Embed Size (px)
Citation preview

Natural insights for chemical biologistsChristopher T Walsh
Chemical biologists studying natural-product pathways encoded in genomes have unearthed new chemistry and insights into the evolution of biologically active metabolites.
Chemical biology has become a contempo-rary research and educational focus of many chemistry departments, as well as a revitalizing theme in biochemistry and biological chemis-try departments in academic medical centers. Although the latter departments have had as their classical purview the study of chemistry that occurs in living systems, the recent fram-ing of chemical biology has seen a greater influx of chemical thinking and chemists into biology. The goal is a seamless application of chemical principles to decipher complexi-ties in biology and bring scientists trained in chemistry to full engagement on biological projects.
Generally, molecular biology has been inar-guably powerful in the manipulation and cha-racterization of macromolecules in biological systems. However, it is the remit of chemical biology to illuminate the ‘other half ’: the small molecules (molecular weight < 2,500 Da) that comprise the metabolome and individually func-tion as specific ligands and signaling molecules. Chemical biology focuses on these small molecules: as probes of particular biological targets, as sources of new leads for pharma-cology, and as molecules used by prokaryotic and eukaryotic organisms for communication and signaling. More insight into the world of natural small molecules and their biological targets will enable new approaches to creat-ing synthetic homologs and analogs. In turn, these compounds will be valuable ligands for defining specificity and selectivity of biologi-cal macromolecules. Indeed, one of the prin-cipal activities of a large swath of the chemical
biology community involves chemical library construction and functional optimization of small-molecule synthetic surrogates in cell-based screening assays to provide molecular tools analogous to specific gene knockouts or RNA knockdowns. Doubtless, these mainline chemical biology efforts in the service of cell biology will be addressed in other commentar-ies in this series by more expert practitioners1.
The bio-logic of small-molecule assemblyI have been particularly impressed over the past decade with the inroads that chemical biology has made in the understanding of natural-product small molecules and the deci-phering of their biosynthetic logic. A remark-
able renaissance in natural products has been brought about by molecular biology, strepto-myces genetics and the sequencing of >300 microbial genomes, leading to the identifica-tion of hundreds of biosynthetic gene clusters for secondary (conditional) metabolite bio-synthesis. This has opened a fantastic window on the logic of small-molecule evolution and assembly principles in nature.
Starting with the sequence of the erythro-mycin gene cluster2,3, the rules for modular assembly of malonyl- and methylmalonyl-CoA monomers into thousands of structural vari-ants of polyketides have been delineated4. The parallel assembly-line logic of nonribosomal peptide assembly into penicillins, vancomycin
Christopher T. Walsh is in the Department of Biological Chemistry and Molecular Pharmacology at Harvard Medical School, Boston, Massachusetts 02115, USA.e-mail: [email protected]
COM M E N TA RY
122 VOLUME 1 NUMBER 3 AUGUST 2005 NATURE CHEMICAL BIOLOGY
Hexaketide
Diels-
a
HH
SO Enz
H
O
OHO
O
OSO Enz
RN
S
NS
S
O
EnzRHN
NH
S
O
Enz
OSH
O
NH3
O
O
NH3
O
S Enz
Cl
NH3
O
O
Lovastatin
Bithiazole
Bleomycin
Coronamic acidL-allo-Isoleucine
Alderase
b
c
SH
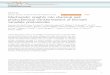
Scheme 1 Intriguing enzymatic chemistry during natural product biosynthesis. (a) The Diels-Alderase activity during lovastatin assembly. (b) The tandem formation of the DNA-binding bithiazole ring system of bleomycin. (c) The conversion of an unactivated methyl of allo-isoleucine to coronamic acid by successive enzymatic chlorination and cyclopropane-forming steps.
© 2
005
Nat
ure
Pub
lishi
ng G
roup
htt
p:/
/ww
w.n
atur
e.co
m/n
atur
eche
mic
alb
iolo
gy

and hundreds of other peptidic frameworks has also been revealed5. The way that nature mixes and matches assembly-line elements, and the underlying chemistry for making polyketide–nonribosomal peptide hybrids such as epothilones, rapamycin, FK506 and bleomycin have become open to inspection and then purposeful manipulation6,7.
Microbes that function as natural-product factories have to coordinate three stages of synthesis. The first is the timely provision of monomers for incorporation by the assem-bly lines. Although the building blocks for polyketides are a very limited set of malonyl-CoA derivatives, there are dozens of nonpro-teinogenic amino acid building blocks that have to be made with ‘just in time’ inventory control when natural-product biosynthetic gene clusters are transcribed. The order of incorporation of monomer units, comprising the second stage, is specified by the order and composition of domains in the multimodular assembly line. For example, the cyclosporine assembly line has 47 domains in a beads-on-a-string array in a single 1.5-MDa polypep-tide8. The third stage is post-assembly-line tailoring and, as noted below, can consist of a set of enzymatic steps by dedicated tailoring enzymes that create biological activity in the final product. The vancomycin and enediyne antitumor antibiotic biosynthetic pathways each have about 50 enzyme-mediated chemi-cal steps9,10.
