Embed Size (px)
Citation preview

Neurocircuitry controlling reward-directed behaviour in rats: Contribution of striatal sub-regions and
prelimbic cortex
PhD. Thesis
By
Christine Stubbendorff M.Sc. Department of Neuroscience, Psychology and Behaviour
University of Leicester Leicester
2016

1
Neurocircuitry controlling reward-directed behaviour in rats: Contribution of striatal sub-regions and prelimbic cortex C. Stubbendorff Rodent striatum is involved in sensory-motor transformations and reward-related learning, with lesion studies suggesting functional differences between striatal subregions. Dorsomedial striatum (DMS) is associated with goal-directed behaviour; dorsolateral striatum (DLS) mediates automated stimulus-response and nucleus accumbens (NAc) is involved in reward expectation. Corticostriatal communication from prelimbic cortex (PrL) to DMS and NAc likely modulates appetitive behaviour. The studies reported here investigated how specific elements of reward-related behaviour are maintained by striatum and cortico-striatal interaction. To better understand the functional significance of DLS sensory responses we developed a novel tactile discrimination task in head-fixed rats. Initial results using this task linked DLS sensory responses to either reward-expectation or motor-initiation but could not distinguish between the two. Next, to separate reward and motor components of striatal neural responses and to examine the role of cortico-striatal interaction, we developed a novel discrimination task requiring rats to either respond or suppress responding to reward-predicting cues. Neuronal responses in DLS, DMS, NAc and PrL were recorded during the discrimination task in overtrained rats. In both striatum and PrL, neuronal responses to cue-onset did not appear to be influenced by differences in reward expectation. However, responses in NAc and DMS showed a possible contribution from motor preparatory processes. Overall, striatal and PrL responses as well as synchronisation between striatal sub-regions and between PrL and striatal sub-regions were greater in error trials (false alarms and misses) than correct response trials (hits and correct rejections). Error responses during performance of an overtrained task may signal trials in which the animal tests the consistency of the learned stimulus response contingencies and thus engage striatal networks associated with goal-directed rather than habitual behaviour. The trial type-dependent differences in synchronisation between PrL and all three striatal subregions may indicate modulation from other brain areas or interactions between different cortico-striatal-thalamic circuits.

2
Acknowledgements
I would like to thank David Jones, Tony Smith, Andrew Warren and Gerald Gutteridge in the
Biomedical Joint Workshop, University of Leicester and Rob Hemmings in School of Psychology,
University of Leicester. Without their ingenuity, skill and patience the work presented here could not
have been produced. I would like to thank Rodrigo Q. Quiroga, Systems Neuroscience Group,
University of Leicester for supplying Matlab based spike sorting code, and Manuel Molano, Center
for Neuroscience and Cognitive Systems, Istituto Italiano di Technologia, Rovereto, Italy, for further
adapting this code to our tetrode recordings. I would also like to thank Tracie Payne, (Department of
Neuroscience, Oberlin College, OH, USA) for advice on the Go/NoGo discrimination task. A big thank
you to my supervisors, Todor V. Gerdjikov and Andrew M.J. Young and my fellow colleagues in the
lab; Rachel E. Rickard, Rosie Parry, Daniel Dautan and Aman Asif-Malik for daily advice and support.
Last, but not least, I would like to thank the rats; Elvis, Sherlock, Darwin, Moses, Herbie, Zorro,
Houdini, Pavlov, Freud, Jung, Han Solo, Obi-wan, Odin, Frej, Loke, Mimer, Lucifer, Nixon, Jekyll, Hyde,
Yoda, Gandalf, Lenin and Mao, for good behaviour.

3
List of contents
Chapter 1: The role of striatum and cortico-striatal circuits in reward-directed behaviour ............... 8 1.1 Introduction ............................................................................................................. 8
1.1.1 The role of striatum in reward-directed behaviour ....................................... 9 1.1.2 The role of medial prefrontal cortex in behavioural control ....................... 10 1.1.3 Cortico-striatal circuits ................................................................................. 11 1.1.4 The role of striatum and mPFC in food seeking and its clinical relevance .. 13 1.1.5 Project overview .......................................................................................... 14
Chapter 2: Contribution of Dorsolateral striatum to tactile processing in the awake rat ................. 17 2.1 Introduction ........................................................................................................... 17
2.1.1 Whisker stimulation in head fixed rats ........................................................ 18 2.1.2 Aims ............................................................................................................. 19
2.2 Methods ................................................................................................................. 19 2.2.1 Animals ........................................................................................................ 19 2.2.2 Initial behavioural screening ........................................................................ 20 2.2.3 Construction of electrode microdrives ........................................................ 20 2.2.4 Surgery ......................................................................................................... 21 2.2.5 Behavioural training and testing .................................................................. 22 2.2.6 Electrophysiological recordings ................................................................... 24 2.2.7 Technical challenges with obtaining single unit recordings ........................ 25
2.3 Results .................................................................................................................... 25 2.3.1 Behaviour ..................................................................................................... 25 2.3.2 Electrophysiological recordings ................................................................... 26
2.4 Discussion ............................................................................................................... 28 2.4.1 Conclusion .................................................................................................... 30
Chapter 3: Exploring motor and reward components of striatal responses to reward-paired auditory cues .................................................................................................................... 32 3.1 Introduction ........................................................................................................... 32
3.1.1 The role of striatum in reward-directed behaviour ..................................... 32 3.1.2 Dorsomedial striatum .................................................................................. 33 3.1.3 Dorsolateral striatum ................................................................................... 34 3.1.4 Nucleus Accumbens ..................................................................................... 35 3.1.5 Interaction between striatal sub-regions .................................................... 36 3.1.6 Study aims .................................................................................................... 37 3.1.7 Hypothesis ................................................................................................... 37
3.2 Methods ................................................................................................................. 38 3.2.1 Animals ........................................................................................................ 38 3.2.2 Apparatus ..................................................................................................... 38 3.2.3 Behavioural training ..................................................................................... 38 3.2.4 Tetrode drives .............................................................................................. 41 3.2.5 Surgery ......................................................................................................... 41 3.2.6 Electrophysiological recordings ................................................................... 43 3.2.7 Verification of tetrode placement ............................................................... 43 3.2.8 Statistical analysis ........................................................................................ 45
3.3 Results .................................................................................................................... 47 3.3.1 Behaviour ..................................................................................................... 48 3.3.2 Firing rate responses .................................................................................... 48 3.3.3 Baseline firing rates ..................................................................................... 50 3.3.4 Effect of previous trial response .................................................................. 51

4
3.3.5 Coherence between striatal subregions ...................................................... 53 3.3.6 Differences between tasks ........................................................................... 55
3.4 Discussion ............................................................................................................... 55 3.4.1 Behaviour ..................................................................................................... 55 3.4.2 Baseline single unit activity .......................................................................... 56 3.4.3 Single unit responses to cue onset .............................................................. 57 3.4.4 Recent behavioural experience predicts neuronal response to cue ........... 60 3.4.5 Striatal subregions collectively respond to cue onset ................................. 61 3.4.6 Coherence between striatal subregions ...................................................... 62 3.4.7 Conclusions .................................................................................................. 64
Chapter 4: Corticostriatal contribution to reward-directed behaviour ............................................. 66 4.1 Introduction ........................................................................................................... 66
4.1.1 Prelimbic cortex and behavioural control ................................................... 66 4.1.2 Prelimbic modulation of striatal processes ................................................. 67 4.1.3 Study aims .................................................................................................... 70
4.2 Methods ................................................................................................................. 70 4.2.1 Animals ........................................................................................................ 71 4.2.2 Apparatus ..................................................................................................... 71 4.2.3 Behavioural training ..................................................................................... 71 4.2.4 Surgery ......................................................................................................... 71 4.2.5 Electrophysiological recordings ................................................................... 72 4.2.6 Verification of tetrode placement ............................................................... 73 4.2.7 Statistical analysis ........................................................................................ 73
4.3 Results .................................................................................................................... 74 4.3.1 Behaviour ..................................................................................................... 74 4.3.2 Firing rate response to cue onset ................................................................ 74 4.3.3 Striatal response to cue onset ..................................................................... 75 4.3.4 Prelimbic cortex baseline firing rates .......................................................... 76 4.3.5 Prelimbic cortex response to cue onset ...................................................... 76 4.3.6 Effect of previous trial response .................................................................. 77 4.3.7 Coherence between prelimbic cortex and striatal subregions .................... 78 4.3.8 Differences between tasks ........................................................................... 80
4.4 Discussion ............................................................................................................... 80 4.4.1 Baseline single unit activity .......................................................................... 80 4.4.2 PrL single unit response to cue onset .......................................................... 81 4.4.3 Effect of previous trial response .................................................................. 82 4.4.4 Coherence between PrL and striatum ......................................................... 83 4.4.5 Conclusions .................................................................................................. 85
Chapter 5: Final Discussion ................................................................................................................ 87 5.1.1 Summary of conclusions from experimental chapters ................................ 87 5.1.2 Comparison between findings in experimental chapters ............................ 89 5.1.3 Future perspectives ..................................................................................... 91
Appendix .......................................................................................................................................... 93 References .......................................................................................................................................... 96

5
List of tables
Table 3-1 Coordinates targeted for recording of single unit responses in striatal subregions. .......... 42
Table 3-2 Number of analysed neurons from each structure. ............................................................ 47
Table 4-1 Number of analysed and significantly responding units in PrL ........................................... 74
Table 4-2 Log transformed coherence between PrL and DLS, DMS and NAc after cue onset. ............ 79

6
List of Figures
Figure 1-1 Projection routes in cortico-striatal-thalamic circuits. ...................................................... 11
Figure 2-1 Illustration of rat somatosensory cortex and apparatus used in the experimental setup.17
Figure 2-2 Discrimination task............................................................................................................. 24
Figure 2-3 Rats trained to discriminate between rewardable (Go) and non-rewardable (NoGo)
whisker stimulation licked more during Go vs. NoGo paired stimulation. ........................................... 26
Figure 2-4 DLS cue-evoked responses during discrimination and reversal in one animal. ................. 27
Figure 3-1 Behavioural paradigm ........................................................................................................ 39
Figure 3-2 Response ratios in the Go-NoGo Plus and Go-NoGo Minus task ....................................... 40
Figure 3-3 Tetrode based spike sorting. .............................................................................................. 41
Figure 3-4 Verification of tetrode placement. ...................................................................................... 44
Figure 3-5 Striatal neuron population respond transiently to cue onset. ............................................ 45
Figure 3-6 Example spike rasters and waveforms from neurons in DLS, DMS and NAc in both tasks. 46
Figure 3-7 Behavioural performance. ................................................................................................... 49
Figure 3-8 Log transformed firing rate responses to cue onset. .......................................................... 51
Figure 3-9 Effect of previous trial response on cue-induced firing. ...................................................... 52
Figure 3-10 Log transformed baseline coherence between striatal subregions. ................................. 54
Figure 4-1 Verification of tetrode placement ...................................................................................... 72
Figure 4-2 PrL neuron population respond transiently to cue onset.................................................... 75
Figure 4-3 Log transformed firing rate responses to cue onset in PrL. ................................................ 76
Figure 4-4 Effect of previous trial response on cue-induced firing. ...................................................... 77
Figure 4-5 Log transformed baseline coherence between PrL and DLS, DMS and NAc. ...................... 78

7
List of abbreviations
DLS dorsolateral striatum
DMS dorsomedial striatum
NAc nucleus accumbens
mPFC medial prefrontal cortex
PrL prelimbic cortex
IL infralimbic cortex
CR correct rejection
FA false alarm
5-CSRTT 5 choice serial reaction time task
MSN Medium spiny neuron

8
Chapter 1: The role of striatum and cortico-striatal circuits in
reward-directed behaviour
1.1 Introduction
The basal ganglia are strongly involved in action selection. This includes both its expression through
adaptive motor control and the processes that lead to movement, including the elements that drive
actions, such as emotions, motivation, and cognition (Haber, 2003, Devan et al., 2011). The basal
ganglia consist of dorsolateral striatum (DLS), dorsomedial striatum (DMS) and nucleus accumbens
(NAc), collectively known as the striatum, as well as the globus pallidus, ventral tegmental area,
substantia nigra pars reticulate and subthalamic nucleus (Devan et al., 2011, Balleine et al., 2009,
Haber, 2003). In rodents, 90–95% of neurons in dorsal striatum and NAc are GABAergic inhibitory
spiny projection neurons, commonly referred to as medium spiny neurons (MSNs), while the
remaining striatal neuronal population consists of interneurons (Gonzales and Smith, 2015). MSNs
project through the basal ganglia via two different routes: the direct pathway project from striatum
to the internal segment of the globus pallidus and the substantia nigra pars reticulate, to the
thalamus and back to cortex (Haber, 2003, Gonzales and Smith, 2015, Joel and Weiner, 2000). The
indirect pathway projects from striatum to the external segment of globus pallidus which connects
reciprocally to the subthalamic nucleus, which in turn project to the internal segment of the globus
pallidus (Haber, 2003, Gonzales and Smith, 2015, Joel and Weiner, 2000). Neurons in the direct
pathway predominantly express D1 dopamine receptors, whereas neurons in the indirect pathway
predominantly express D2 dopamine receptors (Haber, 2003, Gonzales and Smith, 2015, Joel and
Weiner, 2000). However, a number of striatal MSNs projects to both the external and internal globus
pallidus (or substantia nigra pars reticulate) and some MSNs express both D1 and D2 dopamine
receptors (Gonzales and Smith, 2015). The dopaminergic system plays an important modulatory role
in basal ganglia function. Whereas dopamine modulation of the direct pathway is thought to
facilitate movement, dopamine modulation of the indirect pathway is thought to inhibit it (Jin et al.,
2014). However, a recent study found that different subsets of direct and indirect pathway neurons
were engaged during sequence initiation, execution and termination, which suggests that the roles
of the direct and indirect pathway may not be quite so strictly divided (Jin et al., 2014). The striatal
dopaminergic systems have both tonic and phasic patterns of activity. Tonic stimulation of striatal
D2 dopamine receptors by basal dopamine levels is considered essential for normal motor and
cognitive functions of the basal ganglia, whereas sensory-evoked phasic stimulation of D1 dopamine

9
receptors, as seen following appearance of reward-predicting cues, likely provide a teaching signal
for instrumental learning (Marcott et al., 2014, Redgrave et al., 2010, Grace et al., 2007).
Although the majority of striatal cells are medium spiny neurons (MSNs) (Gonzales and Smith,
2015), acetylcholine release from cholinergic interneurons within striatum is thought to modulate
dopamine transmission by acting at both muscarinic and nicotinic acetylcholine receptors (Threlfell
and Cragg, 2011). The subtypes of muscarinic and nicotinic acetylcholine receptors differ between
dorsal striatum and NAc, thus enabling cholinergic interneurons to modulate dopamine transmission
differently in specific striatal sub-regions (Threlfell and Cragg, 2011), which in turn may underlie
differences between striatal sub-regions in their contribution to behaviour (Aoki et al., 2015).
1.1.1 The role of striatum in reward-directed behaviour
The striatum is the main input structure to the basal ganglia and is associated with cognitive and
motivational processing (Haber, 2003) as well as with the execution of motor response (Haber, 2003,
Costa et al., 2004, Pisa and Schranz, 1988) and is considered a key brain region for the regulation of
stimulus-driven behaviour (Yin et al., 2008, Balleine, 2005, 2007). Lesion studies suggest that DLS
(homologue to putamen in humans), DMS (homologue to caudate nucleus in humans) and NAc are
functionally segregated (Balleine et al., 2009, Redgrave et al., 2011). Whereas DMS is considered to
be responsible for acquisition in the early stages of learning and in updating of stimulus-response-
outcome contingencies (Devan et al., 2011, Yin et al., 2005), DLS is primarily associated with
automated stimulus-response behaviour (Yin et al., 2006) and NAc is thought to mainly integrate
motivational aspects of learning (Haber, 2003, Liljeholm and O'Doherty, 2012). Lesions of the DMS in
rats reduce sensitivity to changes in action-outcome contingency as well as post-training outcome
devaluation, suggesting that DMS contributes to behavioural flexibility (Devan et al., 2011) and plays
a key role in the initial phase of goal-directed learning, encoding the association between action and
its specific consequence (Yin et al., 2005). As a task is learned and becomes habitual, responding
becomes dependent on the DLS (Balleine et al., 2009, Tang et al., 2009). When rats are over-trained
on a lever pressing task they become insensitive to changes in outcome value, that is, they continue
pressing the lever even when the reward is devalued (Yin et al., 2006). Several studies suggest DLS
plays a crucial role in the fine tuning of precise motor responses which, through repeated training
and pairings of stimulus-outcome associations, optimise the rat’s motor response toward achieving a
desired outcome (Featherstone and McDonald, 2004, 2005, Balleine et al., 2009, Tricomi and
Lempert, 2015, Pisa and Schranz, 1988, Devan et al., 2011). Rats with lesions in NAc consistently
show a reduction in the vigour of performance during the acquisition of instrumental learning.
However, they remain sensitive to changes in the instrumental contingency (Hart et al., 2014). This

10
suggests that NAc’s involvement in instrumental learning is specific to the modulation of response
vigour or affective arousal. In rats, trained to respond to an auditory cue for reward, neurons in NAc
responded during subsequent exploration of the reward receptacle regardless of whether the
reward was delivered or withheld, whereas uncued entries to the reward receptacle, which were
never rewarded, did not produce excitation in NAc neurons (Nicola et al., 2004b). This finding
demonstrates how NAc reward response can be triggered not just by the actual delivery of reward
but also by conditioned stimuli associated with the reward (Nicola et al., 2004b).
1.1.2 The role of medial prefrontal cortex in behavioural control
Medial Prefrontal cortex (mPFC) plays a crucial role in the organisation of previously acquired
information and in subsequent integration of this information into the planning and execution of
complex behaviour (Groenewegen and Uylings, 2000, Dalley et al., 2004). MPFC is thought to exert
an influence on appetitive behaviour (Riga et al., 2014) via top down control of downstream areas in
nucleus accumbens (NAc) (Riga et al., 2014, Balleine et al., 2009, Christakou et al., 2004, Stefanik et
al., 2015) and medial parts of dorsal striatum (Christakou et al., 2001, Baker and Ragozzino, 2014,
Thorn and Graybiel, 2014). Whereas infralimbic cortex (IL), in ventral mPFC, is associated with habit
formation (Maier, 2015, Smith and Graybiel, 2013), prelimbic cortex (PrL), in dorsal mPFC, is involved
in goal-directed behaviour and complex behaviour that requires flexible switching between different
context-dependent strategies (Riga et al., 2014, Heidbreder and Groenewegen, 2003, Funamizu et
al., 2015).
PrL and IL afferents project mainly from perirhinal, agranular insular and the piriform
cortices, hippocampus and the medial basal forebrain, whereas limbic subcortical information
mainly reaches the PrL and IL via the midline thalamus and the basal nuclei of the amygdala (Hoover
and Vertes, 2007, Vertes et al., 2012, Mattinson et al., 2011, Heidbreder and Groenewegen, 2003).
Furthermore, mPFC is reciprocally connected to the basolateral amygdala (Little and Carter, 2013).
The cell population in mPFC comprises primarily pyramidal neurons, which are excited by glutamate,
cholinergic interneurons as well as inhibitory GABAergic interneurons (Steketee, 2003). Within the
mPFC, dopamine release inhibit pyramidal neurons and stimulates GABA release from GABA
interneurons, which in turn further inhibit pyramidal neurons (Steketee, 2003). PrL projects mainly
to NAc core whereas the NAc shell receives mPFC afferents from IL (Ding et al., 2001, Hart et al.,
2014, Gabbott et al., 2005, Balleine et al., 2009, Groenewegen et al., 1999, Balleine et al., 2007,
Heidbreder and Groenewegen, 2003) as part of the limbic cortico-striatal-thalamic circuit and to
dorsomedial striatum (DMS) as part of the associative cortico-striatal-thalamic circuit (Gabbott et al.,

11
2005, Groenewegen et al., 1999, Hart et al., 2014, Balleine et al., 2007, Balleine and O'Doherty,
2010, Heidbreder and Groenewegen, 2003).
1.1.3 Cortico-striatal circuits
The functional segregation in striatum is further maintained by spatially segregated cortico-striatal-
thalamic circuits, in which DMS, DLS and NAc receive projections from different cortical regions
(Figure 1-1B) and in turn project topographically through the other parts of basal ganglia, to
thalamus, and back to cortex (Redgrave et al., 2011, Haber, 2003, Alexander et al., 1986) (Figure 1-
1A). DMS is part of the associative cortico-striatal-thalamic circuit and receives projections from
medial prelimbic (PrL), cingulate and motor cortex (Van Waes et al., 2012, Balleine et al., 2009). The
associative cortico-striatal-thalamic circuit in involved in acquisition of stimulus-response-outcome
contingencies and behavioural flexibility (Balleine et al., 2007, 2009). In a conditional discrimination
task, contralateral inactivation of PrL and DMS in rats impaired performance in trials when rats had
to change their behaviour to obtain a reward, whereas performance within trial blocks, where no
switching was required, was unaffected (Baker and Ragozzino, 2014). This suggests that
communication between PrL and DMS, as part of the associative cortico-striatal-thalamic circuit,
modulate cue-guided behavioural shifting during tasks that require discrimination between sets of
different stimulus-outcomes.
Figure 1-1 Projection routes in cortico-striatal-thalamic circuits. A. Illustration of cortico-triatal-thalamic circuits, conveying limbic (shown in red), associative (shown in yellow–green) and sensorimotor (shown in blue–white) information (Schematic from Redgrave et al., 2011). B. Cortical input to striatal sub-regions and affiliation of striatal subregions to sensory-motor, associative and limbic cortico-striatal-thalamic circuits. Abbreviations: IL, Infralimbic; PL, Prelimbic; CG, cinculate; M1, motor; M2, medial agranular; SS, somatosensory; I, insular; LO, lateral orbital (schematic from van Waes et al., 2012).
DLS is part of the sensory-motor cortico-striatal-thalamic circuit and receives projections
from primary motor and sensory cortex (Van Waes et al., 2012, Redgrave et al., 2011). The sensory-
motor circuit is involved in automated stimulus-response behaviour (Balleine et al., 2007). In rats,
sensory input to sensory cortex in the form of whisker stimulation led to increased neural activity in

12
DLS (Mowery et al., 2011) and coherence between local field potentials in motor cortex and dorsal
striatum has been found to increase as rats learn an operant task (Koralek et al., 2013), which is
consistent with sensory-motor cortico-striatal-thalamic circuit’s role in over-trained habitual
responding.
NAc is part of the limbic cortico-striatal-thalamic circuit and receives projections from PrL
and IL cortex (Balleine, 2005, Hart et al., 2014, Van Waes et al., 2012). The limbic cortico-striatal-
thalamic circuit maintains motivational aspects of reward-seeking behaviour (Yin et al., 2008,
Balleine, 2005) and disruption of mPFC-NAc core connectivity, through contralateral lesions of mPFC
and NAc core, have been shown to interfere with the planning of responding to reward-paired cues,
but only in trials following immediately after a trial with a rewarded correct response (Christakou et
al., 2004). Thus, the interaction between cortex and NAc is likely to be involved in the updating of
response-outcome contingencies and may be particularly sensitive to recent reinforcement.
Many studies suggest consistent functional differences between striatal sub-regions (Devan
et al., 2011, Balleine et al., 2009). However, adaptive behaviour in a natural environment requires
the ability to associate multiple cues with a variety of possible outcomes and subsequent
implementation of an appropriate behavioural response. Successful behaviour necessitates
integration of reward processing, associative learning and motor planning and thus interaction
between brain regions maintaining these processes (Haber and Knutson, 2010, Joel and Weiner,
2000). Within the striatum, axons and dendrites within each sub-region often cross into other
striatal sub-regions (Haber, 2003). This inter-striatal connectedness, in conjunction with the
striatum’s position in the limbic, associative and somatosensory cortico-striatal-thalamic circuits,
makes the striatum a likely candidate site for interaction and coordination between these circuits,
and organisation of adaptive behavioural output (Liljeholm and O'Doherty, 2012).
As in the striatum, the associative, sensory-motor and the limbic circuits all project to and
from sub-regions of the thalamus (Haber and Calzavara, 2009) and the thalamus may play an
important role in modulation of the joint output of these circuits, with distinct groups of thalamic
nuclei likely contributing to different aspects of sensory, motor, and cognitive processing (Haber and
Calzavara, 2009). Traditionally the thalamus has been regarded primarily as a passive relay station
for sensory and motor signals (Fama and Sullivan, 2015). However, the thalamus is now considered
to also contribute to cognitive processes, including attention, speed of information processing, and
memory (Fama and Sullivan, 2015) and gamma coherence between LFPs in mPFC and mediodorsal
thalamus in relation to reward delivery has been found to increase in rats as a result of instrumental
learning (Yu et al., 2012). The convergence of cortico-striatal-thalamic circuits in sub-regions of the
thalamus makes this structure another possible hub for interaction and coordination between

13
cortico-striatal-thalamic circuits. However, the focus in this thesis will be limited to the role of
striatal subregions and their interaction with prelimbic cortex.
One hypothesis for the role of these integrated cortico-striatal projections is that they
contribute to the regulation of stimulus-driven responses (Mowery et al., 2011, Pidoux et al., 2011)
and may work as a relay station modulated by the salience of the input stimulus, so that the most
salient input is selectively disinhibited. This disinhibition of the input signal may in turn permit
salient sensory signals to dominate the input to targeted motor systems while tonic inhibition of
input from less salient signals is maintained, thereby facilitating the appropriate behavioural
response (Redgrave et al., 2010, 2011, Pidoux et al., 2011). Characterizing the dynamic modulation
of behaviour imposed by the salience of sensory input in behaving animals will be a key step to
understanding the normal function of cortico-striatal projections.
1.1.4 The role of striatum and mPFC in food seeking and its clinical relevance
The use of “natural” rewards such as water or food in an experimental setting allows us to study
neuronal responses to reward directed behaviour in healthy animals, thereby providing vital insight
into how core regions of the brain are affected by reward experience. The presentation of food or
food associated stimuli evokes dopamine release and neuronal activity both in striatum and mPFC
(Carrelli, 2002, Hadad and Knackstedt, 2014, McCutcheon, 2015). The dopamine response to feeding
experience differs between brain areas, possibly underlying the different roles these areas
undertake in the modulation of food seeking and consumption. One study measuring extracellular
dopamine levels using micro-dialysis in rats found that introduction to novel food increases
dopamine release in both mPFC and NAc, whereas repeated exposed to the same food, or to objects
associated with this food, only led to increased dopamine release in mPFC but not in NAc (Bassareo
and Di Chiara, 1997). Another study using fast scan cyclic voltammetry to measure dopamine release
in DMS, DLS and NAc core and shell in rats found that unexpected reward only evoked DA release in
NAc core but not in any other striatal subregion (Brown et al., 2011). Furthermore, reward predictive
cues only evoked dopamine release in NAc core and in DMS but not in NAc shell or DLS (Brown et al.,
2011). In rats trained to associate flavoured saccharin solutions with intragastric infusion of either
sugar or water, injection of D1-like receptor antagonists in NAc, mPFC, the amygdala or lateral
hypothalamus during training, decreased preference for the sugar-associated flavour, further
suggesting that flavour preference is maintained by a network of brain regions (Sclafani et al., 2011).
Obesity, caused by inappropriate feeding habits, is a growing health problem across the
world (Lutsiv et al., 2015, Naef et al., 2015) and has been associated with altered dopamine function
in striatum (Naef et al., 2015), as well as changes in neurotransmitter function in mPFC and thalamus

14
(Blasio et al., 2014, Cole et al., 2015). However, research studying NAc function in relation to feeding
has shown that dopamine release within NAc as well as neuronal firing respond to both “natural”
rewards like food and water and to cocaine self-administration (Carrelli, 2002, Hadad and
Knackstedt, 2014) and dysfunction in neural circuitry involved in food seeking has been associated
with addictive behaviour. Disruption of NAc function in rats produces shift in effort-related choice
behaviour towards decreases willingness to work for food (Nunes et al., 2013) but has also been
found to reduce response inhibition and increase impulsive choice (Feja et al., 2014, Pothuizen et al.,
2005). Dysfunction in the neural processes involved in habituation and reward related learning has
been implicated in several psychiatric disorders related to motivation and attention. Drug addiction
can be defined as a maladaptive compulsive habit and chronic use of cocaine or methamphetamine
has been shown to lead to reorganisation of the dorsal striatum (Belin et al., 2009), (Belin et al.,
2009, Willuhn et al., 2012) and PFC (Hearing et al., 2012). Individuals with antisocial personality
disorder have been shown to have increased volume in the putamen compared with control subjects
and studies suggest that the dorsal striatum in antisocial individuals do not process absence of
reward appropriately, causing it to continuously respond to a stimulus after it has ceased to be
rewarding (Glenn and Yang, 2012). In patients with schizophrenia dopamine transmission in the
striatum is increased during psychotic state and this increased activity is correlated with positive
symptoms such as hallucinations and delusion (Sorg et al., 2012, Goda et al., 2015). Dysregulation of
(mPFC) glutamatergic and cholinergic circuitry has been implicated in disorders such as
schizophrenia, depression and addiction (Mattinson et al., 2011) and Bulimia Nervosa (Hadad and
Knackstedt, 2014). By studying how the brain processes sensory inputs and translates them into a
learned behavioural response, we not only gain a better understanding of processes governing our
everyday behaviour but may also provide important clues to the development of these detrimental
illnesses.
1.1.5 Project overview
Together the subunits of striatum maintain a range of functions crucial for assessing stimulus-
outcome contingencies and optimising the individual’s responses to these cues (Balleine et al., 2009,
Liljeholm and O'Doherty, 2012). Optimising responses to cues requires the retention of learned
stimulus-outcome contingencies, as well as planning and execution of reward-directed motor
responses. Region-specific lesion studies suggest that motivational, motor and cognitive components
of reward-directed behaviour are represented differently in each striatal sub-region (Balleine et al.,
2009, Devan et al., 2011, Hart et al., 2014, Yin et al., 2005, 2006). These lesion studies provide clues
on whether striatal sub-regions are necessary for specific components of reward-related behaviour.

15
However, examining the activity of the three main subareas simultaneously in the non-lesioned
brain allows comparisons between structures within animal and trial as well as an assessment of
how network activity between the sub-regions relates to behavioural choice.
Cortico-striatal communication from medial prefrontal cortex (mPFC) to NAc and DMS likely
play a role in appetitive behaviour, particularly when tasks are demanding and involve shifts
between several stimulus-response-outcome contingencies (Baker and Ragozzino, 2014, Funamizu
et al., 2015, Riga et al., 2014, Heidbreder and Groenewegen, 2003). Examining the activity in mPFC
and striatal sub-regions simultaneously during complex behavioural tasks allows evaluation of the
contribution of cortico-striatal communication to the modulation of behaviour.
Characterizing the dynamic modulation of behaviour imposed by the reward expectation as
well as motor preparation in behaving animals will be a key step to understanding the normal
function of cortico-striatal projections. However, in most standard behavioural paradigms, cues
signalling reward availability also signal to the animal to make a motor response, thereby making
standard behavioural paradigms unable to separate motor and reward component of neural
responses to reward-paired sensory stimuli. However, by developing a novel behavioural paradigm,
in which contribution of these two components can be separated, a greater understanding of striatal
and cortico-striatal modulation of reward-directed behaviour can be achieved.
Overview of aims in experimental chapters
The study reported in Chapter 2 aimed to assess whether the level of salience of sensory input to
DLS affects the sensory representation in structure. To this end, a novel tactile discrimination task
was implemented in head fixed rats, in which sensory stimulation to one whisker is associated with a
reward whereas stimulation of another whisker is associated with reward omission. In animals over-
trained on the discrimination task, DLS evoked tactile responses were expected to be stronger in
response to stimulation of the reward-paired whisker compared with stimulation of the whisker
paired with reward omission.
The study reported in chapter 3 examined the contribution of single unit activity obtained in DMS,
DLS and NAc simultaneously in rats during execution of two comparable conditioned discrimination
tasks; a standard Go-NoGo task (Go-NoGo Minus) and a novel Go-NoGo task (Go-NoGo Plus).
Importantly, in the Go-NoGo Minus task, reward expectancy was exclusively linked to motor
initiation but not with motor suppression, whereas in the Go-NoGo Plus task, reward expectancy
was coupled with either motor initiation or motor suppression in different trials within the same
session. Through comparison of the single unit responses to cue onset in these two tasks, this study
aimed to examine the role of individual striatal sub-regions, as well as communication between sub-

16
regions, on reward expectancy and preparation of motor response during conditioned
discrimination. Striatal sub-regions associated with motor preparation, such as DLS and to lesser
extend NAc, were expected to produce a stronger response to cues signalling motor initiation
compared with DMS, whereas sub-regions modulated by reward expectancy, most notably NAc,
were expected to produce a stronger response to cues signalling the opportunity to obtain a reward
compared with non-rewarded trials.
The study presented in chapter 4 examined the contribution of PrL single unit activity and
synchronisation between PrL and DMS, DLS and NAc during execution of the same two Go-NoGo
behavioural paradigms presented in chapter 3. Because mPFC projects directly to DMS and NAc but
not to DLS, greater task-related synchronisation was expected between PrL and DMS and PrL and
NAc compared with synchronisation between PrL and DLS in response to trial onset cues.

