Embed Size (px)
Citation preview

Supplemental Methods for
Neisseria meningitidis recruits factor H using protein
mimicry of host carbohydrates
Muriel C. Schneider1+*
, Beverly E. Prosser2*
, Joseph J.E. Caesar2, Elisabeth
Kugelberg1, Su Li
1, Qian Zhang
1, Sadik Quoraishi
2, Janet E. Lovett
2, Janet E. Deane
2,
Robert B. Sim3, Pietro Roversi
2, Steven Johnson
2, Christoph M. Tang
1 & Susan M.
Lea2
1Centre for Molecular Microbiology and Infection, Imperial College, London SW7
2AZ, U.K. 2Sir William Dunn School of Pathology, University of Oxford, South Parks
Road, Oxford OX1 3RE, U.K. 3MRC Immunochemistry Unit, Department of
Biochemistry, University of Oxford, South Parks Road, Oxford OX1 3QU, U.K.
*These authors contributed equally to this work
Supplemental Analysis
Structure comparisons: As noted in the main text, when this structure of fHbp is compared
to the earlier structure of the “BC” region of the same protein1 there are significant
differences, particularly in the “B” region (Fig. S2). We interpret these differences as being
genuine differences between the structures due to inappropriate truncation (part way through
the N-terminal β-barrel) in the construct used for the NMR structure, rather than reflecting
differences arising from the different methods of structure determination. By contrast, when
the fH67 structure is compared to the equivalent portion from our earlier fH678/SOS complex2
(Fig. S6), no significant structural differences are seen, presumably reflecting the fact that
truncation of CCP8 does not have adverse effects on the structures of CCPs 6 and 7. No
structural homologue of the novel structure seen in the N-terminal β-barrel of fHbp is found
searching using either DALI3 or SSM 4. If more distant similarities are searched for (SSM
lower limit of acceptable secondary structure matching 40%) using the entire fHbp structure
(and ignoring topological considerations), an intriguing similarity in the arrangement of the two
β-barrels compared to the earlier structure of an engineered avidin5 (PDBID: 2C4I, Fig S10) is
observed. Given this protein’s role in complement related biology, we also note that the serine
protease domain of complement regulator Factor B6 (PDBID: 2OK5) is, in part, a two β-barrel
structure, and the relative arrangement of the two barrels is rather similar in both fHbp and
SUPPLEMENTARY INFORMATION
doi: 10.1038/nature07769
www.nature.com/nature 1

Factor B (Fig. S11). Any biological relevance of these structural similarities is, however,
unclear.
doi: 10.1038/nature07769 SUPPLEMENTARY INFORMATION
www.nature.com/nature 2
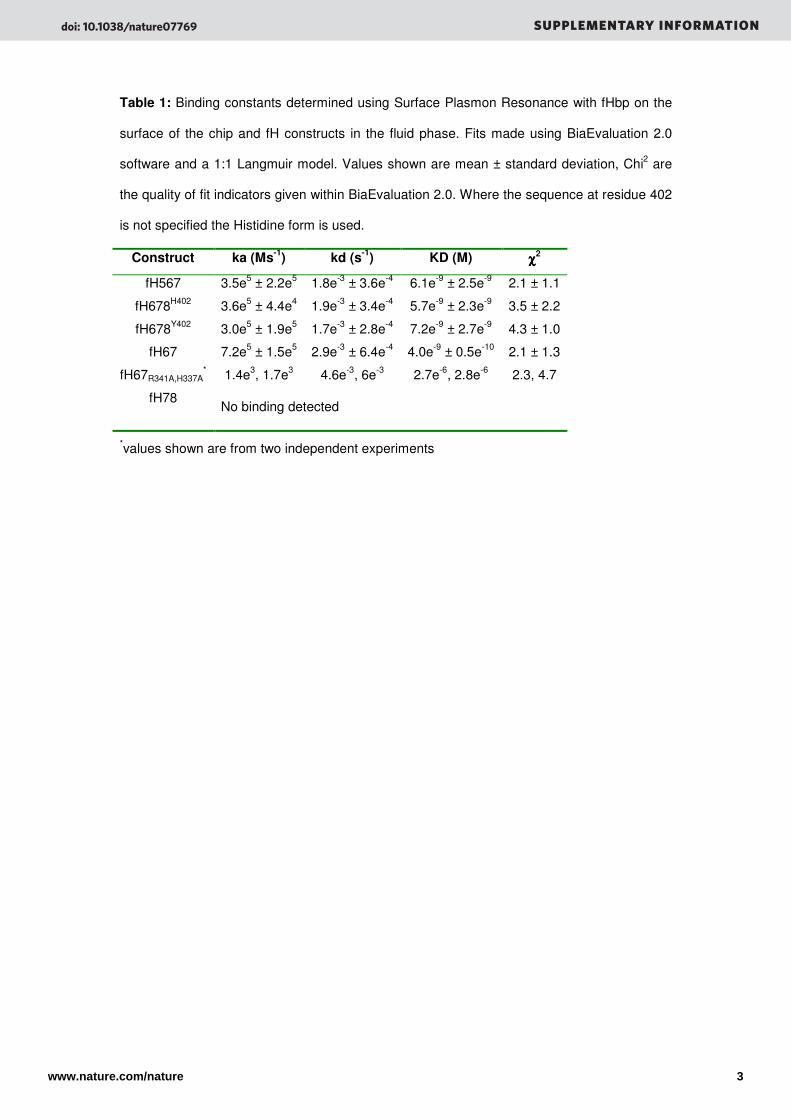
Table 1: Binding constants determined using Surface Plasmon Resonance with fHbp on the
surface of the chip and fH constructs in the fluid phase. Fits made using BiaEvaluation 2.0
software and a 1:1 Langmuir model. Values shown are mean ± standard deviation, Chi2 are
the quality of fit indicators given within BiaEvaluation 2.0. Where the sequence at residue 402
is not specified the Histidine form is used.
