Embed Size (px)
Citation preview

J . Zool., Lond. (1984) 204,241-257
Morphology, growth, food habits and population characteristics of the Smooth snake CoroneZZu uustriucu in southern Britain
P A U L G O D D A R D * Department of Biology, The University, Southampton SO9 5NH
(Accepted 14 February 1984)
(With 7 figures in the text)
A three year ecological study was made of the Smooth snake in the New Forest, Hampshire. The aim was to provide quantitative information on particular aspects ofthe Smooth snake's life-history characteristics. On three study sites, snakes were captured by hand and individu- ally marked. On capture, each snake was identified, sexed, measured, weighed and examined for the presence of food in its stomach and hind gut. In general, mature Smooth snakes showed sexual dimorphism in morphology and growth rates. This was thought to reflect the influence of body size on female reproduction. It was found that Smooth snakes preyed on both small mammals and lizards, suggesting that this species is an opportunist predator. Diet did not vary between study sites. The population sizes were estimated by direct enumeration and densities of one and two snakes per hectare were found. The sex ratios in each study site population were even, and the age structures suggested that both sexes exhibited similar mortality. Consideration was also given to the estimation of the total population size of Smooth snakes in Britain.
Introduction . . . . Materials and methods . .
Study sites . . . . Sample procedures . . Statistical methods . .
Results . . . . . . Morphology . . . . Growth . . . . . . Food habits . . . . Population characteristics
Size and sex ratio . . Density . . . . Age structure . .
Discussion . . . . Morphology . . . . Growth . . . . . . Food habits . . . . Population characteristics
Summary . . . . . . References . . . .
Contents . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
. .
Page 242 242 242 242 243 243 243 245 241 249 249 250 25 1 252 252 253 2 54 254 255 256
"Present address: Brathay Hall Trust, Brathay Hall, Ambleside, Cumbria LA22 OHP, U.K. 24 1
0022-5460/84/ I0024 I + 17 $03.00/0 0 1984 The Zoological Society of London

242 P. GODDARD
Introduction The Smooth snake Coronellu austriuca Laurenti is one of three species of snake indigenous
to Britain. Unlike the Grass snake Nutrix nutrix Lacepbde and the adder Viperu berus L., the Smooth snake is rarely encountered and consequently is not well known. It is a slender, viviparous snake which is found only in the lowland heaths of central southern England. In recent years, concern has been expressed for the conservation of the Smooth snake since its distribution is decreasing in Britain, Scandinavia and throughout Europe. This decline has been attributed to the destruction of its habitat (Honeggar, 1978). In Britain, the Smooth snake is listed as a protected species under schedule five of the Wildlife and Countryside Act 198 1.
Published data on the life-history characteristics of Smooth snakes are sparse. Apart from a detailed post-mortem, study of specimens captured in central France by Rollinat (reported by Duguy, 1961), and a field investigation in southern England by Spellerberg & Phelps (1977), much of the available information is anecdotal in nature. This paper concerns particular aspects of the ecology of Smooth snakes (morphology, pattern of growth, food habits and population characteristics) captured in three areas in the New Forest, Hampshire during the period 1976-1978 inclusive.
Materials and methods Study sites
Three field sites in the New Forest, Hampshire were used in the present study. Site I was an area of heathland which had been afforested and enclosed in 1959. At the time of the study, this site was a mosaic of dense coniferous forest (primarily Pinus spp.) and heathland, comprising heather (CaNuna vulgaris (L.) Hull), Bell heather (Erica cinerea L.), Crossed-leaved heath (Erica tetralix L.) and Purple- moor grass (Molinea caerulea (L.) Moench). Gorse (Ulex europaeus L. and Ulex minor Roth) also provided a large amount of ground cover. The site was dissected by numerous forest rides and paths. Specimens of Smooth snakes were captured in most areas of the site which were not heavily forested. Part of site 1 had been used for an earlier study of Smooth snakes by Spellerberg & Phelps ( 1 977).
Site 2 was situated 2 km from site 1 . It was a less clearly defined area than site 1 since it comprised an area of unenclosed, undulating open heathland. Tree cover was minimal and grazing livestock had free access to all parts of the heath. A major feature of this site was a south-facing embankment which ran through part of the study area. This embankment was covered by a mixture of bare substratum, grass (chiefly Agrostis setacea Curtis), heath species and gorse. Smooth snakes were captured throughout the site but, in particular, on or near the south-facing embankment.
Site 3 was situated in an area of heathland which was changing to woodland as a result of natural pine invasion. The area was heavily grazed by livestock and in places there was evidence of recent damage by fire. On this site, specimens were captured near to a shallow stream.
The sizes of the areas searched on sites I , 2 and 3 were 80 ha, 75 ha and 30 ha, respectively.
Sample procedures Between March 1976 and November 1978 inclusive, 324, 205 and 54 visits were made to sites I ,
2 and 3, respectively. Each site was sampled by walking a predetermined route through the habitat, checking known localities for snakes and visiting additional areas. Snakes were captured by hand, either basking or moving in the open, or hidden beneath tin sheeting, brick tiles and concrete slabs. A number of these objects were strewn across sites I and 2, and these were checked on each visit.
Snakes were individually marked by clipping their ventral scales (Spellerberg, 1977~) . None of the snakes showed any signs of having suffered from the effects of marking. A snake which was too small
THE SMOOTH SNAKE IN SOUTHERN BRITAIN 243
to mark for fear of inflicting injury was subsequently identified according to the pigmentation pattern on the dorsal surface of its neck and head.
On capture, each snake was identified, sexed, measured for snout-vent length and tail length, palpated to cause the regurgitation of any stomach contents or the voiding of any faecal material, and weighed using a “Persola” spring balance. After processing, the snake was released at the site ofcapture.
Statistical methods The statistical procedures in this paper follow those of Sokal & Rohlf (1969) (Mann-Whitney test:
Q Wilcoxon 2 sample test: t ,$) and Meddis (1975) (Fisher’s exact test: Z; Kolmogorov-Smirnov 2 sample test: K ; Mann-Whitney test: R ; Chi squared test: f ) . Where applicable, the data from the study sites have been pooled according to the results of the F(max)-test or Bartlett’s test for the homogeneity of variance (see Goddard, 198 1 a).
Results Morphology
In total, measurements were collected of 22 sexually immature (individuals less than 33.0 cm in snout-vent length) and 92 sexually mature snakes. The snout-vent lengths (SVLs), tail lengths (TLs), relative tail lengths (RTLs calculated as SVL/TL), and body weights (BWTs) of those specimens are summarized in Table 1. The BWTs of mature females were divided into those belonging to breeding or non-breeding condition individuals, depending on whether or not they carried ova in their oviducts either at the time of first capture or at a subsequent date during the same season.
