Embed Size (px)
Citation preview

Multi-objective optimization of structures topology by genetic algorithms
J.F. Aguilar Madeiraa,b,*, H. Rodriguesa, Heitor Pinaa
aIDMEC/IST-Mechanical Engineering Department, Instituto Superior Tecnico, Av. Rovisco Pais, 1049-001 Lisbon, PortugalbISEL-Instituto Superior de Engenharia de Lisboa, Lisbon, Portugal
Received 1 November 2002; accepted 25 July 2003
Available online 24 July 2004
Abstract
This work develops a computational model for topology optimization of linear elastic structures for situations where more than one
objective function is required, each one of them with a different optimal solution.
The method is thus developed for multi-objective optimization problems and is based on Genetic Algorithms. Its purpose is to evolve an
evenly distributed group of solutions (population) to obtain the optimum Pareto set for the given problem.
To reduce computational effort, optimal solutions of each of the single-objective problems are introduced in the initial population.
Two numerical examples are presented and discussed to assess the method.
q 2004 Civil-Comp Ltd and Elsevier Ltd. All rights reserved.
Keywords: Structural topology design; Multi-Objective Optimization; Genetic Algorithm; Evolutionary Algorithms
1. Introduction
In its most general form, the topology optimization
problem of structures can be viewed as the process of
constructing the characteristic (indicator) function of the
domain occupied by the material, i.e.
x ¼1 if material exists
0 if material doesn0t exist
(
for a given total amount of material.
The binary chromosome design and global search
capabilities of the Genetic Algorithm (GA) make is a
powerful tool for solving topology design problem [1,9].
In this work a GA for multi-objective topology
optimization of linear elastic structures, with an equality
constraint on volume, is developed. Due to this constraint all
the chromosomes used in the GA are constructed to ensure
the same volume for all individuals (designs), hence all
of them have the same number of ones (implicitly, the same
number of zeros) along the evolutionary process.
For a typical chromosome defined by L genes there are a
total of 2L possible solutions in the representation scheme.
Considering the volume equality constraint, a significant
decrease of this value can be achieved. If a chromosome
with a total of L genes is considered and K genes being one
ð1 , K , L; ðL 2 KÞ genes being zero), only
CLK ¼
L!
K!ðL 2 KÞ!
possible configurations are possible. The present analysis
explores this fact to reduce the computational effort
required. It is thus necessary to define these chromosomes
and to create accordingly new operators of crossover and
mutation that preserve this constraint.
In contrast to single-objective optimization, where
objective and fitness functions are often identical, both
fitness assignment and selection must allow for several
objectives when multi-objective optimization problems are
considered. Hence, instead of a single optima, multi-
objective optimization problems solutions consist often of
a family of points, known as the Pareto optimal set, where
each objective component of any point along the Pareto-front
can only be improved by degrading at least one of its other
objective components. In the total absence of information
regarding the preferences of objectives, a ranking scheme
based upon the Pareto optimality is regarded as an
appropriate approach to represent the strength of each
individual in an evolutionary algorithm for multi-objective
optimization [7,17].
Since a great diversity of solutions exist, additional
information obtained a priori from classical methods for
0965-9978/$ - see front matter q 2004 Civil-Comp Ltd and Elsevier Ltd. All rights reserved.
doi:10.1016/j.advengsoft.2003.07.001
Advances in Engineering Software 36 (2005) 21–28
www.elsevier.com/locate/advengsoft
* Corresponding author.
E-mail address: [email protected] (J.F.A. Madeira).

single-objective optimization is incorporated in the algo-
rithm to obtain a reduction in the computational effort.
Some mathematical support for this is given in Ref. [13].
This information is introduced in the initial population
through special individuals (see example 1 in Section 6).
