Embed Size (px)
Citation preview

Vegetafio 123: 183-192, 1996. 183 @ 1996 Kluwer Academic Publishers. Printed in Belgium.
Morphological responses to nutrient availability in four clonal herbs
Ming Dong l, Heinjo J. During & Marinus J. A. Werger* Department of Plant Ecology and Evolutionary Biology, Utrecht University, Postbus 800.84, 3508 TB Utrecht, the Netherlands; 1Present address: Institute of Botany, Chinese Academy of Sciences, 141 Xizhimenwai Ave, Beijing 100044, China (* author for correspondence)
Received 1 March 1995; accepted in revised form 17 October 1995
Key words: Carex, Clonal plant, Constraint, Habitat adaptation, Morphological plasticity, Trifolium
Abstract
This paper examines morphological plasticity of clonal plants of contrasting habitats and of contrasting architectures in response to nutrient supply. The hypotheses were tested that plants from rich habitats possess greater plasticity in response to variation in resource supply than species from poor habitats, and that rhizomatous species are less plastic in their response than stoloniferous species. Two sympodial rhizomatous herbs (Carex flacca, C. hirta) and two monopodial stoloniferous herbs (Trifoliumfragiferum, T. repens) were subjected to four levels of nutrient supply in a garden experiment. One of the two species of each genus (C. hirta, T. repens) is from fertile and the other from infertile habitats. We measured 1) whole plant characters: total plant dry weight, number of modules (product of a single apical meristem) and number of ramets; 2) ramet characters: ramet leaf area and ramet height; and 3) spacer characters: branches per module, length per module and length per module internode.
All measured characters in the Trifolium species significantly responded to treatment: the values for all measured characters increased with higher levels of fertilization. The differences in plant characters between fertilization levels were larger in Trifolium repens than in T. fragiferum in terms of whole plant characters, ramet characters and stolon internode length. The two Carex species did not differ in their responses to treatment in terms of most characters measured. In ramet characters and in some whole plant characters the species from fertile habitats were more plastic than those from infertile habitats. In spacer characters this pattern was not found. Foraging could not be demonstrated unequivocally.
Morphological plasticity in the stoloniferous (Trifolium) species was much larger than in the rhizomatous (Carex) species. This seems in accordance with a foremost storage function of rhizomes, as against a foremost explorative function of stolons.
Introduction
Clonal plants may respond to variation in environmen- tal factors by plastic adjustment of their morpholo- gy (Slade & Hutchings 1987a, b; de Kroon & Knops 1990; Evans 1992; Caradus et al. 1993; Dong 1993; Thompson 1993; Dong & de Kroon 1994; Hutchings & de Kroon 1994). Morphological plasticity in spac- er (rhizome, stolon internode, Bell 1984) characters such as branching probability and length as well as in ramet characters such as leaf area and ramet height may be of ecological significance. Plasticity influences
the placement of resource-acquiring structures such as leaves and roots in a heterogeneous environment (Grime et al. 1986; Hutchings & de Kroon 1994), and may enhance the exploitation of patchily distribut- ed essential resources (foraging, Grime et al. 1986; Hutchings & de Kroon 1994). Several authors have recently shown that some clonal species may differ in responses of branching intensity and/or spacer length and that other species are non-plastic (Sutherland & Stillman 1988; Hutchings & de Kroon 1994; de Kroon & Hutchings 1995). We hypothesized that such differ- ences in plasticity may be related to the characteristics

184
of the habitats in which the species evolved. We test aspects of this hypothesis in this paper.
A similar morphological plasticity may not be equally important in all habitats (Grime et al. 1986; Sutherland & Stillman 1988; de Kroon & Schiev- ing 1990; Dong 1995). Nutrient-rich, mesic, well- illuminated habitats can support plants with relatively high growth rates. As leaves and roots of those plants grow out, depletion zones are constantly being created in a rapidly shifting pattern of microsites in the three dimensional space of such habitats, creating a spatially and temporally highly dynamic environment (Grime et al. 1986). In such habitats a high degree of mor- phological plasticity is required to sustain high rates of resource capture by constantly locating new leaves and roots in unoccupied resource-rich microsites.
In nutrient-poor, xeric habitats, however, spa- tial and temporal variation in microsites of different resource availability is expected to be less pronounced and resource capture opportunities for various plant parts seem less variable (cf. Grime et al. 1986). There plants (and plant parts) have to cope with persistent low availability of soil-based resources, but also have relatively little chance of being rapidly outshaded. In such habitats a high degree of morphological plasticity does not seem an asset.
Plasticity of a character can be influenced by nat- ural selection as it apparently has its own genetic basis (Bradshaw 1965; Schlichting 1986; Scheiner 1993). Consequently, morphological plasticity may differ between species that have evolved in different habitats (Grime et al. 1986; Hutchings & de Kroon 1994). We thus expect that clonal plant species of fer- tile habitats have developed a higher degree of morpho- logical plasticity than species of infertile habitats.
