Embed Size (px)
Citation preview

MORPHOLOGY OF MANDIBLE AND LATERALIA IN SIX ENDEMICAMPHIPODS (AMPHIPODA, GAMMARIDEA) FROM LAKE BAIKAL,
IN RELATION TO FEEDING
BY
IRINA V. MEKHANIKOVA1)
Limnological Institute SB RAS, Laboratory of Aquatic Invertebrate Biology, Office 227, 3,Ulan-Batorskaya St., Irkutsk 664033, Russia
ABSTRACT
The morphology of the mandible and stomach lateralia in some Lake Baikal amphipods has beeninvestigated in regard to their food preference. Six species of endemic amphipods with different lifestyles (so-called “life forms”) have been studied by means of light and scanning electron microscopy.The modifications of the mandible are attributed to the food resources used. The position of theincisor and lacinia mobilis, their width and length, the structure of the triturative area of the molar,the wearing of incisor and lacinia mobilis between two moults, and the relative length of the stomachdepend on the type of food. None of the species studied revealed complete reduction of any part ofthe mandible. The most specialized species are the oophage Pachyschesis bazikalovae G. Karaman,1976, the necrophage Ommatogammarus albinus (Dybowsky, 1874), and a species biting holes insponges with a siliceous skeleton, Eulimnogammarus violaceus (Dybowsky, 1874). The morphologyof the lateralia seems not to be connected with the food type, but is probably important from aphylogenetic point of view.
RÉSUMÉ
La morphologie de la mandibule et des pièces latérales de l’estomac de certains amphipodes dulac Baïkal a été étudiée en relation avec leurs préférences alimentaires. Six espèces d’amphipodesendémiques ayant des modes de vie différents (désignés par le terme “life forms”) ont été examinéesau microscope électronique à balayage et au microscope optique. On a montré que les modificationsde la mandibule sont déterminées par le type d’aliments utilisés. La position du processus incisif etde la lacinia mobilis, la largeur et la longueur de ces pièces, la structure de la surface broyeuse duprocessus molaire, l’usure subie par le processus incisif et la lacinia mobilis pendant l’intermue, ainsique la longueur relative de l’estomac dépendent du type de nourriture. Parmi les espèces examinées,aucune n’a subi la réduction totale d’une partie ou d’une autre de la mandibule. Les espècesles plus spécialisées sont l’oophage Pachyschesis bazikalovae G. Karaman, 1976, le nécrophageOmmatogammarus albinus (Dybowsky, 1874) et une espèce inquiline Eulimnogammarus violaceus(Dybowsky, 1874) qui vit dans des cavités qu’elle creuse dans des éponges à squelette siliceux. La
1) e-mail: [email protected]
© Koninklijke Brill NV, Leiden, 2010 Crustaceana 83 (7): 865-887Also available online: www.brill.nl/cr DOI:10.1163/001121610X504289

866 IRINA V. MEKHANIKOVA
morphologie des pièces latérales de l’estomac ne semble pas liée au type de nourriture, mais estprobablement importante du point de vue phylogénétique.
INTRODUCTION
The morphology of the mouthparts of amphipods is closely connected with theirfeeding habits (Sainte-Marie, 1984; Moore & Rainbow, 1989; Coleman, 1991a,b; Steele & Steele, 1993; Watling, 1993). The structure of the digestive tract,in contrast, is more conservative (Johnston et al., 2004). However, the size andstructure of the stomach in some groups of amphipods (fig. 1A) reflect their feedingstrategy (Agrawal, 1964; Coleman, 1992).
In most amphipods, all parts of the mandible are well developed (fig. 1B).This type of mandible is called a basic gammaridean type (Barnard, 1969).A high specialization of the mouth apparatus depending on a particular type offood consumed is observed in some families of Amphipoda: either reduction, orovergrowth of some parts, or even atrophication of the mouth apparatus as a whole(De Broyer & Thurston, 1987; Watling, 1993; Bellan-Santini, 1999). The majorityof the mandible modifications found are attributed to predation or to carrionfeeding (Sainte-Marie, 1984; Coleman, 1989; Steele & Steele, 1993; Dauby et al.,2001; Legezinska, 2008). However, there are aberrations from the basic scheme ofthe structure in herbivorous amphipods (Agrawal, 1964). Subtle differences in thestructure of some parts of the mandible are also attributed to feeding habits. Forexample, the reduction of molar size and increase of densely set short spinules on
Fig. 1. Schematic structure of: A, the stomach; and, B, the mandible of the basic type of amphipod.

MORPHOLOGY AND FEEDING IN BAIKAL GAMMARIDEA 867
the triturative area of Talorchestia species II (family Talitridae) are adaptations tothe grinding of siliceous diatom valves (Johnston et al., 2004).
The amphipod fauna of Lake Baikal is characterized by its high species di-versity: 348 endemic species and subspecies from 41 genera and 6 families(Takhteev, 2000). Amphipods are one of the most species-rich groups of inverte-brates, inhabiting all biotopes and depths of the lake, including the maximal depths.There are very few specialized forms in the Baikal amphipod fauna (Takhteev &Mekhanikova, 1993), and most species are omnivorous (Bazikalova, 1954; Bek-man, 1962; Morino et al., 2000; Mekhanikova, 2001, 2006). However, an analysisof the ratio of carbon isotopes 13C/12C of twenty benthic species of amphipodsshowed that the isotopic composition of food consumed by those varies withinwide ranges, which indicates their different feeding strategies (Melnik et al., 1996).
The structure of Baikal amphipod organs responsible for the capture andprocessing of food in relation to their food preferences, has not been studied pre-viously. Six species of endemic amphipods of different life forms2) were selectedfor this work (fig. 2). Brandtia parasitica (Dybowsky, 1874) lives exclusively onBaikal sponges of the family Lubomirskiidae. For a long time it has been consid-ered a parasitic species, feeding on sponge tissue (Lomonosov & Chekanovsky,1869; Kamaltynov, 1999). However, recently it has been established that this isan omnivorous species (Mekhanikova, 2001). Another symbiont, Eulimnogam-marus violaceus (Dybowsky, 1874) inhabits sponges by carving out holes in theirbodies. Morino et al. (2000) believe that it feeds upon sponge tissue. Brandtialatissima lata (Dybowsky, 1874), a typical inhabitant of the stony littoral zone,is a polyphage (Mekhanikova, 2006). Acanthogammarus grewingkii (Dybowsky,1874), one of the most common species of the deep zone of Lake Baikal, isalso a polyphage; age dependent changes in its feeding were studied in detailby Bazikalova (1954). Pachyschesis bazikalovae G. Karaman, 1976 inhabits thegill cavity of a large nectobenthic amphipod, Garjajewia cabanisii (Dybowsky,1874) and feeds upon its eggs (Takhteev & Mekhanikova, 1993). Ommatogam-marus albinus (Dybowsky, 1874) feeds on carrion and is usually caught with abaited trap containing rotting fish or meat (Dybowsky, 1874; Bazikalova, 1945;Takhteev, 2000).
The morphology of the mandible corpus and some detailed structures of thelateralia of the stomach have now been studied for the first time in Baikal
2) The term “life forms” has been coined by Takhteev (2000). It encompasses life history, habitat, andmorphological adaptations. Its classification is rather complicated, but it gives a more complete viewon a particular species. E.g., “lithophilic, armoured, walking” or “lithophilic, smooth, swimming”,etc. This system of life forms had previously been elaborated for marine amphipods (Kamenskaya,1984; Vinogradov, 1988, 1990).

