one galagonid.Furthermore, galagonids have significantlywider interincisal gaps than most of theirless-gregarious lorid relatives.
If gregarious taxa actually allogroom morefrequently than others, then Rosenbergerand Strasser’s association of anatomicalstrepsirrhinism with the toothcomb offers atleast a partial explanation for their rela-tively large interincisal gap.
Evolution of the lemuriform toothcomb
This study supports Rosenberger andStrasser’s hypothesis that function of thetoothcomb is related to anatomical strepsir-rhinism. However, it does not support theircontention that adapiforms possessed anautapomorphic rostrum.
The shared possession in Tupaia, adapi-forms, and most non-gregarious lemuri-forms of a moderately sized gap between thecentral upper incisors, tethered upper lip,and palatine papilla (Figs. 3A, 3B) supportsthe interpretation that this condition amonglemuriforms is primitive. This is in accordwith Beard’s (1988) interpretation that thebase of the clade containing adapiforms andlemuriforms was not marked by a reconfigu-ration of oronasal morphology.
The inference that early strepsirrhine pri-mates had a plesiomorphic rostrum does notnecessarily invalidate Rosenberger andStrasser’s conclusion that the adapiformrostrum was ‘‘preadaptive’’ to the lemuri-form toothcomb. Primitive characters can,after all, have functional importance and atthe same time be integrated with a novelfunctional regime (e.g., phalangeal curva-ture in early hominids; see Susman et al.,1984). However, without some sort of deci-sive anatomical evidence, it is difficult totest when and how the adapiform rostrumbecame ‘‘preadaptive,’’ and decide whichsimilarly configured non-primate rostra arenot ‘‘preadaptive.’’
It is therefore unclear that allogroomingas a means of collecting pheromones wasthe phylogenetically primary function of thelemuriform toothcomb. Taxa that possessrelatively wide gaps—argued here to beassociated with grooming—appear to be well-nested within the Strepsirrhini (Fleagle,1988; Yoder, 1996); hence, I tentatively re-gard diet-based hypotheses of toothcomborigins (e.g., Martin, 1972; Rose et al., 1981)as plausible explanations.
365STREPSIRRHINISM AND TOOTHCOMB EVOLUTION
Article ID # 920@xyserv1/disk8/CLS_liss/GRP_phan/JOB_phan105-3/DIV_920z16 dawn

Finally, interesting—but anecdotal—infor-mation on dental combs in nonprimates isworth reviewing at this point. To my knowl-edge, the only other mammal with a well-developed dental comb known to be used forallogrooming is the dermopteran Cynocepha-lus (Aimi and Inagaki, 1988). This taxon isinteresting because it possesses a very widediastema in the upper anterior toothrow.The behavior of Cynocephalus in the wild ispoorly known (see Lekagul and McNeely,1977; Lim, 1967; Wharton, 1950). However,the evidence presented here can be used tomake a prediction: i.e., colugos are socialand, as in lemurids, allogrooming occurswith relative frequency and plays a sociallysignificant role. The wide interincisal gap incolugos results from the need for a wide pathfrom the anterior rostrum to the vomerona-sal organ in a taxon that allogrooms, collectsconspecific pheromones, and uses its vomero-nasal organ to process them. Hopefully, suf-ficient behavioral data on living dermopter-ans will someday be collected to test thishypothesis.
CONCLUSIONS
The gap between the central upper inci-sors characteristic of lemuriform primatesand Tupaia is one of several ways in whichcontact between the vomeronasal organ andrhinarium can be established. Other mam-mals have evolved specialized means involv-ing morphology (e.g., the elongate fold ofconnective tissue in some lipotyphlans; seeFig. 3) and behavior (e.g., flehmen in bovids)for this purpose.
Among strepsirrhine primates, gregari-ous lemuriforms have apomorphically largegaps between their central upper incisors(Fig. 5). This observation is consistent withthe functional link between anatomicalstrepsirrhinism and dental grooming pro-posed by Rosenberger and Strasser (1985).However, adapiforms and non-gregarious le-muriforms do not appear to share significantrostral apomorphies. It is therefore difficultto infer the extent to which social factorsinfluenced the origin of the toothcomb inearly toothcomb prosimians.
