Embed Size (px)
Citation preview

Muscling in on the Cause of Tying-Up
Stephanie J. Valberg, DVM, PhD, Diplomate ACVIM, ACVSMR
Author’s address: Professor, Director, University of Minnesota Equine Center, St. Paul, MN 55108;e-mail: [email protected]. © 2012 AAEP.
1. Introduction and History of Tying-Up
Horses are supreme athletes whose beauty and per-formance depends on their powerful musculature.Through careful genetic selection, equine breedershave shaped muscle form and function to producebreeds with muscles fine-tuned for extreme endur-ance, an elegant piaffe, or a bold stretch across thefinish line. Along with selection for positive traits,deleterious traits can also be inadvertently incorpo-rated into the genome. Even minor or intermittentperturbations in muscle function can have a majorimpact in horses because they are constantly beingexercised and pushed toward maximal performance.Fortunately, muscle remains a highly plastic tissuethat is capable of adjusting in response to growth,hormonal influences, exercise, disease, and diet.Thus, horses may be able to compensate for delete-rious traits if appropriate changes in environment,diet, and training are instituted. The past 30 yearshave offered exciting discoveries with regard toequine muscle form and function, genomics, inher-ited causes of muscle disorders, and regimens tomanage these traits. These scientific advancesbuilt up on a wealth of new knowledge of equineexercise physiology, muscle histochemistry, and ge-netics. Many clinician scientists past and presenthave provided the building blocks that have led tothe discovery of specific muscle disorders in horses,the most perplexing of which has been exertional
rhabdomyolysis, also called tying-up. Horses withtying-up develop a shortened, stiff stride, anxiety,pain, and eventually inability to move forward withlight exercise. In this review, I hope to pay tributeto these contributions, to provide insight into theadvances in equine exercise physiology, muscle bi-opsy, and genomics that led to the discovery ofequine muscle disorders, as well as provide an over-view of the current state of knowledge of exertionalrhabdomyolysis in horses.
History of Exertional Rhabdomyolysis
The dogmas of the quiet past are inadequate to thestormy present. The occasion is piled high with dif-ficulty, and we must rise with the occasion. As ourcase is new, so we must think anew and act anew.
—Abraham Lincoln
Exertional rhabdomyolysis (ER) in horses is a cen-turies-old condition that is well described in theliterature of the 19th century. In 1883, the clinicalsigns of ER, or as it was then termed “azoturia,”were described as “sweating and trembling, scarcelyable to turn in the stall, the muscles of the back andloins in a state of spasm, tail quite stiff.”1 Azoturiawas known to develop in “animals rather handsome,well-shaped, and good thrivers, than others differ-ently constituted.”1 It occurred most often in the falland winter in horses fed legumes or corn. Further,veterinarians had already recognized that “azoturia
AAEP PROCEEDINGS � Vol. 58 � 2012 85
FRANK J. MILNE STATE-OF-THE ART LECTURE
NOTES
Orig. Op. OPERATOR: Session PROOF: PE’s: AA’s: 4/Color Figure(s) ARTNO:
1st disk, 2nd beb dainop 15 1,3-8,12,13,15-17 3353

does not, or very rarely, attacks horses roaming atlarge in the fields, whether young or old; also that inall cases it is more apt to occur under conditionssucceeding a period of rather smart or active workthat followed enforced idleness.”1
The variety of terms used for this condition overcenturies is as diverse as its proposed etiologies.Initially “hysteria,” “lumbago,” and “black water”were applied.2 In 1883, the coffee-colored urinefound with severe ER was believed to be due toexcessive nitrogenous waste and thus the term azo-turia was used.1 Veterinarians readily admitted,however, that “in our day azoturia is freely spoken ofas something new or strange, but it has apparentlybeen well-enough recognized by many who have pre-ceded us.”1 “Holiday disease” or “Monday morningdisease” described the common occurrence of ERafter a day or more of rest.3,4 Lay terms “set fast,cording up, and tying-up” well describe the tighten-ing of muscles during episodes. “Paralytichemaglobinuria” was coined in the late 19th century,based on finding heme pigment in the urine, andthis term was replaced with “paralytic myoglobin-uria” when myoglobin was identified in urine.2,4
ER is now the most commonly used term that de-scribes the degeneration of striated skeletal musclewith exercise.
The development of ER was a common and cata-strophic occurrence during the 19th century andearly part of the 20th century before cars replaceddraft and carriage horses as a means of transporta-tion. Mortality rates as high as 50% were reportedin some clinics.2,4 Although it was an importantdisease, little was really known about its etiology,and speculation abounded. In 1917, Steffin statedthat “No one disease [ER] in the horse has beensubject to so many theories, theoretical treatmentsand hypothetical suggestions as this one.”5 Mostearly theories were based on observation. A kidneydisease was initially believed to cause ER because ofthe “black water” passed by horses and pain palpa-ble in the loins over the kidney.6 The higher inci-dence in horses rested, fed a full ration, and thenexercised led to a popular theory of nutritional in-toxication. “A disturbance or perversion of func-tion, not very well understood, produced by asuperabundant supply of a particular nutritive ma-terial, leads to the exaltation and ultimate loss offunction of particular parts of the voluntary muscu-lar system and the extensive disturbance of the ner-vous system.”1 Others proposed a streptococcalinfection as the basis for ER and thought that coldpredisposed horses to the disease.
In the early 20th century, elegant scientific studiesby Birger Carlstrom definitively established thatazoturia was a muscular disease with myoglobinuriaarising from damaged muscle.2,4 Further, Carl-strom was able to reproduce ER in draft-type horsesby feeding 3 kg of molasses during a period of restbefore exercise. Muscle biopsy samples obtainedduring an acute episode of ER had muscle glycogen
concentrations that were twice normal and musclelactate concentrations up to 3 times normal. As aresult of these well-designed studies, Carlstrom’stheory that a period of rest on a high grain dietbrought about a lactic acidosis with exercise thatcoagulated muscle tissue was pervasively held to betrue for all breeds of horses for 60 years. His ob-servations were astute precursors to the discovery ofpolysaccharide storage myopathy in horses.
As lighter breeds of horses increased in popularityin the mid 20th century, confusion abounded as towhether the milder clinical signs of “tying-up” inlighter breeds of horses had a similar basis as azo-turia in draft-type horses. Robertson reported in1886 that “when developed in lighter animals, andthose employed for fast work, the same generalcauses seem to be in operation which we have al-ready indicated as observed in agricultural horses.1
However, a higher incidence was noted in maresthan geldings and as a rule, “the female is moresusceptible of nervous excitement than the male.”Debate ensued among practitioners as to whetherthese were one or two conditions, and opinions oftenvaried based on the type of clinical practice. In1917, Steffin recognized that the development of ERvaried with circumstances, localities, environment,feeding customs, breeds, and individuals.5 Megin-nis in 1957 further substantiated the view that ty-ing-up and azoturia were different muscle disorders,noting from his practice that there were four signif-icant differences between ER in light versus draftbreeds: (1) seasonality (none in racehorses), (2) his-tory (fit racehorses worked daily unaffected by achange in ration), (3) handling (racehorses were lessseverely affected if walked during an attack, and (4)mortality (much lower in light breeds).7 Many oth-ers, however, disagreed and thought that lighterbreeds had a milder form of the same disease, azo-turia, that afflicted heavier breeds.3,8,9 One pointmost authors agreed on was that “until the physio-logic chemists can determine the changed chemistryin the metabolism of the normal and ‘tying-up’horses, prevention and treatment will be merelyguesswork.”7
From 1980 onward, there was a dramatic increasein the number of scientific and clinical investiga-tions of ER in horses. Koterba and Carlson10 es-tablished that light breeds of horses with ER did nothave a lactic acidosis but rather a hypochloremicmetabolic alkalosis. Roneus11 and Lindholm deter-mined that deficiency in Vitamin E, selenium, orboth were not probable causes of ER in Standard-breds. The proposed theory of hypothyroidismcausing ER12 was not substantiated by thyroid-re-leasing hormone stimulation tests,13 T3 activitymeasured when horses were not receiving nonsteroi-dal anti-inflammatories, or by thyroidectomy.14
Electrolyte disturbances including intracellular po-tassium depletion were also proposed to causeER.15,16 Substantial electrolyte imbalances wereapparent in serum samples from endurance horses
86 2012 � Vol. 58 � AAEP PROCEEDINGS
FRANK J. MILNE STATE-OF-THE ART LECTURE
Orig. Op. OPERATOR: Session PROOF: PE’s: AA’s: 4/Color Figure(s) ARTNO:
1st disk, 2nd beb dainop 15 1,3-8,12,13,15-17 3353

but not horses having ER with light exercise.17–19
Fractional excretion of electrolytes measured incatheterized urine samples suggested that bodystores of sodium or calcium were depleted in ERhorses, and dietary supplementation was suggestedto ameliorate episodes of ER.20,21 Subsequentstudies, however, showed that there was markedvariability in fractional excretion of electrolytesfrom day to day and horse to horse and found nodifference in total daily electrolyte excretion be-tween Thoroughbreds with ER and healthy horsesfed the same diet.22,23
A gradual recognition of the complexity of ER thataffected a wide variety of equine breeds arose fromthese studies. To delve deeper into the mystery ofER, we needed a thorough understanding of normalequine muscle responses to exercise; better diagnos-tic techniques to capture physiologic disturbances inER muscle; a better understanding of muscle pathol-ogy; and development of equine genome maps andsequencing of the equine genome to characterizepotential genetic defects. During the later part ofthe 20th century, an explosion of information anddiagnostic techniques in human neuromuscular dis-orders would prove key to unraveling tying-up.Armed with these tools, we moved from theories ofautointoxication or lactic acidosis to a more sophis-ticated understanding of multiple environmentaland genetic causes of equine ER.
II. New Discoveries
1. Muscle Form and FunctionA window into the world of equine muscle openedwhen Arne Lindholm (1974)24 and David Snow(1976)25 adapted the percutaneous needle biopsytechnique for use in horses. A modified 6-mm Ber-grstrom needle (Fig. 1) made it possible to repeat-edly sample unsedated horses and characterize theproperties of equine muscle. The gluteus mediusmuscle was frequently studied in horses because ofits major role in equine locomotion. It became pos-
sible to integrate knowledge from other species withequine studies to obtain a clear picture of the struc-ture and function of equine muscle. Our under-standing was also greatly enhanced when high-speed treadmills were introduced for horses andmetabolic responses to standardized exercise werecaptured by obtaining muscle biopsies before, dur-ing, and immediately after various intensities anddurations of exercise.19 The remarkably athleticability of the horse was apparent, as was the diver-sity of muscle characteristics in different breeds.
Gross Arrangement of Myofibers
Locomotor muscles are supplied by motor nervesthat branch to enervate a set of myofibers. Thenumber of myofibers enervated by a single motornerve varies, depending on the design of the musclefor dexterity (fewer fibers per motor unit) or power(many fibers per motor unit). Motor neurons differin their rates of discharge, with fast-contractingmyofibers supplied by fast-discharging phasic motorneurons and slow-contracting myofibers supplied bytonic motor neurons with a slow discharge rate.For efficiency, myofibers are grouped within themuscle such that the slower-contracting fibers usedfor postural support are frequently located deeper inthe muscle and the faster-contracting fibers used forhigher speeds are located more superficially.26
In most locomotor muscles, however, the distribu-tion of fiber types is always in a mosaic pattern, withvarying percentages of fast and slow fibers along thedepth and length of the muscle.
Structure of a Myofiber
Myofibers possess a number of structural adapta-tions that confer the ability to generate forcethrough contraction. These include a cell mem-brane capable of propagating an electrical potential,a precise alignment of contractile proteins, and in-ternal membrane structures and energy-generatingpathways that can regulate the amount of calcium
Fig. 1. Percutaneous needle biopsy. A, Three components, the outer 6-mm-diameter needle with a cutting window, the internalcutting piece that is used to slice a core of muscle tissue, which then moves into its hollow core, and the smallest-diameter piece usedto express the muscle tissue after the biopsy. B, Dynamic picture showing how a biopsy is obtained from a standardized site in thegluteus medius muscle.
AAEP PROCEEDINGS � Vol. 58 � 2012 87
FRANK J. MILNE STATE-OF-THE ART LECTURE
F1
Orig. Op. OPERATOR: Session PROOF: PE’s: AA’s: 4/Color Figure(s) ARTNO:
1st disk, 2nd beb dainop 15 1,3-8,12,13,15-17 3353

and adenosine triphosphate (ATP) available for ex-citation-contraction coupling.
SarcolemmaA basement membrane surrounds muscle fibers andis directly linked to the sarcolemma. It provides ascaffold for myogenesis and myofiber regenerationas well as structural support. The sarcolemmamaintains the intracellular milieu, actively trans-ports substrates into the myofiber, serves as a dock-ing location for proteins originating in the basementmembrane and cytoskeleton, and also transmitsneural excitatory impulses that lead to muscle con-traction. Facilitated diffusion of glucose across thesarcolemmal occurs via glucose transporters(GLUT). GLUT-1 is constitutively present in thesarcolemmal and provides basal glucose uptake,whereas GLUT-4 is present in endosomes in thesarcoplasm, which migrate to the sarcolemmal whenstimulated by insulin and contraction-dependentprocesses.27 Transport of long-chain fatty acids oc-curs via translocases28 located in the sarcolemmma,whereas short- and medium-chain fatty acids enterthe myofiber by diffusion.29
The sarcolemmal properties of excitation and con-duction are largely due to the presence of mem-brane-spanning, ion-conducting pathways thatregulate the selective and nonselective conductanceof sodium, potassium, calcium, and chloride. Volt-age-gated channels contain additional voltage-sens-ing transmembrane domains and are essential forthe generation and modification of action potentials.Sodium/calcium exchangers and ligand-gated ionchannels set myoplasmic calcium concentrations.Tubular invaginations of the sarcolemma called T-tubules project deep within each myofiber at regularintervals perpendicular to the cell length. The T-tubule membranes contain numerous voltage-gated
calcium channels called dihydropyridine receptors(DHPR) and serve to transmit electrical impulsesinto the interior of the myofiber, where they canalmost simultaneously initiate myofibrillarcontraction.
Sarcoplasmic ReticulumThe sarcoplasmic reticulum (SR) is physically sepa-rate from the sarcolemma and surrounds each myo-fibril in a highly repeating pattern (Fig. 2). The SRmembranes contain a high concentration of calciumATPase, the protein calsequestrin, and the calciumrelease channel called the ryanodine receptor (RYR).This system of membranes sequesters calcium in therelaxed muscle fiber, leaving extremely low con-centrations in the sarcoplasm surrounding themyofibrils.
Excitation-Contraction CouplingExcitation-contraction coupling is the transforma-tion of depolarizing events in the sarcolemma intothe initiation of mechanical shortening of the myo-fibrils. The action potential that is propagated intothe depths of the myofiber via transverse T-tubulestriggers the voltage-gated DHPR located within thetriads. Activation of the DHPR triggers release ofcalcium ions from the terminal cisternae into thesarcoplasm by opening the RYR in the SR mem-brane (Fig. 3). This elevates the calcium ion con-centration surrounding the myofilaments in thesarcoplasm from 10�7 to 10�5 M.
Calcium released into the sarcoplasm binds totroponin, resulting in tropomyosin moving deeperinto the groove of the actin helix and exposing themyosin binding site. Once revealed, the globularhead of myosin forms a cross-bridge with actin atthis binding site, which activates myosin ATPaseand releases ATP. Relaxation of myofibrils occurs
Fig. 2. Section of myofibrils within a muscle fiber showing the strategic alignment of myofilaments that form a contractile unit calledthe sarcomere and the inversion of the sarcolemma to form T-tubules that juxtapose with the terminal cisternae of the sarcoplasmicreticulum (SR). Available at http://www.answers.com/topic/skeletal-muscle#ixzz1kxVua6pE.
88 2012 � Vol. 58 � AAEP PROCEEDINGS
FRANK J. MILNE STATE-OF-THE ART LECTURE
F2
F3
Orig. Op. OPERATOR: Session PROOF: PE’s: AA’s: 4/Color Figure(s) ARTNO:
1st disk, 2nd beb dainop 15 1,3-8,12,13,15-17 3353

through active transport of calcium ions into thelumen of the SR by the SR calcium-ATPase(SERCA).
Perturbations in the regulation of excitation-con-traction coupling can lead to degeneration of musclefibers through elevation in sarcoplasmic calciumconcentrations. Although some excessive calciumcan be sequestered by the mitochondria, eventuallymitochondria become overloaded, and oxidative me-tabolism ceases; oxygen free radicals are generated;phospholipases are activated, inducing the arachi-donic cascade; calcium-dependent proteases arestimulated; and complement is activated all leadingto myofiber degeneration.
Contractile ProteinsThe ability of skeletal muscle to shorten is conferredby a strategic alignment of contractile proteinscalled myofilaments that form a myofibril. The re-peating unit of myofilaments within the myofibrilis referred to as a sarcomere, the fundamental unitof contraction. Muscle contractions occur when,within each sarcomere, thin myofilaments slide overthe thick myofilaments, bringing consecutive ends ofthe sarcomere closer together. Thick myofilamentsconsist of two myosin heavy chains (MHC) arrangedin a double helix. At one end, each heavy chainforms a globular head that has binding sites for bothactin and ATP, as well the enzyme ATPase. Thinmyofilaments consisting primarily of actin have acomplementary binding site for the myosin globular
head. The interaction between myosin globularheads and actin is regulated by tropomyosin andtroponin.
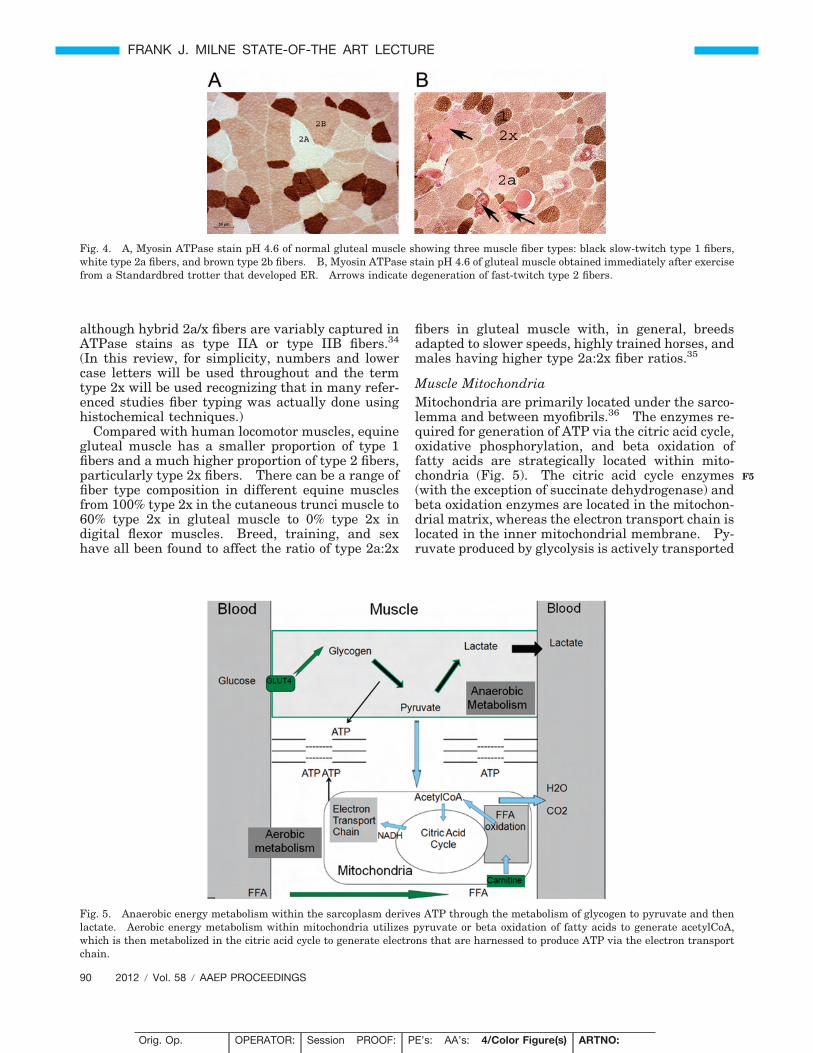
The composition of myosin heavy chains withinsarcomeres varies between individual myofibers andconfers a range of speed of contraction. Originallyequine muscle fiber types were distinguished byATPase stains, which relied on the distinct sensitivityof myosin ATPase enzyme in different fiber types tovariations in pH.24,30 Preincubation of equine muscleat pH 4.6 before ATPase staining reveals three fibertypes, darkly stained slow-twitch type I fibers, lightlystained fast-twitch type IIA fibers, and intermediatestaining fast-twitch type IIB fibers (Fig. 4). (Romannumerals and capitals are used here to distinguishfiber typing using histochemical techniques.) Immu-nohistochemical techniques that use antibodies di-rected against various MHC isoforms also have beendeveloped for fiber typing.31 (Numbers and small-case letters are used here to distinguish fiber typingusing immunohistochemistry.)
Equine StudiesEquine skeletal myofibers may express the followingdistinct MHC isoforms: perinatal (or neonatal),slow, fast-type 2a, fast-type 2x (or 2d), or a hybrid of2a/2x.32,33 The speed of contraction of these myosinheavy-chain isoforms increases in the order listedabove. The gluteal muscle contains type 1, type 2a,and 2x fibers as well as hybrid 2a/x fibers. In gen-eral, type IIB fibers correspond to type 2x fibers,
Fig. 3. Normal: Left-hand section shows the normal course of excitation-contraction coupling where depolarization of the cellmembrane activates the dihydropyridine receptor (DHPR), which triggers release of calcium ions from the terminal cisternae into thesarcoplasm by opening the ryanodine receptor (RYR) in the terminal cysternae of the SR membrane. This raises myoplasmic calciumconcentrations and causes contraction of the myofilaments. Calcium is actively resequestered in the SR through the calcium pump(SERCA), which utilizes ATP and causes relaxation of the contraction. The right-hand section shows how abnormal release of calciumvia the RYR1 can raise myoplasmic calcium excessively, resulting in uptake by mitochondria, interruption of energy metabolism,muscle contracture without relaxation, and activation of lipases, which destroy muscle cell membranes.
AAEP PROCEEDINGS � Vol. 58 � 2012 89
FRANK J. MILNE STATE-OF-THE ART LECTURE
F4
Orig. Op. OPERATOR: Session PROOF: PE’s: AA’s: 4/Color Figure(s) ARTNO:
1st disk, 2nd beb dainop 15 1,3-8,12,13,15-17 3353
although hybrid 2a/x fibers are variably captured inATPase stains as type IIA or type IIB fibers.34
(In this review, for simplicity, numbers and lowercase letters will be used throughout and the termtype 2x will be used recognizing that in many refer-enced studies fiber typing was actually done usinghistochemical techniques.)
Compared with human locomotor muscles, equinegluteal muscle has a smaller proportion of type 1fibers and a much higher proportion of type 2 fibers,particularly type 2x fibers. There can be a range offiber type composition in different equine musclesfrom 100% type 2x in the cutaneous trunci muscle to60% type 2x in gluteal muscle to 0% type 2x indigital flexor muscles. Breed, training, and sexhave all been found to affect the ratio of type 2a:2x
fibers in gluteal muscle with, in general, breedsadapted to slower speeds, highly trained horses, andmales having higher type 2a:2x fiber ratios.35
Muscle MitochondriaMitochondria are primarily located under the sarco-lemma and between myofibrils.36 The enzymes re-quired for generation of ATP via the citric acid cycle,oxidative phosphorylation, and beta oxidation offatty acids are strategically located within mito-chondria (Fig. 5). The citric acid cycle enzymes(with the exception of succinate dehydrogenase) andbeta oxidation enzymes are located in the mitochon-drial matrix, whereas the electron transport chain islocated in the inner mitochondrial membrane. Py-ruvate produced by glycolysis is actively transported
Fig. 4. A, Myosin ATPase stain pH 4.6 of normal gluteal muscle showing three muscle fiber types: black slow-twitch type 1 fibers,white type 2a fibers, and brown type 2b fibers. B, Myosin ATPase stain pH 4.6 of gluteal muscle obtained immediately after exercisefrom a Standardbred trotter that developed ER. Arrows indicate degeneration of fast-twitch type 2 fibers.
Fig. 5. Anaerobic energy metabolism within the sarcoplasm derives ATP through the metabolism of glycogen to pyruvate and thenlactate. Aerobic energy metabolism within mitochondria utilizes pyruvate or beta oxidation of fatty acids to generate acetylCoA,which is then metabolized in the citric acid cycle to generate electrons that are harnessed to produce ATP via the electron transportchain.
90 2012 � Vol. 58 � AAEP PROCEEDINGS
FRANK J. MILNE STATE-OF-THE ART LECTURE
F5
Orig. Op. OPERATOR: Session PROOF: PE’s: AA’s: 4/Color Figure(s) ARTNO:
1st disk, 2nd beb dainop 15 1,3-8,12,13,15-17 3353

across the inner mitochondrial membrane and intothe matrix, where it is oxidized and combined withcoenzyme A. The citric acid cycle oxidizes theacetyl-CoA to carbon dioxide, and, in the process,produces reduced cofactors that are a source of elec-trons for the electron transport chain. The redoxenergy is transferred to oxygen in several steps, andthe electrochemical gradient this produces acrossthe inner mitochondrial membrane is coupled to thegeneration of ATP.
Metabolic Properties of Equine Muscle Fiber TypesMuscle fibers have characteristic oxidative and gly-colytic properties (Table 1). These are determinedbiochemically by measuring the activities of citricacid cycle enzymes such as citrate synthase orsuccinic dehydrogenase and glycolytic enzymes suchas lactate dehydrogenase or subjectively determinedby histochemical staining with nicotinamide ade-nine dinucleotide tetrazolium reductase (NADH-TR).37–40 Across species, type 1 fibers have lowerglycogen, lower glycolytic capacity, and higher lipidand oxidative capacity relative to type 2 fibers.Type 2a fibers have intermediate properties,whereas type 2x fibers have higher glycogen, higherglycolytic, and lower lipid and oxidative capacitythan the other fiber types. It is important to note,however, that the metabolic properties of fibers canvary widely, especially with training.40–42 For ex-ample, type 2x fibers in some trained Thoroughbredshave a greater oxidative capacity than type 1 fibersin an untrained horse.43,44
Responses to Exercise
Muscle Fiber RecruitmentFor smooth, coordinated locomotion, muscles are re-cruited in an orderly manner. Within a particularmuscle, myofibers are selectively recruited in a spe-cific pattern that varies according to the gait, speed,and duration of exercise. This occurs through thedifferential stimulation of alpha motor neurons ofspecific types, in line with the size principle. Thesmallest-diameter motor neurons, which have thelowest threshold, innervate the type 1 fibers,
whereas the largest innervate the type 2x fibers.During low-intensity submaximal exercise, type 1and 2a fibers are recruited; however, if extremelyprolonged low-intensity exercise is performed, type2x myofibers may eventually be recruited. As thespeed of exercises increases, muscle fibers are re-cruited in the order of type 1, type 2a, and type2x.45–47
Energy Metabolism With Submaximal ExerciseMuscle glycogen concentrations fall steadily in type1 and 2a fibers during submaximal exercise as aresult of metabolism of glycogen to pyruvate, oxida-tion of pyruvate in the citric acid cycle, and closecoupling with the electron transport chain to pro-duce ATP.45 High rates of pyruvate oxidation in-hibit phosphofructokinase (PFK) and slow glucoseutilization in favor of free fatty acid (FFA) oxidation.Thus, as exercise progresses, the fall in glycogen isattenuated by oxidation of FFAs.19,48,49 A drop inplasma insulin and rise in cortisol with exercisefacilitates release of FFA from adipose tissue, in-creasing their availability within 15 minutes of ini-tiating submaximal exercise.19 Long-chain FFAsare actively transported into the myofiber by trans-locases and utilize a carnitine transport shuttle totraverse the inner mitochondrial membrane. Car-nitine also serves to buffer excess fatty acid CoAmoieties that may be formed. Beta oxidation ofFFA is an essential process for prolonged exerciseand spares the finite stores of glycogen with skeletalmuscle. Amino acids may also serve as energy sub-strates within skeletal muscle under conditions ofrestricted energy intake or prolonged exercise.50
Fatigue usually occurs with extended submaximalexercise as a result of dehydration, electrolyte deple-tion, hyperthermia, glycogen depletion, or centralfatigue.
Equine StudiesUsing tissue snap-frozen in liquid nitrogen, Lind-holm,24 Snow,25 and many other subsequent re-searchers captured metabolic activity of equinemuscle exactly as it was at rest or when sampled
Table 1. Characteristics of Contractile Myofiber Types of Skeletal Muscle in Untrained Horses
Type 1 Type 2a Type 2x
Fiber size (cross-sectional area/standard depth) Smallest Intermediate LargestSpecific ATP activity (alkaline pH) Low High HighSpecific ATP activity (pH 4.6) High Low IntermediateSpeed for contraction Low Intermediate HighPower output Low Intermediate HighMuscle fibers per motor unit Low High HighResistance to fatigue High Intermediate LowOxidative capacity Low Intermediate HighLipid content High Intermediate LowGlycolytic capacity High Intermediate LowGlycogen content Intermediate High High
AAEP PROCEEDINGS � Vol. 58 � 2012 91
FRANK J. MILNE STATE-OF-THE ART LECTURE
T1
Orig. Op. OPERATOR: Session PROOF: PE’s: AA’s: 4/Color Figure(s) ARTNO:
1st disk, 2nd beb dainop 15 1,3-8,12,13,15-17 3353

