Embed Size (px)
Citation preview

Network, nodes and nexus: systems approach to multitargettherapeuticsDivya Murthy1, Kuldeep Singh Attri1 and Rajesh S Gokhale1,2,3
Available online at www.sciencedirect.com
Systems biology is revealing multiple layers of regulatory
networks that manifest spatiotemporal variations. Since genes
and environment also influence the emergent property of a cell,
the biological output requires dynamic understanding of
various molecular circuitries. The metabolic networks
continually adapt and evolve to cope with the changing milieu
of the system, which could also include infection by another
organism. Such perturbations of the functional networks can
result in disease phenotypes, for instance tuberculosis and
cancer. In order to develop effective therapeutics, it is
important to determine the disease progression profiles of
complex disorders that can reveal dynamic aspects and to
develop mutitarget systemic therapies that can help overcome
pathway adaptations and redundancy.
Addresses1 CSIR-Institute of Genomics and Integrative Biology, Mall Road, Delhi,
India2 National Institute of Immunology, Aruna Asaf Ali Marg, New Delhi, India3 Jawaharlal Nehru Centre for Advanced Scientific Research, Bangalore,
India
Corresponding author: Gokhale, Rajesh S ([email protected])
Current Opinion in Biotechnology 2013, 24:1129–1136
This review comes from a themed issue on Pharmaceutical
biotechnology
Edited by Ajikumar Parayil and Federico Gago
For a complete overview see the Issue and the Editorial
Available online 28th February 2013
0958-1669/$ – see front matter, # 2013 Elsevier Ltd. All rights reserved.
http://dx.doi.org/10.1016/j.copbio.2013.02.009
IntroductionThe drug discovery process has been undergoing a suc-
cession of interesting paradigm shifts during different
periods of human evolution. While ancient practices of
medicine followed holistic approaches through careful
interpretations, the discovery of antimicrobial agents
during the beginning of modern biology era resulted
through serendipitous observations [1]. The next wave
was spurred by understanding of biological dogmas with
emphasis on enzyme functions and specificity of biomo-
lecular interactions. Concurrent increase in the under-
standing of the mechanisms of drug action along with
rapidly developing combinatorial chemistry encouraged
the emergence of target-based approaches for drug dis-
covery. The amalgamation of small molecule assemblages
and high-throughput screening resulted in an incredible
www.sciencedirect.com
growth in compound statistics [2–4]. However, the
benefits from these initiatives have been limited and
there has been an escalation in the cost of new molecular
entities at an annual rate of approximately 13.4% [5].
Although factors such as design of clinical trials and
economic decisions do add to the complexity of drug
discovery development, there is also an implicit need to
rewire the innovation programs based on systems-level
understanding of biological processes.
The advancement in acquiring high throughput data in
conjunction with heuristic network algorithms and
incredible imaging technologies is resetting many para-
digms of biological functionality. The remarkable flexi-
bility and redundancy that is being elucidated in cellular
circuitry implies that biology capitalizes survivability by
exploiting adaptability rather than utilizing the most
efficient systems. It appears that selectivity and speci-
ficity in the output of biological function is built-in
through multiple layers of sieve, many of which could
have interactions of low affinities and even low selectiv-
ity. Here we discuss new approaches and strategies of
systems biology that could be applied to drug discovery
programs. We discuss how understanding of metabolic
network-based study can delineate critical nodes and how
multitargeting could be rationally developed to generate
new classes of ‘systemic’ drugs.
Lacunae in understanding progression ofcomplex diseasesComplex disorders are multifactorial or polygenic dis-
orders whose outcomes could simultaneously involve
multiple perturbations. These phenotypes are further
complicated by lifestyle and environmental factors.
Although complex disorders often cluster in families,
there is no predictable pattern of inheritance. The robust-
ness of biological system that maximizes survival ensues
different downstream outcomes making it even harder to
determine the trigger. The challenge in unraveling such
complexity, therefore, is to define ‘Disease progression
profiles’ (DPPs) as shown in Figure 1a. While the trigger
initiates a cascade of cellular events that lead to onset of
disease (Phase I), the clinical diagnosis as well as disease
management primarily happens during the equilibrium
phase (Phase III). Much of the scientific research with
model systems is primarily performed in the time frame
that is somewhere between these two states (Phase II).
The challenge is to establish correlations between resul-
tant phenotypes and various dynamic interacting con-
stituents. An interesting example of a complex disorder
Current Opinion in Biotechnology 2013, 24:1129–1136
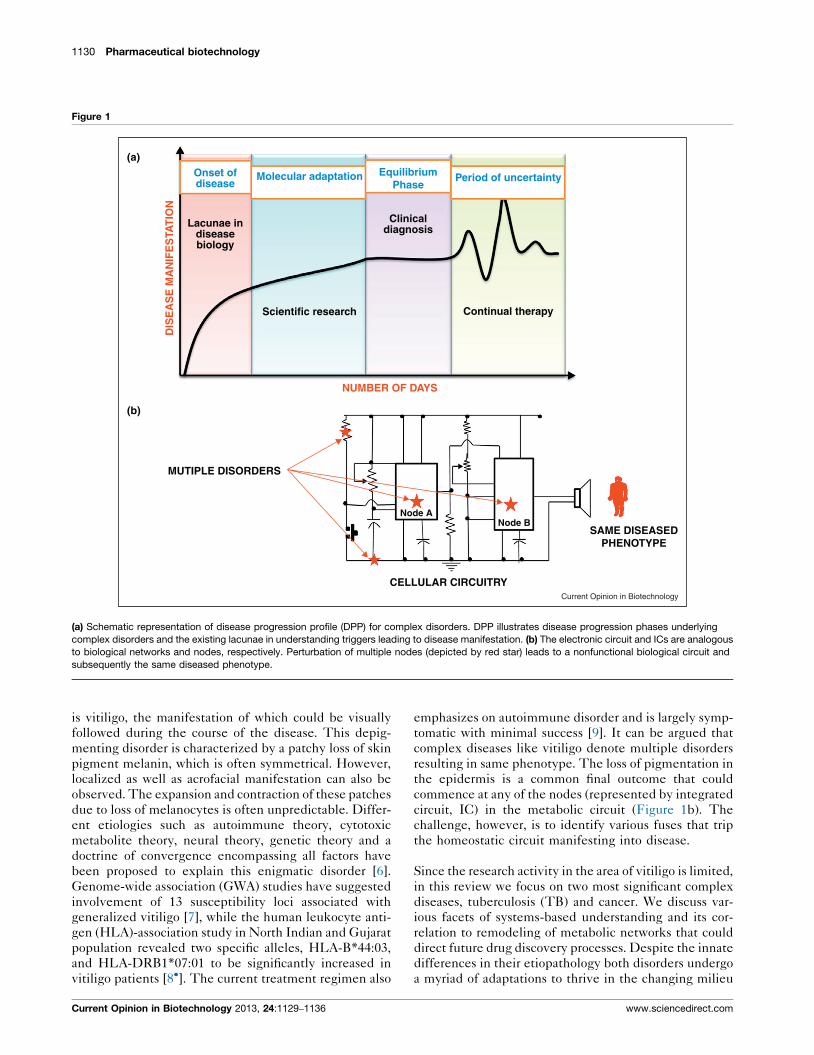
1130 Pharmaceutical biotechnology
Figure 1
(a)
(b)
Onset ofdisease
Molecular adaptation EquilibriumPhase
Period of uncertainty
Clinicaldiagnosis
Scientific research
NUMBER OF DAYS
DIS
EA
SE
MA
NIF
ES
TAT
ION
Continual therapy
Node ANode B
CELLULAR CIRCUITRY
MUTIPLE DISORDERS
SAME DISEASEDPHENOTYPE
Lacunae indiseasebiology
Current Opinion in Biotechnology
(a) Schematic representation of disease progression profile (DPP) for complex disorders. DPP illustrates disease progression phases underlying
complex disorders and the existing lacunae in understanding triggers leading to disease manifestation. (b) The electronic circuit and ICs are analogous
to biological networks and nodes, respectively. Perturbation of multiple nodes (depicted by red star) leads to a nonfunctional biological circuit and
subsequently the same diseased phenotype.