Construct ka (Ms-1
) kd (s-1
) KD (M) χχχχ2
fH567 3.5e5 ± 2.2e5 1.8e-3 ± 3.6e-4 6.1e-9 ± 2.5e-9 2.1 ± 1.1
fH678H402 3.6e5 ± 4.4e4 1.9e-3 ± 3.4e-4 5.7e-9 ± 2.3e-9 3.5 ± 2.2
fH678Y402 3.0e5 ± 1.9e5 1.7e-3 ± 2.8e-4 7.2e-9 ± 2.7e-9 4.3 ± 1.0
fH67 7.2e5 ± 1.5e5 2.9e-3 ± 6.4e-4 4.0e-9 ± 0.5e-10 2.1 ± 1.3
fH67R341A,H337A* 1.4e3, 1.7e3 4.6e-3, 6e-3 2.7e-6, 2.8e-6 2.3, 4.7
fH78 No binding detected
*values shown are from two independent experiments
doi: 10.1038/nature07769 SUPPLEMENTARY INFORMATION
www.nature.com/nature 3
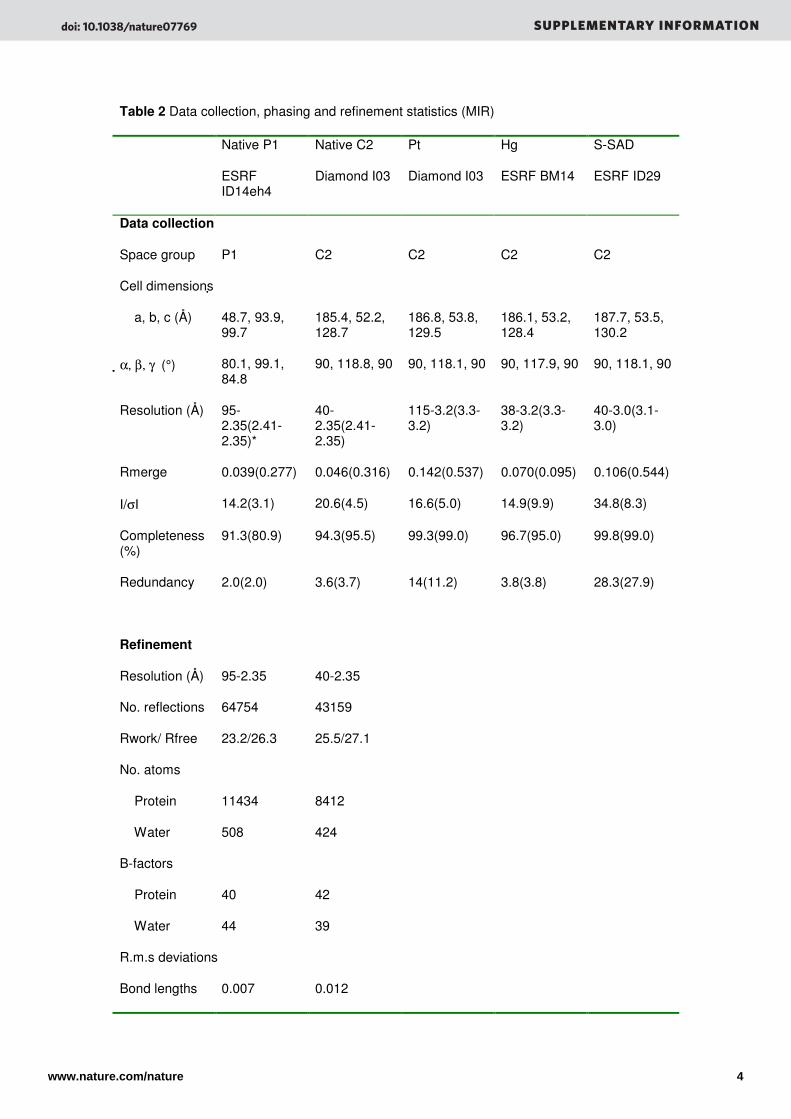
Table 2 Data collection, phasing and refinement statistics (MIR)
Native P1
ESRF ID14eh4
Native C2
Diamond I03
Pt
Diamond I03
Hg
ESRF BM14
S-SAD
ESRF ID29
Data collection
Space group P1 C2 C2 C2 C2
Cell dimensions
a, b, c (Å) 48.7, 93.9, 99.7
185.4, 52.2, 128.7
186.8, 53.8, 129.5
186.1, 53.2, 128.4
187.7, 53.5, 130.2
α, β, γ (°) 80.1, 99.1, 84.8
90, 118.8, 90 90, 118.1, 90 90, 117.9, 90 90, 118.1, 90
Resolution (Å) 95-2.35(2.41-2.35)*
40-2.35(2.41-2.35)
115-3.2(3.3-3.2)
38-3.2(3.3-3.2)
40-3.0(3.1-3.0)
Rmerge 0.039(0.277) 0.046(0.316) 0.142(0.537) 0.070(0.095) 0.106(0.544)
I/σI 14.2(3.1) 20.6(4.5) 16.6(5.0) 14.9(9.9) 34.8(8.3)
Completeness (%)
91.3(80.9) 94.3(95.5) 99.3(99.0) 96.7(95.0) 99.8(99.0)
Redundancy 2.0(2.0) 3.6(3.7) 14(11.2) 3.8(3.8) 28.3(27.9)
Refinement