17
Chapter 2: Contribution of Dorsolateral striatum to tactile
processing in the awake rat
2.1 Introduction
Dorsolateral striatum (DLS) is involved in the learning and execution of automatic stimulus-driven
behaviour (Pidoux et al., 2011) and DLS has been implicated in tactile representations (Hawking and
Gerdjikov, 2013, Mowery et al., 2011) and automatic stimulus-response behaviours (Yin et al., 2006).
DLS lesioned rats have difficulty learning tasks that involve precise motor movement whereas
general movement was left unimpaired (Devan et al., 2011) and several studies suggest DLS play a
crucial role in the fine tuning of precise motor responses which, through repeated training and
pairings of stimulus-outcome associations, optimises the rats motor-dependent behaviour toward
achieving a desired outcome (Balleine et al., 2009, Featherstone and McDonald, 2004, Featherstone
and McDonald, 2005, Tricomi and Lempert, 2015, Pisa and Schranz, 1988, Devan et al., 2011). In rats
trained to nose poke in response to an auditory cue signalling reward availability, neurons that
responded to movement showed increased firing when movement was paired with reward than
when it was unrewarded (Kimchi et al., 2009), suggesting that reward expectation also contribute to
DLS evoked responses.
Figure 2-1 Illustration of rat somatosensory cortex and apparatus used in the experimental setup. A. Arrangement of the cortical columns (barrels) in the left somatosensory cortex of a rat. Whiskers in the D row are shown full length with their corresponding barrels highlighted in the cortical map (schematic from Diamond & Arabzadeh,. 2013). B. Example of a microdrive, proportions are identical to the drives used in the current study (Haiss et al., 2010). C. Rats were trained to run through a fixation tunnel and to be fixed by the headpost to eliminate head movement during electrophysiological recordings.
DLS receives strong projections form primary somatosensory cortex (Mowery et al., 2011,
Alloway et al., 1999, Hoffer et al., 2005). In rats and mice, individual whiskers are represented in

18
somatosensory cortex in segregated cortical columns in a map-like fashion (the barrel cortex)
(Miyashita and Feldman, 2013, Diamond and Arabzadeh, 2013) Figure 2-1A), - an arrangement
unparalleled in other systems. Stimulation of neurons in barrel cortex can be linked directly to
activity in the DLS (Mowery et al., 2011, Hawking and Gerdjikov, 2013). Repeated whisker
stimulation evokes neuronal responses in DLS in anaesthetised rats which is consistent with the
extensive projections from somatosensory cortex to the DLS (Syed et al., 2011, Mowery et al., 2011).
Anterograde labelling of projections from individual cortical columns in barrel cortex to DLS have
shown significantly greater overlap between projections into DLS from cortical columns representing
whiskers positioned within the same horizontal row on the rat’s head compared with projections
from cortical columns representing whiskers in different rows (Alloway et al., 1999). These
corticostriatal projections likely affect striatum-mediated regulation of sensory stimulus-driven
responses (Mowery et al., 2011, Pidoux et al., 2011) and may work as a relay station modulated by
the salience of the input stimulus, so that the most salient input is selectively disinhibited. This
disinhibition of the input signal may in turn permit salient sensory signals to dominate the input to
targeted motor systems, thereby facilitating the appropriate behavioural response (Redgrave et al.,
2010, 2011, Pidoux et al., 2011). Several studies suggest DLS play a crucial role in the fine tuning of
precise motor responses which, through repeated training and pairings of stimulus-outcome
associations, optimised the rat’s motor response toward achieving a desired outcome (Featherstone
and McDonald, 2004, 2005, Balleine et al., 2009, Tricomi and Lempert, 2015, Pisa and Schranz, 1988,
Devan et al., 2011). While some studies have found DLS neuronal activity during reward-directed
behaviour to mainly respond to motor aspects of the task (Tang et al., 2007, 2009) other findings
suggest that differences in value associated with the behavioural responses may also modulate firing
(Samejima et al., 2005, Kimchi et al., 2009).
Characterizing the dynamic modulation of behaviour imposed by the salience of sensory input
in behaving animals will be a key step to understanding the normal function of corticostriatal
projections. However, this “salience model” of striatal regulation has not been tested
experimentally.
2.1.1 Whisker stimulation in head fixed rats
Head fixation in awake rodents it a well-established preparation in which a rat is trained to allow
fixation of its head, successfully prohibiting head movement, thus enabling precise stimulation
(deflection) of individual whiskers (Schwarz et al., 2010). Within this setup, the head fixed rat can be
trained to produce a simple instrumental behaviour, such as lever pressing or licking, in response to
single whisker stimulation (Stuttgen and Schwarz, 2010). The projections from barrel cortex to DLS

19
(Alloway et al., 1999, Mowery et al., 2011) are well in accord with the role of that part of striatum in
stimulus-response learning (Balleine et al., 2007) and with findings that the whisker system supports
reward-related learning (Schwarz et al., 2010). As a model, whisker stimulation in the head fixed rat
is uniquely suited to assessing response properties in discrete corticostriatal projections. The head
fixed preparation enables recording of tactile evoked neuronal responses in awake animals without
the contamination of movement artefacts which can complicate analysis of recordings obtained
from freely moving animals (Schwarz et al., 2010). Behaviourally, the system allows the concurrent
conditioning of discrete barrel cortical columns by stimulating individual whiskers in different rows
and stimulation of one whisker may be paired with reward and stimulation of a second whisker with
reward omission. Thus discrete sensory cortical columns will receive identical sensory information
but each corticostriatal channel will have fundamentally different motivational significance. It is
therefore ideally suited to determining whether presentation of a stimulus engaging a given column
biases cortical input to the striatum towards that column.
2.1.2 Aims
By implementing a novel tactile discrimination task, in which sensory stimulation to one whisker is
associated with a reward whereas stimulation of another whisker is associated with reward
omission, this study aims to assess whether the level of salience of sensory input to DLS affects the
sensory representation in this structure.
Hypothesis
In animals overtrained on the discrimination task, DLS evoked tactile responses are expected
to be stronger in response to stimulation of the reward-paired whisker compared with
stimulation of the whisker paired with reward omission.
2.2 Methods
2.2.1 Animals
6 male Sprague-Dawley rats were purchased from Charles Rivers at bodyweight 250-300g. All
animals were kept on reversed light/dark cycle (12:12h; lights on 7.00h). Animals had access to food
(LabDiet 5LF5, PMI Nutrition Intl, Brentwood, MO) ad libitum and animal welfare was monitored
daily. On training and test days water was removed 14-17h before the first training/test session on
the following day. All experiments were carried out under institutional ethical approval and with
project and personal licence approved by the UK Home Office.

20
2.2.2 Initial behavioural screening
Upon arrival animals were housed in pairs and left undisturbed for 4 days. After this they were
handled and given rodent treats (Pitti Heimtierprodukte GmbH, Willich, Germany) daily. A week
after arrival the animals were introduced to a custom made black acrylic fixation box that
simulates a tunnel (height 11cm, depth 21 cm, width at back end 7cm, width at front end 5cm,
Biomedical Workshop, University of Leicester) (Figure 2.1C), secured to the floor of sound-
attenuated aluminium-plated chamber, which also served as a Faraday cage during
electrophysiological recordings. The rats were trained to run through the tunnel, first for treats and
once this had been learned, for water administered through a 1ml syringe. These early habituation
sessions served as a screening process to remove anxious and untrainable animals from the
experiment. In addition to monitoring the animal’s behaviour a bat detector was used during
sessions to ascertain if the animal was emitting 22kHz ultrasonic distress calls (Litvin et al., 2007).
22kHz ultrasonic vocalisation was used to adjust the training to the pace of the individual rat – if a
rat emitted distress calls, the training session was terminated and training started at an earlier
training stage in the next session.
During this process one rat was deemed unsuitable for further testing due to an unusually high
level of anxiety both in and outside the test box and was subsequently removed from the
experiment. Animals that passed the initial behavioural screening were implanted with head posts
and microelectrodes before commencing their training (see “Surgery” for details).
2.2.3 Construction of electrode microdrives
Electrodes: Electrodes were constructed from quartz glass insulated tungsten wire with an outer
diameter of 80μm and a metal core diameter of 20μm (Thomas Recording, Giessen, Germany). The
electrodes were pulled in a vertical puller equipped with a heater element inside an argon filled
chamber and further ground on a rotating grinding disk (Narishige Co Ltd, Tokyo, Japan) to generate
fine tipped electrodes with an impedance of 1.5-3mΩ. The quartz glass on the back end of the
electrode was cracked to expose the tungsten core which was then attached to a Teflon insulated
silver wire (diameter of silver core 125µm, Science Products GmbH, Hofheim, Germany) either
through soldering or with silver epoxy glue (ITW Chemtronics, Kennesaw, Georgia USA). The exposed
joint was insulated and strengthened with nail varnish and dental cement.
Microdrives: Custom microdrives were constructed to enable vertical movement of the
electrodes after implantation, as described in Haiss et al. (2010) (Figure 2.1B). The thread at the tip
of a stainless M1.2 steel screw was removed and a circular groove was inserted into the smooth part
(Biomedical Workshop, University of Leicester). The screw was then placed between two stainless

21
steel guiding rods (1 mm diameter, 5 mm length; Cooper’s Needleworks Ltd., Birmingham, UK)
and the rods were secured to the screw at either end with light curing dental cement (Henry Schein
Inc, Melville, NY USA). This construction enables the top block (rider) of dental cement to be moved
up or down along the screw, whereas the blunted screw tip only allows sideways rotation, thereby
stopping the screw from penetrating through the bottom dental cement block (anchoring block).
Three microelectrodes were each threaded through a polyimide tubing guide (outer diameter
0.163mm, inner diameter 0.125mm, Cole-Palmer, Vernon Hills, Illinois USA) which were aligned and
secured using epoxy glue (Evo-stik Epoxy Express, Bostik Ltd, Strafford, UK). The epoxy secured
tubings were then attached vertically to the anchoring block. The back part of the electrode was
cemented onto the rider block. Before implantation the electrode tips were cleaned with ethanol.
2.2.4 Surgery
Rats were anaesthetised with 4% v/v isofluorane (Schering-Plough) in O2, and maintained between
2-3%. An intramuscular injection of Glucopyrronium Bromide (40μl/kg bodyweight) was given to
slow down gastrointestinal mucus secretion. A sc injection of Baytril (0.2ml/kg bodyweight) was
given at the beginning of surgery. The animal’s rectal temperature was controlled automatically by a
feedback circuit composed of a rectal probe and a heating pad (Harvard Apparatus, Boston,
Massachusetts, USA) set to 37 oC. During surgery the animal received glucose/saline sc infusion
(3ml/hour) and Lacri-Lube Eye Ointment (Allergan, Wesport, Ireland) was applied to the eyes to
prevent corneal desiccation. The animal was placed in a stereotactic frame and an incision was made
along the midline and the periosteum was retracted to expose the skull. 12% hydrogen-peroxide
solution (Vet Way Ltd; York, UK) was applied to the exposed skull to enable identification of bregma
and lambda. The skull was then treated with light curing etching gel (Henry Schein Inc, Melville, NY
USA) to improve bonding of dental cement to the skull. 11 stainless steel anchoring screws (Morris
Co., Southbridge, Massachusetts, USA, part number 0X 1/8 flat)) were affixed to the cranium (3
screws to the frontal plate, 4 screws to the side of and 3 screws to the top of the parietal plate and 1
screws to the interparietal plate) to enable secure placement of the implant which was built up using
light-curing dental cement (Henry Schein Inc, Melville, NY USA). A silver wire (Science Products
GmbH, Hofheim, Germany) was connected with silver paint to one skull screw in the interparietal
plate and one skull screw in the frontal plate, to ground the animal during the electrophysiological
recordings. A craniotomy was made on the right side of the cranium and the dura was removed
immediately before insertion of 3 recording electrodes into the DLS; AP from bregma: -1.4mm,
ML/DV from bregma: +3.2mm/-3.4mm, +4.0mm/-3.7mm and +4.6mm/-5.0mm (Paxinos and
Watson, 2007). The exposed dura and the cavity around the inserted electrodes were covered with

22
antibacterial ointment (Fuciderm). The anchoring block on the microdrive was secured to the skull
cap with dental cement and a custom made aluminium tower (outer diameter 8mm, inner diameter
6.5mm, Biomedical Workshop, University of Leicester) was placed around the Microdrive to protect
it from grooming. The top of the tower was closed with a custom made screw-cap (Biomedical
Workshop, University of Leicester) to protect the drive from dirt, yet still provide access to the drive.
The grounding wires and the silver wires connected to the recording electrodes were soldered to the
male side of a micro plug microplug (Mill-Max Mfg. Corp, Oyster Bay, NY, USA), which was secured
to the skull cap with dental cement. A head post in the form of a custom made aluminium
(Biomedical Workshop, University of Leicester) was secured to the back part of the skull cap with
dental composite (Henry Schein Inc, Melville, NY USA). The skin and muscle layer around the
exposed skull and skull cap were cleaned with Povidone-Iodine (Animalcwere Ltd, York, UK) and a
layer of Fusiderm (Dechra Veterinary Products A/S, Uldum, Denmark) and the incision at the front
and back of the skull cap was sutured together. Analgesia was administered 2-3 hours before the
end of surgery (Carprieve, 0.1ml/kg bodyweight, s.c.; Norbrook Laboratories, Carlisle, UK). The
animal was removed from the stereotactic frame. However, heating and oxygen were provided until
it recovered from the anaesthesia after which it was returned to its home cage.
The animal was giving analgesia (Carpofen, 0.1ml/kg bodyweight per day) for 3 days and
antibiotics (Baytril, 0.2ml/kg bodyweight per day) for 5 days post op. In addition animals were given
intraperitoneal injections of glucose/saline solution for 3-5 days until the animal was eating dry diet
and gaining bodyweight. Training commenced minimum 7 days after surgery once the animal had
regained its pre-operation body weight.
2.2.5 Behavioural training and testing
Habituation to head fixation apparatus: The rats were reintroduced to the sound attenuated
chamber and to the fixation tunnel and trained to run through the tunnel for water administered
through a 1ml syringe. The rats were habituated to being fixed by the head post upon exiting the
tunnel in incremental stages: Initially the head post was gently post touched by the experimenter
while the rat received water upon exiting the tunnel, incrementally the gentle touch was replaced
with a firmer hold of the head post by the experimenter, briefly limiting head movement and finally
the head post was secured into a custom made bracket (Biomedical Workshop, University of
Leicester) (Figure 2.1C), completely preventing movement of the head. Initially, this head fixation
lasted less than a second while the rat was given water continuously. As the training progressed, the
duration of the head fixation was slowly increased and single droplets of water, with an increasing
waiting period between droplets, replaced the delivery of continuous water during head fixation.

23
Once the rats were habituated to head fixation, the licking spout was introduced. The licking spout
was constructed from a plastic pipette with a steel end, to prevent the plastic from being chewed.
Licks were detected as deflections of the spout recorded by a miniature piezo film sensor (Part nr.
FS-2513P; Farnell, Leeds, UK) glued to the underside of the plastic pipette. The spout was connected
to a container filled with 50ml water attached to the side of the attenuated sound chamber
approximately 20cm above the spout and the amount of water released was controlled by the
opening time of a magnetic valve (Takasago Electric Inc, Nagoya, Japan; WTA-2R-N3F). In the first
training session with the spout, the rats were allowed to explore and drink freely from the spout. In
the following training session, and after, the rat was head fixed as described above and water
rewards were delivered thought the licking spout, with the tip of the spout positioned 3-5 mm in
front of the rat’s lower lip.
Whisker stimulation: The whisker stimulators were constructed from a glass capillary glued
to a piezo actuator (Physik Instrumente, Karlsruhe, Germany). Whiskers on the right side of the rat’s
head were trimmed and maintained at a length of 1cm for the duration of the project. The whisker
stimulator was advanced slowly toward the right side of the rat’s head and the rat was habituated to
the vibration of the stimulator touching its whiskers before threading of the whiskers were
attempted. Once the rat was habituated to the stimulator, a whisker was threaded into the capillary
of the stimulator and the rat continued onto Pavlovian conditioning.
Pavlovian conditioning: Animals were trained to associate single whisker stimulation with
liquid reward delivered from the spout positioned immediately in front of the rat. Each stimulation
was delivered at 60hz and with an amplitude (whisker deflection) of 0.3mm and was always
presented for 1.5 seconds. The stimulation of a single whisker signalled a 1.5 sec response interval
during which licking the spout was rewarded. At the beginning of the Pavlovian conditioning training
the rat was rewarded with a drop of water even if it did not lick during the response interval (the
reward was provided at the end of the response interval) to strengthen the association between the
whisker-stimulation and the reward. Once the rat had learned the association between licking and
obtaining the water reward, only licking within the response interval triggered a reward. Random
licking outside the response interval was discouraged by introducing a dead time before each
stimulus. This entailed delaying the next stimulus by 5-20 seconds if the rat licks outside the
response interval.
Discrimination task: Animals were trained to discriminate between stimulation of two
different whiskers, where licking during stimulation of one whisker (Go) triggered the water reward,
whereas licking during stimulation to the other whisker (NoGo) triggered a LED light (Maplin
Electronics, Wombwell, UK) consisting of 9 LEDs, which served as negative feedback on false

24
alarms (Figure 2-2). In the early training sessions the amplitude of the Go stimulus was high (0.5mm)
compared with the NoGo stimulus (0.1mm). The two target whiskers differed between rats but were
always the same for each rat. During the session the difference in amplitude between the Go and
NoGo stimulation was decreased in small increments until the amplitude reaches 0.3mm for both Go
and NoGo. Once the rat learned to discriminate between stimulation of the two whiskers, sessions
were started with a smaller difference in amplitude (Go = 0.4; NoGo = 0.2) to indicate which whisker
signals reward after which the session was continued with the same amplitude of Go and NoGo. The
rats were considered to be discriminating at criterion when Hit rate (no. correct responses to Go
cue/total no. Go trials) was above 0.75 and False Alarm rate (no. incorrect responses to NoGo
cue/total no. NoGo trials) was below 0.25 within a session for 3 consecutive sessions.
Reversal task: Only one rat continued through to reversal training. The paradigm for the
reversal sessions were the same as used in the discrimination task, except the whisker that signalled
Go cue in the discrimination task now signalled NoGo cue and vice versa.
Figure 2-2 Discrimination task Rats were trained to discriminate between rewardable (Go) and non-rewardable (NoGo) vibrotactile stimulation delivered to individual whiskers.
Online control of the hardware and analysis of the animals' behaviour during the experimental
paradigms were implemented via in-house software written in LabView (National Instruments,
Austin, TX, USA) and a standard multi-purpose AD/DA board (model NI 6229). Animals were tested in
darkness and were constantly monitored by the experimenter via an infrared USB camera (Maplin
Electronics, Wombwell, UK).
2.2.6 Electrophysiological recordings
15-30 minutes prior to each recorded session, each electrode was lowered approximately 0.32mm
(corresponding to a 45 degree turn of the drive screw) in an attempt to sample neuronal responses
from different locations in the target structure. The head fixed rat was connected through a flexible
wire, to a 16 channel head stage (Plexon Inc., Dallas, TX, USA) immediately before recording. During
the discrimination task, wideband signals were acquired continuously via an op-amp based head-
stage amplifier (HST/8o50-G1-GR, 1x gain, Plexon Inc., Dallas, TX, USA), passed through a
preamplifier (PBX2/16wb, 1000x gain; Plexon Inc., Dallas, TX, USA) and digitized at 40,000 Hz.

25
All data processing was done offline. Recorded field potentials were down sampled to 5,000 Hz and
evoked responses extracted from the raw data using a 200 Hz low-pass Butterworth filter.
Timestamps for cue onsets and licking responses were synchronised in neuroexplorer (Nex
Technologies, Madison, AL, USA). Further analyses were calculated using Neuroexplorer and custom-
written Matlab routines.
2.2.7 Technical challenges with obtaining single unit recordings
The headpost and microdrives were implanted in one surgery and the majority of the behavioural
training occurred after the microdrives had been implanted. As the electrophysiological recordings
were made in over-trained animals, this meant that the implanted electrodes were embedded in the
brain of the rat for months before recordings could be obtained. In the presented dataset no single
units were recorded, which we attribute to this long period between implantation and recording
(Prasad et al., 2012). In order to increase the quality of the recording electrodes by decreasing the
duration they were imbedded in tissue before recording, a second group of 8 rats were trained and
implanted using a modified surgical procedure, in which only the skull cap and head post were fixed
after the initial behavioural screening and tetrode tungsten electrodes1 were implanted in a second
surgery, after the rat had successfully learned the discrimination task. However, this change in
procedure caused the skull cap to become structurally unstable and no electrophysiological
recordings were obtained from these rats. Data from this later group of rats will not be presented
here.
2.3 Results
2.3.1 Behaviour
Three rats successfully learned to discriminate between stimulation of the Go and NoGo paired
whisker (Figure 2-3). In addition, one rat (rat 1 in Figure 2-3) also learned to discriminate between
Go and NoGo stimulus after reversal, i.e., when the whisker previously associated with Go cue
instead signalled NoGo cue and the whisker previously associated with NoGo cue instead signalled
Go cue (Reversal) (Figure 2-3C).
1 For details on manufacturing of tetrode electrodes, see Chapter 3, Methods.
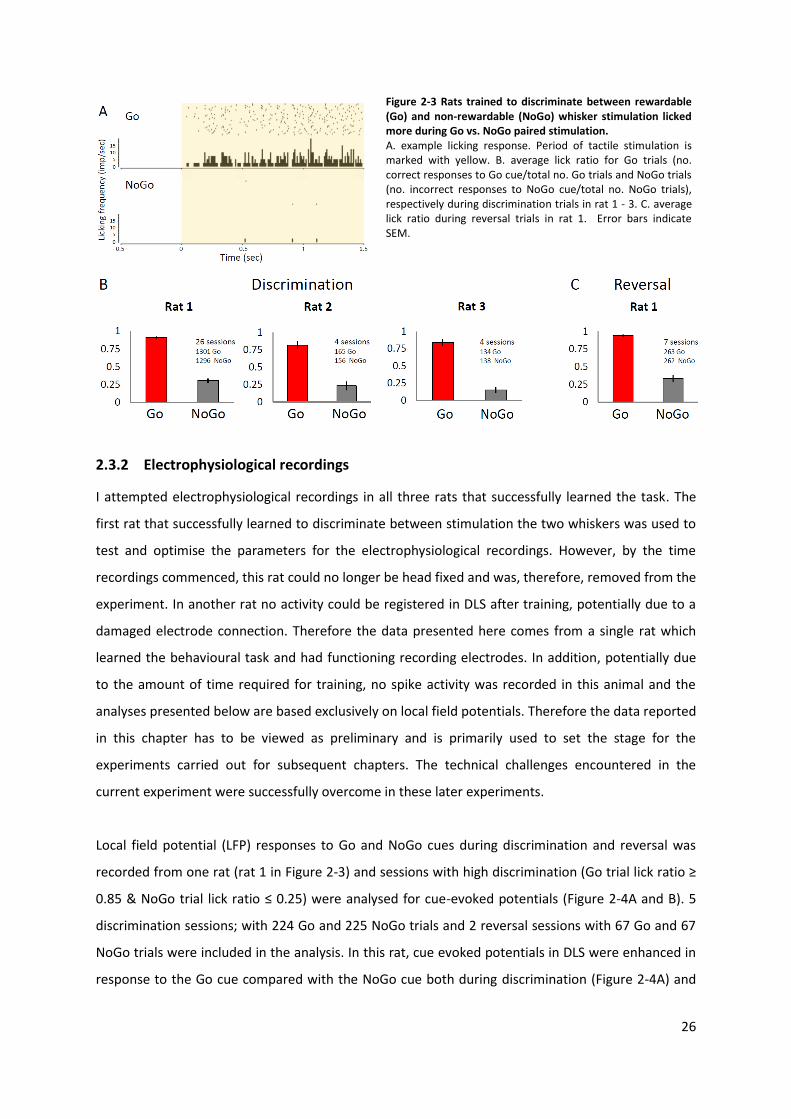
26
Figure 2-3 Rats trained to discriminate between rewardable (Go) and non-rewardable (NoGo) whisker stimulation licked more during Go vs. NoGo paired stimulation. A. example licking response. Period of tactile stimulation is marked with yellow. B. average lick ratio for Go trials (no. correct responses to Go cue/total no. Go trials and NoGo trials (no. incorrect responses to NoGo cue/total no. NoGo trials), respectively during discrimination trials in rat 1 - 3. C. average lick ratio during reversal trials in rat 1. Error bars indicate SEM.
2.3.2 Electrophysiological recordings
I attempted electrophysiological recordings in all three rats that successfully learned the task. The
first rat that successfully learned to discriminate between stimulation the two whiskers was used to
test and optimise the parameters for the electrophysiological recordings. However, by the time
recordings commenced, this rat could no longer be head fixed and was, therefore, removed from the
experiment. In another rat no activity could be registered in DLS after training, potentially due to a
damaged electrode connection. Therefore the data presented here comes from a single rat which
learned the behavioural task and had functioning recording electrodes. In addition, potentially due
to the amount of time required for training, no spike activity was recorded in this animal and the
analyses presented below are based exclusively on local field potentials. Therefore the data reported
in this chapter has to be viewed as preliminary and is primarily used to set the stage for the
experiments carried out for subsequent chapters. The technical challenges encountered in the
current experiment were successfully overcome in these later experiments.
Local field potential (LFP) responses to Go and NoGo cues during discrimination and reversal was
recorded from one rat (rat 1 in Figure 2-3) and sessions with high discrimination (Go trial lick ratio ≥
0.85 & NoGo trial lick ratio ≤ 0.25) were analysed for cue-evoked potentials (Figure 2-4A and B). 5
discrimination sessions; with 224 Go and 225 NoGo trials and 2 reversal sessions with 67 Go and 67
NoGo trials were included in the analysis. In this rat, cue evoked potentials in DLS were enhanced in
response to the Go cue compared with the NoGo cue both during discrimination (Figure 2-4A) and
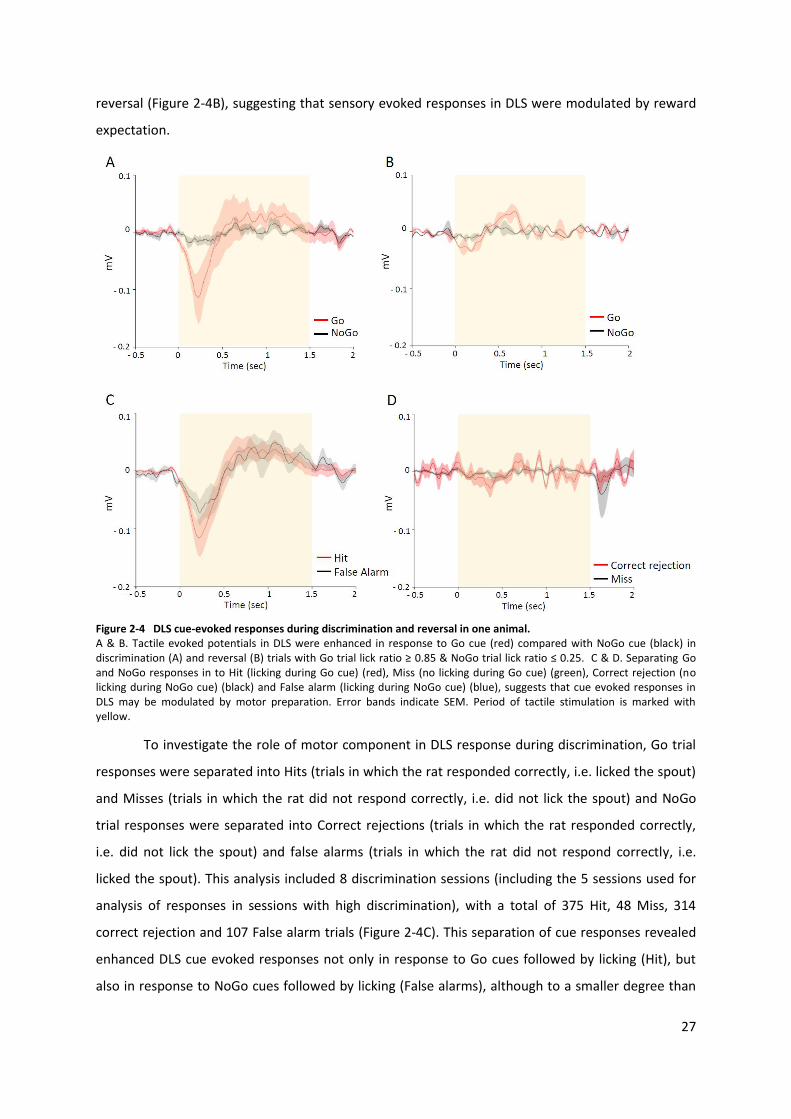
27
reversal (Figure 2-4B), suggesting that sensory evoked responses in DLS were modulated by reward
expectation.
Figure 2-4 DLS cue-evoked responses during discrimination and reversal in one animal. A & B. Tactile evoked potentials in DLS were enhanced in response to Go cue (red) compared with NoGo cue (black) in discrimination (A) and reversal (B) trials with Go trial lick ratio ≥ 0.85 & NoGo trial lick ratio ≤ 0.25. C & D. Separating Go and NoGo responses in to Hit (licking during Go cue) (red), Miss (no licking during Go cue) (green), Correct rejection (no licking during NoGo cue) (black) and False alarm (licking during NoGo cue) (blue), suggests that cue evoked responses in DLS may be modulated by motor preparation. Error bands indicate SEM. Period of tactile stimulation is marked with yellow.
To investigate the role of motor component in DLS response during discrimination, Go trial
responses were separated into Hits (trials in which the rat responded correctly, i.e. licked the spout)
and Misses (trials in which the rat did not respond correctly, i.e. did not lick the spout) and NoGo
trial responses were separated into Correct rejections (trials in which the rat responded correctly,
i.e. did not lick the spout) and false alarms (trials in which the rat did not respond correctly, i.e.
licked the spout). This analysis included 8 discrimination sessions (including the 5 sessions used for
analysis of responses in sessions with high discrimination), with a total of 375 Hit, 48 Miss, 314
correct rejection and 107 False alarm trials (Figure 2-4C). This separation of cue responses revealed
enhanced DLS cue evoked responses not only in response to Go cues followed by licking (Hit), but
also in response to NoGo cues followed by licking (False alarms), although to a smaller degree than

28
in Hit trials, suggesting that motor preparation may constitute a significant component to reward
paired cue evoked responses in DLS.
The observed enhanced responses to Go stimulation compared with NoGo stimulation was
consistent throughout discrimination and reversal sessions in the one recorded. When trials were
further divided into correct and incorrect behavioural response, the enhanced response observed to
stimulation in Hit and False alarm trials were consistent throughout the analysed discrimination
sessions.
2.4 Discussion
To assess whether the level of salience of the sensory input to DLS affects the sensory
representation in this structure, a novel tactile discrimination task was implemented, in which
sensory stimulation to one whisker is associated with a reward whereas stimulation of another
whisker was associated with reward omission. Rats were trained to lick a spout in response to
stimulation of the rewardable (Go) whisker while abstaining from licking when the non-rewardable
(NoGo) whisker was stimulated (Figure 2-2). All three rats presented in this chapter successfully
learned to discriminate between the two stimulated whiskers and to adjust their behaviour to
optimise the outcome (Figure 2-3). A similar two whisker discrimination task was recently
implemented by Ollerenshaw et al. (2014) to examine the role of adaptation on stimulus detection
and discrimination. In their setup, rats were also trained to discriminate between stimulation of two
distinct whiskers associated with either reward or reward omission. However, in their experiment,
these cue-paired discriminative stimulations were either preceded by stimulation of both whiskers
(adaptation) or not stimulated prior to cue-paired discriminative stimulation (no adaptation) and this
adaptation was found to improve discrimination (Ollerenshaw et al., 2014). The above study, along
with the findings presented here, demonstrate the versatility of using two whisker stimulation in
behaving rodents to examine the role of additional factors affecting the processing of sensory input,
be it pre-cue stimulation as in the above study or reward-value as in the current study. The current
study is the first to use this tactile Go-NoGo discrimination task to address the role of salience on
sensory processing in DLS neuronal ensembles.
In terms of the neurophysiological data obtained in the current study, due to technical
challenges no spike activity was detected and I was only able to record local field potential data from
a single rat as detailed in the results section. The data obtained was largely consistent with my
hypotheses and the observed effect of trial type on stimulus-evoked DLS responses was consistent
throughout discrimination and reversal sessions. Given the very limited amount of data however,

29
the current results can only be viewed as preliminary. Here they are used to motivate and set the
stage for the experiments reported in subsequent chapters where the technical challenges
encountered here were overcome successfully.
Analysis of LFP response to stimulus onset in sessions with high accuracy of execution
revealed enhanced evoked potentials in Go trials compared with NoGo trials, suggesting that DLS
evoked potentials are influenced by differences in reward-value (Figure 2-4A). However, when
sessions with lower accuracy were included into the analysis and stimulus-evoked DLS responses
were also divided into correct and incorrect behavioural response, an enhanced evoked response to
stimulus onset was not only observed in Go trials with correct response (Hits), but also in NoGo trials
with incorrect response (False Alarms) (Figure 2-4C). In both Hit and False Alarm trials the rat licked
the spout in response to stimulus onset, suggesting that the enhanced LFP response in DLS may be
associated with movement initiation. In comparison, only a very small response was observed in
NoGo trials in which the rat correctly suppressed licking (Correct Rejections), and no response was
seen in Go trials where the rat failed to lick (Misses) (Figure 2-4D), further suggesting that DLS
response to stimulus onset were not affected by differences in reward-value associated with the two
stimulations. Indeed, previous work examining T-maze choice behaviour in rats have observed an
increase in DLS activity during execution of the task as result of training (Barnes et al., 2011, Root et
al., 2010, Thorn et al., 2010, Kimchi et al., 2009). However, this increased DLS activity was more
strongly associated with movement onset, rather than instructional cue. The cue-evoked DLS
response observed in the current study may be more strongly related to the initiation of movement
immediately after presentation of the cue rather than to the cue itself. In the T-maze studies
mentioned above, the instructional cue was associated with increased activity in dorsomedial
striatum, rather than DLS, as a result of training (Root et al., 2010, Thorn et al., 2010, Horvitz, 2009).
In the T-maze choice task the instruction cue signals to the animal to initiate a motor
response as well as the availability of reward in every trial, thereby making it difficult to analyse the
contribution of reward-value to responses in neurons that are also associated with motor response.
In Monkeys performing a reward-directed motor task, motor-related neurons in putamen
(homologous of DLS in rodents) were found to be modulated by reward probability immediately
before and during initiation of reward-directed movement (Pasquereau et al., 2007, Hassani et al.,
2001), and findings from another study in monkeys executing a similar reward-directed motor task,
suggest that some motor-related neurons in putamen encode the value associated with action
rather than the action itself (Samejima et al., 2005). In rats trained to nose poke in response to an
auditory cue signalling reward availability, neurons that responded to movement showed increased
firing when movement was paired with reward compared with unrewarded movement (Kimchi et

30
al., 2009). Similarly, in the current study, a greater LFP response was observed in Hit trials than in
False alarm trials, suggesting that reward expectation also contribute to DLS evoked responses.
Thus, it is possible that the rat licked in response to the NoGo stimulus in False Alarm trials because
it mistook the NoGo stimulus for a Go stimulus, in which case the enhanced LFP response in False
Alarm trials may still be influenced by reward expectation.
In the standard Go-NoGo task, cues signalling reward availability also signal to the animal to
make a motor response, whereas cues signalling the animal to suppress motor response are not
rewarded. Therefore, it is not possible to separate motor and reward component of striatal
responses to reward-paired sensory stimuli using the Go-NoGo paradigm in its original form. To
separate these two components, I propose a modification of the standard Go-NoGo task, wherein
not only correct responses to Go cue (Hit) are rewarded but also correct responses to NoGo cue
(Correct rejection) are rewarded. With this modification, reward availability would be signalled by
both cue types but only Go cues would signal to the rat to initiate a motor response, which would
enable examination of reward expectation and motor preparatory components of cue-evoked
striatal responses. Previous work suggests that distinct subregions of striatum contribute differently
to reward and motor components of learned reward-directed behaviour (Balleine et al., 2009,
Liljeholm and O'Doherty, 2012). Electrophysiological recording of neurons in DMS and nucleus
accumbens as well as DLS during execution of the proposed modified Go-NoGo task may provide
information about subregional differences in cue-evoked responses a well as information about the
role of communication between striatal subregions during execution of a complex reward-driven
behavioural task.
2.4.1 Conclusion
LFP response to stimulus onset was found to be enhanced in trials in which cue onset was
immediately followed by initiation of motor response, suggesting the observed response is
associated with motor initiation. However, the enhanced response in False Alarm trials may still be
caused by the rat, incorrectly, expecting a reward for its response. Because of the restricted dataset,
the current findings should be viewed as preliminary, however they do support the significance of
considering not only the valence of the conditioned cue (here the tactile stimulus) but also the
associated behavioural response when interpreting cue-triggered neural responses in Go-NoGo
tasks. Further research is needed to investigate the extent to which DLS responses reflect the reward
value of the conditioned cue vs. response initiation associated with reward retrieval. To this end, a
modified version of the standard Go-NoGo task is proposed in the next chapter. To avoid the
technical challenges encountered here the next chapter also moves away from the head-fixed

31
preparation and successfully implements tetrode recordings in freely moving animals using tones
rather than tactile stimuli as conditioned cues.

