Embed Size (px)
Citation preview

Ms. Shivani BhagwatLecturer,School of BiotechnologyDAVV
Transcription of DNA

Transcription:information transfer from DNA to RNA
The information encoded by DNA is transcribed into RNA information of RNA is translated into proteins
Transcription (RNS synthesis) is carried out by RNA polymerases RNA polymerases are large complexes of proteins, not single enzymes
Transcription is more complex in eukaryotes than in prokaryotes, but basic mechanisms are very similar in all living cells
Protein coding sequences constitute only 1-2 % of the human genome!
DNA encodes a number of RNA molecules, which are not mRNAs:ribosomal RNAs, rRNAstransferRNAs, tRNAssmall nuclear RNAs, snRNAssmall nucleolar RNA, snoRNAmicroRNAs, miRNAsanti-sense RNAsRNA molecules, that are parts of enzymes

Information transfer from DNA to RNA
Transcription and translation are closely linked processes.
In prokaryotes they are physically linked, in eukaryotes they mutually regulate each other, sharing controling molecules and mechanisms.
Cells contain 20 times more RNA than DNA
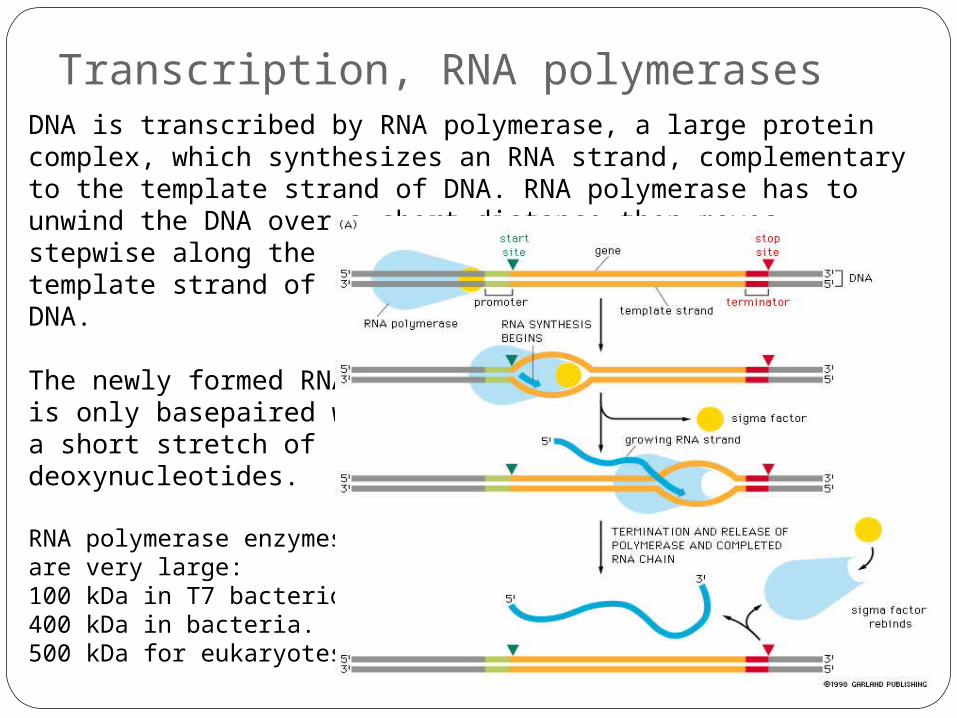
Transcription, RNA polymerasesDNA is transcribed by RNA polymerase, a large protein complex, which synthesizes an RNA strand, complementary to the template strand of DNA. RNA polymerase has to unwind the DNA over a short distance then moves stepwise along the template strand of DNA.
The newly formed RNA is only basepaired with a short stretch of deoxynucleotides.
RNA polymerase enzymes are very large:100 kDa in T7 bacteriophage. 400 kDa in bacteria. 500 kDa for eukaryotes
RNA polymerases
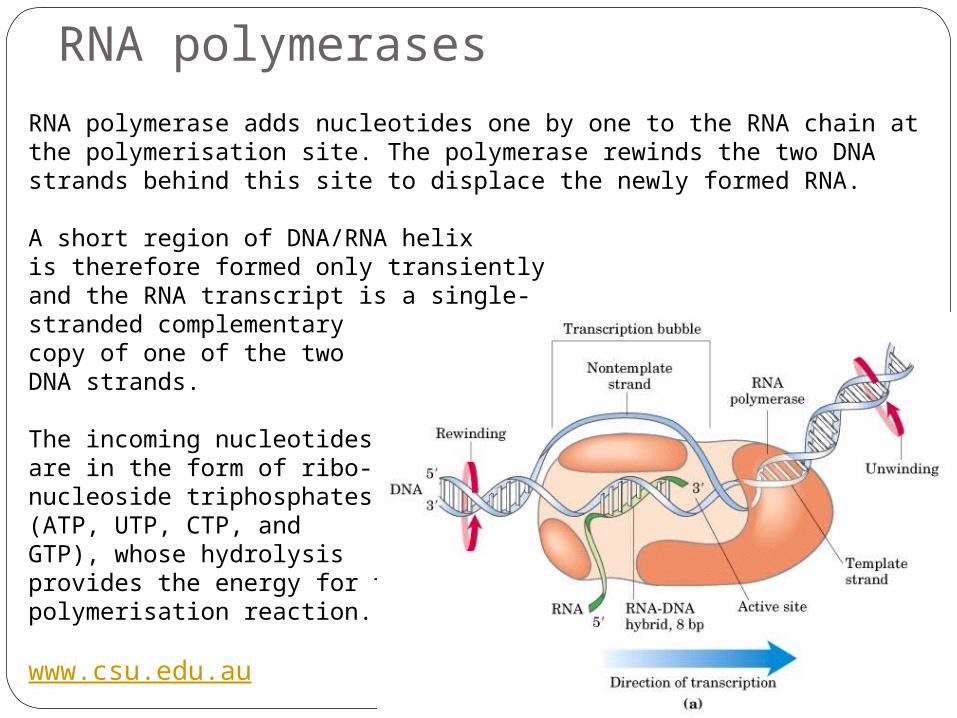
RNA polymerase adds nucleotides one by one to the RNA chain at the polymerisation site. The polymerase rewinds the two DNA strands behind this site to displace the newly formed RNA.
A short region of DNA/RNA helix is therefore formed only transientlyand the RNA transcript is a single-stranded complementary copy of one of the two DNA strands.
The incoming nucleotides are in the form of ribo-nucleoside triphosphates (ATP, UTP, CTP, and GTP), whose hydrolysis provides the energy for the polymerisation reaction.
www.csu.edu.au
RNA polymerases
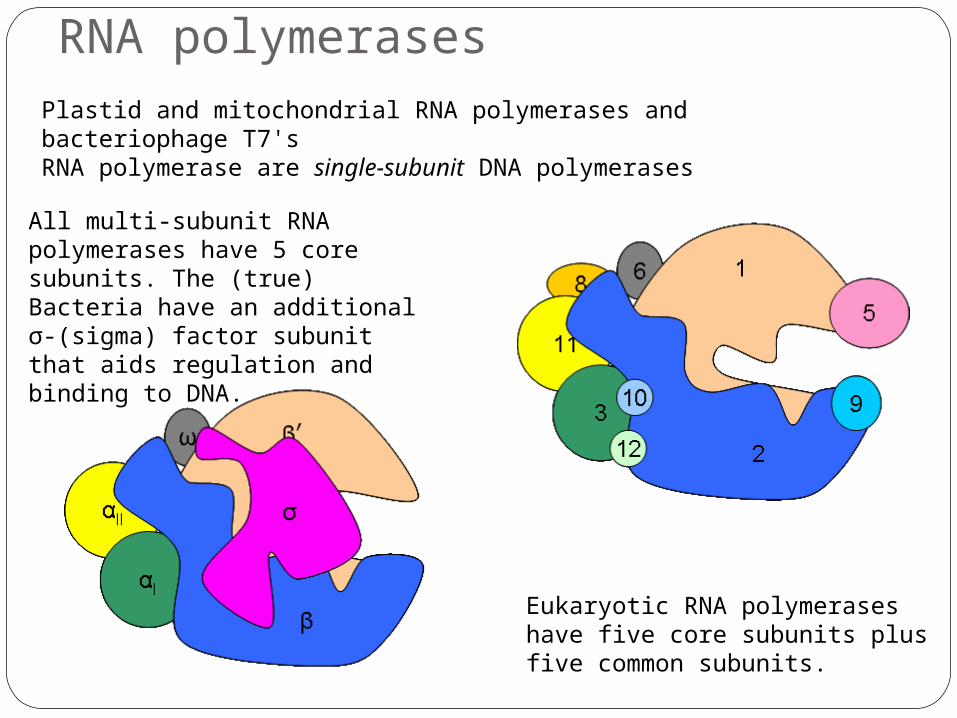
All multi-subunit RNA polymerases have 5 core subunits. The (true) Bacteria have an additional σ-(sigma) factor subunit that aids regulation and binding to DNA.
Eukaryotic RNA polymerases have five core subunits plus five common subunits.
Plastid and mitochondrial RNA polymerases and bacteriophage T7's RNA polymerase are single-subunit DNA polymerases