Immature snakes There were no significant differences in the SVL. TL and BWT measurements between
the sexes (R = 65.50, R = 52.00 and R = 58.00, respectively, n , = 7 and n, = 15, P > 0.05) in
TABLE I Body measurements qj‘rmmalure and mature Smooth snakes
Snout-vent length Tail length Body weight (cm) (cm) Relative tail length (g)
Standard Standard Standard Standard n Mean error Mean error Mean error Mean error
Immature snakes Males 15 25.38 2.45 6.81 0.32 3.86 0.02 10.86 2.91 Females 7 22.30 7.03 4.57 0.36 4.9 1 0.03 6.71 2.75
Mature snakes Males 54 40.28 0.17 11.63 0.03 3.48 0.001 36.88 1.23 Females 38 42.27 0.66 8.81 0.02 4.8 I 0.006 47.68* 6.38 (25)
3 l.46** 8.02 (13)
*Breeding females with sample size in parenthesis. **Nowbreeding females with sample size in parenthesis.
244 P. GODDARD
0 0
0 0 0 0
0.
0 .
0 0
0
0
0
I . 0 0
1 ' " ' 1 8 " '
15 20 25 3C SVLkrn 1
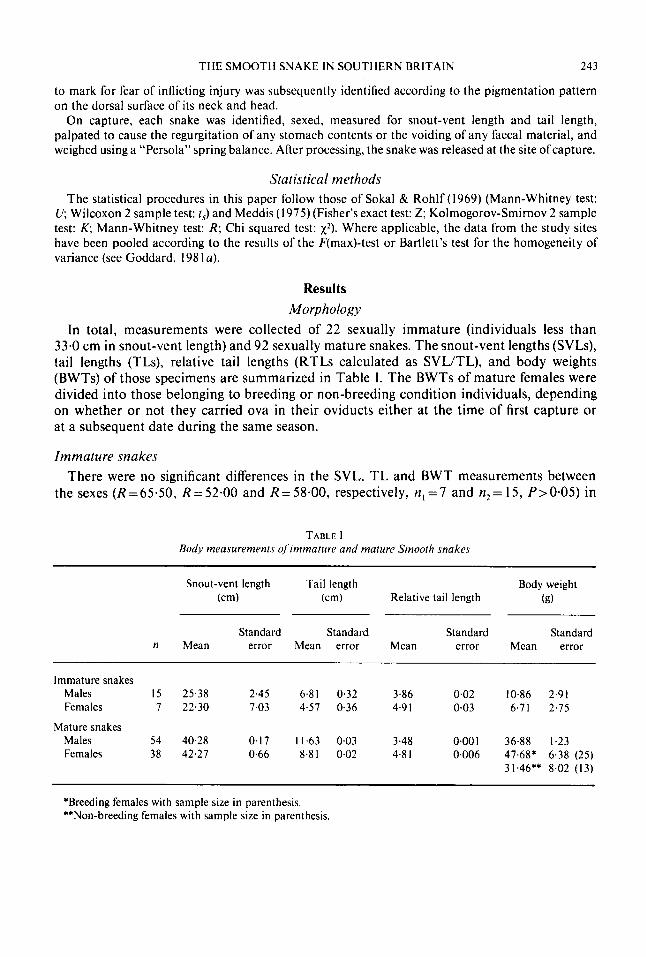
FIG. I . The relationship between snout-vent length (SVL) and body-weight (BWT) in immature snakes (0 males, BWT =0.001 SVL*'*71 and 0 females, BWT=0.006SVL224°).
all cases. However, male snakes had a smaller RTL than female snakes (R=35.00, n , = 7 and nz= 15, P<0.05). There was also evidence of an ontogenetic difference in RTL between immature and mature males. The former had shorter tails in proportion to their SVLs than the latter (t ,=3.01, n = 6 9 , P<O.OOI).
The allometric relationship BWT = aSVL was assessed separately for males and females using Bartlett's three group method for model I1 regression (Sokal & Rohlf, 1969). Double logarithmic plots of this relationship are shown in Fig. 1. In both sexes a linear function is described in which BWT is highly correlated with SVL. Thus large snakes are relatively heavier than small snakes.
Mature snakes In general, females had longer SVLs and shorter TLs than males (t.,= 1.98, P<O.O5 and
t, = 7.85, P> 0.00 1 , respectively, n = 92 in both cases). Furthermore, females had relatively shorter tails than males of an equivalent SVL (t,$= 8.07, n = 92, P> 0.00 1).
There was no significant difference in BWT between non-breeding female and male snakes ( t ,= 1.75, n = 67, P> 0.05). However, breeding females were heavier than both non-breeding
THE SMOOTH SNAKE IN SOUTHERN BRITAIN
55
50
45
40
35
I
0
I- - 30- 2
25
20-
15.
245
* * * * * * * -
- * * *
* * - - * * *,a?
*” a *
* * * * - b
I I I I I 1 1 1 1 1 I I I I I
32 33 34 35 36 37 38 39 40 41 42 43 44 45 46 47 48 SVL I cm)
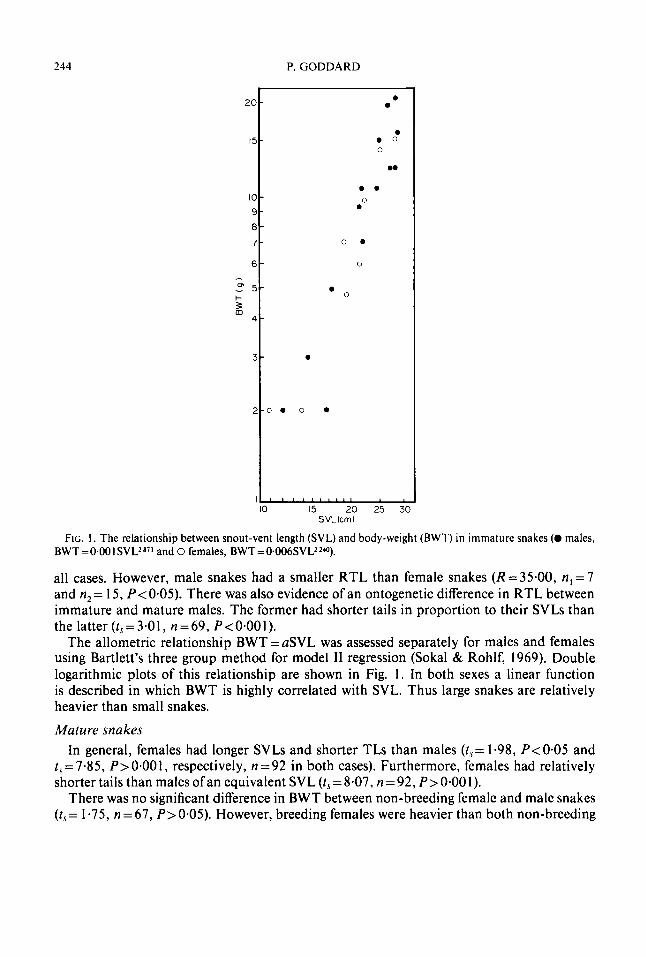
Fic. 2. The relationship between snout-vent length (SVL) and body weight (BWT) in mature males (BWT = 0.005SVL2’384).
females and males (t,=3.63, n=38, and t,=40.39, n=69, respectively, P < O . O O l in both cases).