32
Chapter 3: Exploring motor and reward components of striatal
responses to reward-paired auditory cues
3.1 Introduction
3.1.1 The role of striatum in reward-directed behaviour
Adaptive behaviour in a natural environment requires the ability to associate multiple cues with a
variety of possible outcomes and subsequent implementation of an appropriate behavioural
response. Successful behaviour necessitates integration of reward processing, associative learning
and motor planning and thus interaction between brain regions maintaining these processes (Haber
and Knutson, 2010, Joel and Weiner, 2000). The striatum is part of the reward circuitry (Haber, 2003,
Tricomi and Lempert, 2015), and is associated with cognitive and motivational processing (Tricomi
and Lempert, 2015, Basar et al., 2010, Baldo and Kelley, 2007), as well as with the execution of
motor responses (Costa et al., 2004, Pisa and Schranz, 1988, Haber, 2003). Rat striatum consists
dorsally of dorsomedial striatum (DMS), homologue to caudate nucleus in humans, and dorsolateral
striatum (DLS), homologue to putamen in humans, and ventrally of nucleus accumbens (NAc) and
the olfactory tubercle (Devan et al., 2011, Balleine et al., 2009, Haber, 2003). Together, DMS, DLS
and NAc maintain a range of functions crucial for assessing stimulus-outcome contingencies and
optimising the individual’s responses to these stimuli (Balleine et al., 2009, Liljeholm and O'Doherty,
2012). However, region-specific lesion studies suggest that motivational, motor and cognitive
components of reward-directed behaviour are represented differently in each striatal sub-region
(Balleine et al., 2009, Devan et al., 2011, Hart et al., 2014, Yin et al., 2005, 2006).
In my previous study (presented in chapter 2) I established a tactile Go-NoGo task (Figure 2-
2), in order to assess whether the level of salience of sensory input to DLS affects the sensory
representation in this structure. The initial findings in this study suggested that DLS responses are
related to motor initiation, but may also be modulated by the behavioural significance of sensory
input. However, in a standard Go-NoGo task, cues signalling reward availability also signal to the
animal to make a motor response, thereby making this behavioural paradigm unable to separate
motor and reward component of striatal responses to reward-paired sensory stimuli. To distinguish
between these two processes, I have developed a modified version of the standard Go-NoGo task, in
which correct responses in both trial types were rewarded (Figure 3-1). With this modification,
reward availability was signalled by both cue types but only Go cues signalled to the rat to initiate a

33
motor response, thereby enabling examination of reward expectation and motor initiation
components of cue-evoked striatal responses. Previous work suggests that DLS, DMS and NAc
contribute differently to specific components of learned reward-directed behaviour and adaptive
behaviour requires successful integration of input from all three striatal sub-regions (Balleine et al.,
2009, Liljeholm and O'Doherty, 2012). Examining the activity of DLS, DMS and NAc simultaneously in
the non-lesioned brain may increase our understanding of the function of these sub-regions as well
as how network activity between the subregions relates to behavioural choice.
3.1.2 Dorsomedial striatum
DMS is strongly associated with goal-directed learning (Shiflett et al., 2010, Shan et al., 2014, Yin et
al., 2005, Balleine et al., 2009, Devan et al., 2011, Hilario et al., 2012), and updating of stimulus-
response-outcome contingencies (Devan et al., 2011, Yin et al., 2005). In a conditional discrimination
task where rats relied on visual cues to determine which arm in a maze was rewarded in the
following block of 3-5 trials, inactivation of DMS increased perseverance errors, that is, the rat
required more trials within a new block to switch arm or completely failed to switch arm (Baker and
Ragozzino, 2014). Similarly, when rats were trained to discriminate sets of odours and bedding type
paired with reward from unrewarded odour/bedding-sets in a foraging task, rats with lesions in DMS
failed to modify their behaviour when previously unrewarded odour/bedding sets became rewarded
and vice versa (Lindgren et al., 2013). Recently, single unit activity in DMS during the action selection
phase in a rat decision making task, has been found to code for behavioural response in previous
trial (Ito and Doya, 2015). In fact, numerous studies have demonstrated that lesions or inactivation
of DMS in rats, either before or after training, reduce sensitivity to changes in response-outcome
contingency, as well as post-training outcome devaluation, thus, making action choice habitual and
rigid (Shiflett et al., 2010, Shan et al., 2014, Yin et al., 2005, Balleine et al., 2009, Devan et al., 2011,
Hilario et al., 2012, Hart et al., 2014) strongly implicating DMS processing in the ability to switch
behaviour in response to changes in context-outcome associations (Devan et al., 2011, Lindgren et
al., 2013, Baker and Ragozzino, 2014). DMS and DLS function independently of each other but also
compete for control over stimulus-response behaviour (Hilario et al., 2012, Balleine et al., 2009). As
a behavioural response is learned, and becomes habitual, control of this behaviour shifts from DMS
to DLS (Dias-Ferreira et al., 2009, Balleine et al., 2009, Yin et al., 2005). However, control reverts
back to DMS if DLS function is disrupted or if increased vigilance or reassessment of behaviour
becomes advantageous (Balleine et al., 2009, Yin et al., 2005).
In humans and monkeys, activation of the caudate nucleus, equivalent of DMS in rats have
been found to increase in response to higher probability of reward and decreased when reward was

34
omitted, suggesting activity in the DMS is sensitive to reward expectancy (Tricomi and Lempert,
2015, Fanelli et al., 2013, Kawagoe et al., 1998).
Lesions of DMS do not significantly affect forelimb reaching movement in rats, whereas
lesions of DLS have been found to impair forelimb reaching (Pisa and Schranz, 1988), suggesting that
DMS is not directly involved in execution of motor response. However, rats trained to nose poke for
reward to olfactory cues decreased the vigour of their response when lesioned in the DMS,
suggestion that DMS modulates the force of learned motor-dependent reward-directed responses
(Wang et al., 2013).
3.1.3 Dorsolateral striatum
DLS is associated with sensory processing (Mowery et al., 2011) and learning and execution of
automated stimulus-response behaviour (Devan et al., 2011, Costa et al., 2004, Mowery et al., 2011,
Schmitzer-Torbert et al., 2015, Fanelli et al., 2013, Yin et al., 2006). As a task is learned and becomes
automated, responding becomes dependent on the DLS (Balleine et al., 2009, Dias-Ferreira et al.,
2009, Tang et al., 2009). Rats over-trained on a lever pressing task become insensitive to changes in
outcome value, that is, they continue pressing the lever even when the reward is devalued (Yin et
al., 2006, Balleine et al., 2009). This training induced insensitivity to changes in outcome value is
coupled to changes to plasticity in DLS and post training lesions in DLS reinstate the sensitivity to
outcome value (Balleine et al., 2009).
Previous work has shown the majority of neurons in DLS to be movement-related (Tang et
al., 2007, 2009) and these neurons were found to decrease their firing rate in response to repeated
training on a movement-dependent task while the smaller population of non-movement related
neurons increased or maintained their firing rate (Tang et al., 2007, 2009). This suggests that as an
automated response is learned the response moves from being facilitated by a large number of
neurons to being modulated by a smaller population of stronger firing neurons. DLS lesioned rats
have difficulty learning tasks that involve precise motor movement whereas general movement was
left unimpaired (Devan et al., 2011) and several studies suggest DLS play a crucial role in the fine
tuning of precise motor responses which, through repeated training and pairings of stimulus-
outcome associations, optimised the rats motor-dependent behaviour toward achieving a desired
outcome (Balleine et al., 2009, Featherstone and McDonald, 2004, Featherstone and McDonald,
2005, Tricomi and Lempert, 2015, Pisa and Schranz, 1988, Devan et al., 2011).
Single neurons in DLS have also been found to increase their activity during movement
triggered by external cues (Devan et al., 2011, graybiel et al., 1994). In rats trained to nose poke in
response to an auditory cue signalling reward availability, neurons that responded to movement

35
showed increased firing when movement was paired with reward than when it was unrewarded
(Kimchi et al., 2009), suggesting that reward expectation also contribute to DLS evoked responses.
3.1.4 Nucleus Accumbens
NAc has been shown to regulate motivational (Tricomi and Lempert, 2015, Basar et al., 2010, Baldo
and Kelley, 2007) and reward-related components of behaviour (Tricomi and Lempert, 2015).
In humans, NAc activation has been shown to be unaffected by variation in reward
probability, but is decreased when reward is devalued, suggesting NAc is sensitive to motivational
value of reward (Tricomi and Lempert, 2015). In rats, trained to respond to an auditory cue for
reward, NAc single units responded during subsequent exploration of the reward receptacle
regardless of whether the reward was delivered or withheld, whereas uncued entries to the reward
receptacle, which were never rewarded, did not produce excitation in NAc single units (Nicola et al.,
2004b). This suggests that NAc reward response may be driven by conditioned stimuli associated
with the reward or be associated with reward-seeking motor response (Nicola et al., 2004b).
The choice of behaviour, e.g. initiate or suppress a motor response, is guided by the
assessment of the available cues, the expected outcome of responding to this cue and cost of
responding (Basar et al., 2010). In rats trained on a lever pressing task, optogenetic inhibition of NAc
MSNs after feedback improved responses in next trial (Aquili et al., 2014) implicating NAc in the
updating of response-outcome contingencies. Changes in NAc firing have been linked to reward-
directed motor response, (Roitman et al., 2005) and lesions in NAc, although to lesser extent than
lesions in dorsal striatum, have been found to impair movement (Pisa and Schranz, 1988, Wang et
al., 2013, Hart et al., 2014). A recent study examined the effect of motor and reward component on
NAc single unit responses (Roitman and Loriaux, 2014). This study also used a modified version of
the Go-NoGo task where correct responses in both trial types were rewarded. Rats were trained to
discriminate between light cues presented either above the extended lever (Go trial cue) or on the
opposite side of the pellet magazine (NoGo trial cue) and correct responses in both trial types (lever
press or withholding lever press, respectively) were rewarded with a sugar pellet. In addition to the
visual cue, an auditory cue (white noise) was presented along with either the Go or NoGo trial light
cue. In this paradigm NAc units displayed a greater increase in firing rate or a smaller decrease in
firing rate in response to trial cue in trials where the rats withheld lever press compared to trials
where the rats pressed the lever, regardless of trial type, suggesting that NAc single unit activity
correlates with initiation of motor response rather than outcome expectancy (Roitman and Loriaux,
2014).

36
3.1.5 Interaction between striatal sub-regions
Together the sub-regions of striatum maintain a range of functions crucial for assessing stimulus-
outcome contingencies and optimising the individual’s responses to these stimuli (Balleine et al.,
2009, Liljeholm and O'Doherty, 2012). Although lesion studies suggest consistent functional
differences between striatal subregions (Devan et al., 2011, Balleine et al., 2009), axons and
dendrites within each sub-region often cross into other subregions (Haber, 2003), which in
conjunction with its position in Cortico-striatal-thalamic circuits, may facilitate cross-regional
integration of information (Haber, 2003), and organisation of adaptive behavioural output (Liljeholm
and O'Doherty, 2012).
In a recent study, Ito and Doya (2015) recorded single unit activity simultaneously in DLS,
DMS and ventral striatum in rats during performance of a choice task. In their paradigm, rats were
trained to nose poke into a central port and await another tone cue indicating which of two ports
(“left” or “right”) offered the highest probability of reward. In a subset of trials a third “choice” tone
cue was presented, which offered no information about the probability of reward in the left and
right port (Ito and Doya, 2015). Striatal subregions were found to respond differently to distinct
elements of the task, with ventral striatum displaying the highest firing rate response at the
initiation of the trial, DLS and DMS responding to onset and offset of the cue tone, respectively, and
all three regions responding as the rat approached the left or right port (Ito and Doya, 2015). In this
task all choices involved initiation of motor response and, thus, makes it difficult to distinguish
between striatal activity related to motor initiation and reward expectation. However, this study
demonstrates how recording single units in DLS, DMS and NAc simultaneously in animals that are
executing a complex behavioural task, provide information about how activity within each sub-
region relate to distinct elements of a specific task.
The execution of optimal behaviour in complex tasks such Go-NoGo tasks, requires the
integration of learned stimulus-response-outcome contingencies, flexible switching between motor
initiation and suppression as well as motivational processes, which likely involve contribution from
dorsal striatal subregions as well as NAc and may also require communication between subregions
to facilitate adaptive behavioural outputs. Region-specific lesion studies provide clues on whether
striatal subregions are necessary for reward-related behaviour. However examining the activity of
the three main subareas in the non-lesioned brain allows comparisons between structures within
animal and trial as well as an assessment of how network activity between the subregions relates to
behavioural choice.

37
3.1.6 Study aims
Characterizing the dynamic modulation of behaviour imposed by the reward expectation as well as
motor preparation in behaving animals will be a key step to understanding the normal function of
striatal sub-regions. However, in most standard behavioural paradigms, cues signalling reward
availability also signal to the animal to make a motor response, thereby making standard
behavioural paradigms unable to separate motor and reward component of neural responses to
reward-paired sensory stimuli. To distinguish between these two components of striatal sensory
responses, one group of rats were tested in a modified version of the Go-NoGo task, in which rats
had to press a lever in response to a Go cue and supress responding to the NoGo cue. In the
modified task (hereafter referred to as the “Go-NoGo Plus” task), correct responding in both trial
types were rewarded with a sugar pellet. A second group of rats were trained in a standard Go-NoGo
task (hereafter referred to as the “Go-NoGo Minus” task) where only correct responses to Go cues
were rewarded and correct suppression of lever press in NoGo trials had no programmed
consequence. Evoked responses in DLS, DMS and NAc were recorded simultaneously during these
discrimination tasks in overtrained rats.
Through comparison the single unit responses to cue onset in these two tasks, the current study
aimed to examine the role of individual striatal sub-regions, as well as communication between
subregions, on reward expectancy and preparation of motor response during conditioned
discrimination.
3.1.7 Hypothesis
Striatal sub-regions associated with motor preparation, such as DLS and to lesser extend
NAc, were expected to produce a stronger response to cues signalling motor initiation
compared with DMS.
Striatal sub-regions modulated by reward expectancy, most notably NAc, were expected to
produce a stronger response to cues signalling the opportunity to obtain a reward compared
with non-rewarded trials.

38
3.2 Methods
3.2.1 Animals
Male Lister Hooded rats (n = 7; Charles River, Cambridge, UK) weighing 225-250g on arrival were
kept on reversed light/dark cycle (12:12h; lights on 19.00h). Animals had access to water ad libitum
and access to food (LabDiet 5LF5, PMI Nutrition Intl, Brentwood, MO) for at least 2h per day. All
experiments were carried out under institutional ethical approval and with project and personal
licence approved by the UK Home Office.
3.2.2 Apparatus
All behavioural training and testing was conducted in four identical operant chambers [30 x 31 x 24
cm (height x width x depth); Med Associates Inc., St Albans, VT]. In each chamber a magazine for
delivery of sugar pellets (Dustless Precision Pellets, Bio Serv, Sheffield UK) was fitted in the middle of
one of the chamber walls with a retractable lever positioned to the left of the magazine. Both
magazine and lever were positioned 2.5 cm above the metal grid floor of the chamber and had a
light positioned immediately above each. A speaker was positioned above the magazine just below
the ceiling of the box and a house light was positioned at the top of the opposite wall of the
chamber.
For the electrophysiological recordings a modified operant chamber was used. The modified
operant chamber differed from the standard operant chamber in that all metal surfaces (walls and
grid floor) were covered by Plexiglas sheets to minimise electrical noise. In addition the wall-fitted
magazine was replaced by a custom made square receptacle (2 x 5 x 3 cm (height x width x depth);
Rob Hemmings, School of Psychology, University of Leicester) attached to the grid floor 3.5 cm from
the wall. The magazine light signalling reward was placed on the wall behind and above the floor-
fitted magazine. This alteration was made as previous work in our group had shown that the
Neuralynx tetrode drive restricted the rat’s access to the wall fitted magazine.
3.2.3 Behavioural training
Upon arrival rats were pair housed and left undisturbed for 4 days, followed by 1-2 days of
habituation to handling by the experimenter before beginning the behavioural training. After
handling a handful of sugar pellets (Dustless Precision Pellets, Bio Serv, Sheffield UK) was left in the
home cage, the same sugar pellets were used as reinforcement throughout training. Behavioural
training consisted of the following stages:
Magazine training. On the first training day each rat was allowed to explore the operant
chamber. During the magazine training session no levers were presented but the house light and
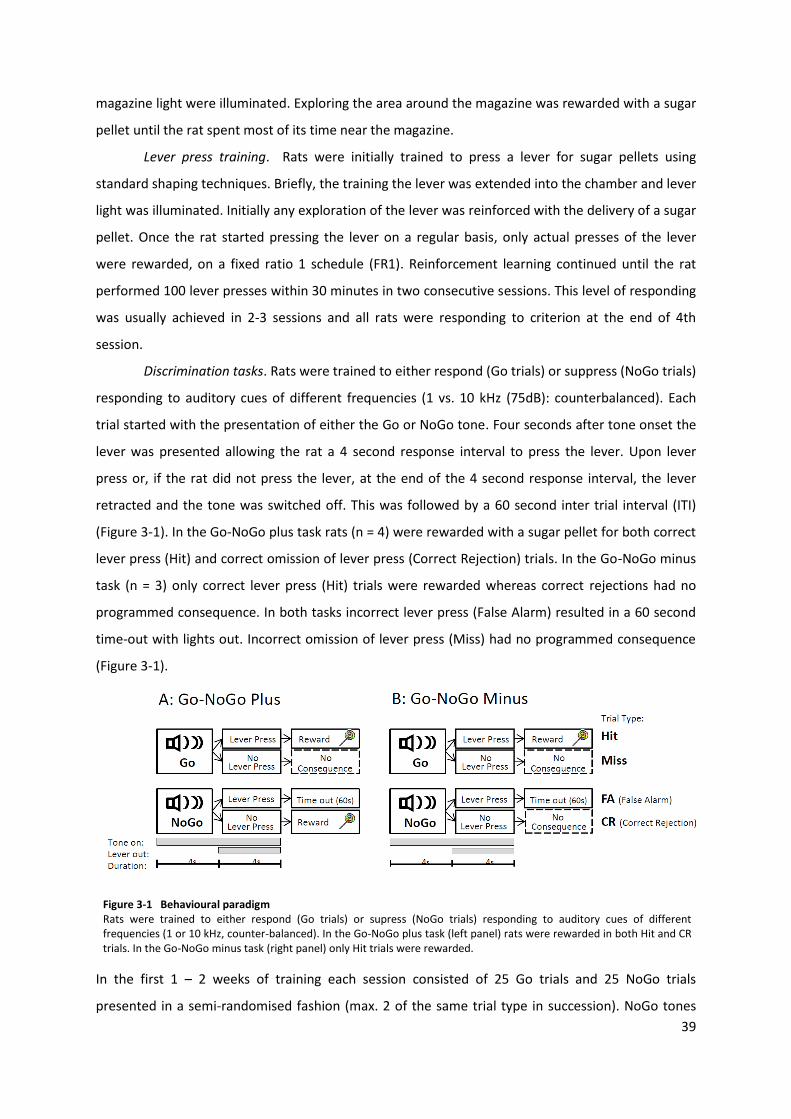
39
magazine light were illuminated. Exploring the area around the magazine was rewarded with a sugar
pellet until the rat spent most of its time near the magazine.
Lever press training. Rats were initially trained to press a lever for sugar pellets using
standard shaping techniques. Briefly, the training the lever was extended into the chamber and lever
light was illuminated. Initially any exploration of the lever was reinforced with the delivery of a sugar
pellet. Once the rat started pressing the lever on a regular basis, only actual presses of the lever
were rewarded, on a fixed ratio 1 schedule (FR1). Reinforcement learning continued until the rat
performed 100 lever presses within 30 minutes in two consecutive sessions. This level of responding
was usually achieved in 2-3 sessions and all rats were responding to criterion at the end of 4th
session.
Discrimination tasks. Rats were trained to either respond (Go trials) or suppress (NoGo trials)
responding to auditory cues of different frequencies (1 vs. 10 kHz (75dB): counterbalanced). Each
trial started with the presentation of either the Go or NoGo tone. Four seconds after tone onset the
lever was presented allowing the rat a 4 second response interval to press the lever. Upon lever
press or, if the rat did not press the lever, at the end of the 4 second response interval, the lever
retracted and the tone was switched off. This was followed by a 60 second inter trial interval (ITI)
(Figure 3-1). In the Go-NoGo plus task rats (n = 4) were rewarded with a sugar pellet for both correct
lever press (Hit) and correct omission of lever press (Correct Rejection) trials. In the Go-NoGo minus
task (n = 3) only correct lever press (Hit) trials were rewarded whereas correct rejections had no
programmed consequence. In both tasks incorrect lever press (False Alarm) resulted in a 60 second
time-out with lights out. Incorrect omission of lever press (Miss) had no programmed consequence
(Figure 3-1).
Figure 3-1 Behavioural paradigm Rats were trained to either respond (Go trials) or supress (NoGo trials) responding to auditory cues of different frequencies (1 or 10 kHz, counter-balanced). In the Go-NoGo plus task (left panel) rats were rewarded in both Hit and CR trials. In the Go-NoGo minus task (right panel) only Hit trials were rewarded.
In the first 1 – 2 weeks of training each session consisted of 25 Go trials and 25 NoGo trials
presented in a semi-randomised fashion (max. 2 of the same trial type in succession). NoGo tones
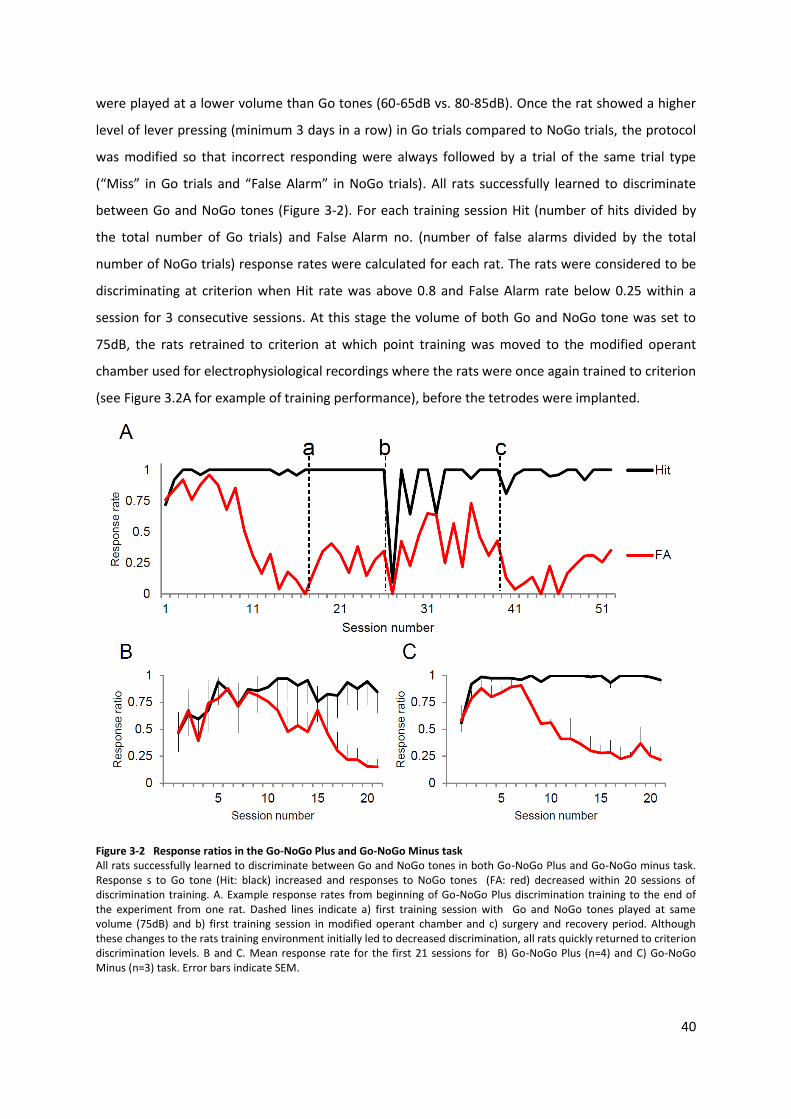
40
were played at a lower volume than Go tones (60-65dB vs. 80-85dB). Once the rat showed a higher
level of lever pressing (minimum 3 days in a row) in Go trials compared to NoGo trials, the protocol
was modified so that incorrect responding were always followed by a trial of the same trial type
(“Miss” in Go trials and “False Alarm” in NoGo trials). All rats successfully learned to discriminate
between Go and NoGo tones (Figure 3-2). For each training session Hit (number of hits divided by
the total number of Go trials) and False Alarm no. (number of false alarms divided by the total
number of NoGo trials) response rates were calculated for each rat. The rats were considered to be
discriminating at criterion when Hit rate was above 0.8 and False Alarm rate below 0.25 within a
session for 3 consecutive sessions. At this stage the volume of both Go and NoGo tone was set to
75dB, the rats retrained to criterion at which point training was moved to the modified operant
chamber used for electrophysiological recordings where the rats were once again trained to criterion
(see Figure 3.2A for example of training performance), before the tetrodes were implanted.
Figure 3-2 Response ratios in the Go-NoGo Plus and Go-NoGo Minus task All rats successfully learned to discriminate between Go and NoGo tones in both Go-NoGo Plus and Go-NoGo minus task. Response s to Go tone (Hit: black) increased and responses to NoGo tones (FA: red) decreased within 20 sessions of discrimination training. A. Example response rates from beginning of Go-NoGo Plus discrimination training to the end of the experiment from one rat. Dashed lines indicate a) first training session with Go and NoGo tones played at same volume (75dB) and b) first training session in modified operant chamber and c) surgery and recovery period. Although these changes to the rats training environment initially led to decreased discrimination, all rats quickly returned to criterion discrimination levels. B and C. Mean response rate for the first 21 sessions for B) Go-NoGo Plus (n=4) and C) Go-NoGo Minus (n=3) task. Error bars indicate SEM.
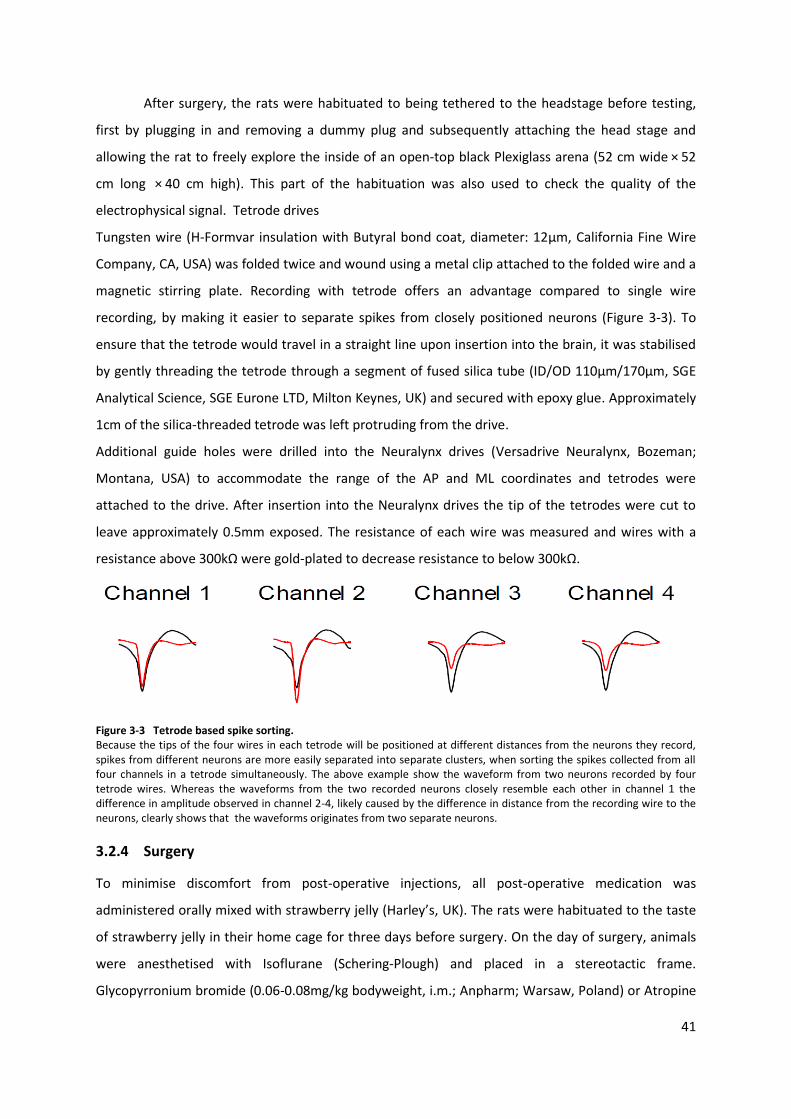
41
After surgery, the rats were habituated to being tethered to the headstage before testing,
first by plugging in and removing a dummy plug and subsequently attaching the head stage and
allowing the rat to freely explore the inside of an open-top black Plexiglass arena (52 cm wide × 52
cm long × 40 cm high). This part of the habituation was also used to check the quality of the
electrophysical signal. Tetrode drives
Tungsten wire (H-Formvar insulation with Butyral bond coat, diameter: 12μm, California Fine Wire
Company, CA, USA) was folded twice and wound using a metal clip attached to the folded wire and a
magnetic stirring plate. Recording with tetrode offers an advantage compared to single wire
recording, by making it easier to separate spikes from closely positioned neurons (Figure 3-3). To
ensure that the tetrode would travel in a straight line upon insertion into the brain, it was stabilised
by gently threading the tetrode through a segment of fused silica tube (ID/OD 110μm/170μm, SGE
Analytical Science, SGE Eurone LTD, Milton Keynes, UK) and secured with epoxy glue. Approximately
1cm of the silica-threaded tetrode was left protruding from the drive.
Additional guide holes were drilled into the Neuralynx drives (Versadrive Neuralynx, Bozeman;
Montana, USA) to accommodate the range of the AP and ML coordinates and tetrodes were
attached to the drive. After insertion into the Neuralynx drives the tip of the tetrodes were cut to
leave approximately 0.5mm exposed. The resistance of each wire was measured and wires with a
resistance above 300kΩ were gold-plated to decrease resistance to below 300kΩ.
Figure 3-3 Tetrode based spike sorting. Because the tips of the four wires in each tetrode will be positioned at different distances from the neurons they record, spikes from different neurons are more easily separated into separate clusters, when sorting the spikes collected from all four channels in a tetrode simultaneously. The above example show the waveform from two neurons recorded by four tetrode wires. Whereas the waveforms from the two recorded neurons closely resemble each other in channel 1 the difference in amplitude observed in channel 2-4, likely caused by the difference in distance from the recording wire to the neurons, clearly shows that the waveforms originates from two separate neurons.
3.2.4 Surgery
To minimise discomfort from post-operative injections, all post-operative medication was
administered orally mixed with strawberry jelly (Harley’s, UK). The rats were habituated to the taste
of strawberry jelly in their home cage for three days before surgery. On the day of surgery, animals
were anesthetised with Isoflurane (Schering-Plough) and placed in a stereotactic frame.
Glycopyrronium bromide (0.06-0.08mg/kg bodyweight, i.m.; Anpharm; Warsaw, Poland) or Atropine
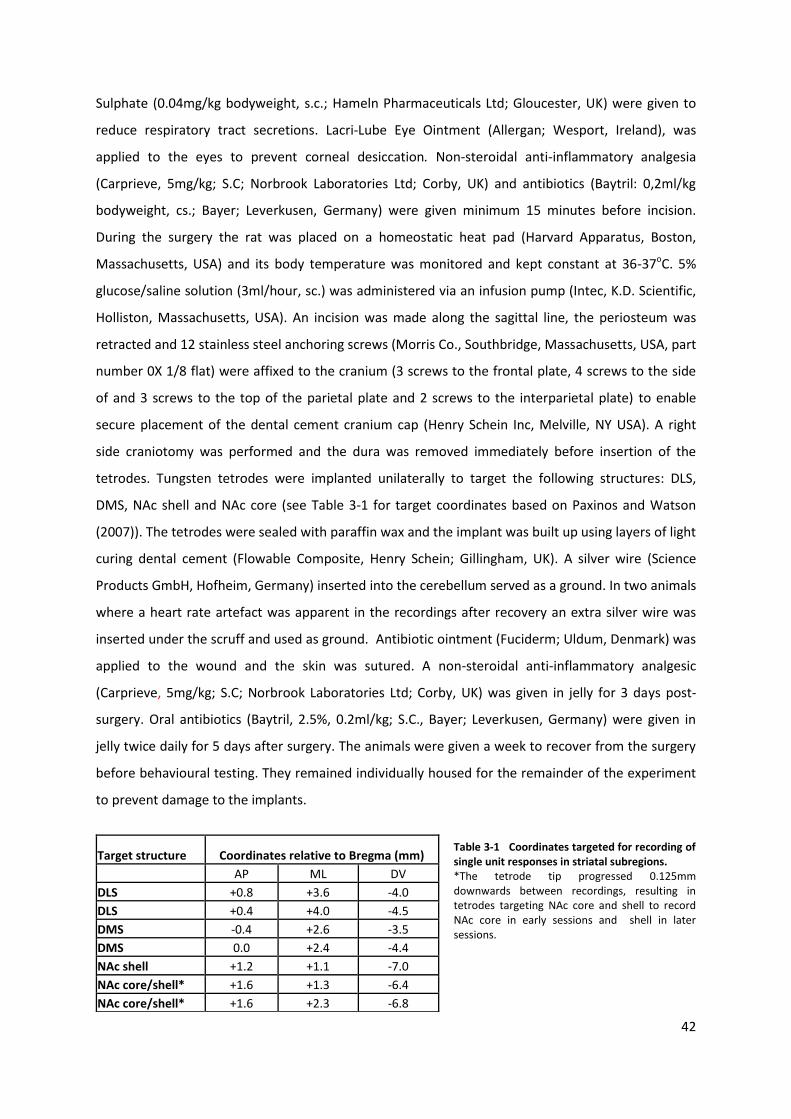
42
Sulphate (0.04mg/kg bodyweight, s.c.; Hameln Pharmaceuticals Ltd; Gloucester, UK) were given to
reduce respiratory tract secretions. Lacri-Lube Eye Ointment (Allergan; Wesport, Ireland), was
applied to the eyes to prevent corneal desiccation. Non-steroidal anti-inflammatory analgesia
(Carprieve, 5mg/kg; S.C; Norbrook Laboratories Ltd; Corby, UK) and antibiotics (Baytril: 0,2ml/kg
bodyweight, cs.; Bayer; Leverkusen, Germany) were given minimum 15 minutes before incision.
During the surgery the rat was placed on a homeostatic heat pad (Harvard Apparatus, Boston,
Massachusetts, USA) and its body temperature was monitored and kept constant at 36-37oC. 5%
glucose/saline solution (3ml/hour, sc.) was administered via an infusion pump (Intec, K.D. Scientific,
Holliston, Massachusetts, USA). An incision was made along the sagittal line, the periosteum was
retracted and 12 stainless steel anchoring screws (Morris Co., Southbridge, Massachusetts, USA, part
number 0X 1/8 flat) were affixed to the cranium (3 screws to the frontal plate, 4 screws to the side
of and 3 screws to the top of the parietal plate and 2 screws to the interparietal plate) to enable
secure placement of the dental cement cranium cap (Henry Schein Inc, Melville, NY USA). A right
side craniotomy was performed and the dura was removed immediately before insertion of the
tetrodes. Tungsten tetrodes were implanted unilaterally to target the following structures: DLS,
DMS, NAc shell and NAc core (see Table 3-1 for target coordinates based on Paxinos and Watson
(2007)). The tetrodes were sealed with paraffin wax and the implant was built up using layers of light
curing dental cement (Flowable Composite, Henry Schein; Gillingham, UK). A silver wire (Science
Products GmbH, Hofheim, Germany) inserted into the cerebellum served as a ground. In two animals
where a heart rate artefact was apparent in the recordings after recovery an extra silver wire was
inserted under the scruff and used as ground. Antibiotic ointment (Fuciderm; Uldum, Denmark) was
applied to the wound and the skin was sutured. A non-steroidal anti-inflammatory analgesic
(Carprieve, 5mg/kg; S.C; Norbrook Laboratories Ltd; Corby, UK) was given in jelly for 3 days post-
surgery. Oral antibiotics (Baytril, 2.5%, 0.2ml/kg; S.C., Bayer; Leverkusen, Germany) were given in
jelly twice daily for 5 days after surgery. The animals were given a week to recover from the surgery
before behavioural testing. They remained individually housed for the remainder of the experiment
to prevent damage to the implants.
Target structure Coordinates relative to Bregma (mm)
AP ML DV
DLS +0.8 +3.6 -4.0
DLS +0.4 +4.0 -4.5
DMS -0.4 +2.6 -3.5
DMS 0.0 +2.4 -4.4
NAc shell +1.2 +1.1 -7.0
NAc core/shell* +1.6 +1.3 -6.4
NAc core/shell* +1.6 +2.3 -6.8
Table 3-1 Coordinates targeted for recording of single unit responses in striatal subregions. *The tetrode tip progressed 0.125mm downwards between recordings, resulting in tetrodes targeting NAc core and shell to record NAc core in early sessions and shell in later sessions.