is vitiligo, the manifestation of which could be visually
followed during the course of the disease. This depig-
menting disorder is characterized by a patchy loss of skin
pigment melanin, which is often symmetrical. However,
localized as well as acrofacial manifestation can also be
observed. The expansion and contraction of these patches
due to loss of melanocytes is often unpredictable. Differ-
ent etiologies such as autoimmune theory, cytotoxic
metabolite theory, neural theory, genetic theory and a
doctrine of convergence encompassing all factors have
been proposed to explain this enigmatic disorder [6].
Genome-wide association (GWA) studies have suggested
involvement of 13 susceptibility loci associated with
generalized vitiligo [7], while the human leukocyte anti-
gen (HLA)-association study in North Indian and Gujarat
population revealed two specific alleles, HLA-B*44:03,
and HLA-DRB1*07:01 to be significantly increased in
vitiligo patients [8�]. The current treatment regimen also
Current Opinion in Biotechnology 2013, 24:1129–1136
emphasizes on autoimmune disorder and is largely symp-
tomatic with minimal success [9]. It can be argued that
complex diseases like vitiligo denote multiple disorders
resulting in same phenotype. The loss of pigmentation in
the epidermis is a common final outcome that could
commence at any of the nodes (represented by integrated
circuit, IC) in the metabolic circuit (Figure 1b). The
challenge, however, is to identify various fuses that trip
the homeostatic circuit manifesting into disease.
Since the research activity in the area of vitiligo is limited,
in this review we focus on two most significant complex
diseases, tuberculosis (TB) and cancer. We discuss var-
ious facets of systems-based understanding and its cor-
relation to remodeling of metabolic networks that could
direct future drug discovery processes. Despite the innate
differences in their etiopathology both disorders undergo
a myriad of adaptations to thrive in the changing milieu
www.sciencedirect.com

Network, nodes and nexus: systems approach to multitarget therapeutics Murthy, Attri and Gokhale 1131
and coping with extreme stress to ward off the host
deployed defense mechanisms. Despite considerable
investment in research and development, there is a
decreased efficiency in the new drug discovery pipeline.
The reductionist approach will yield only limited un-
derstanding of such complex systemic diseases and thus
multitarget interventions based on integrated network
analysis could prove to be an effective therapeutic
strategy.
Systems-based understanding of diseasesThe advent of the molecular biology era in the 1970s
incited the study of individual cellular components and
signaling events as independent cellular processes. How-
ever, it is now becoming clear that the behavior of a living
system may be hard to predict from the properties of the
individual components. Several paradoxical facts regard-
ing biological systems further complicate our ability to
predict deterministic outcome of biological function.
While genome sequencing studies have revealed that
the complexity of an organism cannot be directly corre-
lated to the size of the genome or to the number of genes,
the abundant and tissue-specific expression of noncoding
RNA (ncRNA) provides a different challenge to under-
stand regulatory aspects that shape our genome function
[10]. Many ncRNAs have also been implicated in a variety
of pathological disorders, including cancer and several
infectious diseases [11]. Interestingly, antituberculosis
drug streptomycin was recently shown to bind specifically
to pre-miRNA of miR-21 suggesting that therapeutic
agents could function through this mechanism [12].
While this shift to the significant role of RNA is increas-
ingly evident from various studies, these RNA molecules
could also act as cellular rheostats to fine-tune final
biological functions of proteins. Together, the ensemble
of protein and RNA networks would converge on meta-
bolic pathways. Remodeling of either networks can, in
turn, generate new metabolic adaptations and nodes for
cell survival.
Genome-wide analyses are now starting to reveal an
integrative perspective of disease biology. A recent gen-
ome-wide small interfering RNA (siRNA) screen con-
ducted to identify host factors that regulated the load of
Mycobacterium tuberculosis (Mtb) infection in human
macrophages has unraveled diverse host cell functional
modules that are engaged by the pathogen [13��]. This
interactome subset included immune, inflammatory and
stress pathways. While field strains of dissimilar genotype
showed variability in their disease manifestation by per-
turbing different networks, all the strains showed com-
monality in terms of inhibition of autophagy process. A
consequence of this mycobacterial adaptation is the for-
mation of the foamy macrophage phenotype that is
characterized by accumulation of lipid bodies (LBs)
[14]. More recently, the mechanism by which virulent
Mtb strains induce LB differentiation through rewiring of
www.sciencedirect.com
metabolic pathways was inferred [15�]. Infection in
macrophages leads to secretion of ESAT6 by Mtb, which
stimulates glucose uptake by the infected macrophage
cells. This enhances the flux of glycolysis leading to an
increase in the levels of acetyl coenzyme A that is parti-
tioned to generate more 3-hydroxybutyrate (3HB). 3HB
then activates GPR109A, which reduces the levels of
cyclic adenosine mono phosphate by repressing adenylyl
cyclase activity. Further inhibition of protein kinase A
causes recruitment of nonphosphorylated perilipin on
LBs that protects LBs from lipolysis. The mycobacterium
localizes into the LBs, which could be a new niche for
persistence.