Resolution (Å) 95-2.35 40-2.35
No. reflections 64754 43159
Rwork/ Rfree 23.2/26.3 25.5/27.1
No. atoms
Protein 11434 8412
Water 508 424
B-factors
Protein 40 42
Water 44 39
R.m.s deviations
Bond lengths 0.007 0.012
doi: 10.1038/nature07769 SUPPLEMENTARY INFORMATION
www.nature.com/nature 4

(Å)
Bond angles (°)
0.99 0.94
*Highest resolution shell is shown in parenthesis. Data for each dataset were collected from a
single crystal
doi: 10.1038/nature07769 SUPPLEMENTARY INFORMATION
www.nature.com/nature 5
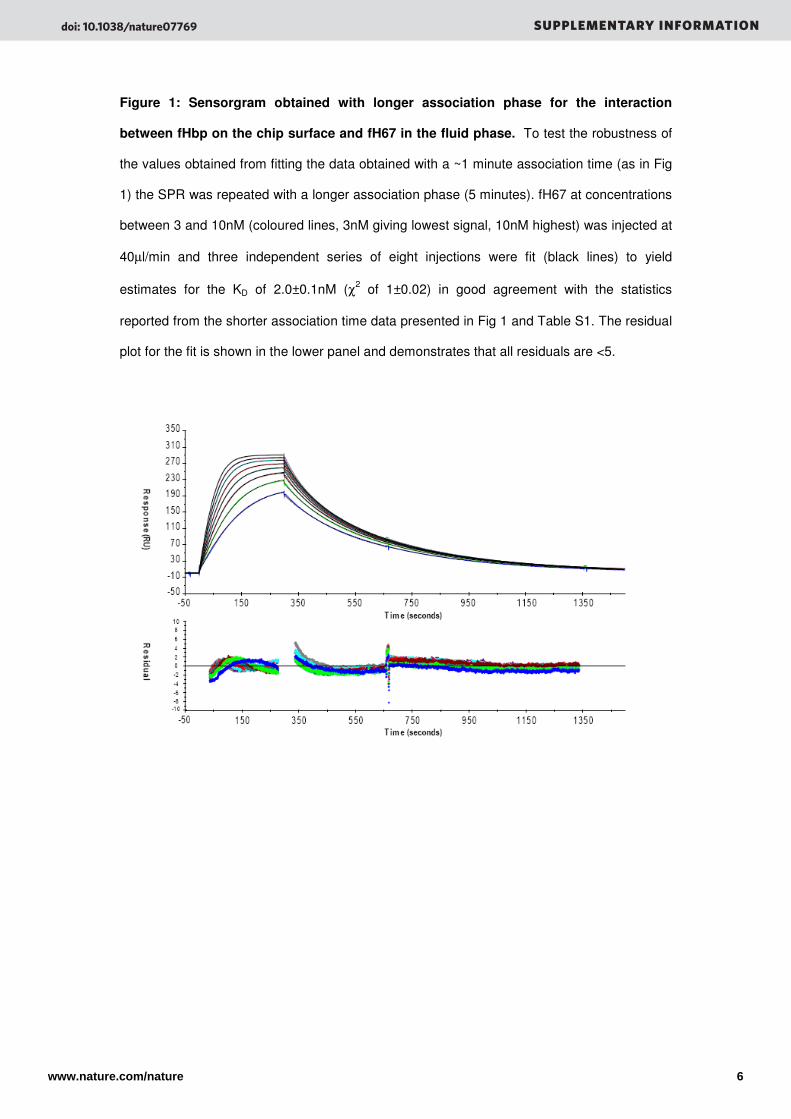
Figure 1: Sensorgram obtained with longer association phase for the interaction
between fHbp on the chip surface and fH67 in the fluid phase. To test the robustness of
the values obtained from fitting the data obtained with a ~1 minute association time (as in Fig
1) the SPR was repeated with a longer association phase (5 minutes). fH67 at concentrations
between 3 and 10nM (coloured lines, 3nM giving lowest signal, 10nM highest) was injected at
40µl/min and three independent series of eight injections were fit (black lines) to yield
estimates for the KD of 2.0±0.1nM (χ2 of 1±0.02) in good agreement with the statistics
reported from the shorter association time data presented in Fig 1 and Table S1. The residual
plot for the fit is shown in the lower panel and demonstrates that all residuals are <5.