The prokaryotic holoenzyme has 2 alpha, 2 beta and an omega subunits. It binds DNA with no sequence specificity.
To find the promoter sequence it needs another subunit: the sigma factor
Formation of the open complex from closed DNA is usually the slowest step in transcription initiation. About 10 nucleotides are polymerized in the open complex before it must undergo promoter clearance. A conformational change must take place that releases the sigma factor.
RNA polymerases

Prokaryotic promoter sequences
Promoter -35 Region spacer -10 Region starttrp operon TTGACA N17 TTAACT N7AtRNA tyr TTACA N16 TATGAT N7AlP2 TTGACA N17 GATACT N6Glac operon TTTACA N17 TATGTT N6Arec A TTGATA N16 TATAAT N7Alex A TTCCAA N17 TATACT N6AT7A3 TTGACA N17 TACGAT N7Aconsensus TTGACA TATAAT
TGAC box (-35) TATAAT Box (-10), This sequence was initially called the Pribnow Box. Do not refer this sequence as a TATA box.
All these promoter sequences are recognized by the sigma70 subunit in E. coli. Note the degree of sequence variation at each position. The consensus sequence has been derived from a much larger database of over 300 well-characterized promoters.The "consensus sequence" is a hypothetical sequence made up of the nucleotides found most often in each position. There may be no single organism with this exact set of nucleotides in the stated positions. The promoter sequences are asymmetrical, the promoter will bind the polymerase in only one orientation, thus determining which strand of the DNA will be transcribed.

