Embed Size (px)
Citation preview

II
UBERSICH TEN
o o
Neurobiologische Mechanismen der Ubertragung von Muskelschmerz* S . Mense lnstitut fiir Anatomic und Zellbiologie der Universitfit Heidelberg
Zusammenfassung. Im Gegensatz zu Hautschmerz wird Muskelschmerz oft in K6rperregionen weit entfernt von der Lfision fibertragen. Ein Beispiel sind Triggerpunkte in den Halsmuskeln, die fibertragene Schmerzen im Kopf ausl6sen k6nnen. Die Konvergenz-Projektions- Theorie von Ruch ist noch immer das zentrale Konzept Rir die Erkl/irung der Schmerziibertragung. Die Grund- lage der Theorie ist der konvergente Antrieb eines Hin- terhornneurons von zwei unterschiedlichen Versor- gungsgebieten; diese Situation ffihrt dazu, dab Neurone im Thalamus den Ursprung der Aktivierung nicht mehr 10kalisieren k6nnen. Im Grunde genommen ist die Ober- tragung von Schmerzen nichts anderes als eine Fehlloka- lisation der Schmerzquelle. Die Konvergenz-Projek- ti0ns-Theorie kann einige Aspekte der Schmerzfibertra- gung bei Patienten nicht erklfiren; daher wird in der v0rliegenden Arbeit ein weiterer Mechanismus vorge- stellt, der in akuten Umschaltvorg/ingen in Hinterhorn- neuronen unter dem Einflul3 eines Muskelschmerzes be- steht. Ergebnisse aus Tierexperimenten deuten darauf hin, dab Hinterhornneurone ineffektive synaptische Ver- bindungen mit der K6rperperipherie besitzen, die bei Einwirkung von Schmerzreizen durchgeschaltet werden und zu einer Fehllokalisation von Schmerzreizen ffihren. Wahrscheinlich ist das Neuropeptid Substanz P an den Verfinderungen der Hinterhornverschaltung bei Muskel- schmerz und Obertragung beteiligt.
Schlfisselsw6rter: Obertragung - Muskelschmerz - Kon- vergenz Projektions-Theorie - Hinterhornneuron - Fehllokalisation - Substanz P
Neurobiological mechanisms of muscle pain referral
Abstract. In contrast to pain from the skin, muscle pain is often referred to regions remote from the lesion. For
* Herrn Professor Dr. M. Zimmermann zum 60. Geburtstag ge- widmet.
instance, trigger points in neck muscles can elicit pain in the head. The convergence-projection theory of Ruch is still the central concept for the explanation of pain referral. The basis of the theory is that a dorsal horn neuron has convergent input from two different body regions. Because of the convergence, thalamic neurons cannot localize the origin of the dorsal horn activation. Basically, the referral of pain is a mislocalization of pain. Some aspects of muscle pain referral in patients cannot be explained by the convergence-projection theory. Therefore, the present paper presents another mecha- nism, which consists in acute changes in dorsal horn synaptic connections following noeiceptive input from muscle. Results from animal experiments indicate that dorsal horn neurons possess ineffective synaptic connec- tions with the body periphery, which become effective under the influence of a painful stimulus and lead to a mislocalization of pain. The neuropeptide substance P is probably involved in the changes in functional orga- nization that occur in the dorsal horn during muscle pain and its referral.
Key words: Referral - Muscle pain - Convergence-pro- jection theory - Dorsal horn neuron - Mislocalization - Substance P
Der Schmerz (1993) 7:241-249 �9 Springer-Verlag 1993
Schmerziibertragung als klinisches Symptom
Im Gegensatz zu Hautschmerz, der genau lokalisiert werden kann, ist Muskelschmerz diffus und schwierig zu lokalisieren, darfiber hinaus wird er oft in K6rperre- gionen weit entfernt vonder Muskell/ision iibertragen [13, 18, 19, 33]. Bei einigen Patienten ist nur der fibertra- gene Schmerz vorhanden; in diesen F/illen mul3 der Arzt nach der L/ision suchen, die die/ibertragenen Schmerzen verursacht. Ob die Fehllokalisation von Schmerzen in

242 Obersichten
eine K6rperregion auBerhalb der Lfision (Ubertragung im engeren Sinne) und die Ausbreitung eines schmerzen- den Gebietes (Ausstrahlung yon Schmerzen) unter- schiedliche Vorg/inge oder zwei Aspekte desselben Phil- nomens darstellen, ist derzeit eine offene Frage.
Da Muskelschmerzen h/iufig ein typisches und konstan- tes Obertragungsmuster zeigen, k6nnen diese Muster dazu verwendet werden, um den Muskel zu lokalisieren, der die Schmerzquelle darstellt. Dies gilt besonders ffir Schmerzen, die durch sogenannte Triggerpunkte verur- sacht werden. Triggerpunkte bestehen aus palpablen und druckempfindlichen Verh/irtungen innerhalb eines kon- trahierten Bandes yon Muskelfasern [35-37]. Ein Bei- spiel sind Triggerpunkte in den Halsmuskeln, die oft iibertragene Schmerzen im Kopf ausl6sen; diese Schmer- zen werden hfiufig als Kopfschmerzen fehldiagnostiziert.
Eine bemerkenswerte Eigenschaft des Muskelschmerzes besteht darin, dab eine Obertragung auch aul3erhalb der Grenzen eines Myotoms stattfinden kann [18], obwohl die Konstanz der Ubertragungsmuster daffir spricht, dab sie definierten anatomischen Wegen folgt [35]. Die eleganten Experimente yon Bogduk (1980) haben aller- dings gezeigt, dab die Obertragung yon Tiefenschmerz aus dem Versorgungsgebiet der Hinterwurzel in das der Vorderwurzel, d.h. innerhalb desselben Segments, be- sonders effektiv ist.
M6gliche Mechanismen der Schmerziibertragung
Verzweigung yon primdr afferenten Fasern
In Tierexperimenten, in denen die Impulsaktivit/it von einzelnen afferenten Fasern des Muskelnerven abgeleitet wird, findet man hfiufig nozizeptive Afferenzen, die 2 separate rezeptive Felder (RFs) haben, d.h. diese Fasern k6nnen von 2 getrennten Gebieten des Muskels aus akti- viert werden [25]. Es wurden auch prim/Jr afferente Fa- sern gefunden, die ein RF in den tiefen somatischen Ge- weben hatten (Muskel, Gelenk, Periost) und ein weiteres in der Haut distal zum tiefen RF [26]. Die anatomische Grundlage ffir diesen Befund sind wahrscheinlich Ver- zweigungen der afferenten Faser nahe ihres Zielgebietes. Fiir die folgenden Oberlegungen mfissen zwei Arten von Verzweigungen unterschieden werden: I. Kurze Endver- zweigungen, wie sic jede freie Nervenendigung in ihrem Versorgungsgebiet hat. 2. Verzweigungen mit langen )ksten, die zu unterschiedlichen Geweben oder separaten RFs ziehen. Fiir ()bertragungsph/inomene sind nur Ver- zweigungen der 2. Art von Bedeutung.