868 IRINA V. MEKHANIKOVA
Fig. 2. Schematic biotope distribution of amphipods of various life forms. 1, Brandtia latissima lata(Dybowsky, 1874); 2, Brandtia parasitica (Dybowsky, 1874); 3, Acanthogammarus grewingkii (Dy-bowsky, 1874); 4, Eulimnogammarus violaceus (Dybowsky, 1874); 5, Ommatogammarus albinus(Dybowsky, 1874); 6, Pachyschesis bazikalovae G. Karaman, 1976. [1-3, 5, according to Bazikalova,
1945; 4, according to Kamaltynov, 1993; 6, according to Takhteev, 2000; with modifications.]
amphipods of different life forms. The relationship between their feeding habitsand the morphology of the organs responsible for the capture and processing of thefood is discussed in this paper.
MATERIAL AND METHODS
Amphipods were sampled with dredges, trawls, baited traps, nets, and by diversin different areas of Lake Baikal during the years 1988-2009. The amphipods werefixed with 4% formalin and stored in 75% ethanol. Analyses of gut contents andfood preferences were performed on 392 specimens. Between 10 and 16 specimens

MORPHOLOGY AND FEEDING IN BAIKAL GAMMARIDEA 869
of each species were dissected to determine the length of the stomach, while 15 to30 (or more) specimens were used for studies of the morphology of the mandiblesand lateralia.
Amphipods were dissected with microscissors under a stereoscopic microscopeMBS-10, and sex and body length were also determined. Temporary preparationsof food components were prepared in water or a mixture of water and glycerin,and examined under an Ergaval compound microscope. Permanent preparationsof mandibles and lateralia were prepared in Fora-Berlese liquid and examinedunder compound light microscopes Axiostar Plus and Axiovert 2000. For scanningelectron microscopy, the mandibles were dehydrated in an ethanol series, dried,glued onto aluminum stubs with carbonic adhesive tape, coated with gold invacuum (Balzers SCD 004), and examined under a scanning electron microscopePhilips SEM 525M.
RESULTS
Brandtia (Brandtia) latissima lata (Dybowsky, 1874)(fig. 3A-F)
All parts of the mandible are well developed. The incisor and lacinia mobilisare directed backwards. The left mandible incisor bears 5 teeth, and the laciniamobilis is slightly smaller with 4 teeth (fig. 3A). The right mandible incisor bears5 teeth, and the lacinia mobilis is in the shape of a short plate with teeth of differentlength (fig. 3B). Both mandibles are topped by a “crown”3) on the toothed edges.The teeth of the incisor and lacinia mobilis in some specimens that were ready formoulting, are slightly truncated, in the others they are extremely dulled (fig. 3E).Different wearing levels of the teeth of the mandible in different individuals of thespecies are likely to be attributed to different food types consumed. The molar ismassive, with rows of teeth (fig. 3C) and a large triturative area.
The stomach length makes up 19.3 ± 0.5% of the body length. The lateralia arewith long, thin spines, without secondary spinules on the surface (fig. 3D).
The gut content of B. latissima lata is diverse and consists of both vegetableand animal matter. The main vegetable components are lower plants and macro-phytes, sometimes pine spores. Among lower plants there are various benthic (fig.3F) and planktonic diatoms, and green and blue-green filamentous algae. Diatomswere found in the form of fragments, valves, and even of intact colonies (e.g.,
3) “Crown” — the teeth of the incisor and lacinia mobilis are of yellow colour in omnivorous and inspongicolous species, and together look like a golden crown on the tooth.
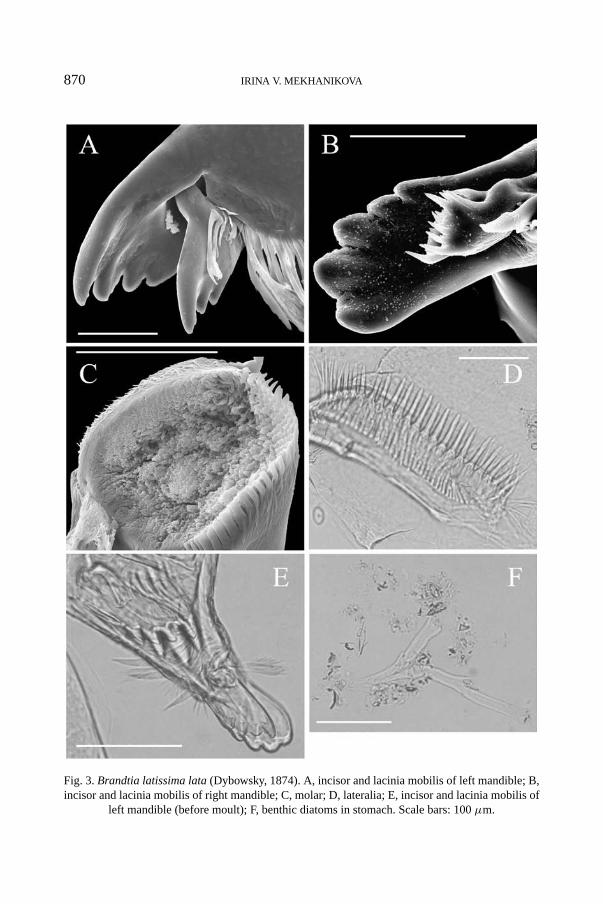
870 IRINA V. MEKHANIKOVA
Fig. 3. Brandtia latissima lata (Dybowsky, 1874). A, incisor and lacinia mobilis of left mandible; B,incisor and lacinia mobilis of right mandible; C, molar; D, lateralia; E, incisor and lacinia mobilis of
left mandible (before moult); F, benthic diatoms in stomach. Scale bars: 100 μm.