This shift in host–pathogen axis that transpires in a switch
of interaction networks can also be exemplified in host-
specific diseases like cancer. Gene expression profiles as
well as genome sequencing comparisons have revealed
expression signatures and networks that can decipher
mechanisms responsible for the neoplastic conversion
of normal cells and response of cancer drugs to antitumor
agents [16]. The Connectivity Map provides a collection
of genome-wide transcriptional expression data from cul-
tured human cells that are treated with bioactive small
molecules [17]. On the basis of pattern-matching algor-
ithms this database enables functional connections be-
tween drugs, genes and diseases. ‘Omics’- platforms also
reveal information with regard to complex molecular
events that characterize cancer development and pro-
gression. For example, sarcosine, an N-methyl derivative
of the amino acid glycine, was identified as a differentially
regulated metabolite that was highly increased during
prostate cancer progression to metastasis [18]. Similarly,
proteomic studies of prostate cancer progression revealed
miR-128 as a potentially important negative regulator of
prostate cancer cell invasion [19]. Somatic mutational
analysis of 21 breast cancers has revealed combination
of substitution mutation and substantial variation in
number and pattern of indels in BRCA1 or BRCA2 genes
[20�]. Three genome sequencing studies have revealed
remarkable heterogeneity in lung cancer. Two of these
studies profiled the genomes of tissue samples from 178
patients with lung squamous cell carcinomas and 183 with
lung adenocarcinomas [21,22]. Both these studies reveal
complex changes across the genome and the challenge is
to convert this cataloged information into interconnected
networks thus permitting classification of patients for
effective personalized treatment. A third study analyzed
17 lung tumors to compare the genomes of smokers and
nonsmokers revealing striking differences of higher and
diverse mutational types in smokers’ tumors [23�]. Inter-
estingly, the patterns of mutations found in lung squa-
mous cell carcinoma more closely resemble those seen in
squamous cell carcinomas of the head and neck instead of
other lung cancers. The tumor classification thus could
follow molecular profiles rather than origin and could be
helpful in deciding right drug therapy.
Current Opinion in Biotechnology 2013, 24:1129–1136
1132 Pharmaceutical biotechnology
Metabolic remodeling and multitargeting‘systemic’ drugsAnother challenge in drug development is to interpret
cellular heterogeneity and understand how ensemble
behavior of population differs from individual cells.
Recently asymmetric cell division was demonstrated in
mycobacterium to give rise to subpopulation of cells that
were differentially susceptible to antibiotics. Such mech-
anisms are proposed to generate cell-to-cell heterogeneity
in the face of environmental stress enabling populations
to better withstand adversities [24]. Analogously tumor
evolution and adaptation also involves intratumor hetero-
geneity. This occurs by not only acquiring genetic
mutations but also by rewiring new pathways and possibly
also by perturbing the dynamic metabolic flux [25]. It
would be of tremendous value to understand the com-
plete bandwidth of metabolic processes that a cell pos-
sesses, which could be utilized during different stages of
stress and adaptations.
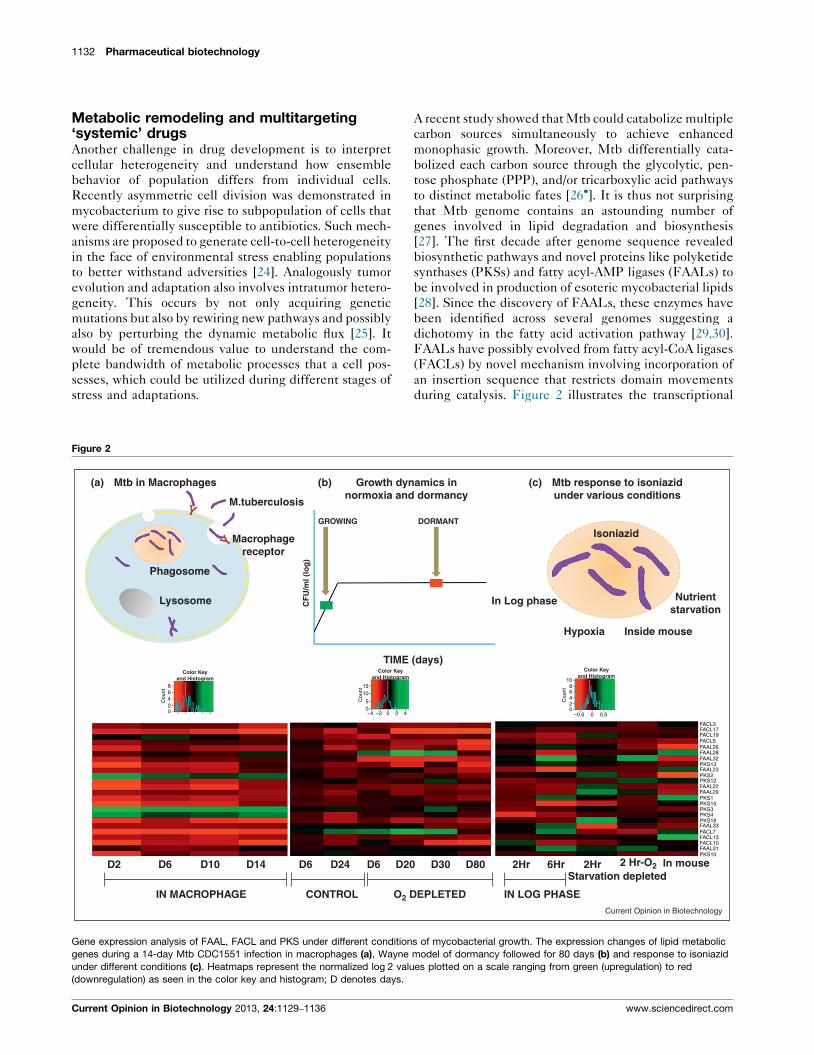
Figure 2
(a) (b) Growth dynnormoxia and
M.tuberculosis
Phagosome
Macrophagereceptor
Lysosome
Color Keyand Histogram
D2 D6 D10 D14 D6 D6D24 D20
CONTROLIN MACROPHAGE O2 D
GROWING
CF
U/m
l (lo
g)
TIME
Mtb in Macrophages
86420
Cou
nt
Color Keyand Histogram
1510
50–4 –2 0 2 4
Cou
nt
Gene expression analysis of FAAL, FACL and PKS under different condition
genes during a 14-day Mtb CDC1551 infection in macrophages (a), Wayne
under different conditions (c). Heatmaps represent the normalized log 2 valu
(downregulation) as seen in the color key and histogram; D denotes days.