Die Sinnesempfindungen, die dutch Aktivitfit in einer Faser mit einem RF in der Haut und einem weiteren in den tiefen Geweben ausgel6st werden, hfingen wahr- scheinlich von den zentralnerv6sen Verbindungen dieser Faser ab. Hat die Faser Verbindungen mit Hinterhorn- neuronen, die Schmerzempfindungen yon der Haut ver- mitteln, so werden wahrscheinlich Schmerzen im Gebiet des kutanen RF empfunden, auch wenn das tiefe RF
gereizt wird. Durch diesen Mechanismus der Fehllokali- sation k6nnte es theoretisch zu einer Ubertragung des Tiefenschmerzes in die Haut kommen.
Diese Erkl/irung trifft jedoch nicht auf Muskelschmerz zu, denn Schmerz von Muskeln und anderen tiefen s0- matischen Geweben wird nicht in die Haut fibertragen, sondern in andere tiefe Gewebe [13, 21]. Die alternative Annahme, dab die Faser Verbindungen zu zentralnerv6- sen Neuronen hat, die Tiefenschmerz vermitteln, wird durch die t/igliche Erfahrung nicht gestfitzt, denn dann mfil3te iibertragener Tiefenschmerz durch Reizung des kutanen RF ausgel6st werden k6nnen. (Natiirlich ist es auch m6glich, dab diese afferenten Fasern mit RFs in unterschiedlichen Geweben fiberhaupt keine subjektiven Empfindungen ausl6sen).
Sinclair et al. [33] haben die Hypothese aufgestellt, dab Aktionspotentiale, die in einem Ast einer Nervenfaser mit langen ~sten entstehen, antidrom in andere Aste derselben Faser einlaufen und hier sog. ,,irritierende" Substanzen fiber den Axonreflex freisetzen. Es wurde weiterhin postuliert, dab die irritierenden Substanzen Nozizeptoren in der Nachbarschaft der antidrom be- einflu[3ten .~ste sensibilisieren. Im Licht neuerer Ergeb- nisse k6nnten Neuropeptide wie Substanz P (SP) und Calcitonin Gen-verwandtes Peptid (CGRP) die Rolle der irritierenden Substanzen spielen. Beide Peptide kommen in freien Nervenendigungen der Haut und des Muskels vor und k6nnen durch Impulsaktivit/it und andere Ein- fliisse freigesetzt werden. Substanz P ist daffir bekannt, dab sic Bradykinin und Serotonin aus Blutbestandteilen freisetzt und Histamin und Prostaglandine aus anderen Gewebszellen. Die meisten dieser Substanzen haben eine sensibilisierende Wirkung auf Nozizeptoren.
Wenn der tiefe Ast einer verzweigten Faser durch einen Schadreiz aktiviert wird, k6nnte die resultierende Sensi- bilisierung der Nozizeptoren in der Nachbarschaft des kutanen Astes eine Hyperalgesie der Haut ausl6sen, die oft den Muskelschmerz begleitet [33]. In diesem Modell geht man davon aus, dab die kutane Hyperalgesie v0n anderen Hautafferenzen vermittelt wird, die die Region versorgen, in der der oberflfichliche Ast der verzweigten Afferenz endet. Es gibt jedoch Berichte, dab Muskel. schmerz aueh mit einer kutanen Hypoalgesie verbunden sein kann [5]; diese Situation wird durch den Axonreflex nicht erkl/irt. Der Axonreflex kann auch nicht als Erklfi- rung ffir solche F/ille herangezogen werden, bei denen eine Lokalan/isthesie des fJbertragungsgebietes die fiber- tragenen Schmerzen nicht beseitigt, oder bei denen eine fJbertragung in eine v611ig denervierte K6rperregion er- folgt [9]. Anatomische und neurophysiologische Unter- suchungen [8, 29] haben gezeigt, dab afferente Fasern mit (langen) Verzweigungen zu unterschiedlichen Ge- webstypen relativ selten sind (wenige % aller afferenten Fasern). Falls diese Fasern tats/ichlich an der IJbertra. gung des Tiefenschmerzes oder der kutanen Hyperalge- sie beteiligt sind, mfiBte eine extrem kleine Zahl v0n Afferenzen diese Symptome verursachen. Es mug daher geschlossen werden, dab die Funktion der verzweigten Afferenzen mit langen Asten fiir die Ubertragung und

(Jbersichten 243
die Begleitph~inomene des Muskelschmerzes fraglich ist. Dagegen ist die Bedeutung der kurzen Endverzweigun- gen einer prim~ir afferenten Faser f/fr den lokalen Axon- reflex (der bei Verletzung zu Schwellung, R6tung und letztendlich zu neurogener Entz/fndung f/ihrt) gut gesi- chert [16, 20].
Eine m6gliche Funktion der nozizeptiven afferenten Fa- sere mit multiplen Endigungen in den tiefen Geweben k6nnte darin bestehen, zur diffusen Natur des Muskel- schmerzes beizutragen, da multiple RFs die r/iumliche Aufl6sung des nozizeptiven Systems vermindern.
Konvergenz auf Ri~ckenmarkebene
Die Konvergenz-Projektions-Theorie von Ruch [30] ist immer noch das zentrale Konzept f/fr die Erkl~irung von fibertragenen Schmerzen. In dieser Hypothese nimmt man an, dab Obertragung dann stattfindet, wenn affe- rente Fasern von 2 unterschiedlichen Versorgungsgebie- ten (z.B. Eingeweide und Haut) synaptische Kontakte mit demselben Hinterhornneuron haben. Wenn das Neuron zu einem spinalen Trakt geh6rt, der Schmerz in der Haut vermittelt, wird es diese Information auch dann an h6here Zentren melden, wenn es von Afferenzen der Eingeweide aktiviert wird. Wegen der konvergenten Verschaltung des Hinterhornneurons k6nnen h6here Zentren den tats~ichlichen Ursprung der Erregung nicht identifizieren. Nach dieser Theorie ist die Fehllokalisa- ti0n des Eingeweideschmerzes dadurch bedingt, dab af- ferente Fasern aus den Eingeweiden a priori mit funktio- nell und topographisch unpassenden Neuronen verbun- den sind, n~imlich mit solchen, die Hautschmerz aus einer K6rperregion vermitteln, die vonde r viszeralen L/ision weit entfernt ist.