MORPHOLOGY AND FEEDING IN BAIKAL GAMMARIDEA 871
Gomphonema). The food of animal origin was represented by oligochaetes, ro-tifers, entomostracans (harpacticoids, ostracods, and cyclopoids) and malacostra-cans (amphipods). Entomostracans were found intact and in fragments, whereasthe amphipods occurred in fragments only (cuticle fragments, extremities, anten-nae, and mouthparts). Detritus, mineral particles, and sponge spicules were com-mon components of the gut contents (Mekhanikova, 2006).
Brandtia (Spinacanthus) parasitica (Dybowsky, 1874)(fig. 4A-F)
All parts of the mandible are well developed. The incisors and lacinia mobilisare directed backwards. The left incisor bears 6 teeth. The lacinia mobilis with 4teeth is slightly shorter than the incisor (fig. 4A). The right incisor is provided with4 teeth. Sometimes there is an additional fifth, short tooth. The second tooth is thelongest. The lacinia mobilis is in the shape of a short plate with teeth (fig. 4B).Both mandibles are topped by a “crown”. The molar is massive, with rows of teeth(fig. 4C, D) and a large grinding area.
The stomach length amounts to 16.7 ± 0.2% of the body length. The lateraliabear short, thick spines, wide at the base, with fine spinules in the distal and middleparts (fig. 4E).
B. parasitica feeds upon vegetable and animal organisms, dwelling on spongesand settling from the water column. Vegetable food components are representedby planktonic (fig. 4F) and benthic diatoms, chrysophycean and filamentous algae,macrophytes, unidentified vegetable remains, and spores of higher plants. Animalorganisms are represented by entomostracans and malacostracans, oligochaetes,rotifers, and ciliates. Sponge spicules and their fragments get into the gut of B.parasitica together with the food consumed. Sponge tissues have not been foundin the gut of any of the specimens studied. A planktonic diatom, Aulacoseirabaicalensis (K. Meyer) Simonsen, is the major food component in B. parasiticain spring during its massive growth. Fragments of diatom valves are more oftenencountered in the food, although intact cells are also found, even in juveniles(Mekhanikova, 2001).
Acanthogammarus grewingkii (Dybowsky, 1874)(fig. 5A-G)
All mandible parts are well developed. The mandible corpus is thick andstrongly chitinized. The incisors and lacinia mobilis are directed backwards. Theleft incisor bears 5 teeth and sometimes a very short additional sixth tooth. Thelacinia mobilis bears 4 teeth, the first one being the longest (fig. 5A, C). The right

872 IRINA V. MEKHANIKOVA
Fig. 4. Brandtia parasitica (Dybowsky, 1874). A, incisor and lacinia mobilis of left mandible; B,incisor and lacinia mobilis of right mandible; C and D, molar; E, lateralia; F, planktonic diatoms in
stomach. Scale bars: 100 μm in A-C, E, and F; 10 μm in D.

MORPHOLOGY AND FEEDING IN BAIKAL GAMMARIDEA 873
Fig. 5. Acanthogammarus grewingkii (Dybowsky, 1874). A and C, incisor and lacinia mobilis of leftmandible; B and D, incisor and lacinia mobilis of right mandible; E, molar (a part only) with diatomfragments; F, lateralia; G, planktonic diatoms in stomach. Scale bars: 1 mm in A and B; 100 μm in
C, D, and G; 10 μm in E; 50 μm in F.
incisor is equipped with 5 teeth, the second being the longest. The lacinia mobilisis bifurcated, with 2 teeth (fig. 5B, D). Some specimens have 1-2 short additionalteeth at the base of the short tooth. Both mandibles are topped by a “crown”. Theteeth of the incisor and lacinia mobilis are worn during the inter-moult period. Themolar is massive, with rows of teeth.

874 IRINA V. MEKHANIKOVA
The stomach length amounts to 11.5 ± 0.3% of the body length. The lateraliahave sharp, smooth spines, without secondary spinules. The spine width at the baseis two times the width in the distal part (fig. 5F).
A. grewingkii consumes different types of food, from detritus to invertebratesand, if possible, carrion. Its main food components are amphipods, entomostra-cans, and detritus (Bazikalova, 1954). Diatoms play an important role in feeding,especially in years of their mass development. In June, when they settle in abun-dance onto the bottom, this omnivorous species feeds almost exclusively on di-atoms (fig. 5G). At this time of the year, the alimentary canal of A. grewingkii isfilled with a green-gray substance: semi-digested diatom cells with a large num-ber of intact valves and their fragments. The food of A. grewingkii consists of thefollowing dominant species: Aulacoseira baicalensis (in volume), Stephanodiscusmeyerii Genkal & Popovskaya (in valve number), Cyclotella, and Aulacoseira is-landica (O. Müller & Simonsen). Acanthogammarus grewingkii breaks chains ofplanktonic diatoms and swallows them without grinding (fig. 5G), but a great por-tion of these diatoms are ground with molar processes as well, which causes wear-ing of rows of teeth on their surface (fig. 5E). The chains of A. baicalensis consistof 2-3 cells (4-6 valves), more rarely of 4 cells (8 valves). Besides diatoms, weencountered sponge spicules, setae of crustaceans, fragments of amphipods, andentire entomostracan crustaceans in the gut as well.
Eulimnogammarus (Eurybiogammarus) violaceus (Dybowsky, 1874)(fig. 6A-G)
The mandible corpus is massive, the incisor and lacinia mobilis are stronglychitinized. The teeth are worn for the period between two moults (fig. 6C, D).In the samples of this species it has been impossible to find two specimens withsimilar mandibles. The incisor and lacinia mobilis are directed downwards andslightly forwards. The left incisor in a specimen after moulting bears 7-8 sharpteeth, the second being the largest. The lacinia mobilis bears 7-8 teeth, the firstbeing the longest (fig. 6A). The left incisor looks like a spatula in specimens duringthe intermoult, sometimes a terminal tooth remains. The incisors have numerouscracks and fissures on their surface. At the beginning of moulting, the laciniamobilis is also provided with rather truncated teeth and numerous cracks. Theright incisor bears smooth teeth. The lacinia mobilis has a long, hook-like, sharptooth, and one tooth is in the shape of a plate with several teeth (fig. 6B). Theright lacinia mobilis is worn during the intermoult and looks like a spatula (“fishtail”) (fig. 6D). Incisors of both mandibles are topped with a massive “crown” (notonly on the toothed edges). The molar is provided with ridges and cone-shapedelevations (fig. 6E, F).

