Embed Size (px)
Citation preview

[CANCER RESEARCH 47, 6324-6328, December I, 1987]
Nerve Growth Factor Stimulation of Mouse Embryonal Carcinoma Cell Migration1Brenda W. Kalian1 and Douglas C. Kramp
Department of Zoology [B. W. K.J and Integrated Microscopy Facility [D. C. K.J. University of Wisconsin, Madison, Wisconsin 53706
ABSTRACT
Embryonal carcinoma cells localize to a specific array of target tissuesincluding the male submaxillary gland following intracardiac injection.These target tissues or conditioned medium derived from them have beenshown to stimulate the in vitro migration of embryonal carcinoma cells.Here we show that in vitro migration of mouse embryonal carcinomacells is induced by the simultaneous presence of two different components.One of these is a specific requirement for a fibronectin substrate. Theother component is present in male submaxillary conditioned mediumand may be nerve growth factor (NGF). Migration of embryonal carcinoma cells on a fibronectin substrate could be induced by purified NGF,but not epidermal growth factor, and submaxillary conditioned mediumcontained very high levels of nerve outgrowth activity which could beblocked by anti-NGF antibody. Only the high molecular weight 7S NGFcomplex was active in inducing migration, while the low molecular weight2.5S NGF inhibited migration. Neither type of NGF or submaxillaryconditioned medium stimulated the in vitro growth of embryonal carcinoma cells. 7S NGF thus differentially affects murine embryonal carcinoma cells by inducing cell motility but not cell division. That embryonalcarcinoma cells require 7S NGF for their migration in vitro raises theinteresting possibility that these cells may respond similarly in vivo, andbe stimulated by tissue-specific high molecular weight NGF moleculesto migrate or extravasate into the parenchyma of target organs.
INTRODUCTION
Embryonal carcinoma stem cells from a variety of teratocar-cinoma tumor lines produce an organotypic metastatic patternfollowing their intracardiac injection into adult mice (1). Amigration assay based on phagokinetic tracks produced in latexbead monolayers demonstrated that tissue pieces or conditionedmedium from five of the six tumor sites could evoke extensivein vitro migration of the EC3 cells. The organs with such activity
included those particularly noted as target organs for the migration of embryonic germ cells (ovary and testis) or the production of neurotrophic factors (male submaxillary gland, iris,and whisker field). This paper further investigates the responsesevoked by conditioned medium from one particular tumor site,the male submaxillary gland. The results indicate that highmolecular weight NGF can stimulate EC cells to migrate invitro and therefore may play a role in the migration of thesecells into specific organs following their injection into the adult.The induction of migration of embryonal stem cells by NGF,which is known to be crucial to the development of sensory andsympathetic neurones, suggests that NGF may serve otherfunctions in the embryo unrelated to supportive developmentof the nervous system.
Received 1/5/87; revised 6/8/87, 9/2/87; accepted 9/10/87.The costs of publication of this article were defrayed in part by the payment
of page charges. This article must therefore be hereby marked advertisement inaccordance with 18 U.S.C. Section 1734 solely to indicate this fact.
1Supported by National Cancer Institute Grant CA33453 and a gift to theUniversity of Wisconsin Foundation.
2To whom requests for reprints should be addressed, at Department of
Zoology, University of Wisconsin, 1117 West Johnson Street, Madison, WI53706.
J The abbreviations used are: EC, embryonal carcinoma; CM, conditionedmedium: NGF, nerve growth factor; DMEM, Dulbecco's modified Eagle's medium; DPBS, Dulbecco's phosphate buffered saline; PCS, fetal calf serum; EGF,
epidermal growth factor.
MATERIALS AND METHODS
Cells and Culture Methods. LT clone 8 cells were propagated asdescribed previously (1). Cells were cultured without feeder cells forone transfer period before being used in migration assays. CM wasproduced as before (1) from the submaxillary glands of 3 to 4-monthold LT or ICR/HSD male mice, except that in some experiments theconcentration of starting gland material was increased to 80 mg wetweight/ml serum-free DMEM. CM protein concentrations were determined by the Lowry method as described in (1).
Migration Assay. The latex bead migration assay used previously wasfurther modified since considerable lot variation in Covasphere beads(types MX, FX, and CX; Covalent Tech. Corp.) was observed. Unsatisfactory lots either failed to attach to the substrate or resulted in littleor no migration in response to active material. All such bead lots couldnot be improved by the changes described below. Satisfactory resultswere obtained using four lots of plain latex beads from PolysciencesInc. or Sigma Chemical Co. Migration results were further improvedand submaxillary CM tracks were essentially equivalent to the best ofthose obtained with Covasphere beads if the plain latex beads were firstcoated with l mg/ml m albumin. The standard protocol developed usedovalbumin (Sigma) precoated 1.0-jim polystyrene microspheres (Poly-sciences no. 15712) added to 7-mm tissue culture wells precoated withhuman plasma fibronectin (generously provided by D. Mosher). Beads(15 ^1/ml) were incubated in 1 mg ovalbumin/ml DPBS lacking Ca2+and Mg2* for l h at 25°Con a reciprocal shaker and centrifugea at
10000 x g for 10 min. The ovalbumin solution was prepared freshweekly. Microwell plates were concurrently incubated for l h with 100Ml/well of fibronectin in 0.05 M carbonate buffer (pH 9.6) in a 5% CO2tissue culture incubator at 37°C.Wells were then rinsed twice with 200
¿il/wellDPBS and once with DMEM, leaving a 50-Mlvolume at eachstep to keep the bottom wet. Beads were filtered through a 3-Mmporesize Nucleopore chemotactic filter (Neuro Probe, Inc.) and added tofibronectin-coated wells in 200-pl serum-free DMEM/well at a concentration of approximately 1¿tlbeads (original volume)/well. An excessivenumber of beads was observed to reduce migration and optimumconcentrations were determined for each lot. Plates were centrifuged at4"C for 15 min at 500 x g after which 200 p\ of medium was removed
per well and replaced by 200 ti\ test solution. Cells were filtered througha 15 /mi nylon cloth and added at 200 cells/well in 50 /il DMEMcontaining 0.2% FCS.