doi: 10.1038/nature07769 SUPPLEMENTARY INFORMATION
www.nature.com/nature 6
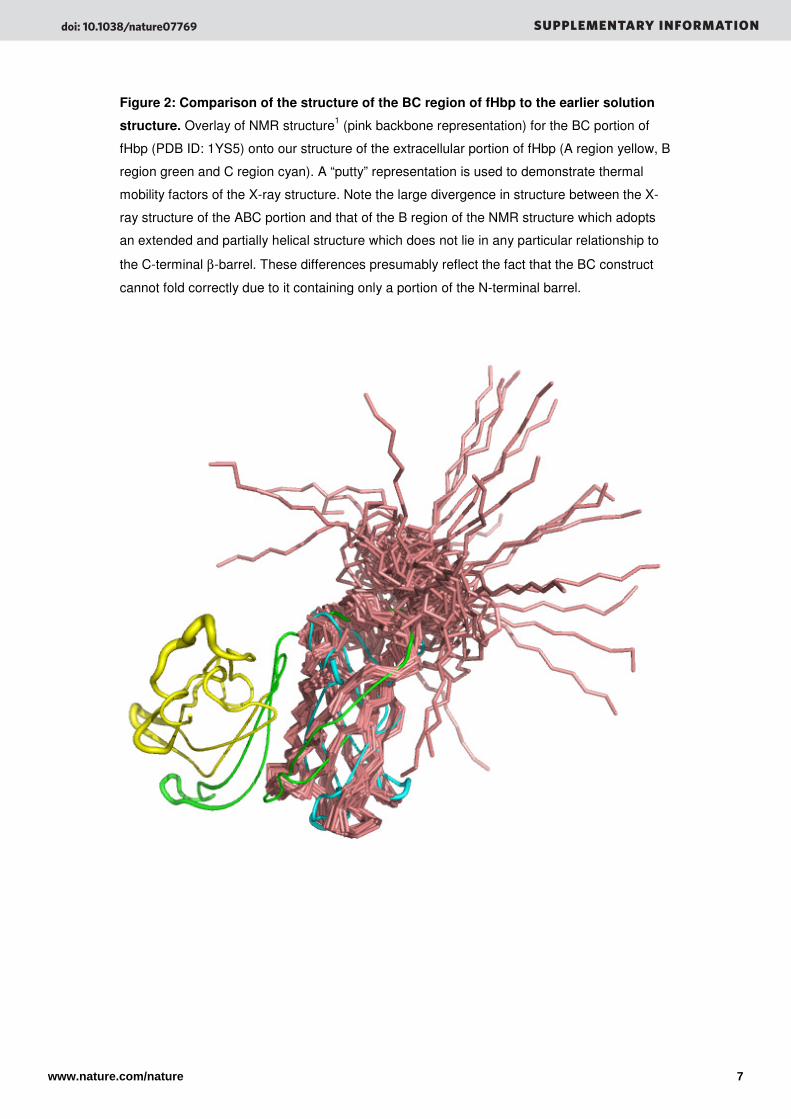
Figure 2: Comparison of the structure of the BC region of fHbp to the earlier solution
structure. Overlay of NMR structure1 (pink backbone representation) for the BC portion of
fHbp (PDB ID: 1YS5) onto our structure of the extracellular portion of fHbp (A region yellow, B
region green and C region cyan). A “putty” representation is used to demonstrate thermal
mobility factors of the X-ray structure. Note the large divergence in structure between the X-
ray structure of the ABC portion and that of the B region of the NMR structure which adopts
an extended and partially helical structure which does not lie in any particular relationship to
the C-terminal β-barrel. These differences presumably reflect the fact that the BC construct
cannot fold correctly due to it containing only a portion of the N-terminal barrel.