MORPHOLOGY AND FEEDING IN BAIKAL GAMMARIDEA 875
Fig. 6. Eulimnogammarus violaceus (Dybowsky, 1874). A, incisor and lacinia mobilis of leftmandible (after moult); B, incisor and lacinia mobilis of right mandible (after moult); C, incisorand lacinia mobilis of left mandible (before moult); D, incisor and lacinia mobilis of right mandible(before moult); E, molar (after moult); F, molar (before moult); G, lateralia. Scale bars: 100 μm in
A-E, and G; 10 μm in F.

876 IRINA V. MEKHANIKOVA
The stomach length amounts to 17.2 ± 0.7% of the body length. The very longand thin spines of the lateralia are hooked at the edges. Secondary spinules arelocated along the entire length of the spines (fig. 6G).
The guts of only two out of 34 dissected animals were filled with spongetissues. The gut of 11 specimens contained fragments of sponges, and the gutsin 21 specimens contained no sponge tissues at all. Besides sponge tissues, somediatoms were found in the gut (Navicula, Ceratoneis, Cocconeis, Gomphonema,and Aulacoseira baicalensis: intact cells and valve fragments), brown or green,uniform, unidentified substance, a rotifer, oligochaete setae, and a fragmentresembling an oligochaete.
Ommatogammarus albinus (Dybowsky, 1874)(fig. 7A-F)
The mandibles are highly specialized, with a wide incisor and a lacinia mobiliswith almost the same width. The incisor and lacinia mobilis are directed down-wards and slightly backwards. The left incisor is composed of 7 teeth located inone plane, and looks like a wide ridge (fig. 7A, C). The lacinia mobilis is also flat,with 6 teeth of almost similar length (fig. 7A, C). The right incisor bears 5-6 teethlocated in the same plane, the second tooth being longer than the others (fig. 7B,D). The lacinia mobilis is falciform. Short and sharp teeth are located between ex-tremely long teeth (fig. 7B, D). Wearing of the teeth of incisor and lacinia mobilishas not been observed for the intermoult period. The teeth of the mandible arewithout a “crown”. The molar is small, weak, and with a row of ridges (fig. 7E).
The stomach is long: 17.2 ± 0.3% of the body length. The spines of the lateraliaare smooth, sharp, and straight, without spinules on the surface (fig. 7F).
Fat droplets (pale yellow, or sometimes orange) are always present in the gutof O. albinus, as well as a whitish, structureless substance (muscles of fishesand mammals). Sometimes the gut content is dark in colour, or it contains darkerinclusions (it is obvious that in this case fish internal organs were consumed). Onlyin three specimens out of 26 the gut contents resembled detritus.
Pachischesis bazikalovae G. Karaman, 1976(fig. 8A-G)
The mouthparts of P. bazikalovae are specialized for feeding on the host’s eggs.The incisor and lacinia mobilis are adapted for piercing eggs (fig. 8A, B), and aredirected vertically downwards. The left incisor is provided with 5 teeth, each ofwhich is located much lower than the previous one, the lacinia mobilis bears 3-4teeth, the location of which is the same (fig. 8A). The right incisor bears 4 teeth

MORPHOLOGY AND FEEDING IN BAIKAL GAMMARIDEA 877
Fig. 7. Ommatogammarus albinus (Dybowsky, 1874). A and C, incisor and lacinia mobilis of leftmandible; B and D, incisor and lacinia mobilis of right mandible; E, molar; F, lateralia. Scale bars:
100 μm in A-D, and F; 10 μm in E.
located as on the left incisor (fig. 8B, C). The lacinia mobilis is bifurcated with twovery long sharp teeth (fig. 8B, D). During the intermoult, the mandible teeth arenot worn, their cusps being slightly truncated (fig. 8A, B). The mandible teeth arenot “crowned”. The molar is reduced; pineal elevations are observed on its surface(fig. 8E, F).
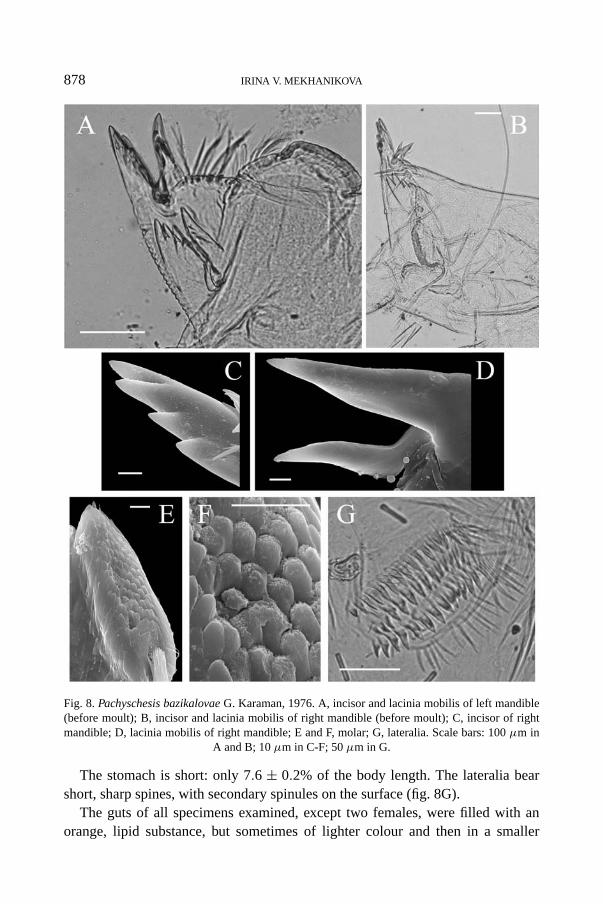
878 IRINA V. MEKHANIKOVA
Fig. 8. Pachyschesis bazikalovae G. Karaman, 1976. A, incisor and lacinia mobilis of left mandible(before moult); B, incisor and lacinia mobilis of right mandible (before moult); C, incisor of rightmandible; D, lacinia mobilis of right mandible; E and F, molar; G, lateralia. Scale bars: 100 μm in
A and B; 10 μm in C-F; 50 μm in G.
The stomach is short: only 7.6 ± 0.2% of the body length. The lateralia bearshort, sharp spines, with secondary spinules on the surface (fig. 8G).
The guts of all specimens examined, except two females, were filled with anorange, lipid substance, but sometimes of lighter colour and then in a smaller