Quantitäten of Migration. After 18-24 h, migration plates wererinsed four times with DPBS, with a 100-^tl volume remaining aftereach rinse in order to leave the monolayer undisturbed, and fixed byadding an equal volume of 25% glutaraldehyde for 15-30 min. Afterfour rinses of distilled water, the wells were totally aspirated and allowedto dry. Plates were placed on a dissecting microscope stage with eachwell centered over an adjustable iris diaphragm so that only a singlewell bottom area was illuminated from below by a standard microscopebulb. Light was adjusted by a regulated power supply (Lambda, I.I'
522 FM), which was independently monitored with a digital voltmeter(Heathkit no. IM-2215). The illuminated well bottom was imaged by aCCD video camera (RCA TC2811). The image was processed intowhite migration tracks versus black background and analyzed by a QX9200 real-time video image processor (Quantex Corp., Sunnyvale, CA).Image analysis gave a summed image intensity, referred to as themigration index, which is an estimate proportional to the percentageof well area over which the cells migrated, typically less than 5% of thetotal well area.
Growth Assay. Cells were inoculated into 96-well plates precoatedwith 5 fig/ml fibronectin at 1 x IO4 cells/well in 0.2 ml DMEM
containing 10% FCS. Plates were incubated for 16 h before being rinsedtwice with serum-free DMEM and renewed with DMEM supplemented
6324
on April 9, 2019. © 1987 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
MIGRATION OF EMBRYONAL CARCINOMA CELLS
with 0.5% FCS. After 24 h, this medium was replaced by test solutionsof DMEM containing 0.5% PCS and various concentrations of NGFor CM. A control growth curve was constructed from wells receiving0.5, 1, 2, 5, or 10% serum at this point. Cells were incubated for anadditional 24 h before [3H]thymidine (1 /tCi/well; 20 Ci/mmol, New
England Nuclear Corp.) was added to each well. After another 24 h,wells were rinsed and left in DMEM containing 1% PCS but no labelfor 2 h before 50 ß\of 40 mM EDTA was added per well for 20 min.The cells were then harvested and rinsed on filters that were dried andcounted in scintillation fluid.
NGF Solutions. 2.5S NGF (Sigma Chemical Co. and CollaborativeResearch, Inc.) was added to each migration or growth assay well froma stock solution made and diluted in DMEM containing 10 mg/mlovalbumin to produce wells having a final concentration of 500 ¿tg/mlovalbumin. Each dilution was performed using a fresh pipette tip andpolystyrene tubes. Reproducible dilutions of submaxillary CM materialdid not require these precautions. 7S NGF (Sigma Chemical Co. andCollaborative Research, Inc.) stock solutions used in growth assays andactive in migration assays were prepared at 200 Mgor more/ml DMEM,pH 7.4, or DPBS and diluted just before use in DMEM containing 100f<g/ml ovalbumin.
Nerve Growth Assays. Dorsal root ganglia from 8-day-old WhiteLeghorn chick embryos were placed in 7-mm tissue culture wellsprecoated with 10 ng/m\ laminin, generously supplied by D. Mosher.Laminili substrates were prepared as described above for fibronectin.Culture medium was DMEM supplemented with 10% calf serum.Ganglia prepared in triplicate for each solution were examined microscopically for neurite outgrowth after 18-22 h.
NGF Antibody. The MQ3-1 hybridoma cell line, secreting rat IgG-1monoclonal antibodies against mouse submaxillary gland ßNGF, wasproduced and the antibody characterized in the laboratory of E. Shooter(2). MC/3-I supernatant was obtained from P. Claude.
RESULTS
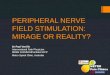
Migration Requires Both a Substrate Bound and a SolubleFactor. Fibronectin; laminin; collagens I, III, and IV; or heparincoated dishes were tested for their ability to support migrationin the presence and absence of 10.3 Mg/ml CM. In the absenceof CM, no migration was observed on any of the substrates(data not shown). However in the presence of CM, fibronectin,but none of the others, was able to support migration (Fig. 1).The response on fibronectin was dose-dependent with the maximum occurring when the dish was coated with 1 Mg/ml orgreater. Coating concentrations above 100 Mg/ml resulted inslightly reduced migration as compared with the peak response.The requirement for CM is specific; no movement occurred ona 5 Mg/ml fibronectin precoat in the presence of 5-500ovalbumin in solution (data not shown).
O FIBRONECTIN•LAMININ
COLLAGEN I
COLLAGEN mCOLLAGEN IVHEPARIN2.5
oZ
2.0
l5-
|..0
0.5
X-
0001 O.I I 10PRECOAT CONCENTRATION (
100
Fig. 1. Substrate requirement for in vitro migration of EC cells. LT clone 8cells were added to 10.3 Mg/ml submaxillary CM on latex bead monolayers inwells precoated with different substrates. Wells were fixed and migration trackareas were quantitated using an image processor and analyzer as described in"Materials and Methods."