2. Background
2.1. A general multi-objective optimization problem
(MOOP)
A general multi-objective optimization problem consist-
ing of k competing objectives and ðm þ p þ qÞ constraints,
defined as functions of decision variable set x; can be
represented as follows:
Minimize=Maximize fðxÞ ¼ ðf1ðxÞ; f2ðxÞ;…; fkðxÞÞ [ Y;
subject to giðxÞ # 0; ;i ¼ 1; 2;…;m;
hjðxÞ ¼ 0; ;j ¼ 1; 2;…; p;
xLl # xl # xU
l ; ;l ¼ 1; 2;…; q;
ð1Þ
where x ¼ ðx1; x2;…; xnÞ [ X:where x is the decision
vector, fðxÞ the multi-objective vector, fjðxÞ the jth objective
function, X denotes as the decision space and Y the
objective space.The constraints giðxÞ and hjðxÞ determine
the set of feasible solutions. The remaining set of constraints
are called variable bonds, restricting each decision variable
xl to values between a lower xLl and an upper xU
l bond.
2.2. Concept of domination
Most multi-objective optimization algorithms use the
concept of domination referred as Pareto optimality [14]. A
non-dominated set of solutions is formally defined as
follows [2]: ‘a feasible solution to a multi-objective problem
is non-dominated if there exists no other feasible solution
that will yield an improvement in one objective without
causing a degradation in at least one other objective’.
The set of solutions of a multi-objective optimization
problem consist then of all decision vectors which cannot be
improved in any objective without degradation in the other
objectives. This vector is know as Pareto-optima. Mathemati-
cally, the concept of Pareto-optima is as follows. Assuming,
without loss of generality, a minimization problem and
considering two decision vectors a; b [ X; we say that a
dominates b (also written as a a b) if and only if
;i[{1;2;…;k} : fiðaÞ# fiðbÞ^’j[{1;2;…;k} : fiðaÞ, fiðbÞ
ð2Þ
All decision vectors which are not dominated by any
other decision vector are called non-dominated or Pareto-
optimal. The family of all non-dominated solutions is
denoted as Pareto-optimal set (Pareto set) or Pareto-optimal
front. It is also customary to write any of the following:
† b is dominated by a;
† a is non-dominated by b; or
† a is non-inferior to b:
2.3. Multi-objective evolutionary algorithms (MOEAs)
A number of multi-objective optimization techniques
using evolutionary algorithms have been suggested since
the pioneering work by Schaffer in Refs. [15,16]. Other
important developments are described in Refs. [7].
Evolutionary optimization algorithms work with a
population of solutions instead of a single solution. Since
multi-objective optimization problems give rise to a set of
Pareto-optimal solutions, evolutionary optimization algo-
rithms are ideal for handling multi-objective optimization
problems.
In order to obtain effective results with an evolutionary
algorithm for multi-objective optimization problems we
need to guide the search toward the Pareto-optimal set and
maintain a diverse population to prevent premature
convergence and to achieve a well distributed population
in the Pareto front.
3. Methodology
As previously stated, the equality constraint on volume
requires that all chromosomes used in the GA lead to the
same volume value, hence all of them should have the same
number of ones along the evolutionary process. It is thus
necessary to properly define these chromosomes and to
create crossover and mutation operators that enforce this
volume constraint.
3.1. The chromosome storage
The topology model for this research is a grid that defines
the design domain. A value of 1 assigned to an element or
cell corresponds to the presence of material and a value 0 to
a void in the domain.Fig. 1 shows the structural topology
represented in the grid model (B) by the chromosome array
(C), the binary chromosome string (D) (containing the
numbering of the elements (A) and the volume value 0 or 1
for each element), and the chromosome viewed as a loop (E)
formed by joining the ends together. A chromosome
considered as a loop can represent more building blocks
for the crossover operator, since they are able to ‘wrap
around’ at the end of the string. These two last represen-
tations depend on the elements numbering.