MORPHOLOGY AND FEEDING IN BAIKAL GAMMARIDEA 879
amount. P. bazikalovae inhabits the gill cavity of Garjajewia cabanisii and feedsexclusively on its eggs. The reproduction period of amphipod host is extended,thus the parasites do not experience shortage of food. Moreover, they periodicallychange host in search of food and sex partners (Takhteev & Mekhanikova, 1993).
DISCUSSION
The mandible morphology of the six species of amphipods studied shows, thatthese species have different feeding strategies (table I), although none of themshowed complete reduction of any of the parts of the mandible. Mandibles in thethree omnivorous species (Acanthogammarus grewingkii, Brandtia latissima lata,and Brandtia parasitica) are of a basic type. The mandible is increasingly modifiedwhen a new feeding strategy is adopted or there is adaptation to any specializedhabitat (Watling, 1993). In Lake Baikal, alternative food sources are the eggs oflarge amphipods, and carcasses of vertebrates (fishes and seals), whereas spongeswith a siliceous skeleton constitute a specialized habitat. Our investigations showthat modification of the mandibles is indeed more expressed in the necrophageOmmatogammarus albinus and the parasitic Pachischesis bazikalovae, feedingon eggs of a large amphipod (table I). The wide incisor and lacinia mobilis ofboth mandibles of O. albinus function as a knife system, while the elongatedincisors and laciniae mobiles of P. bazikalovae are adapted to pierce egg shells.The right lacinia mobilis of P. bazikalovae is the most modified: it looks likea long, two-pronged fork. Describing Pachischesis, Takhteev (2000) noted that“The transition to oophagy did not cause any transformation of the mouthparts.”This viewpoint can hardly be supported. The mandibles of Eulimnogammarusviolaceus, a symbiont of Baikal sponges, must carve out holes, and thus experiencea massive friction and wear when cracking the siliceous spicules. Therefore,between two moults, these teeth are completely worn, and numerous fissures areleft on their surface. The mandibles of this species are adapted to crack thosesiliceous sponge skeletons, and they bear strong “crowns” on both the incisor andlacinia mobilis (unlike polyphagic species, that bear “crowns” only on the toothededges). Morino et al. (2000) note that the mandibular corpus in E. violaceus iswider compared to the palp size. The distal part of the incisor is strongly developedand sponge fragments are found in the stomachs. According to our data, only in6% of the specimens studied the guts were filled with sponge tissues, and in 32%these tissues were present in small amounts. The majority of the guts examinedcontained no sponge material. Therefore, it is still unclear whether those spongetissues are to be considered as food, or that those pieces were bitten out whileconstructing a shelter, and next swallowed accidentally.

880 IRINA V. MEKHANIKOVA
TA
BL
EI
Taxo
nom
icpo
sitio
n,ec
olog
ical
char
acte
rist
ics,
food
stra
tegi
es,a
ndst
ruct
ural
char
acte
rist
ics
ofm
andi
ble
and
stom
ach
ofsi
xsp
ecie
sof
Bai
kala
mph
ipod
s
Spec
ies
Hab
itat
Dep
thin
Lif
efo
rmB
ody
leng
th,
Feed
ing
type
Man
dibl
eSt
omac
hm
mm
Bra
ndtia
(Bra
ndtia)
Lak
eB
aika
l1-
65W
alki
ng,
Up
to18
.7Po
lyph
age
Non
-spe
cial
ized
;R
elat
ive
stom
ach
latiss
ima
lata
(usu
ally
atlit
hoph
ilous
,with
inci
sor
dire
cted
leng
th19
.3%
;(D
ybow
sky,
1874
)2-
40)
arm
edbo
dyba
ckw
ards
;la
tera
liasp
ines
long
(fam
ilym
olar
with
(∼56
μm
),th
in,
Aca
ntho
gam
mar
idae
)to
oth
row
sw
ithou
tsec
onda
rysp
ines
Bra
ndtia
Lak
eB
aika
l1-
60E
pibi
onto
f10
-11.
5Po
lyph
age
Non
-spe
cial
ized
;R
elat
ive
stom
ach
(Spi
naca
nthu
s)ex
cept
the
(sel
dom
atB
aika
lspo
nges
inci
sor
dire
cted
leng
th16
.7%
;pa
rasi
tica
Sele
nga
Riv
er20
0)ba
ckw
ards
;la
tera
liasp
ines
(Dyb
owsk
y,18
74)
shoa
lm
olar
with
shor
t(∼3
4μ
m),
(fam
ilyto
oth
row
sth
ick,
with
Aca
ntho
gam
mar
idae
)se
cond
ary
spin
es
Aca
ntho
gam
mar
usL
ake
Bai
kal
100-
1380
Nec
tobe
nthi
c,U
pto
90Po
lyph
age
Non
-spe
cial
ized
;R
elat
ive
stom
ach
grew
ingk
ii(u
sual
lyst
raig
htin
ciso
rdi
rect
edle
ngth
11.5
%;
(Dyb
owsk
y,18
74)
belo
wsw
imm
ers
back
war
ds;
late
ralia
spin
eslo
ng(f
amily
300-
400)
mol
arw
ith(∼
90μ
m),
thin
,A
cant
hoga
mm
arid
ae)
toot
hro
ws
with
outs
econ
dary
spin
es