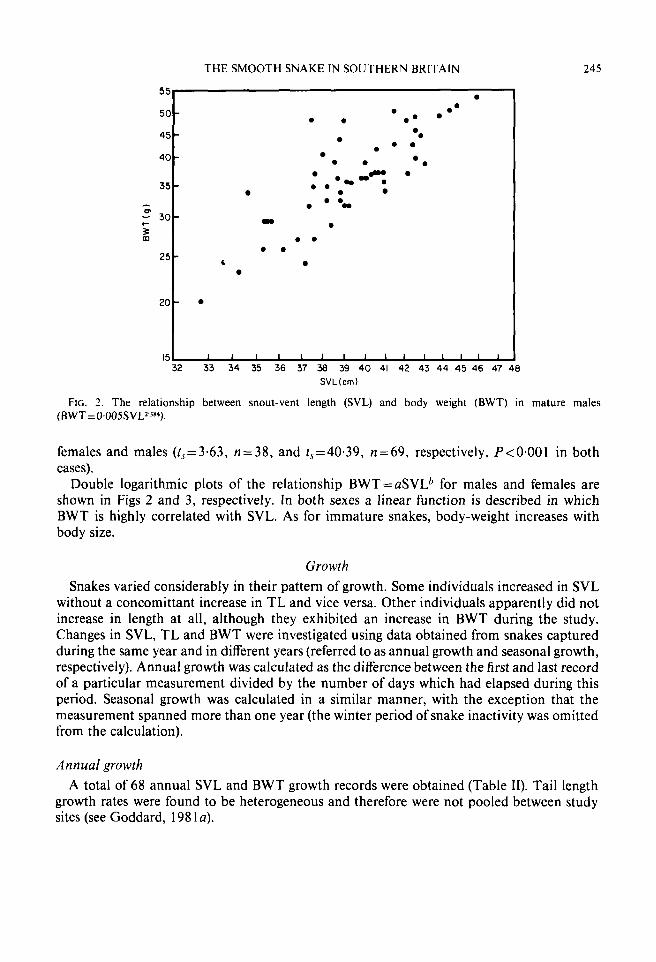
Double logarithmic plots of the relationship BWT=aSVL6 for males and females are shown in Figs 2 and 3, respectively. In both sexes a linear function is described in which BWT is highly correlated with SVL. As for immature snakes, body-weight increases with body size.
Growth Snakes varied considerably in their pattern of growth. Some individuals increased in SVL
without a concomittant increase in TL and vice versa. Other individuals apparently did not increase in length at all, although they exhibited an increase in BWT during the study. Changes in SVL, TL and BWT were investigated using data obtained from snakes captured during the same year and in different years (referred to as annual growth and seasonal growth, respectively). Annual growth was calculated as the difference between the first and last record of a particular measurement divided by the number of days which had elapsed during this period. Seasonal growth was calculated in a similar manner, with the exception that the measurement spanned more than one year (the winter period of snake inactivity was omitted from the calculation).
Annual growth A total of 68 annual SVL and BWT growth records were obtained (Table 11). Tail length
growth rates were found to be heterogeneous and therefore were not pooled between study sites (see Goddard, 198 1 a).
246
45L
40
- 3 5 -
I- 30- 3 m
m -
25
20
P. GODDARD
-
-
-
. 0
b. .
0
0 15 1 0 0
1 I 1 1 0 , 1 1 I I I I 1 1 I I I I I I I 1 I 1 I
32 33 34 35 36 37 30 39 40 41 42 43444546 47 4849 5051 52535455 SVL h)
FIG. 3. The relationship between snout-vent length (SVL) and .body weight (BWT) in breeding (0, BWT=0.039SVLl.888) and non-breeding (0, BWT=0.01 3SVL21M) mature females.
TABLE I 1 Annual growth rates for immature and mature Smooth snakes
Snout-vent length Tail length (cm/da y) (cmlday)
Standard Standard Standard n Mean error Mean error Mean error
Immature snakes Males 7 0.01849 0.00001 0.00529 0.000003 0.03844 0.00026 Females 2 0.02875 0.00038 0.00200 0.000005 0 0
Males 39 0.01 134 0*000009 - - -0.01908 0.00032 Females 20* 0.00567 0.000003 - - - 0.03362 0.000 I 7
2** 0.00208 0.0 17645 - - 0.04166 0.06863
Mature snakes
*Breeding females. **Non-breeding females.
There were no significant differences in the SVL, TL or BWT annual growth rates between immature males and females ( U z 8 . 5 , U= 10.0 and U= 13.5, respectively, n, = 7 and n 2 = 2 , P>0.05 in all cases).