Clonal plants may differ in architectural model (Lovell & Lovell 1985; Bell 1991) specifying 'growth rules' such as the timing and development of modular branches and the length of spacing organs (Bell & Tom- linson 1980; Waller & Steingraeber 1985). Such dif- ferences may constrain morphological plasticity. For example, Prunella vulgaris with longer spacers was morphologically more responsive to the presence of other grasses than Bellis perennis with shorter spacers (Schmid 1986). In Lycopodium annotinum, branching is restricted to the youngest parts of the plants while Carex bigelowii is able to produce different types of axillary tillers from dormant buds. Thus, Carex is more responsive to variation in resource availability than Lycopodium (Carlsson et al. 1990).
Different clonal plant species may possess differ- ent types of spacers such as sympodial rhizomes or monopodial stolons (Leakey 1981). Stolons and rhi- zomes are homologous organs; stolons grow above- ground while rhizomes are subterranean. Stolons lack the considerable storage and regenerative capacities of rhizomes (Grime 1979; Leakey 1981; Hutchings & Mogie 1990). In stolons, generally all or most buds easily grow new ramets while rhizomes contain a great number of dormant buds (de Kroon & Knops 1990). Such differences constrain responses of species to environmental change (de Kroon & van Groenen- dael 1990). It has recently been shown that in grass- es possessing stolons and/or rhizomes, stolons were morphologically more responsive to differential light supply than rhizomes (Dong & de Kroon 1994; Dong & Pierdominici 1994). De Kroon & Hutchings (1995) did not find conclusive evidence for such a consis- tent difference between rhizomatous and stoloniferous species in response to nutrients. However, they did not consider quantified aspects of the responses and inven- toried only characters that are of direct relevance for foraging. Considering the pronounced storage func- tion of rhizomes (for carbohydrates and dormant buds) and the explorative, possibly even forage, function of stolons, we expect that stoloniferous and rhizomatous species differ in plasticity in a number of characters in response to variation in nutrient availability. Specifi- cally, we expect rhizomatous species to be less respon- sive than stoloniferous species in terms of plant size (plant mass and number of ramets), internode length and branching frequency.
These expectations were tested in a garden exper- iment in which four clonal herbaceous species, Carex flacca, C. hirta, Trifolium fragiferum and T. repens, were subjected to four levels of nutrient supply. The two species within each genus have the same branching pattern and spacer type, while they characteristically occur in nutrient-poor and -rich habitats, respectively. These genera differ in branching pattern (monopodial versus sympodial) and in spacer type (stolon versus rhizome).
Material and methods
The species
Trifolium fragiferum L. and Trifolium repens L. are perennial leguminous herbs commonly found in Europe (Burdon 1983; Grime et al. 1988). Trifoli-

um fragiferum grows in grassy and muddy, frequent- ly heavily trampled places, usually on saline ground (Zohary & Heller 1984), and is regarded as an indica- tion for nutrient-poor habitats (Landolt 1977). Trifoli- um repens is most common in meadows, pastures, road verges (Burdon 1983; Grime etal. 1988) and is consid- ered as a good indicator for nutrient-rich sites (Ober- dorfer 1970; Landolt 1977). The two species spread plagiotropically by means of monopodial vegetative stolons, i.e., monopodial modules (product of a single apical meristem, White 1979; Cook 1985) consisting of a number of leaf-ramets (Mogie & Hutchings 1990) interconnected by stolon internodes (Fig. 1A). Each leaf-ramet consists of a leafy stolon node usually with attached adventitious roots. At each stolon node one palmately trifoliate leaf is produced subtending an axil- lary bud which may give rise to either a lateral stolon or a flower bud (Burdon 1983).
Carex flacca (L.) Schreber and Carex hirta L. are perennial rhizomatous sedges with a wide distribution. Carexflacca usually is a calcicole species ( Grime et al. 1988) of chalk grasslands, dune valleys and open forests, always on fairly nutrient-poor sites in Western Europe (Taylor 1956; Oberdorfer 1970). Carex hir- ta usually inhabits damp nutrient-rich sands, which are frequently a little disturbed (Oberdorfer 1970). Both species form sympodial rhizomes, i.e., sympodi- al modules (Fig. 1B) which widely vary in length (de Kroon & Knops 1990). The rhizomes grow plagiotrop- ically before the apices develop into orthotropic shoots (shoot-ramet, Mogie & Hutchings 1990). The base of a shoot consists of a number of compressed internodes from which adventitious roots and new rhizomes may grow out.
Experimental design
Plant material was collected in the Netherlands, in relatively nutrient-rich habitats for Carex hirta (bank of the Amsterdam-Rijn canal near Utrecht) and Trifolium repens (a meadow at the Uithof near Utrecht), and in relatively nutrient-poor habitats for Carexflacca (chalk grassland Vrakelberg in the province of Limburg) and Trifoliumfragiferum (coastal dunes in the province of Zeeland).