Die Konvergenz-Projektions-Theorie kann einige Aspekte der Schmerz/fbertragung nicht erkl/iren, die sch0n seit langer Zeit bekannt sind: 1. die Intensit~it und das Gebiet des/fbertragenen Schmerzes nimmt mit der St/irke der Schmerzen an der Stelle der urspr/fng- lichen L~ision zu [33, 34]. 2. Obertragung von Schmerzen braucht Zeit, n/imlich Minuten bis Stunden [18]. 3. Ober- tragung erfolgt oft in K6rpergebiete auBerhalb des Seg- ments der urspriinglichen L/ision [7, 18]. Die ersten 2 Punkte lassen vermuten, dab die Schmerz/fbertragung eine dynamische Komponente hat; der 3. Punkt spricht daffir, dab eine einfache Konvergenz von afferenten Fa- sern innerhalb desselben R/fckenmarksegments als Er- kl/irung ffir alle F~ille von Obertragung nicht ausrei- r ist.
Neue Ergebnisse aus Tierexperimenten lassen die Kon- vergenz-Projektions-Theorie in neuem Licht erscheinen. Was das Hauptpostulat der Theorie angeht, so kann kein Zweifel daran bestehen, dab auf R/fckenmarkebene eine extensive Konvergenz von Afferenzen besteht. Dies ist in elektrophysiologischen Experimenten an Hinter- h0rnneuronen der Ratte und Katze gezeigt worden, die k0nvergenten Antrieb von den Eingeweiden und der Haut [6, 10] oder von unterschiedlichen somatischen
HTM deep L TM cutaneous o3OS i , t,,JId,,d 0 t
~~ Touch cut.
ep L./
il, lklh bL, L_. IdldJ, Ih I ,.,I,].ml a6/~ Touch Mod. p Nox. p.
Cut. deep deep
Abb. 1. Peristimulus-Zeit-Histogramm der Antworten eines kon- vergenten Hinterhornneurons mit 2 rezeptiven Feldern (RFs). Ein RF war hochschwellig mechanosensitiv (HTM) und befand sich im M. tibialis ant., das andere war niederschwellig mechanosensitiv (LTM) und lag in der Haut der lateralen Pfote. Die Lage und ungef/ihre Gr6Be der RFs sind als Projektionen auf die Umrisse eines Standardhinterbeines dargestellt. Die Anwendung mecha- nischer Reize ist durch kurze Balken unterhalb des Histogramms markiert. Nox, mod, deep: Sch~idlicher (noxious) oder schwacher (moderate) Druck auf das tiefe Gewebe; Touch, cut. : Ber/ihrung des kutanen RFs mit einem Tuschepinsel (aus [14])
Strukturen (Haut, Muskel, Gelenk) [14, 17, 32, 40] besa- Ben. In diesen Experimenten drfickt sich die Konvergenz darin aus, dab ein gegebenes Hinterhornneuron von 2 oder mehr K6rperregionen aktiviert werden kann, d.h. in der Existenz von multiplen RFs (Abb. 1). Neurone mit einem hochschwellig mechanosensitiven (HTM) RF in tiefen Geweben besaBen oft ein zweites HTM RF in der Haut (Tabelle 1 links). Dieser Befund k6nnte als St/ftze ffir die Hypothese angesehen werden, dab Neu- rone mit konvergentem Antrieb an der Entwicklung der kutanten Hyperalgesie bei Muskelschmerz beteiligt sind. Allerdings miissen daf/fr 2 weitere Annahmen gemacht werden: 1. Das Neuron hat Verbindungen mit zentral- nerv6sen Trakten, die Hautschmerz vermitteln. 2. Von dem tiefen RF geht ein stiindiger Impulseinstrom aus, der das Hinterhornneuron gemiiB der Konvergenz-Fazi- litierungs-Theorie von Mac Kenzie [23] sensibilisiert. Hiiufig jedoch war das 2. (kutane) RF niederschwellig mechanosensitiv (LTM). Dies pal3t nur dann zur obigen Interpretation, wenn angenommen wird, dab dieser Neu- ronentyp kutane Hyperfisthesie anstelle von Hyperalge- sie vermittelt.
Einer der neuen Aspekte der Konvergenz-Projektions- Theorie besteht darin, dab zumindest ein Teil der kon- vergenten Antriebe nicht st/indig funktionstfichtig ist, sondern nur dann durchgeschaltet wird, wenn ein Schmerzreiz auf die Nozizeptoren der Peripherie ein- wirkt. In Tierexperimenten, in denen die Aktivit/it von einzelnen Hinterhornneuronen registriert wird, kann das Durchschalten von neuen Verbindungen daran erkannt werden, dab neue RFs nach schmerzhafter Reizung auf- treten. Ein Beispiel ist in Abb. 2 dargestellt. Ein Hinter- hornneuron der Ratte mit einem HTM RF im M. biceps femoris bildete nach Injektion einer schmerzhaften Dosis von Bradykinin (BKN) in den M. tibialis ant. 2 neue RFs aus, eines am Injektionsort und ein weiteres in den tiefen Geweben des Pfotenr/fckens. 15 min nach dem

244 Obersichten
Tabelle I. Eigenschaften der RFs von Hinterhornneuronen, die Antrieb yon Rezeptoren in den tiefen somatischen Geweben (Muskel, Faszie, Sehne, Gelenk) erhalten
Reaktionsarten
Antrieb von Antrieb von tiefen kutanen Rezeptoren Rezeptoren
Anzahl von Neuronen
IntaktesRfickenmark Kfilteblock des Rfickenmarks
Alle tiefen Muskel Alle tiefen Muskel Gewebe Gewebe
D Zellen LTM 4 3 7 4 HTM 10 0 2 0 LTM, LTM 1 1 - - HTM, LTM - - 2 1
C-D Zellen LTM LTM 12 10 2 2 LTM HTM 4 4 4 4 HTM HTM 7 2 2 0 HTM LTM 10 6 7 4 LTM LTM, HTM - - 1 1 HTM HTM, LTM 3 1 HTM LTM, LTM 1 1 - LTM, LTM LTM 3 3 1 1 HTM, HTM LTM - - 1 1 HTM, HTM HTM - - 2 2 HTM, LTM HTM - - 4 2 HTM, HTM LTM, HTM - - 1 1
52 30 39 24
D Zellen: Neurone mit ausschlieglichem Antrieb yon tiefgelegenen Rezeptoren. C-D Zellen: Neurone mit konvergentem Antrieb yon kutanen und tiefen Rezeptoren. Im linken Teil der Tabelle ist die Mechanosensitivit/it der RFs aufgefiihrt. Kombinationen yon LTM und HTM weisen auf die Existenz multipler RFs fiir ein Einzelneuron hin. Die Tabelle vergleicht zwei experimentelle Bedingungen, n/imlich 1. Tiere mit intaktem Riickenmark, und 2. Tiere mit einem K/ilteblock des Rfickenmarks zur Unterbrechung der deszendieren- den Hemmung (spinal cord cooled). Die Kategorie ,,Muskel" kennzeichnet solche Neurone, die Antrieb yon Muskelrezeptoren besaBen, entweder allein oder kombiniert mit einem Antrieb yon anderen Geweben (aus [14]). L T M , Niederschwellig mechanosensitiv; H T M , hochschwellig mechanosensitiv; RF, rezep- tives Feld
Setzen des Schmerzreizes waren die neuformier ten R F s immer noch v o r h a n d e n ; das urspri ingliche R F wies n u n eine Senkung der Reizschwelle au f u n d reagierte bereits au f schwachen Druck. D a dieses R F keinen K o n t a k t mi t B K N hatte, muB die Schwel lensenkung auf eine Stei- gerung der Erregbarkei t des H i n t e r h o r n n e u r o n s zurfick- gehen (zentrale Sensibil isierung).