43
3.2.5 Electrophysiological recordings
15-30 minutes prior to each recorded session, each tetrode was lowered approximately 0.125mm
(corresponding to a 180 degree turn of the drive screw) to ensure that different neurons were
recorded in each session. The rat was connected through a flexible wire, allowing unimpaired
movement, to a 32 channel head stage (Plexon Inc., Dallas, TX, USA) immediately before recording
and placed in an operant chamber positioned in a sound-attenuated aluminium-plated box, which
served as a Faraday cage. During the discrimination task, wideband signals were acquired
continuously at a sampling rate of 25 kHz via an op-amp based head-stage amplifier (HST/32o25-
36P-GR, 1x gain, Plexon Inc., Dallas, TX, USA), and passed through a preamplifier (PBX2/32wb, 1000x
gain; Plexon Inc., Dallas, TX, USA). For spike sorting the raw signal was band-pass filtered offline
300-3,000Hz and single channel recordings were referenced to the average of all recorded channels.
For one rat in the Go-NoGo Minus task a lower signal/noise ratio was found when referencing to a
single channel not showing any spikes, therefore, referencing to a single channel was used in this rat.
Artefacts (identified as events occurring simultaneously in 8 channels) were removed using a custom
made Matlab code supplied by Manuel Molano, Systems Neuroscience Group, University of
Leicester. Spikes from each session with amplitudes above 5x the SD of background noise were
sorted as tetrodes using the Matlab-based Wave_clus software to yield single-unit spike trains
(Quiroga et al., 2004). Timestamps from single unit spike trains and timestamps for cue onset and
lever press obtained from the operant chambers were synchronised in Neuroexplorer (Nex
Technologies, Madison, AL, USA). Further analyses were calculated using Neuroexplorer and custom-
written Matlab routines. All statistical analysis was calculated using SPSS 22 Statistics (IBM SPSS,
Somers, NY, USA).
3.2.6 Verification of tetrode placement
After the last recording session, the rats were anaesthetised with Isoflurane. The area
around each tetrode tip was lesioned by passing a 30µA current through each electrode wire for 15
seconds. The rats were then perfused with 5% formal saline, the brains were removed and kept
refrigerated (5o C) in formal saline for 24 hours, then transferred into a 30% sucrose solution and
kept refrigerated for a further 2-3 days after which they were rapidly frozen using dry ice and hexane
and stored at -20o C. Tetrode placement was verified visually while cutting the frozen brains in 30μm
slices on a cryostat (Figure 3-4). Not all tetrodes could be identified from the brain slices. In these
cases, the medial-lateral (ML) and anterior-posterior (AP) position of the non-observed tetrode was
calculated based on the observed position of neighbouring tetrodes. To verify if the tip of each
observed tetrode had been identified, the observed dorsal-ventral (DV) position of the presumed tip
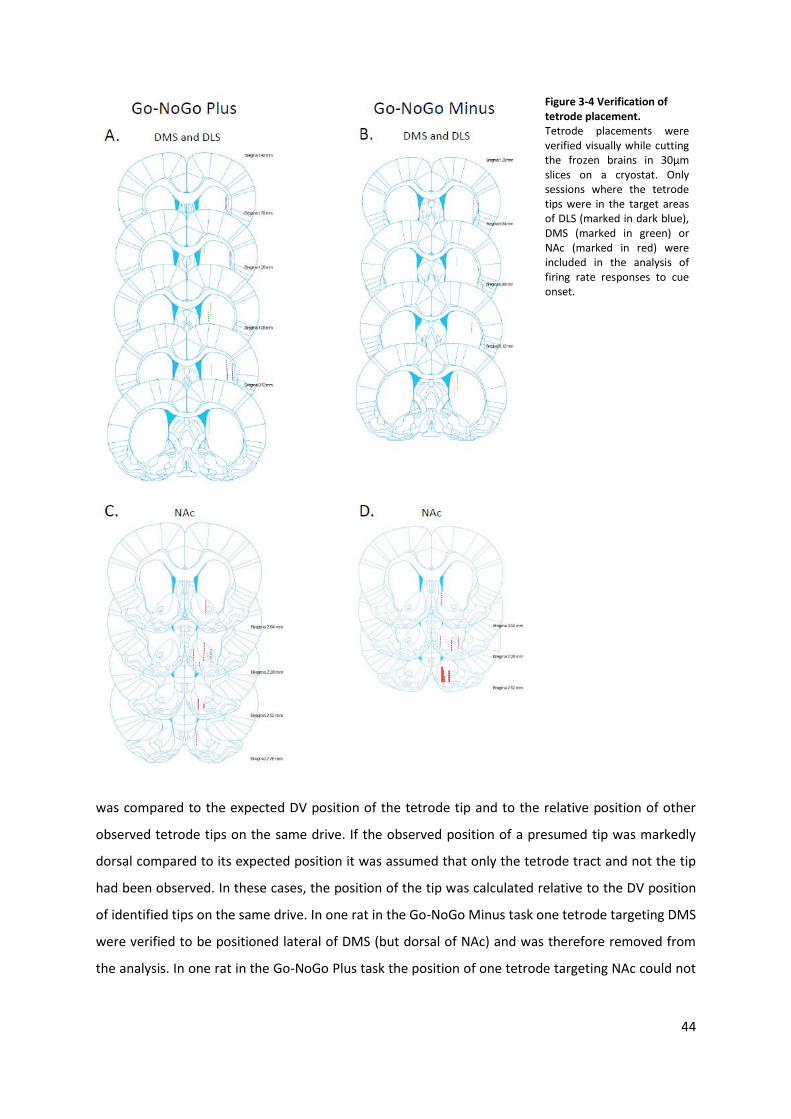
44
Figure 3-4 Verification of tetrode placement. Tetrode placements were verified visually while cutting the frozen brains in 30μm slices on a cryostat. Only sessions where the tetrode tips were in the target areas of DLS (marked in dark blue), DMS (marked in green) or NAc (marked in red) were included in the analysis of firing rate responses to cue onset.
was compared to the expected DV position of the tetrode tip and to the relative position of other
observed tetrode tips on the same drive. If the observed position of a presumed tip was markedly
dorsal compared to its expected position it was assumed that only the tetrode tract and not the tip
had been observed. In these cases, the position of the tip was calculated relative to the DV position
of identified tips on the same drive. In one rat in the Go-NoGo Minus task one tetrode targeting DMS
were verified to be positioned lateral of DMS (but dorsal of NAc) and was therefore removed from
the analysis. In one rat in the Go-NoGo Plus task the position of one tetrode targeting NAc could not
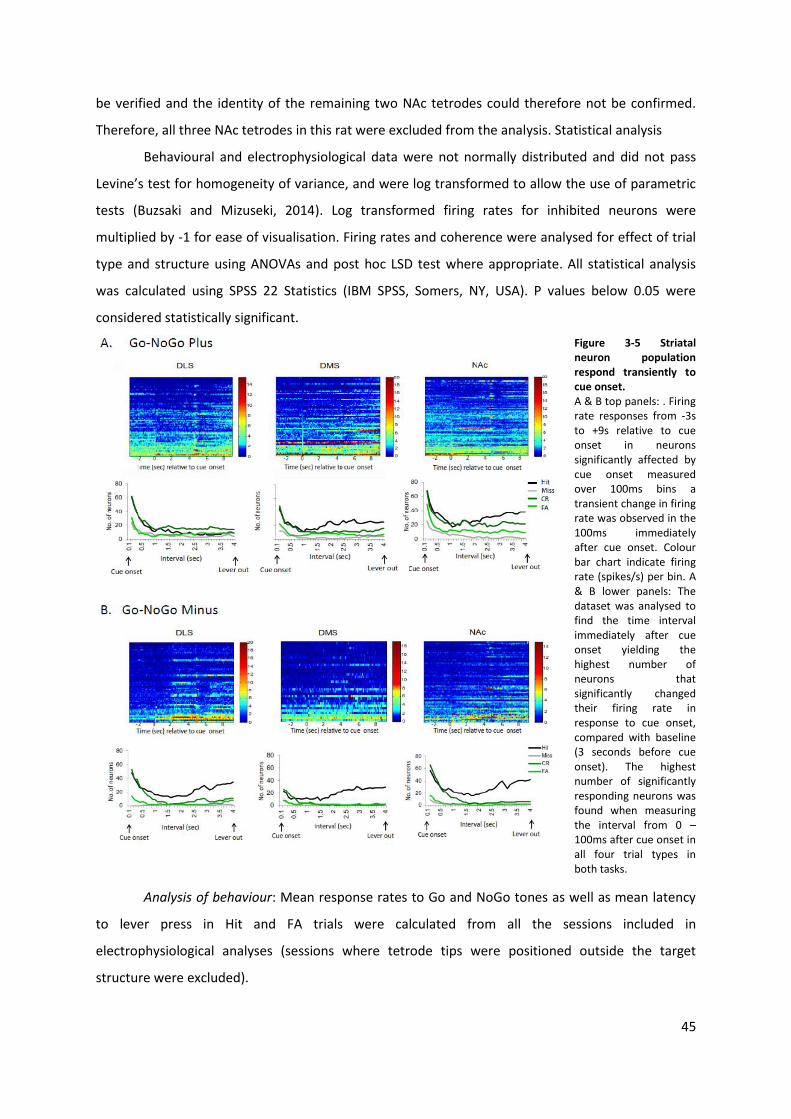
45
be verified and the identity of the remaining two NAc tetrodes could therefore not be confirmed.
Therefore, all three NAc tetrodes in this rat were excluded from the analysis. Statistical analysis
Behavioural and electrophysiological data were not normally distributed and did not pass
Levine’s test for homogeneity of variance, and were log transformed to allow the use of parametric
tests (Buzsaki and Mizuseki, 2014). Log transformed firing rates for inhibited neurons were
multiplied by -1 for ease of visualisation. Firing rates and coherence were analysed for effect of trial
type and structure using ANOVAs and post hoc LSD test where appropriate. All statistical analysis
was calculated using SPSS 22 Statistics (IBM SPSS, Somers, NY, USA). P values below 0.05 were
considered statistically significant.
Figure 3-5 Striatal neuron population respond transiently to cue onset. A & B top panels: . Firing rate responses from -3s to +9s relative to cue onset in neurons significantly affected by cue onset measured over 100ms bins a transient change in firing rate was observed in the 100ms immediately after cue onset. Colour bar chart indicate firing rate (spikes/s) per bin. A & B lower panels: The dataset was analysed to find the time interval immediately after cue onset yielding the highest number of neurons that significantly changed their firing rate in response to cue onset, compared with baseline (3 seconds before cue onset). The highest number of significantly responding neurons was found when measuring the interval from 0 – 100ms after cue onset in all four trial types in both tasks.
Analysis of behaviour: Mean response rates to Go and NoGo tones as well as mean latency
to lever press in Hit and FA trials were calculated from all the sessions included in
electrophysiological analyses (sessions where tetrode tips were positioned outside the target
structure were excluded).
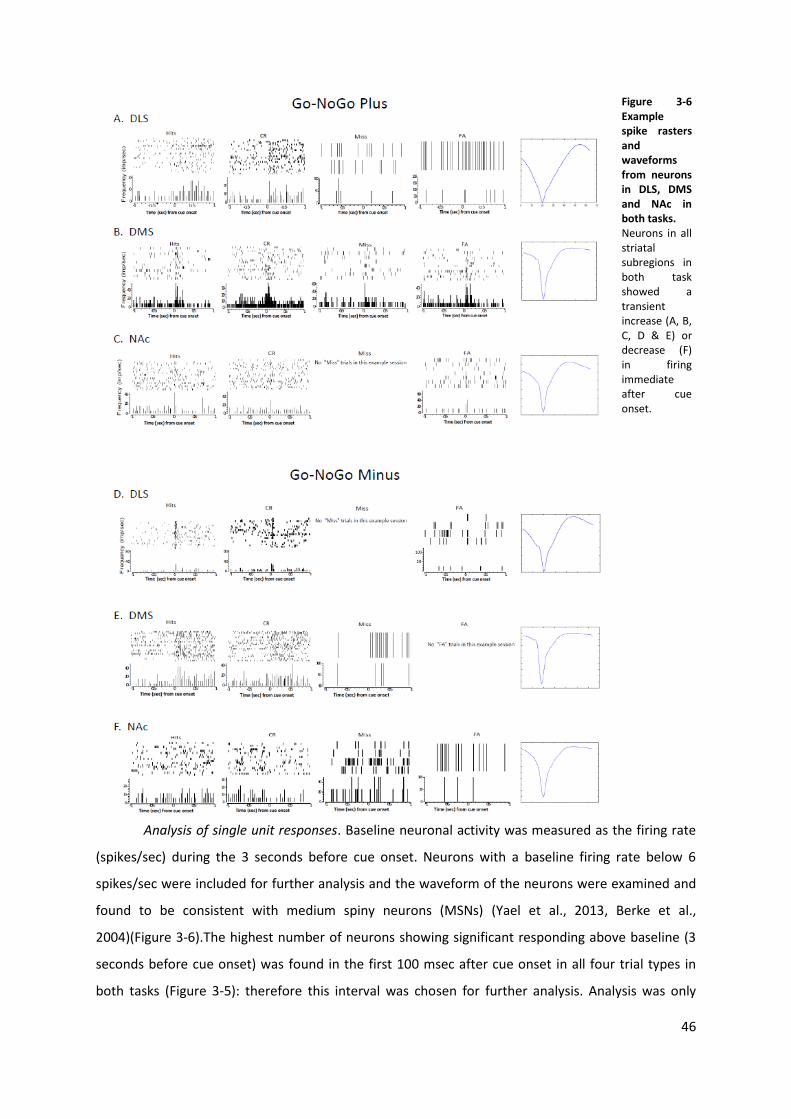
46
Figure 3-6 Example spike rasters and waveforms from neurons in DLS, DMS and NAc in both tasks. Neurons in all striatal subregions in both task showed a transient increase (A, B, C, D & E) or decrease (F) in firing immediate after cue onset.
Analysis of single unit responses. Baseline neuronal activity was measured as the firing rate
(spikes/sec) during the 3 seconds before cue onset. Neurons with a baseline firing rate below 6
spikes/sec were included for further analysis and the waveform of the neurons were examined and
found to be consistent with medium spiny neurons (MSNs) (Yael et al., 2013, Berke et al.,
2004)(Figure 3-6).The highest number of neurons showing significant responding above baseline (3
seconds before cue onset) was found in the first 100 msec after cue onset in all four trial types in
both tasks (Figure 3-5): therefore this interval was chosen for further analysis. Analysis was only
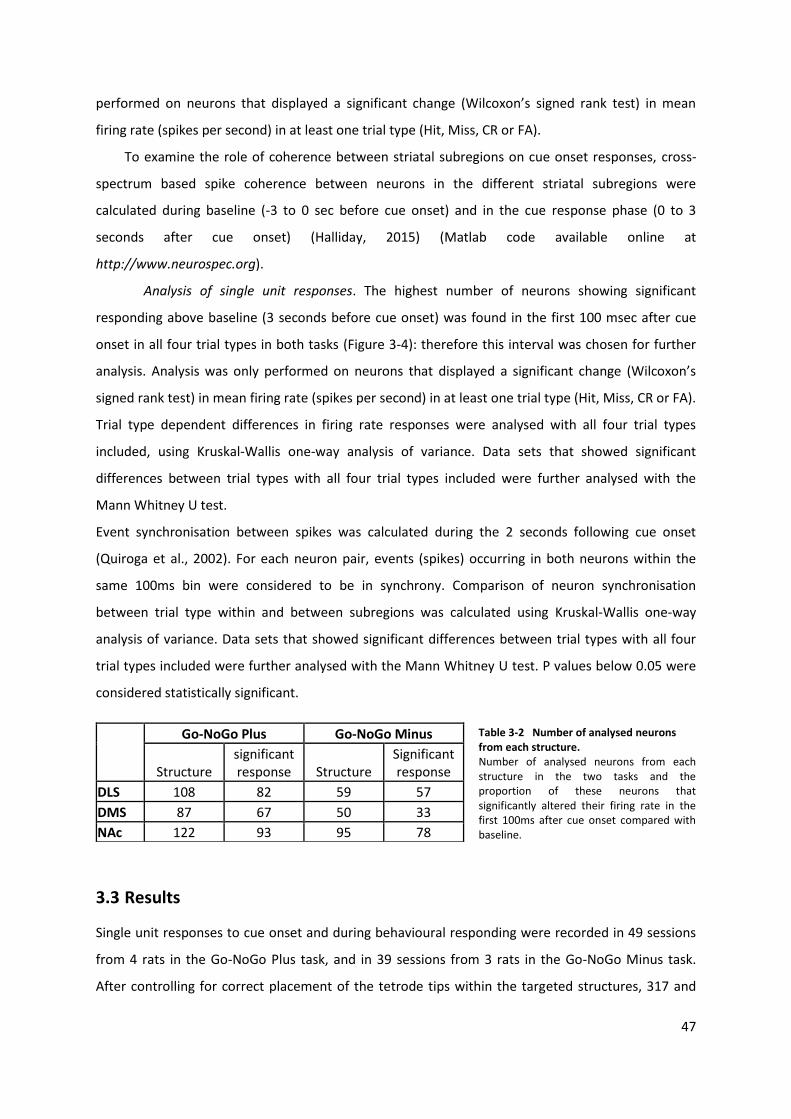
47
performed on neurons that displayed a significant change (Wilcoxon’s signed rank test) in mean
firing rate (spikes per second) in at least one trial type (Hit, Miss, CR or FA).
To examine the role of coherence between striatal subregions on cue onset responses, cross-
spectrum based spike coherence between neurons in the different striatal subregions were
calculated during baseline (-3 to 0 sec before cue onset) and in the cue response phase (0 to 3
seconds after cue onset) (Halliday, 2015) (Matlab code available online at
http://www.neurospec.org).
Analysis of single unit responses. The highest number of neurons showing significant
responding above baseline (3 seconds before cue onset) was found in the first 100 msec after cue
onset in all four trial types in both tasks (Figure 3-4): therefore this interval was chosen for further
analysis. Analysis was only performed on neurons that displayed a significant change (Wilcoxon’s
signed rank test) in mean firing rate (spikes per second) in at least one trial type (Hit, Miss, CR or FA).
Trial type dependent differences in firing rate responses were analysed with all four trial types
included, using Kruskal-Wallis one-way analysis of variance. Data sets that showed significant
differences between trial types with all four trial types included were further analysed with the
Mann Whitney U test.
Event synchronisation between spikes was calculated during the 2 seconds following cue onset
(Quiroga et al., 2002). For each neuron pair, events (spikes) occurring in both neurons within the
same 100ms bin were considered to be in synchrony. Comparison of neuron synchronisation
between trial type within and between subregions was calculated using Kruskal-Wallis one-way
analysis of variance. Data sets that showed significant differences between trial types with all four
trial types included were further analysed with the Mann Whitney U test. P values below 0.05 were
considered statistically significant.
Go-NoGo Plus Go-NoGo Minus
Structure significant response Structure
Significant response
DLS 108 82 59 57
DMS 87 67 50 33
NAc 122 93 95 78
Table 3-2 Number of analysed neurons from each structure. Number of analysed neurons from each structure in the two tasks and the proportion of these neurons that significantly altered their firing rate in the first 100ms after cue onset compared with baseline.
3.3 Results
Single unit responses to cue onset and during behavioural responding were recorded in 49 sessions
from 4 rats in the Go-NoGo Plus task, and in 39 sessions from 3 rats in the Go-NoGo Minus task.
After controlling for correct placement of the tetrode tips within the targeted structures, 317 and

48
204 neurons recorded during performance of the Go-NoGo Plus and Go-NoGo Minus task,
respectively, were included for further analysis (Table 3-2). Behaviour
All rats trained in either the Go-NoGo Plus or the Go-NoGo Minus task successfully learned to
discriminate between the Go and NoGo tone and maintained a high average level of discrimination,
i.e. Go trial Hit rate above 0.75 and NoGo trial FA rate below 0.25, until the end of the experiment
(Figure 3-7A and B). Latency to lever press was not normally distributed and variance and sample
size was different between Hit and FA trials, thus analysis was performed using non-parametric
tests. Mean latency to lever press was longer in FA trials than Hit trials in both tasks (p < 0.001
Mann-Whitney U test) (Figure 3-7C and D).
Examining the distribution of response latencies (Figure 3-7E and F) revealed that Hit trials peaked
within the 1st second of lever presentation, whereas the distribution of response latencies in FA
trials was spread out throughout the 4 second response interval. A higher proportion of Hit
responses were performed during the first second of lever presentation in the Go-NoGo Minus
(73.9%) compared to the Go-NoGo Plus task (43.9%) and comparison between the tasks showed
latency to respond in Hit trials to be significantly different (p < 0.001, Mann-Whitney U). Comparison
between tasks showed no significant difference in latency to respond in FA trials (p = 0.362, Mann-
Whitney U).
3.3.1 Firing rate responses
Intervals of increasing duration after cue onset were analysed to find the interval yielding the
highest number of neurons that significantly changed their firing rate in response to cue onset,
compared with baseline (3 seconds before cue onset). The analysed interval ran from 0- 100ms after
cue onset to 0-4sec after cue onset, in incremental steps of 50ms. The highest number of
significantly responding neurons was found in the first 100 msec after cue onset in all four trial types
in both tasks (Figure 3-5). A transient change in firing rate was also observed shortly after lever
presentation (4 seconds after cue onset) (Figure 3-5). This time interval was subject to a lot of
electrical artefact caused by the rat touching the lever and the pellet magazine, particularly in Hit
and FA trials. Therefore, analysis of spikes isolated from this time interval is unlikely to give a true
representation of neuronal responses to lever presentation. Thus, analysis was limited to the time
interval immediately after cue onset. Neurons showing a significant change in firing rate within the
first 100ms after cue onset, in at least one trial type, were selected for further analysis of effect of
trial type (Table 3-2). Significantly responding neurons were divided into two groups depending on
whether they increased or decreased their firing in response to cue onset. Consecutive 100 msec
windows were examined for effect of trial type during the first 4 sec of cue presentation (before
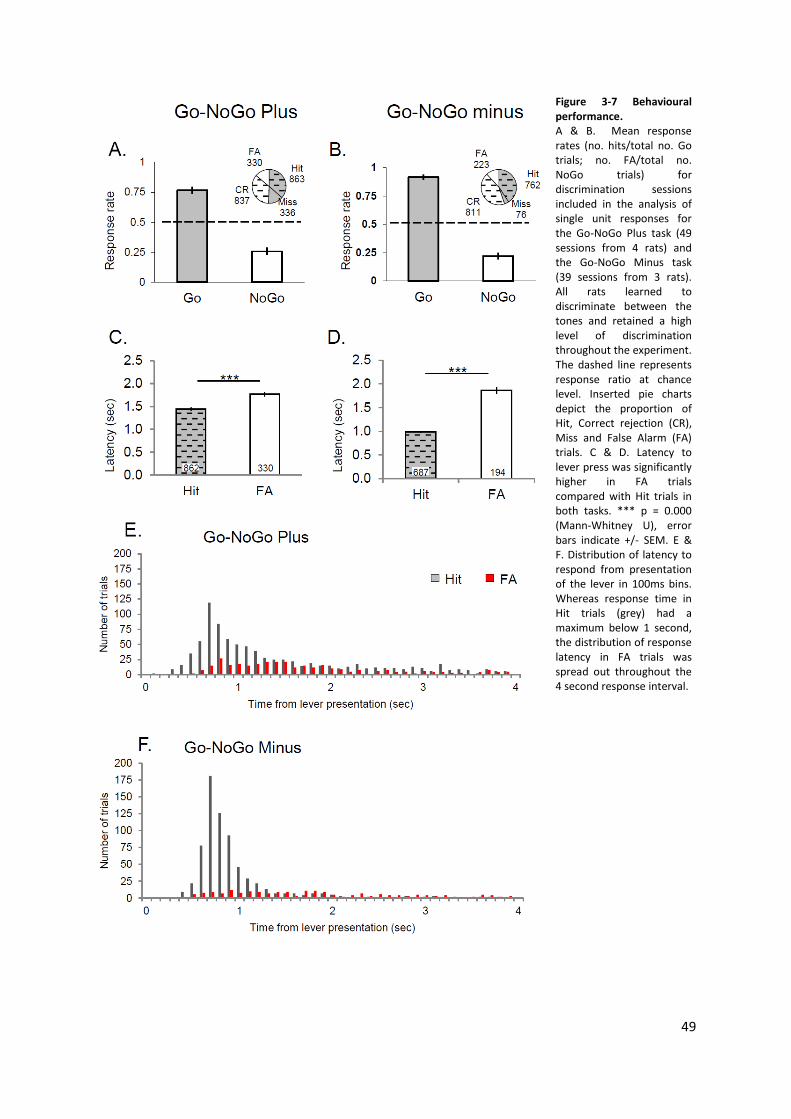
49
Figure 3-7 Behavioural performance. A & B. Mean response rates (no. hits/total no. Go trials; no. FA/total no. NoGo trials) for discrimination sessions included in the analysis of single unit responses for the Go-NoGo Plus task (49 sessions from 4 rats) and the Go-NoGo Minus task (39 sessions from 3 rats). All rats learned to discriminate between the tones and retained a high level of discrimination throughout the experiment. The dashed line represents response ratio at chance level. Inserted pie charts depict the proportion of Hit, Correct rejection (CR), Miss and False Alarm (FA) trials. C & D. Latency to lever press was significantly higher in FA trials compared with Hit trials in both tasks. *** p = 0.000 (Mann-Whitney U), error bars indicate +/- SEM. E & F. Distribution of latency to respond from presentation of the lever in 100ms bins. Whereas response time in Hit trials (grey) had a maximum below 1 second, the distribution of response latency in FA trials was spread out throughout the 4 second response interval.

50
lever extension). The highest number of neurons showing significant responding above baseline (3
seconds before cue onset) was found in the first 100ms after cue onset in all four trial types in both
tasks (Figure 3-5). Therefore, this interval was chosen for further analysis.
3.3.2 Baseline firing rates
As expected, baseline activity did not differ between trial types in either of the tasks in neither
excited neurons (F(3,381)= 0.559, p = 0.642 (Go-NoGo Plus)) and F(3,181)= 1.020, p=0.385 (Go-NoGo
Minus)) nor inhibited neurons (F(3,491)= 2.041, p= 0.107 (Go-NoGo Plus)) and F(3,337)=1.658,
p=0.176 (Go-NoGo Minus)) (Appendix – Table 1). However, in both tasks baseline firing rate
significantly differed between the striatal subregions. In the Go-NoGo Plus task, NAc exhibited a
higher baseline firing rate than DMS in neurons excited by cue onset (F(2,381)=6.374, p=0.002 -
p<0.001 (Post Hoc LSD)), and higher baseline firing rate than both DMS and DLS in neurons inhibited
by cue onset (F(2,491)=8.212, p<0.001 (ANOVA) - p<0.02(Post Hoc LSD)). In the Go-NoGo Minus,
baseline firing rate was higher in DLS than NAc in neurons excited by cue (F(2,181)=3.076, p=0.049 –
p=0.002 (Post Hoc LSD)), whereas NAc displayed a higher baseline firing rate than DMS in neurons
inhibited by cue onset (F(2,337)=3.395, p=0.035 – p=0.035 (Post Hoc LSD)). Firing rate response to
cue onset
Examining the firing rate in the first 100ms following cue onset suggested that firing rate responses
were greater in response to cue onset in trials where the rat subsequently produced an incorrect
behavioural response (Miss and FA) than in trials where the rat subsequently produced an correct
behavioural response (Hit and CR) (Figure 3-8) .
Go-NoGo Plus: In the Go-NoGo Plus task this pattern was particularly clear in inhibited
neurons which displayed significantly greater reduction of firing rate in error trials than correct trials.
Statistical analysis confirmed that both excited (F(3,381)=6.452, p<0.001) and inhibited
(F(3,491)=8.644, p<0.001) neurons responded differently to cue onset depending on trial type
(Figure 3-8A). Pairwise post hoc analysis further revealed that inhibited neurons, indeed, showed a
stronger decrease in firing rate to cue onset in error trials (Miss and FA) than correct trials (Hit and
CR) (p<0.002). Excited neurons displayed a significantly greater response in FA than CR trials
(p=0.038) and also a greater response in Miss trials than in other trial types (p<0.043).
The firing rate response to cue onset did not differ significantly between subregions in
excited neurons (F(2381)=2.494, p=0.084), but a significantly stronger inhibition was observed in
inhibited neurons in NAc compared with inhibited neurons in DMS (F(2,491)=6.820, p=0.001)
(Appendix – Table 2).
Go-NoGo Minus: A significant difference between trial type in response to cue onset was
found in both excited (F(3,181)= 9.108, p<0.001) and inhibited neurons (F(3,337)=10.376, p<0.001),
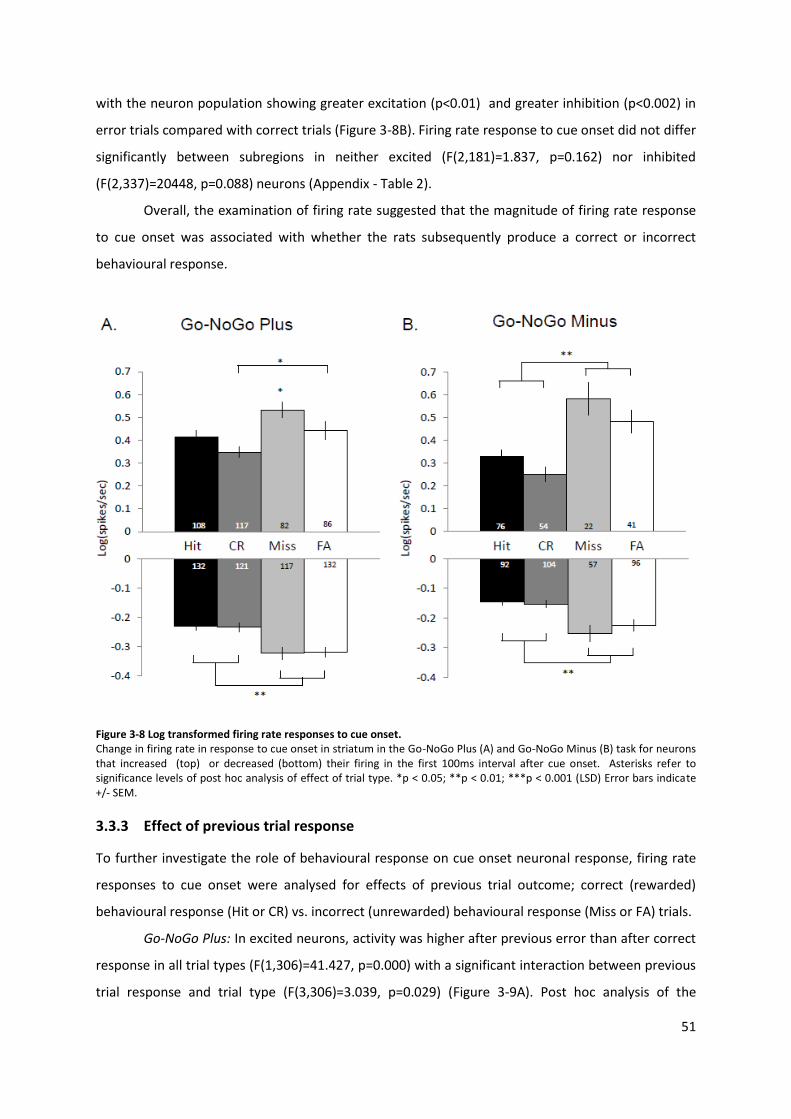
51
with the neuron population showing greater excitation (p<0.01) and greater inhibition (p<0.002) in
error trials compared with correct trials (Figure 3-8B). Firing rate response to cue onset did not differ
significantly between subregions in neither excited (F(2,181)=1.837, p=0.162) nor inhibited
(F(2,337)=20448, p=0.088) neurons (Appendix - Table 2).
Overall, the examination of firing rate suggested that the magnitude of firing rate response
to cue onset was associated with whether the rats subsequently produce a correct or incorrect
behavioural response.
Figure 3-8 Log transformed firing rate responses to cue onset. Change in firing rate in response to cue onset in striatum in the Go-NoGo Plus (A) and Go-NoGo Minus (B) task for neurons that increased (top) or decreased (bottom) their firing in the first 100ms interval after cue onset. Asterisks refer to significance levels of post hoc analysis of effect of trial type. *p < 0.05; **p < 0.01; ***p < 0.001 (LSD) Error bars indicate +/- SEM.
3.3.3 Effect of previous trial response
To further investigate the role of behavioural response on cue onset neuronal response, firing rate
responses to cue onset were analysed for effects of previous trial outcome; correct (rewarded)
behavioural response (Hit or CR) vs. incorrect (unrewarded) behavioural response (Miss or FA) trials.
Go-NoGo Plus: In excited neurons, activity was higher after previous error than after correct
response in all trial types (F(1,306)=41.427, p=0.000) with a significant interaction between previous
trial response and trial type (F(3,306)=3.039, p=0.029) (Figure 3-9A). Post hoc analysis of the
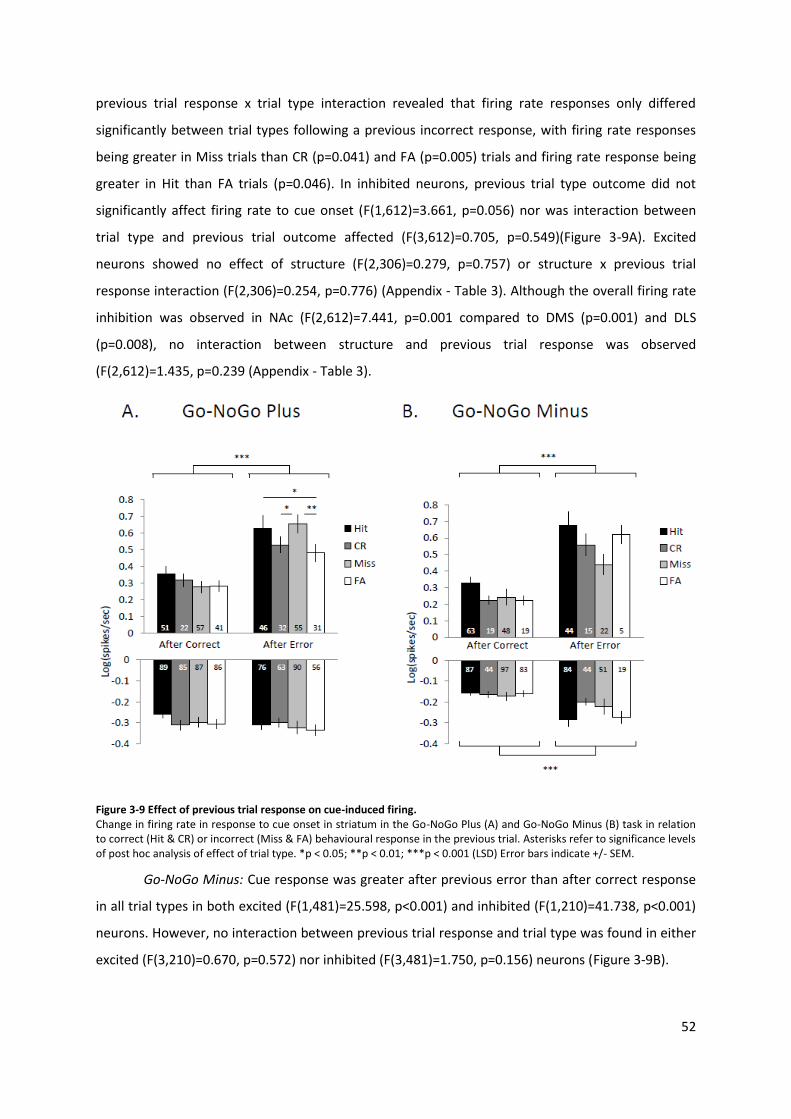
52
previous trial response x trial type interaction revealed that firing rate responses only differed
significantly between trial types following a previous incorrect response, with firing rate responses
being greater in Miss trials than CR (p=0.041) and FA (p=0.005) trials and firing rate response being
greater in Hit than FA trials (p=0.046). In inhibited neurons, previous trial type outcome did not
significantly affect firing rate to cue onset (F(1,612)=3.661, p=0.056) nor was interaction between
trial type and previous trial outcome affected (F(3,612)=0.705, p=0.549)(Figure 3-9A). Excited
neurons showed no effect of structure (F(2,306)=0.279, p=0.757) or structure x previous trial
response interaction (F(2,306)=0.254, p=0.776) (Appendix - Table 3). Although the overall firing rate
inhibition was observed in NAc (F(2,612)=7.441, p=0.001 compared to DMS (p=0.001) and DLS
(p=0.008), no interaction between structure and previous trial response was observed
(F(2,612)=1.435, p=0.239 (Appendix - Table 3).
Figure 3-9 Effect of previous trial response on cue-induced firing. Change in firing rate in response to cue onset in striatum in the Go-NoGo Plus (A) and Go-NoGo Minus (B) task in relation to correct (Hit & CR) or incorrect (Miss & FA) behavioural response in the previous trial. Asterisks refer to significance levels of post hoc analysis of effect of trial type. *p < 0.05; **p < 0.01; ***p < 0.001 (LSD) Error bars indicate +/- SEM.
Go-NoGo Minus: Cue response was greater after previous error than after correct response
in all trial types in both excited (F(1,481)=25.598, p<0.001) and inhibited (F(1,210)=41.738, p<0.001)
neurons. However, no interaction between previous trial response and trial type was found in either
excited (F(3,210)=0.670, p=0.572) nor inhibited (F(3,481)=1.750, p=0.156) neurons (Figure 3-9B).