For each of the species, experimental plants were propagated from four or five original plants, collected in different locations within each habitat that were at least several meters apart. They may or may not differ in genotype. The material was propagated outdoors for six months before the experiment started.
185
The experiment was conducted at the Uithof Exper- imental Garden, Utrecht, the Netherlands, from April 1 to August 31,1991. Initial experimental plants were single ramets each consisting of one leafy stolon node and attached adventitious roots in the Trifolium species, or one orthotropic leafy shoot and attached adventi- tious roots in the Carex species.
Treatments were four levels of nutrient supply. The high nutrient level (H) was equivalent to 200 kg N as NHaNO3, 60 kg P as NaH2POa.2H20 and 200 kg K as KCI per hectare per year. The medium (M), low (L) and extra low (EL) were 50%, 25% and 10% of the highest level, respectively. Nutrient solution was sup- plied twice a month. The nutrient solutions used were similar in volume and only differed in the concentra- tion of nutrients. During the experiment the plants were watered as needed.
Plants were grown in plastic pots (44 cm surface diameter and 50 cm deep) filled with river sand. Each pot contained two replicate plants which were propa- gated from different original plants of the same species. The pots were placed outside in a random arrange- ment. There were eight replicate plants per treatment and per species. To make sure that differences found in response to treatments were not due to genotypic effects, the same number of replicate plants that were propagated from a given original plant were used in all four treatments.
Harvesting and analyses
The plants were harvested after five months of growth. Washed plants were stored in a freezer ( - 2 1 °C) until analysis. For each of the plants, constituent modules were identified and labelled according to branching orders. The number of modules and ramets per plant as well as the number of lateral modules (branches) per module were counted. Two modules (shoot-ramets with rhizomes >5 mm, in Carex) per branching order and per replicate plant were randomly selected to mea- sure detailed morphologicalcharacters. For both Carex and Trifolium, module length and number of nodes were determined. Length of longest leaf and ramet leaf area were measured for Carex, and petiole length and lamina area were measured for all ramets along the Trifolium stolons. Leaf (lamina) area was determined with a Delta-T area meter.
The plants were separated into spacers (stolons or rhizomes) and ramets. The ramets were subdivided into roots, leaves, petioles and inflorescences in Trifolium, and into leaves, "culms" (thickened underground shoot

186
base, cf. de Kroon & Knops 1990), roots and infiores- cences in Carex. After drying at 75 °C for at least 48 h, dry weights of the different components were measured.
The following three groups of characters were derived for comparisons between species: (1) whole plant characters such as whole plant dry weight, num- ber of modules and number of ramets, (2) spacer char- acters including branches per module, length per mod- ule (stolons, fully grown rhizomes >5 mm) and length per module internode, and (3) ramet characters in terms of leaf area per ramet and height per ramet (petiole length in Trifolium and length of the longest leaf in Carex).
All statistical comparisons for spacer and ramet characters were based on the mean values for each of the replicate plants. Effects of treatments and dif- ferences between species were analysed by ANOVA (SAS 1985). Following Schlichting & Levin (1984) and Via (1987) degrees of plasticity were measured as coefficients of variation of the means per treatment (CV). We prefer this measure because it translates as the species' response proportional to the mean, rather than the absolute differences in response as measured by the interaction term in ANOVA.