after various intensities of exercise. The changesin glycogen, ATP, inosine monophosphate (IMP),and lactate concentrations that occurred with exer-cise were completely dependent on the speed, dura-tion, training level, and muscle fiber typecomposition of the horse.19,39,51–55 The informationderived from these studies of normal muscle metab-olism was integral to compare subsequent studies ofmuscle metabolism in horses with ER.
In contrast to humans, horses were found to havehigher resting glycogen concentrations, rangingfrom 450 to 650 mmol/kg dry weight (dw) (human,300 to 400 mmol/kg), with the highest concentra-tions found after long term training35,42,56,57 Fur-ther, in contrast to humans, muscle glycogenconcentrations in healthy horses were found to beresistant to carbohydrate loading and slow to repleteafter exercise.58 Resting equine muscle lactate(equine, 24 to 40 mmol/kg) and ATP (22 to 28mmol/kg dw) concentrations were slightly lowerthan in humans, and resting IMP concentrations inboth species were found to be �0.25 mmol/kg dw.Submaximal exercise studies in horses lasting 90minutes showed a decline of glycogen in the range of25% to 40%, depending on the speed of exercise,whereas repeated bouts of submaximal and sprintexercise at 100% maximum oxygen uptake over 3days produced glycogen depletion of 50%,59 and de-pletion was pronounced at 56% after 80-km endur-ance rides.60
Energy Metabolism With Near-Maximal ExerciseIntensity
For a few seconds of high-speed exercise, creatinephosphate (CP) and ATP serve as small reserves ofimmediate energy until ATP production fromglyocogenolysis begins. The rapid rate of energygeneration required and potentially limited oxygensupply at high speeds drives the conversion of gly-cogen to pyruvate to lactate via anaerobic glycolysis.It is important to note that anaerobic glycolysis isnot the sole source of energy with high-speed exer-cise. It produces energy over and above that simul-taneously being supplied by oxidation of pyruvate inmitochondria. As the muscle reaches its maximumability to generate ATP from oxidative metabolism(at point of maximum oxygen uptake), all three mus-cle fiber types are recruited and anaerobic glycolysisproduces an exponential rise in lactate.45,51 Withmaximal exertion, ATP also may be generated bythe myokinase reaction utilizing adenosine diphos-phate (ADP), produced by the hydrolysis of ATP.61
This metabolic process is facilitated by removingadenosine monophosphate (AMP) to prevent enzymeinhibition accomplished through deamination ofAMP to IMP.62 Fatigue with maximal exertionmay occur as a result of hydrogen ion accumulationand a drop in muscle pH, which affects myosin-actininteractions, reuptake of calcium into the sarcoplas-mic reticulum and PFK activity. In addition, ATP
depletion can occur in individual muscle fibers, caus-ing fatigue.
Equine Studies
Relative to humans, equine muscle has remarkablyhigh muscle glycogen concentrations, high oxidativeand glycolytic capacities, and a remarkable ability toproduce and withstand high lactate and low ATPconcentrations with intense exercise.35 Muscle bi-opsies obtained after near-maximal exercise over800 m show a decline in glycogen of 28% and ATP by30%, an 8-fold increase of IMP, and 9-fold increasein muscle lactate,63 dropping muscle pH to 6.4.64
Even with maximal exertion, only a proportion ofmyofibers in a gluteal biopsy are recruited and thusstudies that measure the general change in sub-strates within a muscle biopsy may underestimatechanges that occur in active fibers. Essen Gustavs-son developed an intricate technique to dissect andtype individual freeze-dried fibers from postexercisemuscle samples.44,65 Single-fiber studies revealedthat whole-muscle biopsies do not accurately reflectthe metabolic events in individual muscle fibers.Of particular note was the fact that whole-muscleATP concentrations of 18.3 � 2.7 mmol/kg dw weremeasured in Standardbreds after a race, whereasATP concentrations as low as 1 mmol/kg dw werefound in individual fibers.66 This single-fiber tech-nique would prove extremely useful for evaluatingpotential disruption in energy metabolism in ER-affected muscle.
Normally, safety mechanisms are in place inhealthy horses to prevent metabolic processes fromproceeding to the point of damaging the internalmilieu of the myofiber. These mechanisms includethe inhibition of PFK by AMP and a low pH, therebyslowing glycolysis and acidosis as well as noscio-ceptors within the muscle that create discomfortand drive a centrally mediated decrease inlocomotion.67,68
Impact of Training on Skeletal Muscle
The major training adaptation of equine muscle isan increase in aerobic capacity.53,69,70 For this ad-aptation to occur, a threshold of training intensity isrequired over a minimum training duration. Thebenefits of training for horses performing at sub-maximal aerobic exercise intensities include en-hanced delivery of oxidative substrates to musclefibers as well as improved oxidative metabolism ofglycogen and free fatty acids. Muscle strength andpower output may also be improved. Especially inendurance horses, greater free fatty acid utilizationleads to a glycogen-sparing effect and prolonged en-durance capacity. The benefits of enhanced oxida-tive capacity for horses performing maximalintensity exercise include increased capacity to gen-erate ATP aerobically, which allows horses toachieve higher speeds before the onset of lactateaccumulation and ATP depletion.71
92 2012 � Vol. 58 � AAEP PROCEEDINGS
FRANK J. MILNE STATE-OF-THE ART LECTURE
Orig. Op. OPERATOR: Session PROOF: PE’s: AA’s: 4/Color Figure(s) ARTNO:
1st disk, 2nd beb dainop 15 1,3-8,12,13,15-17 3353
These advances in exercise physiology in the laterpart of the 20th century fueled an explosion of infor-mation in the field of human and subsequentlyequine neuromuscular disorders. Specialized neu-romuscular laboratories72 were established whereneurologists began to personally evaluate frozensections of muscle biopsy specimens from their pa-tients, using histochemical and biochemical tech-niques, and closely correlate results with theirclinical evaluation.73
2. Neuromuscular Diagnostic LaboratoriesNeuromuscular diagnostic laboratories formed thecornerstone for the identification of specific etiolo-gies for ER in horses. A centralized site to receivemuscle biopsies from across North America facili-tated (1) assimilation of the astute observations oflarge numbers of practitioners submitting biopsies,(2) accumulation of hundreds of cases from which toidentify patterns of disease, (3) banking of biochem-ical and DNA samples that could be used in inves-tigating the pathophysiology of disease subsets, and(4) formation of a research hub where breeders,equine practitioners, physiologists, biochemists, andmolecular biologists could collaborate on muscle dis-eases in horses.
The first neuromuscular diagnostic laboratorythat provided routine diagnostics in veterinary med-icine was developed by Dr. Cardinet III at the Uni-versity of California, Davis, in the late 1970s. Dr.Cardinet’s training at the National Institutes ofHealth and his close collaboration with neurologistDr. Terryl Holliday provided a unique, clinicallyfocused platform to investigate muscle diseases inveterinary medicine.74 Dr. Cardinet’s laboratoryserved as the launching point for both the Compar-ative Neuromuscular Laboratory at the Universityof California, San Diego, and the NeuromuscularDiagnostic Laboratory at the University ofMinnesota.
The interpretation of biopsies at NeuromuscularDiagnostic Laboratories combined findings from
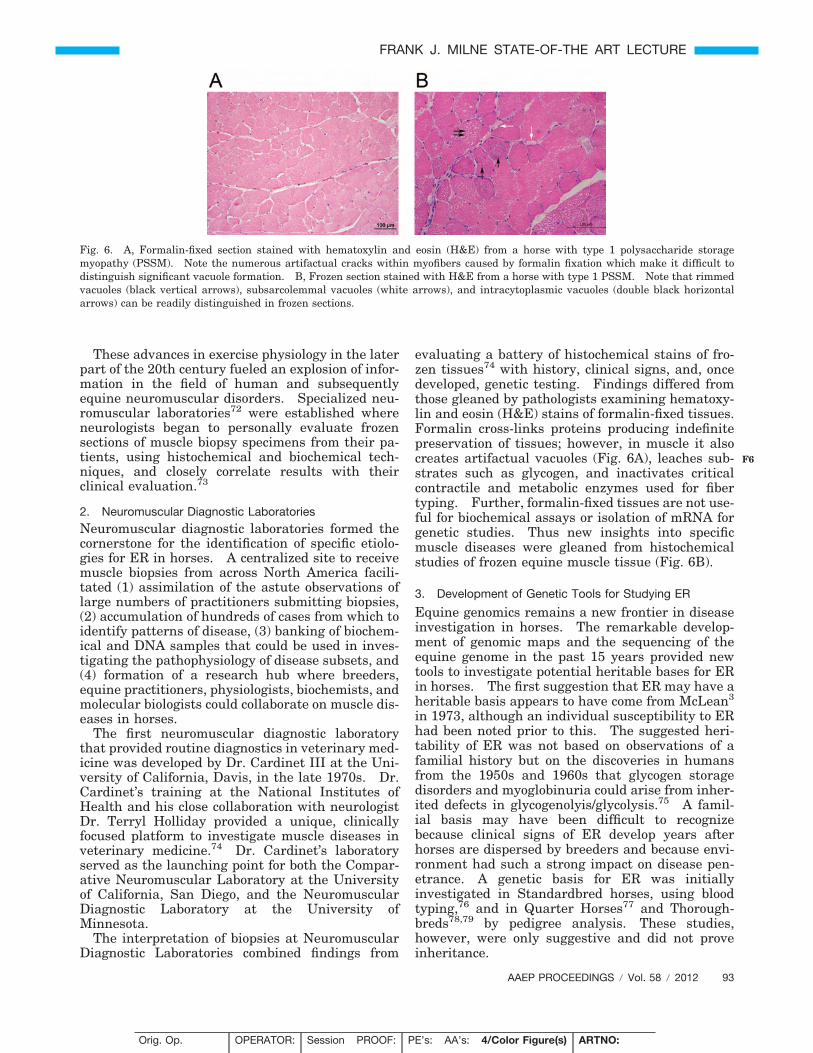
evaluating a battery of histochemical stains of fro-zen tissues74 with history, clinical signs, and, oncedeveloped, genetic testing. Findings differed fromthose gleaned by pathologists examining hematoxy-lin and eosin (H&E) stains of formalin-fixed tissues.Formalin cross-links proteins producing indefinitepreservation of tissues; however, in muscle it alsocreates artifactual vacuoles (Fig. 6A), leaches sub-strates such as glycogen, and inactivates criticalcontractile and metabolic enzymes used for fibertyping. Further, formalin-fixed tissues are not use-ful for biochemical assays or isolation of mRNA forgenetic studies. Thus new insights into specificmuscle diseases were gleaned from histochemicalstudies of frozen equine muscle tissue (Fig. 6B).
3. Development of Genetic Tools for Studying ER
Equine genomics remains a new frontier in diseaseinvestigation in horses. The remarkable develop-ment of genomic maps and the sequencing of theequine genome in the past 15 years provided newtools to investigate potential heritable bases for ERin horses. The first suggestion that ER may have aheritable basis appears to have come from McLean3
in 1973, although an individual susceptibility to ERhad been noted prior to this. The suggested heri-tability of ER was not based on observations of afamilial history but on the discoveries in humansfrom the 1950s and 1960s that glycogen storagedisorders and myoglobinuria could arise from inher-ited defects in glycogenolyis/glycolysis.75 A famil-ial basis may have been difficult to recognizebecause clinical signs of ER develop years afterhorses are dispersed by breeders and because envi-ronment had such a strong impact on disease pen-etrance. A genetic basis for ER was initiallyinvestigated in Standardbred horses, using bloodtyping,76 and in Quarter Horses77 and Thorough-breds78,79 by pedigree analysis. These studies,however, were only suggestive and did not proveinheritance.
Fig. 6. A, Formalin-fixed section stained with hematoxylin and eosin (H&E) from a horse with type 1 polysaccharide storagemyopathy (PSSM). Note the numerous artifactual cracks within myofibers caused by formalin fixation which make it difficult todistinguish significant vacuole formation. B, Frozen section stained with H&E from a horse with type 1 PSSM. Note that rimmedvacuoles (black vertical arrows), subsarcolemmal vacuoles (white arrows), and intracytoplasmic vacuoles (double black horizontalarrows) can be readily distinguished in frozen sections.
AAEP PROCEEDINGS � Vol. 58 � 2012 93
FRANK J. MILNE STATE-OF-THE ART LECTURE
F6
Orig. Op. OPERATOR: Session PROOF: PE’s: AA’s: 4/Color Figure(s) ARTNO:
1st disk, 2nd beb dainop 15 1,3-8,12,13,15-17 3353

Breeding TrialsBreeding trials in Quarter Horses80,81 and Thor-oughbreds79 were required to establish that someforms of ER are passed down through family lines.Breeding trials are ideal when the heritable natureof a trait is in question and the trait is believed to becaused by a single gene. The classic approach is tomate second-generation heterozygotes and analyzethe proportion of affected and unaffected offspring todetermine the probable pattern of inheritance.The numbers of animals required usually makessuch trials cost-prohibitive to perform in veterinarymedicine. Fortunately, however, the AmericanQuarter Horse Association and the Morris AnimalFoundation had the foresight to support breedingherds of Quarter Horses and Thoroughbreds withER that were crucial to establishing the nature ofinheritance of their specific forms of ER. Furtherevidence of the heritable nature of forms of ER inhorses awaited development of equine-specific ge-netic tools.
Candidate Gene ApproachThe first genetic approach available to equine re-searchers prior to development of equine genomemaps was the candidate gene approach. This ap-proach was first used by Dr. Sharon Spier to identifythe genetic basis for hyperkalemic periodic paralysisin Quarter Horses82 and was successfully used toinvestigate the ryanodine receptor 1 (RYR1) gene asa cause for ER by Dr. Monica Aleman.83 A candi-date gene is usually identified comparatively, whereanother species is found to have a disease with sim-ilar clinical signs and a genetic defect has alreadybeen identified in that species. Dr. Aleman recog-nized that the clinical signs she observed in twoQuarter Horses under general anesthesia wereidentical to those of pigs with malignant hyperther-mia (MH).84 Selected exons of the putative candi-date gene, the (RYR1), were sequenced from cDNAprepared from snap-frozen muscle samples fromhealthy and affected horses. Polymerase chain re-action (PCR) primers designed on the basis of knownsequence homology from other species were utilizedto generate templates to sequence. A point muta-tion in exon 46 was identified in horses with MH,and horses with this mutation were subsequentlyfound to show signs of ER.
Whole blood for isolation of genomic DNA andfrozen tissue for mRNA are ideal for candidate geneapproaches to muscle disorders. The availability ofcomplete genome sequence helps to predict intronexon boundaries and sequencing of just the codingexons by PCR from a simple DNA sample can beachieved. Sequence polymorphisms are identifiedwhen comparing normal and affected individuals.It is important to note, however, that many base-pair substitutions are of little consequence becausethey do not change the amino acid code (silent mu-tations). On the other hand, missense, nonsense,frameshift, and splice site junction mutations can all
cause altered structure, function, or synthesis of theencoded protein and potentially be causative of adisease. Roadblocks with this direct sequencingapproach include sequencing the wrong gene andmissing large structural rearrangements of a genethat result in no mRNA production or simply alteredlevels of mRNA. The larger structural rearrange-ments can, however, be detected using Southernblotting techniques on genomic DNA or large-scaleDNA sequencing.
Genome-Wide Association Mapping andSequencingThe ability to perform genome-wide mapping washighly desirable for studying ER because for mostforms of ER, no likely candidate gene were readilyapparent. It requires only the hypothesis that thedisease is inherited and the mutant genes are some-where in the genome. This approach seeks to iden-tify genetic markers closely associated with thechromosome containing the as-yet unknown affectedgene, and, through fine mapping, zero in on likelycandidate genes. The initial limiting factor for itsapplication in horses was the lack of readily avail-able equine genome maps with polymorphic mark-ers evenly spaced across the genome.
Development of equine genome maps began in the1990s, based mainly on short repetitive DNA se-quences called microsatellites, in which alleles differin the number of nucleotide repeat units. Collab-orative international efforts generated two equinegenetic linkage maps, each containing more than740 microsatellite markers.85,86 A 4,000 markerwhole-genome equine-hamster radiation hybrid RHmap including 2,000 microsatellites, and 2,000 geneloci was also developed to enable both microsatelliteand gene-based markers to be efficiently assigned toa horse genome map.87 This achieved the goal of agenome-wide coverage with an average markerspacing of less than 1 Mb and ready comparison ofthe same gene markers on the equine chromosomemaps with their homologues on the chromosomes ofother species. These microsatellite genome mapsproved useful for the whole-genome associationanalysis that identified the genetic mutation re-sponsible for one form of ER in Quarter Horses,polysaccharide storage myopathy.81 A panel ofmarkers spaced across all 31 autosomes was ini-tially screened in 48 control and 48 PSSM-affectedhorses. Associated markers were identified onequine chromosome 10 (ECA10), and reanalysis us-ing a denser panel of markers on ECA10 narrowedthe region to an area that contained an excellentcandidate gene the glycogen synthase 1 (GYS1)gene. Sequencing of this gene revealed the caus-ative genetic mutation.
Linkage AnalysisAnother approach that was also used to study thegenetic basis for ER in horses is linkage analysis.Genetic linkage analysis ideally uses multi-genera-
94 2012 � Vol. 58 � AAEP PROCEEDINGS
FRANK J. MILNE STATE-OF-THE ART LECTURE
Orig. Op. OPERATOR: Session PROOF: PE’s: AA’s: 4/Color Figure(s) ARTNO:
1st disk, 2nd beb dainop 15 1,3-8,12,13,15-17 3353

tion pedigrees with many full or half-sib offspring,whereas genetic association utilizes populations ofrelatively unrelated individuals. Like the associa-tion studies described above, a subset of the DNAmarkers from the genome map are statistically an-alyzed for co-inheritance or association with thetrait in appropriate resource populations. Geneticlinkage would indicate that the gene for the diseaseis located close to the genetic marker and thereforeit does not dissociate from the marker during ran-dom crossovers in meiosis. Due to widespread geo-graphic distribution of related horses and thedifficulties created in phenotyping and sample col-lection, it is very difficult to assemble sufficientlysized pedigrees to perform linkage analysis inhorses. This approach, however, was used in Thor-oughbred horses where samples were collectedthrough breeder cooperation and from a breedingtrial.88 Linkage analysis was used to exclude po-tential candidate genes involved in excitation-con-traction coupling as causative of ER in that cohort ofThoroughbred horses.
Single Nucleotide Polymorphism ChipsIn 2008, whole-genome sequencing of the femaleThoroughbred Twilight was achieved by scientistsat The Broad Institute under the auspices of theNational Human Genome Research Institute.89
This sequence covered 2.42 gigabytes (Gb) of the2.67 Gb horse genome and was of sufficient coverageto give almost complete, or at least partial, sequenceof all the equine genes, as well as define repetitivesequence content and nature. This sequence wasaligned with those of human and other mammaliangenomes to define syntenic segments and predict thelocations of equine genes with insufficient sequencecoverage. To enhance the ability to discover genesassociated with diseases and specific traits inhorses, a denser set of polymorphic markers wasrequired than the microsatellites identified previ-ously. These new markers were selected from theapproximately 1.5 million single nucleotide poly-morphisms (SNPs) that were identified from thesequence of Twilight and seven breed representa-tives, derived from both recently developed andancient breeds. SNPs exist on average at approxi-mately every 1,500 base pairs (bp) of horse sequence.To further facilitate equine genetic discovery, acommercial chip that contained 50,000 SNPs wasdeveloped for automated whole genome mapping.Analysis of such SNP chip genotype data processedfrom affected and control horses searches for statis-tical associations between the frequency of alleles ofSNP markers and the disease phenotype. SNPchips have been used to search for potential genescausing ER in Thoroughbreds. The University ofMinnesota research group identified a region onequine chromosome 16 that was significantly asso-ciated with ER in Thoroughbreds.90 The next stepsinvolve searching for potential candidate genes inthe relatively short genomic regions, based on pro-
posed function (basically as with candidate genesabove). If the function of the genes in the regionare known or their expression in various tissues isknown, their potential likelihood for producing thephysiologic basis for the disease helps to select genesfor subsequent sequencing. Unfortunately, despitean enormous effort in human medicine, the functionof a large fraction of all genes remains unknown,complicating selection of the appropriate gene forsequencing. It is highly likely, however, that fur-ther progress in developing tools for genome analy-sis in horses will lead to the unmasking of moregenetic mutations that cause muscle disorders inhorses.
III. Clinical Approach to Horses With ER
The advances described in the preceding sectionwere all essential for developing a modern approachto evaluating horses with ER. A thorough under-standing of muscle contraction and relaxation aswell as the documentation of metabolic responsesthat normally occur with exercise provided an idealbasis from which to compare muscle function inhorses with ER. The development of a safe musclebiopsy technique and laboratories specialized in in-terpretation of muscle disease in frozen sectionswidened the ability to identify specific diseases.Advances in diagnostic techniques for equine inter-nal medicine opened up new avenues to standardizethe clinical approach to muscle diseases. Finally,the development of genome maps and sequencing ofthe equine genome provided tools to investigate thegenetic basis for ER.
The modern approach to evaluating horses withER that arose from many previous studies is de-scribed in this section.
History
A detailed history is the foundation for evaluatingER because the disorder can be intermittent in na-ture and not evident on physical examination andalso because identifying the environmental stimulithat trigger ER is necessary to appropriately man-age the condition. A careful description of thehorse’s muscle tone, muscle mass, gait, degree ofpain, exercise intolerance, and weakness while ex-periencing clinical signs as well as the duration andfrequency of signs is important. Further, an effortto characterize possible eliciting factors such as thehorse’s temperament, diet, previous performance,exercise schedule, evidence of exercise intolerance,type and duration of exercise performed when expe-riencing episodes, mental state when episodes begin,other possible precipitating factors, and currentmedications should be made. Accumulating histor-ical information on a number of horses of a variety ofbreeds revealed that some horses seem to have anextrinsic cause for ER that was sporadic in nature,whereas in other cases, even under the best man-agement, horses will have repeated episodes of ER.
AAEP PROCEEDINGS � Vol. 58 � 2012 95
FRANK J. MILNE STATE-OF-THE ART LECTURE
Orig. Op. OPERATOR: Session PROOF: PE’s: AA’s: 4/Color Figure(s) ARTNO:
1st disk, 2nd beb dainop 15 1,3-8,12,13,15-17 3353
Thus, a subset of horses appeared to have an intrin-sic abnormality that caused chronic ER.
Physical Examination
Regardless of the cause of ER, horses with acutesigns of ER share features of muscle stiffness,sweating, firm painful muscles, elevated respiratoryrate, and reluctance to move. Even when acutesigns are not present, however, the physical exami-nation may reveal alterations in muscle mass andsymmetry or symmetrical atrophy characteristic of arecent severe acute episode of ER. Inspection of thehorse at a distance for symmetry of muscle masswhile the horse is standing with forelimbs and hindlimbs exactly square is imperative to assess musclemass, symmetry, and signs of atrophy. Subse-quently, palpation of the entire muscle mass of thehorse provides valuable information regarding mus-cle tone, heat, pain, swelling, subtle muscle atrophy,and fasciculations. Firm, deep palpation of thelumbar, gluteal, and semimembranosus and semi-tendinosus muscle may reveal pain, cramps, or fi-brosis. A lameness evaluation, including flexiontests, is often indicated because chronic lamenessmay be present94 that exacerbates ER and must betreated before normal fluid muscle function returns.Muscle pain may be secondary to changes in move-ment caused by lower limb lameness. A neurologicexamination should be performed to determine ifthis is an underlying cause of abnormal movementin which owners have attributed a gait abnormalityto ER.
Ultrasonography is potentially useful for identify-ing focal area of muscle trauma with physical dis-ruption of the muscle.105 Horses with classic signsof ER and no external evidence of muscle stiffnessmay have rhabdomyolysis within the psoas muscles,which can be identified by rectal palpation and ul-trasound. Careful comparisons must be made be-tween similar sites in contralateral limbs in bothtransverse and longitudinal images because the typ-ical striated echogenic pattern varies according tothe muscle group. The appearance of muscle is alsosensitive to the way the horse is standing andwhether the muscle is under tension, so it is impor-tant that the horse is standing squarely and bearingweight evenly. Muscle fascia appears as well-de-fined, relatively echo-dense bands. Care must betaken in identifying large vessels and artifacts cre-ated by them. In an acute injury, muscle fiber dis-ruption is seen as relatively hypoechoic areas withinmuscle, with loss of the normal muscle striation.The jagged edge of the margin of the torn muscle maybe increased in echogenicity. Tears in the musclefascia may be identified. The defect in muscle may befilled by a loculated hematoma that is slowly replacedby hypoechoic granulation tissue. Muscle repairshows a progressive increase in echogenicity. Rela-tively hyperechoic regions may develop due to fibrousscarring. Hyperechoic regions causing shadowing ar-tifacts reflect mineralization.105
Serum Creatine Kinase and Aspartate TransaminaseThe most rapid diagnosis of severe acute ER forhundreds of years has been catheterization of thebladder and examining the urine for pigmenturiathat is Hb-positive on urine stick tests in the ab-sence of intravascular hemolysis and hematuria.Diagnosis was greatly simplified by research thatshowed that elevations in serum creatine kinase(CK) and aspartate transaminase (AST) activitieswere indicative of rhabdomyolysis.106 The degreeof elevation of these enzymes in serum is dependenton the severity of muscle damage as well as thelength of time that has elapsed between samplecollection and the occurrence of muscle damage.91
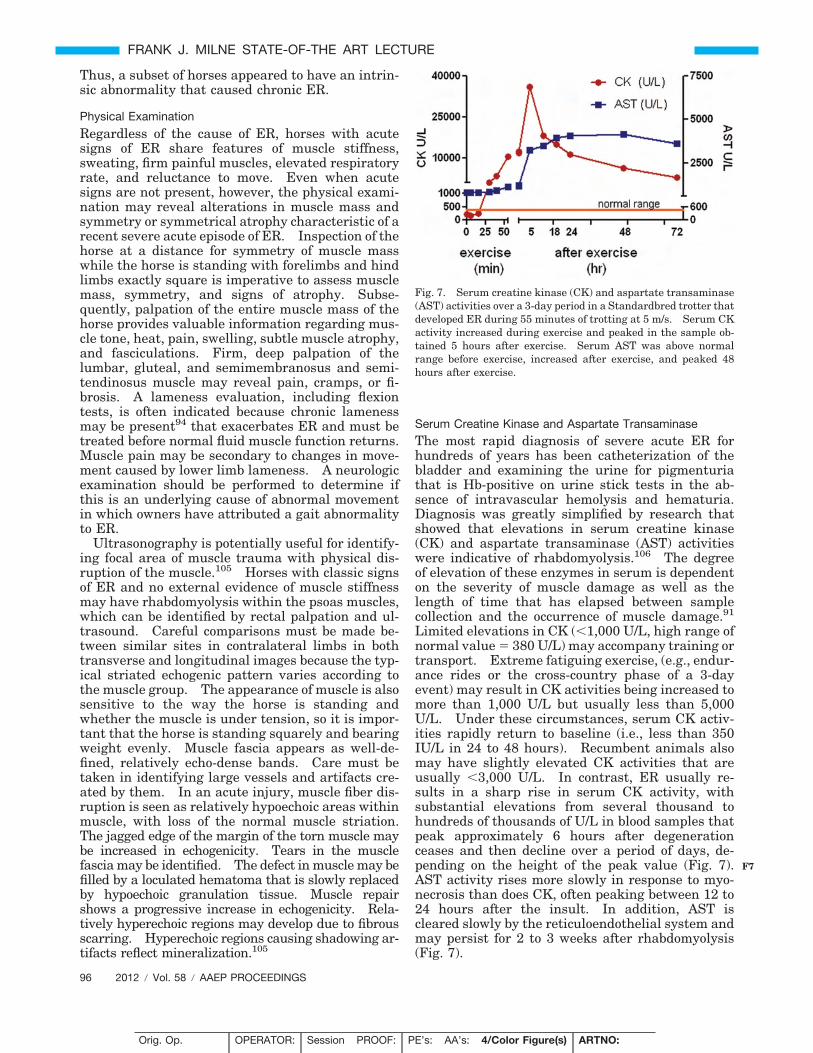
Limited elevations in CK (�1,000 U/L, high range ofnormal value � 380 U/L) may accompany training ortransport. Extreme fatiguing exercise, (e.g., endur-ance rides or the cross-country phase of a 3-dayevent) may result in CK activities being increased tomore than 1,000 U/L but usually less than 5,000U/L. Under these circumstances, serum CK activ-ities rapidly return to baseline (i.e., less than 350IU/L in 24 to 48 hours). Recumbent animals alsomay have slightly elevated CK activities that areusually �3,000 U/L. In contrast, ER usually re-sults in a sharp rise in serum CK activity, withsubstantial elevations from several thousand tohundreds of thousands of U/L in blood samples thatpeak approximately 6 hours after degenerationceases and then decline over a period of days, de-pending on the height of the peak value (Fig. 7).AST activity rises more slowly in response to myo-necrosis than does CK, often peaking between 12 to24 hours after the insult. In addition, AST iscleared slowly by the reticuloendothelial system andmay persist for 2 to 3 weeks after rhabdomyolysis(Fig. 7).
Fig. 7. Serum creatine kinase (CK) and aspartate transaminase(AST) activities over a 3-day period in a Standardbred trotter thatdeveloped ER during 55 minutes of trotting at 5 m/s. Serum CKactivity increased during exercise and peaked in the sample ob-tained 5 hours after exercise. Serum AST was above normalrange before exercise, increased after exercise, and peaked 48hours after exercise.
96 2012 � Vol. 58 � AAEP PROCEEDINGS
FRANK J. MILNE STATE-OF-THE ART LECTURE
F7
Orig. Op. OPERATOR: Session PROOF: PE’s: AA’s: 4/Color Figure(s) ARTNO:
1st disk, 2nd beb dainop 15 1,3-8,12,13,15-17 3353