3.2. Crossover techniques
The selected progenitors exchange between them
their genetic material producing individuals with new
J.F.A. Madeira et al. / Advances in Engineering Software 36 (2005) 21–2822

characteristics. To preserve volume the progenitors and the
descendants must have the same number of ones in their
chromosomes, which requires new types of crossover to be
described below.
3.2.1. Algorithm
To explain these crossover techniques the chromosome’s
representation is considered as a loop (Fig. 2).
In the following crossover operator Ngc; a; p1; p2; n1s1; e
n1s2 are natural numbers representing:
Ngc Number of genes in the chromosome (in Fig. 2,
Ngc ¼ 16Þ;
a Number of genes to exchange in the crossover
operator,
p1 Number of the gene of the first progenitor where
the cut begins,
p2 Number of the gene of the second progenitor,
where the cut begins,
n1s1 Number of ones of the first progenitor existing in
the interval ½p1; p1 þ ða2 1Þ�;
n1s2 Number of ones of the second progenitor existing
in the interval ½p2; p2 þ ða2 1Þ�:
The following algorithm describes a step-by-step pro-
cedure for finding the progenitors gene sequence to use in
the crossover operator:
Step 1 Fix a; the number of genes to exchange.
Step 2 Determine the inicial p1:
Step 3 Determine the inicial p2 (Fig. 3).
Step 4 The number of ones of the first progenitor existing
in the interval ½p1; p1 þ ða2 1Þ� is counted. Let us
say that there are n1s1 ones.
Step 5 The number of ones of the first progenitor existing
in the interval ½p2; p2 þ ða2 1Þ� is counted. Let us
say that there are n1s2 ones.
Step 6 If {ðn1s1 – n1s2Þ ^ ðp2 , NgcÞ} increment p2 by
one, and go to Step 5.
Step 7 If {ðn1s1 – n1s2Þ ^ ðp2 ¼ NgcÞ} increment p1 by
one, and go to Step 4
Step 8 If n1s1 ¼ n1s2; make the exchange.
Fig. 2. Chromosome representation considered as a loop.
Fig. 1. Element numbering (A), structural topology in the grid model (B), binary chromosome array (C), binary chromosome string (D), and chromosome
viewed as a loop (E).
J.F.A. Madeira et al. / Advances in Engineering Software 36 (2005) 21–28 23

3.2.2. Types of crossover with this technique
In the Step 2 and Step 3, the determination of p1 and p2
leads to three possible types of crossover: description
Type 1 Start with p1 ¼ p2 ¼ fixed value; for all the
evolution.
Type 2 Start with p1 ¼ p2 ¼ random value; this value to
be determined whenever the operator is used.
Type 3 p1 and p2 are determined by an independent
random choice
. In the first type, whenever two individuals cross their
descendants will always be the same. In the second type, the
probability to get the same descendants is smaller, so we
have a bigger diversity of descendants than in the previous
one. The third type gives a higher diversity in the
descendant population compared with the previous ones
and is by this reason the one used in Section 6.
3.2.3. Example
We illustrate the method presented with the example of
Fig. 2. Let us consider a ¼ 8; p1 ¼ 1 e p2 ¼ 1: The number
of ones in the marked region is for the first progenitor,
n1s1 ¼ 4 and, for the second, n1s2 ¼ 5 (Fig. 4). We are in
Step 5.
As it happens that {ðn1s1 – n1s2Þ ^ ðp2 , NgcÞ}; then
increment p2 by one (Fig. 5) and return to Step 5.
In summary, with
p2 ¼ 1 ) n1s2 ¼ 5 – n1s1 ¼ 4 and p2 , Ngc;
p2 ¼ 2 ) n1s2 ¼ 5 – n1s1 ¼ 4 and p2 , Ngc;
p2 ¼ 3 ) n1s2 ¼ 5 – n1s1 ¼ 4 and p2 , Ngc;
with p2 ¼ 4 we finally get that, n1s1 ¼ n1s2 ¼ 4; then we
make the exchange of genetic material of the two
progenitors, taking into account the point of the selected
cut, as it is suggested in Fig. 6.