A Monopodi l l l A. Monopodial architecture module :
(Trifoliurn) A leaf-
rarnet :
_ P
% 'o 5~o- - -~-- >
p
,o
B. Sympodial architecture (Carex)
A sympodisl module :
A shoot- tamer :
Results
Responses of whole plant characters
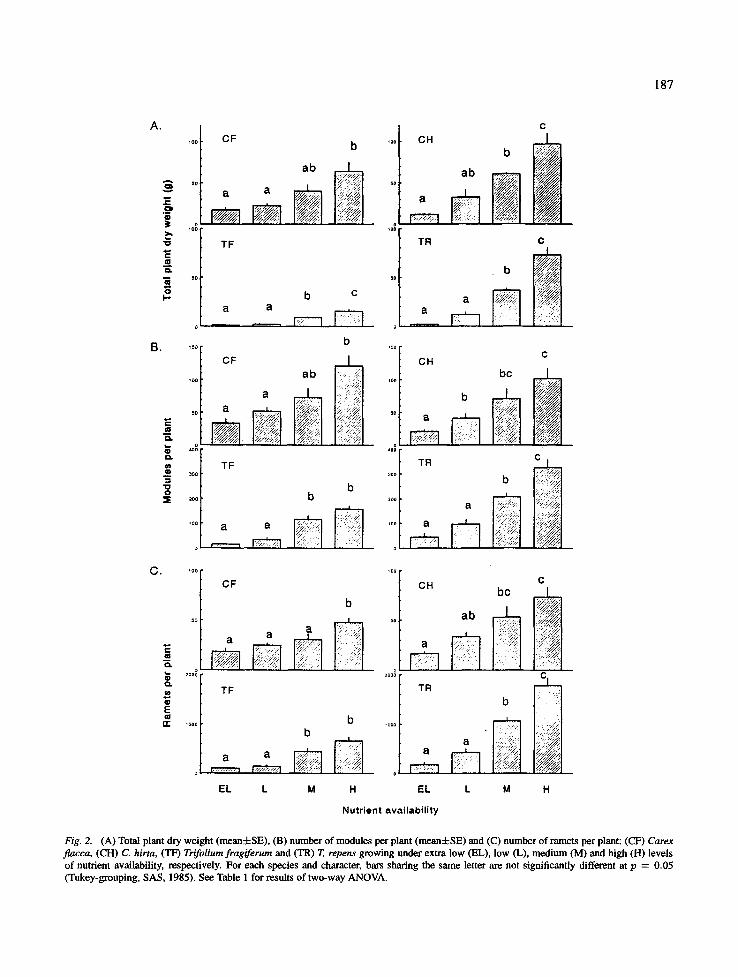
Each of the four species accumulated more biomass (Fig. 2A) and produced more modules (Fig. 2B) and ramets (Fig. 2C) under higher levels of nutrient sup- ply (Table 1). Plants growing under the highest lev- el showed a 3- to 25-fold, 4- to 6-fold and 2- to 9- fold larger total plant dry weight, number of modules and number of ramets, respectively, as compared with plants growing under the extra low level of nutrient supply. Carex hirta tended to respond more strongly than C. flacca (Fig. 2; Table 2), but the species by treatment interaction effects (see Table 1), which are an indication whether the species responded differently to treatment, were not significant at p = 0.05. Trifoli- um repens also responded more strongly to fertilization than T. fragiferum (Table 1) and the species by treat- ment interactions were significant. Except for the num- ber of modules per plant, the CV values for T. repens were also higher than for T. fragiferum (Table 2).
Fig. 1. Diagrammatic representation of the vegetative morphology of (A) monopodial stoloniferous herbs (Tr~folium) and (B) sympodial rhizomatous herbs (Carex) used in the present experiment. A Tri- folium plant consists of a number of monopodial modules (stolons). A monopodial module is composed of a number of leaf-ramets each consisting of a node (solid circle) and a leaf (open circle) and spaced by single internodes (line interconnecting two nodes). A Carex plant consists of a number of sympodial modules (rhizomes). A sympodial module consists of a developing rhizome or of a developed rhizome with a shoot ramet (open rectangle). Arrows depict the apices of modules.
Responses of spacer characters
Carex flacca branched more intensively in terms of branches per module and produced shorter rhizomes with shorter internodes than C. hirta. However, the Carex species did not significantly respond to treat- ment in each of these spacer characters (Fig. 3A-C, Table 1). C. hirta had higher CV values. In contrast, both Trifolium species branched more intensively and produced longer stolons with longer internodes under higher levels of nutrient supply (Fig. 3A-C, Table 1). The response in stolon internode length was signifi- cantly stronger in Trifolium repens than in T. fragifer-
187
A. ~00
Q
lOO
,13
• ~ ~o
I -
B . ,~o
Q .
300
0
CF
a a ~
TF
b c a
cF
ab i ~: ;~
a I ~,: ~
TF
b b
a
I , , ' I
~50
c
b
a ~ TR C
c CH
aoo b
~°° a
( ~ . too
e=
~ooo
o E
CF
b
a
TF
b b
a a ~ N l" ( , , . l , . / . ] I,/.',..J~//~|
EL L M H
C CH
~ ab ~
TR
b
EL L M H
Nutrient availabil ity
Fig. 2. (A) Total plant dry weight (mean:ESE), (B) number of modules per plant (mean+SE) and (C) number of ramets per plant: (CF) Carex flacca, (CH) C. hirta, (TF) Trifoliumfragiferum and (TR) T. repens growing under extra low (EL), low (L), medium (M) and high (H) levels of nutrient availability, respectively. For each species and character, bars sharing the same letter are not significantly different at p = 0.05 (Tukey-grouping, SAS, 1985). See Table 1 for results of two-way ANOVA.

188
A.
B.
C.
0 E
(D 20 e" O
400
E E 2oo
loo
1o o o E =
. c
c
,..1 ~oo
A ~5 E
0 a D 0
_¢ 0 _o
0 E
t -
- J
CF
a a a a
CH
a a a a
TF b
b
CF 500
400
3~C'
200 a a a a
T F ~o
b b ,oo
TR
b b
a
CH
a a a a
TR c C
CF
a a a a
CH a a a
TF
b b
a
EL L M H
• T R
c C
EL L M H Nutr ient avai labi l i ty
Fig. 3. (A) Branches per module (mean-l-SE), (B) length per module (mean=t=SE) and (C) length per module intemode (mean±SE). In Carex, lengths were based on full-grown rhizomes (i.e., rhizomes with a shoot ramet), longer than 5 mm. Species and treatment code as in Figure 2. For each species and character, bars sharing the same letter are not significantly different at p = 0.05 (Tukey-grouping). See Table 1 for results of two-way ANOVA.