By comparing serial activities of CK and AST,information is derived concerning the progression ofmuscle degeneration. Elevations in both CK andAST reflect relatively recent or active myodegenera-tion; persistently elevated serum CK indicates thatmyodegeneration is likely to be continuing. Ele-vated AST activity accompanied by decreasing ornormal CK activity indicates that myodegenerationhas ceased. The degree of elevation of CK and ASTdoes not necessarily reflect the severity of clinicalsigns, as some forms of rhabdomyolysis are morepainful than others.
Serum Chemistry
With severe rhabdomyolysis, electrolyte abnormali-ties such as hyponatremia, hypochloremia, hypo-calcemia, hyperkalemia, and hyperphosphatemiaare identified.107 These derangements result fromlosses in sweat as well as shifting of fluid and elec-trolytes (sodium, chloride, calcium) down a concen-tration gradient into damaged muscle. Release ofelectrolytes such as potassium and phosphorus fromdamaged muscle cells can result in increased serumconcentrations. A metabolic alkalosis was found tobe the most common acid base abnormality with ER,as a compensation for hypochloremia.10 Lactic ac-idosis is rarely, if ever, observed.10,108 Azotemiahas been identified in dehydrated horses from myo-globinuric nephrotoxicity. Azotemia is much lesscommon in horses compared with humans becausetheir alkaline urine provides some protection frommyoglobin precipitation.
Exercise Response Test
An exercise response test proved to be useful forevaluating horses with a possible history of ER thathave normal serum CK and AST activity at the timeof evaluation. Blood samples are taken before ex-ercise and about 4 to 6 hours after a light exercisetest to evaluate peak changes in CK activity. Asubmaximal exercise test is used for detecting ERbecause more consistent evidence of subclinicalrhabdomyolysis was found with this test versusmaximal exercise tests.91 Fifteen minutes of trot-ting or, in unfit horses, 2-minute intervals of walk-ing and trotting for a maximum of 15 minutes, isoften sufficient to produce subclinical muscle dam-age in horses prone to chronic ER.109 If signs ofstiffness develop before this, exercise must be con-cluded. A normal response is less than a 3- to 4-foldincrease from basal CK to 4 to 6 hours after theexercise test.
This exercise test is not only useful to identifysusceptibility to ER, it is also useful in defining theamount of exercise to use when horses are put backinto training. The duration of exercise when begin-ning training should be less than that which pro-duced abnormal elevations in serum CK. SerumCK activity is not measured immediately after exer-cise because at this time point it will not reflect theamount of damage occurring during the exercise
test. A 4- to 6-hour postexercise sample is idealbecause this is when the highest CK activity willoccur in serum after muscle damage. Small fluctu-ations in serum CK activity may occur with exercisedue to enhanced muscle membrane permeability,particularly if exercise is prolonged or strenuous andthe horse is untrained.
Muscle Biopsy
The muscle biopsy technique has now become a rou-tine part of equine practice. Most frequently, prac-titioners utilize an open surgical approach; however,neuromuscular laboratories that have the ability tofreeze muscle biopsies in isopentane use the percu-taneous technique. The percutaneous needle bi-opsy has the advantage of a no layup after biopsy.
Open Surgical Technique
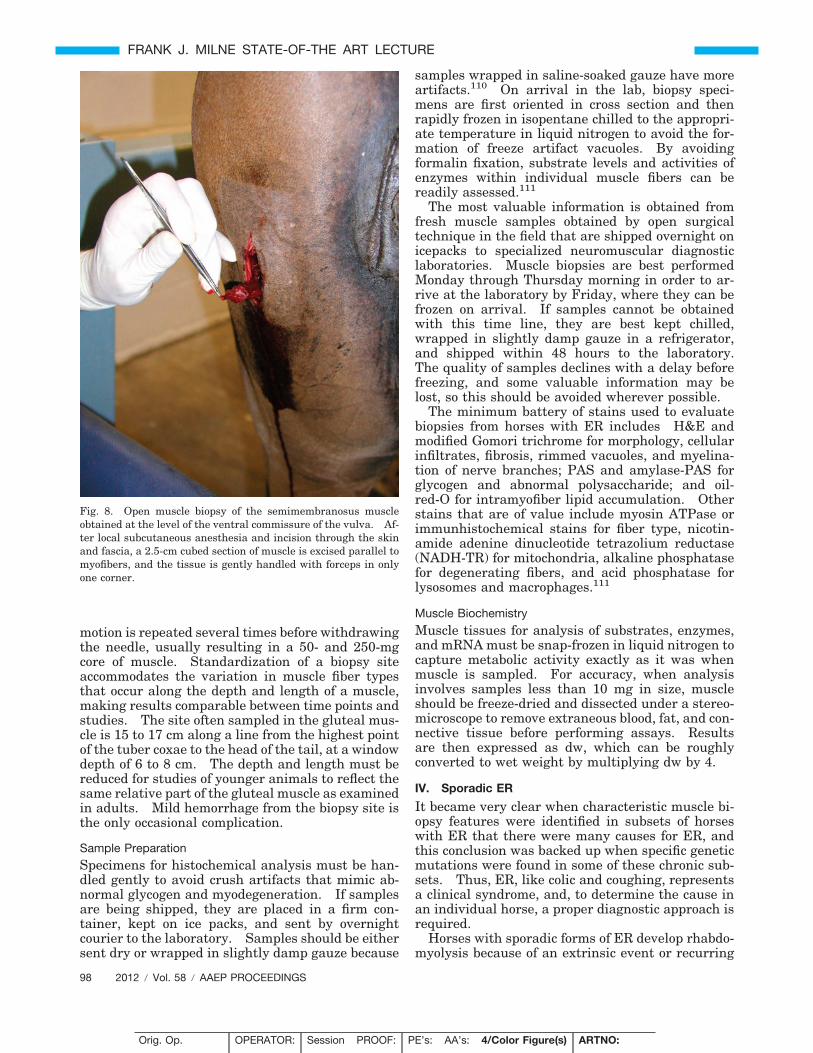
The semimembranosus or semitendinosus muscle isusually selected for the open surgical biopsy becauseof the ability to easily align an incision parallel tothe length of muscle fibers. After wrapping the tailand sterile preparation of the site situated at aboutthe height of the lower commissure of the vulva, alocal anesthetic is placed under the skin (5 mL) butnot into the muscle. The objective is to obtain ap-proximately a half-inch cube of tissue; hence, a suit-ably long skin incision and incision into the fascia isrequired. After undermining the fascia, two paral-lel incisions one-half inch apart are made longitudi-nal to the muscle fibers with a scalpel. The muscleis only handled in one corner using forceps to avoidcrushing the sample and thereby creating artifacts(Fig. 8). The muscle sample is then excised bycross-secting incisions one-half inch apart. Closureof the fascia will prevent muscle hernia, and closureof the subcutaneous layer eliminates dead space pre-venting dehiscence. Interrupted sutures are usedin the skin and can be removed after 14 days. Boxstall rest for a few days is necessary to preventdehiscence.
Percutaneous Technique
This technique is not commonly used in the fieldbecause needles are not readily available, experi-ence is required to obtain an adequate sample size,and this type of sample does not ship as well aslarger surgically excised biopsies. The percutane-ous needle biopsy technique can be used to sample avariety of muscles, but the gluteus medius is mostfrequently evaluated because of its major role inexertion and ease of access. The skin over the siteof the biopsy is sterilely prepared, and 3 mL of localanesthetic is injected. The internal cutting cylin-der of the needle is inserted into the muscle througha 1-cm skin incision and the cutting cylinder is par-tially withdrawn, exposing a window in the needle.The biopsy needle is pressed firmly against the mus-cle to trap a piece of muscle within the window, andthe cylinder is depressed to excise a piece of musclethat moves into the hollow cylinder. The chopping
AAEP PROCEEDINGS � Vol. 58 � 2012 97
FRANK J. MILNE STATE-OF-THE ART LECTURE
F8
Orig. Op. OPERATOR: Session PROOF: PE’s: AA’s: 4/Color Figure(s) ARTNO:
1st disk, 2nd beb dainop 15 1,3-8,12,13,15-17 3353
motion is repeated several times before withdrawingthe needle, usually resulting in a 50- and 250-mgcore of muscle. Standardization of a biopsy siteaccommodates the variation in muscle fiber typesthat occur along the depth and length of a muscle,making results comparable between time points andstudies. The site often sampled in the gluteal mus-cle is 15 to 17 cm along a line from the highest pointof the tuber coxae to the head of the tail, at a windowdepth of 6 to 8 cm. The depth and length must bereduced for studies of younger animals to reflect thesame relative part of the gluteal muscle as examinedin adults. Mild hemorrhage from the biopsy site isthe only occasional complication.
Sample PreparationSpecimens for histochemical analysis must be han-dled gently to avoid crush artifacts that mimic ab-normal glycogen and myodegeneration. If samplesare being shipped, they are placed in a firm con-tainer, kept on ice packs, and sent by overnightcourier to the laboratory. Samples should be eithersent dry or wrapped in slightly damp gauze because
samples wrapped in saline-soaked gauze have moreartifacts.110 On arrival in the lab, biopsy speci-mens are first oriented in cross section and thenrapidly frozen in isopentane chilled to the appropri-ate temperature in liquid nitrogen to avoid the for-mation of freeze artifact vacuoles. By avoidingformalin fixation, substrate levels and activities ofenzymes within individual muscle fibers can bereadily assessed.111
The most valuable information is obtained fromfresh muscle samples obtained by open surgicaltechnique in the field that are shipped overnight onicepacks to specialized neuromuscular diagnosticlaboratories. Muscle biopsies are best performedMonday through Thursday morning in order to ar-rive at the laboratory by Friday, where they can befrozen on arrival. If samples cannot be obtainedwith this time line, they are best kept chilled,wrapped in slightly damp gauze in a refrigerator,and shipped within 48 hours to the laboratory.The quality of samples declines with a delay beforefreezing, and some valuable information may belost, so this should be avoided wherever possible.
The minimum battery of stains used to evaluatebiopsies from horses with ER includes H&E andmodified Gomori trichrome for morphology, cellularinfiltrates, fibrosis, rimmed vacuoles, and myelina-tion of nerve branches; PAS and amylase-PAS forglycogen and abnormal polysaccharide; and oil-red-O for intramyofiber lipid accumulation. Otherstains that are of value include myosin ATPase orimmunhistochemical stains for fiber type, nicotin-amide adenine dinucleotide tetrazolium reductase(NADH-TR) for mitochondria, alkaline phosphatasefor degenerating fibers, and acid phosphatase forlysosomes and macrophages.111
Muscle BiochemistryMuscle tissues for analysis of substrates, enzymes,and mRNA must be snap-frozen in liquid nitrogen tocapture metabolic activity exactly as it was whenmuscle is sampled. For accuracy, when analysisinvolves samples less than 10 mg in size, muscleshould be freeze-dried and dissected under a stereo-microscope to remove extraneous blood, fat, and con-nective tissue before performing assays. Resultsare then expressed as dw, which can be roughlyconverted to wet weight by multiplying dw by 4.
IV. Sporadic ER
It became very clear when characteristic muscle bi-opsy features were identified in subsets of horseswith ER that there were many causes for ER, andthis conclusion was backed up when specific geneticmutations were found in some of these chronic sub-sets. Thus, ER, like colic and coughing, representsa clinical syndrome, and, to determine the cause inan individual horse, a proper diagnostic approach isrequired.
Horses with sporadic forms of ER develop rhabdo-myolysis because of an extrinsic event or recurring
Fig. 8. Open muscle biopsy of the semimembranosus muscleobtained at the level of the ventral commissure of the vulva. Af-ter local subcutaneous anesthesia and incision through the skinand fascia, a 2.5-cm cubed section of muscle is excised parallel tomyofibers, and the tissue is gently handled with forceps in onlyone corner.
98 2012 � Vol. 58 � AAEP PROCEEDINGS
FRANK J. MILNE STATE-OF-THE ART LECTURE
Orig. Op. OPERATOR: Session PROOF: PE’s: AA’s: 4/Color Figure(s) ARTNO:
1st disk, 2nd beb dainop 15 1,3-8,12,13,15-17 3353

extrinsic events that induce muscle damage withexercise. Usually a horse is presumed to have spo-radic ER the first time it develops ER.105 If ERrecurs once the horse has been rested and graduallyreturned to exercise, then an investigation of aninherent chronic form of ER is usually pursued.Causes of sporadic ER include focal or generalizedtrauma to muscle, exercise performed beyond anytraining adaptation or performed to the point ofexhaustion, and dietary imbalances that affect mus-cle function.
Signalment, History, and Diagnosis
Horses with sporadic ER may be of any age, breed,or sex and involved in a wide variety of athleticdisciplines.112 Sporadic ER usually occurs inhorses with a history of adequate performance priorto onset of ER, and a familial history is absent.A diagnosis is made by history, clinical signs, andelevations of serum muscle enzymes. Once exter-nal perturbations that affect muscle function arecorrected, complete resolution and a successful re-turn to performance are expected. Resolution oc-curs after a reasonable period of rest, provision of abalanced diet, and a gradual introduction of a train-ing program matched with performance demands.If over time ER recurs despite reasonable manage-ment, a diagnosis of chronic ER becomes more likely,and further diagnostic testing is appropriate.
Focal or Generalized Trauma
Horses that strain a particular muscle may be veryreluctant to move and show signs resembling moregeneralized ER. This is particularly true whenlumbar, gluteal, psoas, adductor, or semi-mem/ten-dinosus muscles are acutely involved.105 Further,horses that struggle when cast, are caught in afence, or are thrashed by another horse may haveacute signs of rhabdomyolysis due to severe exertionor trauma. A diagnosis of focal trauma is estab-lished by history, physical examination, and serummuscle enzymes. Rectal examination and ultra-sonography may help to localize the extent of muscledamage for focal lesions. On occasion, scintigraphymay help to identify specific muscle involvement.113
Overexertion
A history of an increase in work intensity without afoundation of consistent training for this level ofexercise is usually the basis for suspecting a train-ing imbalance as a cause of ER. Signs of musclestiffness and gait changes range from mild to severe,and severity is reflected by variable elevations ofserum CK activity.105 Overexertion is a well-de-scribed cause of ER in polo horses, with 81% of casesof ER attributed to overexertion and 30% of casesoccurring after a day of rest.114 The incidence ofER is as high as 9% in US polo horses, with most ofthese horses only having one episode of ER early inthe polo season.114
Pathological changes are often not evident in lightmicroscopic evaluation of muscle biopsies from indi-viduals performing unaccustomed exercise, but elec-tron microscopy shows significant disruption of thealignment of muscle contractile proteins withinmuscle fibers.115 In more severe cases, overt seg-mental damage to myofibers may be apparent inmuscle biopsy. Repetitive overuse of muscles, suchas occurs with overtraining, may result in exerciseintolerance and is associated with pathologicchanges such as increased muscle fiber size varia-tion and centrally located myonuclei in musclebiopsies.116
Exhaustion
Exhaustion occurs most commonly in endurancehorses or racehorses exercising in hot, humidweather. Signs of heat exhaustion include weak-ness, ataxia, rapid breathing, muscle fasciculations,sweating, and, in severe cases, collapse. The bodytemperature may be elevated to 105° to 108°F.Muscles are frequently not firm on palpation, al-though serum CK activity can be markedly elevatedand myoglobinuria may be noted.17,18
Dietary Imbalances
Episodes of ER may be triggered by diets with a highnonstructural carbohydrate (NSC) content and lowforage content117,118 or by diets deficient in electro-lytes and may be exacerbated by inadequate sele-nium and vitamin E.
Electrolyte Imbalances
Electrolyte balance within the body is difficult todetermine accurately. One suggested means topractically assess electrolyte balance in horses is tomeasure urinary fractional excretion of electro-lytes.20 Measurement of urinary electrolyte excre-tion as an indicator of electrolyte balance iscomplicated because marked variation can occurfrom diet, exercise, and sampling technique betweenindividuals as well as within individuals from day today.22 Furthermore, the high calcium crystal con-centration of alkaline equine urine requires acidifi-cation to accurately assess calcium and magnesiumcontent.22,119 The high potassium content inter-feres with sodium analysis, using conventional ionspecific electrodes.109 Thus, although popular inthe 1990s16,20,21 urinary fractional excretion is nowrarely performed in the United States unless thereis a strong suspicion that body depletion has oc-curred and if a laboratory can be located with gaschromatography mass spectrometry analysis of acid-ified urine. It is important to ensure that horsesreceive sodium chloride in the diet, with higher con-centrations needed in horses that compete in hothumid weather. Between 30 to 50 g/d combinedwith 15 to 25 g of lite salt containing potassiumchloride is recommended for horses exercising inhot, humid conditions and sweating extensively.
AAEP PROCEEDINGS � Vol. 58 � 2012 99
FRANK J. MILNE STATE-OF-THE ART LECTURE
Orig. Op. OPERATOR: Session PROOF: PE’s: AA’s: 4/Color Figure(s) ARTNO:
1st disk, 2nd beb dainop 15 1,3-8,12,13,15-17 3353