3.3. Mutation
In spite its secondary importance as compared with
the crossover operator, the mutation plays an important
role in the GA procedure as it is responsible for the
maintenance of the population genetic diversity [10] and
to prevent the algorithm to get locked in a local extreme
by making it explore other regions. The method is as
follows:
Step 1 Determine randomly the position of two genes in
the chromosome.
Step 2 If they have equal values, they will remain in the
same position. Return to Step 1.
Step 3 If they have different values, they will exchange
positions.
4. Multi-objective optimization problem (MOOP)
4.1. Formulation
Consider a structural component, occupying the struc-
tural domain V; subjected to applied body forces b;
boundary traction t on Gt and essential boundary conditions
on Gu (Fig. 7).
To introduce the material based formulation, consider the
structural component made of material with variable
volume fraction m:
The k competing objectives for a multi-objective
optimization problem are:
faðxÞ ¼ðV
biuai dVþ
ðGt
tai uai dG
� �witha¼ 1;2;…;k ð3Þ
Fig. 5. a ¼ 8; p1 ¼ 1; p2 ¼ 2; n1s1 ¼ 4 e n1s2 ¼ 5:
Fig. 4. a ¼ 8; p1 ¼ 1; p2 ¼ 1; n1s1 ¼ 4 e n1s2 ¼ 5:
Fig. 3. Example with: a ¼ 8; p1 ¼ 1 e p2 ¼ 4:
J.F.A. Madeira et al. / Advances in Engineering Software 36 (2005) 21–2824

where faðxÞ with a ¼ 1; 2;…; k represents the compliance
(equivalent to the energy norm of the total displacement) for
the load case a (boundary traction ta and body forces b) and
where ua is the respective displacement field.
The multi-objective optimization problem is:
Minimize fðxÞ ¼ ðf1ðxÞ; f2ðxÞ;…; fkðxÞÞ [ Y; ð4Þ
where x ¼ ðx1; x2;…; xnÞ [ X (decision space) and Y is the
objective space, fðxÞ the multi-objective vector, and fjðxÞ the
jth objective function.subjected to the volume constraint,ðVm dV ¼ vol ð5Þ
and where the displacement ua is the solution of the integral
equilibrium equation, in virtual displacement form,ðV
Eijkmðm; qÞeijðuaÞekmðwÞ2 biwi dV2
ðGt
tai wi dG
¼ 0; ;w admissible: ð6Þ
5. Computational model
A flow chart of the general procedure for a optimization
problem with two objectives is outlined in Fig. 8
5.1. Populations
Assuming that the initial population has N individuals
and MOOP has K competing objectives, the initial
population is determined by the following procedure:
(i) Solve K single-objective problems to determine the K
optimal solutions for each objective, such that:
necessary conditions for the optimum are derived
analytically, approximated numerically through a
suitable finite element discretization and solved by a
first order method based on the optimization problem.
(ii) The initial population is formed by these K optimal
solutions of the single-objective problems and by
ðN 2 KÞ different individuals randomly generated.
Hence, the initial population has N different individuals.
Fig. 6. Crossover example.
Fig. 7. General problem of elasticity. Fig. 8. Numerical model flow chart.
J.F.A. Madeira et al. / Advances in Engineering Software 36 (2005) 21–28 25
To achieve low memory demand and computational
effort reduction, the algorithm does not allow two equal
individuals to be present in the population.
5.2. Finite element method (FEM) and parallel processing
Using the finite element method, the work of the external
loads is calculated for each objective. As the work of the
external loads can be calculated independently for each
individual parallel processing was used in its evaluation.
5.3. Fitness assignment, new population for mating
and selection
The concept of calculating an individual’s fitness on the
basis of Pareto dominance was first suggested in Ref. [8].