RNA synthesis by RNA polRNA polymerase synthesizes ribosomal RNAs. In bacteria there are many (in E. coli 7) ribosomal rRNA genes.
RNA polymerase – as all other NA synthesizing enzymes – polymerizes from 5’>3’, DNA is read from 3’>5’
The produced RNA is complementary to template strand,and equivalent to non-template strand RNA polymerase stops at termination or stop signal.Rho factor binds RNA and helps recognizing termination signals (RNA loops).
Palindromic sequences (e.g.AATCG.CGATT) form loops on the RNA and serve assignal termination.

The 16S rRNA
The 16S rRNA
The 16S rRNA is 1.5 kb longand has an elaborate secondary structure

rRNA molecules in the ribosome
Ribosomes are very large RNA-protein complexes, subcellular organelles.
They are responsible for the synthesis of proteins.
Both prokaryotic and eukaryotic ribosomes consist of two subunits and have 3 (4) RNA molecules.
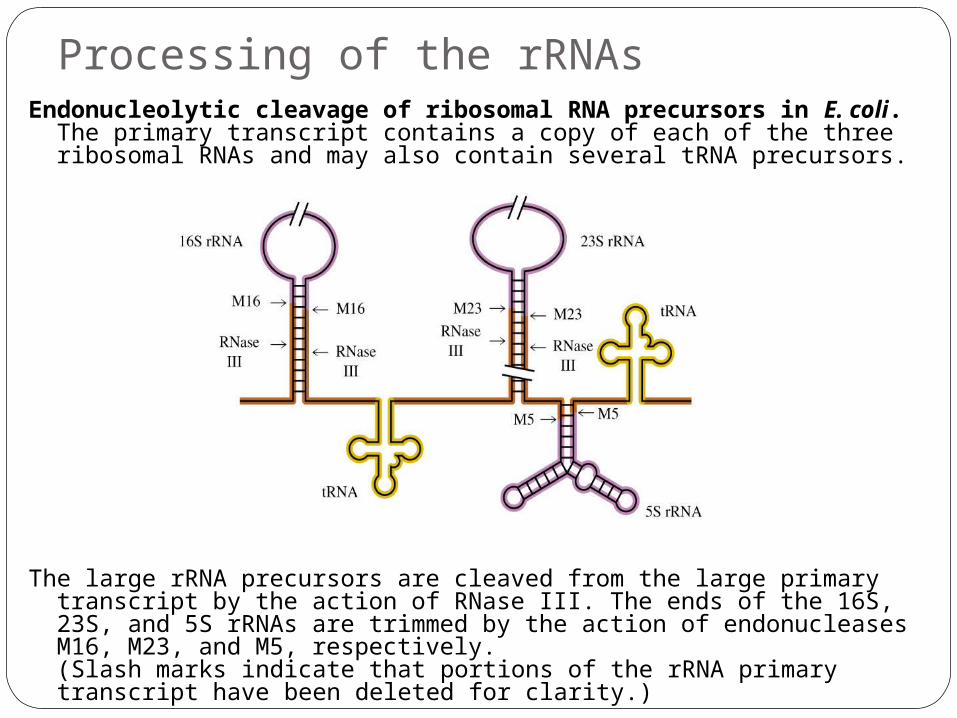
Processing of the rRNAsEndonucleolytic cleavage of ribosomal RNA precursors in E. coli.
The primary transcript contains a copy of each of the three ribosomal RNAs and may also contain several tRNA precursors.
The large rRNA precursors are cleaved from the large primary transcript by the action of RNase III. The ends of the 16S, 23S, and 5S rRNAs are trimmed by the action of endonucleases M16, M23, and M5, respectively. (Slash marks indicate that portions of the rRNA primary transcript have been deleted for clarity.)
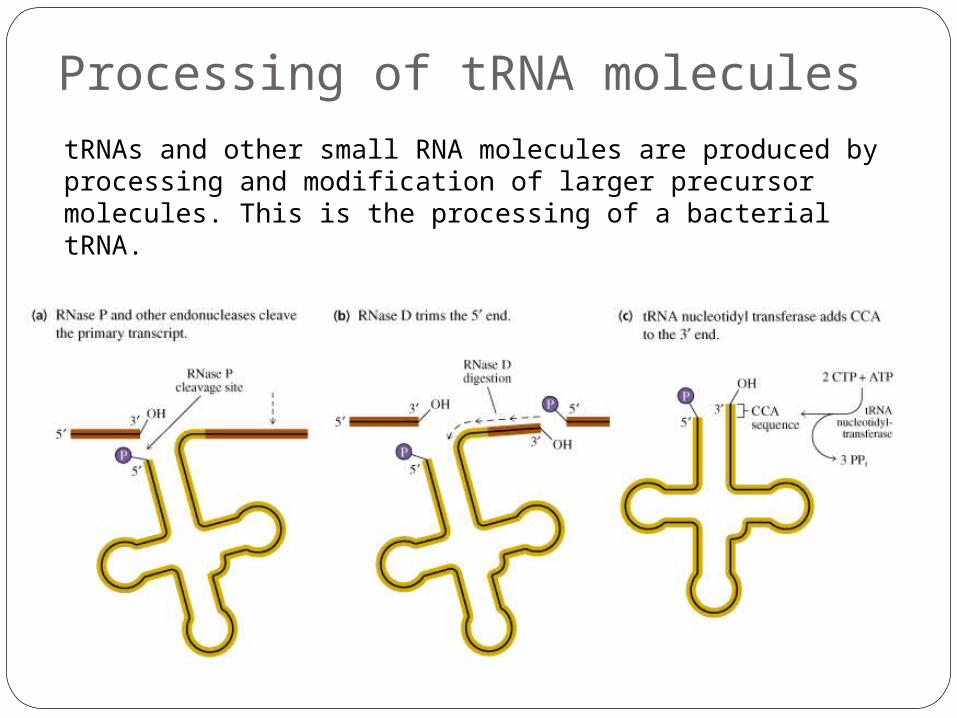
Processing of tRNA molecules tRNAs and other small RNA molecules are produced by processing and modification of larger precursor molecules. This is the processing of a bacterial tRNA.