Current Opinion in Biotechnology 2013, 24:1129–1136
A recent study showed that Mtb could catabolize multiple
carbon sources simultaneously to achieve enhanced
monophasic growth. Moreover, Mtb differentially cata-
bolized each carbon source through the glycolytic, pen-
tose phosphate (PPP), and/or tricarboxylic acid pathways
to distinct metabolic fates [26�]. It is thus not surprising
that Mtb genome contains an astounding number of
genes involved in lipid degradation and biosynthesis
[27]. The first decade after genome sequence revealed
biosynthetic pathways and novel proteins like polyketide
synthases (PKSs) and fatty acyl-AMP ligases (FAALs) to
be involved in production of esoteric mycobacterial lipids
[28]. Since the discovery of FAALs, these enzymes have
been identified across several genomes suggesting a
dichotomy in the fatty acid activation pathway [29,30].
FAALs have possibly evolved from fatty acyl-CoA ligases
(FACLs) by novel mechanism involving incorporation of
an insertion sequence that restricts domain movements
during catalysis. Figure 2 illustrates the transcriptional
amics in dormancy
(c) Mtb response to isoniazidunder various conditions
Isoniazid
Nutrientstarvation
Inside mouseHypoxia
In Log phase
D30 D80 2Hr 6Hr 2Hr In mouseStarvation depleted
IN LOG PHASEEPLETED
2 Hr-O2
DORMANT
(days)Color Key
and Histogram1086420
–0.5 0.50
FACL3FACL17FACL19FACL5FAAL26FAAL28FAAL32
FAAL23
FAAL29
FACL7FACL13FACL10FAAL31
FAAL33
FAAL22
PKS13
PKS2
PKS1PKS15PKS3PKS4PKS18
PKS10
PKS12
Cou
nt
Current Opinion in Biotechnology
s of mycobacterial growth. The expression changes of lipid metabolic
model of dormancy followed for 80 days (b) and response to isoniazid
es plotted on a scale ranging from green (upregulation) to red
www.sciencedirect.com
Network, nodes and nexus: systems approach to multitarget therapeutics Murthy, Attri and Gokhale 1133
regulation of some of the FAALs and FACLs during
varying conditions of Mtb growth [31–33]. Whereas
FAALs are involved in producing complex lipids such
as phthiocerol dimycocerosate, mycobactin, sulfolipids,
mycolic acids, polyacyl trehalose, the functional signifi-
cance of only FACL5, FACL3 and FACL19 has been
deciphered in lipid catabolism. Although complete path-
way is not yet known, mycobacteria can also utilize host
lipids such as triglycerides and cholesterol as nutrient
source. Studies also suggest that mycobacteria could even
utilize their own lipids such as mycolic acids for survival
[34–36]. All these studies emphasize that FAAL and
FACL are critical nodes for carbon metabolism of myco-
bacteria and simultaneous inhibition of these enzymes
may be important for therapeutic intervention. On the
basis of structural and mechanistic similarities within all
34 FAAL and FACL homologues along with acyl-aden-
ylate being a common reaction intermediate a nonhydro-
lysable analog of this bi-substrate was examined as
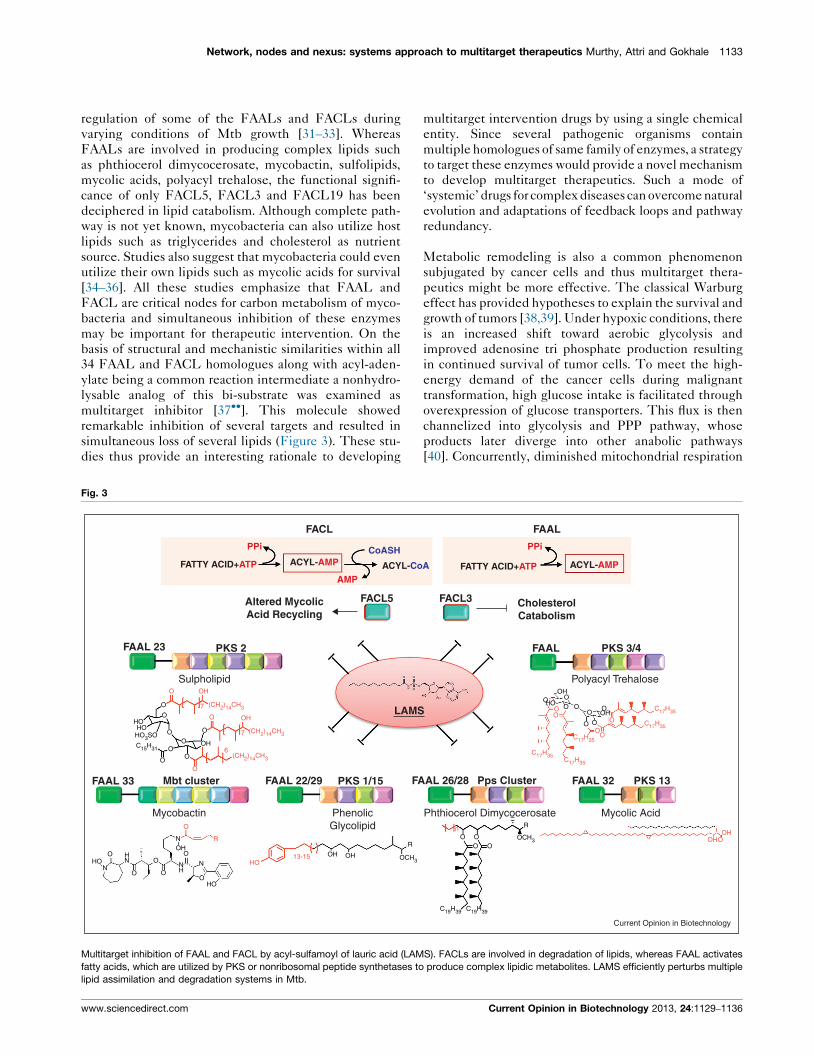
multitarget inhibitor [37��]. This molecule showed
remarkable inhibition of several targets and resulted in
simultaneous loss of several lipids (Figure 3). These stu-
dies thus provide an interesting rationale to developing
Fig. 3
PPi CoASH
FATTY ACID+ATP ACYL-AMP ACYL-Co
FACL
FACL5Altered MycolicAcid Recycling
FAAL 23
FAAL 33 Mbt cluster FAAL 22/29 FAPKS 1/15
PKS 2
Sulpholipid
Mycobactin PhenolicGlycolipid
O
O
O
O
HO13-15
R
OH
OH
7
7
6
(CH2)14CH3
(CH2)14CH3
(CH2)14CH3
LAMS
AMP
O
O
OO
OO
O
O
O
O
O
O
O
HO
HO
OHOH OH
R
OCH3
N
NN N
H
HN
C15 H31
HO3SOHO
HOO
OH
Multitarget inhibition of FAAL and FACL by acyl-sulfamoyl of lauric acid (LAM
fatty acids, which are utilized by PKS or nonribosomal peptide synthetases to
lipid assimilation and degradation systems in Mtb.