53
Excited neurons displayed no significant difference between the two tasks (F(1,516)=0.024,
p=0.878) nor interaction between task and previous trial response (F(1,516)=2.495, p=0.115).
However, cue induced inhibition was significantly different between the two tasks
(F(1,1093)=38.537, p<0.001) and there was an interaction between task and previous trial response
(F(1,1093)=6.697, p=0.010). Post hoc analysis revealed that cue induced inhibition was greater in the
Go-NoGo Plus task than the Go-NoGo Minus task both after previous error (p=0.021) and previous
correct (p<0.001) trials (Figure 3-9B). Excited neurons showed no effect of structure
(F(2,210)=1.177), p=0.310) or previous trial response x structure interaction (F(2,210)=2.007,
p=0.137) (Appendix - Table 3). Likewise, no effect of structure (F(2,481)=1.728), p=0.120) or previous
trial response x structure interaction (F(2,481)=1.432, p=0.240) was found in inhibited neurons
(Appendix - Table 3).
Overall, firing rate response to cue onset was greater after previous incorrect response than
after previous correct response in both the Go-NoGo Plus and Go-NoGo Minus task.
3.3.4 Coherence between striatal subregions
Coherence between pairs of neurons in different striatal subregions was analysed during the
baseline (-3 – 0 seconds before cue onset) and the cue response period (0 – 3 seconds after cue
onset).
Go-NoGo Plus: Baseline coherence was strongly affected by trial type (F(3,2237)=81.421,
p<0.001), structure (F(2,2237)=6.944, p=0.001), and trial type x structure interaction (F(6,2237)=
8.052, p<0.001) (Figure 3-10A). Baseline coherence between NAc and DMS was significantly greater
before Miss trials than all other trial types (p<0.001, LSD post hoc) and greater before FA trials than
before Hit and CR (p<0.001, LSD post hoc), whereas baseline coherence was only significantly
greater before CR than Hit trials (p=0.019, LSD post hoc) (Figure 3-10A). Baseline coherence between
NAc and DLS was significantly greater before trials with incorrect behavioural response (Miss and FA)
than before trials with correct behavioural response (Hit and CR) (p<0.001, LSD post hoc) (Figure 3-
10A). Baseline coherence between DMS and DLS was significantly greater before Miss trials than all
other trial types (p<0.001, LSD post hoc) (Figure 3-10A).
Go-NoGo Minus: Baseline coherence was strongly affected by trial type (F(3,1120)=49.368,
p<0.001), structure (F(2,1120)=12.607, p=0.001), and trial type x structure interaction (F(6,1120)=
8.031, p<0.001) (Figure 3-10B). Baseline coherence between NAc and DMS was greater before FA
trials than all other trial types (p<0.013, LSD post hoc) and greater before Miss trials than before Hit
and CR (p<0.004, LSD post hoc) (Figure 3-10B). Baseline coherence between NAc and DLS was
significantly greater before trials with incorrect behavioural response (Miss and FA) than before
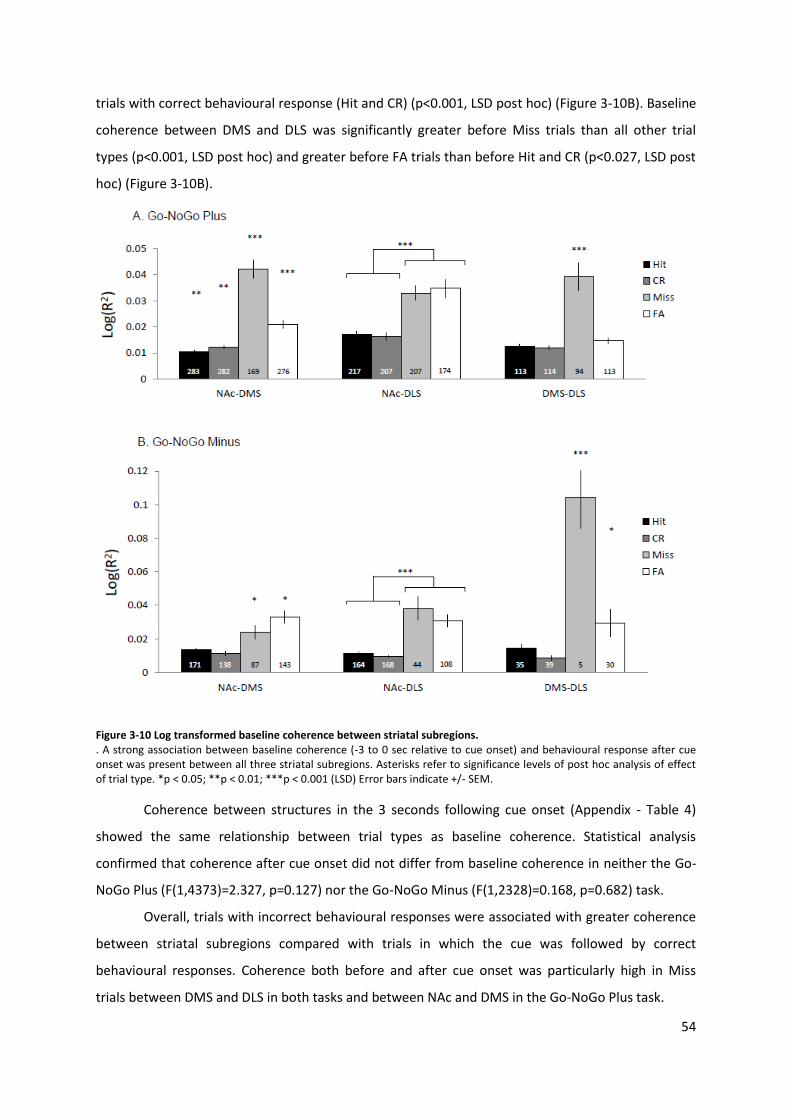
54
trials with correct behavioural response (Hit and CR) (p<0.001, LSD post hoc) (Figure 3-10B). Baseline
coherence between DMS and DLS was significantly greater before Miss trials than all other trial
types (p<0.001, LSD post hoc) and greater before FA trials than before Hit and CR (p<0.027, LSD post
hoc) (Figure 3-10B).
Figure 3-10 Log transformed baseline coherence between striatal subregions. . A strong association between baseline coherence (-3 to 0 sec relative to cue onset) and behavioural response after cue onset was present between all three striatal subregions. Asterisks refer to significance levels of post hoc analysis of effect of trial type. *p < 0.05; **p < 0.01; ***p < 0.001 (LSD) Error bars indicate +/- SEM.
Coherence between structures in the 3 seconds following cue onset (Appendix - Table 4)
showed the same relationship between trial types as baseline coherence. Statistical analysis
confirmed that coherence after cue onset did not differ from baseline coherence in neither the Go-
NoGo Plus (F(1,4373)=2.327, p=0.127) nor the Go-NoGo Minus (F(1,2328)=0.168, p=0.682) task.
Overall, trials with incorrect behavioural responses were associated with greater coherence
between striatal subregions compared with trials in which the cue was followed by correct
behavioural responses. Coherence both before and after cue onset was particularly high in Miss
trials between DMS and DLS in both tasks and between NAc and DMS in the Go-NoGo Plus task.

55
3.3.5 Differences between tasks
Although trial type did not affect baseline firing rates in neither the Go-NoGo Plus nor the Go-NoGo
Minus task, baseline firing rate did differ between tasks, with baseline firing rate being significantly
greater in the Go-NoGo Plus than in the Go-NoGo Minus task both in neurons excited by cue onset
(F(1, 562)=20.414, p<001) and neurons inhibited by cue onset (1, 828)=43.170. p<001).
In excited neurons, firing rate response to cue onset did not differ significantly between the
two tasks (F(1,562)=0.607, p=0.436). However, in inhibited neurons a greater reduction in firing rate
in response to cue onset was observed in the Go-NoGo Plus than the Go-NoGo Minus task
(F(1,828)=30.732. p<0.001). Similarly, when examining the role of task on the effect of previous trial
response, no effect of task (F(1,559)=36.331, p=0.853) or was found in excited neurons, whereas
significant differences in firing rate was found in inhibited neurons (F(1,1136)=77.846, p<0.001).
Baseline coherence was also significantly higher in the Go-NoGo Plus than the Go-NoGo
Minus task (F(1,3294)=22.327, p<0.001), and interaction between task, trial type and structure was
also highly significant (F(6,3294)=11.597, p<0.001).
In summary, the rats in the two tasks displayed significantly different baseline firing rates.
This task effect persisted after cue onset in inhibited neurons and was also manifested in differences
in striatal coherence between the two tasks.
3.4 Discussion
3.4.1 Behaviour
All rats maintained a high level of accuracy in their responses after tetrode implantation in both
tasks, with response rate in Go trials above 75% and NoGo trial response rate below 25% (Figure 3-
5), suggesting that rats were able to learn and retain stimulus-response-outcome contingencies and
to switch successfully between them in both tasks. In both tasks latency to respond was significantly
longer in NoGo trials than Go trials (Figure 3-7C-E). This observation is consistent with previous
studies using the standard Go-NoGo paradigms (Harding et al., 2004, Curzon et al., 1999). Latencies
to respond in Go trials were shorter in the Go-NoGo Minus task than the Go trial latencies in the Go-
NoGo Plus task, whereas no difference in latencies in the NoGo trials were observed between the
Go-NoGo Plus and Go-NoGo Minus task. In the Go-NoGo Minus task, a reward could only be won in
Go trials, as correct suppression of lever press in the NoGo trials had no programmed consequence.
Incorrect lever press to the NoGo cue in the Go-NoGo Minus only resulted in a mild error cue (a one

56
minute timeout), whereas incorrect lever press to the NoGo cue in the Go-NoGo Plus resulted in the
loss of a sugar pellet as well as the timeout. Therefore, pressing the lever in the Go-NoGo Minus may
be perceived as having a lesser adverse consequence than in the Go-NoGo Plus task, which could
create a stronger bias to lever press in the rats trained in the Go-NoGo Minus task compared with
the rats trained in the Go-NoGo Plus task. Interestingly, there was no significant difference between
tasks in latency to respond in FA trials and latency to respond was shorter in Hit compared to FA
trials in both tasks. This suggests that the differences in latency between trial types and between
tasks were not due to deficits in memory or cognitive processing, the rats in both tasks were able to
distinguish between the two stimuli-response contingencies (as suggested by the high level of
accuracy in both tasks). Rather, the rats were less likely to suppress motor initiation to Go cue in the
Go-NoGo Minus than the Go-NoGo Plus task. Studies in monkeys and humans performing a stop-
signal task have showed that movements were initiated if and only if the neural activity in motor
cortex reached a certain activation level. In the majority of neurons recorded in rhesus monkeys,
activity was less likely to reach the necessary threshold in trials where the monkey suppressed
movement compared to trials where movement was initiated (Hanes and Schall, 1996). An fMRI
study in humans showed that efficient response inhibition was associated with greater activation of
inhibitory motor areas in frontal cortex and were negatively correlated with stop-signal reaction
times (Li et al., 2006). This process has been modelled as an interactive race between “go” and
“stop” neurons, which interact though inhibitory connections to control motor initiation in response
to learned stop cues (Boucher et al., 2007, Verbruggen and Logan, 2009, Schall and Godlove, 2012).
Upon hearing the cue tone, the rats in the current study have to decide to either initiate or suppress
motor response. Due to the difference in reward contingencies between the Go-NoGo Plus and Go-
NoGo Minus task, the rats trained in Go-NoGo Plus task stand to lose more than those trained in the
Go-NoGo Minus task, if they incorrectly press the lever. The shorter response latency in Go trial in
the Go-NoGo Minus task may reflect stronger contribution from motor inhibiting units as suggested
in the interactive race model (Boucher et al., 2007, Verbruggen and Logan, 2009), leading to longer
response latencies in the Go-NoGo Plus task.
3.4.2 Baseline single unit activity
Although trial type did not affect baseline firing rates in either of the two tasks, baseline firing rate
was significantly greater in the Go-NoGo Plus than in the Go-NoGo Minus task (Appendix - Table 1)
and this task-effect persisted after cue onset in inhibited neurons. The difference in cue-induced
inhibition is likely influenced by the overall difference in baseline firing which makes comparison of
task effects on cue-induced neuronal responses difficult to interpret. Therefore, the following

57
discussion will mainly focus on trial type and structure dependent differences in firing rate response
within each task. The differences in baseline firing rates may be due to individual differences
between the rats (Cohen, 2007) which have become apparent because of the low sample size.
However, examining the impact of individual differences in neuronal responses falls outside the
scope of the current study.
3.4.3 Single unit responses to cue onset
Overall, the examination of firing rate suggested that the magnitude of firing rate responses to cue
onset in both tasks were associated with whether the rats subsequently produced a correct or
incorrect behavioural response, regardless of whether the response required motor initiation or
suppression. (Figure 3-8). This pattern was particularly clear in the Go-NoGo Minus task, where both
the excitatory and inhibitory response to cue onset was greater in error trials compared with trials
where the rat subsequently responded correctly, as well as in the inhibitory response in the Go-
NoGo Plus task, where neurons displayed significantly greater reduction of firing rate in response to
cue onset in error trials than correct trials.
Previous research in rats has shown that discrimination training in at T-maze paradigm
causes changes in firing patterns in dorsal striatum (Barnes et al., 2011, Barnes et al., 2005). During
the early stages of training, firing rate response to cues signalling location of the reward increased,
but as the rats became overtrained on the task the firing rate response shifted towards, and peaked
at, onset of motor response following the initial cue (Barnes et al., 2011, Barnes et al., 2005). These
paradigms required the rat to always initiate a motor response, and therefore, cannot elaborate on
the role of motor initiation vs. motor suppression. However, they suggest that as a task is learned
and executed with a high level of accuracy, the neuronal ensembles involved in the initial acquisition
of specific elements of the task decrease their activity to cue onset, which could account for the
lower firing rate response observed in the correct trials in the current study. Cue onset in error trials,
where the rat’s behaviour contradicts the overtrained correct stimulus-response contingency, were
associated with greater firing rate response compared with correct trials. This increase in activity
may be an effect of modulation from upstream projection areas such as medial prefrontal cortex
(mPFC). In a conditional fear paradigm, mPFC neuron that responded to initiation or inhibition of
movement showed tonically elevated activity already before the conditioned stimulus, suggesting
that tonic firing rates in these mPFC neurons may bias the rat’s choice to either initiate or inhibit
movement (Halladay and Blair, 2015). In addition, some laboratory rodents trained to run through a
maze for reward have been observed to continue to examine alternative, never rewarded, routes
through the maze, even after the task has been learned (Coppens et al., 2010, Benus et al., 1990).

58
This behaviour is considered a characteristic of a reactive coping style (Coppens et al., 2010, Benus
et al., 1990), which may seem counterproductive in a laboratory setting, when stimulus-response-
outcome contingencies are kept constant, but would be adaptive in a natural environment where
stimuli and outcome may be less constant. It is possible that the comparative increase in firing rate
response in error trials, may reflect a switch from the overtrained habitual response to more goal-
directed approach, like in the early stages of acquisition, in order for the rats to test learned
stimulus-response contingencies. A recent fMRI study in humans performing a rule learning and set-
shifting task, found that whereas activity in ventral striatum, increased in the first trial after a rule
switch had been announced, the “hypothesis testing” phase (after the subject had received error
cues to indicate the direction of the new rule) was associated with increased activity in the posterior
dorsal parts of striatum (Liu et al., 2015), further suggesting that testing known stimulus-response-
outcome contingencies is associated with increased activity in distinct striatal regions.
Although single unit cue responses in the Go-NoGo Minus task and inhibitory cue responses
in the Go-NoGo Plus task were associated with whether the rats subsequently produced a correct or
incorrect behavioural response, regardless of whether the response required motor initiation or
suppression, this pattern was not as clear in excited neurons in the Go-NoGo Plus task. In the Go-
NoGo Plus task, cue induced excitation was greater in Miss trials compared to all other trial types
and excitation in FA trials was only significantly greater than in CR trials (Figure 3-8A). Previous work
has shown that the majority of neurons in DLS are movement-related and change their firing rate in
response to repeated training on movement-dependent task (Tang et al., 2007, 2009). However,
lesion studies also implicate NAc and DMS in the initiation and vigour of movement (Pisa and
Schranz, 1988, Wang et al., 2013, Hart et al., 2014). In the Go-NoGo Plus task in the current study, a
similar excitatory response was observed in Hit and FA trials, and the smallest and largest excitatory
response was found in CR and Miss trials, respectively. Whereas Hit and FA trials involved
movement, no movement was required in Miss and CR trials. A possible explanation for the
observed excitatory responses may be that in overtrained rats, striatal neurons entrained on
movement mask the effect of reward expectation or processing involved in behavioural choice. In
rats trained in a three tone Go-NoGo task, where tone onset signalled the rat to either go left, right
or stay immobile to win a reward, dopamine release in NAc was attenuated until movement was
initiated, suggesting that dopamine release within this structure was triggered by movement rather
than reward expectation (Syed et al., 2016). However, more research is needed to understand the
interaction of reward expectation, motor preparation and behavioural choice on single neuron
responses within the striatum.

59
Interestingly, cue evoked responses appear highly sensitive to variation in Go-NoGo
paradigms as well as the duration of the analysed cue response period. When analysing the first
1000 ms after cue onset in a Go-NoGo paradigm where both Hit and CR responses were rewarded
(similar to the Go-NoGo Plus task in the current study), NAc single unit showed greater increase in
excited units and a smaller increase in inhibited units in CR and Miss trials (where the rats withheld
lever press) than in Hit and FA trials (where the rats pressed the lever) (Roitman and Loriaux, 2014).
The findings in this study suggest that cue responses correlate with initiation of motor response
rather than outcome expectancy. However, another study analysed NAc single unit responses during
the first 500 ms after cue onset in a traditional Go-NoGo paradigm (where only Hit responses were
rewarded – comparable to the Go-NoGo Minus task in the current study) and this study found the
greatest excitation and inhibition in Hit trials compared with other trial types (Nicola et al., 2004a).
Because only correct responses in Go trials yielded a reward in this study, the greater firing rate
response in Hit trials suggests that cue responses are influenced by both motor initiation and reward
expectation. In agreement with the findings of Roitman and Loriaux (2014) the current study found
greater excitation after cue onset in Miss than Hit trials and greater inhibition in FA than CR trials in
both tasks and greater excitation in Miss than FA trials in the Go-NoGo Plus task. However,
excitation was also greater FA than CR in both tasks in the current study whereas Roitman and
Loriaux (2014) reported the opposite. Equally, in agreement with the findings of Nicola et al. (2004a)
the current study found greater excitation in Miss and Fa trial compared with CR trials and no
significant difference between excitation in Hit and CR trials and inhibition in Miss and FA trials in
both tasks as well as no significant difference between Miss and FA trials in the Go-NoGo Minus task.
In contrast, the current study found greater excitation in Miss than Hit trials and greater inhibition in
Miss and FA trials compared with Hit trials in both tasks, where Nicola et al. (2004a) observed the
opposite. Whereas Roitman and Loriaux (2014) and Nicola et al. (2004a) both found greater
inhibition in Hit than Miss trials, they found different relationship between all examined trial type
pairs in excited neurons. In both these studies, the lever was presented at the beginning of the trial,
simultaneously with the cue indicating whether the trial was a Go or NoGo trial, whereas in the
current study the Go vs NoGo trial discrimination cue was always presented four seconds before
presentation of the lever (Figure 1A). Loriaux (2014) and Nicola et al. (2004a) analysed the first
100ms and 500ms after cue onset whereas the current study focused on the first 100ms interval as
this interval yielded the largest number of significantly responding neurons. Together these three
studies underline how subtle differences in paradigms and analysis may in turn reveal different
aspects of neuronal responses associated with behaviour.

60
3.4.4 Recent behavioural experience predicts neuronal response to cue
To further investigate the role of behavioural response on cue onset neuronal response, firing rate
responses to cue onset were analysed for effects of previous trial outcome; correct (rewarded)
behavioural response (Hit or CR) vs. incorrect (unrewarded) behavioural response (Miss or FA) trials.
Excited neurons were found to display a significantly greater response to cue onset in trials following
incorrect response trials (Miss and FA) than correct response trials (Hit and CR) in both tasks (Figure
3-9). This observation was consistent across both tasks, regardless of whether correct rejection of
the NoGo cue was rewarded or not, suggesting that the neuronal response to cue onset were
modulated by recent behavioural experience rather than recently reward experience. Previous work
has also examined the role of previous response-outcome experience on neuronal responses in
striatum. In a decision making task in rats, DMS was found to code for action in previous trial during
action selection phase in current trial but not immediately after cue (Ito and Doya, 2015).In rats
trained on visual discrimination task, neurons in ventral striatum were found to modulate their
activity according to the rat’s actions in the previous trial (Kim et al., 2009). The ability to integrate
recent response-outcome experiences into the planning of future responses is imperative to
optimising behaviour in a changing environment. The observed increased firing rate response to cue
onset following error trials could indicate increased attention in trials following previous
unsuccessful behavioural response. Lesion of DMS decrease accuracy and increase response time in
attentional tasks suggesting that this structure is a vital component organisation of adaptive
behaviour particularly in task requiring flexible responding (Lindgren et al., 2013, Rogers et al.,
2001). Similarly, contralateral lesion of mPFC and NAc (Christakou et al., 2001) or contralateral
inactivation of PrL and DMS (Baker and Ragozzino, 2014) have been show to disrupt planning and
responding in tasks that require switching between multiple stimulus-outcome contingencies. A
recent study found greater firing rate response to cue onset in a directional choice task in rat frontal
cortex after previous correct trial than error trial (Yuan et al., 2015) suggesting that recent
experience is associated with firing rate response to cue onset in both cortex and striatum, although
with opposite effect.
Whereas cue onset in trials following previously correct response showed no differences
between trials in either of the tasks, in the Go-NoGo Plus task trials following previous error
responses were greater to Go cues compared with NoGo cues (Figure 3-9A), suggesting greater
striatal sensitivity to cues following recent incorrect and unrewarded responses which is consistent
with previous findings (Stalnaker et al., 2012, Oyama et al., 2015).

61
3.4.5 Striatal subregions collectively respond to cue onset
Whereas previous studies have recorded single units in NAc in rats performing Go/NoGo tasks
(Roitman and Loriaux, 2014, Nicola et al., 2004a), this study simultaneously recorded single units in
DMS, DLS and NAc to enable comparison of cue responses between these structures. In the current
study, NAc was found to have a greater baseline firing rate compared with dorsal striatum (Appendix
– Table 1) and this difference was maintained after cue onset in inhibited neurons in the Go-NoGo
Plus task, where NAc showed stronger inhibition than DMS (Appendix - Table 2). However, the
current study found no interaction of structure x trial type, suggesting that although overall firing
activity differed between NAc and dorsal striatum, the three striatal subregions did not differ in their
immediate response to cue onset in the current study.
Lesion studies suggest consistent functional differences between striatal subregions (Devan
et al., 2011, Balleine et al., 2009). However, task related information is likely integrated across the
striatal subregions in order to collectively organise adaptive behavioural output and the level of
functional segregation and coordination between subregions may depend on the nature and
complexity of the task (Liljeholm and O'Doherty, 2012). In the current study no structure dependent
differences in cue responses were observed. However, a recent study recording single units in DLS,
DMS and ventral striatum in rats during performance of a choice task showed DLS to respond more
strongly to cue onset than DMS and ventral striatum (Ito and Doya, 2015). In the above experiment
cues always required the rat to initiate a motor response and reward was delivered probabilistically,
whereas in the current study the rat had to either initiate or inhibit a motor response to successfully
complete a trial. Together, these studies indeed suggest that the striatal segregation depend on
distinct elements of the task such as motor requirement and outcome contingencies.
The similarity in cue response between the striatal subregions may suggest that the joint
striatal response to cue in the current task was coordinated by modulatory input from outside the
striatum. MPFC would be a likely candidate area to be exerting such higher level control. Strong
afferents from mPFC connect to both ventral and dorsal regions of striatum (Gabbott et al., 2005,
Groenewegen et al., 1999, Hart et al., 2014, Balleine et al., 2007, Balleine and O'Doherty, 2010,
Heidbreder and Groenewegen, 2003) and several studies have shown mPFC to be involved in the
organisation and planning of complex behaviour (Groenewegen and Uylings, 2000, Dalley et al.,
2004). The findings in this study further emphasise the relevance of recording activity in multiple
structures within relevant neuronal networks in behaving animals to fully understand the role of
these structures in modulating outcome oriented behaviour.

62
3.4.6 Coherence between striatal subregions
When examining the coherence between neurons in different subregions, coherence in both tasks
was found to vary significantly between trial types in the three seconds prior to cue onset (as well as
in the three seconds following cue onset) between neurons in the three striatal subregions (Figure 3-
10) although firing rates showed no effect of trial types during the same time interval. In freely
moving rats, theta band coherence within the striatum and between primary motor cortex and
striatum has been found to be greater during periods of wakefulness than when the rats were
resting (Lepski et al., 2012). In the current study, Miss trials (where the rats failed to respond to the
Go cue) were associated with greater pre-stimulus coherence between striatal subregions compared
with correct response trials (Figure 3-10). Increased coherence may reflect low levels of attention to
external stimuli, causing the rat to miss the cue (Gusnard and Raichle, 2001, Herzog et al., 2014).
Several studies in humans and monkeys have demonstrated that particularly alpha band coherence
between different cortical regions before cue onset affects stimulus detection (Boly et al., 2007,
Sadaghiani et al., 2010, Forstmann et al., 2010, Melloni et al., 2007, Shulman et al., 2002), with some
studies reporting greater coherence to be associated with low detection (Hanslmayr et al., 2007,
Mazaheri et al., 2011, van Dijk et al., 2008), and others report stronger coherence between
structures to predict high stimulus detection (Boly et al., 2007, Super et al., 2003). More recently, a
study in rats performing an auditory detection task found increased theta, alpha and beta band
coherence between frontal and parietal cortex before cue onset in trials where the rats failed to
detect the tone (Herzog et al., 2014), whereas another study in rats found increased phase
synchrony between single unit activity to local field potentials (LFPs) in prelimbic cortex and anterior
cingulate cortex before cue onset in Hit trials compared to Miss trials in a sustained visual attention
task (Totah et al., 2013). These studies use a variety of methods to quantify interaction between
neuronal populations and focus on different combinations of mainly cortical areas. While some
studies show that coherence between their target areas enhance stimulus detections, other studies
show that activity in their target areas decrease stimulus detection, they all demonstrate a
relationship between pre-stimulus coherence and stimulus detection. Fluctuation in activity and
network connectivity has long been linked to attentional state and previous work suggests that low
detection rates following high coherence likely represent functional inhibition within the target
network diverting attention away from external stimuli to focus attention on internal
representations such as working memory (Hanslmayr et al., 2007, Mazaheri et al., 2011, Cooper et
al., 2003, Gusnard and Raichle, 2001, van Dijk et al., 2008). Strong afferents from medial prefrontal
cortex (Gabbott et al., 2005, Groenewegen et al., 1999, Hart et al., 2014, Balleine et al., 2007,
Balleine and O'Doherty, 2010, Heidbreder and Groenewegen, 2003), motor and sensory cortex (Van

63
Waes et al., 2012, Redgrave et al., 2011), and thalamus connect to both ventral and dorsal regions of
striatum. Thus, the striatum is intricately connected to both task-positive and task-negative
networks implicated in the regulation of attention to external stimuli (Sadaghiani et al., 2010). The
high prestimulus coherence between striatal subregions observed in Miss may originate from
modulatory input from task-negative regions of cortex or thalamus.
Overall, prestimulus coherence was greater in FA trials as well as in Miss trials compared
with correct response trials, suggesting that coherence is associated with future behavioural choice.
However, coherence between NAc and DMS and between DMS and DLS differed significantly
between FA and Miss trials in both tasks, suggesting that striatal prestimulus coherence does not
merely code for future correct or incorrect behavioural responses (Figure 3-10). Furthermore, the
relationship between Miss and FA trial coherence also differed greatly between structure pairs, with
Miss coherence being higher than FA coherence between DMS and DLS but similar between NAc and
DLS in both tasks, whereas Miss trial coherence between NAc and DMS being higher than FA trial
coherence in the Go-NoGo Plus task but lower in the Go-NoGo Minus task. This suggests that
coherence between different striatal subregions contributes differently to the modulation of
behavioural choice. However, more research is needed to clarify the role of striatal coherence on
reward directed choice behaviour.
Retrograde and anterograde tracing studies suggest that the information flow within
striatum runs in a spiral from NAc through DMS to DLS (Haber et al., 2000). Coherence between NAc
and dorsal striatum may facilitate integration of motivational aspects associated with NAc function
(Tricomi and Lempert, 2015, Basar et al., 2010, Baldo and Kelley, 2007)with goal directed processes
(Shiflett et al., 2010, Shan et al., 2014, Yin et al., 2005, Balleine et al., 2009, Devan et al., 2011,
Hilario et al., 2012) in DMS and with motor processes behaviour (Devan et al., 2011, Costa et al.,
2004, Mowery et al., 2011, Schmitzer-Torbert et al., 2015, Fanelli et al., 2013, Yin et al., 2006) in DLS
in order to execute the task. Previous research suggests that both dorsal and ventral regions of
striatum contribute to the processing and execution of complex behaviour (Liljeholm and O'Doherty,
2012) and high trial type dependent coherence between the striatal subregions observed in the
current study further suggest that cue responses in this current task to be maintained by interaction
between striatal subregions. The high striatal prestimulus coherence observed in Miss trials is well in
accord with previous studies examining the role of cortical coherence and stimulus detection (Boly
et al., 2007, Sadaghiani et al., 2010, Forstmann et al., 2010, Melloni et al., 2007, Shulman et al.,
2002, Herzog et al., 2014) and the coherence between striatal regions may reflect coherence in
cortical areas projecting to the striatum, with mPFC being a prime candidate for the exertion of such
top down control (Riga et al., 2014, Balleine et al., 2009, Christakou et al., 2004, Stefanik et al., 2015,

64
Baker and Ragozzino, 2014, Thorn and Graybiel, 2014). Future research into the influence of mPFC
modulation on striatal processing during complex behavioural tasks may offer insight into the
precise role of cortico-striatal regulation of complex reward-directed behaviour.
3.4.7 Conclusions
Overall, firing rate response to cue onset was greater in error trials compared with trials with
subsequent correct response. This finding emphasises the importance of contribution of subject’s
choice when analysing effect of reward-paired cues on neuronal activity. We hypothesised that firing
rate response to CR trials compared to other trial types would be different between tasks, as this
was the only trial type for which the outcome differed between tasks. However, firing rate response
to cue onset did not appear to be influenced by differences in reward expectancy. Error responses
during performance of an overtrained task may signal trials in which the animal tests the consistency
of the learned stimulus response contingencies and thus engage striatal networks associated with
goal-directed rather than habitual behaviour. In both tasks, cue induced excitation and inhibition in
Miss trials were greater than in trials with correct behavioural response. However, more research is
needed to elucidate the role of motor- and reward related aspects of neuronal responses associated
with behaviour. Firing rate response to cue onset was significantly greater in trials following error
trials, which may signify increased attention in trials following previous unsuccessful behavioural
response.
Although overall firing rate differed between striatal structures, no interaction between
structure and trial type was found, suggesting that although overall firing activity differed between
NAc and dorsal striatum, the three striatal subregions did not differ in their immediate response to
cue onset. The similarity in cue response between the striatal subregions may suggest that the joint
striatal response to cue in the current task was coordinated by modulatory input from outside the
striatum.
Coherence between striatal subregions was found to be particularly high before and during
Miss trials. Fluctuations in prestimulus coherence between cortical regions have been linked to
attention and the high prestimulus coherence between striatal subregions observed in Miss trials
suggests that the striatum may originate from modulatory input from task-negative regions of cortex
or thalamus. Overall, prestimulus coherence was greater in FA trials as well as in Miss trials
compared with correct response trials, suggesting that coherence is associated with future
behavioural choice. However, the relationship between coherence in Miss and FA trials differed
significantly between structures and tasks, suggesting that striatal prestimulus coherence does not
merely code for future correct or incorrect behavioural responses and that the striatal subregions

65
contribute differently to the modulation of behavioural choice. However, more research is needed
to clarify the role of striatal coherence on reward directed choice behaviour.
Previous research suggests that both dorsal and ventral regions of striatum contribute to the
processing and execution of complex behaviour (Liljeholm and O'Doherty, 2012) and the current
study further suggest that cue responses in this task to be maintained by interaction between
striatal subregions. However, further research on the interaction between striatal subregions during
complex behaviour is warranted to illuminate the role of such interactions on reward-directed
choice behaviour.
The high striatal prestimulus coherence observed in Miss trials is well in accordance with
previous studies examining the role of cortical coherence and stimulus detection (Boly et al., 2007,
Sadaghiani et al., 2010, Forstmann et al., 2010, Melloni et al., 2007, Shulman et al., 2002, Herzog et
al., 2014) and the coherence between striatal regions may reflect coherence in cortical areas
projecting to the striatum, with mPFC being a prime candidate for the exertion of such top down
control (Riga et al., 2014, Balleine et al., 2009, Christakou et al., 2004, Stefanik et al., 2015, Baker and
Ragozzino, 2014, Thorn and Graybiel, 2014). Future research into the influence of mPFC modulation
on striatal processing during complex behavioural tasks may offer insight into the precise role of
cortico-striatal regulation of complex reward-directed behaviour.