First, a ranking strategy based in non-domination properties
is used [3], that is, the non-dominated solution receives
the rank 1 (front 1), then a new non-domination test is made
in the population without these solutions and the new non-
dominated solution receives the rank 2 (front 2). This
procedure is repeated until all solutions are ranked. Second,
the rank of an individual determines its fitness value. It is
worth remarking the fact that individual fitness is related to
the whole population.
In order to avoid a possible premature convergence to a
population where only non-dominated individuals (of front
1) exist, it is imposed that the new population for matting
will be constituted by individuals of several fronts. A
geometric function, proposed in Ref. [5], the NSGA-II
method, is employed for this purpose. This function is
governed by a reduction factor r , 1 which leads to the
introduction of a greater diversity in the population. Let the
number of fronts to use be given by K; the total number of
individuals in the population be N; then the number of
individuals ni in each front i is given by Eq. (7)
ni ¼ N1 2 r
1 2 rKri21 ð7Þ
The mating individuals, in each front, are selected by the
Clustering Method [6] and a tournament selection based in
ranking position is used.
5.4. Population check and new descendants
After crossover and mutation, it is verified if the
descendants already exist in the population and, if they
do, they are eliminated to prevent unnecessary calculations.
The new descendants are finally obtained.
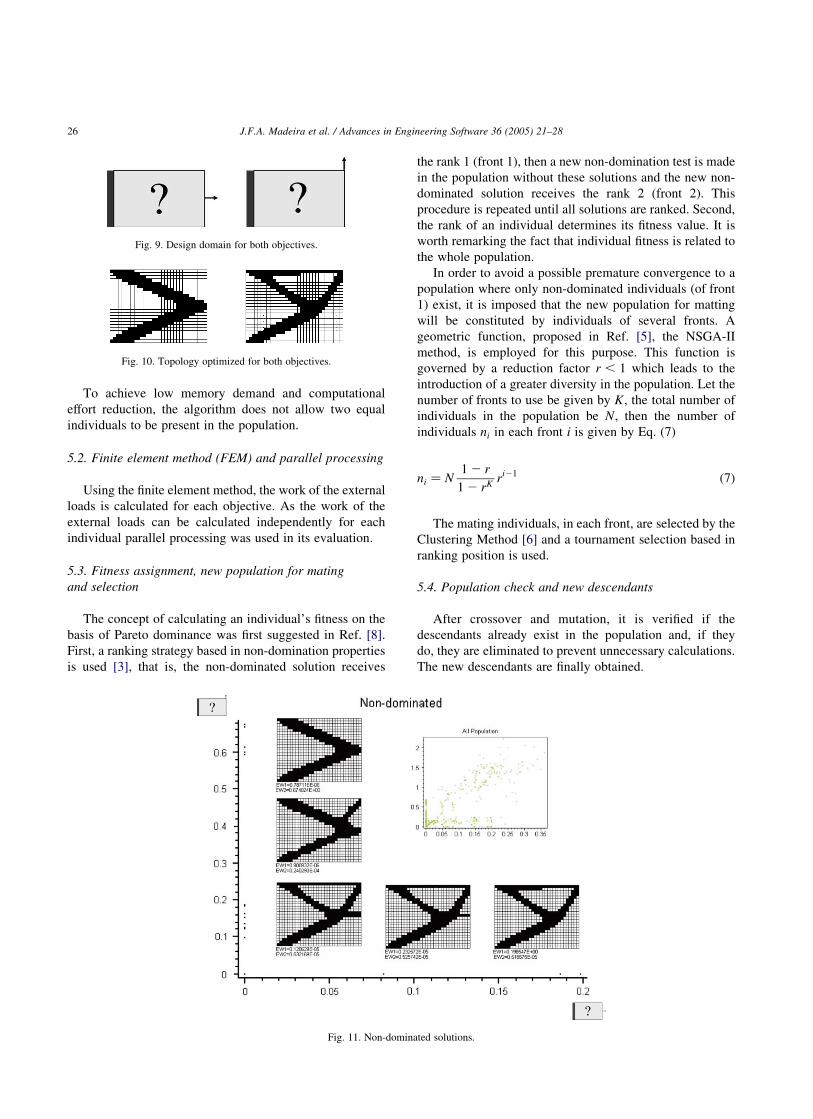
Fig. 9. Design domain for both objectives.