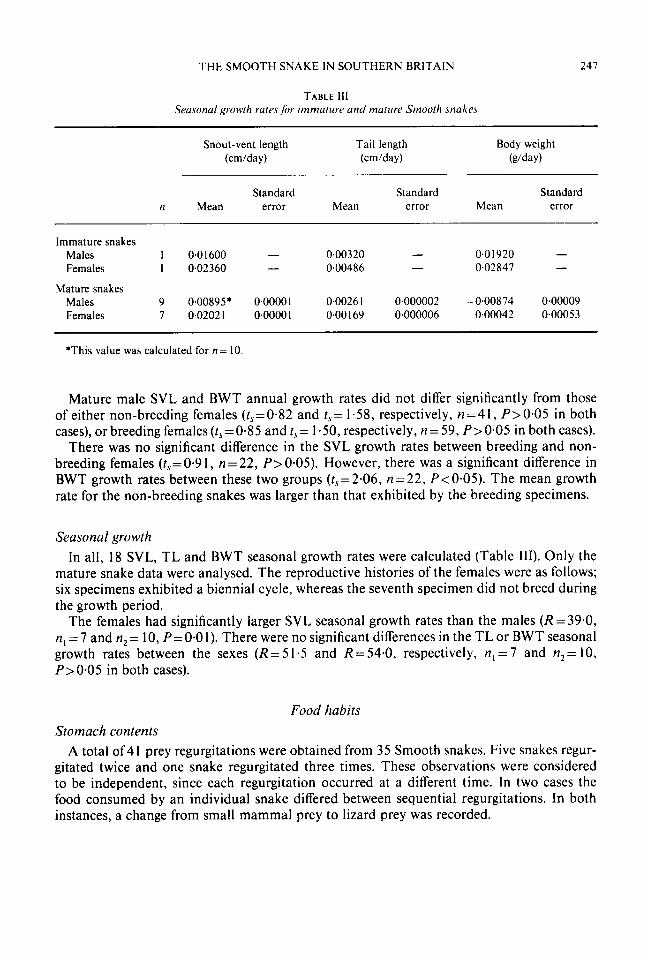
THE SMOOTH SNAKE IN SOUTHERN BRITAIN 247
TABLE I l l Seasonal growth rates fbr immalure and matitre Smooth snakes
Snout-vent length Tail length (cmlday) (cm/da y)
Standard Standard Standard n Mean error Mean error Mean error
Immature snakes 1 0.01600 - 0.00320 - 0.01920 - I 0.02360 - 0.00486 - 0.02847 -
Males Females
Mature snakes Males 9 0,00895* 0.00001 0.0026 1 0.000002 -0’00874 0.00009 Females 7 0.02021 0~00001 0.00 169 0.000006 0.00042 0.00053
*This value was calculated for n= 10.
Mature male SVL and BWT annual growth rates did not differ significantly from those of either non-breeding females (t,=0.82 and t,= 1.58, respectively, n=41, P>O.O5 in both cases), or breeding females (t, = 0.85 and t, = 1.50, respectively, n = 59, P> 0.05 in both cases).
There was no significant difference in the SVL growth rates between breeding and non- breeding females (t,=0.9 1, n = 22, P>0.05). However, there was a significant difference in BWT growth rates between these two groups (t,=2.06, n=22, P<0-05). The mean growth rate for the non-breeding snakes was larger than that exhibited by the breeding specimens.
Seasonal growth In all, 18 SVL, TL and BWT seasonal growth rates were calculated (Table 111). Only the
mature snake data were analysed. The reproductive histories of the females were as follows; six specimens exhibited a biennial cycle, whereas the seventh specimen did not breed during the growth period.
The females had significantly larger SVL seasonal growth rates than the males (R = 39.0, n, = 7 and n2 = 10, P=O.O 1). There were no significant differences in the TL or BWT seasonal growth rates between the sexes ( R = 5 1.5 and R= 54.0, respectively, n , = 7 and n2= 10, P>0.05 in both cases).
Food habits Stomach contents
A total of41 prey regurgitations were obtained from 35 Smooth snakes. Five snakes regur- gitated twice and one snake regurgitated three times. These observations were considered to be independent, since each regurgitation occurred at a different time. In two cases the food consumed by an individual snake differed between sequential regurgitations. In both instances, a change from small mammal prey to lizard prey was recorded.

248 P. GODDARD
TABLE IV Items ofprey regurgitated by Smooth snakes
Site 1 Site 2 Site 3
No. of No. of No. of No. of No. of No. of Prey snakes* items** snakes* items** snakes* items**
Adult shrews 3 3 5 5 I 1 Nestling shrews 1 5 2 7 1 2 Nestling rodents 1 1 1 I 0 0 Small mammals
(unidentified nestlings) 7 14 5 9 0 0 Common lizards 8 9 5 5 I I
*Total number of snakes containing food in which that prey type was found. **Total number of food items recorded which were of that prey type.
No snake regurgitated more than one type of prey on the same occasion. However, it was not uncommon to find more than one individual of a certain prey type being regurgitated. This was particularly true for snakes which had eaten nestling prey (young shrews and young rodents in underground nests). By comparing the state of digestion of each item, it was appar- ent that, in all cases, the prey items had been taken within a short time of each other. The greatest number of items obtained from a single regurgitation were five unidentified nestling prey. The number and type of prey items consumed are summarized in Table IV.
Out of 3 1 adult snakes examined, 26 (84%) regurgitated small mammal prey of which 1 1 (40%) regurgitations were of nestling prey. In contrast, only one of the eight immature snakes examined regurgitated a nestling mammal. The remaining seven immature snakes had consumed lizard prey.
Taking into account the two main dietary groups (small mammals and lizards), there was no significant difference in the food habits of snakes captured on site 1 and those captured on site 2 (Z=0.46, n=38, b 0 . 0 5 ) . There was also no association between the number of ingested items and the locality of the snakes (Z=0.90, n=64, P>0.05).
Faecal material A reference collection of faecal material was prepared prior to the sorting of the field
samples. Twelve captive snakes were fed various items of prey. Their faeces were collected and then checked against the type of prey known to have been consumed.
A total of 54 faecal samples were collected from snakes captured in the field (Table V). There was no significant association between the prey types identified and the site of snake capture (sites 1 and 2) (Z=0.59, n = 5 l ,P>O.O5). An almost equal number of faecal samples contained either small mammal or lizard remains (2 1 and 19, respectively). Seven samples contained both prey items. With the exception of one sample, which contained the remains of a legless lizard Anguisfragilis L., all the lizards recorded were common lizards Lacerta vivipara Jacquin.

THE SMOOTH SNAKE IN SOUTHERN BRITAIN
TABLE V Prey types idenr(jied,fiom Smooth snake jaecal marerial
249
No. of snakes
Prey Site 1 Site 2 Site 3
Small mammals 14 6 1 Small mammals plus lizards 5 2 0 Lizards 9 8 2 Nothing identified 5 I I
TABLE VI Number of individual Smooth snakes captured
Number of individual
snakes marked
Inclusive period Males Females Total
Site 1 1972-1978 41 31 72 Site2 1976-1978 34 20 54 Si te3 1977-1978 5 5 10
Five Smooth snakes regurgitated and defecated on the same occasion. In two cases, lizard prey were identified in the faecal sample whereas small mammal prey had been regurgitated. In the remaining three cases, the prey type identified in the sample matched the regurgitated items.
Faecal material was collected from six immature snakes. Two of the samples contained small mammal remains, a further two samples contained lizard remains, and nothing was identified in the remaining two samples.
Population characteristics Size and sex ratio
The population sizes of Smooth snakes on sites 1 , 2 and 3 were estimated by direct enumeration. The number of individuals captured during the inclusive period 1972-1978 is shown in Table VI. Snakes captured prior to 1976 were marked by Spellerberg (pers. comm.). There was no significant departure from an even sex ratio on site 1 ( x 2 = 1.38, d.f = 1, P > 0.03, site 2 ( x 2 = 3.63, d.f. = I , P > 0.05) or site 3 (even numbers of each sex).