Table I Effect on migration of different ways of adding CMAll wells were precoated with 20 </n'nil fibronectin. CM at 20.7 Mg/ml was
present in the well for 24 h during the assay or applied to the substrate alone orto the substrate and beads for 4 h. In the latter cases, wells were then rinsed sixtimes with DMEM before cells were added to bead monolayers in unconditionedDMEM and incubated for 24 h. Beads alone were coated for 1 h with 1.2 mg/mlCM instead of ovalbumin (see "Materials and Methods"), rinsed twice with
DPBS, and centrifuged to form a monolayer in the well; cells were added inunconditioned DMEM and incubated for 24 h.
Method of CM additionRelative
migration"
CM in well throughout assay 1.00Fibronectin substrate alone coated with CM 0.04Beads alone coated with CM 0.10Fibronectin and beads coated with CM 0.11
" Numbers are average migration index values of quadruplicate wells given as
a percentage of the maximum average obtained.
10SUBMAXILLARY CM (fig protein/ml)
100
Fig. 2. Effect of EGF on EC cell migration induced by submaxillary CM. LTclone 8 cells were added to latex bead monolayers in wells precoated with 20 Mg/ml fibronectin and containing different concentrations of submaxillary CM andEGF. Migration index values are averages of duplicate 7-mm wells. Similarresults were obtained in two other experiments.
The continuous presence of CM is required for migration onfibronectin. Coating either the fibronectin substrate or thebeads, or the beads and the fibronectin substrate with submaxillary CM was insufficient to produce good migration (Table1), and resulted in only 11% or less of the migration inducedwhen submaxillary CM remained in the well for the durationof the assay. EC cell migration thus appears to require twodifferent entities: a fibronectin substrate and a CM factor thatmust be continuously present in the solution. AH further migration experiments described were therefore performed using a5-20-Mg/ml fibronectin precoat in the continued presence oftest solutions.
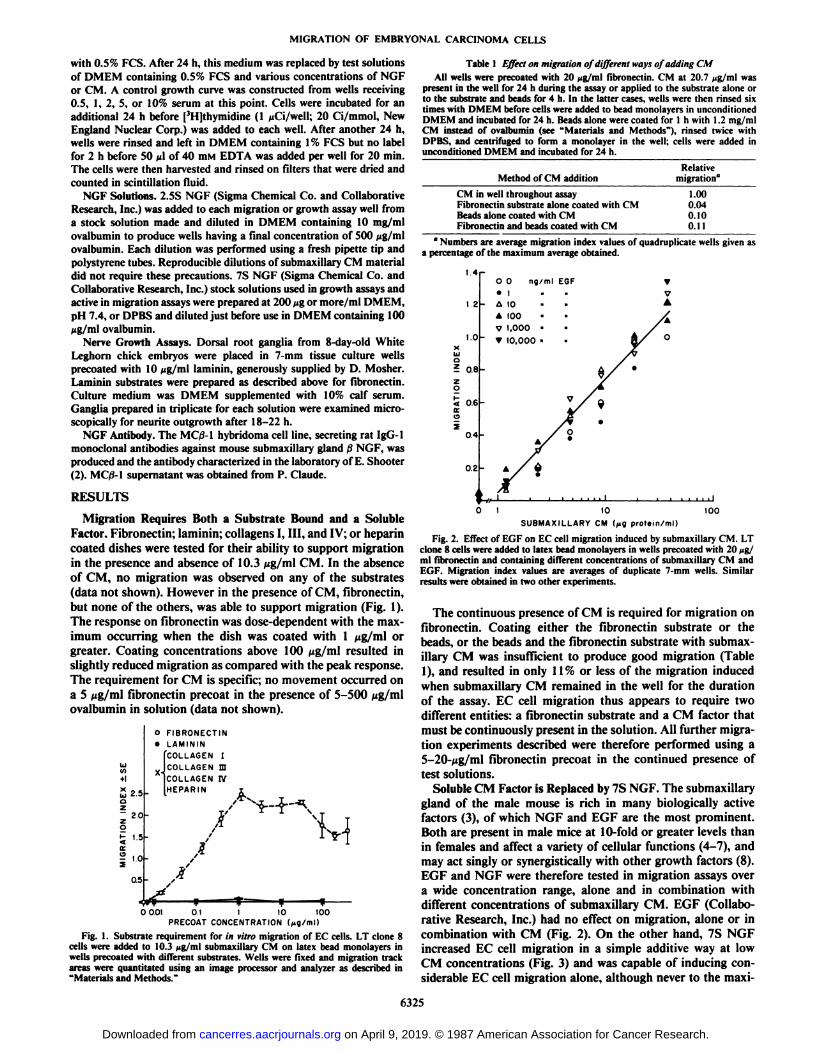
Soluble CM Factor is Replaced by 7S NGF. The submaxillarygland of the male mouse is rich in many biologically activefactors (3), of which NGF and EGF are the most prominent.Both are present in male mice at 10-fold or greater levels thanin females and affect a variety of cellular functions (4-7), andmay act singly or synergistically with other growth factors (8).EGF and NGF were therefore tested in migration assays overa wide concentration range, alone and in combination withdifferent concentrations of submaxillary CM. EGF (Collaborative Research, Inc.) had no effect on migration, alone or incombination with CM (Fig. 2). On the other hand, 7S NGFincreased EC cell migration in a simple additive way at lowCM concentrations (Fig. 3) and was capable of inducing considerable EC cell migration alone, although never to the maxi-
6325
on April 9, 2019. © 1987 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
MIGRATION OF EMBRYONAL CARCINOMA CELLS
ng/ml 7S NGF
1.0 10SUBMAXILLARY CM U g prot«in/ml)
Fig. 3. Effect of 7S NGF on EC cell migration induced by submaxillary CM.LT clone 8 cells were added to latex bead monolayers in wells precoated with 20ut; ml fibronectin and containing different concentrations of submaxillary CMand 7S NGF. Similar results were obtained using four different lots of 7S NGFfrom two suppliers in a total of 10 experiments.