Synthesis of prokaryotic mRNAs
Genes coding for proteins are first transcribed into mRNAs. One gene codes for a protein. In cases of multi-subunit proteins with heterolo-gous subunits, one cistron codes for a polypeptide chain. (A protein with four different subunits is coded by four cistrons.)
In prokaryotes there are single cistronic and polycistronic messages, coding for one protein or a chain of (related) enzymes. Metabolically linked enzymes (e.g. enzymes of a synthetic pathway) are usually controlled together.
Polycistronic mRNAsare characteristic of prokaryotic organisms.
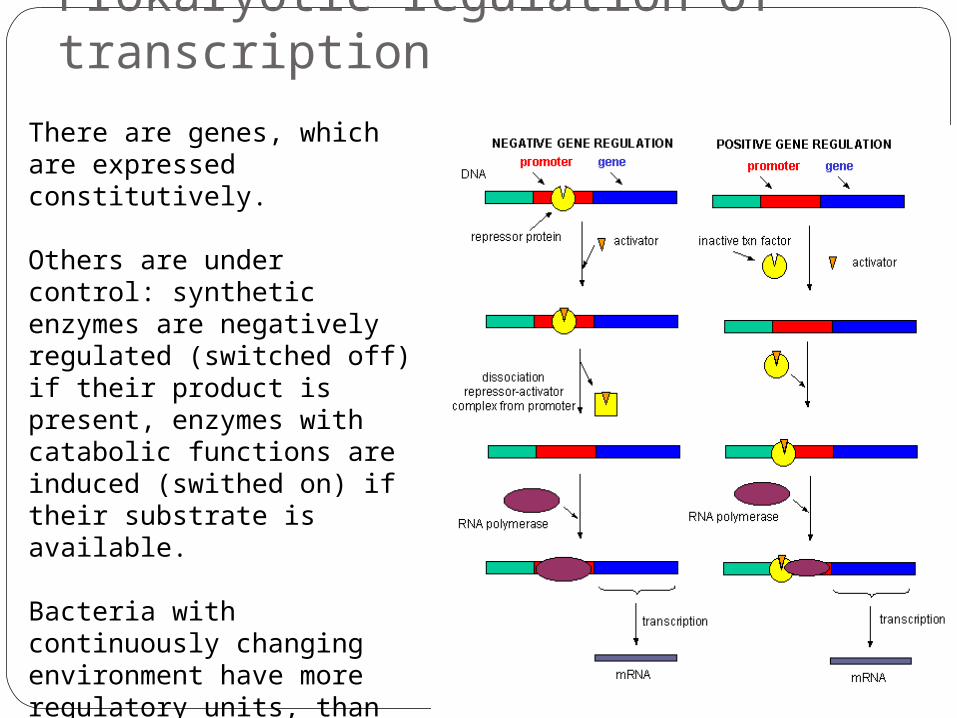
Prokaryotic regulation of transcription
There are genes, which are expressed constitutively.
Others are under control: synthetic enzymes are negatively regulated (switched off) if their product is present, enzymes with catabolic functions are induced (swithed on) if their substrate is available.
Bacteria with continuously changing environment have more regulatory units, than those, which live in more stable niche.