The entire diet should be adequately balanced withcalcium and phosphorus (ration of 2:1 Ca:P is ideal).
Vitamin E and Selenium ConcentrationsWhole-blood selenium concentrations measured inEDTA or heparin tubes are of value in assessingselenium status in animals housed in areas wheresoil is deficient in selenium. In cases where sele-nium has been administered prior to blood collec-tion, glutathione peroxidase activity can be used toassess potential selenium deficiency.120 Vitamin Econcentration should be measured in serum sampleskept chilled and protected from light; however, vari-ability in serum levels can be quite large, and repeatsampling is recommended if marginal levels areidentified in the first sample tested. Horses withER are infrequently deficient in selenium and vita-min E, and, alone, it may not be responsible forER11; however, anecdotal reports suggest that insome cases, supplementation may help to furtherprevent episodes of ER.
ManagementAs described in the section on acute ER, rest withregular access to a paddock once stiffness resides andweekly monitoring of serum CK activity are recom-mended. Horses are much more susceptible to a sec-ond episode of ER in the 2 weeks after an acuteepisode, and allowing horses the ability to calmly de-termine their own exercise through turnout oftenavoids exacerbating rhabdomyolysis. If that is notfeasible, hand-walking must be performed with cau-tion and limited to no more than a few minutes ini-tially. While the horse is rested, the diet can beassessed to ensure that the ration is fed in an amountrecommended by the manufacturer for the correspond-ing level of exercise. This will ensure that the properbalance of vitamins and minerals is provided. If ex-cessive calories are provided when fed at manufactur-er-recommended levels, then the diet should beswitched to a less calorie-dense vitamin/mineral bal-anced diet. A salt block or 30 to 50 g (1 to 3 table-spoons) of salt per day will provide the necessaryadditional sodium chloride, with the amount fed de-pending on the heat, humidity, and intensity of exer-cise. Once serum CK returns to normal, training canbe resumed gradually. A regular exercise schedule,beginning with 20 minutes or less of exercise per day,is gradually increased to eventually match the ex-pected amount of daily exercise to the underlying stateof conditioning.
V. Chronic ER: Intrinsic Causes
Horses that have repeated episodes of ER from ayoung age, or from the time of purchase, or whenthey are put back into training after a long period ofrest, may have an underlying intrinsic abnormalityof muscle function. Many horses with intrinsicmuscle defects will have repeated episodes of ERwith minimal exercise even when the dietary andtraining recommendations for sporadic ER are fol-
lowed. Five specific intrinsic causes of ER havebeen identified to date:
(1) Recurrent exertional rhabdomyolysis (RER)(2) Malignant hyperthermia(3) Type 1 polysaccharide storage myopathy
(PSSM1)(4) Type 2 polysaccharide storage myopathy
(PSSM2)(5) Idiopathic chronic exertional rhabdomyolysis
The idiopathic group represents other causes ofER that have yet to be identified. In all of theseintrinsic forms of chronic ER, it appears that thereare specific environmental stimuli that are neces-sary to trigger muscle necrosis in genetically suscep-tible animals. Horses cannot be cured of theirsusceptibility to this condition, but, if the specificform of ER is identified, changes in managementcan be implemented to minimize episodes ofrhabdomyolysis.
1. Recurrent Exertional Rhabdomyolysis
The term recurrent exertional rhabdomyolysis(RER) describes a subset of ER that is believed to bedue to an abnormality in the regulation of musclecontraction and relaxation.93,109,121,122 Researchinto RER has primarily been performed in Thor-oughbreds and to a lesser extent in Standardbredhorses.93–95,123 Scientific studies of RER have useda small number of horses that share common clinicalsigns of ER and have abnormal muscle contracturetests.93,122,124 Broader epidemiologic and geneticstudies have assumed that the same pathophysio-logic basis for ER exists in the majority of Thorough-bred and Standardbred horses with similar clinicalsigns. Whether this is true are not will requiremore research. There are reports of some Arabianhorses and Warmblood horses103 with ER that mayalso suffer from RER, based on the overlapping his-tories, clinical signs, muscle biopsy findings and re-sponse to management.
Pathogenesis
Rhabdomyolysis is triggered suddenly during exer-cise in RER horses, which results in a sharp rise inserum myoglobin and CK activity (Fig. 7).91 Clini-cally, the triggering event is often associated withexcitement in a horse that already has an underly-ing nervous temperament.94,95,123 Segmental ne-crosis in single fibers or small clusters of necroticfibers scattered throughout fascicles may be appar-ent in fewer than 5% of type 2A and 2B muscle fibersin a gluteus medius muscle biopsy.91,92 Serum cor-tisol concentrations are higher in RER horses thanin normal horses prior to exercise and increase dur-ing an episode of ER.108 Serum concentrations ofepinephrine and norepinephrine are normal prior toan episode but increase dramatically in horses withmarked elevations in serum CK activity.108
100 2012 � Vol. 58 � AAEP PROCEEDINGS
FRANK J. MILNE STATE-OF-THE ART LECTURE
Orig. Op. OPERATOR: Session PROOF: PE’s: AA’s: 4/Color Figure(s) ARTNO:
1st disk, 2nd beb dainop 15 1,3-8,12,13,15-17 3353
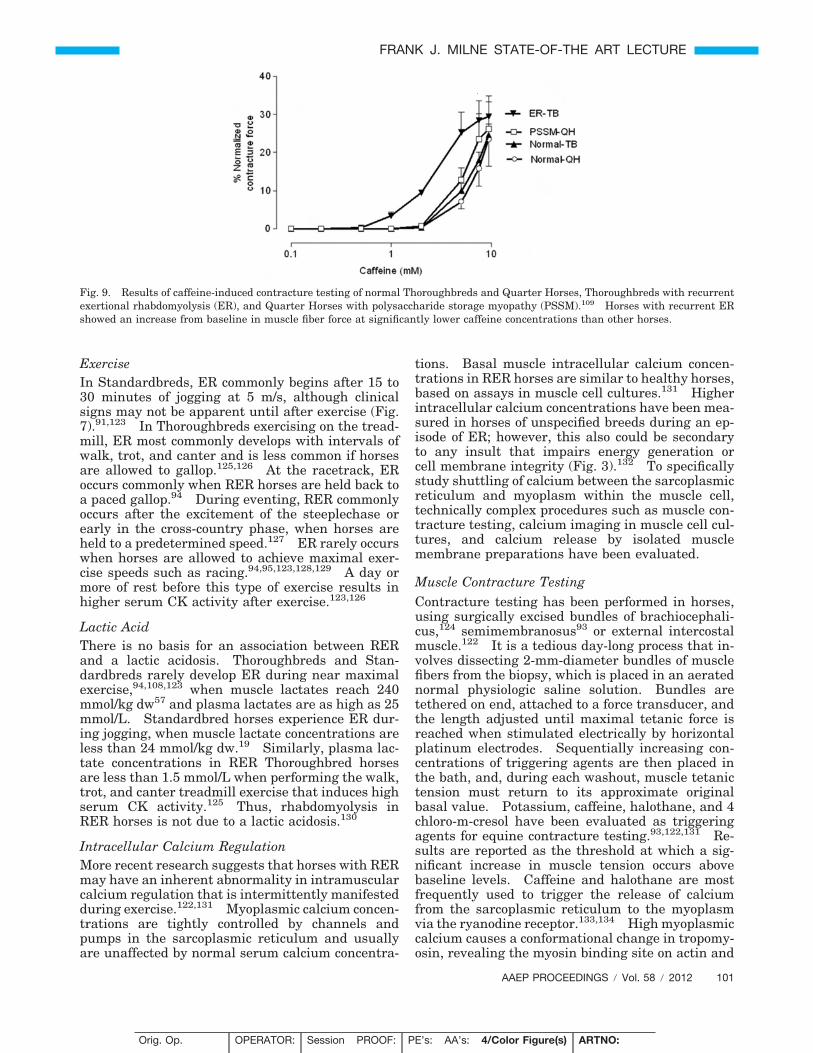
ExerciseIn Standardbreds, ER commonly begins after 15 to30 minutes of jogging at 5 m/s, although clinicalsigns may not be apparent until after exercise (Fig.7).91,123 In Thoroughbreds exercising on the tread-mill, ER most commonly develops with intervals ofwalk, trot, and canter and is less common if horsesare allowed to gallop.125,126 At the racetrack, ERoccurs commonly when RER horses are held back toa paced gallop.94 During eventing, RER commonlyoccurs after the excitement of the steeplechase orearly in the cross-country phase, when horses areheld to a predetermined speed.127 ER rarely occurswhen horses are allowed to achieve maximal exer-cise speeds such as racing.94,95,123,128,129 A day ormore of rest before this type of exercise results inhigher serum CK activity after exercise.123,126
Lactic AcidThere is no basis for an association between RERand a lactic acidosis. Thoroughbreds and Stan-dardbreds rarely develop ER during near maximalexercise,94,108,123 when muscle lactates reach 240mmol/kg dw57 and plasma lactates are as high as 25mmol/L. Standardbred horses experience ER dur-ing jogging, when muscle lactate concentrations areless than 24 mmol/kg dw.19 Similarly, plasma lac-tate concentrations in RER Thoroughbred horsesare less than 1.5 mmol/L when performing the walk,trot, and canter treadmill exercise that induces highserum CK activity.125 Thus, rhabdomyolysis inRER horses is not due to a lactic acidosis.130
Intracellular Calcium RegulationMore recent research suggests that horses with RERmay have an inherent abnormality in intramuscularcalcium regulation that is intermittently manifestedduring exercise.122,131 Myoplasmic calcium concen-trations are tightly controlled by channels andpumps in the sarcoplasmic reticulum and usuallyare unaffected by normal serum calcium concentra-
tions. Basal muscle intracellular calcium concen-trations in RER horses are similar to healthy horses,based on assays in muscle cell cultures.131 Higherintracellular calcium concentrations have been mea-sured in horses of unspecified breeds during an ep-isode of ER; however, this also could be secondaryto any insult that impairs energy generation orcell membrane integrity (Fig. 3).132 To specificallystudy shuttling of calcium between the sarcoplasmicreticulum and myoplasm within the muscle cell,technically complex procedures such as muscle con-tracture testing, calcium imaging in muscle cell cul-tures, and calcium release by isolated musclemembrane preparations have been evaluated.
Muscle Contracture TestingContracture testing has been performed in horses,using surgically excised bundles of brachiocephali-cus,124 semimembranosus93 or external intercostalmuscle.122 It is a tedious day-long process that in-volves dissecting 2-mm-diameter bundles of musclefibers from the biopsy, which is placed in an aeratednormal physiologic saline solution. Bundles aretethered on end, attached to a force transducer, andthe length adjusted until maximal tetanic force isreached when stimulated electrically by horizontalplatinum electrodes. Sequentially increasing con-centrations of triggering agents are then placed inthe bath, and, during each washout, muscle tetanictension must return to its approximate originalbasal value. Potassium, caffeine, halothane, and 4chloro-m-cresol have been evaluated as triggeringagents for equine contracture testing.93,122,131 Re-sults are reported as the threshold at which a sig-nificant increase in muscle tension occurs abovebaseline levels. Caffeine and halothane are mostfrequently used to trigger the release of calciumfrom the sarcoplasmic reticulum to the myoplasmvia the ryanodine receptor.133,134 High myoplasmiccalcium causes a conformational change in tropomy-osin, revealing the myosin binding site on actin and
Fig. 9. Results of caffeine-induced contracture testing of normal Thoroughbreds and Quarter Horses, Thoroughbreds with recurrentexertional rhabdomyolysis (ER), and Quarter Horses with polysaccharide storage myopathy (PSSM).109 Horses with recurrent ERshowed an increase from baseline in muscle fiber force at significantly lower caffeine concentrations than other horses.
AAEP PROCEEDINGS � Vol. 58 � 2012 101
FRANK J. MILNE STATE-OF-THE ART LECTURE
Orig. Op. OPERATOR: Session PROOF: PE’s: AA’s: 4/Color Figure(s) ARTNO:
1st disk, 2nd beb dainop 15 1,3-8,12,13,15-17 3353

inducing a contracture. This test remains an ex-perimental procedure due to the difficulty in obtain-ing accurate and timely results.
Early studies by Hildebrand124 suggested thatsome but not all horses with ER showed a lowerthreshold for caffeine- and halothane-inducedmuscle contractures than healthy horses. Wal-dron-Mease135 and Beech93 found that caffeinecontracture thresholds were lower in semimembra-nosus muscle from Thoroughbreds and Standard-breds susceptible to RER versus breed-matchedcontrols. Lentz et al. chose to study intact externalintercostal muscle fibers, based on concerns thatdamaged ends of transected fibers may cause slightdepolarization and affect the repeatability of thecontracture results.122,136 Intact intercostals mus-cle bundles from six Thoroughbreds with RER had alower contracture threshold for triggering agentssuch as potassium, caffeine, and halothane but not 4chloro-m-cresol compared with control horses (Fig.9).122,131 An abnormality in intracellular calciumregulation was also supported by a muscle cell cul-ture study of RER Thoroughbred horses.131 Alower threshold for induction of myoplasmic calciumrelease by caffeine was found in myotube culturesfrom six RER Thoroughbreds compared with con-trols. Thus, all of these studies supported an ab-normality in intracellular calcium regulation inmost of the Thoroughbred or Standardbred horsesstudied. Contracture testing of PSSM horses didnot reveal any difference in contracture thresholdcompared with controls (Fig. 9).136
Myofibrillar Sensitivity to CalciumA potential alteration in the activation and regula-tion of the myofibrillar contractile apparatus by ion-ized calcium has been studied as a basis for RER.137
No difference was detected between the type II fi-bers of RER-affected and control horses in terms ofcalcium sensitivity of force production in chemicallyskinned muscle fibers or in myofibrillar ATPase ac-tivity. Thus, the basis for pathologic contracturedevelopment in muscles from RER-affected horsesdid not appear to be due to an alteration in myofi-brillar calcium sensitivity.
Isolated Membrane PreparationsTo determine if calcium release and ryanodine bind-ing is abnormal in RER muscle, sarcoplasmic retic-ulum membrane has been isolated from biopsies ofRER and normal horses. An initial study found adifference in calcium-induced calcium release be-tween RER and control horses.93 In another study,the time courses of sarcoplasmic reticulum calcium-induced calcium release and [3H]ryanodine bindingto the ryanodine receptor after incubation withvarying concentrations of ryanodine, caffeine, andionized calcium were similar between muscle mem-branes obtained from control and RER horses.138
Furthermore, the maximal rate of SR calcium-ATPase activity and its affinity for calcium did not
differ between muscle membranes from controlhorses and horses with RER.138 Thus, these stud-ies did not conclusively identify any abnormalities inintracellular calcium regulation, indicating thatthere are distinct differences between RER and dis-orders such as malignant hyperthermia.
Linkage Analysis for Genes Involved inIntracellular Calcium RegulationTo further explore the possibility that RER was dueto an inherited defect in intracellular calcium regu-lation, genetic linkage analysis was performed using96 horses in 4 families of Thoroughbred horses seg-regating for RER.88 No association was detectedlinking RER with the genes RYR1 (ryanodine recep-tor), SERCA (SR ATPase), or DHPR1 (dihydropyri-dine receptor).
Twitch CharacteristicsThe twitch characteristics of muscle from RER Stan-dardbreds and Thoroughbreds have been investi-gated and are different between RER and normalhorses.122 Muscle bundles from RER horses had aslight but significantly faster time to peak tensionand a significantly shorter time to 50% relaxationthan muscle from normal horses.
Thus, despite many detailed investigations of asmall number of horses, the exact defect in regulat-ing muscle contraction in RER horses has yet to beidentified. RER appears to be a novel defect inmuscle excitation-contraction coupling, calcium reg-ulation, or contractility whose basis is yet to bedefined. Alternatively, the increased sensitivity ofRER horses in contracture testing may be a nonspe-cific indicator of an abnormality in other pathwaysgoverning muscle contraction.
Prevalence and Risk FactorsAssuming that the majority of cases of ER at race-tracks are due to RER, the prevalence in Thorough-bred racehorses is remarkably similar around theworld, with estimates ranging from 4.9% in theUnited States,94 5.4% in Australia,112 and 6.7% inthe United Kingdom.95,129 The annual incidence ofER in Standardbred racehorses in Sweden is6.4%.123 Exercise obviously increases the preva-lence of RER in horses, and episodes are observedmore frequently once horses achieve a level offitness.94,123
In Standardbreds and Thoroughbreds, maresmore commonly have RER than males94,95,123; how-ever, no general correlation has been observed be-tween episodes of rhabdomyolysis and stages of theestrus cycle.139 There is an interaction betweenage and sex in RER Thoroughbred horses such thatthe proportion of affected females to males is muchhigher in young horses compared with older agegroups.94 Swedish Standardbreds with RER,which do not begin racing until 3 years of age, didnot show the same age association with RER asThoroughbreds.123 Temperament has a strong ef-
102 2012 � Vol. 58 � AAEP PROCEEDINGS
FRANK J. MILNE STATE-OF-THE ART LECTURE
F9
Orig. Op. OPERATOR: Session PROOF: PE’s: AA’s: 4/Color Figure(s) ARTNO:
1st disk, 2nd beb dainop 15 1,3-8,12,13,15-17 3353
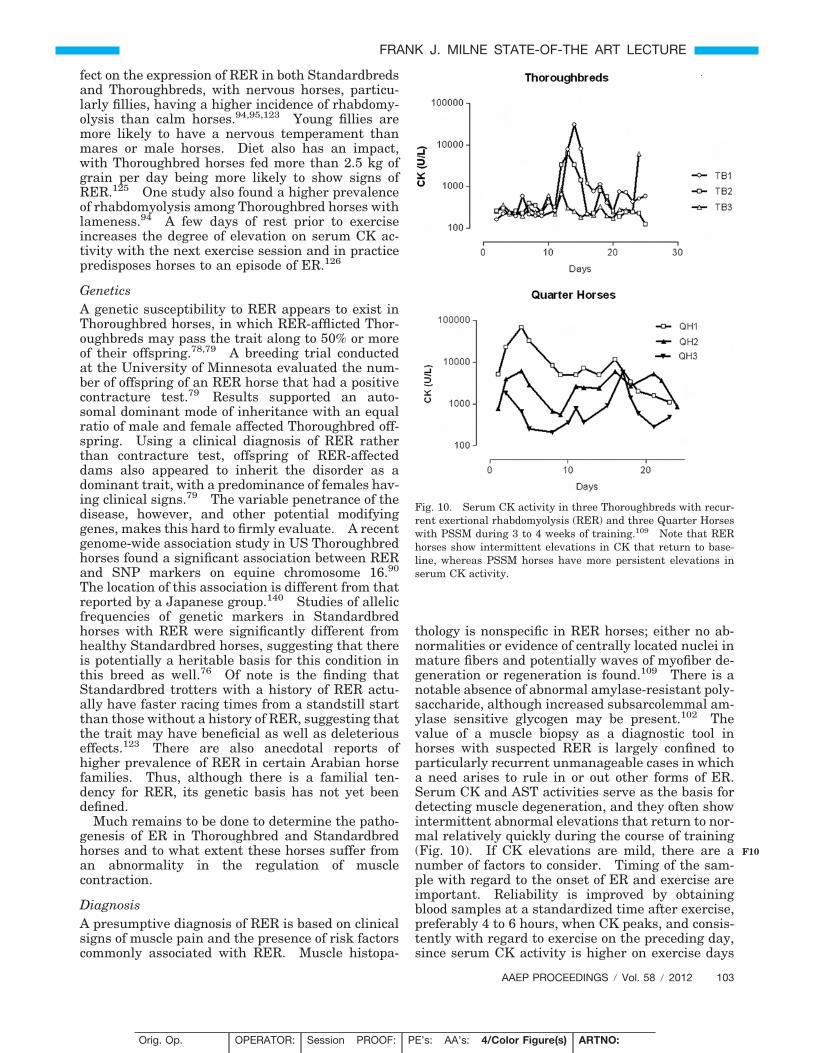
fect on the expression of RER in both Standardbredsand Thoroughbreds, with nervous horses, particu-larly fillies, having a higher incidence of rhabdomy-olysis than calm horses.94,95,123 Young fillies aremore likely to have a nervous temperament thanmares or male horses. Diet also has an impact,with Thoroughbred horses fed more than 2.5 kg ofgrain per day being more likely to show signs ofRER.125 One study also found a higher prevalenceof rhabdomyolysis among Thoroughbred horses withlameness.94 A few days of rest prior to exerciseincreases the degree of elevation on serum CK ac-tivity with the next exercise session and in practicepredisposes horses to an episode of ER.126
GeneticsA genetic susceptibility to RER appears to exist inThoroughbred horses, in which RER-afflicted Thor-oughbreds may pass the trait along to 50% or moreof their offspring.78,79 A breeding trial conductedat the University of Minnesota evaluated the num-ber of offspring of an RER horse that had a positivecontracture test.79 Results supported an auto-somal dominant mode of inheritance with an equalratio of male and female affected Thoroughbred off-spring. Using a clinical diagnosis of RER ratherthan contracture test, offspring of RER-affecteddams also appeared to inherit the disorder as adominant trait, with a predominance of females hav-ing clinical signs.79 The variable penetrance of thedisease, however, and other potential modifyinggenes, makes this hard to firmly evaluate. A recentgenome-wide association study in US Thoroughbredhorses found a significant association between RERand SNP markers on equine chromosome 16.90
The location of this association is different from thatreported by a Japanese group.140 Studies of allelicfrequencies of genetic markers in Standardbredhorses with RER were significantly different fromhealthy Standardbred horses, suggesting that thereis potentially a heritable basis for this condition inthis breed as well.76 Of note is the finding thatStandardbred trotters with a history of RER actu-ally have faster racing times from a standstill startthan those without a history of RER, suggesting thatthe trait may have beneficial as well as deleteriouseffects.123 There are also anecdotal reports ofhigher prevalence of RER in certain Arabian horsefamilies. Thus, although there is a familial ten-dency for RER, its genetic basis has not yet beendefined.
Much remains to be done to determine the patho-genesis of ER in Thoroughbred and Standardbredhorses and to what extent these horses suffer froman abnormality in the regulation of musclecontraction.
DiagnosisA presumptive diagnosis of RER is based on clinicalsigns of muscle pain and the presence of risk factorscommonly associated with RER. Muscle histopa-
thology is nonspecific in RER horses; either no ab-normalities or evidence of centrally located nuclei inmature fibers and potentially waves of myofiber de-generation or regeneration is found.109 There is anotable absence of abnormal amylase-resistant poly-saccharide, although increased subsarcolemmal am-ylase sensitive glycogen may be present.102 Thevalue of a muscle biopsy as a diagnostic tool inhorses with suspected RER is largely confined toparticularly recurrent unmanageable cases in whicha need arises to rule in or out other forms of ER.Serum CK and AST activities serve as the basis fordetecting muscle degeneration, and they often showintermittent abnormal elevations that return to nor-mal relatively quickly during the course of training(Fig. 10). If CK elevations are mild, there are anumber of factors to consider. Timing of the sam-ple with regard to the onset of ER and exercise areimportant. Reliability is improved by obtainingblood samples at a standardized time after exercise,preferably 4 to 6 hours, when CK peaks, and consis-tently with regard to exercise on the preceding day,since serum CK activity is higher on exercise days
Fig. 10. Serum CK activity in three Thoroughbreds with recur-rent exertional rhabdomyolysis (RER) and three Quarter Horseswith PSSM during 3 to 4 weeks of training.109 Note that RERhorses show intermittent elevations in CK that return to base-line, whereas PSSM horses have more persistent elevations inserum CK activity.
AAEP PROCEEDINGS � Vol. 58 � 2012 103
FRANK J. MILNE STATE-OF-THE ART LECTURE
F10
Orig. Op. OPERATOR: Session PROOF: PE’s: AA’s: 4/Color Figure(s) ARTNO:
1st disk, 2nd beb dainop 15 1,3-8,12,13,15-17 3353

that are preceded by a day or more of rest.141 Inaddition, normal values must be adjusted for the ageand sex of horses. Two-year-old fillies generallyhave greater fluctuations in serum CK activity dur-ing race training than 3-year-old fillies orgeldings.141
ManagementPrevention of rhabdomyolysis in horses susceptibleto RER is complex, and multiple factors must bechanged to decrease episodes. These factors in-clude the environment, the exercise regime, anddiet. In addition, medication may be needed attimes to prevent further episodes of muscle damage.
EnvironmentBecause excitement and nervousness often triggerrhabdomyolysis,94,95,123 stressful situations in theenvironment that can be modified must be identi-fied. This may involve a change to a smaller barnwith fewer horses and fewer handlers, compatiblecompanions, and a more consistent daily routine.Many stressed RER-susceptible horses respond to aregular routine including feeding first before otherhorses and training first before other horses, espe-cially if the horse becomes impatient while waiting.Other horses respond to being housed in a quiet areaof the barn next to calm, companionable horses.The use of hot walkers, exercise machines, andswimming pools must be evaluated on an individualbasis, as some horses develop rhabdomyolysis whenusing this type of equipment. Horses that developrhabdomyolysis at specific events, such as horseshows, may need to be reconditioned to decreasethe stress level associated with such events or tran-quillized as part of the accommodation process.Providing daily turnout with other horses seemsto be very beneficial for RER horses and may de-crease anxiety and thereby the likelihood ofrhabdomyolysis.
ExerciseAlthough a period of rest is recommended for spo-radic forms of ER until serum CK normalizes, it isnot recommended for horses with RER.105 Dailyexercise is important in preventing episodes of rhab-domyolysis, and therefore when serum CK is lessthan about 3,000 U/L, horses are returned to regulardaily exercise. Horses with more severe episodes ofrhabdomyolysis may require additional time off in apaddock before gradually resuming exercise. Onceback in training, some form of daily exercise is rec-ommended. Avoiding fighting to hold racehorses toslower speeds during galloping may decrease rhab-domyolysis in RER-susceptible Thoroughbreds.Interval training and reduction of jog miles to nomore than 15 minutes per session will benefit Stan-dardbreds. For horses under saddle, a relaxedwarm-up with intermittent periods of long, lowstretching may be of benefit, and continued breaksto stretch between episodes of collection are recom-
mended. Event horses may require training thatincorporates calm exposure to speeds achieved dur-ing the steeplechase or cross-country phase, espe-cially when used in an interval training program.
DietA nutritionally balanced diet with appropriate ca-loric intake and adequate vitamins and minerals isthe core element of managing RER. For RER Thor-oughbreds and Standardbreds in training, the chal-lenge is usually supplying enough calories in ahighly palatable form to meet their daily energydemands. This is in part because they often re-quire �30 MCal of digestible energy (DE) a day (5 kgsweet feed, 1.5% of body weight in hay), and, be-cause of their nervous temperament, they may bemore discriminating in their eating habits. Out ofthe total daily calories required by RER horses, re-search suggests that less than 20% DE be suppliedby nonstructural carbohydrates (NSC � starch andsugar) and at least 20% of DE be supplied byfat.126,142
Selection of ForageThoroughbred horses do not appear to show thesame significant increase in serum insulin concen-trations in response to consuming hay with an NSCof 17%, as seen in Quarter Horses (Borgia L. 2010PhD thesis, University of Minnesota). This factcombined with the high caloric requirements of race-horses may mean that it is not as important to selecthay with very low NSC content in RER Thorough-breds as it is in PSSM horses. Anecdotally, sometrainers find horses with RER have more frequentepisodes on alfalfa hay. The nervous disposition ofsome RER horses may predispose them to gastriculcers, and thus frequent provision of hay with amoderate NSC and mixed alfalfa content may beindicated.
Low Starch, High Fat ConcentratesWhen RER Thoroughbreds are fed a moderate ca-loric intake (24 MCal/d) in the form of high starchconcentrates (2.5 kg of corn/oats/wheat middlings/molasses) they show very little elevation in serumCK activity with exercise.125 Most Thoroughbredracehorses, however, are fed at least 5 kg of highstarch concentrate/d at 30 MCal or more per horseper day, and, at this level of feeding, postexerciseserum CK activity rises significantly.94 The discov-ery that substitution of fat for starch in a high cal-orie ration significantly reduces muscle damage inexercising RER horses was a major advance in nu-tritional management of RER.126 Practically, how-ever, it was difficult to achieve the desired caloricintake of racehorses because the maximum amountof fat that finicky Thoroughbreds will happily con-sume is limited often to 600 mL for vegetable oil or5 lb/d of balanced rice bran. Management of RERhorses was significantly improved when a palatablemeans to provide the amount of fat required by fit
104 2012 � Vol. 58 � AAEP PROCEEDINGS
FRANK J. MILNE STATE-OF-THE ART LECTURE
Orig. Op. OPERATOR: Session PROOF: PE’s: AA’s: 4/Color Figure(s) ARTNO:
1st disk, 2nd beb dainop 15 1,3-8,12,13,15-17 3353

finicky RER Thoroughbreds was developed. A con-trolled trial using a specialized feed developed forRERa (13% fat by weight and 9% NSC) determinedthat NSC should be no greater than 20% of daily DEand 20% to 25% of daily DE should be provided byfat for optimal management of RER horses fed 30MCal or more per day.126 No beneficial effect onserum CK activity occurred when sodium bicarbon-ate (4.2% of the pellet) was added to a high starchpellet feed.126 The amount of fat in the diet of anRER horse depends on its caloric requirements.Over and above hay at 1.5% to 2% of body weightand 2.5 kg of grain, any additional calories should besupplied by fat.
The benefit of a fat diet does not appear to be achange in muscle metabolism. Pre- and postexer-cise muscle glycogen and lactate concentrations arethe same in RER horses fed a low starch, high fatdiet compared with a high starch.143 Rather, lowstarch, high fat diets in RER horses may decreasemuscle damage by assuaging anxiety and excitabil-ity, which are tightly linked to developing rhabdo-myolysis in susceptible horses. High fat, low starchdiets fed to fit RER horses produce lower glucose,insulin and cortisol responses and led to a calmerdemeanor and lower pre-exercise heart rates.144
Neurohormonal changes may develop in response tohigh serum glucose, insulin, and cortisol concentra-tions, resulting in an anxious demeanor. While acalm demeanor is desired during training, someracehorse trainers feeding low starch, high fat feedsprefer to supplement with a titrated amount of grain3 days prior to a race to potentially increase liverglycogen and increase a horse’s energy during therace.
Expectations
Studies in RER horses show that significant reduc-tions or normalization of postexercise serum CK ac-tivity occurs within a week of commencing a lowstarch, high fat diet.126 Days off training in a stallare discouraged because postexercise CK activity ishigher after 2 days of rest compared with valuestaken when performing consecutive days of thesame amount of submaximal exercise.
Supplements
Horses require daily dietary supplementation withsodium and chloride either in the form of loose salt(30 to 50 g/d) or a salt block. Additional electrolytesupplementation is indicated in hot humid condi-tions. A myriad of supplements are sold that arepurported to decrease lactic acid buildup in skeletalmuscle of RER horses. These include sodium bicar-bonate, B vitamins, branched chain amino acids,and dimethylglycine. Since lactic acidosis is no lon-ger implicated as a cause for rhabdomyolysis, it isdifficult to find a rationale for their use.
MedicationsLow doses of tranquillizers, such as acepromazine,prior to exercise have been used in RER horsesprone to excitement.145 A dose of 7 mg IV 20 min-utes before exercise is reported to make horses morerelaxed and manageable.61 Reserpine and flu-phenazine, which have a longer duration of effect,have also been used for this purpose. Horses givenfluphenazine may occasionally exhibit prolonged bi-zarre behavior. Use of tranquillizers may only benecessary when horses are in their initial phase oftraining and accommodation to a new environment.Horses obviously cannot compete on these medica-tions, and withdrawal times must be observed.
Dantrium sodium acts to decrease release of cal-cium from the ryanodine receptor in skeletal muscleand is used to treat malignant hyperthermia.146
Studies of intact equine intercostal muscle from ERhorses show that dantrolene lowers already elevatedmyoplasmic calcium concentrations.132 Controlledand field studies have also shown that oral dan-trolene can significantly decrease signs of rhabdo-myolysis in RER horses.147,148 One study indicatedthat horses must be fasted to achieve measurableblood levels of dantrolene,148 but more recent con-trolled trials suggest that absorption of dantrium issuperior in fed versus fasted horses.149 Adminis-tration of 4 mg/kg of dantrium PO 1 hour beforeexercise to RER-susceptible horses fed high graindiets resulted in significantly lower serum CK activ-ity 4 hours after exercise compared with placebo(Fig. 11).148 A dose of 800 mg of dantrium wasgiven to Thoroughbred horses in the United King-dom 1 hour prior to exercise and resulted in signif-
Fig. 11. Effect of oral administration of 4 mg/kg dantrolene 60minutes before treadmill exercise in Thoroughbred horses withrecurrent exertional rhabdomyolysis.148 Bars indicate meanvalues. Four-hour postexercise serum creatine kinase (CK) ac-tivity was significantly lower when dantrolene was administered.
AAEP PROCEEDINGS � Vol. 58 � 2012 105
FRANK J. MILNE STATE-OF-THE ART LECTURE
F11
Orig. Op. OPERATOR: Session PROOF: PE’s: AA’s: 4/Color Figure(s) ARTNO:
1st disk, 2nd beb dainop 15 1,3-8,12,13,15-17 3353