www.sciencedirect.com
multitarget intervention drugs by using a single chemical
entity. Since several pathogenic organisms contain
multiple homologues of same family of enzymes, a strategy
to target these enzymes would provide a novel mechanism
to develop multitarget therapeutics. Such a mode of
‘systemic’ drugs for complex diseases can overcome natural
evolution and adaptations of feedback loops and pathway
redundancy.
Metabolic remodeling is also a common phenomenon
subjugated by cancer cells and thus multitarget thera-
peutics might be more effective. The classical Warburg
effect has provided hypotheses to explain the survival and
growth of tumors [38,39]. Under hypoxic conditions, there
is an increased shift toward aerobic glycolysis and
improved adenosine tri phosphate production resulting
in continued survival of tumor cells. To meet the high-
energy demand of the cancer cells during malignant
transformation, high glucose intake is facilitated through
overexpression of glucose transporters. This flux is then
channelized into glycolysis and PPP pathway, whose
products later diverge into other anabolic pathways
[40]. Concurrently, diminished mitochondrial respiration
PPi
FATTY ACID+ATP ACYL-AMPA
CholesterolCatabolism
FACL3
FAAL
FAAL PKS 3/4
AL 26/28 Pps Cluster FAAL 32 PKS 13
Phthiocerol Dimycocerosate Mycolic Acid
Polyacyl Trehalose
O
OO
O
O
nOH
OHO
C17 H35C17 H35
C17 H35
C17 H35
C17 H35
HOO OO
O
O O
OO
OH
OH
O OO O
R
OCH3
C19 H39 C19 H39
Current Opinion in Biotechnology
S). FACLs are involved in degradation of lipids, whereas FAAL activates
produce complex lipidic metabolites. LAMS efficiently perturbs multiple
Current Opinion in Biotechnology 2013, 24:1129–1136
1134 Pharmaceutical biotechnology
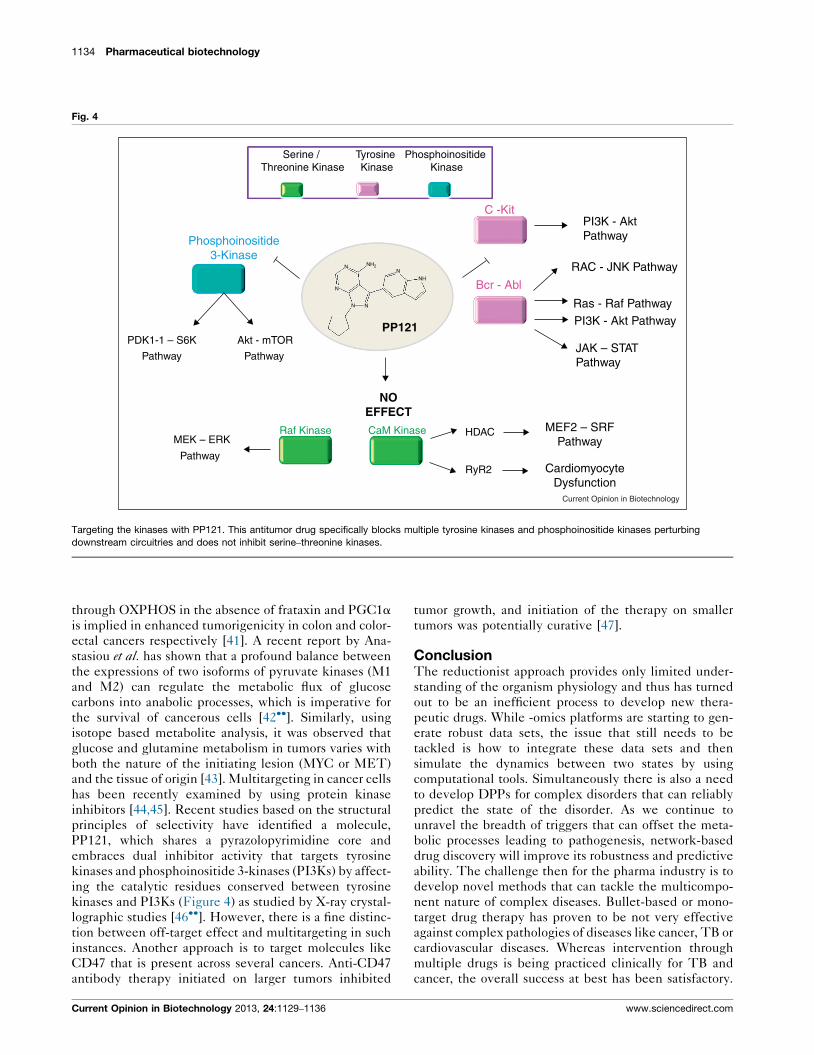
Fig. 4
PDK1-1 – S6K Akt - mTOR
Pathway Pathway
MEK – ERKRaf Kinase CaM Kinase HDAC
RyR2
MEF2 – SRFPathway
JAK – STATPathway
PI3K - AktPathway
C -Kit
Bcr - Abl
Phosphoinositide3-Kinase
Serine / Threonine Kinase
Tyrosine Kinase
N
N
N N
NNH
Phosphoinositide Kinase
PI3K - Akt Pathway
Ras - Raf Pathway
RAC - JNK Pathway
CardiomyocyteDysfunction
Pathway
NOEFFECT
PP121
Current Opinion in Biotechnology
Targeting the kinases with PP121. This antitumor drug specifically blocks multiple tyrosine kinases and phosphoinositide kinases perturbing
downstream circuitries and does not inhibit serine–threonine kinases.