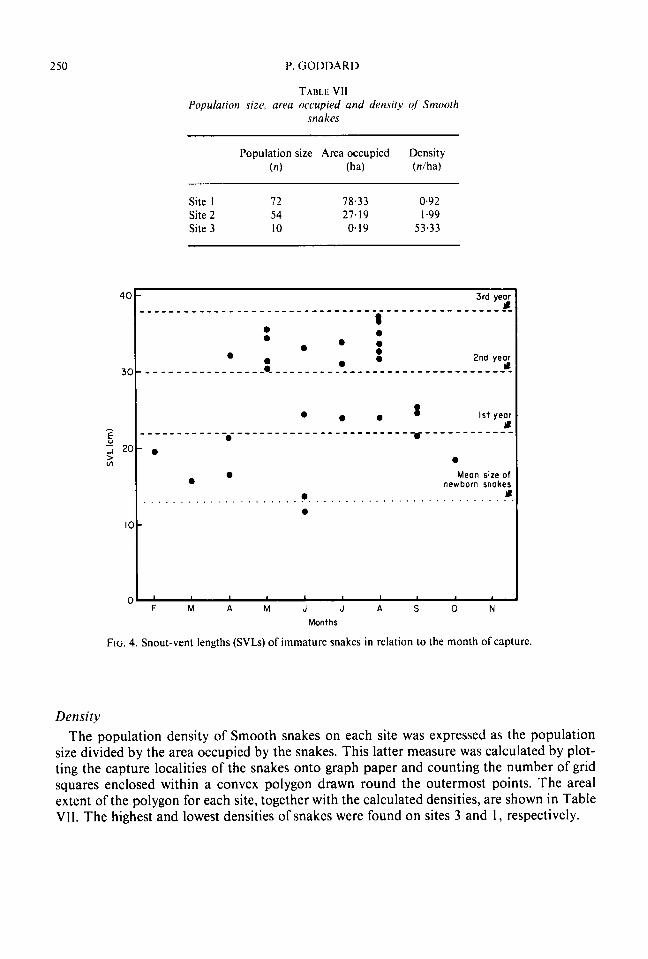
250 P. GODDARD
TABLE VII Population size. area occupied and density of Smooth
snakes
Population size Area occupied Density ( n ) (ha) (nlha)
Site 1 12 78.33 0.92 Site 2 54 27.19 1.99 Site 3 10 0.19 53.33
Density The population density of Smooth snakes on each site was expressed as the population
size divided by the area occupied by the snakes. This latter measure was calculated by plot- ting the capture localities of the snakes onto graph paper and counting the number of grid squares enclosed within a convex polygon drawn round the outermost points. The areal extent of the polygon for each site, together with the calculated densities, are shown in Table VII. The highest and lowest densities of snakes were found on sites 3 and 1, respectively.
T H E S M O O T H SNAKE IN SOUTHERN BRITAIN 25 I
1976 1977 I 1978
I ~ l l l l l l l r l l l r l l l l l l r l 1 0 2 3 2 1 0 1 2 5 4 3 2 1 0 1 2 3 4 5
Number of snokes
FIG. 5 . Population age structure of snakes captured on site I in 1976, 1977 and 1978 (B males, 0 females).
1977
10+
1978
0
a
3
2
I
I I I I I I I I I I 1 1 1 1 1 1 1 1 1 1 1 3 2 1 0 1 2 3 4 6 5 4 3 2 1 0 1 2 3 4
Number of snakes
FIG. 6. Population age structure of snakes captured on site 2 in 1977 and 1978 (m males, 0 females).
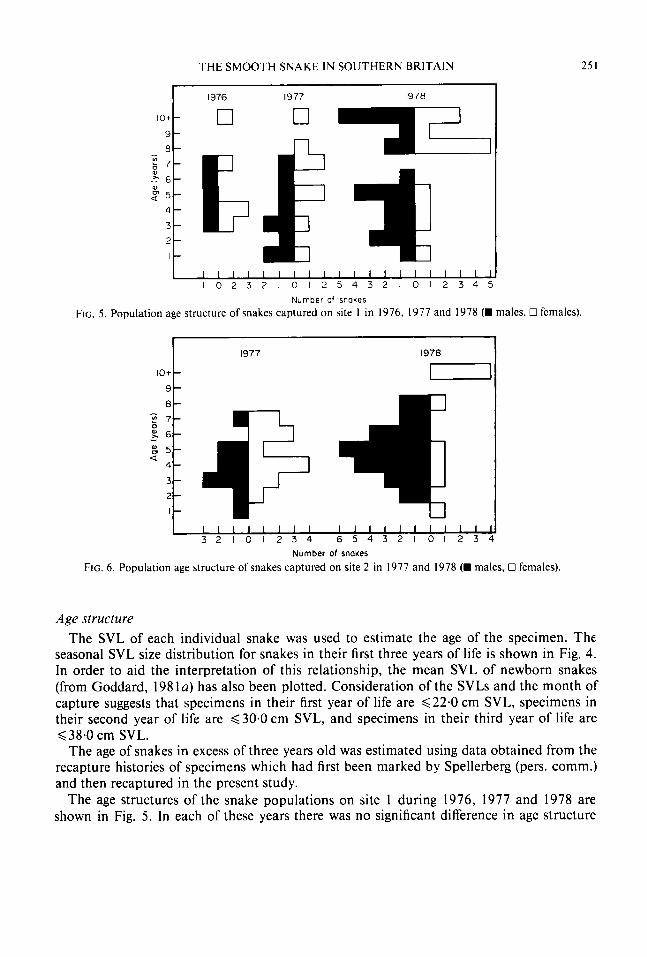
Age structure The SVL of each individual snake was used to estimate the age of the specimen. The
seasonal SVL size distribution for snakes in their first three years of life is shown in Fig. 4. In order to aid the interpretation of this relationship, the mean SVL of newborn snakes (from Goddard, 198 la) has also been plotted. Consideration of the SVLs and the month of capture suggests that specimens in their first year of life are 622.0 cm SVL, specimens in their second year of life are d30.0 cm SVL, and specimens in their third year of life are 638.0 cm SVL.
The age of snakes in excess of three years old was estimated using data obtained from the recapture histories of specimens which had first been marked by Spellerberg (pers. comm.) and then recaptured in the present study.