doi: 10.1038/nature07769 SUPPLEMENTARY INFORMATION
www.nature.com/nature 7
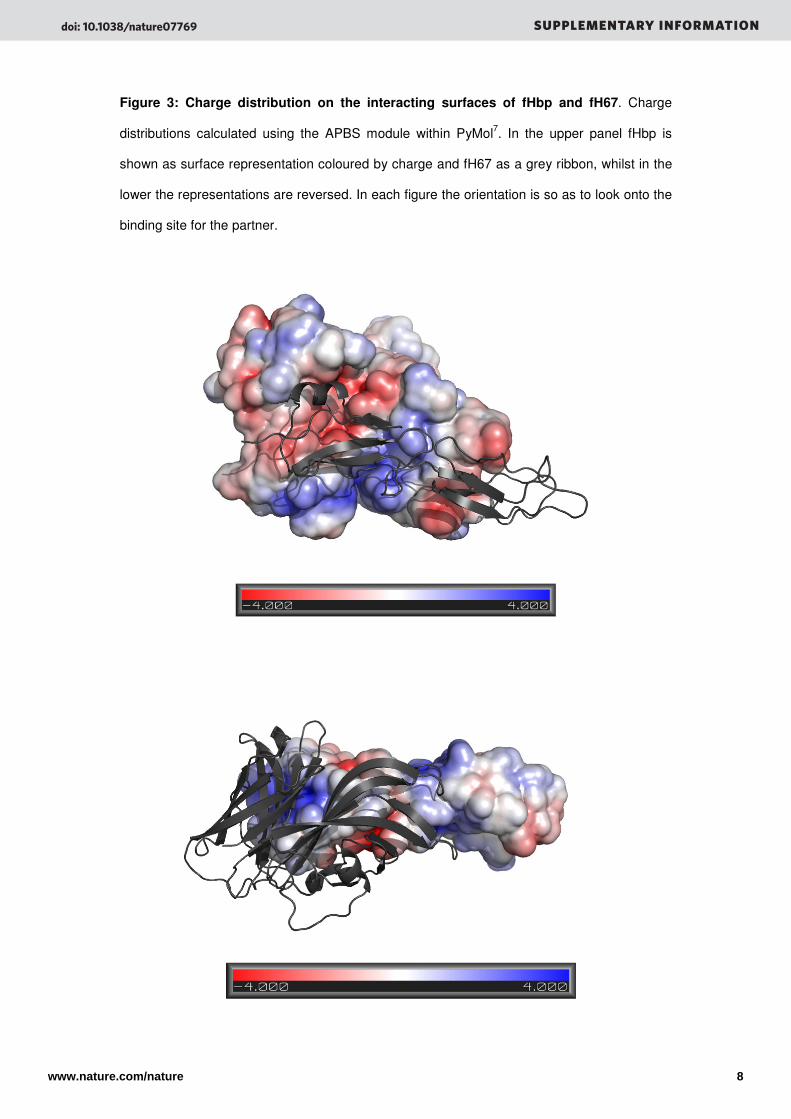
Figure 3: Charge distribution on the interacting surfaces of fHbp and fH67. Charge
distributions calculated using the APBS module within PyMol7. In the upper panel fHbp is
shown as surface representation coloured by charge and fH67 as a grey ribbon, whilst in the
lower the representations are reversed. In each figure the orientation is so as to look onto the
binding site for the partner.
doi: 10.1038/nature07769 SUPPLEMENTARY INFORMATION
www.nature.com/nature 8

Figure 4: Flexibility in the fH67/fHbp complex. Overlay of the seven, crystallographically
independent copies of the fH67/fHbp complex from the two crystal forms (four copies of the
complex in the P1 form and 3 copies in the C2 form). The high degree of similarity despite
very different packing environments indicates that the complex is biologically relevant.
Rearrangement of the proteins within the complex is limited to a small amount of movement
in the position of CCP7 of fH67 relative to CCP6 and fHbp. The loops of fHbp which pack
closest to CCP7 are also relatively mobile as indicated by their higher temperature factors
(displayed here as a “thicker” putty representation).