through OXPHOS in the absence of frataxin and PGC1a
is implied in enhanced tumorigenicity in colon and color-
ectal cancers respectively [41]. A recent report by Ana-
stasiou et al. has shown that a profound balance between
the expressions of two isoforms of pyruvate kinases (M1
and M2) can regulate the metabolic flux of glucose
carbons into anabolic processes, which is imperative for
the survival of cancerous cells [42��]. Similarly, using
isotope based metabolite analysis, it was observed that
glucose and glutamine metabolism in tumors varies with
both the nature of the initiating lesion (MYC or MET)
and the tissue of origin [43]. Multitargeting in cancer cells
has been recently examined by using protein kinase
inhibitors [44,45]. Recent studies based on the structural
principles of selectivity have identified a molecule,
PP121, which shares a pyrazolopyrimidine core and
embraces dual inhibitor activity that targets tyrosine
kinases and phosphoinositide 3-kinases (PI3Ks) by affect-
ing the catalytic residues conserved between tyrosine
kinases and PI3Ks (Figure 4) as studied by X-ray crystal-
lographic studies [46��]. However, there is a fine distinc-
tion between off-target effect and multitargeting in such
instances. Another approach is to target molecules like
CD47 that is present across several cancers. Anti-CD47
antibody therapy initiated on larger tumors inhibited
Current Opinion in Biotechnology 2013, 24:1129–1136
tumor growth, and initiation of the therapy on smaller
tumors was potentially curative [47].
ConclusionThe reductionist approach provides only limited under-
standing of the organism physiology and thus has turned
out to be an inefficient process to develop new thera-
peutic drugs. While -omics platforms are starting to gen-
erate robust data sets, the issue that still needs to be
tackled is how to integrate these data sets and then
simulate the dynamics between two states by using
computational tools. Simultaneously there is also a need
to develop DPPs for complex disorders that can reliably
predict the state of the disorder. As we continue to
unravel the breadth of triggers that can offset the meta-
bolic processes leading to pathogenesis, network-based
drug discovery will improve its robustness and predictive
ability. The challenge then for the pharma industry is to
develop novel methods that can tackle the multicompo-
nent nature of complex diseases. Bullet-based or mono-
target drug therapy has proven to be not very effective
against complex pathologies of diseases like cancer, TB or
cardiovascular diseases. Whereas intervention through
multiple drugs is being practiced clinically for TB and
cancer, the overall success at best has been satisfactory.
www.sciencedirect.com

Network, nodes and nexus: systems approach to multitarget therapeutics Murthy, Attri and Gokhale 1135
Metabolic remodeling and network analysis can reveal
important functional cellular circuits that regulate diverse
phenotypes and thus could overcome the present pro-
blem of developed drug resistance. A multitargeting
approach for multifamily proteins could prove to be a
useful rational approach to developing systemic drugs.
The traditional knowledge of multicomponent herbal
drugs that by nature are multitargeting could provide
important resources for network-based drug discovery
efforts. At the same time gut microbiota as well as other
modes of delivering active ingredients could provide
guidance for combination regimes [48]. Finally, strategic
mindset changes by adopting new models like open
source drug discovery (OSDD) for certain diseases could
prove beneficial for successfully affording quality health-
care in future [49,50].
AcknowledgementsDM and KSA are CSIR Senior Research Fellows. RSG thanks Council ofScientific and Industrial Research (CSIR) and Department of Biotechnology(DBT) for grants to CSIR-IGIB and NII.
References and recommended readingPapers of particular interest, published within the period of review,have been highlighted as:
� of special interest
�� of outstanding interest
1. Newman DJ, Cragg GM, Snader KM: The influence of naturalproducts upon drug discovery. Nat Prod Rep 2000, 17:215-234.
2. Christophe T, Jackson M, Jeon HK, Fenistein D, Contreras-Dominguez M, Kim J, Genovesio A, Carralot JP, Ewann F, Kim EHet al.: High content screening identifies decaprenyl-phosphoribose 20 epimerase as a target for intracellularantimycobacterial inhibitors. PLoS Pathog 2009, 5:e1000645.
3. Ananthan S, Faaleolea ER, Goldman RC, Hobrath JV, Kwong CD,Laughon BE, Maddry JA, Mehta A, Rasmussen L, Reynolds RCet al.: High-throughput screening for inhibitors ofMycobacterium tuberculosis H37Rv. Tuberculosis (Edinb) 2009,89:334-353.
4. Garnett MJ, Edelman EJ, Heidorn SJ, Greenman CD, Dastur A,Lau KW, Greninger P, Thompson IR, Luo X, Soares J et al.:Systematic identification of genomic markers of drugsensitivity in cancer cells. Nature 2012, 483:570-575.
5. Munos B: Lessons from 60 years of pharmaceutical innovation.Nat Rev Drug Discov 2009, 8:959-968.
6. Le Poole IC, Das PK, van den Wijngaard RM, Bos JD,Westerhof W: Review of the etiopathomechanism of vitiligo: aconvergence theory. Exp Dermatol 1993, 2:145-153.
7. Jin Y, Birlea SA, Fain PR, Ferrara TM, Ben S, Riccardi SL, Cole JB,Gowan K, Holland PJ, Bennett DC et al.: Genome-wideassociation analyses identify 13 new susceptibility loci forgeneralized vitiligo. Nat Genet 2012, 44:676-680.
8.�
Singh A, Sharma P, Kar HK, Sharma VK, Tembhre MK, Gupta S,Laddha NC, Dwivedi M, Begum R, Gokhale RS et al.: HLA allelesand amino-acid signatures of the peptide-binding pocketsof HLA molecules in vitiligo. J Invest Dermatol 2012,132:124-134.
The role of alleles of HLA class I and class II loci was examined tounderstand the manifestation of vitiligo in two distinct Indian populationsand subsequently identifying three specific alleles associated with vitiligo.
9. Schallreuter KU, Bahadoran P, Picardo M, Slominski A,Elassiuty YE, Kemp EH, Giachino C, Liu JB, Luiten RM, Lambe Tet al.: Vitiligo pathogenesis: autoimmune disease, geneticdefect, excessive reactive oxygen species, calcium
www.sciencedirect.com
imbalance, or what else? Exp Dermatol 2008, 17:139-140discussion 141–160.