Es ist zu beachten, dab die B K N - I n j e k t i o n bewuBt au- Berhalb des urspr i ingl ichen R F s der Zelle gesetzt wurde ; daher wurde das N e u r o n auch nicht durch die In jek t ion erregt. Offenbar f/ihrt ein Schmerzreiz i rgendwo in der K6rperper ipher ie zu ausgedehnten Erregbarkei ts / inde- rungen im Hin te rho rn . Im Fal l des N e u r o n s in Abb. 2 war die Erregbarkei t gesteigert, u n d ursprfinglich inef- fektive synaptische V e r b i n d u n g e n mit der Peripherie wurden effektiv. Das Ergebnis war die A u s b i l d u n g von neuen RFs , die eine hohe mechanische Schwelle ha t t en u n d in tiefen somat ischen Geweben lagen.
Der obige Befund stellt ein kritisches P rob lem fiir Tier- experimente im al lgemeinen dar, weil die vorbere i tenden Opera t ionen des Tieres Schadreize darstellen, die den
A B
5 rain 15 rain
Nox.p. ~ ~" N o x . p . ~ C N~ ~ C deep ~ \ deep
deep ~ " b
~ a
Abb. 2A und B. Verfinderungen in den Eigenschaften der RFs eines Hinterhornneurons nach intramuskul/irer Injektion einer schmerz- haften Dosis von Bradykinin (BKN). A Lage und Gr6Be des ur- spriinglichen RFs (schwarz ausgefiillt) vor der Injektion yon BKN. Das Neuron konnte nur durch schmerzhafte Druckreizung (n0x. p. deep) des proximalen M. biceps femoris aktiviert werden. Der Pfeil zeigt auf die Injektionsstelle im M. tibialis ant. (TA). B (links) 5 min nach der BKN Injektion waren 2 neue RFs vorhanden, die in den tiefen Geweben lagen und eine hohe mechanische Schwelle hatten. B (rechts): 15 min nach der BKN Injektion wies das ur. sprfingliche RF eine gesenkte mechanische Schwelle auf und rea- gierte nun auf schwachen, unsch~idlichen Druck (mod. p. deep) (aus [1 51)

(Jbersichten 245
Grad der Konvergenz auf Hinterhornebene steigern [14]. Wenn ein Neuron mit 2 RFs in einem Experiment zum erstenmal angetroffen wird, muB often bleiben, ob beide rezeptiven Felder auch im intakten Tier vorhanden ge- wesen w/iren.
Ein weiterer Faktor, der die Antriebskonvergenz be- einflugt, ist das deszendierende antinozizeptive System (vgl. [2]). Eine Unterbrechung der deszendierenden Bah- hen durch K/ihlung des Rfickenmarks rostral der Ableit- stelle ffihrt zu einem Anstieg der Zahl der RFs in Hinter- h0rnneuronen (rechte H/ilfte der Tabelle 1) [14]. Diese Befunde demonstrieren, dal3 viele synaptische Verbin- dungen des Hinterhorns in ihrer Effektivit/it nicht kon- stant find, sondern dutch verschiedene Einfltisse ver/in- dert werden k6nnen.
In dem Neuron in Abb. 2 ben6tigte die Bildung der neuen RFs weniger als 5 rain; in anderen Experimenten war die Latenz zwischen Setzen des Schmerzreizes und Auftauchen yon neuen RFs 1/inger als 15 min. Die neu- f0rmierten RFs blieben I/inger bestehen als der Schmerz- reiz andauerte. Eine m6gliche Interpretation dieser Er- gebnisse ist, dab die Bildung yon neuen RFs auf neuro- plastischen Ver/inderungen in den Verbindungen der Uinterhornneurone beruht (vgl. auch [15]).
Hat die Bildung yon neuen RFs in Tierexperimenten elwas mit der Ubertragung yon Muskelschmerz bei Pa- tienten zu tun ? Eine etwas spekulative Antwort auf diese Frage ist folgende: Unter der Annahme, dab die Entla- dungen eines nozizeptiven Neurons auch Informationen fiber den Oft der L/ision enthalten, kann das Auftauchen eines neuen RFs im M. tibialis ant. distal vom urspriing- lichen RF im M. biceps femoris (Abb. 2) als Ausdruck einer Schmerziibertragung nach proximal gewertet wer- den: Das Neuron meldet im urspr/inglichen Zustand Schmerzen im M. biceps femoris; es wird diese Informa- tion auch dann an h6here Zentren senden, wenn nach Bildung der neuen RFs ein Schmerzreiz auf den M. tibia- lis ant. einwirkt. Wenn weiterhin angenommen wird, dai3 Aktivit/it in nozizeptiven Neuronen Schmerzen ausl6st, so k6nnte die obige Situation zu einer fJbertragung des Muskelschmerzes vom M. tibialis ant. in den M. biceps fem0ris f/ihren. Bei einer L/ision des M. tibialis ant. wer- den lokale Schmerzen in diesem Gebiet auftreten (ver- mittelt durch nozizeptive Neurone mit RFs im M. tibia- lis ant.) und/ibertragene Schmerzen im M. biceps femo- ks (vermittelt von Neuronen, die Schmerzen aus diesem Muskel vermitteln und neue RFs im M. tibialis ant. aus- bilden).
Nach dieser Interpretation k6nnte die Senkung der Reiz- ~hwelle des urspriinglichen RFs zu fibertragener Druck- empfindlichkeit (referred tenderness) im M. biceps femo- ris f/ihren: Nach schmerzhafter Reizung des M. tibialis ant. und erfolgter Ver/inderung der RFs wird eine ~hwache Druckreizung des M. biceps femoris Schmer- ~n in diesem Muskel ausl6sen, da das hiergelegene RF nun eine niedrige Schwelle hat, das Neuron aber immer n0ch Verbindungen mit schmerzvermittelnden zentral- nerv6sen Trakten hat.