30
•/ SUBMAXILLARY CM
O 2 9S N6FA 70S NSF
10
PROTEIN/ml
Fig. 4. Comparison of EC migration induced by submaxillary CM, 7S NGFor 2.SS NGF. LT clone 8 cells were plated on latex bead monolayers in wellsprecoated with S pg/ml fibronectin and containing different concentrations ofsuhmaxillary CM. 7S NGF or 2.5S NGF.
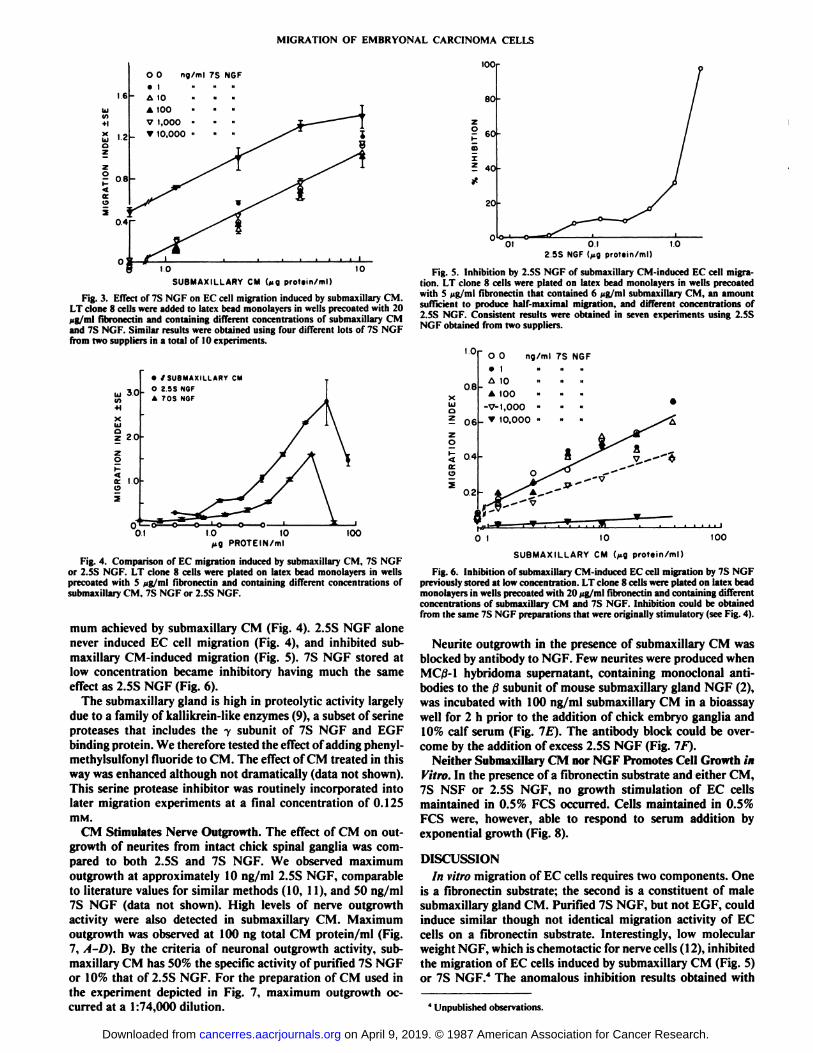
mum achieved by submaxillary CM (Fig. 4). 2.5S NGF alonenever induced EC cell migration (Fig. 4), and inhibited sub-maxillary CM-induced migration (Fig. 5). 7S NGF stored atlow concentration became inhibitory having much the sameeffect as 2.5S NGF (Fig. 6).
The submaxillary gland is high in proteolytic activity largelydue to a family of kallikrein-like enzymes (9), a subset of serineproteases that includes the y subunit of 7S NGF and EGFbinding protein. We therefore tested the effect of adding phenyl-methylsulfonyl fluoride to CM. The effect of CM treated in thisway was enhanced although not dramatically (data not shown).This serine protease inhibitor was routinely incorporated intolater migration experiments at a final concentration of 0.125IBM.