Novel enzyme-mediated chemistrySorting out nature’s assembly-line logic for this vast cornucopia of bioactive natural pro- ducts has revealed new principles of modular enzymology11 and spawned several strategies for gene shuffling and module replacement to effect combinatorial biosynthesis of focused libraries of new congeners12. These studies have also revealed new chemistry (Scheme 1), from biological Diels-Alder cyclizations13, enzymatic conversion of peptide bonds to oxazoles and thiazoles14, and biological func-tionalization of unactivated carbon sites for chlorination and cyclopropanation15. Analysis of natural routes to complex molecules, such as paclitaxel, have revealed that nature makes the taxadiene hydrocarbon framework first and then stereospecifically and regiospecifi-cally hydroxylates eight carbon sites around the periphery16, pointing to as yet unrealized goals for synthetic chemists.
Natural tailoring of architectural scaffoldsThe interplay of enzymatic and chemical synthesis is also illuminated by the realiza-tions that thousands of natural products, released from polyketide synthases and non-ribosomal peptide synthetases, gain useful therapeutic activity only after the periphery is decorated by suites of dedicated tailoring enzymes. These tailoring enzymes perform a multitude of modifications, such as the oxida-tion of reduced carbon sites, as in paclitaxel
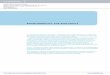
maturation from taxadiene, or oxidative cross-linking of the vancomycin heptapeptide scaffold. Glycosylation is also a favorite tailoring strategy, where select deoxyhexoses are added to natural-product scaffolds17. We have recently shown that enzymatic glycosylation or chemoenzymatic sugar decoration are promis-ing late-stage diversification strategies in three distinct scaffolds: vancomycin and teicoplanin antibiotics, anthracycline antitumor antibiot-ics and the aminocoumarin family of antibi-otics18–20 (Scheme 2). The deoxysugars in all three molecular classes are key pharmaco-phores involved in target recognition and thus are important elements for rational alteration to elicit new therapeutic activity21.
Although natural products have fallen out of current favor in the pharmaceutical industry (with infectious disease and cancer therapeu-tics as notable exceptions), there is no doubt that the architecture and functional-group density of natural scaffolds reflect incom-pletely exploited starting points for biologi-cally relevant structures22.
An incomplete molecular inventoryOne aspect of note is the observation that much, perhaps most, of the secondary metabolite synthetic capacity of producer microorganisms is cryptic under normal cul-ture conditions. For example, Streptomyces avermitilis, the producer of the widely used drug avermectin, has 25 predicted biosyn-thetic clusters in its genome23 At most three of the 25 clusters make detectable products in laboratory culture conditions yet reported24; the identity of the other predicted secondary metabolites is unknown. Many could be sig-naling molecules that require particular con-ditions to elicit expression of the responsible gene clusters. This ten-fold underrepresenta-tion of the biosynthetic capacity of the few hundred sequenced microbes is dwarfed by the estimation that 99 to 99.9% of microbes are not yet culturable25. Thus, there could be between 100- and 1000-fold more biological and chemical diversity to microbial small molecules lying ahead of investigators attempt-ing metagenome characterizations, environ-mental DNA isolation and manipulation of new functional small molecules26.
Nearest neighbors for chemical biologyAlthough genomics and molecular biology have enabled chemical biologists to move with some confidence into deeper investiga-tions of biology’s logic, advances in physical methods are the other drivers for innovation in this field. Perhaps the single most important technology has been the biological application of mass spectrometry. The power of modern
COM M E N TA RY
NATURE CHEMICAL BIOLOGY VOLUME 1 NUMBER 3 AUGUST 2005 123
NH
HN
NH
HN
O
O
O
NH
Acyclic peptide precursor Chloroeremomycin
NH2
OO
HO
O
OHOH HO
OH
OO
O
O
OOHOH
OHO
Cl
Cl
O O
OHNH2
HOO O
OHHN
O
OO
OO
NH
OH
Cl
R
O
Glycosylation
4'-O-Methylation
3'-O-Acylation
Acylation
Chlorination
Clorobiocin
ChlorinationChlorination
Glycosylation
Glycosylation
H3C
a
O
H3NHO
O
H3NHOOxidative
cyclization
Oxidativecyclization
b
Scheme 2 Post-assembly line enzymatic tailoring of natural product scaffolds. (a) Chlorinations, oxidative cyclizations and glycosylations during maturation of the vancomycin family glycopeptide antibiotic chloroeremomycin. (b) Chlorination, O-glycosylation, O-methylation and O, N-acylations during enzymatic conversion of the coumarin core to the antibiotic clorobiocin.