Diese Vorg/inge k6nnen als dynamische Komponente der Konvergenz-Theorie angesehen werden; sie haben einen Zeitverlauf, der dem von tibertragenen Schmerzen bei Patienten /ihnelt. In den Tierexperimenten lag die grol3e Mehrzahl der neuformierten RFs distal von den urspr/inglichen RFs. Im Sinne der obigen Interpretation k6nnte diese Anordnung eine Erkl/irung f/Jr eine fJber- tragung nach proximal liefern. Bei Patienten und Ver- suchspersonen wird Muskelschmerz oft nach distal fiber- tragen [19, 36]; jedoch wird auch proximale fJbertra- gung beobaehtet [34].
In Tierexperimenten kann ein anhaltender nozizeptiver Antrieb dutch Muskelafferenzen dadurch erzeugt wer- den, dab eine experimentelle Myositis durch Injektion von Carageenan in den Muskel ausgel6st wird. Dieses Vorgehen f/ihrt innerhalb von 2 h zu einer histologisch erkennbaren Entz/indung und zu einer gesteigerten Akti- vit/it in Nozizeptoren des entz/indeten Muskels [3]. Die systematische Kartographierung yon Hinterhornneuro- nen, die auf elektrische Reizung peripherer Nerven rea- gieren, hat gezeigt, dab im Verlauf einer solchen experi- mentellen Myositis die Neuronenpopulation an Gr613e zunimmt, die durch Afferenzen von dem entz/indeten Muskel aktiviert werden kann. Mit anderen Worten, der effektive Einflul3bereich eines Muskelnerven im Riicken- mark dehnt sich aus, wenn der Muskel entziindet ist [4]. Die Ausdehnung erfolgt vorwiegend in das laterale Hinterhorn und in das Segment rostral des" ursprfing- lichen Einflul3bereichs. Die Ausdehnung in das rostral benachbarte Segment zeigt, dab Ver/inderungen in den Verbindungen yon Hinterhornneuronen nicht auf das Segment der L/ision beschr/inkt sind. So erhielten Neu- rone im lateralen Teil des Segments L3 neue mechano- sensitive RFs im M. gastrocnemius-soleus, wenn in die- sem ein Entzfindungsreiz gesetzt wurde. Bei Tieren mit intaktem Muskel fehlte ein solcher Antrieb in L3 v611ig; der Haupteinflul3bereich des Gastrocnemius-Soleus- Muskelnerven war das mediale Hinterhorn in den Seg- menten L4 und L5.
Da im Hinterhorn eine Somatotopie fiir Neurone mit nozizeptivem Antrieb von tiefen Geweben besteht (je la- teraler die Lage des Neurons im Hinterhorn, desto proxi- maler die Lage des RFs) [41], wiirde eine laterale Aus- dehnung des Einflul3bereichs eines distalen Muskelnervs bedeuten, dab Neurone im lateralen Hinterhorn, die ur- spriinglich Antrieb vom proximalen Bein haben, nun ei- nen zusiitzlichen Antrieb von der distalen Extremit/it er- halten. Unter diesen Bedingungen k6nnte eine distale L/ision fibertragene Schmerzen in der proximalen Extre- mit/it ausl6sen. Die beobachtete Ausbreitung der Erreg- barkeitssteigerung in benachbarte Neuronenpopulatio- nen des Hinterhorns k6nnte die subjektive Empfindung der Schmerzausbreitung oder -ausstrahlung erzeugen. In den Myositis-Experimenten dehnten sich die Ver/inde- rungen der neuronalen Erregbarkeit auch in das rostral benachbarte Segment aus. Dieser Befund k6nnte ffir die klinische Beobachtung von Bedeutung sein, dal3 Muskel- schmerz oft in K6rperregionen aul3erhalb des Segments der ursprfinglichen L/ision iibertragen wird, oder sich nach dort ausbreitet [36].
246 Obersichten
Umschaltungsvorgiinge im Hinterhorn
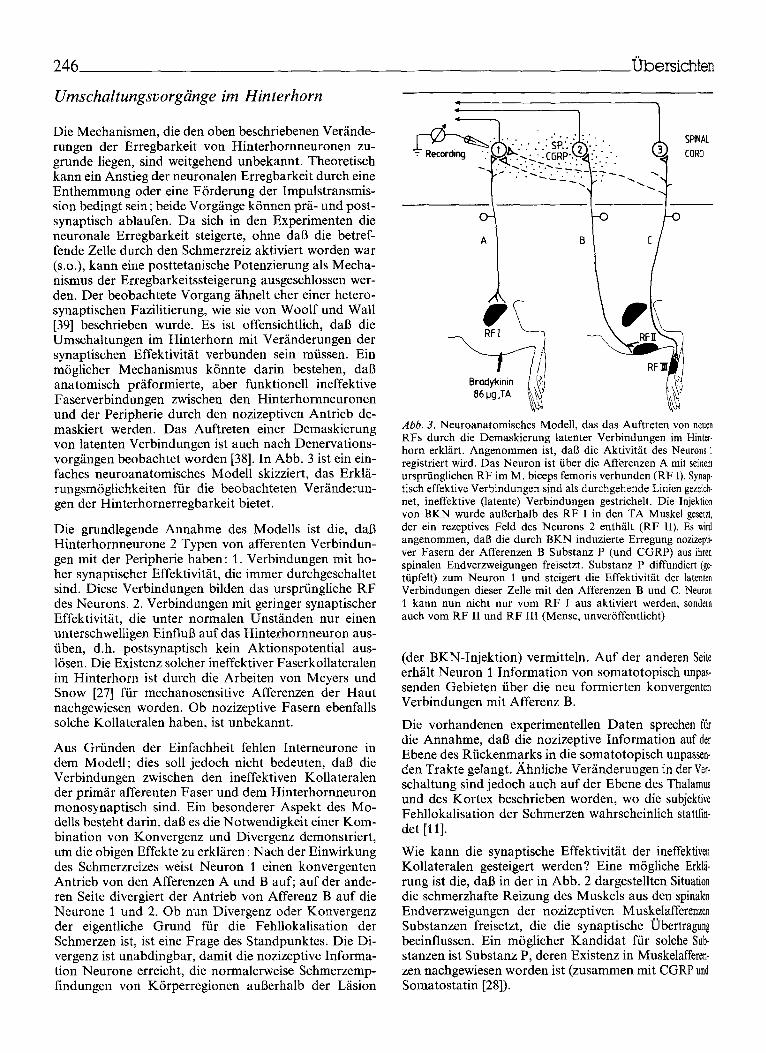
Die Mechanismen, die den oben beschriebenen Ver/inde- rungen der Erregbarkeit von Hinterhomneuronen zu- grunde liegen, sind weitgehend unbekannt. Theoretisch kann ein Anstieg der neuronalen Erregbarkeit durch eine Enthemmung oder eine F6rderung der Impulstransmis- sion bedingt sein; beide Vorg/inge k6nnen pr~i- und post- synaptisch ablaufen. Da sich in den Experimenten die neuronale Erregbarkeit steigerte, ohne dab die betref- fende Zelle dutch den Schmerzreiz aktiviert worden war (s.o.), kann eine posttetanische Potenzierung als Mecha- nismus der Erregbarkeitssteigerung ausgeschlossen wer- den. Der beobachtete Vorgang/ihnelt eher einer hetero- synaptischen Fazilitierung, wie sie von Woolf und Wall [39] beschrieben wurde. Es ist offensichtlich, dab die Umschaltungen im Hinterhorn mit VerS, nderungen der synaptischen Effektivit~it verbunden sein mfissen. Ein m6glicher Mechanismus k6nnte darin bestehen, dab anatomisch pr/iformierte, abet funktionell ineffektive Faserverbindungen zwischen den Hinterhornneuronen und der Peripherie durch den nozizeptiven Antrieb de- maskiert werden. Das Auftreten einer Demaskierung von latenten Verbindungen ist auch nach Denervations- vorg/ingen beobachtet worden [38]. In Abb. 3 ist ein ein- faches neuroanatomisches Modell skizziert, das Erkl/i- rungsm6glichkeiten fiir die beobachteten Ver~inderun- gender Hinterhornerregbarkeit bietet.