Fig. 10. Topology optimized for both objectives.
Fig. 11. Non-dominated solutions.
J.F.A. Madeira et al. / Advances in Engineering Software 36 (2005) 21–2826
6. Examples
To test the model, two numerical examples were solved.
The optimization problem, for these two numerical
examples, is:
Minimize fðxÞ ¼ ðf1ðxÞ; f2ðxÞÞ; ð8Þ
where x ¼ ðx1; x2Þ; f1ðxÞ and f2ðxÞ are the objective
functions, representing the compliance (or external work
(EW)) for different boundary tractions, subjected to the
volume constraint (5).
In the subsequent examples, the domain has 40 £ 30 cm
and is discretized with 768 (24 £ 32) isoparametric Q8
plane stress finite elements.
The unit cell material is assumed isotropic with Young’s
modulus equal to 210 £ 109 Pa and Poisson coefficient
equal to 0.3.
The material volume is approximately 30% of the total
design volume as a result of employing a chromosome of
length 768 bits with 236 ‘ones’ and 532 ‘zeros’. The explicit
introduction of the volume equality constraint in the
chromosome de_finition lead in the examples to a reduction
of the search possibilities from <10232 to <10205, a non
negligible improvement in relative terms. The new
crossover and mutation operators were designed to exploit
this advantage.
6.1. Example 1
For the present example, domain geometry and boundary
conditions are shown in Fig. 9.
Loading conditions for both of the two objectives are also
presented in the same figure. Two load cases are considered
in the present application, for each one of the objectives:
1. A point load in the x direction with a magnitude of
200 N, for the first objective.
2. A point load in the y direction with a magnitude of
200 N, for the second objective.
Initially, the proposed single-objective problems were
solved, and the resulting solutions, for each one of the
objectives are, respectively (Fig. 10):
These single-objective optimal solutions were introduced
in the initial population. The initial population was
composed of 200 individuals (in each generation only 100
individuals were selected for matting). The solutions
obtained after 600 generations are presented in Fig. 11.
To assess the influence of the introduction of single
objective solutions in the initial population, this example
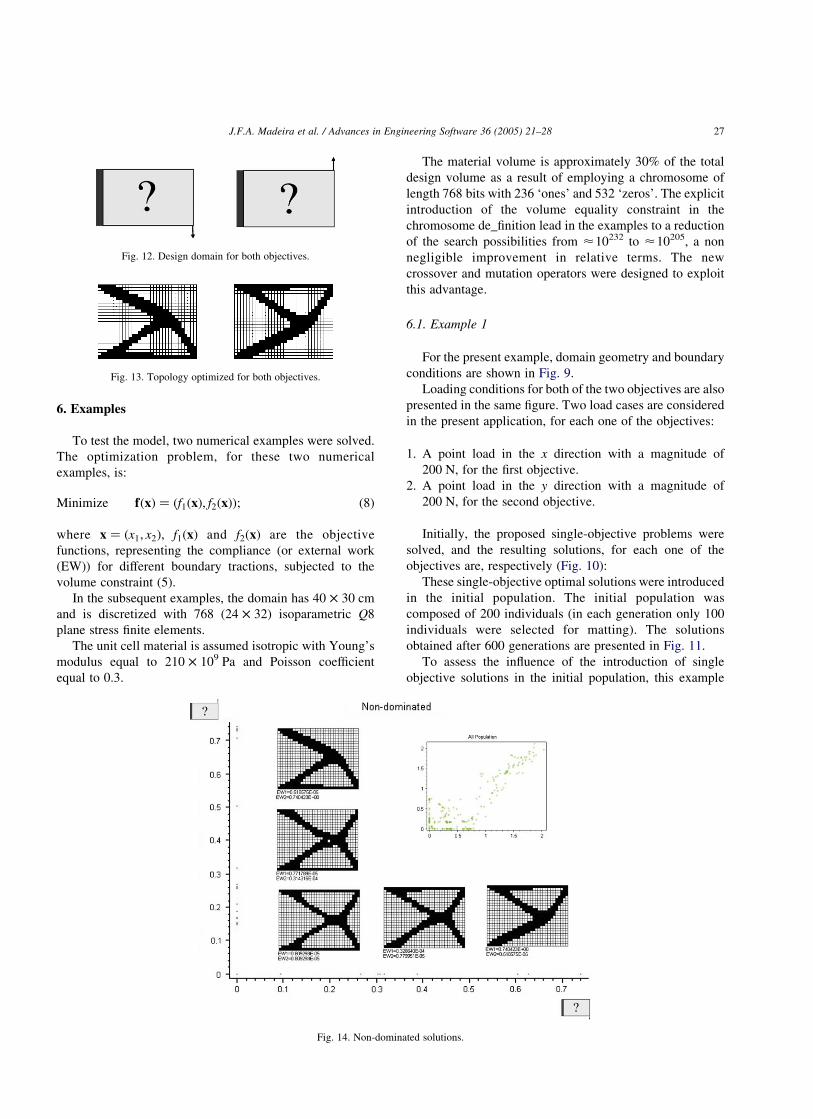
Fig. 13. Topology optimized for both objectives.
Fig. 12. Design domain for both objectives.
Fig. 14. Non-dominated solutions.
J.F.A. Madeira et al. / Advances in Engineering Software 36 (2005) 21–28 27

was tested with two different initial populations: (1) totally
random and (2) with single objective solutions introduced.