fH CCP6
fH CCP7
fHbp
doi: 10.1038/nature07769 SUPPLEMENTARY INFORMATION
www.nature.com/nature 9
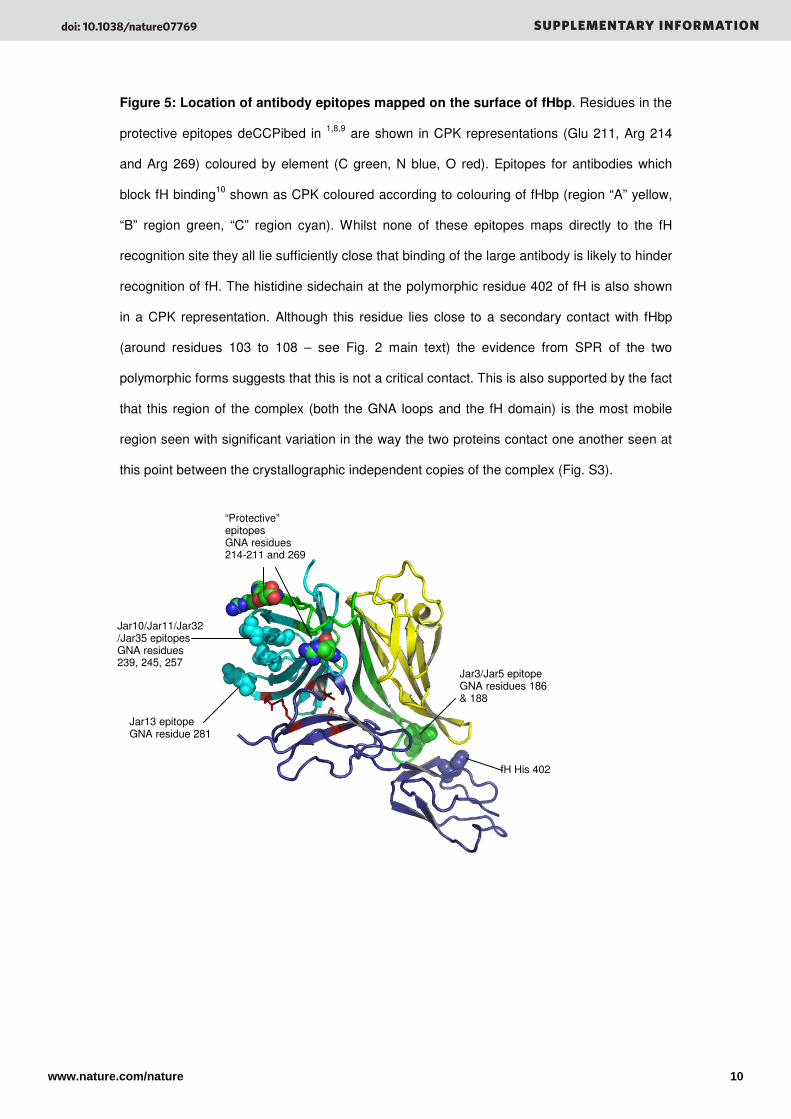
Figure 5: Location of antibody epitopes mapped on the surface of fHbp. Residues in the
protective epitopes deCCPibed in 1,8,9 are shown in CPK representations (Glu 211, Arg 214
and Arg 269) coloured by element (C green, N blue, O red). Epitopes for antibodies which
block fH binding10 shown as CPK coloured according to colouring of fHbp (region “A” yellow,
“B” region green, “C” region cyan). Whilst none of these epitopes maps directly to the fH
recognition site they all lie sufficiently close that binding of the large antibody is likely to hinder
recognition of fH. The histidine sidechain at the polymorphic residue 402 of fH is also shown
in a CPK representation. Although this residue lies close to a secondary contact with fHbp
(around residues 103 to 108 – see Fig. 2 main text) the evidence from SPR of the two
polymorphic forms suggests that this is not a critical contact. This is also supported by the fact
that this region of the complex (both the GNA loops and the fH domain) is the most mobile
region seen with significant variation in the way the two proteins contact one another seen at
this point between the crystallographic independent copies of the complex (Fig. S3).
fH His 402
Jar3/Jar5 epitope GNA residues 186 & 188
Jar10/Jar11/Jar32/Jar35 epitopes GNA residues 239, 245, 257
Jar13 epitope GNA residue 281
“Protective” epitopes GNA residues 214-211 and 269
doi: 10.1038/nature07769 SUPPLEMENTARY INFORMATION
www.nature.com/nature 10

Figure 6: Comparison of the structures of fH67 in complex with fHbp and fH678 in
complex with SOS. Overlay of the seven, crystallographically independent copies of the
fH67 portion of the fh67:fHbp complex with the structure of fH6782 (in green, PDB ID:2UWN).
The position of CCP7 in the structure of fH678 is seen to lie in the middle of the range of
variation with respect to CCP6 seen in the structure of fH67 complexed to fHbp. The pattern
of thermal mobility factors is rather different (displayed as the thickness of the putty
representation) with the CCP6 helix that is buried in the fHbp complex seen to be less mobile
in the complex structure than in the earlier structure of fH678 in complex with sucrose
octasulphate.
fH CCP6 fH CCP7 fH CCP8
doi: 10.1038/nature07769 SUPPLEMENTARY INFORMATION
www.nature.com/nature 11

Figure 7: SOS inhibits binding of fH to N. meningitidis expressing fHbp on their
surface. Wild-type MC58 (fHbp+) or an isogenic mutant lacking fHbp (fHbp-) were incubated
with purified fH (225 mg/ml) with or without SOS as indicated. Binding was measured by
FACS analysis with an anti-human fH pAb and expressed as the mean fluorescence index
(MFI). SOS (100 mM, final concentration) inhibited binding of full length fH to bacteria
(Student’s T test, p<0.001).