ACKNOWLEDGMENTS
The following individuals deserve thanksfor discussion, access to equipment, and/orhelp in accessing literature: Chris Beard,Brigitte Demes, John Fleagle, Bill Jungers,David Krause, Charles Lockwood, OsbjornPearson, Beth Peterson, Tab Rasmussen,David Reim, Brian Richmond, Alfie Rosen-berger, Callum Ross, Randy Susman, ChiaTan, Ben Walcott, and Roshna Wunderlich.Two anonymous reviewers helped to im-prove this manuscript considerably. I thankClare Flemming, Wolfgang Fuchs, and RossMacPhee for access to specimens at theAMNH, Maria Rutzmoser for access to collec-tions at the MCZ, and SUNY Stony Brookand Sigma Xi for financial support.
LITERATURE CITED
Aimi M and Inagaki H (1988) Grooved lower incisors inflying lemurs. J Mammal 69:138–140.
Bailey K (1978) Flehman in the ring tailed lemur(Lemur catta). Behaviour 65:309–319.
Beard KC (1988) The phylogenetic significance of strep-sirhinism in Paleogene primates. Int. J. Primatol.9:83–96.
Bearder SK (1987) Lorises, bushbabies, and tarsiers:diverse societies in solitary foragers. In BB Smuts, DLCheyney, RM Seyfarth, RW Wrangham, and TTStrushaker (eds.): Primate Societies. Chicago: Univer-sity of Chicago Press, pp. 9–24.
Buettner-Janusch A and Andrew RM (1962) Use of theincisors by primates in grooming. Am. J. Phys. Anthro-pol. 20:127–129.
Cave AJE (1973) The primate nasal fossa. Biol. J.Linnean Soc. 5:377–387.
Charles-Dominique P (1977) Ecology and Behavior ofNocturnal Primates: Prosimians of Equatorial WestAfrica. Translated by RD Martin. New York: ColumbiaUniversity Press, 277 pp.
Charles-Dominique P (1978) Solitary and gregariousprosimians: evolution of social structures in primates.In DJ Chivers and KA Joysey (eds.): Recent Advancesin Primatology. London: Academic Press, pp. 139–149.
Eisenberg JF and Gould E (1970) The tenrecs: a study inmammalian behavior and evolution. Smithsonian Con-trib. Zool. 27:1–137.
Estes R (1972) The role of the vomeronasal organ inmammalian reproduction. Mammalia 36:315–341.
Fleagle JG (1988) Primate Evolution and Adaptation.San Diego: Academic Press, 486 pp.
Hofer HO (1976) Preliminary study of the comparativeanatomy of the external nose of South Americanmonkeys. Folia Primatol. 25:193–214.
Hofer HO (1977) The anatomical relations of the ductusvomeronasalis and the occurrence of taste buds in thepapilla palatina of Nycticebus coucang (Primates,Prosimiae) with remarks on strepsirrhinism. Gegen-baurs Morphol. Jahrb. 123:836–856.
Hofer HO (1982) Anatomy of the oro-nasal region ofsome species of Tenrecidae and considerations oftupaiids and lemurids. Gegenbaurs Morphol. Jahrb.4:588–613.
Jenkins PD (1987) Catalogue of Primates in the BritishMuseum (Natural History) and Elsewhere in the
366 R.J. ASHER
Article ID # 920@xyserv1/disk8/CLS_liss/GRP_phan/JOB_phan105-3/DIV_920z16 dawn

British Isles. Part IV. Suborder Strepsirrhini, includ-ing the Subfossil Madagascan lemurs and the FamilyTarsiidae. London: British Museum (Natural His-tory).
Krishtalka L, Stucky RK, and Beard KC (1990) Theearliest fossil evidence for sexual dimorphism in pri-mates. Proc. Natl. Acad. Sci. U.S.A. 87:5223–5226.