MORPHOLOGY AND FEEDING IN BAIKAL GAMMARIDEA 881
TA
BL
EI
(Con
tinue
d)
Spec
ies
Hab
itat
Dep
thin
Lif
efo
rmB
ody
leng
th,
Feed
ing
type
Man
dibl
eSt
omac
hm
mm
Eul
imno
gam
mar
usL
ake
Bai
kal
1.5-
140
Sym
bion
tbiti
ng18
-30
Spon
giop
hage
?Sp
ecia
lized
for
bitin
gR
elat
ive
stom
ach
(Eur
ybio
gam
mar
us)
exce
ptth
e(u
sual
lyat
hole
sin
Bai
kal
outs
pong
etis
sues
;le
ngth
17.2
%;
viol
aceu
sSe
leng
aR
iver
10-3
0)sp
onge
sin
ciso
rdi
rect
edla
tera
liasp
ines
long
(Dyb
owsk
y,18
74)
shoa
lsl
ight
lyfo
rwar
d;(∼
128
μm
),th
in,
(fam
ilyG
amm
arid
ae)
mol
arw
ithro
ws
ofcu
rved
atth
een
ds,
ridg
esan
dw
ithse
cond
ary
cone
-sha
ped
spin
esel
evat
ions
Om
mat
ogam
mar
usO
pen
part
ofL
ake
47-1
641
Ben
thic
-pel
agic
,20
-25
Obl
igat
eSp
ecia
lized
;inc
isor
Rel
ativ
est
omac
hal
binu
sB
aika
l,se
ldom
(usu
ally
necr
opha
gene
crop
hage
dire
cted
slig
htly
leng
th17
.2%
;(D
ybow
sky,
1874
)Pr
oval
Bay
belo
wba
ckw
ards
;mol
arla
tera
liasp
ines
long
(fam
ilyG
amm
arid
ae)
200-
400)
with
row
sof
(∼20
0μ
m),
with
out
ridg
esse
cond
ary
spin
es
Pach
ysch
esis
Sout
hern
Bai
kal,
100-
1050
Para
sitic
onFe
mal
eO
opha
geSp
ecia
lized
;inc
isor
Rel
ativ
est
omac
hba
zika
lova
eM
aloy
eM
ore,
amph
ipod
sup
to13
.8,
dire
cted
vert
ical
lyle
ngth
7.6%
;G
.Kar
aman
,197
6B
argu
zin
Bay
,m
ales
dow
nwar
ds;m
olar
late
ralia
spin
es(f
amily
Chi
vyrk
uyB
ay,
upto
8.4
with
pine
alsh
ort(
∼25
μm
),Pa
chys
ches
iidae
)sl
ope
ofPo
sols
kan
del
evat
ions
thic
k,w
ithSo
snov
skB
anka
s,se
cond
ary
spin
essl
ope
ofSe
leng
aR
iver
delta
fron
t
Not
e:Ta
xono
my
and
char
acte
risa
tion
oflif
efo
rms
are
afte
rTak
htee
v(2
000)
;dis
trib
utio
n,ha
bita
tdep
th,a
ndbo
dyle
ngth
from
Baz
ikal
ova
(194
5),T
akht
eev
(200
0),a
ndK
amal
tyno
v(2
001)
.

882 IRINA V. MEKHANIKOVA
Differences among polyphages, oligophages, and spongicolous E. violaceushave also been recorded in the structure of the triturative area of the molar. On themolar surface of all polyphages there are rows of teeth, which serve as a rasp, andin oligophages and E. violaceus there are rows of ridges and/or pineal elevations(table I).
The modifications of the mandible are revealed not only by the reduction or,in contrast, by the extreme development of its separate parts, but also by theposition of the mandible in the head (Watling, 1993). The incisors of Baikalomnivorous amphipods are directed backwards, in the oophage P. bazikalovaevertically downwards, in the necrophage O. albinus slightly backwards, and inthe spongicolous E. violaceus slightly forward.
Variations in stomach structure depending on the type of food have beendescribed for many marine species (Coleman, 1994). The lateralia of the Baikalspecies E. violaceus are equipped with long and very thin, serrated spines, whichare hooked on the distal edges. In the other species studied, the lateralia spinesare not hooked on the distal edges; in some they are smooth (B. latissima lata, A.grewingkii, O. albinus), and in others they have secondary spinules (B. parasitica,P. bazikalovae); in the latter species the lateralia bear triangular spines arranged inthree rows, with remarkably sharp edges. Variations in the length of the spines ofthe lateralia were also recorded (table I).
It has previously been demonstrated in marine species, that the highest diversityof stomach size is characteristic of animals with a specialized type of feeding. Forexample, in the necrophage Waldekia obesa (Chevreux, 1905) (family Lysianas-sidae) the stomach length makes up about 34% of the body length, in Andanioteslinearis K. H. Barnard, 1932 (family Stegocephalidae), feeding on cnidarians, thelength of the stomach is over 45% of the total body length. The stretching of thestomach in the first species is due to its adaptation to long intervals between foodconsumption, and in the other, due to consuming low-calory food. The stomach inmarine detritophages is short, less than 10%, in carnivores it varies from 2-5 to 16-17% of the total body length (Coleman, 1991a, 1992). Such significant variationsin the relative length of the stomach have not been registered in Baikal amphipods,although there is variation here, as well. The largest relative stomach length isencountered in the omnivorous B. latissima lata (19.3% of the body length), inthe spongicolous E. violaceus (17.2%), and in the necrophage O. albinus (17.2%),the smallest relative stomach length occurs in the oophage P. bazikalovae (7.6%).Probably, this species is able to find enough food, at least during the major part ofthe year.
Baikal polyphagic amphipods consume any available organic material. It isshown in the case of the three omnivorous species studied, that larger and moremobile species have a broader spectrum of feeding. The similar food choice of

MORPHOLOGY AND FEEDING IN BAIKAL GAMMARIDEA 883
different species is attributed to the similarity of their mode of life and a sharedhabitat in the same biotopes. The differences in food composition in specimens ofthe same species from the same sample are explained rather by food availability,i.e., a patchiness in the distribution of vegetable and animal organisms in thebiocenoses, than by food preference. Food composition also varies depending onseason: in winter the percentage of animal matter in the feeding of polyphagicspecies increases (Mekhanikova, 2001, 2006; original data).
During their mass development, diatoms play an important role in the feedingof amphipods (Carey & Boudrias, 1987; Quigley & Vanderploeg, 1991; Werner,1997; Polterman, 2001). At that time of the year, even the marine necrophages andcarnivores feed on diatoms (Carey & Boudrias, 1987). Many amphipods digest di-atoms, bacteria, and fungi more effectively than any other organic food such asdetritus, macroalgae, and higher plants, which serve them as a substratum (Kosta-los & Seymour, 1976; Harrison, 1977; Zimmerman et al., 1979). Diatoms are agood food source: they are preferred and digested faster than green filamentousmacroalgae (Giani & Laville, 1995). The selectivity in feeding of invertebrates isoften attributed to different availabilities of the unicellular algae, due to their dif-ferent adhesion to a substratum (Moore, 1977; Hudon, 1983). Most invertebratesprefer planktonic filamentous algae (Fragilaria, Melosira, and Diatoma), as wellas attached bushy algae (Gomphonema, Cymbella, and Rhoicosphenia), and, to alesser extent, they consume Cocconeis, Achnantes, and Amphora that are all firmlyattached to the substratum (Moore, 1975, 1977; Hudon & Bourget, 1981). A sim-ilar selectivity for different diatoms has not been observed in the Baikal speciesstudied. Both planktonic (solitary and colonial) and benthic (bushy and solitary at-tached) diatoms were present in different proportions in the food of the polyphages.Large attached diatoms (e.g., Didymosphenia) are likely to be actively consumedby amphipods, whereas small attached forms are swallowed together with the sub-stratum on which they dwell. Amphipods break up diatoms that form filamentouscolonies (Aulacoseira baicalensis) at their girdle band with the toothed incisor, andat the frustule openings with their molar teeth. In many cases, diatoms are reducedto such tiny pieces that it is impossible to identify their species or even genus.
Thus, according to the results found in the six species of Baikal endemicamphipods analysed in this paper, the relationship between their feeding habits andthe structure of their mandible has been established. The following characteristicsof amphipod mandibles and stomach are evidence of the varied composition ofthe food consumed (including diatoms with siliceous valves): the incisors andlacinia mobilis are directed backwards, there are dense rows of fine teeth on thetriturative surface of the molar, the wearing of the incisor and lacinia mobilisbetween two moults is obvious, there is a “crown” present on the incisor, and theyhave a relatively long stomach. In species feeding on soft food (amphipod eggs