189
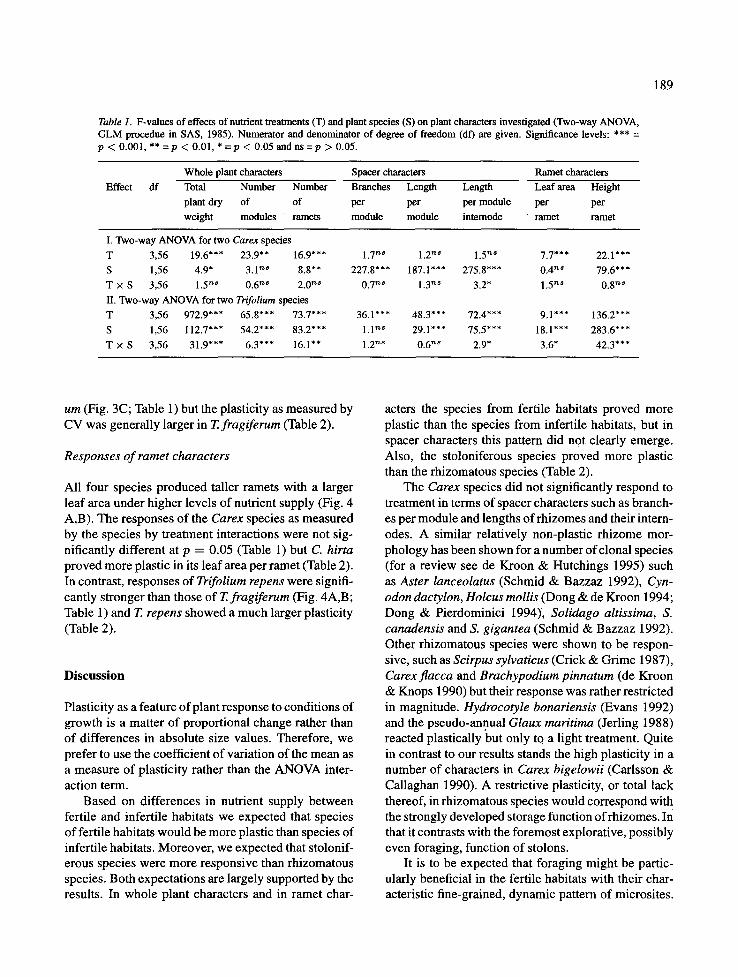
Table 1. F-values of effects of nutrient treatments (T) and plant species (S) on plant characters investigated (Two-way ANOVA, GLM procedue in SAS, 1985). Numerator and denominator of degree of freedom (df) are given. Significance levels: *** = p < 0.001, ** =p < 0.01, * =p < 0.05 and ns =p > 0.05.
Effect df Whole plant characters Spacer characters Ramet characters Total Number Number Branches Leng th Length Leaf area Height plant dry of of per per per module per per weight modules ramets module m o d u l e internode ramet ramet
I. Two-way ANOVA for two Carex species T 3,56 19.6"** 23.9** 16.9"** 1.7 '~s 1.2 ns 1.5 '~8 7.7*** 22.1"** S 1,56 4.9* 3.1 '~s 8.8** 227.8*** 187.1"** 275.8*** 0.4 '~s 79.6*** T x S 3,56 1.5 T M 0.6 '~s 2.0 '~8 0.7 ~ 1.3 '~ 3.2* 1,5 ~s 0.8 '~s II. Two-way ANOVA for two Tr~folium species T 3,56 972.9*** 65.8*** 73.7*** 36.1"** 48.3*** 72.4*** 9.1"** 136.2"** S 1,56 112.7"** 54.2*** 83.2*** 1.1 '~s 29.1"** 75.5*** 18.1"** 283.6*** T × S 3 , 5 6 31.9"** 6.3*** 16.1"* 1.2 ~s 0.6 ns 2.9* 3.6* 42.3***
um (Fig. 3C; Table 1) but the plasticity as measured by CV was general ly larger in T. fragiferum (Table 2).
Responses of ramet characters
All four species produced miler ramets with a larger leaf area under higher levels of nutrient supply (Fig. 4 A,B). The responses of the Carex species as measured by the species by treatment interactions were not sig- nificantly different at p = 0.05 (Table 1) but C. hirta proved more plastic in its leaf area per ramet (Table 2). In contrast, responses of Trifolium repens were signifi- cantly stronger than those of T. fragiferum (Fig. 4A,B; Table 1) and T. repens showed a much larger plasticity (Table 2).
Discussion
Plasticity as a feature of plant response to conditions of growth is a matter of proport ional change rather than of differences in absolute size values. Therefore, we prefer to use the coefficient of variation of the mean as a measure of plasticity rather than the ANOVA inter- action term.