doi: 10.1038/nature07769 SUPPLEMENTARY INFORMATION
www.nature.com/nature 12
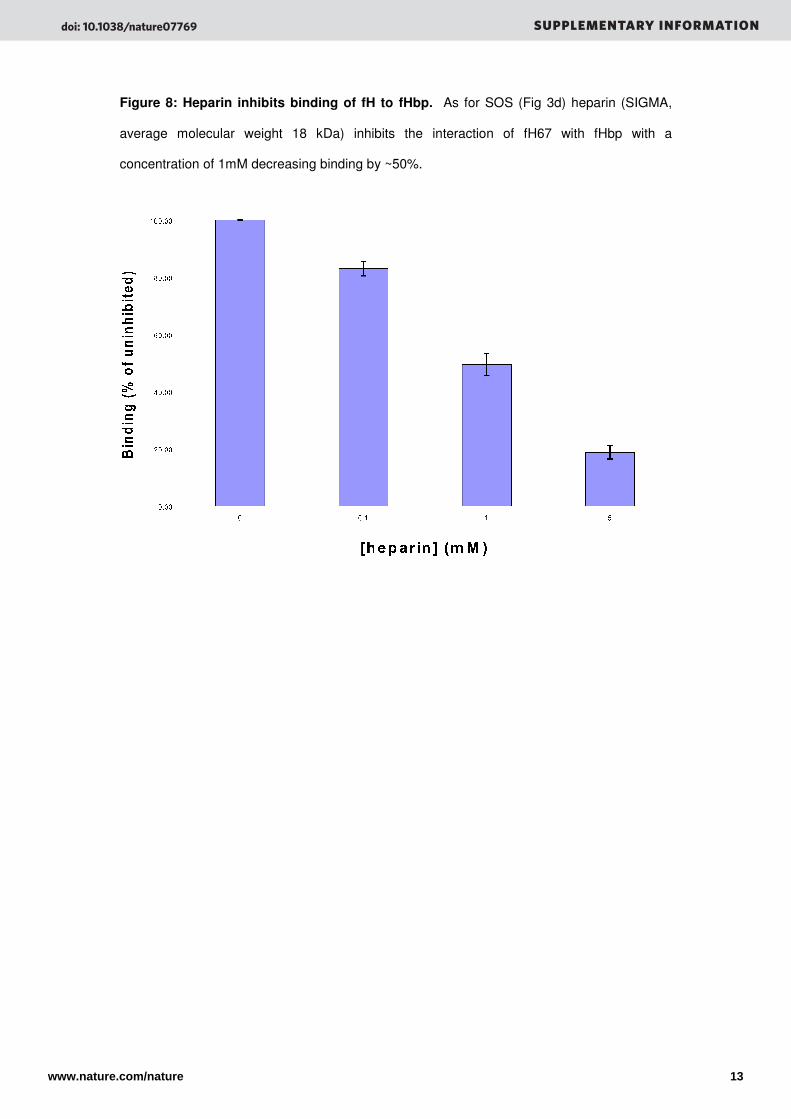
Figure 8: Heparin inhibits binding of fH to fHbp. As for SOS (Fig 3d) heparin (SIGMA,
average molecular weight 18 kDa) inhibits the interaction of fH67 with fHbp with a
concentration of 1mM decreasing binding by ~50%.
doi: 10.1038/nature07769 SUPPLEMENTARY INFORMATION
www.nature.com/nature 13
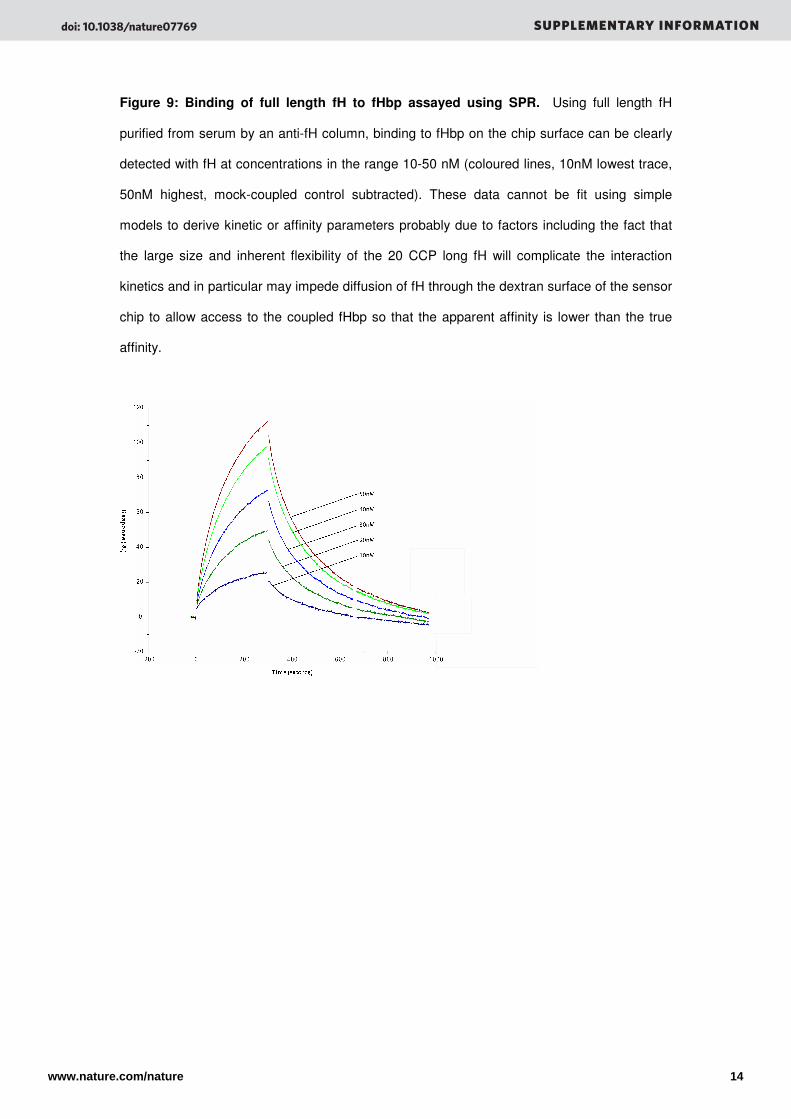
Figure 9: Binding of full length fH to fHbp assayed using SPR. Using full length fH
purified from serum by an anti-fH column, binding to fHbp on the chip surface can be clearly
detected with fH at concentrations in the range 10-50 nM (coloured lines, 10nM lowest trace,
50nM highest, mock-coupled control subtracted). These data cannot be fit using simple
models to derive kinetic or affinity parameters probably due to factors including the fact that
the large size and inherent flexibility of the 20 CCP long fH will complicate the interaction
kinetics and in particular may impede diffusion of fH through the dextran surface of the sensor
chip to allow access to the coupled fHbp so that the apparent affinity is lower than the true
affinity.