884 IRINA V. MEKHANIKOVA
and rotten fish) the mandible teeth are without a “crown” and are not obviouslyworn during the period between two moults. The modifications of an oophage(P. bazikalovae) mandible are observed in the lengthening of the incisors thatare also directed vertically downwards, and in the presence of pineal elevationson the molar surface instead of rows of teeth. The most pronounced mandiblespecialization is expressed in the necrophage, O. albinus: the incisors are wide,their teeth are located in one plane, and on the molar surface there are rows ofridges. As for the spines of the lateralia, their sizes and shapes and the presenceor absence of secondary spinules on their surface are likely to be attributed, firstof all, to phylogenetic characteristics, and only in some species they express foodstrategies (the oophage P. bazikalovae). An unusual form of the lateralia spines inE. violaceus, a Baikal symbiont biting out holes in sponges, is probably evidencefor its separate taxonomic position.
ACKNOWLEDGEMENTS
This work was supported by the Integration Project SB RAS No. 95 “Studies ofCo-Adaptation in the System “Producer-Consumer” on the “Example of Diatomsand Their Consumers in Freshwater Ecosystems”.
I would like to express my thanks to the divers, the crews of the scientificresearch vessels, and to researchers from the Limnological Institute and IrkutskState University for their assistance in amphipod sampling. I thank S. Vorobyovafor consultations in identifying diatoms, V. Starostin, V. Egorov, A. Lopatin, andK. Arsentyev for help with light and scanning electron microscopy, G. Filippovafor help in finding reprints, E. Likhoshway and T. Sitnikova for revision of themanuscript and useful comments, G. Nagornaya for translation of this manuscript,S. Weinberg for his valuable comments on the manuscript and for the improvementof the English language, and A. Shirokaya for drawings and help with illustrations.I am very grateful to Dr. J. C. von Vaupel Klein and Dr. D. Platvoet for their criticalcomments on the manuscript.
REFERENCES
AGRAWAL, V. P., 1964. The digestive system of some British amphipods — III. The alimentarycanal. Proceedings of the National Academy of Sciences (India), (B, Biological) 34: 429-458.
BARNARD, J. L., 1969. The families and genera of marine gammaridean Amphipoda. U.S. NationalMuseum Bulletin, 271: 1-539.
BAZIKALOVA, A. YA., 1945. The amphipods of Lake Baikal. Proceedings of Baikal LimnologicalStation, 11: 1-440. [In Russian.]
— —, 1954. Some data on biology of Acanthogammarus (Brachyuropus) grewingki (Dyb.).Proceedings of Baikal Limnological Station, 14: 312-326. [In Russian.]

MORPHOLOGY AND FEEDING IN BAIKAL GAMMARIDEA 885
BEKMAN, M. YU., 1962. Ecology and production of Micruropus possolskii Sow. and Gmelinoidesfasciatus Stebb. Proceedings of Limnological Institute, 22 (1): 141-155. [In Russian.]
BELLAN-SANTINI, D., 1999. Ordre des Amphipodes (Amphipoda Latreille, 1816). Traité deZoologie. Anatomie, Systématique, Biologie, 7 (3A): 93-186. (Mémoires de l’InstitutOcéanographique, Monaco, 19).
CAREY, A. G. & M. A. BOUDRIAS, 1987. Feeding ecology of Pseudalibrotus (= Onisimus)litoralis Krøyer (Crustacea: Amphipoda) on the Beaufort Sea inner continental shelf. PolarBiology, 8: 29-33.
COLEMAN, C. O., 1989. On the nutrition of the two Antarctic Acanthonotozomatidae (Crustacea:Amphipoda). Gut contents and functional morphology of mouthparts. Polar Biology, 9: 287-294.
— —, 1991a. Comparative fore-gut morphology of Antarctic Amphipoda (Crustacea) adapted todifferent food sources. Hydrobiologia, 223: 1-9.
— —, 1991b. Redescription of Anchiphimedia dorsalis (Crustacea, Amphipoda, Iphimediidae) fromthe Antarctic, and functional morphology of mouthparts. Zoologica Scripta, 20: 367-374.
— —, 1992. Foregut morphology of Amphipoda (Crustacea). An example of its relevance forsystematics. Ophelia, 36: 135-150.
— —, 1994. Comparative anatomy of the alimentary canal of hyperiid amphipods. Journal ofCrustacean Biology, 14: 346-370.
DAUBY, P., Y. SCAILTEUR & C. DE BROYER, 2001. Trophic diversity within the eastern WeddellSea amphipod community. Hydrobiologia, 443: 69-86.
DE BROYER, C. & M. H. THURSTON, 1987. New Atlantic material and redescription of the typespecimens of the giant abyssal amphipod Alicella gigantea Chevreux (Crustacea). ZoologicaScripta, 16: 335-350.
DYBOWSKY, B. N., 1894. Beiträge zur näheren Kenntnis der in dem Baikal-See vorkommendenniederen Krebse aus der Gruppe der Gammariden: 1-190. (Herausgegeben von der Russ.Entomol. Gesellsch. zu St. Petersburg; Buchdr. von W. Besobrasoff und Comp., St. Petersburg).
GIANI, N. & H. LAVILLE, 1995. Réseau trophique benthique. In: R. POURRIOT & M. MEYBECK
(eds.), Limnologie générale: 565-587. (Masson Éd., Paris, Milan, Barcelona).HARRISON, P. G., 1977. Decomposition of macrophytes detritus in seawater effects of grazing by
amphipods. Oikos, 28: 165-169.HUDON, C., 1983. Selection of unicellular algae by the littoral amphipods Gammarus oceanicus
and Calliopius laeviusculus (Crustacea). Marine Biology, Berlin, 78: 59-67.HUDON, C. & E. BOURGET, 1981. Initial colonization of artificial substrate: community develop-
ment and structure studied by scanning electron microscopy. Canadian Journal of Fisheries andAquatic Sciences, 38: 1371-1384.
JOHNSTON, M. D., D. J. JOHNSTON & A. M. M. RICHARDSON, 2004. Mouthpart and digestivetract structure in four talitrid amphipods from a translittoral series in Tasmania. Journal of theMarine Biological Association of the U.K., 84: 717-726.
KAMALTYNOV, R. M., 1999. On the evolution of Lake Baikal amphipods. Crustaceana, 72 (8):921-931.
— —, 2001. Amphipods (Amphipoda, Gammaroidea). Index of animal species inhabiting LakeBaikal and its catchment area, 1 (1): 572-831. (Nauka, Novosibirsk). [In Russian.]
KAMALTYNOV, R. M., V. I. CHERNYKH, Z. V. SLUGINA & E. B. KARABANOV, 1993. Theconsortium of the sponge Lubomirskia baicalensis in Lake Baikal, east Siberia. Hydrobiologia,271: 179-189.
KAMENSKAYA, O. E., 1984. Ecological classification of deepwater amphipods. Proceedings of P.Shirshov’s Institute of Oceanology, 119: 154-160. [In Russian.]
KOSTALOS, M. & R. L. SEYMOUR, 1976. Role of microbial enriched detritus in the nutrition ofGammarus minus (Amphipoda). Oikos, 27: 512-516.