Die grundlegende Annahme des Modells ist die, dab Hinterhornneurone 2 Typen von afferenten Verbindun- gen mit der Peripherie haben: 1. Verbindungen mit ho- her synaptischer Effektivit/it, die immer durchgeschaltet sind. Diese Verbindungen bilden das urspriingliche RF des Neurons. 2. Verbindungen mit geringer synaptischer Effektivit/it, die unter normalen Unst/inden nur einen unterschwelligen EinfluB auf das Hinterhornneuron aus- fiben, d.h. postsynaptisch kein Aktionspotential aus- 16sen. Die Existenz solcher ineffektiver Faserkollateralen im Hinterhorn ist durch die Arbeiten von Meyers und Snow [27] fiir mechanosensitive Afferenzen der Haut nachgewiesen worden. Ob nozizeptive Fasern ebenfalls solche Kollateralen haben, ist unbekannt.
Aus Grfinden der Einfachheit fehlen Interneurone in dem Modell; dies soll jedoch nicht bedeuten, dab die Verbindungen zwischen den ineffektiven Kollateralen der prim/ir afferenten Faser und dem Hinterhornneuron monosynaptisch sind. Ein besonderer Aspekt des Mo- dells besteht darin, dab es die Notwendigkeit einer Kom- bination von Konvergenz und Divergenz demonstriert, um die obigen Effekte zu erkl/iren: Nach der Einwirkung des Schmerzreizes weist Neuron 1 einen konvergenten Antrieb von den Afferenzen A und B auf; auf der ande- ren Seite divergiert der Antrieb von Afferenz B auf die Neurone 1 und 2. Ob nun Divergenz oder Konvergenz der eigentliche Grund fiir die Fehllokalisation der Schmerzen ist, ist eine Frage des Standpunktes. Die Di- vergenz ist unabdingbar, damit die nozizeptive Informa- tion Neurone erreicht, die normalerweise Schmerzemp- findungen yon K6rperregionen auBerhalb der Lfision
- Recording . . - . SRNAL
CORO
A I B ] C
B: dy?:!o l,, t
Abb. 3. Neuroanatomisches Modell, das das Auftreten yon neuen RFs durch die Demaskierung latenter Verbindungen im Hinter- horn erkliirt. Angenommen ist, dab die Aktivitiit des Neurons 1 registriert wird. Das Neuron ist fiber die Afferenzen A mit seinem ursprtinglichen RF im M. biceps femoris verbunden (RF I). Synap- tisch effektive Verbindungen sind als durchgehende Linien gezeich- net, ineffektive (latente) Verbindungen gestrichelt. Die lnjekti0n yon BKN wurde augerhalb des RF I in den TA Muskel geseta, der ein rezeptives Fe|d des Neurons 2 enthglt (RF II). Es wird angenommen, dab die durch BKN induzierte Erregung nozizepti- ver Fasern der Afferenzen B Substanz P (und CGRP) aus ihren spinalen Endverzweigungen freisetzt. Substanz P diffundiert (ge- tfipfelt) zum Neuron 1 und steigert die Effektivitiit der latenten Verbindungen dieser Zelle mit den Afferenzen B und C. Neuron 1 kann nun nicht nur vom RF I aus aktiviert werden, sondern auch vom RF II und RF III (Mense, unver6ffentlicht)
(der BKN-Injektion) vermitteln. Auf der anderen Scitr erh/ilt Neuron 1 Information yon somatotopisch unpas- senden Gebieten fiber die neu formierten konvergenten Verbindungen mit Afferenz B.
Die vorhandenen experimentellen Daten sprechen ffir die Annahme, dab die nozizeptive Information auf der Ebene des Rfickenmarks in die somatotopisch unpassen- den Trakte gelangt. ,~hnliche Veriinderungen in der Ver- schaltung sind jedoch auch auf der Ebene des Thalamus und des Kortex beschrieben worden, wo die subjektive Fehllokalisation der Schmerzen wahrscheinlich stattfin- det [I1].
Wie kann die synaptische Effektivitiit der ineffektiven Kollateralen gesteigert werden? Eine m6gliche Erkl~- rung ist die, dab in der in Abb. 2 dargestellten Situation die schmerzhafte Reizung des Muskels aus den spinalen Endverzweigungen der nozizeptiven Muskelafferenzen Substanzen freisetzt, die die synaptische Obertragung beeinflussen. Ein m6glicher Kandidat ffir solche Sub- stanzen ist Substanz P, deren Existenz in Muskelafferen- zen nachgewiesen worden ist (zusammen mit CGRP und Somatostatin [28]).

Obersichten 247
Tibial n. Common peroneal n. Sural n.
(- 82 mY)
SV sv
SP IO01~M
5rain (-TgmV)
3Stain F92r~)
~ I.O5V 5v
~/,OmV ~ ~ S V
! !