The lac operon

Regulatory elements of the eukaryotic promoter
A TATA Box, located approximately 25 bp upstream of the startpoint of transcription is found in many promoters. The consenus sequence of this element is TATAAAA, it resembles the TATAAT sequence of the prokaryotic -10 region (please do not mix them up!)
TATA box appears to be more important for selecting the startpoint of transcription (i.e. positioning the enzyme) than for defining the promoter. The Initiator is a sequence that is found in many promoters and defines the startpoint of transcription.
The GC box is a common element in eukaryotic class II promoters. Its consensus sequence is GGGCGG. It may be present in one or more copies which can be located between 40 and 100 bp upstream of the startpoint of transcription. The transcription factor Sp1 binds to the GC box. The CAAT box - consensus sequence CCAAT - is also often found between 40 and 100 bp upstream of the startpoint of transcription. The transcription factor CTF or NF1 binds to the CAAT box.
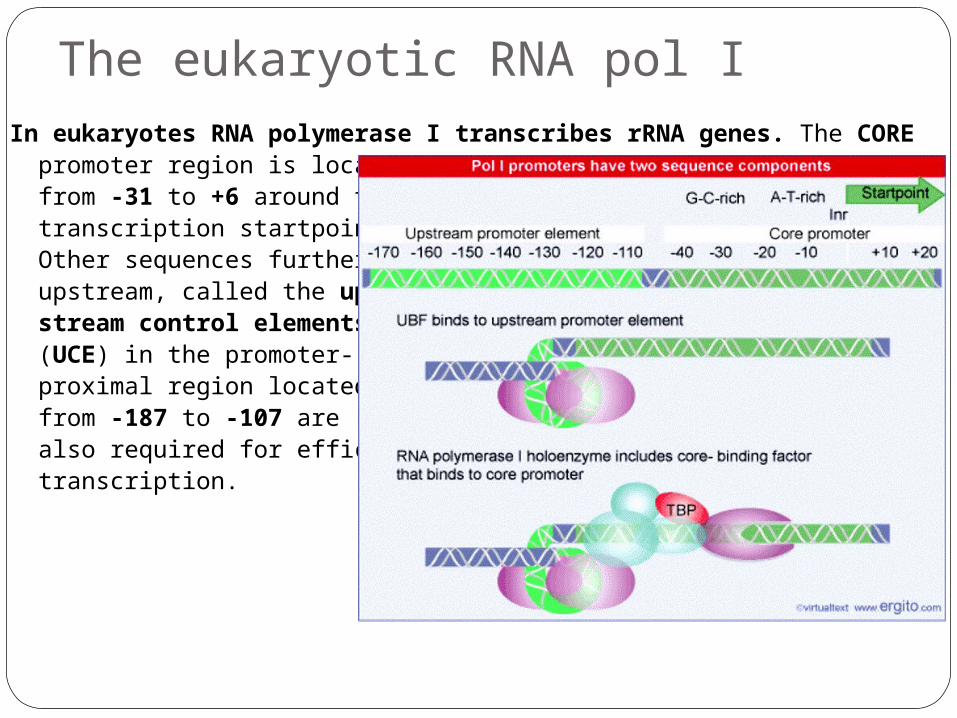
The eukaryotic RNA pol IIn eukaryotes RNA polymerase I transcribes rRNA genes. The CORE
promoter region is located from -31 to +6 around the transcription startpoint. Other sequences further upstream, called the up-stream control elements (UCE) in the promoter-proximal region located from -187 to -107 are also required for efficient transcription.
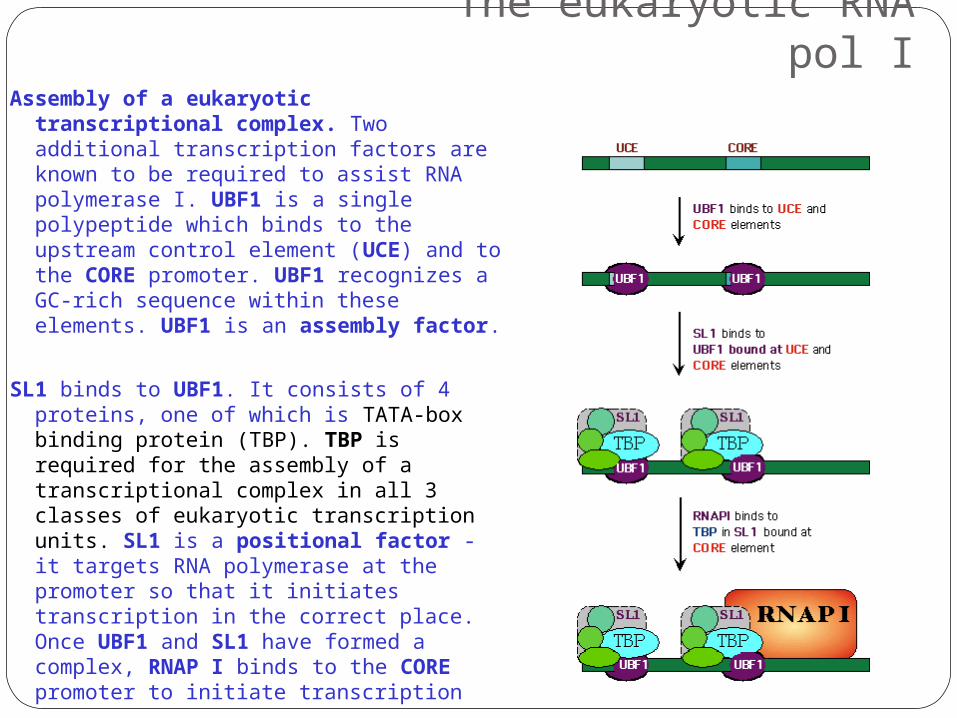
The eukaryotic RNA pol IAssembly of a eukaryotic
transcriptional complex. Two additional transcription factors are known to be required to assist RNA polymerase I. UBF1 is a single polypeptide which binds to the upstream control element (UCE) and to the CORE promoter. UBF1 recognizes a GC-rich sequence within these elements. UBF1 is an assembly factor.
SL1 binds to UBF1. It consists of 4 proteins, one of which is TATA-box binding protein (TBP). TBP is required for the assembly of a transcriptional complex in all 3 classes of eukaryotic transcription units. SL1 is a positional factor - it targets RNA polymerase at the promoter so that it initiates transcription in the correct place. Once UBF1 and SL1 have formed a complex, RNAP I binds to the CORE promoter to initiate transcription