icantly lower postexercise CK activity than aplacebo.147
Phenytoin (7.7 to 11 mg/kg PO BID) has beenreported to be effective in preventing rhabdomyoly-sis in horses with RER.150 Phenytoin acts on anumber of ion channels within muscle and nerves,including sodium and calcium channels. Thera-peutic levels vary, so oral doses are adjusted bymonitoring serum levels to achieve 8 �g/mL and notto exceed 12 �g/mL.151 Drowsiness and ataxia areevidence that the dose of phenytoin is too high andthe dose should be decreased by half. Initial dos-ages start at 6 to 8 mg/kg orally twice a day for 3 to5 days. If the horse is still experiencing rhabdomy-olysis but is not drowsy, the dose can be increased by1 mg/kg increments every 3 to 4 days. Phenytoin isa monoaminoxidase activator and can affect dosagesof other medications. Unfortunately, long-termtreatment with dantrolene or phenytoin is expen-sive, and these drugs must be withdrawn prior tocompetition.
Hormone TherapyVarious hormones, ranging from thyroxine to pro-gesterone and testosterone, have been given tohorses with RER. Initial studies linking low T3and T4 concentrations in horses with ER have notbeen substantiated.12,14 Some mares appear to ex-hibit signs of rhabdomyolysis during estrus, and itmay be of benefit in these horses to suppress estrusbehavior by using progesterone injections. Testos-terone and anabolic steroids have been used at race-tracks to prevent signs of RER, but their use is nolonger permitted.
Adjunct TherapiesMassage, myofascial release, mesotherapy, stretch-ing, aqua-treadmills, and hot/cold therapy per-formed by experienced therapists may be of benefitin individual cases.152–154 Their use may promoterelaxation and normal muscle tension and buildmuscle strength.
2. Malignant HyperthermiaMalignant hyperthermia (MH) is a genetic disorderthat occurs in Quarter Horses and Paint Horses as aresult of a mutation in the ryanodine receptor 1 gene(RYR1).83,155 It was discovered by Dr. Monica Ale-man, who investigated two research horses that de-veloped classic clinical signs of MH when anesthesiawas induced by inhalation of halothane by maskduring a research project.84,155
PathogenesisA point mutation in the RYR1 gene lowers the acti-vation and heightens the deactivation threshold ofthe ryanodine receptor. When triggered, the ab-normal ryanodine receptor remains open, causing adrastic efflux of calcium from the SR and an increasein myoplasmic calcium, producing a contracture.156
The process of reabsorbing excess myoplasmic cal-
cium consumes large amounts of ATP and generatesexcessive heat. Myofibers are damaged by the de-pletion of ATP and possibly the high temperatures(Fig. 3).
Prevalence and Risk FactorsMalignant hyperthermia is a rare disorder in Quar-ter Horses and Paint Horses, affecting �1% of thebreed. Halter and pleasure horse lines have thehighest prevalence. Rhabdomyolysis may be in-duced by exercise and anesthesia but can be ex-tremely intermittent in nature.83
GeneticsAn autosomal dominant mutation exists in exon 46of the skeletal muscle RYR1 gene on ECA 10.
Clinical SignsClinical signs of hyperthermia (40.5°C, 104.9°F), hy-percapnea (PaCO2, 274 mm Hg) and acidosis (pH of6.72) were reported in an experimental protocol ap-proximately 60 minutes after anesthesia was in-duced by delivering halothane via a facemaskwithout premedication.70 Death occurred from car-diopulmonary arrest, and profound rigor mortis waspresent almost immediately. Hematologic changesmeasured 2 minutes after death included hemocon-centration, hyperkalemia, hypercalcemia, hyper-phosphatemia, hyperglycemia, and elevatedcreatinine. Serum CK activity was mildly elevatedat 843 U/L, and myoglobin was 10� higher than thereference range in this horse.
Horses with the RYR1 mutation may intermit-tently show signs of ER and high body tempera-tures.96 Some MH-affected horses have diedsuddenly after an episode of ER. Horses with theMH mutation may also have the GYS1 mutationassociated with type 1 PSSM.62 Such horses havemore severe episodes of ER, higher serum CK activ-ity after exercise, and a more moderated response tothe diet and exercise regimes recommended for type1 PSSM.62
DiagnosisGenetic testing is recommended in Quarter Horsesand Paint Horses with difficult-to-manage forms ofPSSM or a family history of postanesthetic compli-cations. Testing is available through the Veteri-nary Diagnostic Laboratory at the University ofMinnesota (http://www.vdl.umn.edu/vdl/ourservices/neuromuscular.html) and at the University of Califor-nia, Davis.
Muscle biopsy is not particularly useful for diag-nosing MH. Biopsies may contain mild myopathicchanges, including increased variation in fiber sizes,centrally located nuclei, fiber necrosis, glycogen de-pletion, and ringbinden fibers.
TreatmentThe most successful outcome for a horse with sus-pected malignant hyperthermia would be pretreat-
106 2012 � Vol. 58 � AAEP PROCEEDINGS
FRANK J. MILNE STATE-OF-THE ART LECTURE
Orig. Op. OPERATOR: Session PROOF: PE’s: AA’s: 4/Color Figure(s) ARTNO:
1st disk, 2nd beb dainop 15 1,3-8,12,13,15-17 3353
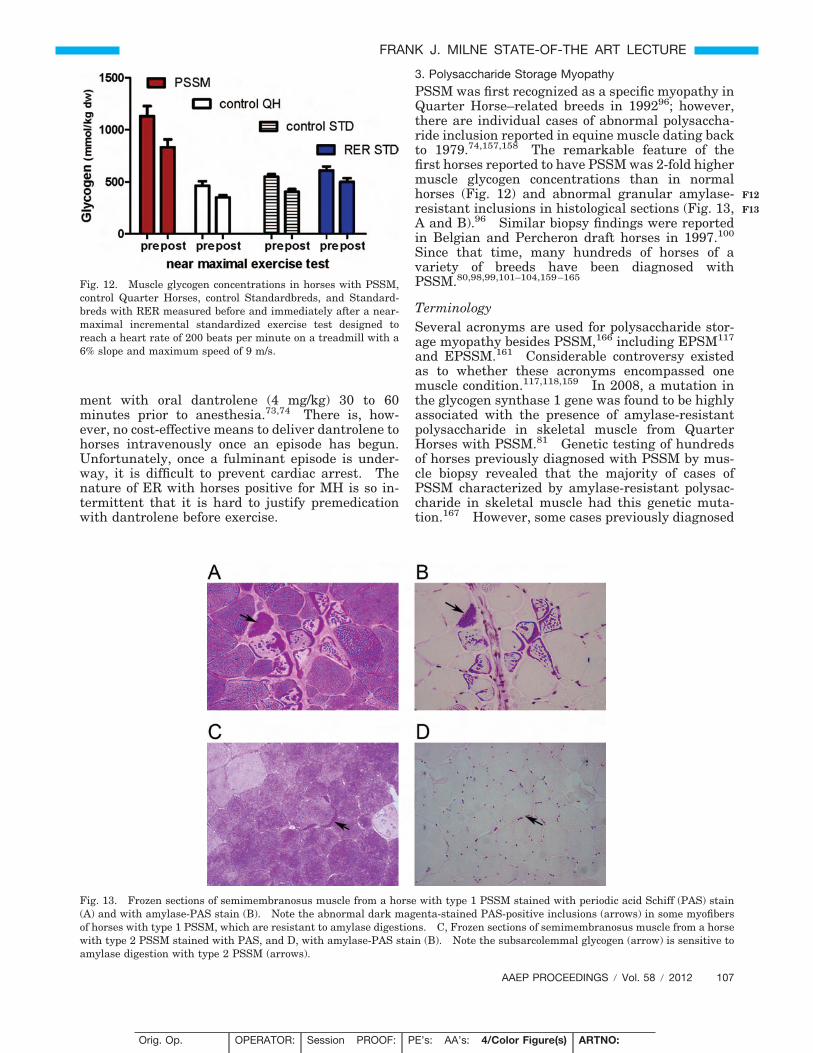
ment with oral dantrolene (4 mg/kg) 30 to 60minutes prior to anesthesia.73,74 There is, how-ever, no cost-effective means to deliver dantrolene tohorses intravenously once an episode has begun.Unfortunately, once a fulminant episode is under-way, it is difficult to prevent cardiac arrest. Thenature of ER with horses positive for MH is so in-termittent that it is hard to justify premedicationwith dantrolene before exercise.
3. Polysaccharide Storage Myopathy
PSSM was first recognized as a specific myopathy inQuarter Horse–related breeds in 199296; however,there are individual cases of abnormal polysaccha-ride inclusion reported in equine muscle dating backto 1979.74,157,158 The remarkable feature of thefirst horses reported to have PSSM was 2-fold highermuscle glycogen concentrations than in normalhorses (Fig. 12) and abnormal granular amylase-resistant inclusions in histological sections (Fig. 13,A and B).96 Similar biopsy findings were reportedin Belgian and Percheron draft horses in 1997.100
Since that time, many hundreds of horses of avariety of breeds have been diagnosed withPSSM.80,98,99,101–104,159–165
TerminologySeveral acronyms are used for polysaccharide stor-age myopathy besides PSSM,166 including EPSM117
and EPSSM.161 Considerable controversy existedas to whether these acronyms encompassed onemuscle condition.117,118,159 In 2008, a mutation inthe glycogen synthase 1 gene was found to be highlyassociated with the presence of amylase-resistantpolysaccharide in skeletal muscle from QuarterHorses with PSSM.81 Genetic testing of hundredsof horses previously diagnosed with PSSM by mus-cle biopsy revealed that the majority of cases ofPSSM characterized by amylase-resistant polysac-charide in skeletal muscle had this genetic muta-tion.167 However, some cases previously diagnosed
Fig. 12. Muscle glycogen concentrations in horses with PSSM,control Quarter Horses, control Standardbreds, and Standard-breds with RER measured before and immediately after a near-maximal incremental standardized exercise test designed toreach a heart rate of 200 beats per minute on a treadmill with a6% slope and maximum speed of 9 m/s.
Fig. 13. Frozen sections of semimembranosus muscle from a horse with type 1 PSSM stained with periodic acid Schiff (PAS) stain(A) and with amylase-PAS stain (B). Note the abnormal dark magenta-stained PAS-positive inclusions (arrows) in some myofibersof horses with type 1 PSSM, which are resistant to amylase digestions. C, Frozen sections of semimembranosus muscle from a horsewith type 2 PSSM stained with PAS, and D, with amylase-PAS stain (B). Note the subsarcolemmal glycogen (arrow) is sensitive toamylase digestion with type 2 PSSM (arrows).
AAEP PROCEEDINGS � Vol. 58 � 2012 107
FRANK J. MILNE STATE-OF-THE ART LECTURE
F12
F13
Orig. Op. OPERATOR: Session PROOF: PE’s: AA’s: 4/Color Figure(s) ARTNO:
1st disk, 2nd beb dainop 15 1,3-8,12,13,15-17 3353

with PSSM by muscle biopsy, particularly thosewith excessive amylase-sensitive glycogen, did notpossess the genetic mutation. This suggested thatthere are at least two forms of PSSM.167,168 Forclarity, the form of PSSM caused by a glycogen syn-thase 1 (GYS1) gene mutation is now termed type 1(PSSM1), whereas the form of PSSM that is notcaused by the GYS1 mutation and whose origin isyet unknown is now termed type 2 (PSSM2).168
PSSM1 probably is the same disorder described as“azoturia” or “Monday morning disease” in workhorses in the 19th and 20th centuries.2–4
Type 1 PSSM
PrevalencePSSM1 is estimated to have emerged approximately1600 years ago when the great horse was beingdeveloped from European draft and light horsebreeds to carry knights with heavy armor into bat-tle.81 The highest prevalence of PSSM1 appears tooccur in draft horses derived from Continental Eu-ropean breeds (Table 2).169 In fact, many Conti-nental European drafts are homozygous for thedominant PSSM1 trait (90% prevalence of PSSM inTrekpaards, with 40% of tested Belgian Trekpaardshomozygous). North American Belgians andPercherons also have a high prevalence of PSSM1,at 36% and 54% of horses affected.170 In contrast,the prevalence of PSSM is very low in United King-dom–derived breeds such as Shires and Clydes-dales,170 but PSSM1 is present in other UK horse
breeds such as Irish Drafts, Cob, and Conne-mara.163,171 Numerous other breeds, more than20, in fact, have the GYS1 mutation responsible forPSSM1.163,165,167,169,170
Prevalence estimates of PSSM1 in Quarter Horsesrange from 6% to 10% of the breed and 6% to 8% forAmerican Paint and Appaloosa Horses.97,172 Thehighest frequency of PSSM1 occurs in halter Quar-ter Horses (28% affected) and the lowest frequencyin racing Quarter Horses.172 The GYS1 mutationhas been identified in approximately 72% of QuarterHorses diagnosed with PSSM by muscle biopsy andin 18% of Warmbloods of a variety of types diag-nosed with PSSM by muscle biopsy.98 The preva-lence of PSSM1 is very low to nonexistent in lighthorse breeds such as Arabians,170 Standardbreds,123
and Thoroughbreds170 (Table 2).
Risk FactorsSome horses with PSSM1 are asymptomatic,whereas other horses routinely show stiffness withexercise.99,104 In a family of Warmbloods sired by astallion with the GYS1 mutation, the mutation re-sulted in a 7-fold higher risk of developing ER,with 34% of horses with the GYS1 mutation lackingclinical signs of PSSM1.104 The reason for vari-ability in clinical signs of PSSM1 probably lies inthe influence of diet, exercise, other environmentalfactors, and other genes on the expression of thedisease.117,173,174 There is no significant tempera-ment, body type, or sex predilection for PSSM1.8
The most common trigger for clinical signs of
Table 2. Number of North American and European Horses Tested and the Percentage of Horses Testing Positive (Heterozygous and Homozygous)for the GYS1 Mutation in Breeds Sampled Using Random Sampling and Nonrandom Sampling
Breed Number Tested Prevalence, %
Random sampling, North AmericaMcCue et al., 2010 Percheron 149 62
Belgian 149 39Clydesdale* 132 0Shire 195 0.5Quarter Horse 335 6.6Paint 195 7.7Appaloosa 152 5.9Morgan 214 0.9Thoroughbred 96 0Arabian 100 0Norwegian Fjord 46 0Icelandic Pony 45 0
Nonrandom sampling, Europe Belgian Trekpaard 38 92Baird et al., 2010 Netherlands Trekpaard 23 74
Comtois 88 80Breton 51 63Swedish Ardenner 29 38Rheinisch-Deutsches Kaltblut 44 68Sud-Deutsches Kaltblut 265 20Hannoverian 214 0
Isgren et al., 2011 Standardbred 36 0
*Both randomized and nonrandomized samples.
108 2012 � Vol. 58 � AAEP PROCEEDINGS
FRANK J. MILNE STATE-OF-THE ART LECTURE
T2
Orig. Op. OPERATOR: Session PROOF: PE’s: AA’s: 4/Color Figure(s) ARTNO:
1st disk, 2nd beb dainop 15 1,3-8,12,13,15-17 3353

PSSM1 is less than 20 minutes of exercise.104,162
Clinical signs occur most commonly at a walk andtrot, particularly if the horse has been rested forseveral days prior to exercise or isunfit.104,104 Diets high in NSC also increase therisk of muscle pain and stiffness in PSSM1horses.173,175 About 40% of owners feel there is aseasonal incidence to the development of clinicalsigns, which some have attributed to quality of grassavailable at the time.162,171 Although exercise is byfar the most common trigger for PSSM, Quarter andPaint Horse foals and weanlings may develop rhab-domyolysis in conjunction with a systemic illness.176
Genetics
A small breeding trial of 4 horses combined with ped-igree analysis of �20 PSSM Quarter Horses initiallypointed to a recessive pattern of inheritance forPSSM.77,80 Conclusions were based on the fact thatfoals from PSSM � PSSM matings developed PSSM,and consanguinity was present in pedigrees. Furtherexpansion of this breeding herd by outcrossing PSSMQuarter Horses to normal stallions indicated that adominant rather than recessive pattern of inheritancewas most likely because affected foals arose from mat-ing PSSM affected to normal horses.81 Whole-genome analysis of Quarter Horses with PSSM1 iden-tified an autosomal dominant single base-pair muta-tion in glycogen synthase 1 (GYS1), which results inan arginine substitution for histidine at codon 309 inglycogen synthase.81 The effect of this amino acidsubstitution is a higher than normal activity of glyco-gen synthase both at basal states and when activatedby G-6-P. This gain-of-function mutation in GYS1 isa novel genetic mutation that has never previouslybeen proposed as a cause for myopathy or exertionalrhabdomyolysis.
Diagnosis
A high degree of suspicion of a diagnosis of PSSM1arises from clinical signs and persistent elevationsin serum CK and AST activities (Fig. 10).103,104,177
If horses have normal serum muscle enzymes atrest, an exercise test consisting of a maximum of 15minutes lunging at a walk and trot may help todetermine if subclinical ER is present.109 Support-ive evidence would include a minimum of a 3-foldelevation in CK activity 4 hours after exercise.
Muscle biopsy provides one means to diagnosetype 1 PSSM.96,178,179 The distinctive features ofPSSM1 in muscle biopsy samples are numerous sub-sarcolemmal vacuoles (Fig. 6B) and dense, crystal-line periodic acid Schiff (PAS)-positive, amylase-resistant inclusions in fast-twitch fibers (Fig. 13, Aand B). A diagnosis can be made irrespective ofdiet and proximity of sampling to recent episodes ofrhabdomyolysis. A false-negative diagnosis of type1 PSSM by muscle biopsy may occur if biopsy sam-ples are small or if horses are less than 1 year ofage.80
The gold standard for diagnosis of PSSM1 is ge-netic testing for the GYS1 mutation performed onwhole blood or hair root samples at the Universityof Minnesota in North America (http://www.vdl.umn.edu/vdl/ourservices/neuromuscular.html), in agenetic testing panel offered through the AQHA orin Europe by Laboklin (http://www.laboklin.co.uk/laboklin/GeneticDiseases.jsp). Other laboratoriesare not licensed to perform this test.
It is important to note that in draft horses ofContinental European origin, the very high preva-lence of PSSM in essence means that there is a highchance that any clinical sign could be falsely associ-ated with the disease PSSM1.99 Thus, clinicaljudgment is required to determine whether the clin-ical signs of the animal could reasonably be associ-ated with a myopathic process.
Acute Clinical SignsHorses usually show signs of PSSM1 at, on average,5 years of age; however, this can range from 1 to 14years of age.104,162 Acute clinical signs occur whenhorses are calm and include tucking up of the abdo-men, fasciculations in the flank, muscle stiffness,sweating, reluctance to move forward, and overtfirm muscle contractures.162 The hindquarters arefrequently most affected, but back muscles, abdo-men, and forelimb muscles may also be involved.Signs of pain can last for more than 2 hours, andabout 10% of cases become recumbent.162 Lesscommon signs of PSSM in Quarter Horses includegait abnormalities, mild colic, and muscle wasting.During an acute episode of ER, horses with PSSM1often have markedly elevated serum CK activity of�35,000 U/L, and myoglobinuria may be present.Severe colic-like pain after exercise and myoglobin-uric renal failure are less common presentingcomplaints.177,180
Chronic Clinical SignsLight breeds: Chronic signs of PSSM1 in ridinghorses include a lack of energy when under saddle,reluctance to move forward, stopping and stretchingout as if to urinate, and a sour attitude towardexercise.103,162 Horses may have a combination oflow-grade reluctance to exercise, poor performance,and repeated episodes of ER.164 PSSM1 may alsopresent in dressage and show jumpers as chronicback pain, failure to round over fences, and fascicu-lations or pain on palpation of lumbar muscles.102
The range of severity of clinical signs of PSSM1 canbe wide, with complete incapacitation in rare cases.Serum CK activities are often elevated in unman-aged horses, even when horses are rested.177 Themedian CK and AST activity for all PSSM1 QuarterHorses with muscle biopsies submitted to the Uni-versity of Minnesota was 2,809 and 1,792 U/L,respectively.
A small number of Quarter Horses and PaintHorses have both the GYS1 mutation and the ge-netic mutation for malignant hyperthermia
AAEP PROCEEDINGS � Vol. 58 � 2012 109
FRANK J. MILNE STATE-OF-THE ART LECTURE
Orig. Op. OPERATOR: Session PROOF: PE’s: AA’s: 4/Color Figure(s) ARTNO:
1st disk, 2nd beb dainop 15 1,3-8,12,13,15-17 3353
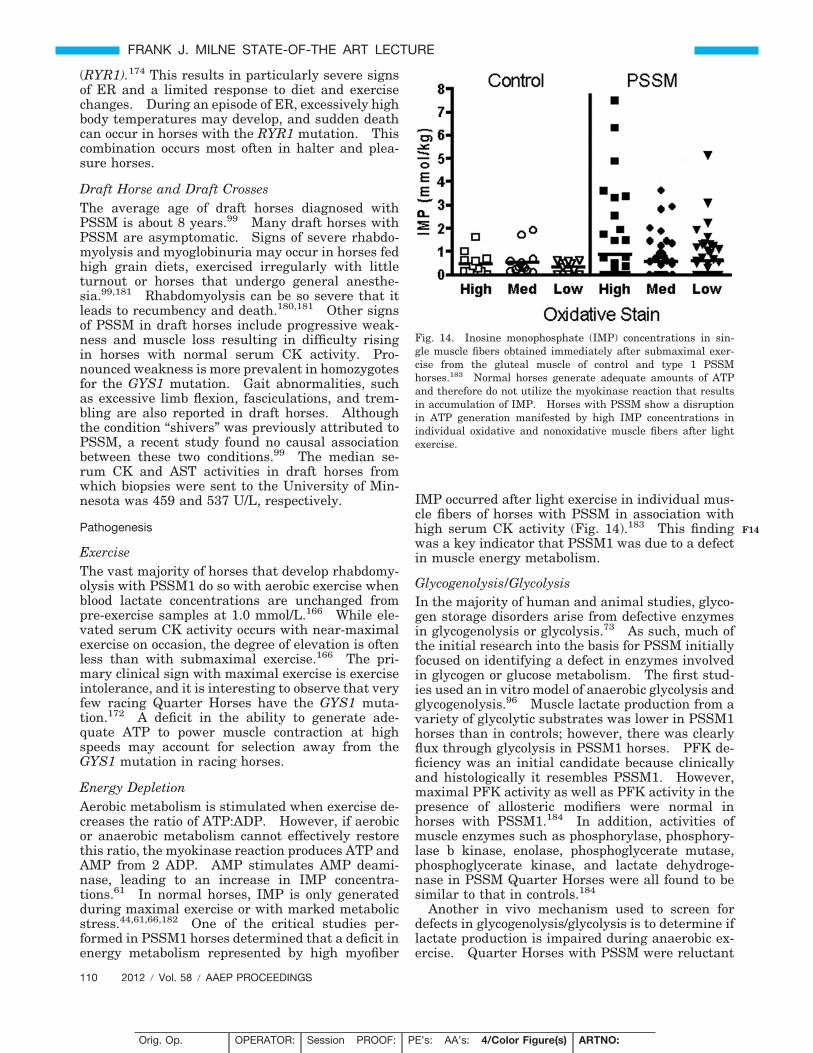
(RYR1).174 This results in particularly severe signsof ER and a limited response to diet and exercisechanges. During an episode of ER, excessively highbody temperatures may develop, and sudden deathcan occur in horses with the RYR1 mutation. Thiscombination occurs most often in halter and plea-sure horses.
Draft Horse and Draft CrossesThe average age of draft horses diagnosed withPSSM is about 8 years.99 Many draft horses withPSSM are asymptomatic. Signs of severe rhabdo-myolysis and myoglobinuria may occur in horses fedhigh grain diets, exercised irregularly with littleturnout or horses that undergo general anesthe-sia.99,181 Rhabdomyolysis can be so severe that itleads to recumbency and death.180,181 Other signsof PSSM in draft horses include progressive weak-ness and muscle loss resulting in difficulty risingin horses with normal serum CK activity. Pro-nounced weakness is more prevalent in homozygotesfor the GYS1 mutation. Gait abnormalities, suchas excessive limb flexion, fasciculations, and trem-bling are also reported in draft horses. Althoughthe condition “shivers” was previously attributed toPSSM, a recent study found no causal associationbetween these two conditions.99 The median se-rum CK and AST activities in draft horses fromwhich biopsies were sent to the University of Min-nesota was 459 and 537 U/L, respectively.
Pathogenesis
ExerciseThe vast majority of horses that develop rhabdomy-olysis with PSSM1 do so with aerobic exercise whenblood lactate concentrations are unchanged frompre-exercise samples at 1.0 mmol/L.166 While ele-vated serum CK activity occurs with near-maximalexercise on occasion, the degree of elevation is oftenless than with submaximal exercise.166 The pri-mary clinical sign with maximal exercise is exerciseintolerance, and it is interesting to observe that veryfew racing Quarter Horses have the GYS1 muta-tion.172 A deficit in the ability to generate ade-quate ATP to power muscle contraction at highspeeds may account for selection away from theGYS1 mutation in racing horses.
Energy DepletionAerobic metabolism is stimulated when exercise de-creases the ratio of ATP:ADP. However, if aerobicor anaerobic metabolism cannot effectively restorethis ratio, the myokinase reaction produces ATP andAMP from 2 ADP. AMP stimulates AMP deami-nase, leading to an increase in IMP concentra-tions.61 In normal horses, IMP is only generatedduring maximal exercise or with marked metabolicstress.44,61,66,182 One of the critical studies per-formed in PSSM1 horses determined that a deficit inenergy metabolism represented by high myofiber
IMP occurred after light exercise in individual mus-cle fibers of horses with PSSM in association withhigh serum CK activity (Fig. 14).183 This findingwas a key indicator that PSSM1 was due to a defectin muscle energy metabolism.
Glycogenolysis/GlycolysisIn the majority of human and animal studies, glyco-gen storage disorders arise from defective enzymesin glycogenolysis or glycolysis.73 As such, much ofthe initial research into the basis for PSSM initiallyfocused on identifying a defect in enzymes involvedin glycogen or glucose metabolism. The first stud-ies used an in vitro model of anaerobic glycolysis andglycogenolysis.96 Muscle lactate production from avariety of glycolytic substrates was lower in PSSM1horses than in controls; however, there was clearlyflux through glycolysis in PSSM1 horses. PFK de-ficiency was an initial candidate because clinicallyand histologically it resembles PSSM1. However,maximal PFK activity as well as PFK activity in thepresence of allosteric modifiers were normal inhorses with PSSM1.184 In addition, activities ofmuscle enzymes such as phosphorylase, phosphory-lase b kinase, enolase, phosphoglycerate mutase,phosphoglycerate kinase, and lactate dehydroge-nase in PSSM Quarter Horses were all found to besimilar to that in controls.184
Another in vivo mechanism used to screen fordefects in glycogenolysis/glycolysis is to determine iflactate production is impaired during anaerobic ex-ercise. Quarter Horses with PSSM were reluctant
Fig. 14. Inosine monophosphate (IMP) concentrations in sin-gle muscle fibers obtained immediately after submaximal exer-cise from the gluteal muscle of control and type 1 PSSMhorses.183 Normal horses generate adequate amounts of ATPand therefore do not utilize the myokinase reaction that resultsin accumulation of IMP. Horses with PSSM show a disruptionin ATP generation manifested by high IMP concentrations inindividual oxidative and nonoxidative muscle fibers after lightexercise.
110 2012 � Vol. 58 � AAEP PROCEEDINGS
FRANK J. MILNE STATE-OF-THE ART LECTURE
F14
Orig. Op. OPERATOR: Session PROOF: PE’s: AA’s: 4/Color Figure(s) ARTNO:
1st disk, 2nd beb dainop 15 1,3-8,12,13,15-17 3353