10. Mattick JS: Challenging the dogma: the hidden layer of non-protein-coding RNAs in complex organisms. Bioessays 2003,25:930-939.
11. Plummer PN, Freeman R, Taft R, Vider J, Sax M, Umer BA, Gao D,Johns CA, Mattick JS, Wilton SD et al.: MicroRNAs regulatetumor angiogenesis modulated by endothelial progenitorcells. Cancer Res 2012, 73:341-352.
12. Bose D, Jayaraj G, Suryawanshi H, Agarwala P, Pore SK,Banerjee R, Maiti S: The tuberculosis drug streptomycin as apotential cancer therapeutic: inhibition of miR-21 function bydirectly targeting its precursor. Angew Chem Int Ed Engl 2012,51:1019-1023.
13.��
Kumar D, Nath L, Kamal MA, Varshney A, Jain A, Singh S, Rao KV:Genome-wide analysis of the host intracellular network thatregulates survival of Mycobacterium tuberculosis. Cell 2010,140:731-743.
This is the first report for constructing functional networks throughmicroarray and genome-wide siRNA screen in macrophages infectedwith Mycobacterium tuberculosis.
14. Peyron P, Vaubourgeix J, Poquet Y, Levillain F, Botanch C,Bardou F, Daffe M, Emile JF, Marchou B, Cardona PJ et al.: Foamymacrophages from tuberculous patients’ granulomasconstitute a nutrient-rich reservoir for M. tuberculosispersistence. PLoS Pathog 2008, 4:e1000204.
15.�
Singh V, Jamwal S, Jain R, Verma P, Gokhale R, Rao KV:Mycobacterium tuberculosis driven targeted recalibration ofmacrophage lipid homeostasis promotes the foamyphenotype. Cell Host Microbe 2012, 12:669-681.
This report deciphers mechanism for foamy phenotype in macrophagesby diverting glycolytic pathway toward ketone body synthesis. Thisderegulation results in activation of GPR109A leading to perturbationof lipid homeostasis.
16. Rhodes DR, Yu J, Shanker K, Deshpande N, Varambally R,Ghosh D, Barrette T, Pandey A, Chinnaiyan AM: Large-scalemeta-analysis of cancer microarray data identifies commontranscriptional profiles of neoplastic transformation andprogression. Proc Natl Acad Sci U S A 2004, 101:9309-9314.
17. Lamb J, Crawford ED, Peck D, Modell JW, Blat IC, Wrobel MJ,Lerner J, Brunet JP, Subramanian A, Ross KN et al.: Theconnectivity map: using gene-expression signatures toconnect small molecules, genes, and disease. Science 2006,313:1929-1935.
18. Sreekumar A, Poisson LM, Rajendiran TM, Khan AP, Cao Q, Yu J,Laxman B, Mehra R, Lonigro RJ, Li Y et al.: Metabolomic profilesdelineate potential role for sarcosine in prostate cancerprogression. Nature 2009, 457:910-914.
19. Khan AP, Poisson LM, Bhat VB, Fermin D, Zhao R, Kalyana-Sundaram S, Michailidis G, Nesvizhskii AI, Omenn GS,Chinnaiyan AM et al.: Quantitative proteomic profiling ofprostate cancer reveals a role for miR-128 in prostate cancer.Mol Cell Proteomics 2010, 9:298-312.
20.�
Nik-Zainal S, Alexandrov LB, Wedge DC, Van Loo P,Greenman CD, Raine K, Jones D, Hinton J, Marshall J,Stebbings LA et al.: Mutational processes molding thegenomes of 21 breast cancers. Cell 2012, 149:979-993.
A catalog of somatic mutation from 21 breast cancers was generated andmutational signatures of the underlying processes of oncogenesis weredetermined using mathematical methods.
21. Hammerman PS, Hayes DN, Wilkerson MD, Schultz N, Bose R,Chu A, Collisson EA, Cope L, Creighton CJ, Getz G et al.:Comprehensive genomic characterization of squamous celllung cancers. Nature 2012, 489:519-525.
22. Imielinski M, Berger AH, Hammerman PS, Hernandez B, Pugh TJ,Hodis E, Cho J, Suh J, Capelletti M, Sivachenko A et al.: Mappingthe hallmarks of lung adenocarcinoma with massively parallelsequencing. Cell 2012, 150:1107-1120.
23.�
Govindan R, Ding L, Griffith M, Subramanian J, Dees ND,Kanchi KL, Maher CA, Fulton R, Fulton L, Wallis J et al.: Genomiclandscape of non-small cell lung cancer in smokers and never-smokers. Cell 2012, 150:1121-1134.
Current Opinion in Biotechnology 2013, 24:1129–1136

1136 Pharmaceutical biotechnology
This report identifies 10-fold increased mutational frequency in smokersversus never smokers. Their deep digital sequencing revealed variousnovel alterations in genes involved in chromatin.
24. Aldridge BB, Fernandez-Suarez M, Heller D, Ambravaneswaran V,Irimia D, Toner M, Fortune SM: Asymmetry and aging ofmycobacterial cells lead to variable growth and antibioticsusceptibility. Science 2012, 335:100-104.
25. Pine SR, Ryan BM, Varticovski L, Robles AI, Harris CC:Microenvironmental modulation of asymmetric cell division inhuman lung cancer cells. Proc Natl Acad Sci U S A 2010,107:2195-2200.
26.�
de Carvalho LP, Fischer SM, Marrero J, Nathan C, Ehrt S, Rhee KY:Metabolomics of Mycobacterium tuberculosis revealscompartmentalized co-catabolism of carbon substrates.Chem Biol 2010, 17:1122-1131.
This report revealed that mycobacteria can utilize multiple carbonsources simultaneously for enhanced monophasic growth by cataboliz-ing different sources through the glycolytic, pentose phosphate, and/ortricarboxylic acid pathways.
27. Cole ST, Brosch R, Parkhill J, Garnier T, Churcher C, Harris D,Gordon SV, Eiglmeier K, Gas S, Barry CE 3rd et al.: Decipheringthe biology of Mycobacterium tuberculosis from the completegenome sequence. Nature 1998, 393:537-544.
28. Trivedi OA, Arora P, Sridharan V, Tickoo R, Mohanty D,Gokhale RS: Enzymic activation and transfer of fatty acids asacyl-adenylates in mycobacteria. Nature 2004, 428:441-445.