RNA polymerase IIRNA polymerase II All genes that are transcribed and expressed via
mRNA are transcribed by RNA polymerase II. Transcription copies the DNA code of a gene and converts it to high mol mass nuclear RNA (hnRNA), which is precessed to mRNA. The mRNA will be used at the ribosome to make polypeptides (proteins).
RNA polymerase II is a multisubunit enzyme-complex. The yeast enzyme has 12 subunits. The largest subunit is the homologue of the beta subunit of the prokaryotic enzyme, it contains the catalytic activity. The enzyme is very sensitive to amanitin, a mushroom toxin.
RNA polymerase II can transcribe RNA from nicked dsDNA templates or from ssDNA templates. However, it cannot initiate transcription at a promoter. In this respect, it resembles the core form of bacterial RNA polymerase.
The promoters used by RNA polymerase II have different structures depending upon the particular combination of transcription factors that are required to build a functional transcriptional complex at each promoter. Nevertheless, these different structures can be viewed as a combination of a relatively limited number of specific sequence elements.
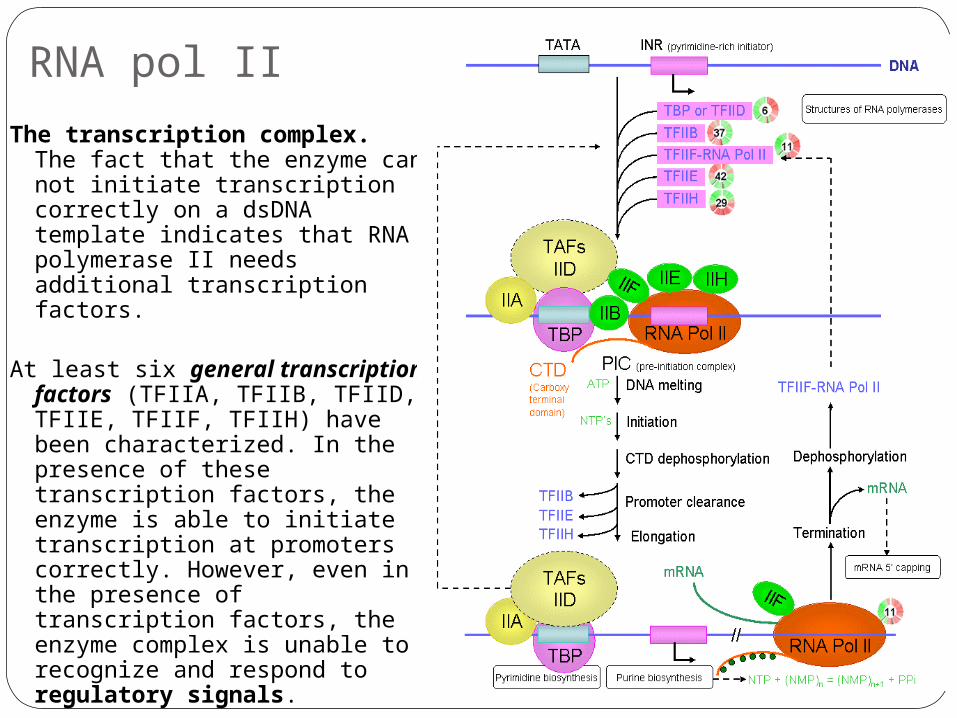
RNA pol II
The transcription complex. The fact that the enzyme can not initiate transcription correctly on a dsDNA template indicates that RNA polymerase II needs additional transcription factors.
At least six general transcription factors (TFIIA, TFIIB, TFIID, TFIIE, TFIIF, TFIIH) have been characterized. In the presence of these transcription factors, the enzyme is able to initiate transcription at promoters correctly. However, even in the presence of transcription factors, the enzyme complex is unable to recognize and respond to regulatory signals.

The eukaryotic RNA pol II
rotation of DNA:www.k2.phys.waseda.ac.jp/Rnapmovies/RNAP.htm

The RNA pol II transcription complexIn addition to the general transcription factors, the eukaryotic
transcriptional complex will also be affected by the presence of an promoter-proximal regulatory sequences (transcription elements or motifs) and the presence of transcription factors that bind to those sequences.
Such factors may be present in some cells/tissues but not in others.
.