With a chromosome of length 128 bits (128 isoparametric
Q8 plane stress finite elements) and a volume constraint of
30% (52 ‘ones’) the results obtained at generation 15, with
single objective solutions introduced in the initial popu-
lation, were better than those obtained with a totally random
initial population at generation 500. This benefit improves
with problem size and thus this feature was retained in all
subsequent analysis.
6.2. Example 2
For the present example, the domain geometry and
boundary conditions are shown in Fig. 12.
Loading conditions for both of the two objectives are also
presented at the same figure. Two load cases are considered
in the present application, for each one of the objectives:
1. A point load in the y direction with a magnitude of
2200 N, for the first objective.
2. A point load in the y direction with a magnitude of
200 N, for the second objective.
Initially, the proposed single-objective problems were
solved, and the resulting solutions, for each one of the
objectives are, respectively (Fig. 13):
These single-objective optimal solutions were intro-
duced in the initial population. The initial population
was composed of 200 individuals (in each generation
only 100 individuals were selected for matting). The
solutions obtained after 600 generations are presented in
Fig. 14.
7. Conclusion and future work
A new method was presented and applied to a special
type of chromosome which was defined and for which it was
necessary to create new operators of crossover and
mutation.
To reduce computational effort, a priori optimal solutions
for each of the single-objective problems were introduced in
the initial population.
The volume constraint is automatically verified due to
the chromosome information structure and to the special
crossover and mutation operators developed.
In the future we intend to extend the present algorithm to
the three-dimensional case, to compare the solutions so
obtained with those available from classic methodologies
and to develop new crossover and mutation operators
applicable to this type of problems.
Acknowledgements
This work was supported by Portuguese Foundation for
Science and Technology through: Project POC-
TI/EME/43531/99. Partial support from project RTA P129
is also gratefully acknowledge.
References
[1] Chapman CD, Saitou K, Jakiela MJ. Genetic Algorithms as an
Approach to Configuration and Topology Design. ASME J Mech Des
1994;.
[2] Cohon JL. Multiobjective programming and planning. Math Sci
Engng, vol. 140. London: Academic Press, Inc; 1978.