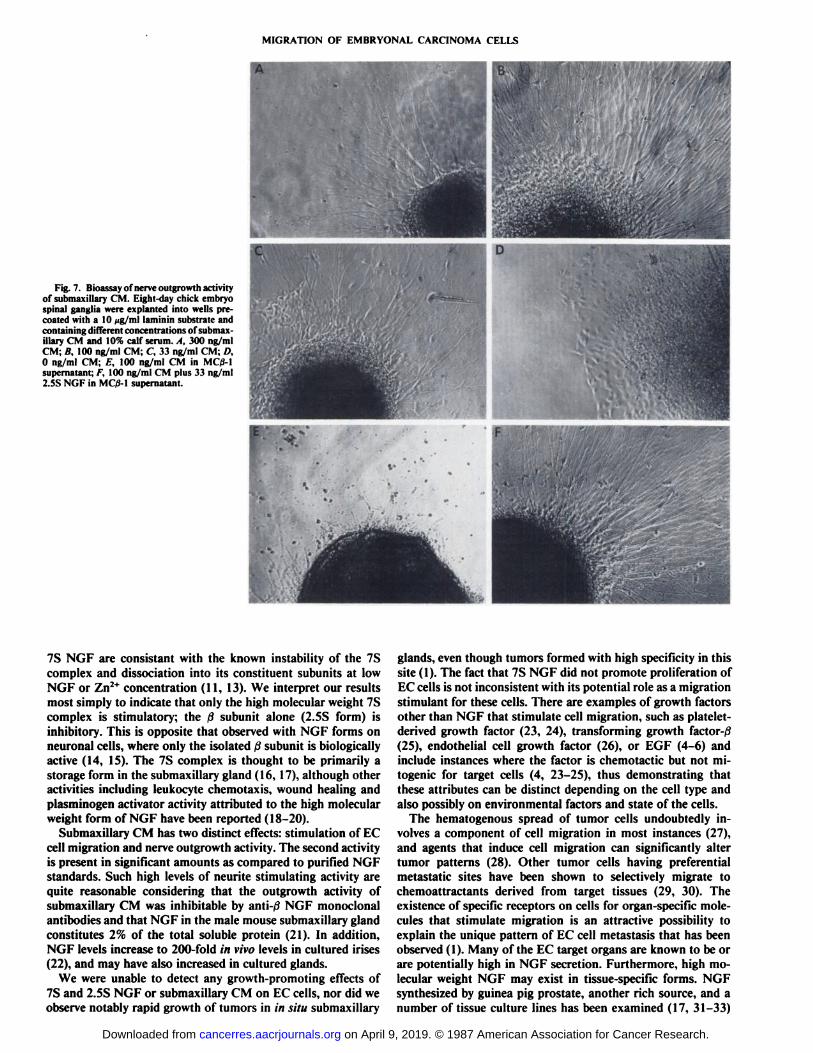
CM Stimulates Nerve Outgrowth. The effect of CM on outgrowth of neurites from intact chick spinal ganglia was compared to both 2.5S and 7S NGF. We observed maximumoutgrowth at approximately 10 ng/ml 2.5S NGF, comparableto literature values for similar methods (10, 11), and 50 ng/ml7S NGF (data not shown). High levels of nerve outgrowthactivity were also detected in submaxillary CM. Maximumoutgrowth was observed at 100 ng total CM protein/ml (Fig.7, A-D). By the criteria of neuronal outgrowth activity, submaxillary CM has 50% the specific activity of purified 7S NGFor 10% that of 2.5S NGF. For the preparation of CM used inthe experiment depicted in Fig. 7, maximum outgrowth occurred at a 1:74,000 dilution.
(OOr
80-
mE? 40f
20
O1«01 01 1.0
2 5S NGF( protein/ml)
Fig. 5. Inhibition by 2.SS NGF of submaxillary CM-induced EC cell migration. LT clone 8 cells were plated on latex bead monolayers in wells precoatedwith S fig/ml fibronectin that contained 6 Mg/ml submaxillary CM, an amountsufficient to produce half-maximal migration, and different concentrations of2.5S NGF. Consistent results were obtained in seven experiments using 2.5SNGF obtained from two suppliers.
tooSUBMAXILLARY CM protein/ml)
Fig. 6. Inhibition of submaxillary CM-induced EC cell migration by 7S NGFpreviously stored at low concentration. LT clone 8 cells were plated on latex beadmonolayers in wells precoated with 20 pg/ml fibronectin and containing differentconcentrations of submaxillary CM and 7S NGF. Inhibition could be obtainedfrom the same 7S NGF preparations that were originally stimulatory (see Fig. 4).
Neurite outgrowth in the presence of submaxillary CM wasblocked by antibody to NGF. Few neurites were produced whenMC/3-1 hybridoma supernatant, containing monoclonal antibodies to the ßsubunit of mouse submaxillary gland NGF (2),was incubated with 100 ng/ml submaxillary CM in a bioassaywell for 2 h prior to the addition of chick embryo ganglia and10% calf serum (Fig. IE). The antibody block could be overcome by the addition of excess 2.5S NGF (Fig. IF).