hnRNA and mRNA
All of the code contained in the hnRNA molecule is not needed to produce the polypeptide. The sections of hnRNA which do not code for translation of polypeptide are called introns and untranslated region.
As the mRNA readies itself to leave the nucleus, enzymes cut out and remove the introns. The remaining exons are spliced back together again by a different enzyme. This modified mRNA is what comes to the ribosome to be translated into polypeptides. Eukaryotic transcription and mRNA synthesis is not taking place in discrete steps: transcription, capping, tailing, splicing and export from the nucleus for translation. The contemporary view of eukaryotic gene expression entails simultaneous transcription and processing..
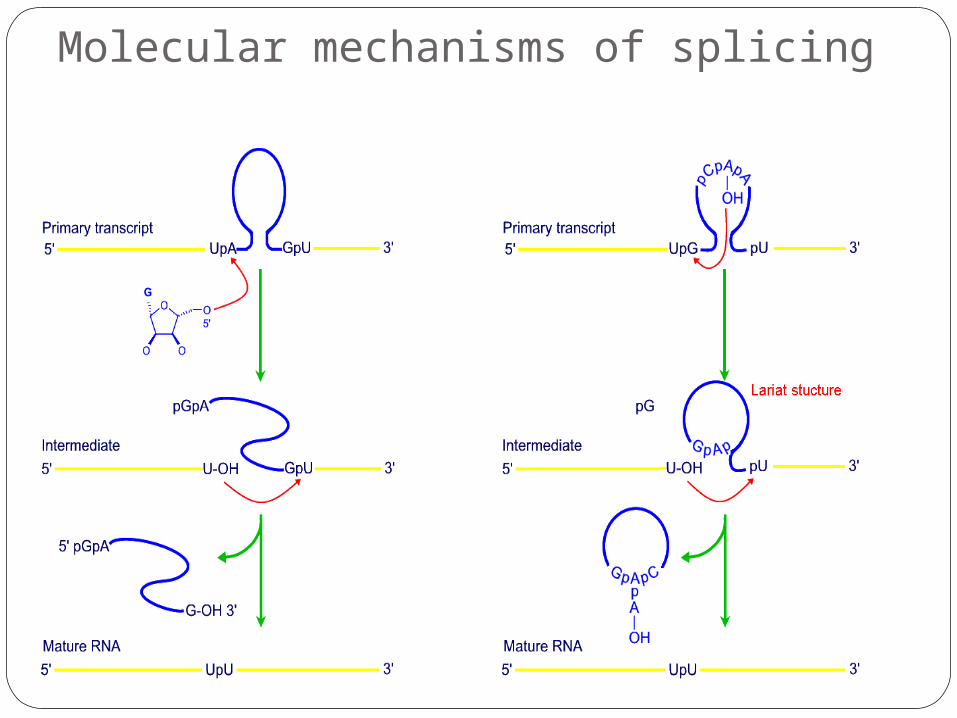
Molecular mechanisms of splicing
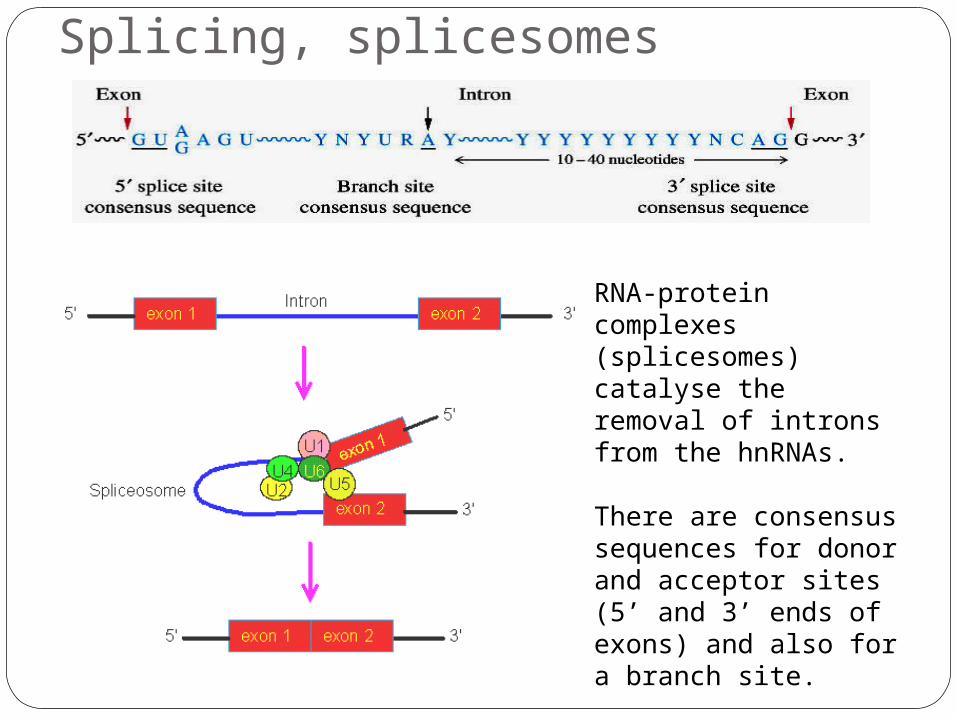
Splicing, splicesomes
RNA-protein complexes (splicesomes) catalyse the removal of introns from the hnRNAs.
There are consensus sequences for donor and acceptor sites (5’ and 3’ ends of exons) and also for a branch site.
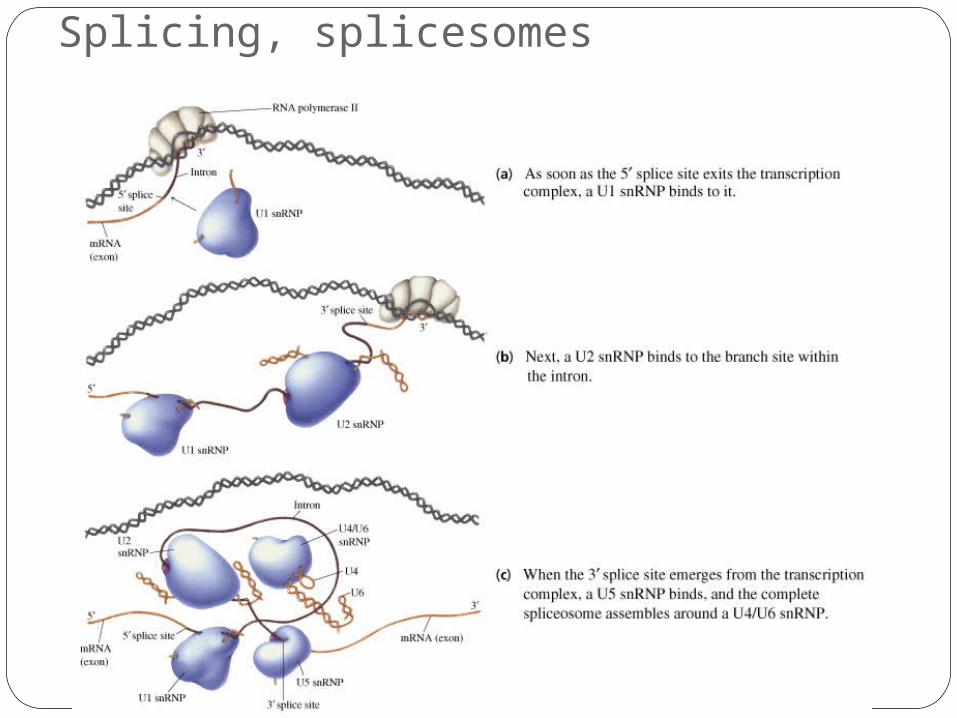
Splicing, splicesomes

mRNAs usually contain many intronsOnly eukaryotic mRNAs do
contain exons and introns. The are single intron mRNAs, but most proteins are coded by multi-intron mRNAs.
Certain introns (or homologues) can be found in many proteins of similar functions.
In different tissues (or in developmental stages) different splice variants of the same mRNA can be found (alternative splicing).