[3] Deb K. Evolutionary algorithms for multi-criterion optimization in
engineering design. Proceedings of Evolutionary Algorithms in
Engineering and computer Science, (EUROGEN’99); 1999.
[4] Deb K, Pratap A, Agrawal S, Meyarivan T. A fast elitist non-
dominated sorting genetic algorithm for multi-objective: NSGA-II.
Proceedings of the Parallel Problem Solving from Nature VI
conference 2000;846–58.
[5] Deb K, Goel T. Controlled elitist non-dominated sorting genetic
algorithms for better convergence. Evolutionary multi-criterion
optimization, EMO, Berlin: Springer; 2001. pp. 67–81.
[6] Deb K. Multi-objective optimization using evolutionary algorithms.
London: Wiley; 2002.
[7] Fonseca CM, Fleming PJ. Genetic algorithms for multi-objective
optimization: formulation, discussion, and generalization. Proceed-
ings of the Fifth International Conference on Genetic Algorithms
1993;416–23.
[8] Goldberg DE. Genetic algorithms in search, optimization, and
machine learning, reading. Massachusetts: Addison Wesley; 1989.
[9] Hajela P. Genetic algorithms in structural topology optimization,
topology design of structures. NATO Advanced Research Workshop,
Sesimbra, Portugal; 1992.
[10] Holland J. Adaptation in natural and artificial systems. Ann Arbor:
University of Michigan Press; 1975.
[11] Horn J, Nafploitis N, Goldberg DE. A niched Pareto genetic algorithm
for multi-objective optimization. In proceedings of the First IEEE
Conference on Evolutionary Computation, IEEE World Congress on
Computational Intelligence, 1, 1994, 82–87.
[12] Knowles J, Corne D. The Pareto archived evolution strategy: a new
baseline algorithm for multi-objective optimization. Proceedings of
the Congress on Evolutionary Computation, Piscataway, NewJersey:
IEEE Service Center; 1999. pp. 98–105.
[13] Miettinen KM. Nonlinear multiobjective optimization. Norwell,
Massachusetts 02061 USA: Kluwer Academic Publishers; 1999.
[14] Pareto V. Cours D’Economie Politique, Vol. I and II, Rouge F.,
Lausanne; 1896.
[15] Schaffer JD. Multiple objective optimization with vector evaluated
genetic algorithms. PhD Thesis, Vanderbilt University; 1984.
[16] Schaffer JD. Multiple objective optimization with vector evaluated
genetic algorithms and their applications. Proceedings of the First
International Conference on Genetic Algorithms; 1985. pp. 93–100.
[17] Srinivas N, Deb K. Multi-objective funcion optimization using non-
dominated sorting genetic algorithms. Evol Comput 1995;(2):
221–48.
[18] Zitzler E, Theiele L. Multiobjective evolutionary algorithm: a
comparative case study and the strength Pareto approach. IEEE
Trans Evol Comput 1999;3(4):257–71.
J.F.A. Madeira et al. / Advances in Engineering Software 36 (2005) 21–2828



![A NEW HYBRID ALGORITHM FOR TOPOLOGY …ijoce.iust.ac.ir/article-1-220-en.pdf · genetic algorithms [8 and 9], particle swarm optimization [10], simulated annealing algorithms [11],](https://img.pdfslide.net/doc/110x75/5aa9c8da7f8b9a72188d5d61/a-new-hybrid-algorithm-for-topology-ijoceiustacirarticle-1-220-enpdfgenetic.jpg)








![Topology optimization of acoustic-structure interaction ... print.pdf · in [13] topology optimization using genetic algorithms, was applied to acoustic-structure interaction problems](https://img.pdfslide.net/doc/110x75/5e3ded29a3623257be6a2ce3/topology-optimization-of-acoustic-structure-interaction-printpdf-in-13.jpg)