Abb. 4A-C. Intrazellulfire Ableitung der Aktivitfit eines Hinter- h0rnneurons vor und w~ihrend Superfusion des Rfickenmarks mit Substanz P (100 I~M). Die Erregbarkeit der Zelle wurde durch elek- trische Reizung der Nn. tibialis, peroneus comm. und suralis an den durch Pfeile gekennzeichneten Zeitpunkten bestimmt. Der Zeitraum der Superfusion mit Substanz P ist durch den vertikalen Balken markiert; die Zeit vom Beginn der Superfusion ist in Minu- ten angegeben, das Ruhemembranpotential des Neurons in Milli- v01t (mV). A Das Neuron reagierte mit einem Aktionspotential auf die Reizung des N. tibialis mit einer Reizstfirke von 360 mV
(beide Registrierungen wurden bei Reizung mit dem Schwellenwert erhalten), bei Reizung der anderen Nerven zeigte es nur unter- schwellige EPSPs trotz einer hohen Reizstfirke yon 5 V. B, C W~ih- rend der Superfusion mit Substanz P feuerte das Neuron auch ein Aktionspotential bei Reizung des N. peroneus comm. (der Schwellenwert betrug 1,05 V in B und 1,3 V in C), der Antrieb vom N. suralis blieb jedoch weiterhin unterschwellig. Das Mem- branpotential zeigte keine deutlichen Ver~inderungen unter dem Einflul~ von Substanz P (Hoheisel und Mense, unver6ffentlicht)
In hoher Dosierung aktiviert Substanz P nozizeptive Neurone, in niedrigen Konzentrat ionen 16st es eine lang anhaltende Depolarisation der Zelle aus [31, 42]. Die Depolarisation ist die Vorbedingung ffir die Beseitigung des Magnesiumblocks des N M D A (N-methyl-D-Aspar- tat)-gesteuerten Ionenkanals. Nach Beseitigung des Ma- gnesiumblocks 6ffnet sich der Kanal , und Kalziumionen fliel3en in die Zelle, die als second messenger wirken [22]. Auf diese Weise k6nnen viele Funkt ionen des Neurons einschliel31ich seiner Erregbarkeit verfindert werden. Substanz P bindet an Rezeptormolekfile in der Zellmem- bran (haupts~ichlich an den Neurokinin- l -Rezeptor) [24]; die Wirkung des Neuropeptids scheint daher recht spezifisch zu sein.
Der EinfluB von Substanz P auf die synaptische Erreg- barkeit von Hinterhornneuronen kann in Experimenten untersucht werden, in denen die Aktivitfit der Zellen wahrend der Superfusion des Riickenmarks mit einer Substanz-P-L6sung intrazellulfir abgeleitet wird (Abb. 4). Vor Gabe von Substanz P hatte das gezeigte Neuron einen iiberschwelligen erregenden Antrieb aus- schlieBlich vom N. tibialis; N. peroneus comm. und N. suralis 16sten auch bei starker elektrischer Reizung (5 V) nur unterschwellige erregende postsynaptische Poten- tiale (EPSPs) aus. Unter dem Einflul3 von Substanz P wurde der Antrieb vom N. peroneus comm. fiberschwel- lig, d.h. die Effektivitfit des synaptischen Antriebs von
diesem Nerven war gesteigert. Diese Ergebnisse zeigen, dab Substanz-P-Synapsen demaskiert und daher an Ver- finderungen der Hinterhornverschaltung beteiligt sein kann, wie sie wahrscheinlich bei Muskelschmerz und Ober t ragung auftreten.
Im Gegensatz zu Aminosfiuretransmittern diffundiert Substanz P fiber weite Entfernungen im Rfickenmark, nachdem es von den spinalen Endigungen der afferenten Fasern freigesetzt worden ist. Daher wird Substanz P auch als ein Volumentransmit ter angesehen [1], der im Gegensatz zu Aminosfiuretransmittern in der Lage ist, eine grol3e Populat ion von Neuronen in der N~he des Freisetzungsortes zu beeinflussen. Diese Eigenschaft ist wahrscheinlich ffir die Ausl6sung der ausgedehnten Ver- finderungen der Hinterhornerregbarkei t wfihrend einer akuten Myositis wesentlich (s.o.); ebenso k6nnte sie fiir die Uber t ragung des Muskelschmerzes in K6rpergebiete aul3erhalb des Myotoms der L~ision von Bedeutung sein.
Schluflf olgerungen
Verzweigungen des prim/Jr afferenten Neurons scheinen keine zufriedenstellende Erkl~irung fiir die Uber t ragung von Muskelschmerz zu liefern. Die Konvergenz von Af- ferenzen auf der Ebene des Hinterhorns, wie sie von der Konvergenz-Projektions-Theorie vorgeschlagen

248 Uber s i ch ten
wird, ist weiterhin das zentrale Konzept fiir das Ver- st/indnis der Schmerziibertragung. Allerdings ist die Konvergenz-Projektions-Theorie zu statisch und erkl/irt den Zeitverlauf von Ubertragungsph~inomenen bei Pa- tienten nicht. Neuere Ergebnisse aus Tierexperimenten deuten darauf hin, daB schmerzhafte Reize in der Ktr- perperipherie auf Riickenmarkebene zu synaptischen Ver/inderungen ffihren, die an den dynamischen Aspek- ten der Obertragung beteiligt sein ktnnten. Die Freiset- zung von Substanz P von nozizeptiven afferenten Fasern scheint ein wichtiger Faktor zu sein, der zur Demaskie- rung von ursprfinglich ineffektiven Verbindungen im Hinterhorn ffihrt.
Zwei Hauptaspekte sollten der Konvergenz-Projektions- Theorie hinzugeffigt werden, wenn sie auf die Obertra- gung von Muskelschmerz angewendet wird:
1. Viele konvergente Verbindungen von Hinterhornneu- ronen werden wahrscheinlich erst durch die schmerz- hafte Reizung von Skelettmuskeln durchgeschaltet.
2. Die Obertragung von Muskelschmerz in Myotome auBerhalb der L/ision ktnnte durch die Ausbreitung einer zentralen Sensibilisierung in benachbarte spinale Segmente bedingt sein. Beide Prozesse benttigen Zeit und sind vonder Intensitiit der Schmerzen abh/ingig.