Lekagul B and McNeely J (1977) Mammals of Thailand.Bangkok: Sahakarnbhat, 758 pp.
Lim BL (1967) Observations on the food habits andecological habitat of the Malayan flying lemur. Int.Zoo Yrbk. 7:196–197.
Maier W (1991) Aspects of ontogenetic development ofnasal and facial skeletons in primates. In G Pfeifer(ed.): Craniofacial abnormalities and clefts of the lip,alveolus, and palate. Stuttgart: Thieme Verlag, pp.115–124.
Martin RD (1972) Adaptive variation of behavior ofMalagasy lemurs. Philos. Trans. R. Soc. Lond. [Biol.]26:295–352.
Martin RD (1990) Primate Origins and Evolution: APhylogenetic Reconstruction. Princeton: PrincetonUniversity Press, 804 pp.
Moorman SJ and Fleagle JG (1980) The nasal fossa inextinct strepsirhines. Am. J. Phys. Anthropol. 52:260A.
Nowak RM (1991) Walker’s Mammals of the World, 5th ed.Baltimore: Johns Hopkins University Press, 1629 pp.
Pocock RI (1918) On the external characters of the lemursand of Tarsius. Proc. Zool. Soc. Lond. 1918:19–53.
Rasmussen DT (1986) Anthropoid origins: A possiblesolution to the Adapidae—Omomyidae paradox. J.Hum. Evol. 15:1–12.
Richard AF (1987) Malagasy Prosimians: Female Domi-nance. In BB Smuts, DL Cheyney, RM Seyfarth, RWWrangham, and TT Strushaker (eds.): Primate Societ-ies. Chicago: University of Chicago Press, pp. 25–33.
Rose KD, Walker A, and Jacobs LL (1981) Function ofthe mandibular toothcomb in living and extinct mam-mals. Nature 289:583–585.
Rosenberger AL and Strasser E (1985) Toothcomb ori-gins: Support for the grooming hypothesis. Primates26:73–84.
Rosenberger AL and Szalay FS (1980) On the tarsiiformorigins of Anthropoidea. In RL Ciochon and AB Chi-arelli (eds.): Evolutionary Biology of the New WorldMonkeys and Continental Drift. New York: Plenum,pp. 139–157.
Rosenberger AL, Strasser E, and Delson E (1985) Ante-rior dentition of Notharctus and the adapid-anthro-poid hypothesis. Folia Primatol. 44:15–39.
Sall J, Ng K, Hecht M, Tilley D, and Potter R (1994)JMP: Statistics Made Visual (computer program).Cary, NC: SAS Institute.
Schilling A (1970) L’organe de Jacobson de lemurienMalgache Microcebus murinus (Miller 1777). Memoi-res de Museum Nationale d’Histoire Naturelle (serieA) 61:203–280.
Sokal RR and Rohlf FJ (1995) Biometry, 3rd ed. NewYork: W.H. Freeman, 887 pp.
Susman RL, Stern JT, and Jungers WL (1984) Arboreal-ity and bipedality in the Hadar hominids. Folia Prima-tol. 43:113–156.
Szalay FS and Seligsohn D (1977) Why did the strepsi-rhine toothcomb evolve? Folia Primatol. 27:75–82.
Verberne G (1976) Chemocommunication among domes-tic cats, mediated by the olfactory and vomeronasalsenses. Z. Tierpsychol. 42:113–128.
Wharton CH (1950) Notes on the life history of the flyinglemur. J. Mammal. 31:269–273.
Yoder AD, Cartmill M, Ruvolo M, Smith K, and VilgalysR (1996) Ancient single origin for Malagasy primates.Proc. Natl. Acad. Sci. U.S.A. 93:5122–5126.
367STREPSIRRHINISM AND TOOTHCOMB EVOLUTION
Article ID # 920@xyserv1/disk8/CLS_liss/GRP_phan/JOB_phan105-3/DIV_920z16 dawn





