to perform anaerobic exercise, but at their maxi-mum speed, they depleted glycogen by 25% (Fig. 12)and generated higher concentrations of lactic acidthan normal horses performing the same exercisetest.166 Thus, combined, this research indicatedthat a glycogenolytic/glyolytic defect was improbablein PSSM1. Since glycogen degradation was unim-paired with maximal exercise, research to uncoverthe basis for excessive glycogen storage in PSSM1shifted to potential novel defects that enhanced glu-cose uptake or glycogen synthesis.
Glucose Uptake Into Muscle CellsGlucose transport occurs via GLUT1, which is con-stitutively present in the cell membrane, andGLUT4, which translocates to the muscle cell mem-brane in response to insulin secretion and exercise.Oral and intravenous glucose tolerance testingshowed that blood glucose clearance occurred 1.5times faster in PSSM1 Quarter Horses comparedwith control horses despite their lower insulin con-centrations.185 Enhanced insulin sensitivity inPSSM1 Quarter Horses was confirmed by euglyce-mic hyperinsulinemic clamping.186 This enhancedsensitivity to insulin was breed-specific, however, asBelgian Draft horses with PSSM did not show sim-ilar enhanced insulin sensitivity.187 Enhanced in-sulin sensitivity and glucose uptake did not appearto be responsible for ER in PSSM horses, as dexa-methasone treatment suppressed insulin sensitivitywithout any effect on serum CK responses to exer-cise.188 When the Quarter Horse data was reeval-uated once their genotypes were discovered, thosehorses with both GYS1 and RYR1 mutations had thegreatest increase in insulin sensitivity. This sug-gests a possible role of the RYR1 gene in enhancingthe insulin sensitivity of PSSM horses. The con-tent of GLUT1 or GLUT4 in muscles from horseswith PSSM1 was evaluated by Western blot, and nodifferences were found compared with controls.186
An increase in PSSM1 skeletal muscle glucoseuptake in response to exercise was suggested by thesignificantly higher free glucose concentrations inmuscle cells of PSSM horses after light exercise,which was not present in either basal resting condi-tions or in control horses after exercise.189 Onceinside the muscle cell, glucose is phosphorylated toform glucose-6-phosphate (G-6-P) by hexokinase.No difference in hexokinase activity was identifiedin PSSM versus control horse muscle96; however,variably high G-6-P concentrations have been foundin PSSM skeletal muscle.166,175,186
Glycogen SynthesisThe finding that PSSM horses rapidly resynthesizeglycogen even when postexercise concentrations be-come as low as pre-exercise concentrations inhealthy horses suggested that glycogen synthesiswas abnormally regulated in PSSM horses (Fig.12).185 Glycogen synthesis begins with a protein(glycogenin) and grows to form at least 12 tiers of
branched glucose residues forming a highly folded�-glycogen particle. Glucose residues are added toglycogen via UDP glucose through �1,4 linkagescatalyzed by the enzyme glycogen synthase andstimulated by insulin and G-6-P (Fig. 15). Branch-ing enzyme, which is not tightly regulated, addsbranch points to the growing glycogen molecule with�1,6 linkages (Fig. 15). Together with branchingenzyme, GS forms a low-molecular-weight (400 kDa)glycogen granule termed proglycogen that growslarger (up to 10,000 kDa) to form macroglycogen.190
Horses with PSSM1 were found to have high activ-ities of glycogen synthase (in basal and G6P-stimu-lated states), high branching enzyme activity, andan increase in both pro- and macro-glycogen concen-trations.81,186,189 The effect of the autosomal dom-inant mutation in the GYS1 gene encoding glycogensynthase is a unique gain of function.81 The high-er-than-normal activity in both basal states andwhen activated by insulin and G-6-P appears tohave a novel link to the regulation of energy gener-ation in muscle fibers during exercise under certaindietary conditions that increase insulin and muscleG-6-P concentrations.
Abnormal Polysaccharide FormationThe iodine absorption spectra of muscle samples fromPSSM1 horses indicated that the abnormal polysac-charide in PSSM1 horse muscle fibers has a lessbranched structure than normal glycogen.186 This isconfirmed by ultrastructural studies that indicate ab-normal polysaccharide consists of both beta glycogenparticles and filamentous material.96,168 The forma-tion of a less branched abnormal polysaccharide isprobably related to chronic enhanced activity of glyco-gen synthase enzyme in the presence of elevated G-6-Pand insulin concentrations without the same relativeactivation of branching enzyme.191 Over years, thisleads to a less highly branched polysaccharide, whichbecomes resistant to amylase digestion. When ab-normal polysaccharide accumulates in large amountsin muscle cells, it is associated with autophagicrimmed vacuoles179 and is tagged for degradation byubiquitin.192 Foals and yearlings with the GYS1 mu-tation rarely have abnormal polysaccharide in skeletalmuscle, yet they have rhabdomyolysis.80 Thus, ab-normal polysaccharide itself is an indicator of PSSM1but not the direct cause for rhabdomyolysis.
Diet and GYS1A high NSC diet increases the propensity to developmuscle pain with aerobic exercise in PSSM1horses.117,173,175 A high NSC versus low NSC, highfat diet results in higher muscle lactate and citrateconcentrations, lower G-6-P, and similar muscle py-ruvate concentrations with aerobic exercise inPSSM1 horses.175 This suggests that the enhancedglycogen synthase activity in PSSM horses may im-pair oxidative metabolism of substrates such as py-ruvate and fatty acids. While the exact mechanismfor this is not known, it may involve a link between
AAEP PROCEEDINGS � Vol. 58 � 2012 111
FRANK J. MILNE STATE-OF-THE ART LECTURE
F15
Orig. Op. OPERATOR: Session PROOF: PE’s: AA’s: 4/Color Figure(s) ARTNO:
1st disk, 2nd beb dainop 15 1,3-8,12,13,15-17 3353

glycogen synthase or glycogen concentrations andmetabolic switches and nutrient sensors.193–195
One of the key sensors and regulators of substrateflux in skeletal muscles is AMP kinase. When en-ergy supply is deemed to be insufficient (2 [ATP]/[ADP]), activation of AMP kinase leads to theincreased transcription of genes involved in oxida-tive metabolism, stimulation of fatty acid oxidation,and activation of pyruvate dehydrogenase, whichincreases carbohydrate oxidation.193–195 We spec-ulate that excessive stimulation of glycogen synthe-sis resulting from the GYS1 mutation (stimulatedfurther by insulin) is interpreted by nutrient sensorsin the cell as an indication not to activate glycogen-olysis and lipolysis. A potential scenario in PSSM1horses on high NSC diets could be that nutrientswitches do not fully activate enzymes such as py-ruvate dehydrogenase during exercise, limiting ad-equate acetyl CoA for oxidative metabolism (Fig. 5).
Although acetyl CoA could also be supplied by fattyacid oxidation, PSSM horses on high NSC diets havelow plasma nonesterified FFA concentrations, pos-sibly due to suppression of lipolysis by high insu-lin.173 A further factor reducing fatty acidoxidation in PSSM horses on high NSC diets couldbe that high muscle citrate concentrations directacetyl CoA away from the citric acid cycle towardmalonyl CoA, the committed step for fatty acid syn-thesis.196 In addition, accumulation of malonylCoA causes inhibition of carnitine palmitoyltrans-ferase, the key enzyme necessary to transport long-chain fatty acids into the mitochondria for�-oxidation.197,198 Thus, PSSM horses fed highNSC diets may be unable to generate sufficientacetyl CoA from either carbohydrate or fat metabo-lism to fuel muscle contraction during submaximalexercise. The common occurrence of clinical signsduring the first 20 minutes of exercise may be be-
Fig. 15. Glycogen synthesis begins with a protein (glycogenin) and grows to form at least 12 tiers of branched glucose residuesforming a highly folded � glycogen particle. Glucose residues are added to glycogen via UDP glucose through �1,4 linkages catalyzedby the enzyme glycogen synthase and stimulated by insulin and G-6-P. Branching enzyme adds branch points to the growingglycogen molecule with �1,6 linkages. Source: Wikipedia.
112 2012 � Vol. 58 � AAEP PROCEEDINGS
FRANK J. MILNE STATE-OF-THE ART LECTURE
Orig. Op. OPERATOR: Session PROOF: PE’s: AA’s: 4/Color Figure(s) ARTNO:
1st disk, 2nd beb dainop 15 1,3-8,12,13,15-17 3353

cause at this stage of exercise, muscles rely heavilyon glycogen/glucose for energy. Low dietary starchand fat supplementation decrease muscle citrateconcentrations and increase plasma FFA concentra-tions, which may well facilitate muscle fat metabo-lism in PSSM1 horses.175
Type 2 PSSMThere is much less known about type 2 PSSM be-cause as it turns out, previous research on PSSMhas largely involved horses with type 1 PSSM.Current knowledge of type 2 PSSM is based on ret-rospective evaluation of cases diagnosed with PSSMby muscle biopsy103 that are now known to be free ofthe GYS1 mutation and a few years of prospectiveclinical cases.
PrevalenceApproximately 28% of cases of PSSM diagnosed bymuscle biopsy in Quarter Horses have PSSM2, as theydo not have the GYS1 mutation.167 Type 2 PSSMseems to be common in both high-performance Quar-ter Horse types such as barrel racing, reining, andcutting horses as well as pleasure and halter horses.In the United Kingdom, approximately 35% of PSSMcases diagnosed by muscle biopsy have PSSM2.163
It also occurs in Paint, Appaloosa, and Morgan Horses.About 80% of cases of PSSM diagnosed by biopsy inWarmbloods have PSSM2.167 Breeds affected in-clude Dutch Warmbloods (highest prevalence), Swed-ish Warmbloods, Selle Francais, Hanoverians,Friesians, Westfalian, Canadian Warmblood, IrishSport Horse, Gerdlander, Hussien, and IcelandicHorses. Many other light breeds have also been di-agnosed with PSSM2, including Standardbreds andThoroughbreds. Type 2 PSSM also occurs in Arabi-ans; however, in my experience, this breed is distinctin that it often has amylase-resistant rather than am-ylase-sensitive polysaccharide but is negative for theGYS1 mutation.
PathogenesisThe cause of PSSM2 is currently unknown. Muscleglycogen concentrations are not as high as thosefound in type 1 PSSM, and glycogenolytic enzymedefects have not been identified in the few cases inwhich enzymes were measured (Valberg, unpub-lished data). To date, the only reported abnormal-ity in muscle ultrastructure is an increase in normalbeta glycogen particles.179 It may well be thatthere is a group of conditions currently capturedunder the term type 2 PSSM that have separateetiologies but share common findings of glycogenaccumulation and poor performance. A heritablepredisposition is suspected in Quarter Horses, butyet to be proven.
Acute Clinical SignsHorses with PSSM2 do not necessarily have thesame calm temperament as horses with PSSM1.In adults, acute clinical signs of rhabdomyolysis are
similar between type 1 and type 2 PSSM.163,167,168
Muscle atrophy after rhabdomyolysis is a commoncomplaint in Quarter Horses with PSSM2. Thereare more Quarter Horses less than 1 year of agereported with PSSM2 than PSSM1, and these foalsmay present with an inability to rise or a stiff hindlimb gait.168
Chronic Clinical Signs
Chronic signs of PSSM2 are often most closely re-lated to poor performance rather than recurrent ERand elevations in serum CK activity. An undiag-nosed gait abnormality, sore muscles, and drop inenergy level and willingness to perform after 5 to 10minutes of exercise are common complaints inhorses with PSSM2. Warmbloods with PSSM2may also have painful, firm back and hindquartermuscles, reluctance to collect, canter, and engagethe hindquarters, poor rounding over fences, gaitabnormalities, and slow onset of atrophy. Themean age of onset of clinical signs in Warmbloods isbetween 8 and 11 years of age, with the median CKand AST activity being 323 and 331 U/L, respec-tively, according to Neuromuscular Diagnostic Lab-oratory records.
Diagnosis
Type 2 PSSM must be diagnosed by muscle biopsywhere increased or abnormal PAS-positive materialthat is usually amylase-sensitive is apparent, par-ticularly in subsarcolemmal locations (Fig. 13, C andD).179 Determination of what constitutes an abnor-mal amount of amylase-sensitive glycogen is subjec-tive; the specificity of the diagnosis may be low, andfalse-positive diagnoses may occur. Considerationshould be made for state of training, as highlytrained horses have increased glycogen storages as anormal response to training. Further, glycogen iscommonly found near capillaries in normal horsesand PAS-positive sarcoplasmic masses can be foundin 60% of healthy horses, so these should not bediagnostic criteria for PSSM2.199 Formalin-fixedsections show a greater deposition of subsarcolem-mal glycogen and precipitation of glycogen in onearea of a fiber even in healthy horses, and theseshould not be considered diagnostic criteria forPSSM2.178 Other histopathologic features thatmay be present with PSSM2 include centrally lo-cated nuclei, subsarcolemmal vacuoles, musclenecrosis, macrophage infiltration of myofibers, re-generative fibers, and fiber atrophy.179 Some labo-ratories grade polysaccharide accumulation as mild,moderate, and severe, where mild accumulation rep-resents a category that has a higher chance of beinga false-positive diagnosis.98,163 Mild PSSM casesin particular should receive a full physical examina-tion to ensure that there are not other underlyingcauses for performance problems.
AAEP PROCEEDINGS � Vol. 58 � 2012 113
FRANK J. MILNE STATE-OF-THE ART LECTURE
Orig. Op. OPERATOR: Session PROOF: PE’s: AA’s: 4/Color Figure(s) ARTNO:
1st disk, 2nd beb dainop 15 1,3-8,12,13,15-17 3353

Management of Type 1 and Type 2 PSSMOwners must be aware that any horse diagnosedwith PSSM will always have an underlying predi-lection for muscle soreness. The best that can bedone is to manage horses in the most appropriatefashion to minimize clinical signs. With adherenceto both diet and exercise recommendations providedbelow, at least 70% of horses show notable improve-ment in clinical signs and many return to acceptablelevels of performance.103,117,162 There is, however,a wide range in the severity of clinical signs shownby horses with PSSM; those horses with severe orrecurrent clinical signs will require more stringentadherence to diet and exercise recommendations toregain muscle function.
RestPSSM horses that are confined for days to weeksafter an episode of rhabdomyolysis often have per-sistently elevated serum CK activity.80 In con-trast, PSSM horses kept on pasture with little grainsupplementation often show few clinical signs ofrhabdomyolysis and have normal serum CK activ-ity.80 As a result, a common recommendation forhorses with PSSM is to limit stall confinement toless than 48 hours after an episode of rhabdomyol-ysis and then provide turnout in paddocks of grad-ually increasing size. Providing horses with asmuch free exercise as possible on pasture appears tobe beneficial in the long term. If pasture is lush, agrazing muzzle may be needed.164 After an acuteepisode, excitable horses may require tranquiliza-tion prior to turnout to avoid excessive galloping.Hand-walking horses initially recovering from anepisode of PSSM for more than 5 to 10 minutes at atime may trigger another episode of rhabdomyolysis.
ExerciseThe beneficial response to low starch, fat supple-mented diets only occurs if it is instituted in con-junction with a regular incremental exerciseprogram.173 Regular daily exercise in PSSMhorses over a 3-week period has been shown to pro-duce a dramatic decrease in serum CK responses toexercise (Fig. 16), whereas stall confinement oftencauses elevations in serum CK activity after exer-cise.80,173 One common adaptation to daily train-ing is an increase in oxidative capacity in skeletalmuscle. The oxidative capacity of skeletal musclein PSSM Quarter Horses was found to be very low,based on markers such as citrate synthase activityand �-fatty acid oxidative marker HAD.173 The ac-tivity of these enzymes, however, was equally low incontrol Quarter Horses. Whether the beneficial ef-fect of daily exercise on PSSM horses is a result ofimproved oxidative enzyme capacities or enhancedsubstrate flux or both has not been elucidated.
Exercise RegimesImportant principles to follow when starting exer-cise programs in PSSM horses include (1) provide
adequate time for adaptation to a new diet prior tocommencing exercise, (2) recognize that the dura-tion of exercise, not the intensity, is of primary im-portance, (3) ensure that the program is graduallyintroduced and consistently performed, and (4) min-imize any days without some form of exercise. Ifhorses have experienced a moderate to marked epi-sode of rhabdomyolysis recently, 2 weeks of turnoutand diet change are often beneficial prior to recom-mencing exercise. Exercise should be very relaxed,and the horse should achieve a long, low frame with-out collection. For many horses, this is most read-ily done in a round pen or on a lunge-line but can bedone under saddle if needed. Successive daily ad-dition of 2-minute intervals of walk and trot begin-ning with only 4 minutes of exercise and working upto 30 minutes after 3 weeks is often recom-mended.105,177 If horses had minor elevations in4-hour postexercise CK with the 15-minute exercisetest, horses may begin at 15 minutes of exercise.Owners often do not recognize that walking thehorse for 10 minutes or more initially can triggermuscle soreness in PSSM horses. Advancing thehorse too quickly often results in an episode of rhab-domyolysis and repeated frustration for the owner.Work can usually begin under saddle after 3 weeksof ground work and can gradually be increased byadding 2-minute intervals of collection or canter tothe initial relaxed warm-up period at a walk andtrot. Unless a horse shows an episode of overtrhabdomyolysis during the initial first 4 weeks ofexercise, reevaluating serum CK activity is not help-ful for the first month. This is because it is verycommon to have subclinical elevations in CK activitywhen exercise is re-introduced, and a return to nor-mal levels often requires 4 to 6 weeks of gradualexercise.177 Keeping horses with PSSM fit seems
Fig. 16. Three isocaloric diets were formulated to provide a totaldigestible energy of 50 kCal/kg per day either in the form of haysupplemented with grain, corn oil on hay cubes (1.5 mL/kg BWper day), or 2.5 kg of a low starch, high fat concentratea.200 Theoil provided 30% of digestible energy (DE) as fat, whereas thehigh fat low NSC concentrate provided 15% of DE as fat. PSSM1horses (n � 8) were fed the diets for 3 weeks each and exercisedfor up to 20 minutes at a walk and trot 5 days per week. Notethat 15% DE as fat significantly reduced serum CK activity inPSSM horses that were exercised and that it was not necessary toprovide more than 25% of DE as fat to have a significantly lowerserum CK activity relative to grain.
114 2012 � Vol. 58 � AAEP PROCEEDINGS
FRANK J. MILNE STATE-OF-THE ART LECTURE
F16
Orig. Op. OPERATOR: Session PROOF: PE’s: AA’s: 4/Color Figure(s) ARTNO:
1st disk, 2nd beb dainop 15 1,3-8,12,13,15-17 3353

the best prevention against further episodes ofrhabdomyolysis.
Diets for PSSMThe basis for designing PSSM diets is the belief thatlowering the daily starch and sugar intake and in-creasing dietary fat content will decrease the glu-cose load, increase the availability of NEFA formuscle metabolism, and lower serum insulin con-centrations.173,200,201 Quarter Horses naturallyhave very little lipid stored within muscle fibers.The end result would thus be decreased uptake ofglucose into muscles cells, less substrate availablefor—and stimulation of—glycogen synthesis andnormalization of substrate flux. Owners reportthat this type of diet improves clinical signs of mus-cle pain, stiffness, and exercise tolerance in drafthorses, Warmbloods, Quarter Horses, and otherbreeds.103,104,117,162 Dietary change appears tohave lesser impact on alleviating gait changes suchas shivers.103
Caloric BalanceThe horse’s caloric requirements at an ideal bodyweight are the most important considerations indesigning the diet for PSSM (Table 3). This is be-cause many horses with PSSM are easy keepers andmay be overweight at the time of diagnosis. Addingexcessive calories in the form of fat to an obese horsemay produce metabolic syndrome and is contraindi-cated. If necessary, caloric intake should be re-duced by using a grazing muzzle during turnout,feeding hay with a low NSC content at 1% to 1.5% ofbody weight, providing a low-calorie ration balancer,and gradually introducing daily exercise. Ratherthan provide dietary fat to an overweight horse,fasting for 6 hours before exercise can be used toelevate plasma FFAs prior to exercise and alleviateany restrictions in energy metabolism in muscle.
Selection of ForageQuarter Horses have been shown to develop a sig-nificant increase in serum insulin concentrations in
response to consuming hay with an NSC of 17%,whereas insulin concentrations are fairly stablewhen fed hay with 12% or 4% NSC content.175
Because insulin stimulates the already overactiveenzyme glycogen synthase in the muscle of PSSM1horses, selecting hay with 12% or less NSC is advis-able. The degree to which the NSC content of hayshould be restricted below 12% NSC depends on thecaloric requirements of the horse. Feeding a lowNSC hay of 4% provides room to add an adequateamount of fat to the diet of easy keepers withoutexceeding the daily caloric requirement and induc-ing excessive weight gain. For example, a 500-kghorse on a routine of light exercise generally re-quires 18 MCal/d of DE. Fed at 2% of body weight,a 12% NSC mixed grass hay almost meets theirdaily caloric requirement by providing 17.4 MCal/d.Thus with a 12% NSC hay, there is only room for0.6 MCal of fat per day (72 mL of vegetable oil) toachieve 18 MCal of energy. In contrast, a 4% NSCBlue Grama hay would provide 13.5 MCal/d, whichwould allow a reasonable addition of 4.5 MCal of fatper day (538 mL of vegetable oil).
Selection of Fat SourceHigh fat diets increase plasma FFA concentrationsand thus the availability of fats for oxidation inskeletal muscle.173 Long-chain fat diets also ap-pear to increase glycogenolytic/glycolytic and oxida-tive flux in PSSM1 muscle as shown by higherG-6-P, lower lactate, and higher pyruvate concentra-tions in muscle of PSSM1 horses fed and trained ona commercial high fat dieta compared with sweetfeed.175 Thus, one means to overcome limitationsin delivery of oxidative substrates in PSSM1 horsesis to provide ample long-chain fat in the diet. Themajor sources of dietary fat for horses are vegetable-based, including vegetable oils and rice bran oranimal based fat (tallow, lard, fish oil). Vegetableoils are highly unsaturated, very digestible (90%to 100%), and very energy-dense. Suitable formsinclude soybean, corn, safflower, canola, flaxseed,linseed, peanut, and coconut. Controlled research
Table 3. Feeding Recommendations for an Average-Sized 500-kg Horse With Exertional Rhabdomyolysis Due to Recurrent Exertional Rhabdomy-olysis or Polysaccharide Storage Myopathy
RER PSSM
Exercise intensity Light Moderate Intense Light Moderate IntenseCaloric intake 20 25 33 18 23 30Percentage of daily digestible energy as NSC 15–20 15–20 15–20 �10 �10 �10Percentage of daily digestible energy as fat 15 15–20 20–25 15 15–20 15–20Selenium, mg/d 2.2 2.8 3.1 2.2 2.8 3.1Vitamin E,* mg/d 1,400 1,800 2,000 1,400 1,800 2,000Daily amount, kg, of 12% NSC, 10% fat concentrate† 2.5 3.5 5 2 3 4Grass hay, kg, with �12% NSC by weight 7.5–10 7.5–10 7.5–10 7.5–10 7.5–10 7.5–10Sodium chloride, g 34 34 41 34 34 41
RER indicates recurrent exertional rhabdomyolysis; PSSM, polysaccharide storage myopathy.*Natural vitamin is preferable to synthetic vitamin E due to better absorption.†Concentratea for horses with exertional myopathies
AAEP PROCEEDINGS � Vol. 58 � 2012 115
FRANK J. MILNE STATE-OF-THE ART LECTURE
T3
Orig. Op. OPERATOR: Session PROOF: PE’s: AA’s: 4/Color Figure(s) ARTNO:
1st disk, 2nd beb dainop 15 1,3-8,12,13,15-17 3353