The age structures of the snake populations on site 1 during 1976, 1977 and 1978 are shown in Fig. 5. In each of these years there was no significant difference in age structure

252 P. GODDARD
1 0 1 2 1 0 1 2 Number of snakes
FIG. 7. Population age structure of snakes captured on site 3 in 1977 and 1978 (m males, 0 females).
between the sexes (K=0.29, n= 12; K=0.37, n=20 and K= 1.05, n = 3 4 , respectively, P> 0.05 in all cases).
The age structures of the snake populations on site 2 in 1977 and 1978 are shown in Fig. 6. There was no significant difference between the sexes in either year (K= 0.68, n = 23 and K= 1.2 I , n = 36, respectively, P> 0.05 in both cases).
The age structures of the snake populations on site 3 in 1977 and 1978 are shown in Fig. 7.
In general, on all three sites in each year of the study, the number of one-year-old and two-year-old snakes captured was low. Snakes which were three or more years of age pre- dominated in the populations. There was no significant difference in the age structure of the populations between sites 1 and 2 in either 1977 (K=0.92, n = 4 3 , P>0.05) or 1978
Discussion Morphology
Sexual dimorphism in SVL is not an uncommon occurrence in snakes (Fitch, 1970), although it has not been noted before in the Smooth snake. Generally, in those species exhi- biting sexual dimorphism, females are larger than males. This has been attributed to female fitness being size-dependent. It has been postulated that natural selection favours a large female SVL because fecundity is proportional to body size (Shine, 1978~). Support for this hypothesis is equivocal in the present species. The number of young produced by female Smooth snakes varies little over a wide range of body sizes (Goddard & Spellerberg, 1980). However, two alternative explanations may be suggested.
Large female Smooth snakes produce large offspring (Goddard, 198 1 a). Compared with smaller individuals, large offspring are more likely to have an increased chance of survival by virtue of (i) more adequate post-natal food reserves and (ii) an ability to tackle a wide size range of prey. Thus, in females, fitness may be expected to be size-dependent due to the
(K= 1.09, n = 68, P> 0.05).

T H E S M O O T H SNAKE IN S O U T H E R N BRITAIN 253
increased advantages accruing to large offspring. This appears to be the case in vipers in which large newborn snakes have a lower mortality than their smaller counterparts (Saint Girons & Naulleau, 198 1).
A second explanation is that there may be a greater likelihood of courtship and mating among large rather than small female snakes. Hawley & Aleksiuk (1976) have shown that the amount of courtship among garter snakes ( Tharnnophis) is positively correlated with SVL. Large females produce a stronger olfactory stimulus than small females, hence they are more attractive to males for the purposes of mating. Female Smooth snakes which are in a reproductive condition also produce an olfactory stimulus (Brodin, 195 1 ; Duguy, I96 1) and its strength may be similarly size-dependent.
The occurrence of sexual dimorphism in the TL of the Smooth snake is well documented (Duguy, 1961; Bruno, 1966; Smith, 1973; AndrCn & Nilson, 1976; Spellerberg & Phelps, 1977). The large length ofthe tail in male specimens is presumably required to accommodate the hemipenes and their associated musculature. It may also aid the male in holding onto the female during mating. Similarly, the ontogenetic difference in RTL among males rather than females suggests that this, too, is related to the growth of the hemipenes.
The difference in BWT between non-breeding and breeding female Smooth snakes is not unexpected. The non-breeding snakes were probably still replenishing energy reserves which had been expended during reproduction. As a result, these specimens were lighter than the breeding females which had recuperated their energy reserves.
The mean difference in BWT between non-breeding and breeding condition snakes was equivalent to 50% of the mean non-breeding BWT. This figure is a reflection of the high cost of reproduction to the females. The difference in mean BWT between males and breed- ing females supports the suggestion by Gregory ( 1 977) and Shine ( I 978b) that the energy costs incurred by males as a consequence of spermatogenesis are less than those incurred through vitellogenesis and gestation by breeding females.
Growth As might be expected, the annual growth rates of Smooth snakes were highest for immature
specimens and lowest for mature specimens. This has also been noted in similar estimates for other species of snake (Platt, 1969; Clark, 1970; Bauerle, 1972) and illustrates the trend among reptiles for growth to decrease once sexual maturity has been attained (Bellairs, 1969).
Shine ( I 980) has suggested that a major reproductive cost among snakes may arise from the reallocation of energy reserves away from bodily growth to reproduction. If this reap- portioning of energy does occur, it should be apparent in the pattern of growth shown by females in different reproductive conditions. In contrast to the biennial reproductive cycle of the adder (Prestt, 197 l), the Smooth snake in Britain has an irregular reproductive cycle (Goddard & Spellerberg, 1980). Specimens may breed once every year, once every two years, or once every three years. Some evidence that growth is depressed during reproduction is provided by the results of the present study. The annual SVL growth rates of breeding con- dition females were similar to those shown by males. However, the seasonal growth rates of females, in this study snakes which exhibited a biennial or triennial reproductive cycle, were higher than those shown by males. Thus the sexual dimorphism in SVL is a result of an increase in female growth during the resting phases of their reproductive cycles. This suggests that females may be able to offset the cost of reproduction through an allocation

254 P. GODDARD
of energy reserves to bodily growth in non-breeding years. Thus some individuals may “choose” to grow rather than breed in any particular year, because of the influence that body size has on fitness. This reallocation of energy may be as important as the availability of energy (Aldridge, 1979) in controlling the timing of reproductive cycles in temperate zone snakes.
Food habits The prominence of small mammal prey, particularly nestling individuals, was unexpected.
The Smooth snake, although known to consume rodents and shrews (Steward, 1971), is generally considered to be a specialist predator on reptiles (Smith, 1973; Prestt, Cooke et al., 1974; Arnold & Burton, 1978). Contrary to this view, the results above indicate that the snakes were feeding both above and below ground, adopting a foraging mode (Huey & Pianka, 1981) which changed from that of a predator feeding on sedentary, clumped prey (nestling mammals) to that of one feeding on mobile prey (lizards and shrews). In the absence of information relating to prey selection, the simplest interpretation of these data is that the Smooth snake is an opportunist predator, feeding without preference on both types of prey according to their availability.
The evidence that has been used in support of a reptile/prey dependence for this species in Britain originates from three main sources: (i) information from continental workers (for example, Duguy, 196 I ; Bruno, 1966; Andren & Nilson, 1976, 1979); (ii) observations made on captive specimens (for example, Pitmen, 1940; Broadley, 1958; Appleby, I97 I ; Spellerberg, 1977b); and (iii) the pooling of miscellaneous field observations on snakes cap- tured in a variety of localities in Britain (Corbett in Nature Conservancy Council, 1983). Although these sources provide interesting information concerning the spectrum of prey types eaten, they can easily lead to an unrealistic view of diet due to local differences in snake food habits (Kephart, 1982) and variations in annual prey abundance (Kephart & Arnold, 1982). In view of these findings, the present author questions earlier reports of a widespread specialization in diet by this species in Britain, albeit biased towards small mammal prey (Spellerberg & Phelps, 1977) or reptile prey (Beebee, 1978).