886 IRINA V. MEKHANIKOVA
LEGEZINSKA, J., 2008. Food resource partitioning among Arctic sublittoral lysianassoid amphipodsin summer. Polar Biology, 31: 663-670.
LOMONOSOV, A. M. & A. L. CHEKANOVSKY, 1869. On gases discharged from Lake Baikalbottom. News of Imperial Russian Geographical Society, 5 (2): 67-75. [In Russian.]
MEKHANIKOVA, I. V., 2001. Composition and seasonal dynamics of feeding of Brandtia parasitica(Dyb.) (Crustacea, Amphipoda) from Lake Baikal. Fauna Studies of East Siberian WaterBodies: 62-70. (Irkutsk State University, Irkutsk). [In Russian.]
— —, 2006. Comparative studies on feeding of Baicalasellus angarensis (Dyb.) (Crustacea, Isopoda)and Brandtia latissima (Dyb.) (Crustacea, Amphipoda) under conditions of their sharedhabitation in the littoral zone of southern Baikal. Proceedings of Biology and Soil Departmentof Irkutsk State University, 6: 83-93. (Irkutsk State University, Irkutsk). [In Russian.]
MELNIK, N. G., O. A. TIMOSHKIN, V. G. SIDELEVA, N. A. BONDARENKO, N. E. GUSEL-NIKOVA, I. V. MEKHANIKOVA, P. N. ANOSHKO, E. V. DZYUBA, S. G. SHUBENKOV & E.WADA, 1996. Studies on trophic structure of Lake Baikal ecosystem (stable isotope method).Abstract Book of the 7 Congress of All-Union Hydrobiological Society: 53-54. (Kazan). [InRussian.]
MOORE, J. W., 1975. The role of algae in the diet of Asellus aquaticus L. and Gammarus pulex L.Journal of Animal Ecology, 44: 719-729.
— —, 1977. Some aspects of the feeding biology of benthic herbivores. Hydrobiologia, 53: 139-146.MOORE, P. G. & P. S. RAINBOW, 1989. Feeding biology of the mesopelagic gammaridean
amphipod Parandania boecki (Stebbing, 1888) (Crustacea: Amphipoda: Stegocephalidae) fromthe Atlantic Ocean. Ophelia, 3: 1-19.
MORINO, H., R. M. KAMALTYNOV, K. NAKAI & K. MASHIKO, 2000. Phenetic analysis,trophic specialization and habitat partitioning in the Baikal amphipod genus Eulimnogammarus(Crustacea). In: Ancient Lakes: biodiversity, ecology and evolution. Advances in EcologicalResearch, 31: 355-376. (Academic Press, London).
POLTERMAN, M., 2001. Arctic sea ice as feeding ground for amphipods — food sources andstrategies. Polar Biology, 24: 89-96.
QUIGLEY, M. A. & H. A. VANDERPLOEG, 1991. Ingestion of live filamentous diatoms by theGreat Lakes amphipod, Diporeia sp.: a case of the limited value of gut contents analysis.Hydrobiologia, 223: 141-148.
SAINTE-MARIE, B., 1984. Morphological adaptations for carrion feeding in four species of littoralor circalittoral lysianassid amphipods. Canadian Journal of Zoology, 62: 1668-1674.
STEELE, D. H. & V. J. STEELE, 1993. Biting mechanisms of the amphipod Anonyx (Crustacea:Amphipoda: Lysianassoidea). Journal of Natural History, London, 27: 851-860.
TAKHTEEV, V. V., 2000. Essays on Lake Baikal amphipods: systematics, comparative ecology,evolution: 1-355. (Irkutsk State University, Irkutsk). [In Russian.]
TAKHTEEV, V. V. & I. V. MEKHANIKOVA, 1993. Some data on biology and ecology of Baikal par-asitic amphipods of the genus Pachyschesis (Amphipoda, Gammaridae). Zoological Journal,72 (4): 18-28. [In Russian.]
VINOGRADOV, G. M., 1988. System of life forms of pelagic amphipods. Reports of the USSRAcademy of Sciences, 298 (4): 1509-1512. [In Russian.]
— —, 1990. Life forms of coastal White Sea amphipods (Crustacea, Amphipoda) from VelikayaSalma area. Biological Sciences, 8: 77-85. [In Russian.]
WATLING, L., 1993. Functional morphology of the amphipod mandible. Journal of Natural History,London, 27: 837-849.
WERNER, I., 1997. Grazing of Arctic under-ice amphipods on sea-ice algae. Marine EcologyProgress Series, 160: 93-99.

MORPHOLOGY AND FEEDING IN BAIKAL GAMMARIDEA 887
ZIMMERMAN, R., R. GIBSON & J. HARRINGTON, 1979. Herbivory and detritivory amonggammaridean amphipods from a Florida seagrass community. Marine Biology, Berlin, 54 (1):41-47.
First received 29 January 2010.Final version accepted 16 April 2010.