doi: 10.1038/nature07769 SUPPLEMENTARY INFORMATION
www.nature.com/nature 14

Figure 10: Comparison of the structures of fHbp with that of an engineered avidin SSM4
was used to overlay secondary structure elements (cutoff of 40% match) between the
structure of fHbp (green) and the earlier structure of an engineered avidin5 (blue). Although
the topology of the N-terminal β-barrel of fHbp is very different to that of the avidin barrels, the
relative arrangement of the pairs of barrels is seen to be grossly similar. This suggests that
packing of two β-barrels selects for particular crossing angles for maximum stability.
doi: 10.1038/nature07769 SUPPLEMENTARY INFORMATION
www.nature.com/nature 15

Figure 11: Comparison of the structure of fHbp with that of an complement Factor B
SSM was used to overlay secondary structure elements between the structure of fHbp
(green) and the earlier structure of complement Factor B 6 (red for overlaid portion and grey).
Although the topologies of the β-barrels are different, the relative arrangement of the pairs of
barrels is seen (again) to be grossly similar.
doi: 10.1038/nature07769 SUPPLEMENTARY INFORMATION
www.nature.com/nature 16

Supplemental References
1. Cantini, F. et al. Solution structure of the immunodominant domain of
protective antigen GNA1870 of Neisseria meningitidis. J Biol Chem 281,
7220-7 (2006).
2. Prosser, B.E. et al. Structural basis for complement factor H linked age-related
macular degeneration. J Exp Med 204, 2277-83 (2007).
3. Holm, L. & Sander, C. Dali: a network tool for protein structure comparison.
Trends Biochem Sci 20, 478-80 (1995).
4. Krissinel, E. & Henrick, K. Secondary-structure matching (SSM), a new tool
for fast protein structure alignment in three dimensions. Acta Crystallogr D
Biol Crystallogr 60, 2256-68 (2004).
5. Hytonen, V.P. et al. Controlling quaternary structure assembly: subunit
interface engineering and crystal structure of dual chain avidin. J Mol Biol
359, 1352-63 (2006).
6. Milder, F.J. et al. Factor B structure provides insights into activation of the
central protease of the complement system. Nat Struct Mol Biol 14, 224-8
(2007).
7. DeLano, W.L. on World Wide Web http://www.pymol.org(2002).
8. Giuliani, M.M. et al. The region comprising amino acids 100 to 255 of
Neisseria meningitidis lipoprotein GNA 1870 elicits bactericidal antibodies.
Infect Immun 73, 1151-60 (2005).
9. Welsch, J.A., Rossi, R., Comanducci, M. & Granoff, D.M. Protective activity
of monoclonal antibodies to genome-derived neisserial antigen 1870, a
Neisseria meningitidis candidate vaccine. J Immunol 172, 5606-15 (2004).
10. Beernink, P.T. & Granoff, D.M. Bactericidal antibody responses induced by
meningococcal recombinant chimeric factor H-binding protein vaccines. Infect
Immun 76, 2568-75 (2008).
doi: 10.1038/nature07769 SUPPLEMENTARY INFORMATION
www.nature.com/nature 17





![Analysis differences betweenNeisseria meningitidis Neisseria … · seria meningitidis (Nm)]withthatofthegonococcus[Neisseria gonorrhoeae (Ng)]. These two human pathogens are very](https://img.pdfslide.net/doc/110x75/5ea11fa2b5452c63b84dc792/analysis-differences-betweenneisseria-meningitidis-neisseria-seria-meningitidis.jpg)