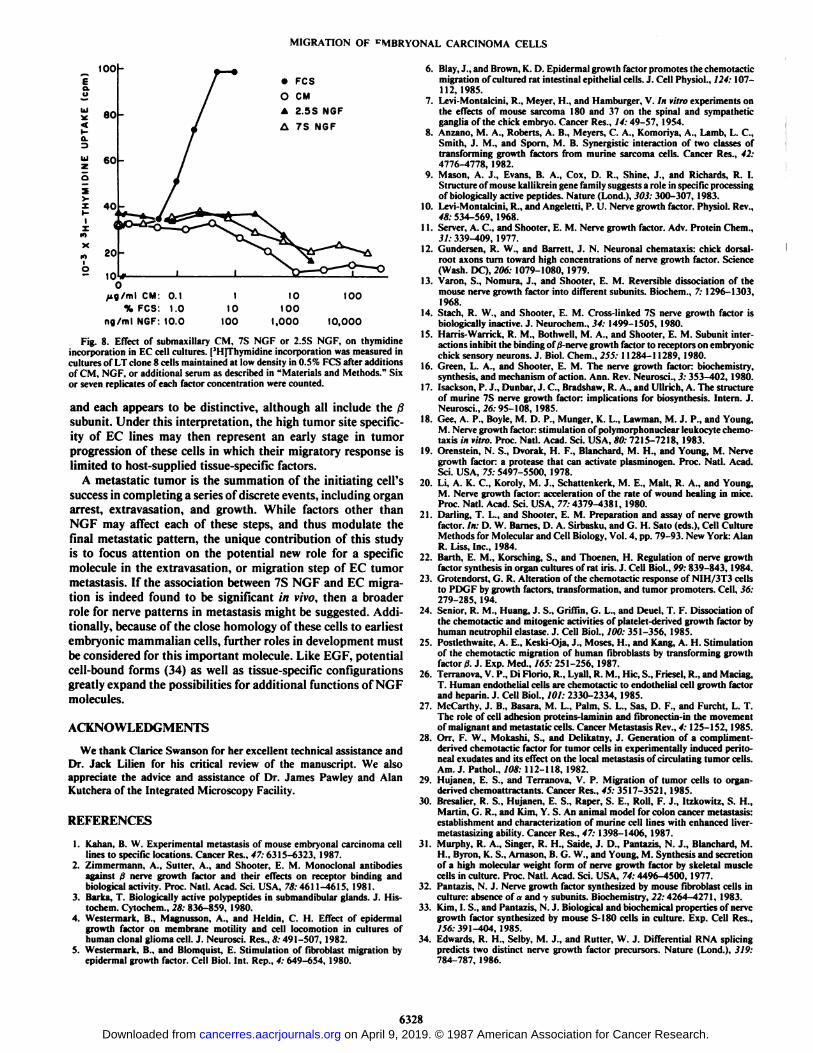
Neither Submaxillary CM nor NGF Promotes Cell Growth inVitro. In the presence of a fibronectin substrate and either CM,7S NSF or 2.5S NGF, no growth stimulation of EC cellsmaintained in 0.5% PCS occurred. Cells maintained in 0.5%PCS were, however, able to respond to serum addition byexponential growth (Fig. 8).
DISCUSSIONIn vitro migration of EC cells requires two components. One
is a fibronectin substrate; the second is a constituent of malesubmaxillary gland CM. Purified 7S NGF, but not EGF, couldinduce similar though not identical migration activity of ECcells on a fibronectin substrate. Interestingly, low molecularweight NGF, which is chemotactic for nerve cells ( 12), inhibitedthe migration of EC cells induced by submaxillary CM (Fig. 5)or 7S NGF.4 The anomalous inhibition results obtained with
' Unpublished observations.
on April 9, 2019. © 1987 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
MIGRATION OF EMBRYONAL CARCINOMA CELLS
Fig. 7. Bioassay of nerve outgrowth activityof submaxillary CM. Eight-day chick embryospinal ganglia were explanted into wells pre-coated with a 10 Mg/ml laminin substrate andcontaining dilTerent concentrations of submaxillary CM and 10% calf serum. A, 300 ng/mlCM; B, 100 ng/ml CM; C. 33 ng/ml CM; D,0 ng/ml CM; E, 100 ng/ml CM in MC/3-1supernatant; F, 100 ng/ml CM plus 33 ng/ml2.5S NGF in MC/3-1 supernatant.
7S NGF are consistant with the known instability of the 7Scomplex and dissociation into its constituent subunits at lowNGF or Zn2+ concentration (11, 13). We interpret our results
most simply to indicate that only the high molecular weight 7Scomplex is stimulatory; the ßsubunit alone (2.5S form) isinhibitory. This is opposite that observed with NGF forms onneuronal cells, where only the isolated ßsubunit is biologicallyactive (14, 15). The 7S complex is thought to be primarily astorage form in the submaxillary gland (16, 17), although otheractivities including leukocyte chemotaxis, wound healing andplasminogli activator activity attributed to the high molecularweight form of NGF have been reported (18-20).
Submaxillary CM has two distinct effects: stimulation of ECcell migration and nerve outgrowth activity. The second activityis present in significant amounts as compared to purified NGFstandards. Such high levels of neunte stimulating activity arequite reasonable considering that the outgrowth activity ofsubmaxillary CM was inhibitable by anti-0 NGF monoclonalantibodies and that NGF in the male mouse submaxillary glandconstitutes 2% of the total soluble protein (21). In addition,NGF levels increase to 200-fold in vivo levels in cultured irises(22), and may have also increased in cultured glands.
We were unable to detect any growth-promoting effects of7S and 2.5S NGF or submaxillary CM on EC cells, nor did weobserve notably rapid growth of tumors in in situ submaxillary
glands, even though tumors formed with high specificity in thissite (1). The fact that 7S NGF did not promote proliferation ofEC cells is not inconsistent with its potential role as a migrationstimulant for these cells. There are examples of growth factorsother than NGF that stimulate cell migration, such as platelet-derived growth factor (23, 24), transforming growth factor-^(25), endothelial cell growth factor (26), or EGF (4-6) andinclude instances where the factor is chemotactic but not mi-togenic for target cells (4, 23-25), thus demonstrating thatthese attributes can be distinct depending on the cell type andalso possibly on environmental factors and state of the cells.