© 2
005
Nat
ure
Pub
lishi
ng G
roup
htt
p:/
/ww
w.n
atur
e.co
m/n
atur
eche
mic
alb
iolo
gy

analytical chemistry can be exemplified in various ways, but DNA sequencers are joined by mass spectrometers in revolutionizing the acquisition of data, the scope of inquiry and the need for accurate and sensitive analysis. This is most clear in the dominance of mass spectrometry for characterization of the stoi-chiometry and location of post-translational modification reactions of proteins, resulting in millions of molecular variants of proteins in a typical eukaryotic cell27. Continued robust development of chemical biology will require close ties to bioengineering and advances in analytic chemistry.
Intellectually, chemical biologists should also be closely paired with systems biologists for maximal understanding of collections of small molecules mapped onto high-density matrices of biological data points for pattern recognitions. For example, such a pairing would be ideal for working out the compo-sition of phosphoprotein proteomes and ubi-quitylated proteomes in different cells under varying physiologic challenges. One particularly exciting opportunity for chemi-cal biologists will be interfacing with devel-opmental biologists to further understand the
decision trees affected by both large and small molecules during organ and tissue develop-ment. This will include directed differentiation of immature cells down particular lineages under the guidance of natural or synthetic small molecules.
For chemical biology to realize its most satisfying future, its practitioners will have to speak the many dialects of chemistry but be firmly embedded in biological cultures and microenvironments.
ACKNOWLEDGMENTSI thank M. Fischbach for the artwork in this Commentary.
1. Schreiber, S.L. Nat. Chem. Biol. 1, 65–66 (2005).2. Donadio, S., Staver, M.J., McAlpine, J.B., Swanson,
S.J. & Katz, L. Science 252, 675–679 (1991).3. Cortes, J., Haydock, S., Roberts, G.A., Bevitt, D.J. &
Leadlay, P.F. Nature 348, 176–178 (1990).4. Katz, L. & Ashley, G. Chem. Rev. 105, 499–528
(2005).5. Schwarzer, D. Firking, R., Marahiel, M.A. Nat. Prod.
Rep. 20, 275–287 (2003).6. Tang, L. et al. J. Antibiot. (Tokyo) 58, 178–184
(2005).7. Lowden, P.A., Bohm, G.A., Metcalfe, S., Staunton, J.
& Leadlay, P.F. ChemBioChem 5, 535–538 (2004).8. Weber, G., Shorgendorfer, K., Schneider-Scherzer, E.
& Leitner, E. Curr. Genet. 26, 120–125 (1994).
9. Hubbard, B.K. & Walsh, C.T. Angew. Chem. Int. Edn Engl. 42, 730–765 (2003).
10. Liu, W., Christensen, S., Standage, S. & Shen, B. Science 297, 1170–1173 (2002).
11. Walsh, C. Science 303, 1805–1810 (2004).12. McDaniel, R. et al. Proc. Natl. Acad. Sci. USA 96,
1846–1851 (1999).13. Auclair, K. et al. J. Am. Chem. Soc. 122, 11519–11520
(2000).14. O’Connor, S., Chen, H. & Walsh, C. Biochemistry 41,
5685–5694 (2002).15. Vaillancourt, F., Yeh, E., Vosburg, D., O’Connor, S. &
Walsh, C. Nature (in the press) (2005).16. Clardy, J. & Walsh, C. Nature 432, 829–837 (2004).17. Liu, H.W. & Thorson, J.S. Annu. Rev. Microbiol. 48,
223–256 (1994).18. Losey, H.C. et al. Walsh, C. Biochemistry 40, 4745–
4755 (2001).19. Freel Meyers, C., Oberthur, M., Heide, L., Kahne, D. &
Walsh, C. Biochemistry 43, 15022–15036 (2004).20. Lu, W. et al. Chem. Biol. 12, 527–534 (2005).21. Walsh, C., Freel Meyers, C. & Losey, H.C. J. Med. Chem.
46, 3425–3436 (2003).22. Koehn, F.E. & Carter, G.T. Nat. Rev. Drug Discov. 4,
206–220 (2005).23. Omura, S. et al. Proc. Natl. Acad. Sci. USA 98, 12215–
12220 (2001).24. Zazopoulos, E. et al. Nat. Biotechnol. 21, 187–190
(2003).25. Handelsman, J. Microbiol. Mol. Biol. Rev. 68, 669–685
(2004).26. Brady, S., Chao, C.J. & Clardy, J. J. Am. Chem. Soc.
124, 9968–9969 (2002).27. Walsh, C. Posttranslational Modification of Proteins:
Expanding Nature’s Inventory (Roberts & Co Press, Englewood, Colorado, USA, 2005).
COM M E N TA RY
124 VOLUME 1 NUMBER 3 AUGUST 2005 NATURE CHEMICAL BIOLOGY
© 2
005
Nat
ure
Pub
lishi
ng G
roup
htt
p:/
/ww
w.n
atur
e.co
m/n
atur
eche
mic
alb
iolo
gy