Based on differences in nutrient supply between fertile and infertile habitats we expected that species of fertile habitats would be more plastic than species of infertile habitats. Moreover, we expected that stolonif- erous species were more responsive than rhizomatous species. Both expectat ions are largely supported by the results. In whole plant characters and in ramet char-
acters the species from fertile habitats proved more plastic than the species from infertile habitats, but in spacer characters this pattern did not clearly emerge. Also, the stoloniferous species proved more plastic than the rhizomatous species (Table 2).
The Carex species did not significantly respond to treatment in terms of spacer characters such as branch- es per module and lengths of rhizomes and their intern- odes. A similar relatively non-plastic rhizome mor- phology has been shown for a number of clonal species (for a review see de Kroon & Hutchings 1995) such as Aster lanceolatus (Schmid & Bazzaz 1992), Cyn- odon dactylon, Holcus moUis (Dong & de Kroon 1994; Dong & Pierdominici 1994), Solidago altissima, S. canadensis and S. gigantea (Schmid & Bazzaz 1992). Other rhizomatous species were shown to be respon- sive, such as Scirpus sylvaticus (Crick & Grime 1987), Carex flacca and Brachypodium pinnatum (de Kroon & Knops 1990) but their response was rather restricted in magnitude. Hydrocotyle bonariensis (Evans 1992) and the pseudo-annual Glaux maritima (Jerling 1988) reacted plastically but only to a light treatment. Quite in contrast to our results stands the high plasticity in a number of characters in Carex bigelowii (Carlsson & Callaghan 1990). A restrictive plasticity, or total lack thereof, in rhizomatous species would correspond with the strongly developed storage function ofrhizomes. In that it contrasts with the foremost explorative, possibly even foraging, function of stolons.
It is to be expected that foraging might be partic- ularly beneficial in the fertile habitats with their char- acteristic fine-grained, dynamic pattern of microsites.

190
A .
B .
CF
a
E ~* ~ E
o
~t~ 4o0
TF 350
:~oo
--I ~ 5 0
~ 0 ~
1 5 0
~o [ - . , - ]
~° OF
a
°
a b b a
b c a
b a ab
~_. ro ~ TF ° ~ 60
x-- 5O
~: '° C E ,o b
a a ~ ' 1 o
EL L M H
1 oo OH
a
"°° TR ~ o
~ o o
1 5 o a
i o
OH
*o lel
1o
,o TR
~o
4o
~o
EL
a b a b
b
I ~k%~ ~;,g?7~;.;," :~ .... , / . , ,~:;<
b
C
b
; ' • , ff;; '72, :
d e
; ::::~ ,:7(:7"1 b + , k , i
":;',: :2:26";71
L M H
Nutrient ava i l ab i l i ty
Fig. 4. (A) Leaf area per ramet (mean-I-SE) and (13) height per ramet (mean-t-SE). Species and treatment code as in Figure 2. For each species and character, bars sharing the same letter are not significantly different at p = 0.05 (Tukey-grouping). See Table 1 for results of two-way ANOVA.
But al though we found higher plasticity in whole plant and ramet characters in plants from fertile habitats as compared to those from infertile habitats, the results for the spacer characters, which are among the most impor- tant attributes of foraging plants, are equivocal.
Spacers al low clonal plants to reposit ion them- selves within the habitat. It has been suggested that more intensive branching and the production of short- er spacers under higher levels of resource supply will enable clonal plants to place ramets selectively into the more favourable patches (Hutchings & Slade 1988;

Table 2. Coefficients of variation of means per treatment and species.
191
Whole plant characters Spacer characters T o t a l Number Number Branches plant dry of of per
weight modules ramets module
Ramet characters Length Length per Leaf area Height
per module per per module internode ramet rarnet
C. flacca 0.59 0.54 0.41 0.05 C hirta 0.73 0.60 0.55 0.08
T.fragiferum 0.91 0.84 0.79 0.45
T. repens 1.02 0.74 0.84 0.30
0.09 0.06 0.18 0.20 0.10 0.10 0.41 0.19
0.42 0.28 0.29 0.34
0.39 0.30 0.58 0.56
Sutherland & Stillman 1988; de Kroon & Schieving 1990; Oborny 1994). Responses of both Carex and Tr/-
folium species are different from those needed for the selective placement of ramets in the more favourable patches. Apparently, all four species do not forage for nutrient-rich patches by plasticity in spacer mor- phology, and while Carex hirta seems more plastic than C. flacca in its spacer characters, in Trifolium the species from the infertile habitat seems more plastic. These results corroborate the conclusion by de Kroon & Hutchings (1995) that apparently very few clonal plants species respond to fertilization by a shortening in spacer length. Thus, foraging cannot unequivocally be demonslrated.
As an alternative to foraging there is another pos- sible interpretation for the results on spacer and ramet characters in Trifolium. While Caradus et al. (1993) showed that stolon internodes of T. repens expressed a hump-backed response curve to seven levels of phos- phorus supply, we found that the spacer responses in Trifolium are similar to that for stolon internodes of Cynodon dactylon (Dong & de Kroon 1994). Such a response to higher nutrient supply by forming larg- er structures has preliminarily been considered to be growth-related (see Hutchings & de Kroon 1994; Cain 1994).