studies in exercising PSSM horses have shown adecrease in muscle pain and serum CK in responseto the addition of corn oil117,162,175 and also soybeanoil and rice brana (Fig. 16).173,175 The amount of oiladded in these trials constituted at least 13% ofdaily DE. Some veterinarians have advocated asmuch as 25% of DE be fed in the form of fat to PSSMhorses.118 As discussed above, the principle consid-eration here should be whether this provides exces-sive calories and additional weight gain becausefeeding 13% DE as fat may well be effective in re-ducing muscle pain.
Limited research has been performed on the formof oil to feed PSSM horses. An odd carbon 7 chainfat (triheptanoin) fed to PSSM horses had a detri-mental effect on muscle pain, exercise tolerance, andserum CK activity, whereas in the same study, long-chain fats fed in the form of corn oil or a rice bran/soy oil supplemented feed had a beneficial effect onlowering serum CK activity.175 Whether there isany direct beneficial effect on skeletal muscle ofproviding energy in the form of Omega 3 versusOmega 6 fatty acids has yet to be determined.Corn oil, sunflower oil, and safflower oil are high inOmega-6 and lower in Omega-3, whereas soybeanand canola oils are moderately high in Omega-6 andOmega-3, and flax seed (linseed) and fish oils con-tain more Omega-3 than Omega-6. It is usuallycost-prohibitive to provide sufficient energy to aPSSM horse each day in the form of dense Omega 3fat supplements. Soybean and canola oils provide arelatively affordable alternative with moderatelyhigh Omega-6, or a mix of these oils and flax or fishoil can be provided. Due to the potential additionaloxidant stress of fats, vitamin E (1000 t 5000 U/d)should be fed to horses receiving high oil diets.
Low Starch, High Fat ConcentratesWhile oils are energy-dense and inexpensive, theyhave the disadvantages of being messy, unpalatableto some horses, prone to rancidity in warm temper-atures, and difficult to feed in large amounts. Assuch, a number of concentrates have been developedthat contain fats. The important principle to bemet by such feeds is that the starch and sugar com-ponents are low enough and fat supplementationhigh enough to ensure that in the total diet, thecalories supplied by NSC comprise no more than10% to 15% of the daily DE and the calories suppliedby fat comprise about 12% to 15% of daily DE.142,173
Common fat sources used in such concentrates in-clude, in addition to the oils mentioned above, sta-bilized rice bran or animal fats. Rice bran and itsproducts are palatable to most horses, have a mod-erate NSC content �25% by weight, contain �20%fat by weight as well as vitamin E, and are naturallyhigh in phosphorus.142 The NSC component of ricebran can vary if the manufacturing process is notcareful to exclude the white rice grain. Commercialrice bran products occur as powder or an extrudedpellet and are considerably more stable in warm
temperatures compared with animal fat and vegeta-ble oils. A number of well-balanced low starch,high fat commercial diets are suitable for horseswith PSSM.142,173 Some commercial feeds meetthe recommended nutritional needs of PSSM horsesin one pelleted ration. These feeds typically con-tain at least 10% to 15% fat by weight and less than20% NSC by weight. Some feed companies offersimilar nutritional content by blending two or moremanufactured feeds or by supplementing with oils orrice bran. At present, the NSC content of equinefeed products is not consistently listed on the feedtag, and consultation with the feed manufacturer isnecessary to obtain this information. Nutritionalsupport is available through most feed manufactur-ers in designing an appropriate diet. There is agreat deal of variation in individual tolerance todietary starch; however, horses with more severeclinical signs of PSSM appear to require the greatestrestriction in starch intake.
ExpectationsThe beneficial effect of low starch, high fat dietsrequire that horses are trained daily to enhanceenzymes involved in fat and glucose metabolism.It is important to note that a horse diagnosed withPSSM will always have an underlying predilectionfor muscle soreness, and the best that can be done isto manage horses to minimize clinical signs. Withadherence to both the diet and exercise recommen-dations 70% to 75% of Quarter Horses and Warm-bloods show notable improvement in clinical signs,and many return to acceptable levels of perfor-mance.103,118,162,164 There is, however, a widerange in the severity of clinical signs shown byhorses with PSSM; those horses with severe or re-current clinical signs will require more stringentadherence to diet and exercise recommendations toregain muscle function. PSSM horses that alsohave the mutation for malignant hyperthermia donot respond as well to diet and exercise recommen-dations and may continue to develop ER with thepossibility of a fatal episode.174
SupplementsThere are no specific supplements that have beenshown to benefit horses with PSSM. Chromium,which is reported to increase glucose absorption,may be contraindicated in PSSM horses. Carnitineis a dietary supplement that may replenish musclecarnitine stores if depleted and assist in transport offat into the mitochondria. A deficiency of plasmacarnitine has not been identified in PSSM1 horses(Valberg, unpublished data; PSSM1 plasma-freecarnitine; PSSM1 range, 10.24 to 27.11 �M; normalrange, 13.20 to 25.63 �M).
VI. Mitochondrial Myopathies
Disorders or aerobic metabolism reported to date inhorses have not been characterized by rhabdomyol-ysis but rather most profoundly by exercise intoler-
116 2012 � Vol. 58 � AAEP PROCEEDINGS
FRANK J. MILNE STATE-OF-THE ART LECTURE
Orig. Op. OPERATOR: Session PROOF: PE’s: AA’s: 4/Color Figure(s) ARTNO:
1st disk, 2nd beb dainop 15 1,3-8,12,13,15-17 3353
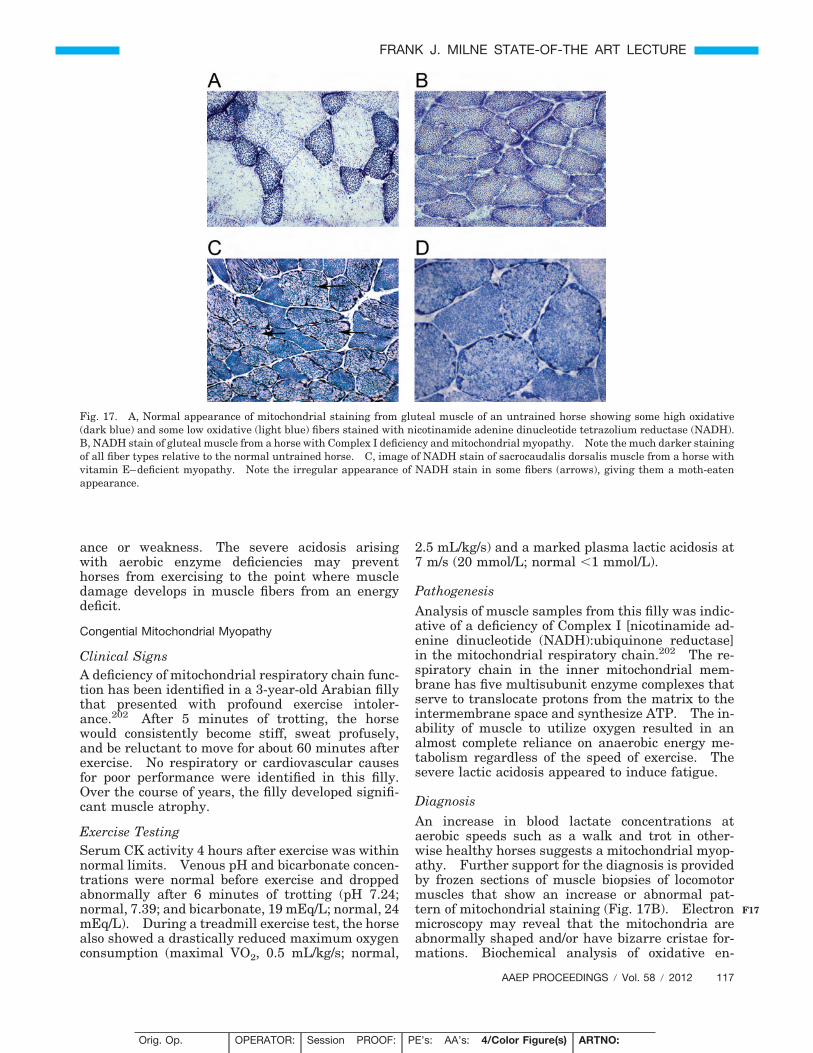
ance or weakness. The severe acidosis arisingwith aerobic enzyme deficiencies may preventhorses from exercising to the point where muscledamage develops in muscle fibers from an energydeficit.
Congential Mitochondrial Myopathy
Clinical SignsA deficiency of mitochondrial respiratory chain func-tion has been identified in a 3-year-old Arabian fillythat presented with profound exercise intoler-ance.202 After 5 minutes of trotting, the horsewould consistently become stiff, sweat profusely,and be reluctant to move for about 60 minutes afterexercise. No respiratory or cardiovascular causesfor poor performance were identified in this filly.Over the course of years, the filly developed signifi-cant muscle atrophy.
Exercise TestingSerum CK activity 4 hours after exercise was withinnormal limits. Venous pH and bicarbonate concen-trations were normal before exercise and droppedabnormally after 6 minutes of trotting (pH 7.24;normal, 7.39; and bicarbonate, 19 mEq/L; normal, 24mEq/L). During a treadmill exercise test, the horsealso showed a drastically reduced maximum oxygenconsumption (maximal VO2, 0.5 mL/kg/s; normal,
2.5 mL/kg/s) and a marked plasma lactic acidosis at7 m/s (20 mmol/L; normal �1 mmol/L).
PathogenesisAnalysis of muscle samples from this filly was indic-ative of a deficiency of Complex I [nicotinamide ad-enine dinucleotide (NADH):ubiquinone reductase]in the mitochondrial respiratory chain.202 The re-spiratory chain in the inner mitochondrial mem-brane has five multisubunit enzyme complexes thatserve to translocate protons from the matrix to theintermembrane space and synthesize ATP. The in-ability of muscle to utilize oxygen resulted in analmost complete reliance on anaerobic energy me-tabolism regardless of the speed of exercise. Thesevere lactic acidosis appeared to induce fatigue.
DiagnosisAn increase in blood lactate concentrations ataerobic speeds such as a walk and trot in other-wise healthy horses suggests a mitochondrial myop-athy. Further support for the diagnosis is providedby frozen sections of muscle biopsies of locomotormuscles that show an increase or abnormal pat-tern of mitochondrial staining (Fig. 17B). Electronmicroscopy may reveal that the mitochondria areabnormally shaped and/or have bizarre cristae for-mations. Biochemical analysis of oxidative en-
Fig. 17. A, Normal appearance of mitochondrial staining from gluteal muscle of an untrained horse showing some high oxidative(dark blue) and some low oxidative (light blue) fibers stained with nicotinamide adenine dinucleotide tetrazolium reductase (NADH).B, NADH stain of gluteal muscle from a horse with Complex I deficiency and mitochondrial myopathy. Note the much darker stainingof all fiber types relative to the normal untrained horse. C, image of NADH stain of sacrocaudalis dorsalis muscle from a horse withvitamin E–deficient myopathy. Note the irregular appearance of NADH stain in some fibers (arrows), giving them a moth-eatenappearance.
AAEP PROCEEDINGS � Vol. 58 � 2012 117
FRANK J. MILNE STATE-OF-THE ART LECTURE
F17
Orig. Op. OPERATOR: Session PROOF: PE’s: AA’s: 4/Color Figure(s) ARTNO:
1st disk, 2nd beb dainop 15 1,3-8,12,13,15-17 3353

zymes is needed to identify a deficiency in oxidativeenzymes or respiratory chain malfunction.
TreatmentThere is no known treatment for congenital mito-chondrial myopathies.
Acquired Mitochondrial Abnormality and Vitamin EDeficiency
Clinical SignsAn acquired disorder affecting mitochondria has re-cently been reported in horses with low muscle vita-min E concentrations.203 Horses show a decrease inperformance due to exercise intolerance, generalizedlocomotor muscle wasting, muscle fasciculations, andweakness. The distinction between this disorder andequine motor neuron disease is the presence of myo-pathic rather than neuropathic features in sacrocau-dalis dorsalis muscle biopsies and the remarkableresponse to treatment with vitamin E.
Exercise TestingExercise testing beyond a neurologic examinationhas not been reported for acquired cases associatedwith low vitamin E. This is because horses oftenpresent with such severe weakness that it is notclinically practical to perform.
PathogenesisThe observed generalized weakness in the horseswith vitamin E–deficient myopathy is suggested tobe due to a reversible manifestation of skeletal mus-cle mitochondrial oxidative stress associated withvitamin E deficiency. Vitamin E–deficient myop-athy may be an entity unto itself or a predecessor todevelopment of equine motor neuron disease.
DiagnosisAcquired vitamin E–deficient myopathy is diag-nosed by confirming low serum or muscle vitamin Econcentrations and identifying a moth-eaten stain-ing pattern in frozen biopsies of the sacrocaudalisdorsalis muscle (Fig. 17C). Mitochondrial stains ofmuscle such as the gluteus medius are normal,whereas highly oxidative muscles such as the sacro-caudalis show an irregular abnormal moth-eatenstaining pattern for mitochondria. In some horses,serum vitamin E may be normal, whereas musclevitamin E may be low.
TreatmentAcquired vitamin E–deficient myopathy is highlyresponsive to treatment with natural vitamin E(5000 IU/d per 500-kg horse) and a very gradualreintroduction to exercise once strength returns.203
Conclusions
The past 30 years have offered exciting discoveriesof several new equine-specific muscle diseases,many of which manifest exclusively with exercise.
I hope within this review that I have paid tribute tothe many clinician scientists past and present whosefascination with muscle disorders provided thebuilding blocks for these advances and inspiredother young scientists to take up the call. As ourknowledge of equine exercise physiology, muscle his-topathology, biochemistry, and genetics continues toexpand and young clinician scientists are drawn tothe field, other new discoveries are likely imminent.
I received amazing mentoring during my PhD inUppsala, Sweden, by Birgitta Essen-Gustavssonand Sune Persson, who taught me the discipline andthe thrill of science. Drs. Arne Lindholm, GaryCarlson, and John Madigan shaped my vision of theclinician scientists, melding clinical medicine withbasic sciences for the betterment of the horse. Dr.Cardinet III shared with me his insights into whatmuscles look like from the inside looking outwardand how to run a diagnostic service with the higheststandards. I have had many essential collabora-tors over the years that I am indebted to. Dr. JohnBaird and Dr. Billi DiMauro have been staunch sup-porters and invaluable resources for more than 20years. I have a huge debt of gratitude to my friendand colleague Jim Mickelson, who has been at thecalm and reasoned core of all of the muscle workperformed at the University of Minnesota and to myultra-organized technician Michelle Lucio. I havebeen inspired and encouraged by my many academicdaughters and sons whose sharp minds, diligentgraduate work, and humor have made coming towork fun. My family has been my rock throughstormy times; to John, you make life worth living; tomy son Niels, you are the light of my life; and to mydaughter Annika, I miss you every single day.
Conflict of interest statement: Drs. Valberg, Mickelson,and McCue own the license for PSSM testing and receive salesincome from its use. Their financial and business interests havebeen reviewed and managed by the University in accordance withits conflict of interest policies. The University of Minnesota haslicensed the sale of Re-Leve and Dr. Valberg receives royaltiesfrom Kentucky Equine Research.
References and Footnote1. Robertson W. azoturia. A Textbook of the Practice of
Equine Medicine. London: Bailliere, Tindall and Cox;1883:368–381.
2. Carlstrom B. Uber die atiologie und pathogenese der kreu-zlahme des pferdes (Haemaglobinaemia paralytica). ScandArchiv 1932;62:1–62.
3. McLean JG. Equine paralytic myoglobinuria (“azoturia”):a review. Aust Vet J 1973;49:41–43.
4. Carlstrom B. Uber die atiologie und pathogenese dePferdes (Haemoglobinaemia paralytica). Scand ArchivPhysiol 1931;61:161–224.
5. Steffin MR. Special Equine Therapy. Chicago: Ameri-can Veterinary Publishing Co; 1917.
6. Williams W. Azoturia. Edinburgh: John Menzies andCo; 1884:435–440.
7. Meginnis P. Myositis (tying up) in race horses. J Am VetMed Assoc 1957;130:237–239.
8. Brennan BF, Marshak RR, Keown GHAOR, Lundvall RL,Churchill EA. Panel discussion-tying up syndrome, in Pro-ceedings. Am Assoc Equine Pract 1959;6:157–169.
118 2012 � Vol. 58 � AAEP PROCEEDINGS
FRANK J. MILNE STATE-OF-THE ART LECTURE
Orig. Op. OPERATOR: Session PROOF: PE’s: AA’s: 4/Color Figure(s) ARTNO:
1st disk, 2nd beb dainop 15 1,3-8,12,13,15-17 3353

9. Farrow WA, Roloff DH, Westman CW, et al. Treatment forazoturia and tying-up. Mod Vet Pract 1976;57:413–416.
10. Koterba A, Carlson GP. Acid-base and electrolyte altera-tions in horses with exertional rhabdomyolysis. J Am VetMed Assoc 1982;180:303–306.
11. Roneus B, Hakkarainen J. Vitamin E in serum and skel-etal muscle tissue and blood glutathione peroxidase activityfrom horses with the azoturia-tying-up syndrome. Acta VetScand 1985;26:425–427.
12. Waldron-Mease E. Hypothyroidism and myopathy in rac-ing Thoroughbreds and Standardbreds. J Equine Vet Sci1979;3:124–128.
13. Harris P, Marlin D, Gray J. Equine thyroid function tests:a preliminary investigation. Br Vet J 1992;148:71–80.
14. Vischer CM, Foreman JH, Constable PD, et al. Hemody-namic effects of thyroidectomy in sedentary horses. Am JVet Res 1999;60:14–21.
15. Bain FT, Merritt AM. Decreased erythrocyte potassiumconcentration associated with exercise-related myopathy inhorses. J Am Vet Med Assoc 1990;196:1259–1261.
16. Beech J, Lindborg S, Braund KG. Potassium concentra-tions in muscle, plasma and erythrocytes and urinary frac-tional excretion in normal horses and those with chronicintermittent exercise-associated rhabdomyolysis. Res VetSci 1993;55:43–51.
17. Foreman JH. The exhausted horse syndrome. Vet ClinNorth Am Equine Pract 1998;14:205–219.
18. Carlson GP. Medical Problems Associated With ProtractedHeat and Work Stress in Horses, in Proceedings. Am AssocEquine Sports Med. Reno, NV. 1985:84–99.
19. Valberg S, Gustavsson BE, Lindholm A, et al. Blood chem-istry and skeletal muscle metabolic responses during andafter different speeds and durations of trotting. Equine VetJ 1989;21:91–95.
20. Harris P, Colles C. The use of creatinine clearance ratiosin the prevention of equine rhabdomyolysis: a report offour cases. Equine Vet J 1988;20:459–463.
21. Harris PA, Snow DH. Role of electrolyte imbalances inthe pathophysiology of the equine rhabdomyolysis syndrome.In: Persson S, Lindholm A, Jeffcott LB, editors. Equine Ex-ercise Physiology. 3rd edition. Davis, CA: ICEEP Publica-tions; 1991:435–442.
22. McKenzie EC, Valberg SJ, Godden SM, et al. Comparisonof volumetric urine collection versus single-sample urinecollection in horses consuming diets varying in cation-anionbalance. Am J Vet Res 2003;64:284–291.
23. McKenzie EC, Valberg SJ, Godden SM, et al. Plasma andurine electrolyte and mineral concentrations in Thorough-bred horses with recurrent exertional rhabdomyolysis afterconsumption of diets varying in cation-anion balance. Am JVet Res 2002;63:1053–1060.
24. Lindholm A, Piehl K. Fibre composition, enzyme activityand concentrations of metabolites and electrolytes in mus-cles of Standardbred horses. Acta Vet Scand 1974;15:287–309.
25. Snow DH, Guy PS. Percutaneous needle muscle biopsy inthe horse. Equine Vet J 1976;8:150–155.
26. Rivero JL, Serrano AL, Diz AM, et al. Changes in cross-sectional area and capillary supply of the muscle fiber pop-ulation in equine gluteus medius muscle as a function ofsampling depth. Am J Vet Res 199354:32–37.
27. Gould GW, Bell GI. Facilitative Glucose Transporters:An Expanding Family. Amsterdam: Elsevier; 1990:18–23.
28. van Dam KG, van Breda E, van Ginneken MM, et al.Investigation of the expression of glucose transporter 4 andfatty acid translocase/CD36 in equine skeletal mus-cle. Am J Vet Res 2004 1;65:951–956.
29. Kiens B. Skeletal muscle lipid metabolism in exercise andinsulin resistance. Physiol Rev 2006;86:205–243.
30. Snow DH, Guy PS. Fiber type and enzyme activities of thegluteus medius muscle of different breeds of horse. In: Poort-mans J, Nisert G, editors. Biochemistry of Exercise. Balti-more: University Park Press; 1981:275.
31. Gorza L. Identification of a novel type 2 fiber population inmammalian skeletal muscle by combined use of histochem-ical myosin ATPase and anti-myosin monoclonal antibodies.J Histochem Cytochem 1990;38:257–265.
32. Dingboom EG, Dijkstra G, Enzerink E, et al. Postnatalmuscle fibre composition of the gluteus medius muscle ofDutch Warmblood foals: maturation and the influence ofexercise. Equine Vet J Suppl 1999;31:95–100.
33. Eizema K, van den Burg MM, de Jonge HW, et al. Myosinheavy chain isoforms in equine gluteus medius mus-cle: comparison of mRNA and protein expression profiles.J Histochem Cytochem 2005;53:1383–1390.
34. Rivero JL, Talmadge RJ, Edgerton VR. A sensitive electro-phoretic method for the quantification of myosin heavychain isoforms in horse skeletal muscle: histochemical andimmunocytochemical verifications. Electrophoresis 1997;18:1967–1972.
35. Snow DH, Valberg SJ. Muscle-anatomy: adaptations toexercise and training. In: Rose RJ, Hodgson DH, editors.The Athletic Horse. 1st edition. 1994:145–179.
36. Kayar SR, Hoppeler H, Essen-Gustavsson B, SchwerzmannK. The similarity of mitochondrial distribution in equineskeletal muscles of differing oxidative capacity. J Exp Biol1988;137:253–263.
37. Essen-Gustavsson B, Karlstrom K, Lindholm A. Fibretypes, enzyme activities and substrate utilisation in skeletalmuscles of horses competing in endurance rides. EquineVet J 1984;16:197–202.
38. Essen-Gustavsson B, Lindholm A, Persson S, et al. Skele-tal muscle characteristics of young Standardbreds in rela-tion to growth and early training. In: Snow DH, PerssonS, Rose RJ, editors. Equine Exercise Physiology. Am-bridge: Granta Editions; 1983:200–210.
39. Hodgson DR, Rose RJ, DiMauro J, et al. Effects of a sub-maximal treadmill training programme on histochemicalproperties, enzyme activities and glycogen utilisation ofskeletal muscle in the horse. Equine Vet J 1985;17:300–305.
40. Snow DH, Guy PS. The effect of training and detraining onseveral enzymes in horse skeletal muscle. Arch Int PhysiolBiochim 1979;87:87–93.
41. Essen-Gustavsson B, Lindholm A. Muscle fibre character-istics of active and inactive Standardbred horses. EquineVet J 1985;17:434–438.
42. Essen-Gustavsson B, McMiken D, Karlstrom K, et al.Muscular adaptation of horses during intensive training anddetraining. Equine Vet J 1989;21:27–33.
43. Valberg S, Essen GB, Skoglund WH. Oxidative capacity ofskeletal muscle fibres in racehorses: histochemical versusbiochemical analysis. Equine Vet J 1988;20:291–295.
44. Valberg SJ, Essen-Gustavsson B. Metabolic response toracing determined in pools of type I, IIA and IIB fibers. In:Gillespie JR, Robinson NE, editors. Equine Exercise Phys-iology 2. Davis, CA: ICEEP Publications; 1987:209–301.
45. Valberg S. Glycogen depletion patterns in the muscle ofStandardbred trotters after exercise of varying intensitiesand durations. Equine Vet J 1986;18:479–484.
46. Lindholm A, Bjerneld H, Saltin B. Glycogen depletion pat-tern in muscle fibres of trotting horses. Acta Physiol Scand1974;90:475–484.
47. Hodgson DR, Rose RJ, Allen JR, et al. Glycogen depletionpatterns in horses performing maximal exercise. Res VetSci 1984;36:169–173.
48. Geor RJ, McCutcheon LJ, Hinchcliff KW, et al. Training-induced alterations in glucose metabolism during moderate-intensity exercise. Equine Vet J Suppl 2002;34:22–28.
49. Geor RJ, Hinchcliff KW, Sams RA. Glucose infusion atten-uates endogenous glucose production and enhances glucoseuse of horses during exercise. J Appl Physiol 2000;88:1765–1776.
50. Rennie MJ, Bohe J, Smith K, et al. Branched-chain aminoacids as fuels and anabolic signals in human muscle. JNutr 2006;136(1 Suppl):264S–268S.
AAEP PROCEEDINGS � Vol. 58 � 2012 119
FRANK J. MILNE STATE-OF-THE ART LECTURE
Orig. Op. OPERATOR: Session PROOF: PE’s: AA’s: 4/Color Figure(s) ARTNO:
1st disk, 2nd beb dainop 15 1,3-8,12,13,15-17 3353

51. Harris RC, Marlin DJ, Snow DH, et al. Muscle ATP lossand lactate accumulation at different work intensities in theexercising Thoroughbred horse. Eur J Appl Physiol OccupPhysiol 1991;62:235–244.
52. Valberg S, Essen-Gustavsson B, Lindholm A, et al. Energymetabolism in relation to skeletal muscle fibre propertiesduring treadmill exercise. Equine Vet J 1985;17:439–444.
53. Hodgson DR. Muscular adaptations to exercise and train-ing. Vet Clin North Am Equine Pract 1985;1:533–548.
54. Hodgson DR, Rose RJ. Effects of a nine-month endurancetraining programme on muscle composition in thehorse. Vet Rec 1987;121:271–274.
55. Hodgson DR, Rose RJ, DiMauro J, et al. Effects of trainingon muscle composition in horses. Am J Vet Res 1986;47:12–15.
56. Roneus M, Essen-Gustavsson B, Aranson T. Racing perfor-mance and longitudinal changes in muscle characteristics inStandardbred trotters. J Equine Vet Sci 1993;13:355.
57. Valberg SJ. Metabolic response to racing and fiber proper-ties of skeletal muscle in Standardbred and Thoroughbredhorses. J Equine Vet Sci 1987;7:6.
58. Lacombe VA, Hinchcliff KW, Kohn CW, et al. Effects offeeding meals with various soluble-carbohydrate content onmuscle glycogen synthesis after exercise in horses. Am JVet Res 2004;65:916–923.
59. Lacombe V, Hinchcliff KW, Geor RJ, et al. Exercise thatinduces substantial muscle glycogen depletion impairs sub-sequent anaerobic capacity. Equine Vet J Suppl 1999;30:293–297.
60. Snow DH, Baxter P, Rose RJ. Muscle fibre compositionand glycogen depletion in horses competing in an enduranceride. Vet Rec 1981;108:374–378.
61. Sewell DA, Harris RC. Adenine nucleotide degradation inthe Thoroughbred horse with increasing exercise duration.Eur J Appl Physiol Occup Physiol 1992;65:271–277.
62. Cutmore CM, Snow DH, Newsholme EA. Effects of train-ing on enzyme activities involved in purine nucleotide me-tabolism in Thoroughbred horses. Equine Vet J 1986;18:72–73.
63. Harris RC, Marlin DJ, Snow DH. Metabolic response tomaximal exercise of 800 and 2,000 m in the Thoroughbredhorse. J Appl Physiol 1987;63:12–19.
64. Marlin DJ, Harris RC. Titrimetric determination of musclebuffering capacity (beta mtitr) in biopsy samples. Equine VetJ 1991;23:193–197.
65. Essen-Gustavsson B, Henriksson J. Enzyme levels inpools of microdissected fibres of identified type: adaptiveresponse to exercise. Acta Physiol Scand 1984;120:505–515.
66. Essen-Gustavsson B, Roneus N, Poso AR. Metabolic re-sponse in skeletal muscle fibres of Standardbred trottersafter racing. Comp Biochem Physiol B Biochem Mol Biol1997;117:431–436.
67. Noakes TD, St Clair GA, Lambert EV. From catastrophe tocomplexity: a novel model of integrative central neuralregulation of effort and fatigue during exercise in humans.Br J Sports Med 2004;38:511–514.
68. Noakes TD, St Clair GA. Logical limitations to the “catas-trophe” models of fatigue during exercise in humans. Br JSports Med 2004;38:648–649.
69. Rivero JL. A scientific background for skeletal muscle con-ditioning in equine practice. J Vet Med A Physiol PatholClin Med 2007;54:321–332.
70. Cutmore CM, Snow DH, Newsholme EA. Activities of keyenzymes of aerobic and anaerobic metabolism in middlegluteal muscle from trained and untrained horses. EquineVet J 1985;17:354–356.
71. Schuback K, Essen-Gustavsson B, Persson SG. Incremen-tal treadmill exercise until onset of fatigue and its relation-ship to metabolic response and locomotion pattern. EquineVet J Suppl 1999;30:337–341.
72. Dubowitz V, Brooke MH. Muscle Biopsy: A Modern Ap-proach. Philadelphia: WB Saunders; 1973.
73. DiMauro S, Tsujino S, Shanske S, et al. Biochemistry andmolecular genetics of human glycogenoses: an over-view. Muscle Nerve 1995;3:S10–S17.
74. Cardinet GH III, Holliday TA. Neuromuscular diseases ofdomestic animals: a summary of muscle biopsies from 159cases. Ann N Y Acad Sci 1979;317:290–313.
75. Cornelio F, Donato S. Myopathies due to enzyme deficien-cies. J Neurol 1985;232:329–340.
76. Collinder E, Lindholm A, Rasmuson M. Genetic markersin Standardbred trotters susceptible to the rhabdomyolysissyndrome. Equine Vet J 1997;29:117–120.
77. Valberg SJ, Geyer C, Sorum SA, et al. Familial basis ofexertional rhabdomyolysis in Quarter Horse-related breeds.Am J Vet Res 1996;57:286–290.
78. MacLeay JM, Valberg SJ, Sorum SA, et al. Heritability ofrecurrent exertional rhabdomyolysis in Thoroughbred race-horses. Am J Vet Res 1999;60:250–256.
79. Dranchak PK, Valberg SJ, Onan GW, et al. Inheritanceof recurrent exertional rhabdomyolysis in Thoroughbreds.J Am Vet Med Assoc 2005;227:762–767.
80. De La Corte FD, Valberg SJ, MacLeay JM, Mickelson JR.Developmental onset of polysaccharide storage myopathy in4 Quarter Horse foals. J Vet Intern Med 2002;16:581–587.
81. McCue ME, Valberg SJ, Miller MB, et al. Glycogen syn-thase (GYS1) mutation causes a novel skeletal muscle gly-cogenosis. Genomics 2008;91:458–466.
82. Rudolph JA, Spier SJ, Byrns G, et al. Periodic paralysis inQuarter Horses: a sodium channel mutation disseminatedby selective breeding. Nat Genet 1992;2:144–147.
83. Aleman M, Nieto JE, Magdesian KG. Malignant hyper-thermia associated with ryanodine receptor 1 (C7360G) mu-tation in Quarter Horses. J Vet Intern Med 2009;23:329–334.
84. Aleman M, Brosnan RJ, Williams DC, et al. Malignanthyperthermia in a horse anesthetized with halothane. JVet Intern Med 2005;19:363–366.
85. Penedo MC, Millon LV, Bernoco D, et al. InternationalEquine Gene Mapping Workshop Report: a comprehensivelinkage map constructed with data from new markers andby merging four mapping resources. Cytogenet Genome Res2005;111:5–15.
86. Swinburne J, Gerstenberg C, Breen M, et al. First com-prehensive low-density horse linkage map based on two3-generation, full-sibling, cross-bred horse reference fami-lies. Genomics 2000;66:123–134.
87. Chowdhary BP, Raudsepp T, Kata SR, et al. The first-generation whole-genome radiation hybrid map in the horseidentifies conserved segments in human and mouse ge-nomes. Genome Res 2003;13:742–751.
88. Dranchak PK, Valberg SJ, Onan GW, et al. Exclusionof linkage of the RYR1, CACNA1S, and ATP2A1 genes torecurrent exertional rhabdomyolysis in Thorough-breds. Am J Vet Res 2006;67:1395–1400.
89. Wade CM, Giulotto E, Sigurdsson S, et al. Genome se-quence, comparative analysis, and population genetics of thedomestic horse. Science 2009;326:865–867.
90. Fritz KL, McCue ME, Valberg SJ, et al. Genetic mappingof recurrent exertional rhabdomyolysis in a population ofNorth American Thoroughbreds. Anim Genet March 23,2012 [Epub ahead of print].
91. Valberg S, Jonsson L, Lindholm A, et al. Muscle histopa-thology and plasma aspartate aminotransferase, creatinekinase and myoglobin changes with exercise in horses withrecurrent exertional rhabdomyolysis. Equine Vet J 1993;25:11–16.
92. Lindholm A, Johansson HE, Kjaersgaard P. Acute rhab-domyolysis (“tying-up”) in Standardbred horses: a morpho-logical and biochemical study. Acta Vet Scand 1974;15:325–339.
93. Beech J, Lindborg S, Fletcher JE, et al. Caffeine contrac-tures, twitch characteristics and the threshold for Ca(2)-induced Ca2 release in skeletal muscle from horses withchronic intermittent rhabdomyolysis. Res Vet Sci 1993;54:110–117.
120 2012 � Vol. 58 � AAEP PROCEEDINGS
FRANK J. MILNE STATE-OF-THE ART LECTURE
Orig. Op. OPERATOR: Session PROOF: PE’s: AA’s: 4/Color Figure(s) ARTNO:
1st disk, 2nd beb dainop 15 1,3-8,12,13,15-17 3353