The hematogenous spread of tumor cells undoubtedly involves a component of cell migration in most instances (27),and agents that induce cell migration can significantly altertumor patterns (28). Other tumor cells having preferentialmetastatic sites have been shown to selectively migrate tochemoattractants derived from target tissues (29, 30). Theexistence of specific receptors on cells for organ-specific molecules that stimulate migration is an attractive possibility toexplain the unique pattern of EC cell metastasis that has beenobserved (1). Many of the EC target organs are known to be orare potentially high in NGF secretion. Furthermore, high molecular weight NGF may exist in tissue-specific forms. NGFsynthesized by guinea pig prostate, another rich source, and anumber of tissue culture lines has been examined (17, 31-33)
on April 9, 2019. © 1987 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from
MIGRATION OF ^MBRYONAL CARCINOMA CELLS
100 -
•PCS
O CM
A 2.5S NGF
7S NGF
CM: 0.1% PCS: 1.0
ng/ml NGF: 10.0
I10
100
10
100
1,000
100
10,000
Fig. 8. Effect of submaxillary CM, 7S NGF or 2.5S NGF. on thymidineincorporation in EC cell cultures. [JH]Thymidine incorporation was measured in
cultures of LT clone 8 cells maintained at low density in 0.5% FCS after additionsof CM, NGF, or additional serum as described in "Materials and Methods." Six
or seven replicates of each factor concentration were counted.
and each appears to be distinctive, although all include the .;subunit. Under this interpretation, the high tumor site specificity of EC lines may then represent an early stage in tumorprogression of these cells in which their migratory response islimited to host-supplied tissue-specific factors.
A metastatic tumor is the summation of the initiating cell's
success in completing a series of discrete events, including organarrest, extravasation, and growth. While factors other thanNGF may affect each of these steps, and thus modulate thefinal metastatic pattern, the unique contribution of this studyis to focus attention on the potential new role for a specificmolecule in the extravasation, or migration step of EC tumormetastasis. If the association between 7S NGF and EC migration is indeed found to be significant /// vivo, then a broaderrole for nerve patterns in metastasis might be suggested. Additionally, because of the close homology of these cells to earliestembryonic mammalian cells, further roles in development mustbe considered for this important molecule. Like EGF, potentialcell-bound forms (34) as well as tissue-specific configurationsgreatly expand the possibilities for additional functions of NGFmolecules.
ACKNOWLEDGMENTS
We thank Clarice Swanson for her excellent technical assistance andDr. Jack Lilien for his critical review of the manuscript. We alsoappreciate the advice and assistance of Dr. James Pawley and AlanKutchera of the Integrated Microscopy Facility.
REFERENCES
1. Kahan, B. W. Experimental metastasis of mouse embryonal carcinoma celllines to specific locations. Cancer Res., 47: 6315-6323, 1987.
2. Zimmermann, A., Suiter, A., and Shooter. E. M. Monoclonal antibodiesagainst .; nerve growth factor and their effects on receptor binding andbiological activity. Proc. Nati. Acad. Sci. USA, 78:4611-4615, 1981.
3. Barka. T. Biologically active polypeptides in submandibular glands. J. His-tochem. Cytochem., 2«:836-859, 1980.
4. Westermark, B., Magnusson, A., and Heldin, C. H. Effect of epidermalgrowth factor on membrane motility and cell locomotion in cultures ofhuman clonal glioma cell. J. Neurosci. Res., fi: 491-507, 1982.
5. Westermark, B., and Blomquist, E. Stimulation of fibroblast migration byepidermal growth factor. Cell Biol. Int. Rep., 4:649-654, 1980.
6. Blay, J., and Brown, K. D. Epidermal growth factor promotes the chemotacticmigration of cultured rat intestinal epithelial cells. J. Cell Physiol., 124:107-112, 1985.
7. Levi-Montalcini, R., Meyer, H., and Hamburger, V. In vitro experiments onthe effects of mouse sarcoma 180 and 37 on the spinal and sympatheticganglia of the chick embryo. Cancer Res., 14:49-57, 1954.
8. Anzano, M. A., Roberts, A. B., Meyers, C. A., Komoriya, A., Lamb, L. C.,Smith, J. M., and Sporn, M. B. Synergistic interaction of two classes oftransforming growth factors from murine sarcoma cells. Cancer Res., 42:4776-4778, 1982.
9. Mason, A. J., Evans, B. A., Cox, D. R., Shine, J., and Richards, R. I.Structure of mouse kallikrein gene family suggests a role in specific processingof biologically active peptides. Nature (Lond.), 303: 300-307, 1983.
10. Levi-Montalcini, R., and Angeletti, P. U. Nerve growth factor. Physiol. Rev.,48: 534-569, 1968.
11. Server, A. C., and Shooter, E. M. Nerve growth factor. Adv. Protein Chem.,31: 339-409, 1977.
12. Gundersen, R. W., and Barrett, J. N. Neuronal chemataxis: chick dorsal-root axons turn toward high concentrations of nerve growth factor. Science(Wash. DC), 206: 1079-1080, 1979.
13. Varón, S., Nomura, J., and Shooter, E. M. Reversible dissociation of themouse nerve growth factor into different subunits. Biochem., 7: 1296-1303,1968.
14. Stach, R. W., and Shooter, E. M. Cross-linked 7S nerve growth factor isbiologically inactive. J. Neurochem., 34: 1499-1505, 1980.