Population characteristics The density estimates for Smooth snakes on sites 1 and 2 are much lower than that for
site 3. This reflects the lack of information concerning the movements of snakes on this latter site. Thus the size of the area calculated to be occupied by the snakes is too small. The result is an unrealistically high density. On sites 1 and 2, the movements of individual specimens were investigated (Goddard, 198 1 b) and, therefore, the extent of the population areas are more precisely known. It is interesting to note that the two densities on sites 1 and 2 (approxi- mately 1 and 2 snakedha, respectively) are much lower than the 1 1-1 7 snakedha previously reported for this species (Spellerberg & Phelps, 1977): It would appear that these higher densities refer to local concentrations of snakes rather than site populations, since individuals which exhibited more wide ranging movements have been excluded.
Why site 2 should support twice as many snakes per unit area than site 1 is unclear. On the former site, snakes tended to be captured on or around a south-facing embankment, resulting in a more clumped distribution of specimens than that found on site 1. Many of the snakes were gravid females and it appeared that they were seeking warm areas in which

THE SMOOTH SNAKE IN SOUTHERN BRITAIN 255
to bask and thereby facilitate the incubation of their ova. The male snakes were presumably attracted to the embankment by the presence of the females. No equivalent topographic feature existed on site 1 , and it is therefore possible that the difference in density between the two study sites simply reflected the influence of the embankment on local distribution. Further field observations are required to confirm this.
The 1 : 1 ratio and similar age structures for male and female Smooth snakes found in the present study suggest that the even ratio at birth (Goddard & Spellerberg, 1980) is per- petuated throughout life. Both sexes would therefore appear to exhibit similar mortality, though the causes of death are difficult to discern. An indication that mortality is higher among juvenile than adult Smooth snakes is provided by the observation that few one-year- old and two-year-old snakes were captured. However, this observation must be treated with caution since such individuals may not necessarily be captured in proportion to their true abundance. Their small size and cryptic colouration makes recognition in the field difficult, hence they may be easily overlooked.
Under the terms of the Wildlife and Countryside Act 198 1, there is a requirement for the periodic review of the endangered status of protected species. Unfortunately, the objective assessment of snake population size is fraught with difficulties (see Turner (1977) for a review). Consequently, estimates of the population size of Smooth snakes in Britain have been based on anecdotal evidence. In 1974, Prestt, Cooke et al. suggested that there was a declining population of between 1000 and 3000 snakes in Britain. In 1983, the British Herpetological Society (in NCC, 1983) reported that the current total population size was 2000 snakes. The interpretation of this information is problematical. For example, these figures could suggest that since 1974 the population of Smooth snakes has further declined, remained stable, or increased. At best, these estimates simply suggest a small population size.
An alternative to this particular approach to population estimation is to calculate an approximate value using the following information, (i) the density of Smooth snakes found on sites 1 and 2, and (ii) the areal extent of heathland habitat in central southern England (26,656 ha) as calculated from figures pertaining to unburnt heath in 1976 (taken from Bibby (1976)). This yields an estimated 26,656-53,312 Smooth snakes in Britain. However, it is unlikely that all of the available habitat in central southern England is occupied by Smooth snakes. For example, areas that have been recently burnt are less likely to support Smooth snakes than areas that have been left undisturbed. Nevertheless, as information becomes available on the extent of habitat likely to be occupied by Smooth snakes, the figures yielded by this method of estimation may be adjusted accordingly.
Summary
The morphology, growth, food habits and population characteristics of the Smooth snake Coronella austriaca Laurenti were studied in southern England. On capture, each snake was identified, sexed, measured, weighed and examined for the presence of food in its stomach and hind gut.
In total, measurements of 22 sexually immature and 92 sexually mature snakes were collected. There was little evidence for sexual dimorphism among immature snakes, though males had relatively longer tails than females. Among mature snakes, females had longer snout-vent lengths and shorter tails than males. The body weights of breeding females were heavier than those for both non-breeding females and males. Regardless of age or sex, body

256 P. GODDARD
weight was positively correlated with snout-vent length. An analysis of 68 growth measure- ments revealed that, in common with other species of snake, immature specimens had higher growth rates than mature specimens. Growth within one year did not differ appreci- ably between males and females. Conversely, growth over more than one year was greatest for female snakes. These differences in morphology and growth were ascribed to the influence that body size has on reproductive success. Large females (i) produce large offspring, and (ii) may have an increased chance of courtship and mating. During non-breeding years it is suggested that females offset the cost of reproduction through an allocation of energy reserves to bodily growth. Thus individuals may “choose” to grow rather than to breed in particular years. Hence, females in the resting phase of their reproductive cycles may exhibit high growth rates.
The Smooth snakes preyed on lizards and small mammals. Nestling prey (young rodents and young shrews taken from underground nests) were the most important dietary items. It was suggested that the Smooth snake is less specialized in its choice of prey than has been previously reported. It was considered to be an opportunist predator, selecting prey according to its availability.
Population size was estimated by direct enumeration, and densities of about one and two snakes per hectare were considered representative for this species in Britain. Attention was drawn to the influence that site factors and snake behaviour may have on these calculations. Few immature snakes were captured. It was unclear whether this reflected a higher mortality among these specimens, or whether it was more difficult to find them. The even sex ratio and stable age structure for males and females suggested that mortality was not dependent on the sex of the snakes. Adequate data are lacking for a more comprehensive examination of mortality to be made.
An alternative method to the anecdotal estimation of the total population size of Smooth snakes in Britain has been suggested.
I wish to record my thanks to Dr I. F. Spellerberg for the advice and support he gave throughout the course of this study. I would also like to thank the Forestry Commission and the Nature Conservancy Council for their permission to undertake this work. Financial support was provided by a University of Southampton Post-graduate Scholarship. Miss E. C. Meredith and two anonymous referees provided useful comments on parts of this manuscript.
REFERENCES Aldridge, R. D. (1979). Female reproductive cycles of the snakes Arizona elegans and Crotalus viridis. Herpetologica
Andren, C. & Nilson, G. (1976). Hasselsnoken (Coronella austriaca)-en utrotningshotad ormart! Fauna Flora.