Alternative splicing
Alternative splicing makes the genome more flexible and economical: with one gene many proteins of different functions can be coded. One example is sex determination in Drosophila.

Capping and polyadenylatingmRNA molecules undergo other types of modifications.
To increase stability, a „cap” structure (a mG of reverse orientation) is added to the 5’ end: this way the molecule has only 3’ ends!
The 3’ polyadenylation sequence is bound by a cleavage and polyadenylation specificity factor (CPSF). An endonuclease binds to CPSF and cleaves the transcript. Finally a polymerase binds to CPSF, adding up to 250 adenylate residues to the 3' end of the transcript. (This polymerase does not need a template!) The length of the polyA tail regulates translational activity of the mRNA.
Some viral mRNAs do not contain caps: instead, a protein is linked to the mRNA (covalently).

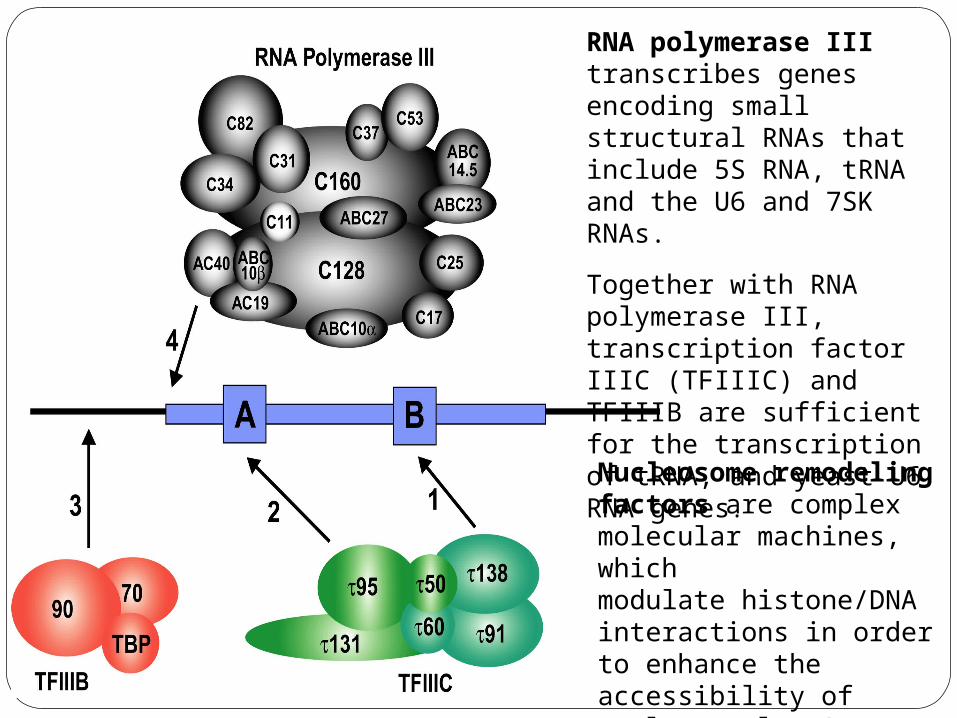
RNA polymerase III
RNA polymerase transcribes tRNAs and other low MM RNAs, usually in a precursor form.
tRNA molecules are adaptorseach amino acid has at least one tRNA, which brings aa’s to the ribosomes.
Aminoacyl-tRNA synthetases charge specific tRNAs with cognate amino acids
More tRNAs serve one amino acid

Processing of the tRNAs
The transcription product, the pre-tRNA, contains additional RNA sequences at both the 5’ and 3’-ends. The additional nucleotides at the 5’-end are removed by an unusual RNA containing enzyme called ribonuclease P (RNase P).
An intron is removed from the tRNA mol.
All mature tRNAs contain the trinucleotide CCA at their 3’-end. These bases are not coded for by the tRNA gene. Instead, these nucleotides are added to the pre-tRNA transcript. The enzyme responsible for the addition of the CCA-end is tRNA nucleotidyl transferase
Mature tRNAs can contain up to 10% bases other than the usual A, G, C and U. These base modifications are introduced into the tRNA at the final processing step.

tRNA moleculesModels of tRNA. The tRNAs contain special bases (not found in other NAs), these are modified after synthesis. The bases denoted psi (pseudouridine) and D (dihydro-uridine) are derived from uracil.
Functional domains of the tRNA molecule. The acceptor stem is aminoacylated, the anticodon arm recognizes the codon of the mRNA.The figure shows how a tRNA molecule is shaped due to basepair interactions. Note the specificity due to the anticodon and due to the fact that each tRNA is charged with a specific amino acid.
Nucleosome remodeling factors are complex molecular machines, whichmodulate histone/DNA interactions in order to enhance the accessibility of nucleosomal DNA.
RNA polymerase III transcribes genes encoding small structural RNAs that include 5S RNA, tRNA and the U6 and 7SK RNAs.
Together with RNA polymerase III, transcription factor IIIC (TFIIIC) and TFIIIB are sufficient for the transcription of tRNA, and yeast U6 RNA genes.

MicroRNAs, antisense and gene silencing
The discovery of microRNAs and the phenomenon of RNA interference might change our whole idea of gene expression and gene regulation.
The human genome codes for a large number of microRNA sequences, their importance is just emerging.
It was known that only a small fraction of mRNAs leave the nucleus and participate in protein synthesis.





