66
Chapter 4: Corticostriatal contribution to reward-directed behaviour
4.1 Introduction
4.1.1 Prelimbic cortex and behavioural control
Medial Prefrontal cortex (mPFC) plays a crucial role in the organisation of previously acquired
information and in subsequent integration of this information into the planning and execution of
complex behaviour (Groenewegen and Uylings, 2000, Dalley et al., 2004). MPFC is thought to exert
an influence on appetitive behaviour (Riga et al., 2014) via top down control of downstream areas in
nucleus accumbens (NAc) (Riga et al., 2014, Balleine et al., 2009, Christakou et al., 2004, Stefanik et
al., 2015) and medial parts of dorsal striatum (Christakou et al., 2001, Baker and Ragozzino, 2014,
Thorn and Graybiel, 2014). Whereas infralimbic cortex (IL), in ventral mPFC, is associated with habit
formation (Maier, 2015, Smith and Graybiel, 2013), prelimbic cortex (PrL), in dorsal mPFC, is involved
in goal-directed behaviour and complex behaviour that requires flexible switching between different
context-dependent strategies (Riga et al., 2014, Heidbreder and Groenewegen, 2003, Funamizu et
al., 2015).
Lesion of PrL impairs acquisition but not expression of instrumental learning tasks (Ostlund
and Balleine, 2005, Tran-Tu-Yen et al., 2009, Dalley et al., 2004) and PrL inactivation does not impair
reward seeking behaviour (Burgos-Robles et al., 2013), suggesting that although PrL is involved in
the regulation of appetitive behaviour it may not be directly involved in reward seeking. PrL lesion
impairs choice accuracy, specifically by increasing perseverance errors (Dalley et al., 2004) and also
cause impairment in tasks with delayed response contingencies (Heidbreder and Groenewegen,
2003), suggesting that PrL is not directly involved in storing and maintaining information long term
but more likely contributes to organisation and planning of flexible behaviour, based on previously
acquired information (Dalley et al., 2004, Hart et al., 2014).
Several studies implicate PrL in the encoding of stimulus-response-outcome associations and
in successful switching between behavioural strategies depending on context (Mulder et al., 2003,
Hosking et al., 2015, Halladay and Blair, 2015, Moorman and Aston-Jones, 2015). When rats were
trained in a two-lever Go-NoGo task, PrL firing rate was significantly affected during correct lever
press after the light cue and subsequent entry into the reward receptacle (Mulder et al., 2003).
However, neither light cue alone, nor incorrect lever press or nose poke (which were never
rewarded) was associated with a change in firing rate (Mulder et al., 2003). This suggests that PrL
contribute to the encoding of stimulus-response-outcome associations and may be involved in the
formation and control of behavioural strategies (Mulder et al., 2003).

67
In a cognitive effort task in rats, where the rats had to choose between performing an low
effort task (with low attentional demands) for low reward and a high effort task (with high
attentional demands) for high reward, inhibition of PrL increased the choice of low effort task and
increased errors in both tasks. This suggests a role for PrL in attentional processes, particularly when
the task involves a choice between behavioural strategies (Hosking et al., 2015). In a conditional fear
paradigm, mPFC neurons that responded to initiation or inhibition of movement showed tonically
elevated activity already before the conditioned stimulus, suggesting that tonic firing rates in these
mPFC neurons may bias the rat’s choice to either initiate or inhibit movement (Halladay and Blair,
2015), further implicating PrL in the planning and execution of behavioural strategies.
In a two lever Go-NoGo task, where rats were trained to press one lever in response to a
visual cue to obtain a reward but where responding to another visual cue by pressing the other lever
had no consequence, PrL firing rate was affected by both reward-paired and no-reward-paired cues
(Moorman and Aston-Jones, 2015). However, the strength of PrL signalling immediately after cue
onset was greater when the rat subsequently responded correctly (Hit and correct rejection (CR)
trials) compared to trials where the rat responded incorrectly (Miss and false alarm (FA) trials)
(Moorman and Aston-Jones, 2015), suggesting that PrL incorporates contextual information into the
regulation of behaviour, rather than strictly promote or inhibit behavioural responding.
4.1.2 Prelimbic modulation of striatal processes
PrL sends strong projections to both core and shell of the NAc (Ding et al., 2001, Hart et al., 2014,
Gabbott et al., 2005, Balleine et al., 2009, Groenewegen et al., 1999, Balleine et al., 2007,
Heidbreder and Groenewegen, 2003) as part of the limbic cortico-striatal-thalamic circuit and to
dorsomedial striatum (DMS) as part of the associative cortico-striatal-thalamic circuit (Gabbott et al.,
2005, Groenewegen et al., 1999, Hart et al., 2014, Balleine et al., 2007, Balleine and O'Doherty,
2010, Heidbreder and Groenewegen, 2003).
The limbic cortico-striatal-thalamic circuit maintains motivational aspects of reward-seeking
behaviour (Yin et al., 2008, Balleine, 2005) and interaction between mPFC and NAc is likely to be
involved in the updating of response-outcome contingencies. Neither lesion of mPFC nor NAc core
alone affect the ability of rats to stop an already initiated motor response in a stop-signal reaction
time task (Eagle and Robbins, 2003). Similarly, NAc core lesions alone had no significant effect on
incorrect responding in the 5 choice serial reaction time task (5-CSRTT) (Christakou et al., 2004).

68
Contralateral2 lesions of mPFC and NAc core increased the number of incorrect responses in the 5-
CSRTT but only in trials following trials with correct response, whereas trials following incorrect
response were unaffected (Christakou et al., 2004). Furthermore, these errors were unaffected my
manipulation of stimulus discriminability (Christakou et al., 2004), suggesting that these errors were
not attributable to attentional impairment but rather that disruption of mPFC-NAc core connectivity
interfered with the planning of responding in subsequent trials immediately after positive
reinforcement. In a recent study, single units were recorded in either mPFC or NAc in rats during
performance in the 5-CSRTT. In this study ramping activity was observed in both structures between
nose poke initiated trial start and subsequent nose poke into one of the 5 ports, regardless of
whether the second nose poke was correct, incorrect or premature, whereas ramping activity was
absent in mPFC and reduced in NAc in omission trials when trial start was not followed by a nose
poke (Donnelly et al., 2015). This suggests a role for both structures in response initation, with the
ramping activity representing internal time representation, that begins too early in trials with
premature responses, and the absence of ramping activity in mPFC in conjunction with reduction in
NAc in omission trials, suggests this activity may be correlates with task management, possibly
imposed top down from mPFC to NAc (Donnelly et al., 2015).
As part of the associative cortico-striatal-thalamic circuit, communication between mPFC and DMS
likely modulate cue-guided behavioural shifting during tasks that require discrimination between
sets of different stimulus-outcomes, particularly when context increases attentional demands
(Christakou et al., 2004, Christakou et al., 2001, Baker and Ragozzino, 2014). In humans, mPFC –
DMS synchronisation have been found to predict successful performance in an active coping task,
where mild electric shock was given in some error trials (Collins et al., 2014). In rats, contralateral
lesions of mPFC and DMS increased the number of errors and latency to respond in the 5-CSRTT, but
left locomotion and Pavlovian approach behaviour intact, while premature responses returned to
control level when the duration of the stimulus was increased. Taken together, these observations
suggest impairment of attentional rather than motivational processes (Christakou et al., 2001).
Interaction between PrL and DMS may coordinate switching between behavioural strategies
in response to context. In a conditional discrimination task where rats relied on visual cues to
determine which arm in a maze was rewarded in the following block of 3-5 trials, inactivation of
DMS increased perseverance errors, i.e. the rat required more trials within a new block to switch
2 As mPFC projections to striatum are primarily ipsilateral, unilateral lesion of mPFC and NAc leaves the
connection between these structures unimpaired in one hemisphere, whereas contralateral lesions disconnects the structures in both hemispheres (Christakou et al. 2001).

69
arm or completely failed to switch arm, whereas inactivation of PrL led to turn bias, where rats
consistently chose the right or left arm, ignoring visual cues (Baker and Ragozzino, 2014). In this
study, ipsilateral inactivation of PrL and DMS together did not affect performance. However,
contralateral inactivation of communication between structures impaired performance specifically in
trials when rats have to switch from one arm to another, whereas performance within trial blocks,
where no switching was required, was unaffected (Baker and Ragozzino, 2014). This suggests that
ipsilateral communication between PrL and DMS modulate cue-guided behavioural shifting during
tasks that require discrimination between sets of different stimulus-outcomes. In a choice task,
where trials switched between variable and fixed reward conditions, single neurons in both PrL and
DMS were found to track action-reward associations across trial types (Funamizu et al., 2015),
further implicating both Prl and DMS processing in the ability to switch behaviour in response to
changes in context-outcome associations.
Dorsolateral striatum (DLS) receives projections from primary motor and sensory cortex (Van Waes
et al., 2012, Redgrave et al., 2011) as part of the sensory-motor cortico-striatal-thalamic circuit and
labelling studies suggest that there are no direct anatomical connection between these structures
(Voorn et al., 2004). Thus, the research on PrL to striatum interaction has so far focused on
interaction effects with DMS and NAc. However, complex behavioural strategies often require
repeated training over time making the behaviour increasingly automated, drawing on increasing
contribution from DLS (Dias-Ferreira et al., 2009, Balleine et al., 2009, Yin et al., 2005, Hilario et al.,
2012). Complex behavioural tasks that involve switching between behavioural strategies in response
to context likely rely on DMS processing (Funamizu et al., 2015) as well as DLS processing (Balleine et
al., 2009, Tang et al., 2009) even in over-trained animals. Successful management of a task with
multiple stimulus-response-outcome contingencies requires some level of executive control of DMS
vs. DLS modulation of the behavioural output (Baker and Ragozzino, 2014, Funamizu et al., 2015,
Riga et al., 2014, Heidbreder and Groenewegen, 2003, Christakou et al., 2004). MPFC, with its
extensive efferents to both ventral and dorsal striatum is in a key position to exert such control
(Christakou et al., 2001, Baker and Ragozzino, 2014, Thorn and Graybiel, 2014), and projections
between dorsal and ventral part of mPFC allows PrL and IL to coordinate mPFC regulation of habitual
vs. goal-directed striatal processes (Riga et al., 2014, Moorman and Aston-Jones, 2015). However, to
fully elucidate mPFC contribution to striatal processing during complex behaviour a more integrated
approach is needed, ideally examining mPFC interaction with distinct dorsal and ventral striatal sub-
regions simultaneously.

70
4.1.3 Study aims
Corticostriatal communication from PrL to both ventral and dorsal striatum likely play a role in
appetitive behaviour, particularly when tasks are demanding and involve shifts between several
stimulus-response-outcome contingencies (Baker and Ragozzino, 2014, Funamizu et al., 2015, Riga
et al., 2014, Heidbreder and Groenewegen, 2003). Successful behaviour in such tasks likely require
executive control exerted by discrete mPFC regions, such as PrL, onto striatal subregions, most
notably NAc and DMS (Christakou et al., 2001, Baker and Ragozzino, 2014, Thorn and Graybiel,
2014).
The study presented in this chapter examined the contribution of PrL single unit activity and
synchronisation between PrL and DMS, DLS and NAc during execution of two comparable
conditioned discrimination tasks; the Go-NoGo Plus and the Go-NoGo Minus task. In both tasks rats
were trained to press a lever in response to an auditory Go cue and to suppress responding to an
auditory NoGo cue. Correct lever press was rewarded with a sugar pellets in both tasks, whereas
correct response suppression (in NoGo trials) was only rewarded in the Go-NoGo Plus task (Figure 3-
1). In both tasks, the rats have to discriminate between stimulus-outcome contingencies and initiate
or suppress motor response accordingly, enabling us to study mPFC-striatal interaction in response
to such cognitive demands. In the standard Go-NoGo (Minus) task cues signalling reward availability
also signal to the animal to make a motor response: therefore, the modified Go-NoGo Plus task was
introduced to enable separation of motor and reward component of cue evoked responses.
Hypothesis
Because mPFC projects directly to DMS and NAc but not to DLS, greater task-related
synchronisation was expected between PrL and DMS and PrL and NAc compared with
synchronisation between PrL and DLS in response to trial onset cues.
4.2 Methods
The data presented in this chapter were collected from the same animals as in chapter 3. Therefore
the majority of the methods employed are identical, and are only summarised briefly. Full details of
the methodology can be found in Chapter 3.

71
4.2.1 Animals
Male Lister Hooded rats (n = 7; Charles River, Cambridge, UK) weighing 225-250g on arrival were
kept on reversed light/dark cycle (12:12h; lights on 19.00h). Animals had access to water ad libitum
and access to food for at least 2h per day. All experiments were carried out under institutional
ethical approval and with project and personal licence approved by the UK Home Office.
4.2.2 Apparatus
All behavioural training and testing was conducted in four identical operant chambers, comprising a
metal grid floor, with a magazine for delivery of sugar pellets and a retractable lever positioned to
the left of the magazine. A light was positioned immediately above each and a speaker was
positioned above the magazine and a house light was positioned at the top of the opposite wall of
the chamber (for full details see paragraph 3.2.2). Electrophysiological recordings were carried out in
a similar chamber, with minor modifications to facilitate recordings (Plexiglas cover and custom
made food receptacle: see paragraph 3.2.2 for details).
4.2.3 Behavioural training
Behavioural procedures were as described in Chapter 3 – paragraph 3.2.3). Briefly, rats were trained
to either respond (Go trials) or suppress (NoGo trials) responding to auditory cues of different
frequencies (1 vs. 10 kHz (75dB): counterbalanced). Each trial started with the presentation of either
the Go or NoGo tone. After 4 seconds the lever was presented allowing the rat a 4 second response
interval to press the lever. Upon lever press, or at the end of the 4 second response interval, the
lever retracted and the tone was switched off. There was a 60 second inter trial interval (ITI)
between trials (Figure 3-1). In the Go-NoGo plus task rats (n = 4) were rewarded with a sugar pellet
for both correct lever press (Hit) and correct omission of lever press (Correct Rejection) trials. In the
Go-NoGo minus task (n = 3) only correct lever press (Hit) trials were rewarded whereas correct
rejections had no programmed consequence. In both tasks incorrect lever press (False Alarm)
resulted in a 60 second time-out with lights out. Incorrect omission of lever press (Miss) had no
programmed consequence (Figure 3-1). The rats were considered to be discriminating at criterion
when Hit rate (number of Hit trials divided by number of Go trials) was above 0.8 and False Alarm
rate (number of False Alarm trials divided by number of NoGo trials) below 0.25 within a session for
3 consecutive sessions.
4.2.4 Surgery
Once rats were discriminating at criterion, a, affixed to Neuralynx drives, was implanted to target PrL
in the left hemisphere at +3.2AP, +1.1ML and -2.6DV. In addition 7 tungsten tetrode recording

72
electrodes were implanted to target DLS, DMS, NAc shell, NAc core(for striatal coordinates, see
Table 3-1). The tetrodes were sealed with paraffin wax and the implant was built up using layers of
light curing dental cement. A silver wire inserted into the cerebellum served as a ground. Surgical
procedures and post-operative care are described in full in Chapter 3 – paragraph 3.2.5).
4.2.5 Electrophysiological recordings
15-30 minutes prior to each recorded session, each tetrode was lowered approximately 0.125mm
(corresponding to a 180 degree turn of the drive screw) to ensure that different neurons were
recorded in each session. The rat was connected through a flexible wire, allowing unimpaired
movement, to a 32 channel head stage (Plexon Inc., Dallas, TX, USA) immediately before recording
and placed in an operant chamber placed in a sound-attenuated aluminium-plated box, which
served as a Faraday cage. During the discrimination task, wideband signals were acquired
continuously at a sampling rate of 25000hz via an op-amp based head-stage amplifier (HST/32o25-
36P-GR, 1x gain, Plexon Inc., Dallas, TX, USA), and passed through a preamplifier (PBX2/32wb, 1000x
gain; Plexon Inc., Dallas, TX, USA).
Filtering, artefact removal, spike sorting and synchronisation of spike activity with behaviour
were carried out using Matlab-based routines, as previously described (paragraph 3.2.6) and
statistical analysis was calculated using SPSS 22 Statistics (IBM SPSS, Somers, NY, USA).
Figure 4-1 Verification of tetrode placement Tetrode placements were verified visually while cutting the frozen brains in 30μm slices on a cryostat. Only sessions where the tetrode tips were in the PrL (marked in red) were included in the analysis of firing rate responses to cue onset.

73
4.2.6 Verification of tetrode placement
After the last recording session, the rats were anaesthetised, perfused and the brain removed using
the same procedure as described in chapter 3, Methods. Tetrode placement was verified visually
while cutting the perfused frozen brains in 30μm slices on a cryostat (Figure 3-4 and Figure 4-1). For
those tetrodes, for which precise location of the tip could not be identified visually, a presumed
position was calculated using the same criteria as described in chapter 3, Methods. One tetrode
targeting DMS (Go-NoGo Minus task), one tetrode targeting PrL and three tetrodes targeting NAc
(Go-NoGo Plus task) were excluded from the analysis due to incorrect placement.
4.2.7 Statistical analysis
Behavioural and electrophysiological data were not normally distributed and did not pass Levine’s
test for homogeneity of variance, and were log transformed to allow the use of parametric tests
(Buzsaki and Mizuseki, 2014). Log transformed firing rates for inhibited neurons were multiplied by -
1 for ease of visualisation. Firing rates and coherence were analysed for effect of trial type and
structure using ANOVAs and post hoc LSD test where appropriate. All statistical analysis was
calculated using SPSS 22 Statistics (IBM SPSS, Somers, NY, USA). P values below 0.05 were
considered statistically significant.
Analysis of behaviour: Mean response rates to Go and NoGo tones as well as mean latency
to lever press in Hit and FA trials were calculated from session included in electrophysiological
analyses (sessions where tetrode tips where positioned outside the target structure were excluded).
Analysis of single unit responses. The highest number of significantly responding neurons
compared with baseline (3 seconds before cue onset) was found in the first 100 msec after cue onset
in all four trial types in both tasks, wherefore this interval was chosen for further analysis. Analysis
was only performed on neurons that displayed a significant change (Wilcoxon’s signed rank test) in
mean firing rate (spikes per second) in at least one trial type (Hit, Miss, CR or FA). Baseline neuronal
activity was measured as the firing rate (spikes/sec) during the 3 seconds before cue onset. Neurons
with a baseline firing rate below 6 spikes/sec were included for further analysis.
To examine the role of coherence between striatal subregions on cue onset responses, cross-
spectrum based spike coherence between neurons in the different striatal subregions were
calculated during baseline (-3 to 0 sec before cue onset) and in the cue response phase (0 to 3
seconds after cue onset) (Halliday, 2015) (Matlab code available online at
http://www.neurospec.org).

74
4.3 Results
Single unit responses to cue onset and during behavioural response were recorded in 49 sessions
from 4 rats in the Go-NoGo Plus task, and in 39 sessions from 3 rats in the Go-NoGo Minus task.
After controlling for correct placement of the tetrode tips within the targeted structures, 317 and
204 neurons in striatum (for details on included neurons from each sub-region, see Table 3-2) and 48
and 70 neurons in PrL (Table 4-1) recorded during performance of the Go-NoGo Plus and Go-NoGo
Minus task, respectively, were included for further analysis.
Go-NoGo Plus Go-NoGo Minus
Analysed neurons
significant response
Analysed neurons
Significant response
48 29 70 51
Table 4-1 Number of analysed and significantly responding units in PrL Number of analysed neurons recorded from PrL in the two tasks and the proportion of these neurons that significantly altered their firing rate in the first 100ms after cue onset compared with baseline.
4.3.1 Behaviour
Behavioural results are reported Chapter 3. In summary, all rats trained in either the Go-NoGo Plus
or the Go-NoGo Minus task successfully learned to discriminate between the Go and NoGo tone and
maintained a high average level of discrimination, i.e. Go trial Hit ratio above 0.75 and NoGo trial FA
ratio below 0.25, until the end of the experiment (Figure 3-7A & B).
4.3.2 Firing rate response to cue onset
Intervals of increasing duration after cue onset was analysed to find the interval yielding the highest
number of neurons that significantly changed their firing rate in response to cue onset, compared
with baseline (3 seconds before cue onset). The analysed interval ran from 0- 100ms after cue onset
to 0-4sec after cue onset, in incremental steps of 50ms. The highest number of significantly
responding neurons was found in the first 100 msec after cue onset in all four trial types in both
tasks (Figure 3-5 & 4-2). Therefore, neurons showing a significant change in firing rate within the first
100ms after cue onset, in at least one trial type, were selected for further analysis of effect of trial
type (Table 4-1). Significantly responding neurons were divided into two groups depending on
whether they increased or decreased their firing in response to cue onset. Consecutive100ms
windows were examined for effect of trial type during the first 4 seconds of cue presentation (before
lever extension). Trial type dependent differences in firing rate response to cue onset were only
statistically significant in the PRL within the first 100ms interval after cue. Therefore, only data from
the interval “0-100ms” from cue onset is presented here (Figure 4-2).
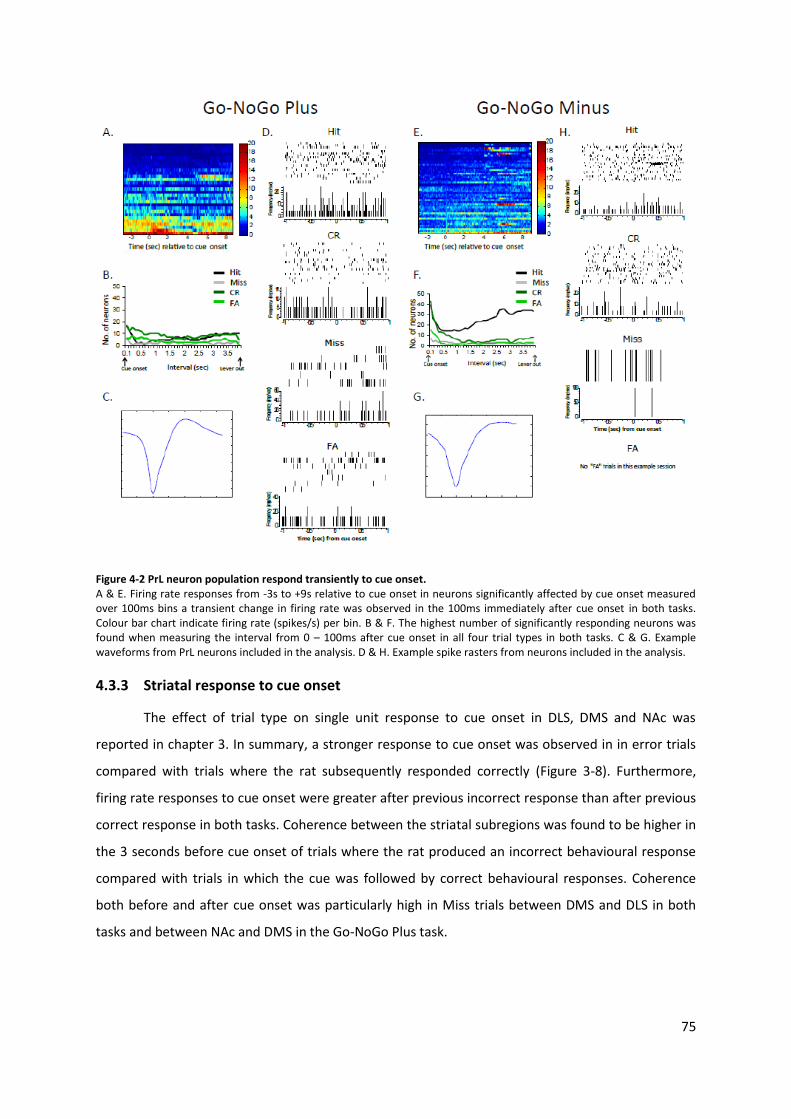
75
Figure 4-2 PrL neuron population respond transiently to cue onset. A & E. Firing rate responses from -3s to +9s relative to cue onset in neurons significantly affected by cue onset measured over 100ms bins a transient change in firing rate was observed in the 100ms immediately after cue onset in both tasks. Colour bar chart indicate firing rate (spikes/s) per bin. B & F. The highest number of significantly responding neurons was found when measuring the interval from 0 – 100ms after cue onset in all four trial types in both tasks. C & G. Example waveforms from PrL neurons included in the analysis. D & H. Example spike rasters from neurons included in the analysis.
4.3.3 Striatal response to cue onset
The effect of trial type on single unit response to cue onset in DLS, DMS and NAc was
reported in chapter 3. In summary, a stronger response to cue onset was observed in in error trials
compared with trials where the rat subsequently responded correctly (Figure 3-8). Furthermore,
firing rate responses to cue onset were greater after previous incorrect response than after previous
correct response in both tasks. Coherence between the striatal subregions was found to be higher in
the 3 seconds before cue onset of trials where the rat produced an incorrect behavioural response
compared with trials in which the cue was followed by correct behavioural responses. Coherence
both before and after cue onset was particularly high in Miss trials between DMS and DLS in both
tasks and between NAc and DMS in the Go-NoGo Plus task.
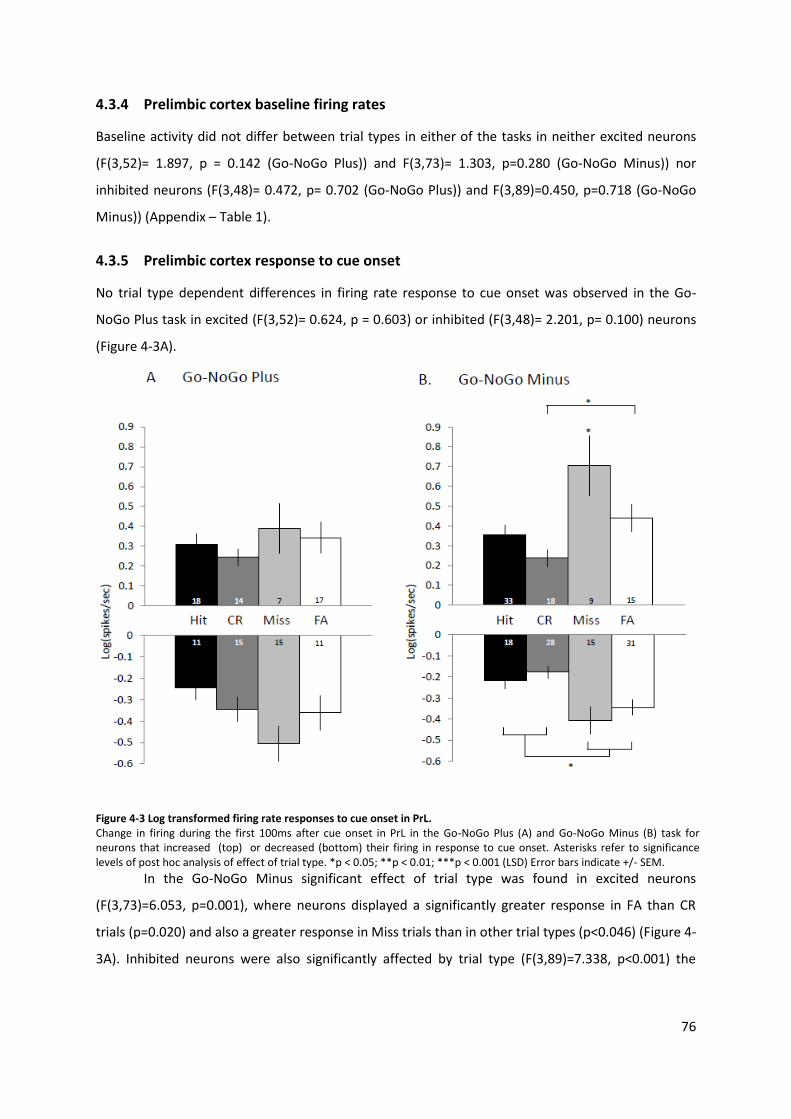
76
4.3.4 Prelimbic cortex baseline firing rates
Baseline activity did not differ between trial types in either of the tasks in neither excited neurons
(F(3,52)= 1.897, p = 0.142 (Go-NoGo Plus)) and F(3,73)= 1.303, p=0.280 (Go-NoGo Minus)) nor
inhibited neurons (F(3,48)= 0.472, p= 0.702 (Go-NoGo Plus)) and F(3,89)=0.450, p=0.718 (Go-NoGo
Minus)) (Appendix – Table 1).
4.3.5 Prelimbic cortex response to cue onset
No trial type dependent differences in firing rate response to cue onset was observed in the Go-
NoGo Plus task in excited (F(3,52)= 0.624, p = 0.603) or inhibited (F(3,48)= 2.201, p= 0.100) neurons
(Figure 4-3A).
Figure 4-3 Log transformed firing rate responses to cue onset in PrL. Change in firing during the first 100ms after cue onset in PrL in the Go-NoGo Plus (A) and Go-NoGo Minus (B) task for neurons that increased (top) or decreased (bottom) their firing in response to cue onset. Asterisks refer to significance levels of post hoc analysis of effect of trial type. *p < 0.05; **p < 0.01; ***p < 0.001 (LSD) Error bars indicate +/- SEM.
In the Go-NoGo Minus significant effect of trial type was found in excited neurons
(F(3,73)=6.053, p=0.001), where neurons displayed a significantly greater response in FA than CR
trials (p=0.020) and also a greater response in Miss trials than in other trial types (p<0.046) (Figure 4-
3A). Inhibited neurons were also significantly affected by trial type (F(3,89)=7.338, p<0.001) the
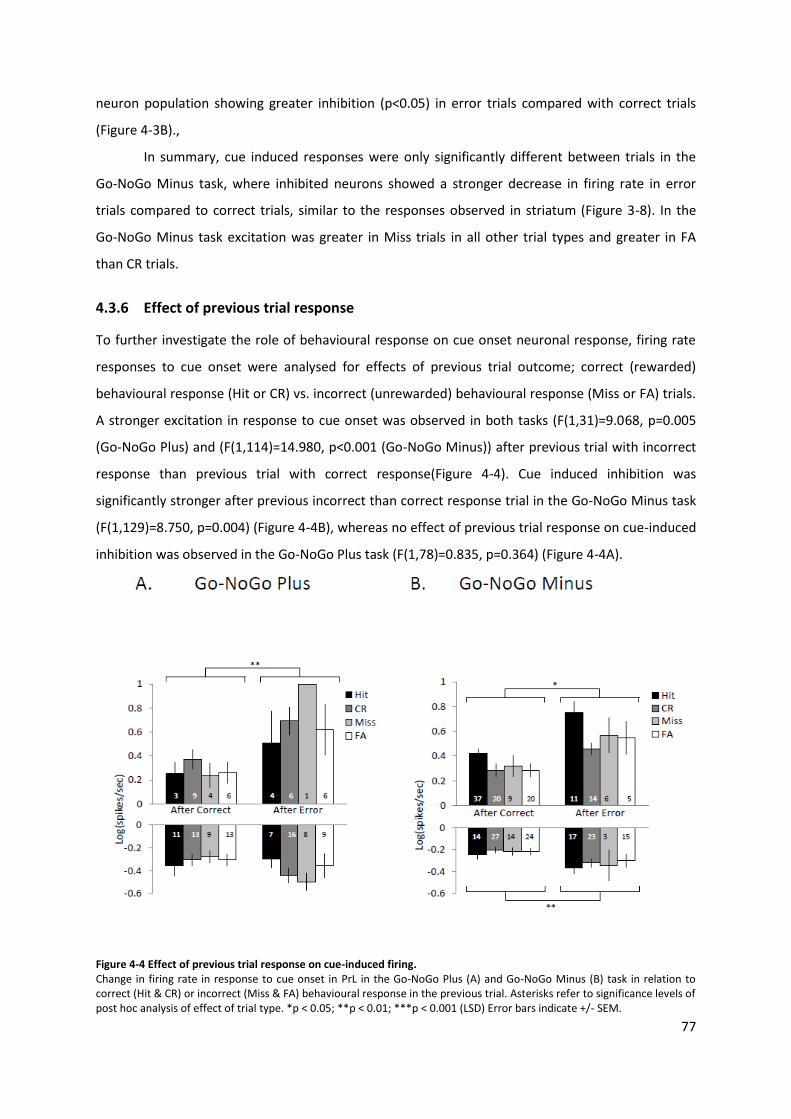
77
neuron population showing greater inhibition (p<0.05) in error trials compared with correct trials
(Figure 4-3B).,
In summary, cue induced responses were only significantly different between trials in the
Go-NoGo Minus task, where inhibited neurons showed a stronger decrease in firing rate in error
trials compared to correct trials, similar to the responses observed in striatum (Figure 3-8). In the
Go-NoGo Minus task excitation was greater in Miss trials in all other trial types and greater in FA
than CR trials.
4.3.6 Effect of previous trial response
To further investigate the role of behavioural response on cue onset neuronal response, firing rate
responses to cue onset were analysed for effects of previous trial outcome; correct (rewarded)
behavioural response (Hit or CR) vs. incorrect (unrewarded) behavioural response (Miss or FA) trials.
A stronger excitation in response to cue onset was observed in both tasks (F(1,31)=9.068, p=0.005
(Go-NoGo Plus) and (F(1,114)=14.980, p<0.001 (Go-NoGo Minus)) after previous trial with incorrect
response than previous trial with correct response(Figure 4-4). Cue induced inhibition was
significantly stronger after previous incorrect than correct response trial in the Go-NoGo Minus task
(F(1,129)=8.750, p=0.004) (Figure 4-4B), whereas no effect of previous trial response on cue-induced
inhibition was observed in the Go-NoGo Plus task (F(1,78)=0.835, p=0.364) (Figure 4-4A).
Figure 4-4 Effect of previous trial response on cue-induced firing. Change in firing rate in response to cue onset in PrL in the Go-NoGo Plus (A) and Go-NoGo Minus (B) task in relation to correct (Hit & CR) or incorrect (Miss & FA) behavioural response in the previous trial. Asterisks refer to significance levels of post hoc analysis of effect of trial type. *p < 0.05; **p < 0.01; ***p < 0.001 (LSD) Error bars indicate +/- SEM.
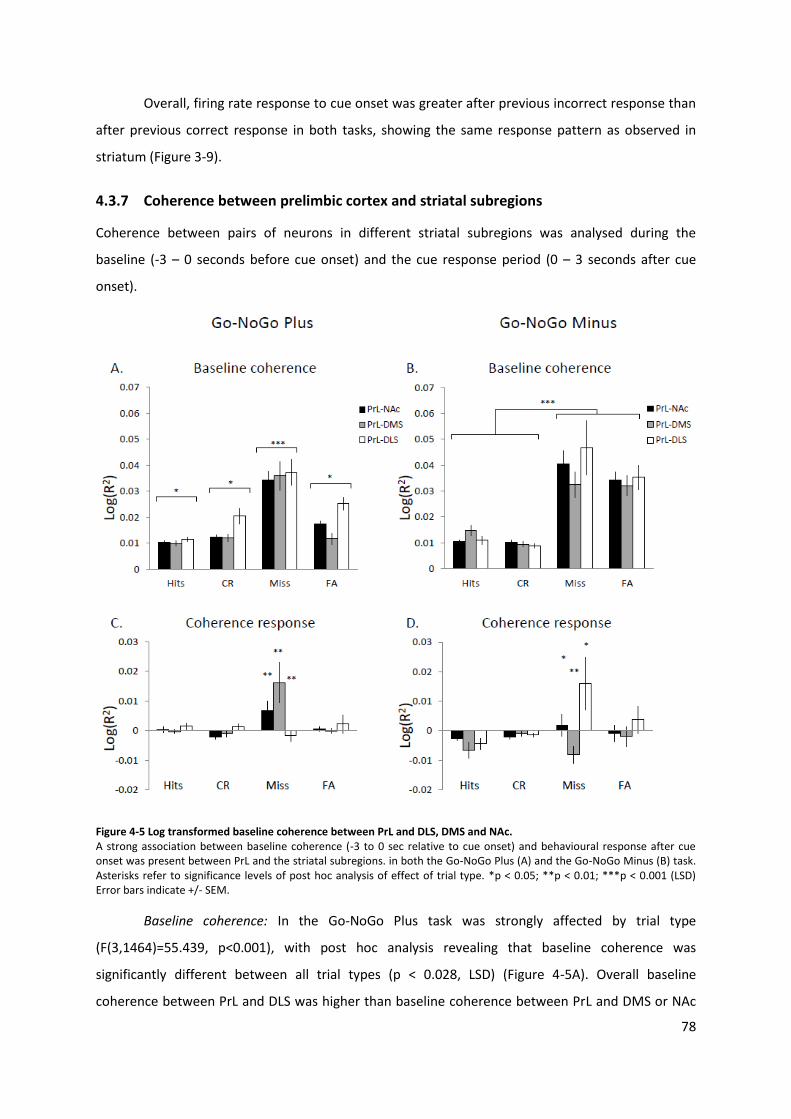
78
Overall, firing rate response to cue onset was greater after previous incorrect response than
after previous correct response in both tasks, showing the same response pattern as observed in
striatum (Figure 3-9).
4.3.7 Coherence between prelimbic cortex and striatal subregions
Coherence between pairs of neurons in different striatal subregions was analysed during the
baseline (-3 – 0 seconds before cue onset) and the cue response period (0 – 3 seconds after cue
onset).
Figure 4-5 Log transformed baseline coherence between PrL and DLS, DMS and NAc. A strong association between baseline coherence (-3 to 0 sec relative to cue onset) and behavioural response after cue onset was present between PrL and the striatal subregions. in both the Go-NoGo Plus (A) and the Go-NoGo Minus (B) task. Asterisks refer to significance levels of post hoc analysis of effect of trial type. *p < 0.05; **p < 0.01; ***p < 0.001 (LSD) Error bars indicate +/- SEM.
Baseline coherence: In the Go-NoGo Plus task was strongly affected by trial type
(F(3,1464)=55.439, p<0.001), with post hoc analysis revealing that baseline coherence was
significantly different between all trial types (p < 0.028, LSD) (Figure 4-5A). Overall baseline
coherence between PrL and DLS was higher than baseline coherence between PrL and DMS or NAc
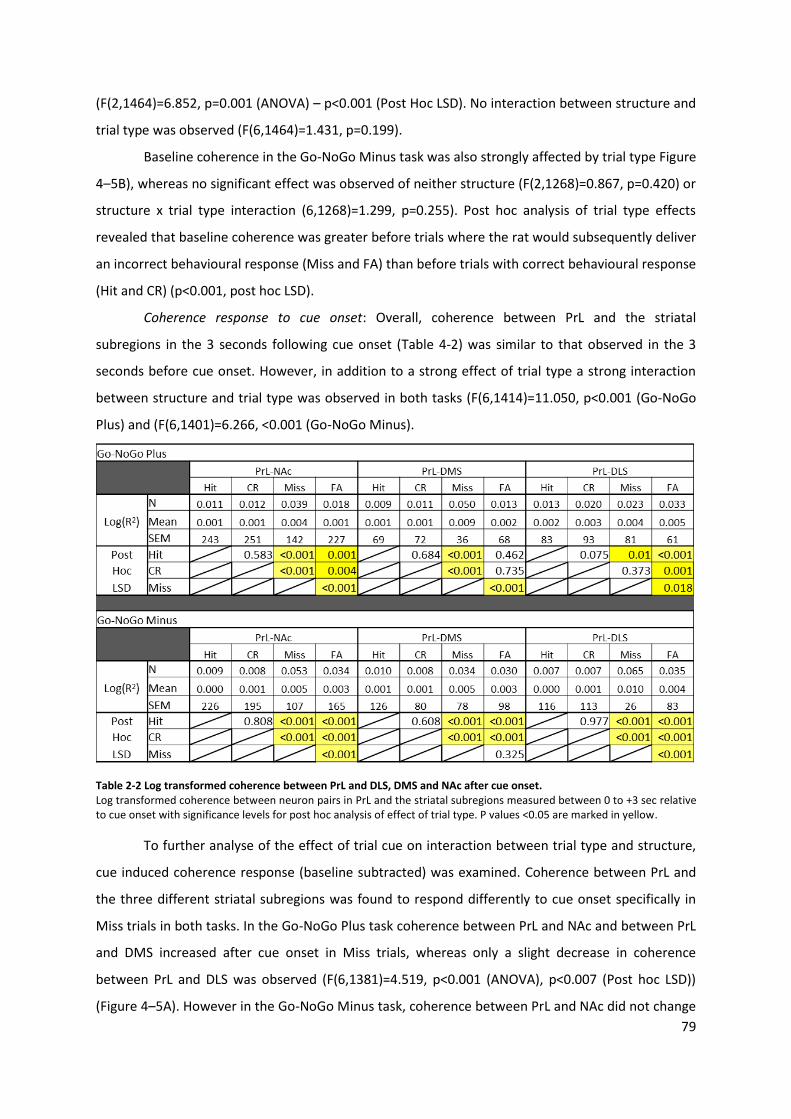
79
(F(2,1464)=6.852, p=0.001 (ANOVA) – p<0.001 (Post Hoc LSD). No interaction between structure and
trial type was observed (F(6,1464)=1.431, p=0.199).
Baseline coherence in the Go-NoGo Minus task was also strongly affected by trial type Figure
4–5B), whereas no significant effect was observed of neither structure (F(2,1268)=0.867, p=0.420) or
structure x trial type interaction (6,1268)=1.299, p=0.255). Post hoc analysis of trial type effects
revealed that baseline coherence was greater before trials where the rat would subsequently deliver
an incorrect behavioural response (Miss and FA) than before trials with correct behavioural response
(Hit and CR) (p<0.001, post hoc LSD).
Coherence response to cue onset: Overall, coherence between PrL and the striatal
subregions in the 3 seconds following cue onset (Table 4-2) was similar to that observed in the 3
seconds before cue onset. However, in addition to a strong effect of trial type a strong interaction
between structure and trial type was observed in both tasks (F(6,1414)=11.050, p<0.001 (Go-NoGo
Plus) and (F(6,1401)=6.266, <0.001 (Go-NoGo Minus).
Table 2-2 Log transformed coherence between PrL and DLS, DMS and NAc after cue onset. Log transformed coherence between neuron pairs in PrL and the striatal subregions measured between 0 to +3 sec relative to cue onset with significance levels for post hoc analysis of effect of trial type. P values <0.05 are marked in yellow.
To further analyse of the effect of trial cue on interaction between trial type and structure,
cue induced coherence response (baseline subtracted) was examined. Coherence between PrL and
the three different striatal subregions was found to respond differently to cue onset specifically in
Miss trials in both tasks. In the Go-NoGo Plus task coherence between PrL and NAc and between PrL
and DMS increased after cue onset in Miss trials, whereas only a slight decrease in coherence
between PrL and DLS was observed (F(6,1381)=4.519, p<0.001 (ANOVA), p<0.007 (Post hoc LSD))
(Figure 4–5A). However in the Go-NoGo Minus task, coherence between PrL and NAc did not change