Andren, C. & Nilson, G. (1979). Hasselsnoken (Coronella austriaca) i Norden-n isolerad och ekologiskt sarstalld
Appleby, L. G. (I97 I ) . British snakes. London: John Baker. Arnold, E. N. & Burton, J. A. (1978). Afield guide to the reptiles and amphibians of Briluin and Europe. London:
Collins. Bauerle, B. A. (1972). Biological productivity ofsnakes qf the Pawnee sites. (US. I.B.P. Grassland Biome Technical
report No. 207) Colorado State University. Beebee,.T. J. C. (1978). An attempt to explain the distribution of the rare herptiles Bulo calamia. Lacerta agilis and
Coronella auslriaca in Britain. Br. J. Herpet. 5 : 763-770. Bellairs, A. d’A. (1969). The Ige qfrepfiles. (2 Vols). London: Wiedenfeld & Nicolson. Bibby, C. J. (1976). Legacy o f a British summer. New Scient. 7 2 : 8-9.
35: 256-261.
Stockh. 71: 61-76.
ras? Fauna Flora, Stockh. 74: 89-96.

THE SMOOTH SNAKE IN SOUTHERN BRITAIN 257
Broadley, D. G. (1958). Some ecological notes on the British reptiles with particular reference to their feeding habits.
Brodin, T. (1951). Hasselsnoken. In Natur i Va.srer,yotlund: 376422. Swanberg. P. 0. & Curry-Lindahl. K. (Eds).
Bruno, S. (1966). Sulk specie del genere Coroncllu Laurenti viventi in Italia. .-Itti Accad. giocnia. Sc,i. nut. (6) 18:
Clark, D. R. (1970). Ecological study of the worm snake Carphophis w m i s (Kennicott). Univ. Kans. Puhh. Mus.
Duguy. R. (I96 I ) . Le cycle annuel d'activite de Corotic~lka airstriuca Laur.; d'apres les observations manuscrites
Fitch, H. S. (1970). Reproductive cycles of'lizards and snakes. Utiiv. K m s . : V r c . c . nut. Hist. Miic. Puh1.s. 52: 1-247. Goddard, P. (I98 la). Ecology ($the .smooth snak.c. Coronetla austriaca Lairrrnti in Britain. Ph.D. thesis. Unpub-
Goddard, P. (198 16). Limited movement areas and spatial behaviour in the smooth snake ('oronella airstriaca in
Goddard, P. & Spellerberg. I. F. (1980). Reproduction as a factor in the conservation of Coronrh aii.striacn in
Gregory, P. T. (1977). Life-history parameters of the red-sided garter snake (Thamnop/ii.s sirtalis parieralis) in an
Hawley. A. W. L. & Aleksiuk. M. (1976). Sexual receptivity in the female red-sided garter snake (Thumnophis
Honeggar, R. ( 1 978). Threatened amphibians and reptiles in Europe. Strasbourg Council for Europe. Huey, R. B. & Pianka, E. R. (198 I ) . Ecological consequences of foraging mode. Ecology 62: 99 1-999. Kephart, D. G. ( 1 982). Microgeographic variation in the diets ofgarter snakes. Oecologia 52: 287-29 I . Kephart, D. G. & Arnold, S. J. ( I 982). Garter snake diets in a fluctuating environment: A seven year study. Ecology
Meddis, R. ( 1 975). A statistical handbook /hr noti-.\tutistrcian.s. Maidenhead: McGraw-Hill. Nature Conservancy Council (1983). The ecolqyj, trnd conservation ol'amphihian and reptile species endangered
Pitmen, R. (1940). The smooth snake (Coronella Iucvis) in Wilts. Wilts. archucol. nut. Hi.\t. Mag. 49: 238-239. Platt, D. R. (1969). Natural history of the hognose snakes Hctcrodon platyrhinos and Heterodon nasicirs. Uriiv.
Prestt, 1. (1971). An ecological study ofthe viper I'iperu hcrus in southern Britain. J . Zool.. Lorid. 164: 373418. Prestt, I., Cooke, A. S. & Corbett. K. F. (1974). British amphibians and reptiles. In Thc c,hangin,g flora andfuunu
Saint Girons, H. & Naulleau, G. ( I 98 I ). Poids des noveau-nb et strategies reproductrices des Viperes europeennes
Shine, R. (1978~) . Sexual size dimorphism and male combat in snakes. Oecologia 33: 269-278. Shine, R. (19786). Growth rates and sexual maturation in six species of Australian elapid snakes. Herpetologica
Shine, R. (1980). "Costs" of reproduction in reptiles. Oecologia 46: 92-100. Smith, M. (1973). The British amphibians and reptiles (5th ed.). Revised by Bellairs, A. d'A. & Frazer, J. F. D.
Sokal, R. R. & Rohlf, F. J . (1969). Biometry. San Francisco: W. H. Freeman. Spellerberg, 1. F. (1977~) . Marking live snakes for identification of' individuals in population studies. J . appl . Ecol.
Spellerberg, I . F. (19776). Behaviour of a young smooth snake. Coronrlla uirstriaca Laurenti. Biol. J . Linn. Soc.
Spellerberg, I. F. & Phelps, T. E. (1977). Biology, general ecology and behaviour of the snake Curonella uirstriaca.
Steward, J . W. (1971). The .snu!ies o/Eitrope. Newton Abbot: David &Charles. Turner, F. B. (1977). The dynamics of populations of Squamates, Crocodilians and Rhynchocephalians. In Biolo,y,~*
Br. J . Herpet. 2: 126-129.
Goteberg.
99-1 17.
nut. Hist. 19: 85-194.
inedites de Raymond Rollinat. Terre C'ir 15: 401435 .
lished. Southampton University.
southern England. In Proc. Euro. Herp. Svmp C'. W. L. P. Oxford 1980: 2 5 4 0 . Coburn, J. (Ed.).
southern England. Bid/ . Eid. 11: 535-541.
extreme environment, the interlake region of Manitoba. Pitbls Zool. natn. Mus. nut. Sci. Can. No. 13: 1 4 4 .
sirtalis pariefalis). Copeia 1976: 40 1 4 0 4 .
63: 1232-1236.
in Britain. London: Nature Conservancy Council.
Kans. Pirbls Miis. nut. Hist. 18: 253420.
ofBritain: 229-254. Hawkesworth, D. L. (Ed.). London: Academic Press.
Terre Vie 35: 597-6 16.
34: 73-79.
London: Collins.
14: 137-138.
9: 323-330.
Bid . J . Linn. Soc. 9: 133-164.
ol'the Rcyiilia 7: 157-264. Cans, C. & Tinkle. D. W. (Eds). London: Academic Press.





