94. MacLeay JM, Sorum SA, Valberg SJ, et al. Epidemiologicanalysis of factors influencing exertional rhabdomyolysis inThoroughbreds. Am J Vet Res 1999;60:1562–1566.
95. McGowan CM, Fordham T, Christley RM. Incidence andrisk factors for exertional rhabdomyolysis in Thoroughbredracehorses in the United Kingdom. Vet Rec 2002;151:623–626.
96. Valberg SJ, Cardinet GH III, Carlson GP, et al. Polysac-charide storage myopathy associated with recurrent exer-tional rhabdomyolysis in horses. Neuromuscul Disord1992;2:351–359.
97. McCue ME, Valberg SJ. Estimated prevalence of polysac-charide storage myopathy among overtly healthy QuarterHorses in the United States. Am J Vet Res 2007;231:746–750.
98. McCue ME, Ribeiro WP, Valberg SJ. Prevalence of poly-saccharide storage myopathy in horses with neuromusculardisorders. Equine Vet J Suppl 2006;36:340–344.
99. Firshman AM, Baird JD, Valberg SJ. Prevalences andclinical signs of polysaccharide storage myopathy and shiv-ers in Belgian draft horses. J Am Vet Med Assoc 2005;227:1958–1964.
100. Valentine BA, Credille KM, Lavoie JP, et al. Severe poly-saccharide storage myopathy in Belgian and Percherondraught horses. Equine Vet J 1997;29:220–225.
101. Valentine BA. Polysaccharide storage myopathy: a com-mon metabolic disorder of horses. Vet Pathol 2002;39:630.
102. Quiroz-Rothe E, Novales M, Guilera-Tejero E, et al. Poly-saccharide storage myopathy in the M longissimus lum-borum of showjumpers and dressage horses with back pain.Equine Vet J 2002;34:171–176.
103. Hunt LM, Valberg SJ, Steffenhagen K, et al. An epidemi-ological study of myopathies in Warmblood horses. EquineVet J 2008;40:171–177.
104. Johlig L, Valberg SJ, Mickelson JR, et al. Epidemiologicaland genetic study of exertional rhabdomyolysis in a Warm-blood horse family in Switzerland. Equine Vet J 2011;43:240–245.
105. Valberg SJ, Dyson S. Skeletal muscle and lameness. In:Ross M, Dyson S, editors. Lameness in the Horse. 1st edi-tion. Philadelphia: Saunders; 2003:723–743.
106. Cardinet GH, Littrell JF, Freedland RA. Comparative in-vestigations of serum creatine phosphokinase and glutamic-oxaloacetic transaminase activities in equine paralyticmyoglobinuria. Res Vet Sci 1967;8:219–226.
107. Perkins G, Valberg SJ, Madigan JM, et al. Electrolytedisturbances in foals with severe rhabdomyolysis. J VetIntern Med 1998;12:173–177.
108. Valberg S, Haggendal J, Lindholm A. Blood chemistry andskeletal muscle metabolic responses to exercise in horseswith recurrent exertional rhabdomyolysis. Equine Vet J1993;25:17–22.
109. Valberg SJ, Mickelson JR, Gallant EM, et al. Exertionalrhabdomyolysis in quarter horses and thoroughbreds: onesyndrome, multiple aetiologies. Equine Vet J Suppl 1999;30:533–538.
110. Stanley RLMC, Piercy RJ. Storage-associated artifacts inequine muscle biopsy samples. Equine Vet J 2009;41:82–86.
111. Cumming WJK, Fulthorpe JJ, Hudgson P, et al. Color Atlasof Muscle Pathology. 1st edition. London, UK: Mosby-Wolfe; 1994.
112. Cole FL, Mellor DJ, Hodgson DR, et al. Prevalence anddemographic characteristics of exertional rhabdomyolysis inhorses in Australia. Vet Rec 2004;155:625–630.
113. Steckel RR. The role of scintigraphy in the lameness eval-uation. Vet Clin North Am Equine Pract 1991;7:207–239.
114. McGowan CM, Posner RE, Christley RM. Incidence of ex-ertional rhabdomyolysis in polo horses in the USA and theUnited Kingdom in the 1999/2000 season. Vet Rec 2002;150:535–537.
115. Stauber WT, Smith CA. Cellular responses in exertion-induced skeletal muscle injury. Mol Cell Biochem 1998;179:189–196.
116. Grobler LA, Collins M, Lambert MI, et al. Skeletal musclepathology in endurance athletes with acquired training in-tolerance. Br J Sports Med 2004;38:697–703.
117. Valentine BA, Van Saun RJ, Thompson KN, et al. Role ofdietary carbohydrate and fat in horses with equine polysac-charide storage myopathy. J Am Vet Med Assoc 2001;219:1537–1544.
118. Valentine BA, Hintz HF, Freels KM, et al. Dietary controlof exertional rhabdomyolysis in horses. J Am Vet MedAssoc 1998;212:1588–1593.
119. Morris DD, Divers TJ, Whitlock RH. Renal clearance andfractional excretion of electrolytes over a 24-hour period inhorses. Am J Vet Res 1984;45:2431–2435.
120. Valberg SJ. Diseases of muscle. In: Smith BP, editor.Large Animal Internal Medicine. 3rd edition. St. Louis:Mosby; 2002:1272–1273.
121. Beech J. Chronic exertional rhabdomyolysis. Vet ClinNorth Am Equine Pract 1997;13:145–168.
122. Lentz LR, Valberg SJ, Balog EM, et al. Abnormal regula-tion of muscle contraction in horses with recurrent exer-tional rhabdomyolysis. Am J Vet Res 1999;60:992–999.
123. Isgren CM, Upjohn MM, Fernandez-Fuente M, et al.Epidemiology of exertional rhabdomyolysis susceptibility inStandardbred horses reveals associated risk factors and un-derlying enhanced performance. PLoS One 2010;5:e11594.
124. Hildebrand SV, Arpin D, Cardinet G III. Contracturetest and histologic and histochemical analyses of musclebiopsy specimens from horses with exertional rhabdomyol-ysis. J Am Vet Med Assoc 1990;196:1077–1083.
125. MacLeay JM, Valberg SJ, Pagan JD, et al. Effect of rationand exercise on plasma creatine kinase activity and lactateconcentration in Thoroughbred horses with recurrent exer-tional rhabdomyolysis. Am J Vet Res 2000;61:1390–1395.
126. McKenzie EC, Valberg SJ, Godden SM, et al. Effect ofdietary starch, fat, and bicarbonate content on exercise re-sponses and serum creatine kinase activity in equine recur-rent exertional rhabdomyolysis. J Vet Intern Med 2003;17:693–701.
127. Kohn CW, Hinchcliff KW. Physiological responses to theendurance test of a 3-day-event during hot and cool weather.Equine Vet J Suppl 1995;20:31–36.
128. Upjohn MM, Archer RM, Christley RM, et al. Incidenceand risk factors associated with exertional rhabdomyolysissyndrome in National Hunt racehorses in Great Brit-ain. Vet Rec 2005;156:763–766.
129. Evans DL, Priddle TL, Davie AJ. Plasma lactate and uricacid responses to racing in pacing Standardbreds and rela-tionships with performance. Equine Vet J Suppl 2002;34:131–134.
130. Lentz LR, Valberg SJ, Herold LV, et al. Myoplasmic cal-cium regulation in myotubes from horses with recurrentexertional rhabdomyolysis. Am J Vet Res 2002;63:1724–1731.
131. Lopez JR, Linares N, Cordovez G, et al. Elevated myoplas-mic calcium in exercise-induced equine rhabdomyolysis.Pflugers Arch 1995;430:293–295.
132. Az-Sylvester PL, Porta M, Copello JA. Halothane modula-tion of skeletal muscle ryanodine receptors: dependence onCa2, Mg2, and ATP. Am J Physiol Cell Physiol 2008;294:C1103–C1112.
133. Copello JA, Barg S, Sonnleitner A, et al. Differential acti-vation by Ca2, ATP and caffeine of cardiac and skeletalmuscle ryanodine receptors after block by Mg2. J MembrBiol 2002;187:51–64.
134. Waldron-Mease E, Klein LV, Rosenberg H, et al. Malignanthyperthermia in a halothane-anesthetized horse. J Am VetMed Assoc 1981;179:896–89.
135. Lentz LR, Valberg SJ, Mickelson JR, et al. In vitro con-tractile responses and contracture testing of skeletal musclefrom Quarter Horses with exertional rhabdomyoly-sis. Am J Vet Res 1999;60:684–688.
136. Mlekoday JA, Mickelson JR, Valberg SJ, et al. Calciumsensitivity of force production and myofibrillar ATPase ac-
AAEP PROCEEDINGS � Vol. 58 � 2012 121
FRANK J. MILNE STATE-OF-THE ART LECTURE
AQ: A
Orig. Op. OPERATOR: Session PROOF: PE’s: AA’s: 4/Color Figure(s) ARTNO:
1st disk, 2nd beb dainop 15 1,3-8,12,13,15-17 3353

tivity in muscles from Thoroughbreds with recurrent exer-tional rhabdomyolysis. Am J Vet Res 2001;62:1647–1652.
137. Ward TL, Valberg SJ, Gallant EM, et al. Calcium regula-tion by skeletal muscle membranes of horses with recurrentexertional rhabdomyolysis. Am J Vet Res 2000;61:242–247.
138. Fraunfelder HC, Rossdale PD, Rickets SW. Changes inserum muscle enzyme levels in associated with trainingschedules and stages of oestrus cycle in Thoroughbred race-horses. Equine Vet J 1986;18:371–374.
139. Tozaki T, Hirota K, Sugita S, et al. A genome-wide scan fortying-up syndrome in Japanese Thoroughbreds. AnimGenet 2010;41(Suppl 2):80–86.
140. Harris PA, Snow DH, Greet TR, et al. Some factors influ-encing plasma AST/CK activities in Thoroughbred race-horses. Equine Vet J Suppl 1990;9:66–71.
141. McKenzie EC, Firshman AM. Optimal diet of horses withchronic exertional myopathies. Vet Clin North Am EquinePract 2009;25:121–135.
142. MacLeay JM, Valberg SJ, Pagan JD, et al. Effect of diet onThoroughbred horses with recurrent exertional rhabdomy-olysis performing a standardised exercise test. Equine VetJ Suppl 1999;30:458–462.
143. Finno CJ, McKenzie E, Valberg SJ, et al. Effect of fitnesson glucose, insulin and cortisol responses to diets varying instarch and fat content in Thoroughbred horses with recur-rent exertional rhabdomyolysis. Equine Vet J 2010;42(Suppl 38):323–328.
144. Freestone JF, Wolfsheimer KJ, Kamerling SG, et al. Exer-cise induced hormonal and metabolic changes in Thorough-bred horses: effects of conditioning and acepromazine. EquineVet J 1991;23:219–223.
145. Wang R, Zhong X, Meng X, et al. Localization of the dan-trolene-binding sequence near the FK506-binding protein-binding site in the three-dimensional structure of theryanodine receptor. J Biol Chem 2011;286:12202–12212.
146. Edwards JG, Newtont JR, Ramzan PH, et al. The efficacyof dantrolene sodium in controlling exertional rhabdomyol-ysis in the Thoroughbred racehorse. Equine Vet J 2003;35:707–711.
147. McKenzie EC, Valberg SJ, Godden SM, et al. Effect of oraladministration of dantrolene sodium on serum creatine ki-nase activity after exercise in horses with recurrent exer-tional rhabdomyolysis. Am J Vet Res 2004;65:74–79.
148. McKenzie EC, Garrett RL, Payton ME, et al. Effect of feedrestriction on plasma dantrolene concentrations in horses.Equine Vet J 2010;42(Suppl 38):613–617.
149. Beech J, Fletcher JE, Lizzo F, et al. Effect of phenytoin onthe clinical signs and in vitro muscle twitch characteristicsin horses with chronic intermittent rhabdomyolysis andmyotonia. Am J Vet Res 1988;49:2130–2133.
150. Beech J. Treating and preventing chronic intermittentrhabdomyolysis. Vet Med 1994;89(5):458–461.
151. Tiidus PM. Manual massage and recovery of muscle func-tion following exercise: a literature review. J OrthopSports Phys Ther 1997;25:107–112.
152. Weerapong P, Hume PA, Kolt GS. The mechanisms ofmassage and effects on performance, muscle recovery andinjury prevention. Sports Med 2005;35:235–256.
153. Zainuddin Z, Newton M, Sacco P, et al. Effects of massageon delayed-onset muscle soreness, swelling, and recovery ofmuscle function. J Athl Train 2005;40:174–180.
154. Aleman M, Riehl J, Aldridge BM, et al. Association ofa mutation in the ryanodine receptor 1 gene with equinemalignant hyperthermia. Muscle Nerve 2004;30:356–365.
155. Mickelson JR, Louis CF. Malignant hyperthermia: exci-tation-contraction coupling, Ca2 release channel, and cellCa2 regulation defects. Physiol Rev 1996;76:537–592.
156. Arighi M, Hulland TJ. Equine exertional rhabdomyolysis.Compend Cont Educ Pract Vet 1984;6:S726–S732.
157. Andrews FM, Spurgeon TL, Reed SM. Histochemicalchanges in skeletal muscles of four male horses with neuro-muscular disease. Am J Vet Res 1986;47:2078–2083.
158. Valentine BA, Cooper BJ. Incidence of polysaccharide stor-age myopathy: necropsy study of 225 horses. Vet Pathol2005;42:823–827.
159. Valentine BA, Habecker PL, Patterson JS, et al. Incidenceof polysaccharide storage myopathy in draft horse-relatedbreeds: a necropsy study of 37 horses and a mule. J VetDiagn Invest 2001;13:63–68.
160. Valentine BA, McDonough SP, Chang YF, et al. Polysac-charide storage myopathy in Morgan, Arabian, and Stan-dardbred related horses and Welsh-cross ponies. VetPathol 2000;37:193–196.
161. Firshman AM, Valberg SJ, Bender JB, et al. Epidemio-logic characteristics and management of polysaccharidestorage myopathy in Quarter Horses. Am J Vet Res 2003;64:1319–1327.
162. Stanley RL, McCue ME, Valberg SJ, et al. A glycogensynthase 1 mutation associated with equine polysaccharidestorage myopathy and exertional rhabdomyolysis occurs ina variety of UK breeds. Equine Vet J 2009;41:597–601.
163. McGowan CM, Menzies-Gow NJ, McDiarmid AM, et al.Four cases of equine polysaccharide storage myopathy inthe United Kingdom. Vet Rec 2003;152:109–112.
164. Herszberg B, McCue ME, Larcher T, et al. A GYS1 genemutation is highly associated with polysaccharide storagemyopathy in Cob Normand draught horses. Anim Genet2009;40(1):94–96.
165. Valberg SJ, MacLeay JM, Billstrom JA, et al. Skeletalmuscle metabolic response to exercise in horses with ‘tying-up’ due to polysaccharide storage myopathy. Equine Vet J1999;31:43–47.
166. McCue ME, Valberg SJ, Lucio M, et al. Glycogen synthase1 (GYS1) mutation in diverse breeds with polysaccharidestorage myopathy. J Vet Intern Med 2008;22(5):1228–1233.
167. McCue ME, Armien AG, Lucio M, et al. Comparative skel-etal muscle histopathologic and ultrastructural featuresin two forms of polysaccharide storage myopathy in horses.Vet Pathol 2009;46:1281–1291.
168. Baird JD, Valberg SJ, Anderson SM, et al. Presence ofthe glycogen synthase 1 (GYS1) mutation causing type 1polysaccharide storage myopathy in continental Europeandraught horse breeds. Vet Rec 2010;167:781–784.
169. McCue ME, Anderson SM, Valberg SJ, et al. Estimatedprevalence of the type 1 polysaccharide storage myopathymutation in selected North American and European breeds.Anim Genet 2010;41(Suppl 2):145–149.
170. McGowan CM, McGowan TW, Patterson-Kane JC. Preva-lence of equine polysaccharide storage myopathy and othermyopathies in two equine populations in the United King-dom. Vet J 2009;180:330–336.
171. Tryon RC, Penedo MC, McCue ME, et al. Evaluation ofallele frequencies of inherited disease genes in subgroups ofAmerican Quarter Horses. J Am Vet Med Assoc 2009;234:120–125.
172. Ribeiro WP, Valberg SJ, Pagan JD, et al. The effect ofvarying dietary starch and fat content on serum creatinekinase activity and substrate availability in equine polysac-charide storage myopathy. J Vet Intern Med 2004;18:887–894.
173. McCue ME, Valberg SJ, Jackson M, et al. Polysaccharidestorage myopathy phenotype in Quarter Horse-related breedsis modified by the presence of an RYR1 mutation. NeuromuscDisord 2009;19(1):37–43.
174. Borgia LA, Valberg SJ, McCue ME, et al. Effect of dietaryfats with odd or even numbers of carbon atoms on metabolicresponse and muscle damage with exercise in QuarterHorse-type horses with type 1 polysaccharide storage myop-athy. Am J Vet Res 2010;71:326–336.
175. Byrne E, Jones SL, Valberg SJ, et al. Rhabdomyolysisin two foals with polysaccharide storage myopathy and con-current pneumonia. Comp Cont Educ Pract Vet 2000;22:503–507.
176. Valberg SJ, MacLeay JM, Mickelson JR. Polysaccharidestorage myopathy associated with exertional rhabdomyoly-
122 2012 � Vol. 58 � AAEP PROCEEDINGS
FRANK J. MILNE STATE-OF-THE ART LECTURE
Orig. Op. OPERATOR: Session PROOF: PE’s: AA’s: 4/Color Figure(s) ARTNO:
1st disk, 2nd beb dainop 15 1,3-8,12,13,15-17 3353

sis in horses. Comp Cont Educ Pract Vet 1997;19:1077–1086.
177. Firshman AM, Valberg SJ, Bender JB, et al. Comparisonof histopathologic criteria and skeletal muscle fixation tech-niques for the diagnosis of polysaccharide storage myopathyin horses. Vet Pathol 2006;43:257–269.
178. McCue ME, Valberg SJ, Lucio M, et al. A comparison ofskeletal muscle histopathologic features of two forms ofpolysaccharide storage myopathy in horses. Vet Pathol2009;46(4):1281–1291.
179. Sprayberry KA, Madigan J, Lecouteur RA, et al. Renalfailure, laminitis, and colitis following severe rhabdomyoly-sis in a draft horse-cross with polysaccharide storage myop-athy. Can Vet J 1998;39:500–50.
180. Bloom BA, Valentine BA, Gleed RD, et al. Postanaestheticrecumbency in a Belgian filly with polysaccharide storagemyopathy. Vet Rec 1999;144:73–75.
181. Essen-Gustavsson B, Gottlieb-Vedi M, Lindholm A. Mus-cle adenine nucleotide degradation during submaximaltreadmill exercise to fatigue. Equine Vet J Suppl 1999;30:298–302.
182. Annandale EJ, Valberg SJ, Essen-Gustavsson B. Effects ofsubmaximal exercise on adenine nucleotide concentrationsin skeletal muscle fibers of horses with polysaccharide stor-age myopathy. Am J Vet Res 2005;66:839–845.
183. Valberg SJ, Townsend D, Mickelson JR. Skeletal muscleglycolytic capacity and phosphofructokinase regulation inhorses with polysaccharide storage myopathy. Am J VetRes 1998;59:782–785.
184. De La Corte FD, Valberg SJ, Mickelson JR, et al. Bloodglucose clearance after feeding and exercise in polysaccha-ride storage myopathy. Equine Vet J Suppl 1999;30:324–328.
185. Annandale EJ, Valberg SJ, Mickelson JR, et al. Insulinsensitivity and skeletal muscle glucose transport in horseswith equine polysaccharide storage myopathy. Neuromus-cul Disord 2004;14:666–674.
186. Firshman AM, Valberg SJ, Baird JD, et al. Insulin sensi-tivity in Belgian horses with polysaccharide storage myop-athy. Am J Vet Res 2008;69:818–823.
187. Firshman AM, Valberg SJ, Karges TL, et al. Serum crea-tine kinase response to exercise during dexamethasone-induced insulin resistance in Quarter Horses withpolysaccharide storage myopathy. Am J Vet Res 2005;66:1718–1723.
188. Brojer JT, Essen-Gustavsson B, Annandale EJ, et al.Proglycogen, macroglycogen, glucose, and glucose-6-phosphateconcentrations in skeletal muscles of horses with polysac-charide storage myopathy performing light exercise. Am JVet Res 2006;67:1589–1594.
189. Brojer JT, Stampfli HR, Graham TE. Analysis of progly-cogen and macroglycogen content in muscle biopsy speci-mens obtained from horses. Am J Vet Res 2002;63:570–575.
190. Raben N, Danon M, Lu N, et al. Surprises of genetic en-gineering: a possible model of polyglucosan body disease.Neurology 2001;56:1739–1745.
191. Valentine BA, Cooper BJ. Development of polyglucosaninclusions in skeletal muscle. Neuromuscul Disord 2006;16:603–607.
192. Hardie DG. The AMP-activated protein kinase pathway:new players upstream and downstream. J Cell Sci 2004;117(Pt 23):5479–5487.
193. Hardie DG. AMP-activated protein kinase: a key systemmediating metabolic responses to exercise. Med Sci SportsExerc 2004;36:28–34.
194. Smith AC, Bruce CR, Dyck DJ. AMP kinase activationwith AICAR simultaneously increases fatty acid and glucoseoxidation in resting rat soleus muscle. J Physiol 2005;565(Pt 2):537–546.
195. Saha AK, Laybutt DR, Dean D, et al. Cytosolic citrateand malonyl-CoA regulation in rat muscle in vivo. Am JPhysiol 1999;276(6 Pt 1):E1030–E1037.
196. Ruderman NB, Saha AK, Vavvas D, et al. Malonyl-CoA,fuel sensing, and insulin resistance. Am J Physiol Endo-crinol Metab 1999;276:E1–E18.
197. McGarry JD, Brown NF. The mitochondrial carnitinepalmitoyltransferase system: from concept to molecularanalysis. Eur J Biochem 1997;244:1–14.
198. Aleman M, Lecouteur RA, Nieto JE, et al. Sarcoplasmicmasses in equine skeletal muscle. Neuromuscul Disord2005;15:147–153.
199. Borgia L, Valberg SJ, McCue ME, et al. The effect of di-etary odd and even carbon fat on metabolic response andmuscle damage with exercise in Quarter Horse-relatedbreeds with type 1 polysaccharide storage myopathy. Am JVet Res 2010;71(3):326–336.
200. Valberg SJ, McCue ME, Mickelson JR. The interplay ofgenetics, exercise and nutrition in polysaccharide storagemyopathy. J Equine Vet Sci 2011;31:205–210.
201. Valberg SJ, Carlson GP, Cardinet GH III, et al. Skeletalmuscle mitochondrial myopathy as a cause of exercise intol-erance in a horse. Muscle Nerve 1994;17:305–312.
202. Bedford H, Valberg SJ, Firshman AM, et al. Reversiblemuscle atrophy and weakness associated with sacrocaudalisdorsalis medialis myopathy and vitamin E (alpha toco-pherol) deficiency in horses. J Am Vet Med Assoc 2012(in press).
aRe-Leve, Kentucky Equine Research, Versailles, KY 40383.
AAEP PROCEEDINGS � Vol. 58 � 2012 123
FRANK J. MILNE STATE-OF-THE ART LECTURE
Orig. Op. OPERATOR: Session PROOF: PE’s: AA’s: 4/Color Figure(s) ARTNO:
1st disk, 2nd beb dainop 15 1,3-8,12,13,15-17 3353





