Longer spacers formed under higher nutrient lev- els in the Trifolium species ensure a longer distance between interconnected ramets along a stolon and may thus reduce ramet density in nutrient-rich patches. Such responses may moderate to some extent the high- er intraclonal competition for light due to formation of larger ramets with larger leaf area under nutrient- rich conditions (see Sutherland & Stillman 1988). This interpretation may apply in all cases where plants have ramets with diameters that are similar in size or larger than the average spacer length. Such an interpretation
would also accord with the strong and highly plastic response in leaf area and ramet height (petiole length) in Trifolium repens as compared to T. fragiferum and in leaf area response of Carex hirta as compared to C. flacca.
Acknowledments
We would like to thank Hans de Kroon and an anony- mous reviewer for valuable comments on an earlier version of the manuscript. We are grateful to Jan van Rheenen, Ren6 Kwant, Leonard P.M. Bik for assis- tance during the experiment.
References
Bell, A.D. 1984. Dynamic morphology: a contribution to plant pop- ulation ecology, pp. 48-56. In: Dirzo, 13. & Sarukhan, J. (eds), Perspectives in plant population ecology. Sunderland, MA, Sin- auer.
Bell, A.D. 1991. Plant form: an illustrated guide to flowering plant morphology. Oxford University Press, New York.
Bell, A.D. & Tomlinson, P.B. 1980. Adaptive architecture in rhi- zomatous plants. Botanical Journal of the Linnean Society 80: 125-160.
Bradshaw, A.D. 1965. Evolutionary significance of phenotypic plas- ticity in plants. Advances in Genetics 13:115-156.
Burdon, J.J. 1983. Biological flora of the british isles: Trifolium repens L. Journal of Ecology 71: 307-330.
Cain, M.L. 1994. Consequences of foraging in clonal plant species. Ecology 75: 933-944.
Caradus, J.R., Hay, M.J.M,, Mackay, A.D., Thomas, V.J., Dunlop, J., Lambert, M.G., Hart, A.L., van den Bosch, J. & Wewala, S. 1993. Variation within white clover (Trifolium repens L.) for phenotypie plasticity of morphological and yield related characters, induced by phosphorus supply. The New Phytologist 123: 175-184.
Carlsson, B.A. & Callaghan, T.V. 1990. Programmed tiller differ- entiation, intraclonal density regulation and nutrient dynamics in Carex bigelowii. Oikos 58: 219-230.

192
Carlsson, B.A., Jonsdottir, I.S., Svensson, B.M. & Callaghan, T.V. 1990. Aspects of clonality in the arctic: a comparison between Lycopodium annotinum and Carex bigelowii, pp. 131-151. In: van Groenendael, J. & de Kroon, H. (eds), Clonal growth in plants: regulation and function. SPB Academic Publishing, The Hague.
Cook, R.E. 1985. Growth and development in clonal plant popula- tion. pp. 259-296. In: Jackson, J.B.C., Buss, L.W. & Cook, R.E. (eds), Population biology and evolution ofclonal organisms. Yale University Press, New Haven.
Crick, J.C. & Grime, J.P. 1987. Morphological plasticity and mineral nutrient capture in two herbaceous species of contrasted ecology. The New Phytologist 107: 403-414.
de Kroon, H. & Hutchings, M.J. 1995. Morphological plasticity in clonal plants: The foraging concept reconsidered. Journal of Ecology 83: 143-152.
de Kroon, H. & Knops, J. 1990. Habitat exploration through mor- phological plasticity in two chalk grassland perennials. Oikos 59: 39-49.
de Kroon, H. & Schieving, F. 1990. Resource partitioning in relation to clonal growth strategy, pp. 113-130. In: van Groenendael, J. & de Kroon, H. (eds), Clonal growth in plants: regulation and function. SPB Academic Publishing, The Hague.
de Kroon, H. & van Groenendael, J. 1990. Regulation and function of clonal growth in plants: an evaluation, pp. 177-186. In: van Groenendael, J. & de Kroon, H. (eds), Clonal growth in plants: regulation and function. SPB Academic Publishing, The Hague.
Dong, M. 1993. Morphological plasticity of the clonal herb Lami- astrum galeobdolon (L.) Ehrend. & Polatschek in response to partial shading. The New Phytologist 124: 291-300.
Dong, M. 1995. Morphological responses to local light conditions in clonal herbs from contrasting habitats, and their modification due to physiological integration. Oecologia 101: 282-288.
Dong, M. & de Kroon, H. 1994. Plasticity in morphology and biomass allocation in Cynodon dactylon, a grass species forming stolons and rhizomes. Oikos 70: 99-106.
Dong, M. & Pierdominici, M.G. 1994. Morphology and growth of stolons and rhizomes in three clonal grasses, as affected by different light supply. Vegetatio 116: 25-32.
Evans, J.E 1992. The effect of local resource availability and clon- al integration on ramet functional morphology in Hydrocotyle bonariensis. Oecologia 89: 265-276.