80
in response to cue onset in Miss trials whereas coherence between coherence between PrL and DMS
decreased and coherence between PrL and DLS increase after cue onset (F(6,1225)=2.272, p=0.018
(ANOVA), p<0.020 (Post hoc LSD)) (Figure 4-5B).
In summary, baseline coherence in the Go-NoGo Plus task showed significant differences
between all trial types, with the highest corticostriatal coherence being observed before Miss trials.
Baseline coherence in the Go-NoGo Minus task was significantly greater before error trials than
correct trials. Further analysis of cue induced changes in corticostriatal coherence revealed that
coherence between PrL and the three striatal subregions responded differently to cue onset in Miss
trials. Differences between tasks
Although trial type did not affect baseline firing rates in neither the Go-NoGo Plus nor the Go-NoGo
Minus task, baseline firing rate did differ between tasks, with baseline firing rate being significantly
higher in the Go-NoGo Plus task than the Go-NoGo Minus task in neurons inhibited by cue onset
(F(1,137)= 10.165, p = 0.002), whereas no different between tasks was found in neurons excited by
cue onset (F(1,125)= 1.267, p = 0.262).
After cue onset, overall excitation was significantly greater in the Go-NoGo Minus task
(F(1,125)= 5.520, p = 0.020), whereas overall inhibition was greater in the Go-NoGo Plus task
(F(1,137)= 4.222, p = 0.042). When examining the role of task on the effect of previous trial
response, no significant effect of task was found neither in excited neurons (F(1,145)=0.129,
p=0.721) nor in inhibited neurons (F(1,207)=3.369, p=0.068).
Baseline coherence was also significantly higher in the Go-NoGo Minus than the Go-NoGo
Plus task (F(1,2732)=12.151, p<0.001), and interaction between task and trial type was also highly
significant (F(3,2732)=17.619, p<0.001) whereas no interaction with structure (F(2,2732)=0.809,
p=0.445) or interaction between task, structure and trial type (F(6.2732)=1.454, p=0.190) was found.
Cue induced change in coherence differed between tasks (F(1,2606)=7.107, p=0.008 and also
showed a significant interaction between task, structure and trial type (F(6,2606)=5.691, p<0.001).
In summary, the rats in the two tasks displayed significantly different baseline firing rates.
This task effect persisted after cue onset in inhibited neurons and was also manifested in differences
in striatal coherence between the two tasks.
4.4 Discussion
4.4.1 Baseline single unit activity
Although trial type did not affect baseline firing rates in either of the two tasks, baseline firing rate
was significantly greater in the Go-NoGo Plus than in the Go-NoGo Minus task in neurons inhibited

81
by cue onset (Appendix - Table 1) and this task-effect persisted after cue onset. Similarly, coherence
also differed between task, with both pre and peristimulus coherence being higher in the Go-NoGo
Minus task than in the Go-NoGo Plus task (Figure 4-3A & B and Table 4-2). The task related
differences in cue-induced inhibition and peristimulus coherence are likely influenced by the overall
difference in baseline values, which makes comparison of task effects on cue-induced neuronal
responses difficult to interpret. Therefore, the following discussion will mainly focus on trial type and
structure dependent differences in firing rate response and coherence within each task.
4.4.2 PrL single unit response to cue onset
Firing rate response to cue onset in PrL were only significantly different between trial types in the
Go-NoGo Minus task, where greater inhibition was observed in error trials compared with correct
trials (Figure 4-3B). The greater inhibitory response in PrL to error trials compared with trials with
correct response is similar to the inhibitory firing rate responses observed in striatal subregions
(reported in Chapter 3 – Figure 3-8), suggesting that mPFC and striatal subregions interact during
complex behaviour. This is in accord with lesioning studies implicating interaction between mPFC
and DMS and NAc in planning and execution of tasks that require switching between multiple
stimulus-outcome contingencies (Christakou et al., 2001, Baker and Ragozzino, 2014).
In the Go-NoGo Minus task, cue induced excitation was greater in Miss trials than in correct
trials, whereas excitation in FA trials was significantly greater than CR but smaller than Miss trial
excitation. Increased firing rate response to cue onset in error trials compared with correct trials
observed in the Go-NoGo Minus task, may be caused by increased attentional load associated with
the decision to contradict the overtrained correct stimulus-response contingency. A similar, but
opposite, relationship between PrL firing and trials with correct and incorrect response was
observed in rats trained in a two-lever Go-NoGo task. In this task the strength of PrL signalling
immediately after cue onset was greater in Hit trials compared with Miss trials and in CR trials
compared with FA trials (Moorman and Aston-Jones, 2015). The behavioural setup in the above
study is more complex than the Go-NoGo Minus task presented in this thesis, in that the rats
discriminated between two levers as well as two tones – with only one tone and one lever being
rewarded. The findings in the above study as well as in the current study suggest that PrL
incorporates contextual information into the regulation of behaviour, rather than strictly promoting
or inhibiting behavioural responding. However, more research is needed to determine if differences
in task complexity affect the direction of firing rate responses to context. Stronger excitation in Miss
than FA trials may indicate that in tasks where incorrect responses require motor inhibition, such
motor inhibition is associated with greater PrL activation than when the incorrect response requires

82
motor initiation. Previous research has shown that inactivation of PrL in rats increases incorrect
responses, suggesting PrL activity contributes to inhibitory response control (Jonkman et al., 2009)
as well as organisation and planning of flexible behaviour, based on previously acquired information
(Dalley et al., 2004). However, more research is needed to elucidate the role of PrL single unit
responses in relation to reward-directed behaviour.
Cue induced excitation was greater in the Go-NoGo Minus task than in the Go-NoGo Plus
task, whereas no difference between tasks was observed at baseline in cue-excited neurons. This
difference in cue induced excitation may be related to the difference in reward contingency between
the two tasks. However, given that no significant effect of trial was found in the Go-NoGo Plus task,
it is difficult to draw conclusions about how differences in task set up may have influenced cue
induced neuronal responses in the current study.
4.4.3 Effect of previous trial response
To further investigate the role of behavioural response on cue onset neuronal response, firing rate
responses to cue onset were analysed for effects of previous trial outcome; correct (rewarded)
behavioural response (Hit or CR) vs. incorrect (unrewarded) behavioural response (Miss or FA) trials.
Excited neurons were found to display a significantly greater response to cue onset in trials following
incorrect response trials (Miss and FA) than correct response trials (Hit and CR) in both tasks (Figure
4-4). Inhibition was also found to be greater in trials following previous incorrect response than trials
following previous correct response, although this difference only reached significance in the Go-
NoGo Minus task (Figure 4-4). The similarity between responses in the two tasks suggests that the
neuronal responses to cue onset were modulated by recent behavioural experience rather than
recent reward experience. Furthermore, the same pattern was observed in the striatal subregions
(Figure 3-9), suggesting that PrL and striatum interact to update and fine tune behavioural responses
based on recent experience. Disruption of mPFC-NAc have been shown to interfere with the
planning of responding to reward-paired cues, but only in trials following immediately after a trial
with a rewarded correct response (Christakou et al., 2004), further implicating interaction between
mPFC and striatum in in the updating of response-outcome contingencies. A recent study in which
rats where trained on a directional choice task, found greater firing rate response to cue onset in the
agranular areas of frontal cortex after previous correct trials than after error trials (Yuan et al.,
2015), suggesting that recent experience is associated with firing rate response to cue onset in both
of these cortical regions, as well as striatum, although with opposite effect. Updating of behaviour
based on recent experience may be maintained by a network of cortical and subcortical regions,
requiring coordination of excitation and inhibition in different brain areas in order to optimise

83
output. However, more research on the role of recent experience on activity in cortical and
subcortical networks is warranted to understand its implication on future behavioural choice.
4.4.4 Coherence between PrL and striatum
Overall, prestimulus coherence was higher before error trials than correct response trials in both
tasks and highest in Miss trials (where the rats failed to respond to the Go cue) (Figure 4-5).
Increased coherence may reflect low levels of attention to external stimuli, causing the rat to miss
the cue (Gusnard and Raichle, 2001, Herzog et al., 2014). Several studies in humans and monkeys
have demonstrated that particularly alpha band coherence between different cortical regions before
cue onset affects stimulus detection (Boly et al., 2007, Sadaghiani et al., 2010, Forstmann et al.,
2010, Melloni et al., 2007, Shulman et al., 2002). More recently, a study in rats performing an
auditory detection task found increased theta, alpha and beta band coherence between frontal and
parietal cortex before cue onset in trials where the rats failed to detect the tone (Herzog et al.,
2014). Fluctuation of activity and network connectivity has long been linked to attentional state and
previous work suggests that low detection rates following high coherence likely represent functional
inhibition within the target network diverting attention away from external stimuli to focus attention
on internal representations such as working memory (Hanslmayr et al., 2007, Mazaheri et al., 2011,
Cooper et al., 2003, Gusnard and Raichle, 2001, van Dijk et al., 2008). Through the associative,
sensory-motor and limbic cortico-striatal-thalamic circuits (Van Waes et al., 2012, Redgrave et al.,
2011), the PrL and striatal subregions are intricately connected to both task-positive and task-
negative networks implicated in the regulation of attention to external stimuli (Sadaghiani et al.,
2010). The high prestimulus coherence between striatal subregions observed in Miss may be driven
by modulatory input from task-negative regions of cortex or thalamus. More studies, recording from
multiple sites in cortical and subcortical networks, are needed to investigate the role of coherence
within these networks on future behavioural choice.
In both tasks, prestimulus coherence was greater in FA trials as well as in Miss trials
compared with correct response trials (Figure 4-5), suggesting that coherence is associated with
future behavioural choice (Figure 4-5), consistent with the overall pattern of coherence observed
between the striatal subregions (Figure 3-10). The high level of coherence between neurons in PrL
and in striatal subregions is well in accord with labelling studies showing strong projections from PrL
to ventral and dorsomedial parts of striatum (Gabbott et al., 2005, Groenewegen et al., 1999,
Heidbreder and Groenewegen, 2003), and further suggest that complex flexible behaviour require
interaction between mPFC and striatum. Both contralateral lesion of mPFC and NAc (Christakou et
al., 2001) or contralateral inactivation of PrL and DMS (Baker and Ragozzino, 2014) have been show

84
to disrupt planning and responding in tasks that require switching between multiple stimulus-
outcome contingencies. However, more research is needed to establish the exact role of this
interaction, in particular to establish if this interaction is strictly top down or if striatal subregions
also influence mPFC activity. Previous work on corticostriatal coherence in humans (Cohen et al.,
2012) and in anaesthetised rats (Sharott et al., 2005) found that cortical structures influenced basal
ganglia activity whereas basal ganglia exerted little or no control of cortical activity. However, a
recent study found that although coherence between primary motor cortex and dorsal striatum was
strictly unidirectional in resting and anesthetised rats, corticostriatal coherence was bidirectional in
awake rats (Nakhnikian et al., 2014) and analysis of causal connectivity in a MRI study in awake
monkeys found striatum to exert a stronger overall influence of striatum on PFC (rather than from
PFC to striatum) during category learning (Antzoulatos and Miller, 2014). These recent findings
suggest that corticostriatal interaction in awake behaving subjects may not exclusively be in the form
of top down control from cortex to striatum but may also involve bottom up modulation from
striatum to cortex. More research is needed to elucidate the precise role of mPFC-striatal interaction
during complex behaviour to establish the direction of this interaction and whether the direction
differs between striatal subregions.
The finding that baseline coherence between PrL and DLS was higher than baseline
coherence between PrL and NAc and DMS was unexpected, because labelling studies suggest that
there are no direct anatomical connection between these structures (Voorn et al., 2004). DLS
received strong projections from primary motor cortex (Koralek et al., 2013) and a recent
anterograde labelling study found evidence of projections from both PrL and IL to primary motor and
primary somatosensory cortex (Bedwell et al., 2014). In addition, similar task-bracketing single unit
activity developed in DLS and IL in rats as a result of training, while task-related activity in PrL
declined (Smith and Graybiel, 2013), which has been suggested to indicate functional connectivity
between IL and DLS (Riga et al., 2014). Such connectivity would enable PrL to influence DLS activity
either through projection via primary motor cortex or through simultaneous modulation from IL of
both areas (Riga et al., 2014, Moorman and Aston-Jones, 2015). The unexpected coherence between
PrL and DLS as well as the overall similarity between trial type dependent baseline coherence
between PrL and all three striatal subregions may indicate joint modulation by other brain areas.
However, more research is needed, incorporating single unit recordings from thalamic and cortical
regions that are part of the limbic, associative and motor cortico-striatal-thalamic circuits, as well as
striatum, to fully elucidate the role of corticostriatal interaction during complex behaviour.
Examining the effect of cue onset on corticostriatal coherence revealed that coherence
between PrL and the three striatal subregions was affected differently by cue onset in Miss trials. In

85
the Go-NoGo Plus task coherence between PrL and NAc and between PrL and DMS increased after
cue onset, whereas only a minor decrease in coherence between PrL and DLS was observed (Figure
4-3A). PrL projects directly to NAc (Ding et al., 2001, Hart et al., 2014, Gabbott et al., 2005, Balleine
et al., 2009, Groenewegen et al., 1999, Balleine et al., 2007, Heidbreder and Groenewegen, 2003)
and DMS (Gabbott et al., 2005, Groenewegen et al., 1999, Hart et al., 2014, Balleine et al., 2007,
Balleine and O'Doherty, 2010, Heidbreder and Groenewegen, 2003) but not to DLS (Voorn et al.,
2004) and the observed increased coherence between directly connected structures likely
represents increased top down regulation associated with task management. In the Go-NoGo Minus
the coherence response to cue onset also differed between structure pairs. However in this task, cue
onset did not appear to affect coherence between PrL and NAc, whereas coherence between PrL
and DMS decreased and coherence between PrL and DLS increased after cue onset (Figure 4-3B). In
the current study coherence differed between tasks already at baseline, which may have influenced
the magnitude and direction of the response to cue onset. However, the task dependent difference
in baseline coherence makes task dependent differences in coherence after cue onset difficult to
interpret. More research is needed to examine the role of behavioural task setup on corticostriatal
coherence.
4.4.5 Conclusions
Firing rate response to cue onset in PrL were only significantly different between trial types in the
Go-NoGo Minus task, where greater inhibition was observed in error trials compared with correct
trials and excitation in both error trials were greater than in CR trials but only excitation in Miss trials
was greater than in Hit trials(Figure 4-3B). Furthermore, both excitatory and inhibitory responses to
cue onset were greater in trials following incorrect response trials (Miss and FA) than correct
response trials (Hit and CR) in both tasks (Figure 4-4). The same pattern was observed in the striatal
subregions (Figure 3-9 & 3-9), suggesting that PrL incorporates contextual information and recent
experience into the regulation of behaviour, rather than strictly promoting or inhibiting behavioural
responses. However, more research is needed to elucidate the role and direction of corticostriatal
interaction in the planning and execution of complex tasks.
The current study is, to my knowledge, the first to simultaneously investigate the role of single
unit coherence between mPFC and DLS, DMS and NAc during execution of complex reward-directed
behaviour. Overall, prestimulus coherence was higher before error trials than correct response trials
in both task and highest in Miss trials (where the rats failed to respond to the Go cue) (Figure 4-5),
suggesting that corticostriatal coherence is associated with future behavioural choice (Figure 4-5),
consistent with the overall pattern of coherence observed between the striatal subregions (Figure 3-

86
10). The high level of coherence between neurons in PrL and in striatal subregions is well in accord
with labelling studies showing strong projections from PrL to ventral and dorsomedial parts of
striatum (Gabbott et al., 2005, Groenewegen et al., 1999, Heidbreder and Groenewegen, 2003). The
current study also found coherence between PrL and DLS, which together with the overall similarity
between trial type dependent baseline coherence between PrL and all three striatal subregions may
indicate joint modulation from other brain areas or interaction between different cortico-striatal-
thalamic circuits. The high prestimulus coherence between PrL and the striatal subregions observed
in Miss trials may reflect low levels of attention to external stimuli (Gusnard and Raichle, 2001,
Herzog et al., 2014) and may be driven by modulatory input from task-negative regions of cortex or
thalamus. However, more research is needed, incorporating single unit recordings from thalamic and
cortical regions that are part of the limbic, associative and motor cortico-striatal-thalamic circuits, as
well as striatum, to fully elucidate the role of corticostriatal interaction during complex behaviour.

87
Chapter 5: Final Discussion
Striatum function is pivotal to the learning and execution of reward-related tasks, with lesion studies
suggesting functional differences between DLS, DMS and NAc (Balleine et al., 2009, Redgrave et al.,
2011). In addition, corticostriatal communication from prelimbic cortex (PrL) to DMS and NAc likely
play a role in the regulation of appetitive behaviour (Groenewegen and Uylings, 2000, Dalley et al.,
2004). Characterizing the dynamic modulation of behaviour imposed by the reward expectation as
well as motor preparation in behaving animals will be a key step to understanding the normal
function of striatum and cortico-striatal projections. The aim of the presented thesis was to examine
single unit responses within PrL, DLS, DMS and NAc, as well as network activity between these
regions, are associated with salience, motor and reward components of cue-responses during
execution of a reward-directed behavioural task.
5.1.1 Summary of conclusions from experimental chapters
In chapter 2, stimulus-evoked LFP responses in DLS were recorded in head fixed rats during
execution of a tactile Go-NoGo task with the aim of assessing whether the level of salience of
sensory input to DLS affects the sensory representation in this structure. In this study, LFP response
to stimulus onset was found to be enhanced in trials in which cue onset was immediately followed
by initiation of motor response (Figure 2-4C), suggesting the observed response is associated with
motor initiation. This accords well with previous research showing that an increase in DLS activity in
rats during execution of the task as result of training (Barnes et al., 2011, Root et al., 2010, Thorn et
al., 2010, Kimchi et al., 2009). However, the enhanced response in False Alarm trials may still have
been caused by the rat, incorrectly, expecting a reward for its response, as previous research has
shown that DLS neurons that responded to movement increased their firing when movement was
paired with reward (Kimchi et al., 2009).
Due to technical challenges, only a limited dataset was collected in this setup. However, the
preliminary results obtained in this chapter did expose limitations of the standard Go-NoGo task: the
cue signalling reward availability also signals to the animal to initiate movement, motor and reward
component of cue-evoked neural responses cannot be distinguished. In addition, the findings in this
chapter emphasise the importance of considering the contribution of the subject’s choice when
analysing effect of reward-paired cues on neuronal activity.
In chapter 3, the observations made in chapter 2 were used to develop a modified version of
the standard Go-NoGo task (Figure 3-1), in which correct responses in both Go and NoGo trials were

88
rewarded (Go-NoGo Plus), allowing comparison of trials with rewarded motor initiation and
rewarded motor suppression, respectively. A second group of rats were trained in the standard Go-
NoGo task (Go-NoGo Minus), where only correct responses in Go trials were rewarded. Single unit
responses were obtained simultaneously from DMS, DLS and NAc to compare responses as well as
coherence between striatal subregions during execution of either of the two Go-NoGo tasks.
Through comparison of the single unit responses to cue onset in these two tasks, this study aimed to
examine the role of individual striatal sub-regions, as well as communication between sub-regions,
on reward expectancy and preparation of motor response during conditioned discrimination. We
hypothesised that neural responses to cue onsets on correct rejection trials compared to other trial
types would be different between tasks, as this was the only trial type for which the outcome
differed between tasks. However, firing rate response to cue onset did not appear to be influenced
by differences in reward expectancy. Instead, firing rate responses to cue onset were overall greater
in error trials compared with trials with subsequent correct response in both tasks (Figure 3-8). A
recent fMRI study in humans found that testing of known stimulus-response-outcome contingencies
is associated with increased activity in distinct striatal regions (Liu et al., 2015). It is possible that the
comparative increase in firing rate response in error trials may reflect a switch from an over-trained
habitual response to more goal-directed approach in order for the rats to test the consistency of
learned stimulus-response contingencies. Firing rate response to cue onset was significantly greater
in trials following error trials (Figure 3-9), which may signify increased attention in trials following
previous unsuccessful behavioural response.
Overall, prestimulus coherence was greater in FA trials as well as in Miss trials compared
with correct response trials, suggesting that coherence is associated with future behavioural choice
(Figure 3-10). Coherence between striatal subregions was found to be particularly high before and
during Miss trials (Figure 3-10). Fluctuations in prestimulus coherence between cortical regions have
been linked to attention (Gusnard and Raichle, 2001, Herzog et al., 2014). The high striatal
prestimulus coherence observed in Miss trials is well in accordance with previous studies examining
the role of cortical coherence and stimulus detection (Boly et al., 2007, Sadaghiani et al., 2010,
Forstmann et al., 2010, Melloni et al., 2007, Shulman et al., 2002, Herzog et al., 2014) and the
coherence between striatal regions may reflect coherence in cortical areas projecting to the
striatum, with mPFC being a prime candidate for the exertion of such top down control (Riga et al.,
2014, Balleine et al., 2009, Christakou et al., 2004, Stefanik et al., 2015, Baker and Ragozzino, 2014,
Thorn and Graybiel, 2014).

89
To address this issue, the study presented in chapter 4 examined the contribution of PrL single unit
activity and coherence between PrL and DMS, DLS and NAc during execution of the same two Go-
NoGo behavioural paradigms presented in chapter 3. Firing rate response to cue onset in PrL were
only significantly different between trial types in the Go-NoGo Minus task, where greater inhibition
was observed in error trials compared with correct trials and excitation in both error trials were
greater than in CR trials but only excitation in Miss trials was greater than in Hit trials (Figure 4-3B).
Furthermore, both excitatory and inhibitory responses to cue onset were greater in trials following
incorrect response trials (Miss and FA) than correct response trials (Hit and CR) in both tasks (Figure
4-4). The same pattern was observed in the striatal subregions (Figure 3-9 & 3-9), suggesting that
mPFC and striatal subregions interact during complex behaviour. This is in accord with lesioning
studies implicating interaction between mPFC and DMS and NAc in planning and execution of tasks
that require switching between multiple stimulus-outcome contingencies (Christakou et al., 2001,
Baker and Ragozzino, 2014).
Overall, prestimulus coherence was higher before error trials than correct response trials in
both task and highest in Miss trials (where the rats failed to respond to the Go cue) (Figure 4-3),
suggesting that corticostriatal coherence is associated with future behavioural choice (Figure 4-3),
consistent with the overall pattern of coherence observed between the striatal subregions (Figure 3-
10). The high level of coherence between neurons in PrL and in striatal subregions is well in accord
with labelling studies showing strong projections from PrL to ventral and dorsomedial parts of
striatum (Gabbott et al., 2005, Groenewegen et al., 1999, Heidbreder and Groenewegen, 2003). The
current study also found coherence between PrL and DLS, which together with the overall similarity
between trial type dependent baseline coherence between PrL and all three striatal subregions may
indicate joint modulation from other brain areas or interaction between different cortico-striatal-
thalamic circuits. The high prestimulus coherence between striatal subregions observed in Miss trials
may reflect low levels of attention to external stimuli (Gusnard and Raichle, 2001, Herzog et al.,
2014) and may be driven by modulatory input from task-negative regions of cortex or thalamus.
5.1.2 Comparison between findings in experimental chapters
Chapter 2 and 3 both examined the contribution of DLS to trial-cues in the Go-NoGo task. In the
study presented in chapter 2, clear trial type dependent differences in cue-evoked LFP responses
were observed, which could be related to either motor initiation or reward expectation. In chapter 3
a modified version of the standard Go-NoGo task was introduced with the expectation that this
modified task would enable verification of whether the response observed in chapter 2 was related
to the motor component of the task or to the reward component of the task. However, in the study

90
presented in chapter 3, trial type dependent differences in firing rate appeared more closely linked
to subsequent correct or incorrect behavioural response than to either motor or reward component.
The electrophysiological data presented in chapter 2 were collected from a single animal and
therefore should be viewed as preliminary. In addition, only LFPs were recorded in chapter 2,
whereas the electrophysiological data presented in chapter 3 and 4 were from single unit recordings,
which in itself limits the comparability between the two datasets (Kajikawa and Schroeder, 2011).
However, the contrasting patterns in trial type dependent differences in response to cue onset may
indicate important differences between the role of the cue in the Go-NoGo task presented in charter
2 and those presented in chapter 3 and 4. In the Go-NoGo task presented in chapter 2 the onset of
the cue signalling trial type, also signalled the beginning of the 1.5 second response interval.
Therefore the motor response (if the rat chose to respond) would always be initiated immediately
after cue onset. In contrast in both Go-NoGo tasks presented in chapter 3 and 4, the onset of the cue
signalling trial type was presented four seconds before the lever was presented, thus a 4 second
waiting period always preceded the 4 second response interval in these tasks. Therefore, vigorous
motor response was not required immediately after cue onset in the tasks presented in chapter 3
and 4. In fact, cue onset was almost always immediately followed by the rat rearing or becoming
immobile before eventually responding (if the rat chose to respond) (observation made by the
experimenter). Whereas the DLS response observed in chapter 2 likely reflects actual motor
initiation, the DLS response observed in chapter 3 may in fact reflect processing related to decision
making prior to initiation or suppression of motor response. In rats trained in a T-maze
discrimination task where direction of the reward was signalled at the beginning of each trial before
the rat was given access to the maze, firing rate response to the initial cues increased in the early
stages of training, but shifted towards the onset of motor response as the rats became over-trained
on the task (Barnes et al., 2011, Barnes et al., 2005). In this T-maze paradigm the rat was always
required to initiate a motor response, and therefore this paradigm cannot elaborate on the role of
motor initiation vs. motor suppression. However, these findings, along with the findings presented in
this thesis, demonstrate how temporal disassociation between cues requiring reward-directed
decision making and task elements associated with motor processing within the experimental design
is pivotal to understanding the neural substrate for reward-directed decision making and behaviour.
The electrophysiological data presented in chapter 3 and 4 were obtained within the same
behavioural task, with chapter 3 focusing on striatal responses and coherence and chapter 4
incorporating single unit recordings from PrL to examine the role of interaction between PrL and the
striatal sub-regions. In both PrL and the striatal sub-regions significant differences between trial
types were primarily observed between trials with subsequent correct and incorrect behavioural

91
response and firing rate responses to cue onset were greater in trials following incorrect response
trials (Miss and FA) than correct response trials (Hit and CR) in both PrL and the striatum. Overall,
prestimulus coherence was higher before error trials than correct response trials in both task and
highest in Miss trials both between striatal subregions (Figure 3-10) and between PrL and the striatal
subregions (Figure 4-3). The similarity in trial type dependent responses to cue onset as well as in
the coherence between PrL and the striatal subregions, which may indicate joint modulation from
other brain areas or interaction between different cortico-striatal-thalamic circuits. The high
prestimulus coherence observed in Miss trials may reflect low levels of attention to external stimuli
(Gusnard and Raichle, 2001, Herzog et al., 2014) and may be driven by modulatory input from task-
negative regions of cortex or thalamus. However, more research is needed, incorporating single unit
recordings from thalamic and cortical regions that are part of the limbic, associative and motor
cortico-striatal-thalamic circuits, as well as striatum, to fully elucidate the role of corticostriatal
interaction during complex behaviour.
The findings in this thesis emphasise the importance of considering the contribution of the
subject’s choice when analysing effect of reward-paired cues on neuronal activity. Preliminary
findings presented in chapter 2 indicated that LFP response in DLS were more strongly associated
with motor initiation than to trial type. Further investigation of single unit responses in DLS, DMS,
NAc and PrL showed that significant differences in response to trial onset cue were predominantly
found between trials with subsequent correct and incorrect behavioural response, possibly
suggesting modulation from other brain areas or interaction between different cortico-striatal-
thalamic circuits.
5.1.3 Future perspectives
Projections from cortex to striatum constitute key segments in the associative, sensory-motor and
limbic cortico-striatal-thalamic circuits (Van Waes et al., 2012, Balleine et al., 2009, Redgrave et al.,
2011). However, to fully understand the role of these circuits, more research into the contribution of
other structures in these circuits is warranted. Like in the striatum, the associative, sensory-motor
and the limbic circuits all project to and from sub-regions of the thalamus (Haber and Calzavara,
2009) and the thalamus may play an important role in modulation of the joint output of these
circuits, with distinct groups of thalamic nuclei likely contributing to different aspects of sensory,
motor, and cognitive processing (Haber and Calzavara, 2009). Traditionally the thalamus has been
regarded primarily as a passive relay station for sensory and motor signals (Fama and Sullivan, 2015).
However, the thalamus is now considered to also contribute to cognitive processes, including
attention, speed of information processing, and memory (Fama and Sullivan, 2015) and gamma