Literatur
1. Agnati LF, Fuxe K, Zoli M, Zini L, Toffano G, Ferraguti F (1986) A correlation analysis of the regional distribution of central enkephalin and beta-endorphin immunoreactive termi- nals and of opiate receptors in adult and old male rats. Evidence for the existence of two main types of communication in the central nervous system: the volume transmission and the wiring transmission. Acta Physiol Scand 128:201
2. Basbaum AI, Fields HL (1984) Endogenous pain control sys- tems: Brainstem spinal pathways and endorphin circuitry. Annu Rev Neurosci 7:309
3. Berberich P, Hoheisel U, Mense S (1988) Effects of a carra- geenan-induced myositis on the discharge properties of group III and IV muscle receptors in the cat. J Neurophysiol 59:1395
4. Beylich G, Hoheisel U, Koch K, Mense S (1993) Neuroplastic changes induced in dorsal horn neurones by an experimental myositis and muscle nerve axotomy. Pfliigers Arch 422 [Suppl 1]:R61
5. Bogduk N (1980) Lumbar dorsal ramus syndrome. Med J Austr 2:537
6. Cervero F (1983) Somatic and visceral inputs to the thoracic spinal cord of the cat: effects of noxious stimulation of the biliary system. J Physiol 337 : 51
7. Coderre TJ, Katz J, Vaccarino AL, Melzack R (1993) Contri- bution of central neuroplasticity to pathological pain: review of clinical and experimental evidence. Pain 52:259
8. Devor M, Wall PD, McMahon SB (1984) Dichotomizing so- matic nerve fibers exist in rats but they are rare. Neurosci Lett 49:187
9. Doran FSA, Ratcliffe AH (1954) The physiological mechanism of referred shoulder-tip pain. Brain 77:427
10. Foreman RD, Blair RW, Weber RN (1984) Viscerosomatic convergence onto T2-T4 spinoreticular, spinoreticular-spino- thalamic, and spinothalamic tract neurons in the cat. Exp Neu- rol 85 : 597
11. Guilbaud G (1991) Central neurophysiological processing of joint pain on the basis of studies performed in normal animals
and in models of experimental arthritis. Can J Physiol Pharma- col 69 : 637
12. Herdegen T, T611e TR, Bravo R, Zieglgfinsberger W, Zimmer- mann M (1991) Sequential expression of JUN B, JUN D and FOS B proteins in rat spinal neurons: cascade of transcripti0nal operations during nociception. Neurosci Lett 129:221
13. Hockaday JM, Whitty CWM (1967) Patterns of referred pain in the normal subject. Brain 90:481
14. Hoheisel U, Mense S (1990) Response behaviour of cat dorsal horn neurones receiving input from skeletal muscle and other deep somatic tissues. J Physiol 426 : 265
15. Hoheisel U, Mense S (1993) Appearance of new receptive fields in rat dorsal horn neurones following noxious stimulation of skeletal muscle: a model for referral of muscle pain? Neur0sci Lett 153 : 9
16. Holzer P (1988) Local effector functions of capsaicin-sensitive sonsory nerve endings: involvement of tachykinins, calcit0nin gene-related peptide and other neuropeptides. Neuroscience 24: 739
17. Hu JW, Sessle BJ, Raboisson P, Dallel R, Woda A (1992) Stimulation of craniofacial muscle afferents induces prolonged facilitatory effects in trigeminal nociceptive brainstem neu. tones. Pain 48: 53
18. Inman VT, Saunders JB, de CM (1944) Referred pain from skeletal structures. J Nerv Ment Dis 99:660
19. Kellgren JH (1938) Observations on referred pain arising from muscle. Clin Sci 3 : 175
20. Lembeck F, Holzer P (1979) Substance P as neurogenic media- tor of antidromic vasodilation and neurogenic plasma extrava- sation. Naunyn-Schmiedeberg's Arch Pharmacol 310:175
21. Lewis T (1942) Pain. The Macmillan Company, London 22. MacDermott AB, Mayer ML, Westbrook GL, Smith SJ,
Barker JL (1986) NMDA-receptor activation increases cyto- plasmic calcium concentration in cultured spinal cord neur0nes. Nature 321:519 MacKenzie J (1909) Symptoms and their interpretation. Shaw and Sons, London Maggi CA, Patacchini R, Rovero P, Giachetti A (1993) Ta- chykinin receptors and tachykinin receptor antagonists. J Au- tonom Pharmacol 13 : 23 Mense S, Meyer H (1985) Different types of slowly conducting afferent units in cat skeletal muscle and tendon. J Physi01 363:403 Mense S, Light AR, Perl ER (1981) Spinal terminations of subcutaneous high-threshold mechanoreceptors. In: Brown AG, R&helyi M (eds) Spinal cord sensation. Scottish Academic Press, Edinburgh, p 79 Meyers DER, Snow PJ (1984) Somatotopical inappropriate projections of single hair follicle afferent fibres to the cat spinal cord. J Physiol 347: 59 Molander C, YGGE I, Dalsgaard C-J (1987) Substance P., somatostatin-, and calcitonin gene-related peptide-like imm~- noreactivity and fluoride resistant acid phosphatase-activity in relation to retrogradely labelled cutaneous, muscular and visceral primary sensory neurons in the rat. Neurosci Lett 74:37 Pierau F-K, Fellmer G, Taylor DCM (1984) Somato-visceral convergence in cat dorsal root ganglion neurones demonstrated by double-labelling with fluorescent tracers. Brain Res 321:63 Ruch TC (1949) Visceral sensation and referred pain. In: Ful- ton JF (ed) Howell's textbook of physiology, 16th edn. Saunders, Philadelphia, p 385 Sastry BR (1979) Substance P effects on spinal nociceptive neu- rones. Life Sci 24:2169 Schaible H-G, Schmidt RF, Willis WD (1987) Convergent in- puts from articular, cutaneous and muscle receptors onto as- cending tract cells in the cat spinal cord. Exp Brain Res 66:479 Sinclair DC, Weddell G, Feindel WH (1948) Referred pain and associated phenomena. Brain 71:184 Torebj6rk HE, Ochoa JL, Schady W (1984) Referred pain from intraneural stimulation of muscle fascicles in the median nerve. Pain 18 : 145
23.
24.
25.
26.
27.
28.
29.
30.
31.
32.
33.
34.

Obersichten 249
35. Travell J, Rinzler SH (1952) The myofascial genesis of pain. Postgrad Med 11:425
36. TraveU JG, Simons DG (1983) Myofascial pain and dysfunc- tion. The trigger point manual. Williams and Wilkins, Balti- more London
37. Travell JG, Simons DG (1992) Myofascial pain and dysfunc- tion. The trigger point manual, vol 2. The lower extremities. Williams and Wilkins, Baltimore London
38. Wall PD (1977) The presence of ineffective synapses and cir- cumstances wich unmask them. Phil Trans R Soc Lond [Biol] 278: 361
39. Woolf C J, Wall PD (1986) Relative effectiveness of C primary afferent fibers of different origins in evoking a prolonged facili- tation of the flexor reflex in the rat. J Neurosci 6:1433
40. u X-M, Mense S (1990) Response properties and descending
control of rat dorsal horn neurones with deep receptive fields. Neurosci 39: 823
41. Yu X-M, Mense S (1990) Somatotopical arrangement of rat spinal dorsal horn cells processing input from deep tissues. Neu- rosci Lett 108 : 43
42. Zieglgfinsberger W, Tulloch IF (1979) Effects of substance P on neurones in the dorsal horn of the spinal cord of the cat. Brain Res 166: 273
Prof. Dr. S. Mense Institut f/Jr Anatomie und Zellbiologie der Universit~it Im Neuenheimer Feld 307 D-69120 Heidelberg





