Grime, J.E 1979. Plant strategies and vegetation processes. John Wiley & Sons, Chichester.
Grime, J.E, Crick, J.C. & Rincon, J.E. 1986. The ecological signif- icance of plasticity, pp. 5-29. In: Jennings, D.H. and Trewavas, A.J. (eds), Plasticity in plants. Biologists Limited, Cambridge.
Grime, J.E, Hodgson, J.G. & Hunt, R. 1988. Comparative plant ecology. Unwin Hyman, London.
Hutchings, M.J. & de Kroon, H. 1994. Foraging in plants: the role of morphological plasticity in resource acquisition. Advances in Ecological Research 25: 159-238.
Hutchings, M.J. & Mogie, M. 1990. The spatial structure of clonal plants: control and consequences, pp. 57-76. In: van Groenen- dael, J. & de Kroon, H. (eds), Clonal growth in plants: Regulation and function. SPB Academic Publishing, The Hague.
Hutchings, M.J. &Slade, A.J. 1988. Morphological plasticity, for- aging and integration in clonal perennial herbs, pp. 83-109. In: Davy, A.J., Hutchings, M.J. & Watkinson, A.R. (eds), Plant population ecology. BlackweU Scientific Publications, Oxford.
Jerling, L. 1988. Clone dynamics, population dynamics and vege- tation pattern of Glaux maritima on a Baltic sea shore meadow. Vegetatio 74: 171-185.
Landolt, E. 1977. Okologische Zeigerwerte zur Schweizer Flo- ra. VerOffentlichungen des Geobotanischen Institutes der Eidg. Techn. Hochschnle, Stiftung Rtibel, ZUrich. Vol. 64.
Leakey, R.R.B. 1981. Adaptive biology of vegetatively regenerating weeds. Advances in Applied Biology 6: 57-90.
Lovell, P.H. & LoveD, P.J. 1985. The importance of plant form as a determining factor in competition and habitat exploitation. pp. 209-221. In: White, J.(ed), Studies on plant demography. Academic Press Inc., London.
Mogie, M. & Hutchings, M.J. 1990. Phylogeny, ontogeny and clonal growth in vascular plants, pp. 3-22. In: van Groenendael, J. & de Kroon, H. (eds), Clonal growth in plants: regulation and function. SPB Academic Publishing, The Hague.
Oberdorfer, E. 1970. Pflanzensoziologische Exknrsionsflora fur Silddeutschland. Eugen Ulmer, Stuttgart.
Obomy, B. 1994. Growth rules in clonal plants and environmental predictability: A simulation study. Journal of Ecology 82: 341- 351.
SAS. 1985. SAS/STAT Guide for personal computers Version 6 Edition. SAS institute Inc., Cary.
Scheiner, S.S. 1993. Genetics and evolution of phenotypic plasticity. Annual Review of Ecology and Systematics 24: 35-68.
Schlichting, C.D. 1986. The evolution of phenotypic plasticity in plants. AnnualReview of Ecology and Systematics 17: 667-693.
Schlichting, C.D. & Levin, D.A. 1984. Phenotypic plasticity of annual Phlox: Tests of some hypotheses. American Journal of Botany 71: 252-260.
Schmid, B. 1986. Spatial dynamics and integration within clones of grassland perennials with different growth form. Proceedings of the Royal Society, London, B 228: 173-186.
Schmid, B. & Bazzaz, EA. 1992. Growth responses of rhizomatous plants to fertilizer application and interference. Oikos 65:13-24.
Slade, A.J. & Hutchlngs, M.J. 1987a. The effect of nutrient availabil- ity on foraging in the clonal herb Glechoma hederacea. Journal of Ecology 75: 95-112.
Slade, A.J. & Hutchings, M.J. 1987b. The effect of light intensity on foraging in the clonal herb Glechoma hederacea. Journal of Ecology 75: 639-650.
Sutherland, W.J. & Stillman, R.A. 1988. The foraging tactics of plants. Oikos 52: 239-244.
Taylor, F.J. 1956. Biological flora: Carexflacca Schreb. Journal of Ecology 44: 281-290.
Thompson, L. 1993. The influence of natural canopy density on the growth of white clover, Trifolium repens. Oikos 67: 321-324.
Via, S. 1987. Genetic constraints on the evolution of phenotypic plasticity, pp. 47-71 in: Loeschke, V. (ed.), Genetic constraints on adaptive evolution. Springer, Berlin.
Waller, D.M. & Steingraeber, D.A. 1985. Branching and modular growth: theoretical models and empirical patterns, pp. 225-257. In: Jackson, B.C., Buss, L.W. & Cook, R.E. (eds), Population biology and evolution of clonal organisms. Yale University Press, New Haven.
White, J. 1979. The plant as metapopulation. Annual Review of Ecology and Systematics 10: 109-145.
Zohary, M. & Heller, D. 1984. The genus Trifolium. Ahva Printing Press, Jerusalem.