15. Harris-Warrick, R. M., Bothwell, M. A., and Shooter, E. M. Subunit interactions inhibit the binding of 0-nerve growth factor to receptors on embryonicchick sensory neurons. J. Biol. Chem., 255: 11284-11289, 1980.
16. Green, L. A., and Shooter, E. M. The nerve growth factor: biochemistry,synthesis, and mechanism of action. Ann. Rev. Neurosci., 3:353-402, 1980.
17. Isackson, P. J., Dunbar, J. C., Bradshaw, R. A., and Ullrich, A. The structureof murine 7S nerve growth factor: implications for biosynthesis. Intern. J.Neurosci., 26:95-108, 1985.
18. Gee, A. P., Boyle, M. D. P., Munger, K. L., Lawman, M. J. P., and Young,M. Nerve growth factor: stimulation of polymorphonuclear leukocyte chemo-taxis in vitro. Proc. Nati. Acad. Sci. USA, 80:7215-7218, 1983.
19. Orenstein, N. S., Dvorak, H. F., Blanchard, M. H., and Young, M. Nervegrowth factor: a protease that can activate plasminogen. Proc. Nati. Acad.Sci. USA, 75: 5497-5500, 1978.
20. Li, A. K. C., Koroly, M. J., Schattenkerk, M. E., Malt, R. A., and Young,M. Nerve growth factor: acceleration of the rate of wound healing in mice.Proc. Nati. Acad. Sci. USA, 77:4379-4381, 1980.
21. Darling, T. L., and Shooter, E. M. Preparation and assay of nerve growthfactor. In: D. W. Barnes, D. A. Sirbasku, and G. H. Sato (eds.), Cell CultureMethods for Molecular and Cell Biology, Vol. 4, pp. 79-93. New York: AlanR. Liss, Inc., 1984.
22. Barth, E. M., Korsching, S., and Thoenen, H. Regulation of nerve growthfactor synthesis in organ cultures of rat iris. J. Cell Biol., 99:839-843,1984.
23. Grotendorst, G. R. Alteration of the chemotactic response of NIH/3T3 cellsto PDGF by growth factors, transformation, and tumor promoters. Cell, 36:279-285, 194.
24. Senior, R. M., Huang, J. S., Griffin, G. L., and Deuel, T. F. Dissociation ofthe chemotactic and mitogenic activities of platelet-derived growth factor byhuman neutrophil elastase. J. Cell Biol., 100: 351-356, 1985.
25. Postlethwaile, A. E., Keski-Oja, J., Moses, H., and Kang, A. H. Stimulationof the chemotactic migration of human fibroblasts by transforming growthfactor 0. J. Exp. Med., 165: 251-256, 1987.
26. Terranova, V. P., Di Florio, R., Lyall, R. M., Hie, S., Friesel, R., and Maciag,T. Human endothelial cells are chemotactic to endothelial cell growth factorand heparin. J. Cell Biol., 101: 2330-2334, 1985.
27. McCarthy, J. B., Basara, M. L., Palm, S. L., Sas, D. F., and Furcht, L. T.The role of cell adhesion proteins-laminin and fibronectin-in the movementof malignant and metastatic cells. Cancer Metastasis Rev., 4:125-152,1985.
28. Orr, F. W., Mokashi, S., and Delikatny, J. Generation of a compliment-derived chemotactic factor for tumor cells in experimentally induced peritoneal exúdalesand its effect on the local metastasis of circulating tumor cells.Am. J. Pathol., 108:112-118, 1982.
29. Hujanen, E. S., and Terranova, V. P. Migration of tumor cells to organ-derived chemoattractants. Cancer Res., 45: 3517-3521, 1985.
30. Bresalier, R. S., Hujanen, E. S., Râper,S. E., Roll, F. J., Itzkowitz, S. H.,Martin, G. R., and Kim, Y. S. An animal model for colon cancer metastasis:establishment and characterization of murine cell lines with enhanced liver-metastasizing ability. Cancer Res., 47: 1398-1406, 1987.
31. Murphy, R. A., Singer, R. H., Saide, J. D., Pantazis, N. J., Blanchard, M.H., Byron, K. S., Arnason, B. G. W., and Young, M. Synthesis and secretionof a high molecular weight form of nerve growth factor by skeletal musclecells in culture. Proc. Nati. Acad. Sci. USA, 74:4496-4500, 1977.
32. Pantazis, N. J. Nerve growth factor synthesized by mouse fibroblast cells inculture: absence of a and y subunits. Biochemistry, 22:4264-4271, 1983.
33. Kim, I. S., and Pantazis, N. J. Biological and biochemical properties of nervegrowth factor synthesized by mouse S-180 cells in culture. Exp. Cell Res.,756:391-404, 1985.
34. Edwards, R. H., Selby, M. J., and Rutler, W. J. Differential RNA splicingpredicts two distinct nerve growth factor precursors. Nature (Lond.), 319:784-787, 1986.
6328
on April 9, 2019. © 1987 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from

1987;47:6324-6328. Cancer Res Brenda W. Kahan and Douglas C. Kramp Carcinoma Cell MigrationNerve Growth Factor Stimulation of Mouse Embryonal
Updated version
http://cancerres.aacrjournals.org/content/47/23/6324
Access the most recent version of this article at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://cancerres.aacrjournals.org/content/47/23/6324To request permission to re-use all or part of this article, use this link
on April 9, 2019. © 1987 American Association for Cancer Research. cancerres.aacrjournals.org Downloaded from