29. Goyal A, Verma P, Anandhakrishnan M, Gokhale RS,Sankaranarayanan R: Molecular basis of the functionaldivergence of fatty acyl-AMP ligase biosynthetic enzymes ofMycobacterium tuberculosis. J Mol Biol 2012, 416:221-238.
30. Zhang Z, Zhou R, Sauder JM, Tonge PJ, Burley SK,Swaminathan S: Structural and functional studies of fatty acyladenylate ligases from E. coli and L. pneumophila. J Mol Biol2011, 406:313-324.
31. Rohde KH, Veiga DF, Caldwell S, Balazsi G, Russell DG: Linkingthe transcriptional profiles and the physiological states ofMycobacterium tuberculosis during an extended intracellularinfection. PLoS Pathog 2012, 8:e1002769.
32. Voskuil MI, Visconti KC, Schoolnik GK: Mycobacteriumtuberculosis gene expression during adaptation to stationaryphase and low-oxygen dormancy. Tuberculosis (Edinb) 2004,84:218-227.
33. Karakousis PC, Williams EP, Bishai WR: Altered expression ofisoniazid-regulated genes in drug-treated dormantMycobacterium tuberculosis. J Antimicrob Chemother 2008,61:323-331.
34. Daniel J, Maamar H, Deb C, Sirakova TD, Kolattukudy PE:Mycobacterium tuberculosis uses host triacylglycerol toaccumulate lipid droplets and acquires a dormancy-likephenotype in lipid-loaded macrophages. PLoS Pathog 2011,7:e1002093.
35. Griffin JE, Pandey AK, Gilmore SA, Mizrahi V, McKinney JD,Bertozzi CR, Sassetti CM: Cholesterol catabolism byMycobacterium tuberculosis requires transcriptional andmetabolic adaptations. Chem Biol 2012, 19:218-227.
36. Rafidinarivo E, Laneelle MA, Montrozier H, Valero-Guillen P,Astola J, Luquin M, Prome JC, Daffe M: Trafficking pathways ofmycolic acids: structures, origin, mechanism of formation,and storage form of mycobacteric acids. J Lipid Res 2009,50:477-490.
Current Opinion in Biotechnology 2013, 24:1129–1136
37.��
Arora P, Goyal A, Natarajan VT, Rajakumara E, Verma P, Gupta R,Yousuf M, Trivedi OA, Mohanty D, Tyagi A et al.: Mechanistic andfunctional insights into fatty acid activation in Mycobacteriumtuberculosis. Nat Chem Biol 2009, 5:166-173.
The report demonstrates the structural basis for enzymatic difference inactivation of fatty acids by FAAL and FACL. Targeting of FAALs byinhibitors can lead to blockade of multiple biosynthetic pathways.
38. Zhao Y, Liu H, Riker AI, Fodstad O, Ledoux SP, Wilson GL, Tan M:Emerging metabolic targets in cancer therapy. Front Biosci2011, 16:1844-1860.
39. Ferreira LM, Hebrant A, Dumont JE: Metabolic reprogrammingof the tumor. Oncogene 2012, 31:3999-4011.
40. Gordan JD, Thompson CB, Simon MC: HIF and c-Myc: siblingrivals for control of cancer cell metabolism and proliferation.Cancer Cell 2007, 12:108-113.
41. Schulz TJ, Thierbach R, Voigt A, Drewes G, Mietzner B,Steinberg P, Pfeiffer AF, Ristow M: Induction of oxidativemetabolism by mitochondrial frataxin inhibits cancer growth:Otto Warburg revisited. J Biol Chem 2006, 281:977-981.
42.��
Anastasiou D, Yu Y, Israelsen WJ, Jiang JK, Boxer MB, Hong BS,Tempel W, Dimov S, Shen M, Jha A et al.: Pyruvate kinase M2activators promote tetramer formation and suppresstumorigenesis. Nat Chem Biol 2012, 8:839-847.
Cancer cells have modified metabolic networks with altered glucosemetabolism. The switch of Pyruvate kinase from PKM2 to PKM1 isoformmay lead to repressed tumor growth and subsequently decreased chan-neling of glycolytic substrates for anabolic pathways of the cell to meetthe cancer cell’s requirements.
43. Yuneva MO, Fan TW, Allen TD, Higashi RM, Ferraris DV,Tsukamoto T, Mates JM, Alonso FJ, Wang C, Seo Y et al.: Themetabolic profile of tumors depends on both the responsiblegenetic lesion and tissue type. Cell Metab 2012, 15:157-170.
44. Knight ZA, Shokat KM: Features of selective kinase inhibitors.Chem Biol 2005, 12:621-637.
45. Sebolt-Leopold JS, English JM: Mechanisms of drug inhibitionof signalling molecules. Nature 2006, 441:457-462.
46.��
Apsel B, Blair JA, Gonzalez B, Nazif TM, Feldman ME,Aizenstein B, Hoffman R, Williams RL, Shokat KM, Knight ZA:Targeted polypharmacology: discovery of dual inhibitors oftyrosine and phosphoinositide kinases. Nat Chem Biol 2008,4:691-699.
In this paper, a multitarget kinase inhibitor screening identified PP121,which possesses dual specificity to tyrosine and phosphotidylinositolkinases.
47. Willingham SB, Volkmer JP, Gentles AJ, Sahoo D, Dalerba P,Mitra SS, Wang J, Contreras-Trujillo H, Martin R, Cohen JD et al.:The CD47-signal regulatory protein alpha (SIRPa) interactionis a therapeutic target for human solid tumors. Proc Natl AcadSci U S A 2012, 109:6662-6667.
48. Akin D, Sturgis J, Ragheb K, Sherman D, Burkholder K,Robinson JP, Bhunia AK, Mohammed S, Bashir R: Bacteria-mediated delivery of nanoparticles and cargo into cells. NatNanotechnol 2007, 2:441-449.
49. Mashelkar R: India’s ‘‘science for all’’ academy. Science 2012,335:891.
50. Vashisht R, Mondal AK, Jain A, Shah A, Vishnoi P, Priyadarshini P,Bhattacharyya K, Rohira H, Bhat AG, Passi A et al.: Crowdsourcing a new paradigm for interactome driven drug targetidentification in Mycobacterium tuberculosis. PLoS One2012:e39808.
www.sciencedirect.com




