92
coherence between LFPs in mPFC and mediodorsal thalamus in relation to reward delivery has been
found to increase in rats as a result of instrumental learning (Yu et al., 2012). Simultaneous
electrophysiological recordings in striatal sub-regions and thalamic nuclei during execution of
reward-directed learning tasks may provide information on the role of this segment of the cortico-
striatal-thalamic circuits and improve our understanding of how cortico-striatal-thalamic circuits
contribute to the processing of salient sensory input and to the optimisation of behavioural
responses.
Although the majority of striatal cells are medium spiny neurons (MSNs) (Gonzales and
Smith, 2015), acetylcholine release from cholinergic interneurons within striatum is thought to
modulate dopamine transmission by acting at both muscarinic and nicotinic acetylcholine receptors
(Threlfell and Cragg, 2011). The subtypes of muscarinic and nicotinic acetylcholine receptors differ
between dorsal striatum and NAc, thus enabling cholinergic interneurons to modulate dopamine
transmission differently in specific striatal sub-regions (Threlfell and Cragg, 2011), which in turn may
underlie differences between striatal sub-regions in their contribution to behaviour (Aoki et al.,
2015). In rats trained in a set-shifting task, cell type specific lesion of cholinergic interneurons in
either DMS or ventral striatum differentially affected the behaviour of the rats, with DMS lesions
particularly impairing the rat’s ability to pay attention to previously irrelevant cues and ventral
striatum lesions specifically impairing the rat’s ability to pay attention to novel cues (Aoki et al.,
2015). This finding suggests that i) cholinergic interneurons may modulate the behavioural output in
tasks that require retention and shifting between multiple stimulus-response-outcome
contingencies and ii) this modulation may play different roles within distinct striatal sub-regions. The
findings presented in this thesis suggest that changes in neural activity striatal sub-regions are
associated with behavioural choice. However, because relationships between behaviour and
electrophysiological data such as those presented in this thesis are correlational, they cannot
provide definite evidence of causality between the neural activity and behaviour. A logical follow up
to the studies presented in the previous chapters would be to use an optogenetic approach to
specifically deactivate cholinergic interneurons within distinct striatal sub-regions in animals during
choice behaviour, while also recording single unit activity in striatal sub-regions. Selectively
deactivating cholinergic interneurons through optogenetic stimulation in a subset of trials in each
behavioural session would allow examination of cholinergic modulation on both behaviour and cue-
evoked single unit responses.
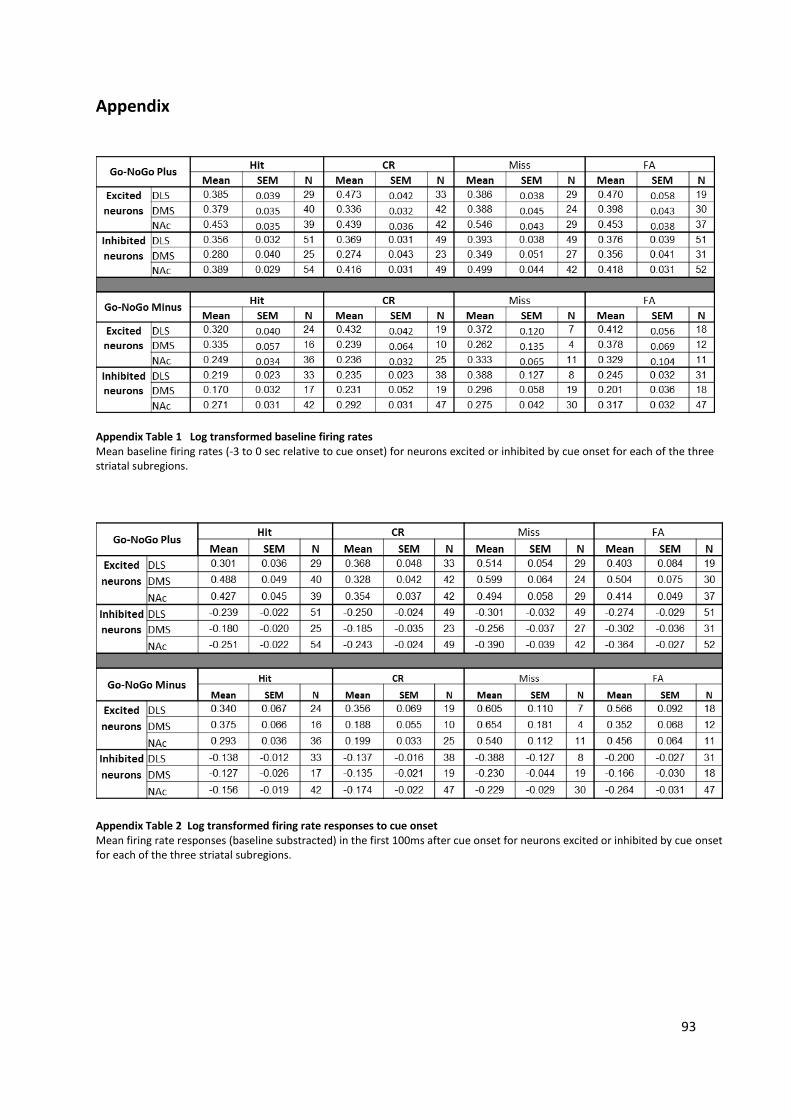
93
Appendix
Appendix Table 1 Log transformed baseline firing rates Mean baseline firing rates (-3 to 0 sec relative to cue onset) for neurons excited or inhibited by cue onset for each of the three striatal subregions.
Appendix Table 2 Log transformed firing rate responses to cue onset Mean firing rate responses (baseline substracted) in the first 100ms after cue onset for neurons excited or inhibited by cue onset for each of the three striatal subregions.
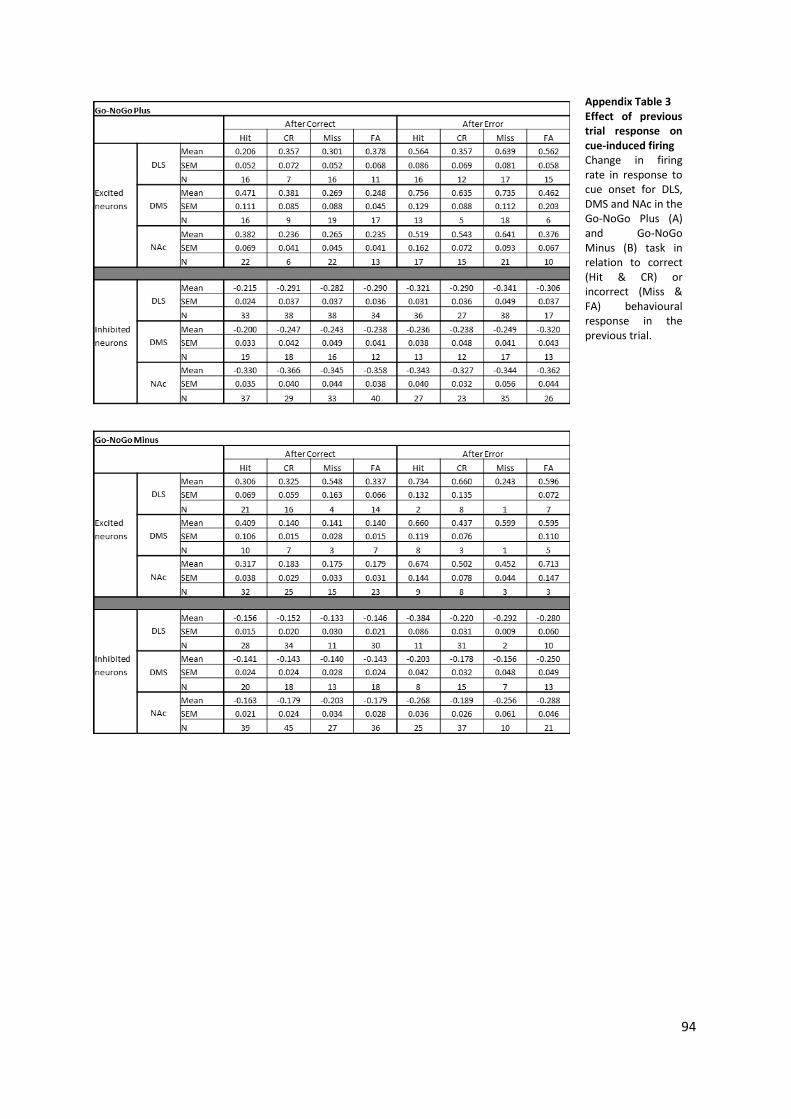
94
Appendix Table 3 Effect of previous trial response on cue-induced firing Change in firing rate in response to cue onset for DLS, DMS and NAc in the Go-NoGo Plus (A) and Go-NoGo Minus (B) task in relation to correct (Hit & CR) or incorrect (Miss & FA) behavioural response in the previous trial.
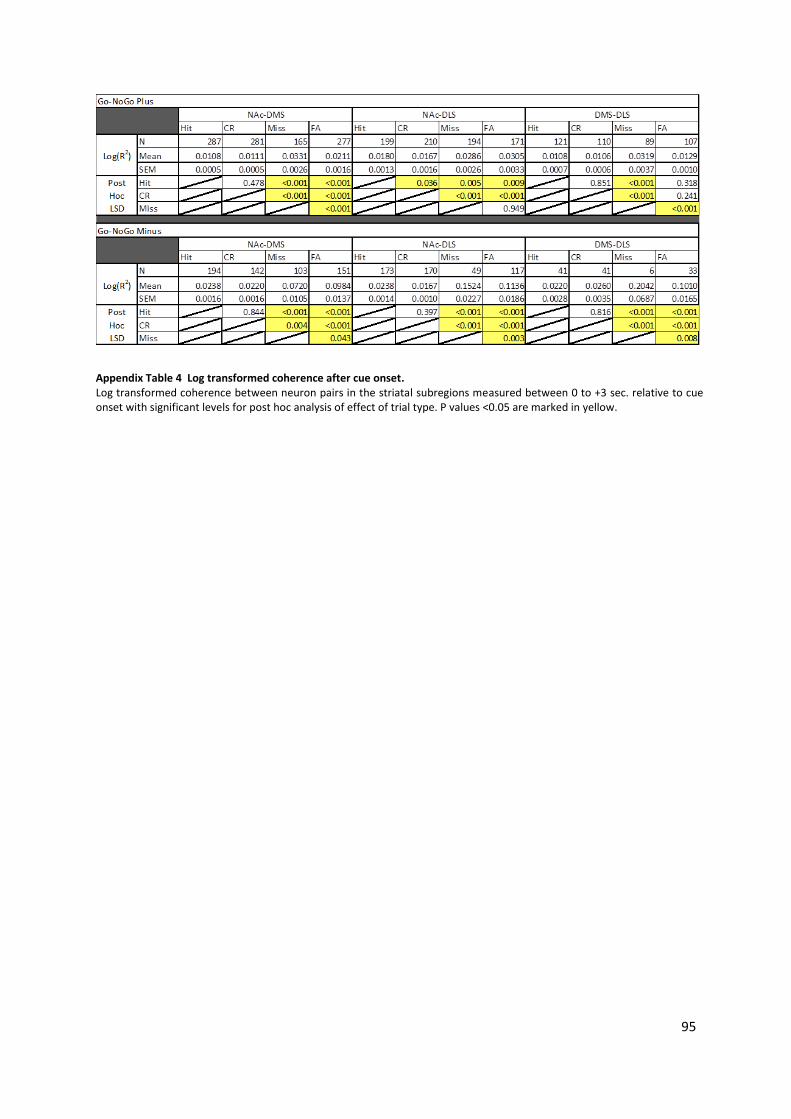
95
Appendix Table 4 Log transformed coherence after cue onset. Log transformed coherence between neuron pairs in the striatal subregions measured between 0 to +3 sec. relative to cue onset with significant levels for post hoc analysis of effect of trial type. P values <0.05 are marked in yellow.

96
References
ALEXANDER, G. E., DELONG, M. R. & STRICK, P. L. 1986. Parallel organization of functionally segregated circuits linking basal ganglia and cortex. Annual Review of Neuroscience, 9, 357-381.
ALLOWAY, K. D., CHRIST, J., MUTIC, J. J. & ROY, S. A. 1999. Corticostriatal Projections from Rat Barrel Cortex Have an Anisotropic Organization that Correlates with Vibrissal Whisking Behavior. Journal of Neuroscience, 19, 10908-10922.
ANTZOULATOS, E. G. & MILLER, E. K. 2014. Increases in functional connectivity between prefrontal cortex and striatum during category learning. Neuron, 83, 216-25.
AOKI, S., LIU, A. W., ZUCCA, A., ZUCCA, S. & WICKENS, J. R. 2015. Role of Striatal Cholinergic Interneurons in Set-Shifting in the Rat. J Neurosci, 35, 9424-31.
AQUILI, L., LIU, A. W., SHINDOU, M., SHINDOU, T. & WICKENS, J. R. 2014. Behavioral flexibility is increased by optogenetic inhibition of neurons in the nucleus accumbens shell during specific time segments. Learn Mem, 21, 223-31.
BAKER, P. M. & RAGOZZINO, M. E. 2014. Contralateral disconnection of the rat prelimbic cortex and dorsomedial striatum impairs cue-guided behavioral switching. Learn Mem, 21, 368-79.
BALDO, B. A. & KELLEY, A. E. 2007. Discrete neurochemical coding of distinguishable motivational processes: insights from nucleus accumbens control of feeding. Psychopharmacology (Berl), 191, 439-59.
BALLEINE, B. W. 2005. Neural bases of food-seeking: affect, arousal and reward in corticostriatolimbic circuits. Physiol Behav, 86, 717-30.
BALLEINE, B. W., DELGADO, M. R. & HIKOSAKA, O. 2007. The role of the dorsal striatum in reward and decision-making. J Neurosci, 27, 8161-5.
BALLEINE, B. W., LILJEHOLM, M. & OSTLUND, S. B. 2009. The integrative function of the basal ganglia in instrumental conditioning. Behav Brain Res, 199, 43-52.
BALLEINE, B. W. & O'DOHERTY, J. P. 2010. Human and rodent homologies in action control: corticostriatal determinants of goal-directed and habitual action. Neuropsychopharmacology, 35, 48-69.
BARNES, T. D., KUBOTA, Y., HU, D., JIN, D. Z. & GRAYBIEL, A. M. 2005. Activity of striatal neurons reflects dynamic encoding and recoding of procedural memories. Nature, 437, 1158-61.
BARNES, T. D., MAO, J. B., HU, D., KUBOTA, Y., DREYER, A. A., STAMOULIS, C., BROWN, E. N. & GRAYBIEL, A. M. 2011. Advance cueing produces enhanced action-boundary patterns of spike activity in the sensorimotor striatum. J Neurophysiol, 105, 1861-78.
BASAR, K., SESIA, T., GROENEWEGEN, H., STEINBUSCH, H. W., VISSER-VANDEWALLE, V. & TEMEL, Y. 2010. Nucleus accumbens and impulsivity. Prog Neurobiol, 92, 533-57.
BASSAREO, V. & DI CHIARA, G. 1997. Differential influence of associative and nonassociative learning mechanisms on the responsiveness of prefrontal and accumbal dopamine transmission to food stimuli in rats fed ad libitum. J Neurosci, 17, 851-861.
BEDWELL, S. A., BILLETT, E. E., CROFTS, J. J. & TINSLEY, C. J. 2014. The topology of connections between rat prefrontal, motor and sensory cortices. Front Syst Neurosci, 8, 177.
BELIN, D., JONKMAN, S., DICKINSON, A., ROBBINS, T. W. & EVERITT, B. J. 2009. Parallel and interactive learning processes within the basal ganglia: relevance for the understanding of addiction. Behav Brain Res, 199, 89-102.
BENUS, R. F., DEN DAAS, S., KOOLHAAS, J. M. & VAN OORTMERSSEN, G. A. 1990. Routine formation and flexibility in social and non-social behaviour of aggressive and non-aggressive male mice. Behaviour, 112, 176-193.
BERKE, J. D., OKATAN, M., SKURSKI, J. & EICHENBAUM, H. B. 2004. Oscillatory entrainment of striatal neurons in freely moving rats. Neuron, 43, 883-96.
BLASIO, A., STEARDO, L., SABINO, V. & COTTONE, P. 2014. Opioid system in the medial prefrontal cortex mediates binge-like eating. Addict Biol, 19, 652-62.

97
BOLY, M., BALTEAU, E., SCHNAKERS, C., DEGUELDRE, C., MOONEN, G., LUXEN, A., PHILLIPS, C., PEIGNEUX, P., MAQUET, P. & LAUREYS, S. 2007. Baseline brain activity fluctuations predict somatosensory perception in humans. Proc Natl Acad Sci U S A, 104, 12187-92.
BOUCHER, L., PALMERI, T. J., LOGAN, G. D. & SCHALL, J. D. 2007. Inhibitory control in mind and brain: an interactive race model of countermanding saccades. Psychol Rev, 114, 376-97.
BROWN, H. D., MCCUTCHEON, J. E., CONE, J. J., RAGOZZINO, M. E. & ROITMAN, M. F. 2011. Primary food reward and reward-predictive stimuli evoke different patterns of phasic dopamine signaling throughout the striatum. Eur J Neurosci, 34, 1997-2006.
BURGOS-ROBLES, A., BRAVO-RIVERA, H. & QUIRK, G. J. 2013. Prelimbic and infralimbic neurons signal distinct aspects of appetitive instrumental behavior. PLoS One, 8, e57575.
BUZSAKI, G. & MIZUSEKI, K. 2014. The log-dynamic brain: how skewed distributions affect network operations. Nat Rev Neurosci, 15, 264-78.
CARRELLI, R. M. 2002. Nucleus accumbens cell firing during goal-directed behaviors for cocaine vs. 'natural' reinforcement. Physiol Behav, 76, 379-387.
CHRISTAKOU, A., ROBBINS, T. W. & EVERITT, B. J. 2001. Functional disconnection of a prefrontal cortical-dorsal striatal system disrupts choice reaction time performance: Implications for attentional function. Behavioral Neuroscience, 115, 812-825.
CHRISTAKOU, A., ROBBINS, T. W. & EVERITT, B. J. 2004. Prefrontal cortical-ventral striatal interactions involved in affective modulation of attentional performance: implications for corticostriatal circuit function. J Neurosci, 24, 773-80.
COHEN, M. X. 2007. Individual differences and the neural representations of reward expectation and reward prediction error. Soc Cogn Affect Neurosci, 2, 20-30.
COHEN, M. X., BOUR, L., MANTIONE, M., FIGEE, M., VINK, M., TIJSSEN, M. A., VAN ROOTSELAAR, A. F., VAN DEN MUNCKHOF, P., SCHUURMAN, P. R. & DENYS, D. 2012. Top-down-directed synchrony from medial frontal cortex to nucleus accumbens during reward anticipation. Hum Brain Mapp, 33, 246-52.
COLE, S., MAYER, H. S. & PETROVICH, G. D. 2015. Orexin/Hypocretin-1 Receptor Antagonism Selectively Reduces Cue-Induced Feeding in Sated Rats and Recruits Medial Prefrontal Cortex and Thalamus. Sci Rep, 5, 16143.
COLLINS, K. A., MENDELSOHN, A., CAIN, C. K. & SCHILLER, D. 2014. Taking action in the face of threat: neural synchronization predicts adaptive coping. J Neurosci, 34, 14733-8.
COOPER, N. R., CROFT, R. J., DOMINEY, S. J. J., BURGESS, A. P. & GRUZELIER, J. H. 2003. Paradox lost? Exploring the role of alpha oscillations during externally vs. internally directed attention and the implications for idling and inhibition hypotheses. Int J Psychophysiol, 47, 65-74.
COPPENS, C. M., DE BOER, S. F. & KOOLHAAS, J. M. 2010. Coping styles and behavioural flexibility: towards underlying mechanisms. Philos Trans R Soc Lond B Biol Sci, 365, 4021-8.
COSTA, R. M., COHEN, D. & NICOLELIS, M. A. 2004. Differential corticostriatal plasticity during fast and slow motor skill learning in mice. Curr Biol, 14, 1124-34.
CURZON, P., BANNON, A. W. & DECKER, M. W. 1999. Effect of 192IgG-saporin injection into the nucleus basalis magnocellularis on acquisition and performance of a go/no-go procedure in the rat. Psychobiology, 27, 114-122.
DALLEY, J. W., CARDINAL, R. N. & ROBBINS, T. W. 2004. Prefrontal executive and cognitive functions in rodents: neural and neurochemical substrates. Neurosci Biobehav Rev, 28, 771-84.
DEVAN, B. D., HONG, N. S. & MCDONALD, R. J. 2011. Parallel associative processing in the dorsal striatum: segregation of stimulus-response and cognitive control subregions. Neurobiol Learn Mem, 96, 95-120.
DIAMOND, M. E. & ARABZADEH, E. 2013. Whisker sensory system - from receptor to decision. Prog Neurobiol, 103, 28-40.
DIAS-FERREIRA, E., SOUSA, J. C., MELO, I., MORGADO, P., MESQUITA, A. R., CERQUEIRA, J. J., COSTA, R. M. & SOUSA, N. 2009. Chronic stress causes frontostriatal reorganization and affects decision-making. Science, 325, 621-5.

98
DING, D. C., GABBOTT, P. L. & TOTTERDELL, S. 2001. Differences in the laminar origin of projections from the medial prefrontal cortex to the nucleus accumbens shell and core regions in the rat. brain Research, 917, 81-89.
DONNELLY, N. A., PAULSEN, O., ROBBINS, T. W. & DALLEY, J. W. 2015. Ramping single unit activity in the medial prefrontal cortex and ventral striatum reflects the onset of waiting but not imminent impulsive actions. Eur J Neurosci, 41, 1524-37.
EAGLE, D. M. & ROBBINS, T. W. 2003. Lesions of the medial prefrontal cortex or nucleus accumbens core do not impair inhibitory control in rats performing a stop-signal reaction time task. Behav Brain Res, 146, 131-144.
FAMA, R. & SULLIVAN, E. V. 2015. Thalamic structures and associated cognitive functions: Relations with age and aging. Neurosci Biobehav Rev, 54, 29-37.
FANELLI, R. R., KLEIN, J. T., REESE, R. M. & ROBINSON, D. L. 2013. Dorsomedial and dorsolateral striatum exhibit distinct phasic neuronal activity during alcohol self-administration in rats. Eur J Neurosci, 38, 2637-48.
FEATHERSTONE, R. E. & MCDONALD, R. J. 2004. Dorsal striatum and stimulus–response learning: lesions of the dorsolateral, but not dorsomedial, striatum impair acquisition of a simple discrimination task. Behavioural Brain Research, 150, 15-23.
FEATHERSTONE, R. E. & MCDONALD, R. J. 2005. Lesions of the dorsolateral striatum impair the acquisition of a simplified stimulus-response dependent conditional discrimination task. Neuroscience, 136, 387-95.
FEJA, M., HAYN, L. & KOCH, M. 2014. Nucleus accumbens core and shell inactivation differentially affects impulsive behaviours in rats. Prog Neuropsychopharmacol Biol Psychiatry, 54, 31-42.
FORSTMANN, B. U., ANWARDER, A., SCHAFER, A., NEUMANN, J., BROWN, S., WAGENMAKERS, E., BOGACZ, R. & TURNER, R. 2010. Cortico-striatal connections predict control over speed and accuracy in perceptual decision making. PNAS, 107, 15916-15920.
FUNAMIZU, A., ITO, M., DOYA, K., KANZAKI, R. & TAKAHASHI, H. 2015. Condition interference in rats performing a choice task with switched variable- and fixed-reward conditions. Front Neurosci, 9, 27.
GABBOTT, P. L., WARNER, T. A., JAYS, P. R., SALWAY, P. & BUSBY, S. J. 2005. Prefrontal cortex in the rat: projections to subcortical autonomic, motor, and limbic centers. J Comp Neurol, 492, 145-77.
GLENN, A. L. & YANG, Y. 2012. The potential role of the striatum in antisocial behavior and psychopathy. Biol Psychiatry, 72, 817-22.
GODA, S. A., OLSZEWSKI, M., PIASECKA, J., REJNIAK, K., WHITTINGTON, M. A., KASICKI, S. & HUNT, M. J. 2015. Aberrant high frequency oscillations recorded in the rat nucleus accumbens in the methylazoxymethanol acetate neurodevelopmental model of schizophrenia. Prog Neuropsychopharmacol Biol Psychiatry, 61, 44-51.
GONZALES, K. K. & SMITH, Y. 2015. Cholinergic interneurons in the dorsal and ventral striatum: anatomical and functional considerations in normal and diseased conditions. Ann N Y Acad Sci, 1349, 1-45.
GRACE, A. A., FLORESCO, S. B., GOTO, Y. & LODGE, D. J. 2007. Regulation of firing of dopaminergic neurons and control of goal-directed behaviors. Trends Neurosci, 30, 220-7.
GRAYBIEL, A. M., AOSAKI, T., FLAHERTY, A. W. & KIMURA, M. 1994. The Basal ganglia and adaptive motor control. Science, 23, 1826-1831.
GROENEWEGEN, H. J. & UYLINGS, H. B. 2000. The prefrontal cortex and the integration of sensory, limbic and autonomic information. Prog Brain Res, 126, 3-28.
GROENEWEGEN, H. J., WRIGHT, C. I., BEIJER, A. V. J. & VOORN, P. 1999. Convergence and segregation of ventral striatal inputs and outputs. Ann N Y Acad Sci, 877, 49-63.
GUSNARD, D. A. & RAICHLE, M. E. 2001. Searching for a baseline: functional imaging and the resting human brain. Nat Rev Neurosci, 2, 685-694.

99
HABER, S. N. 2003. The primate basal ganglia: parallel and integrative networks. Journal of Chemical Neuroanatomy, 26, 317-330.
HABER, S. N. & CALZAVARA, R. 2009. The cortico-basal ganglia integrative network: the role of the thalamus. Brain Res Bull, 78, 69-74.
HABER, S. N., FUDGE, J. L. & MCFARLAND, N. R. 2000. Striatonigrostriatal Pathways in Primates Form an Ascending Spiral from the Shell to the Dorsolateral Striatum. Journal of Neuroscience, 20, 2369-82.
HABER, S. N. & KNUTSON, B. 2010. The reward circuit: linking primate anatomy and human imaging. Neuropsychopharmacology, 35, 4-26.
HADAD, N. A. & KNACKSTEDT, L. A. 2014. Addicted to palatable foods: comparing the neurobiology of Bulimia Nervosa to that of drug addiction. Psychopharmacology (Berl), 231, 1897-912.
HAISS, F., BUTOVAS, S. & SCHWARZ, C. 2010. A miniaturized chronic microelectrode drive for awake behaving head restrained mice and rats. J Neurosci Methods, 187, 67-72.
HALLADAY, L. R. & BLAIR, H. T. 2015. Distinct ensembles of medial prefrontal cortex neurons are activated by threatening stimuli that elicit excitation versus inhibition of movement. J Neurophysiol, jn 00656 2014.
HALLIDAY, D. M. 2015. Nonparametric directionality measures for time series and point process data. J Integr Neurosci, 14, 253-77.
HANES, D. P. & SCHALL, J. D. 1996. Neural control of voluntary movement initiation. Science, 274, 427-430.
HANSLMAYR, S., ASLAN, A., STAUDIGL, T., KLIMESCH, W., HERRMANN, C. S. & BAUML, K. H. 2007. Prestimulus oscillations predict visual perception performance between and within subjects. Neuroimage, 37, 1465-73.
HARDING, E. J., PAUL, E. S. & MENDL, M. 2004. Animal behaviour: Cognitive bias and affective state. Nature, 22, 312.
HART, G., LEUNG, B. K. & BALLEINE, B. W. 2014. Dorsal and ventral streams: the distinct role of striatal subregions in the acquisition and performance of goal-directed actions. Neurobiol Learn Mem, 108, 104-18.
HASSANI, O. K., CROMWELL, H. C. & SCHULTZ, W. 2001. Influence of Expectation of Different Rewards on Behaviour-Related Neuronal Activity in the Striatum. J Neurophysiol, 85, 2477-89.
HAWKING, T. G. & GERDJIKOV, T. V. 2013. Populations of striatal medium spiny neurons encode vibrotactile frequency in rats: modulation by slow wave oscillations. J Neurophysiol, 109, 315-20.
HEARING, M. C., ZINK, A. N. & WICKMAN, K. 2012. Cocaine-induced adaptations in metabotropic inhibitory signaling in the mesocorticolimbic system. Rev Neurosci, 23, 325-51.
HEIDBREDER, C. A. & GROENEWEGEN, H. J. 2003. The medial prefrontal cortex in the rat: evidence for a dorso-ventral distinction based upon functional and anatomical characteristics. Neuroscience & Biobehavioral Reviews, 27, 555-579.
HERZOG, L., SALEHI, K., BOHON, K. S. & WIEST, M. C. 2014. Prestimulus frontal-parietal coherence predicts auditory detection performance in rats. J Neurophysiol, 111, 1986-2000.
HILARIO, M., HOLLOWAY, T., JIN, X. & COSTA, R. M. 2012. Different dorsal striatum circuits mediate action discrimination and action generalization. Eur J Neurosci, 35, 1105-14.
HOFFER, Z. S., ARANTES, H. B., ROTH, R. L. & ALLOWAY, K. D. 2005. Functional circuits mediating sensorimotor integration: quantitative comparisons of projections from rodent barrel cortex to primary motor cortex, neostriatum, superior colliculus, and the pons. J Comp Neurol, 488, 82-100.
HOOVER, W. B. & VERTES, R. P. 2007. Anatomical analysis of afferent projections to the medial prefrontal cortex in the rat. Brain Struct Funct, 212, 149-79.
HORVITZ, J. C. 2009. Stimulus-response and response-outcome learning mechanisms in the striatum. Behav Brain Res, 199, 129-40.

100
HOSKING, J. G., COCKER, P. J. & WINSTANLEY, C. A. 2015. Prefrontal Cortical Inactivations Decrease Willingness to Expend Cognitive Effort on a Rodent Cost/Benefit Decision-Making Task. Cereb Cortex.
ITO, M. & DOYA, K. 2015. Distinct neural representation in the dorsolateral, dorsomedial, and ventral parts of the striatum during fixed- and free-choice tasks. J Neurosci, 35, 3499-514.
JIN, X., TECUAPETLA, F. & COSTA, R. M. 2014. Basal ganglia subcircuits distinctively encode the parsing and concatenation of action sequences. Nat Neurosci, 17, 423-30.
JOEL, D. & WEINER, I. 2000. The connections of the dopaminergic system with the striatum in rats and primates: an analysis with respect to the functional and compartmental organization of the striatum. Neuroscience, 96, 451-474.
JONKMAN, S., MAR, A. C., DICKINSON, A., ROBBINS, T. W. & EVERITT, B. J. 2009. The rat prelimbic cortex mediates inhibitory response control but not the consolidation of instrumental learning. Behav Neurosci, 123, 875-85.
KAJIKAWA, Y. & SCHROEDER, C. E. 2011. How local is the local field potential? Neuron, 72, 847-58. KAWAGOE, R., TAKIKAWA, Y. & HIKOSAKA, O. 1998. Expectation of reward modulates cognitive
signals in the basal ganglia. Nature Neuroscience, 1, 411-416. KIM, H., SUL, J. H., HUH, N., LEE, D. & JUNG, M. W. 2009. Role of striatum in updating values of
chosen actions. J Neurosci, 29, 14701-12. KIMCHI, E. Y., TORREGROSSA, M. M., TAYLOR, J. R. & LAUBACH, M. 2009. Neuronal correlates of
instrumental learning in the dorsal striatum. J Neurophysiol, 102, 475-89. KORALEK, A. C., COSTA, R. M. & CARMENA, J. M. 2013. Temporally precise cell-specific coherence
develops in corticostriatal networks during learning. Neuron, 79, 865-72. LEPSKI, G., ARÉVALO, A., VALLE, A. C. D., BALLESTER, G. & GHARABAGHI, A. 2012. Increased
coherence among striatal regions in the theta range during attentive wakefulness. Brazilian Journal of Medical and Biological Research, 45, 763-770.
LI, C. S., HUANG, C., CONSTABLE, R. T. & SINHA, R. 2006. Imaging response inhibition in a stop-signal task: neural correlates independent of signal monitoring and post-response processing. J Neurosci, 26, 186-92.
LILJEHOLM, M. & O'DOHERTY, J. P. 2012. Contributions of the striatum to learning, motivation, and performance: an associative account. Trends Cogn Sci, 16, 467-75.
LINDGREN, H. S., WICKENS, R., TAIT, D. S., BROWN, V. J. & DUNNETT, S. B. 2013. Lesions of the dorsomedial striatum impair formation of attentional set in rats. Neuropharmacology, 71, 148-53.
LITTLE, J. P. & CARTER, A. G. 2013. Synaptic mechanisms underlying strong reciprocal connectivity between the medial prefrontal cortex and basolateral amygdala. J Neurosci, 33, 15333-42.
LITVIN, Y., BLANCHARD, D. C. & BLANCHARD, R. J. 2007. Rat 22kHz ultrasonic vocalizations as alarm cries. Behav Brain Res, 182, 166-72.
LIU, Z., BRAUNLICH, K., WEHE, H. S. & SEGER, C. A. 2015. Neural networks supporting switching, hypothesis testing, and rule application. Neuropsychologia.
LUTSIV, O., MAH, J., BEYENE, J. & MCDONALD, S. D. 2015. The effects of morbid obesity on maternal and neonatal health outcomes: a systematic review and meta-analyses. Obes Rev, 16, 531-46.
MAIER, S. F. 2015. Behavioral control blunts reactions to contemporaneous and future adverse events: medial prefrontal cortex plasticity and a corticostriatal network. Neurobiol Stress, 1, 12-22.
MARCOTT, P. F., MAMALIGAS, A. A. & FORD, C. P. 2014. Phasic dopamine release drives rapid activation of striatal D2-receptors. Neuron, 84, 164-76.
MATTINSON, C. E., BURMEISTER, J. J., QUINTERO, J. E., POMERLEAU, F., HUETTL, P. & GERHARDT, G. A. 2011. Tonic and phasic release of glutamate and acetylcholine neurotransmission in sub-regions of the rat prefrontal cortex using enzyme-based microelectrode arrays. J Neurosci Methods, 202, 199-208.

101
MAZAHERI, A., DIQUATTRO, N. E., BENGSON, J. & GENG, J. J. 2011. Pre-stimulus activity predicts the winner of top-down vs. bottom-up attentional selection. PLoS One, 6, e16243.
MCCUTCHEON, J. E. 2015. The role of dopamine in the pursuit of nutritional value. Physiol Behav, 152, 408-15.
MELLONI, L., MOLINA, C., PENA, M., TORRES, D., SINGER, W. & RODRIGUEZ, E. 2007. Synchronization of neural activity across cortical areas correlates with conscious perception. J Neurosci, 27, 2858-65.
MIYASHITA, T. & FELDMAN, D. E. 2013. Behavioral detection of passive whisker stimuli requires somatosensory cortex. Cereb Cortex, 23, 1655-62.
MOORMAN, D. E. & ASTON-JONES, G. 2015. Prefrontal neurons encode context-based response execution and inhibition in reward seeking and extinction. PNAS, 112, 9472-9477.
MOWERY, T. M., HARROLD, J. B. & ALLOWAY, K. D. 2011. Repeated whisker stimulation evokes invariant neuronal responses in the dorsolateral striatum of anesthetized rats: a potential correlate of sensorimotor habits. J Neurophysiol, 105, 2225-38.
MULDER, A. B., NORDQUIST, R. E., ÖRGÜT, O. & PENNARTZ, C. M. A. 2003. Learning-related changes in response patterns of prefrontal neurons during instrumental conditioning. Behavioural Brain Research, 146, 77-88.
NAEF, L., PITMAN, K. A. & BORGLAND, S. L. 2015. Mesolimbic dopamine and its neuromodulators in obesity and binge eating. CNS Spectr, 20, 574-83.
NAKHNIKIAN, A., REBEC, G. V., GRASSE, L. M., DWIEL, L. L., SHIMONO, M. & BEGGS, J. M. 2014. Behavior modulates effective connectivity between cortex and striatum. PLoS One, 9, e89443.
NICOLA, S. M., YUN, I. A., WAKABAYASHI, K. T. & FIELDS, H. L. 2004a. Cue-evoked firing of nucleus accumbens neurons encodes motivational significance during a discriminative stimulus task. j Neurophysiol, 91, 1840-1865.
NICOLA, S. M., YUN, I. A., WAKABAYASHI, K. T. & FIELDS, H. L. 2004b. Firing of Nucleus Accumbens Neurons During the Consummatory Phase of a Discriminative Stimulus Tak Depends on Previous Reward Predictive Cues. J Neurophysiol, 91, 1866-82.
NUNES, E. J., RANDALL, P. A., PODURGIEL, S., CORREA, M. & SALAMONE, J. D. 2013. Nucleus accumbens neurotransmission and effort-related choice behavior in food motivation: effects of drugs acting on dopamine, adenosine, and muscarinic acetylcholine receptors. Neurosci Biobehav Rev, 37, 2015-25.
OLLERENSHAW, D. R., ZHENG, H. J., MILLARD, D. C., WANG, Q. & STANLEY, G. B. 2014. The Adaptive Trade-Off between Detection and Discrimination in Cortical Representations and Behavior. Neuron, 81, 1152-64.
OSTLUND, S. B. & BALLEINE, B. W. 2005. Lesions of medial prefrontal cortex disrupt the acquisition but not the expression of goal-directed learning. J Neurosci, 25, 7763-70.
OYAMA, K., TATEYAMA, Y., HERNADI, I., TOBLER, P. N., IIJIMA, T. & TSUTSUI, K. 2015. Discrete coding of stimulus value, reward expectation, and reward prediction error in the dorsal striatum. J Neurophysiol, 114, 2600-15.
PASQUEREAU, B., NADJAR, A., ARKADIR, D., BEZARD, E., GOILLANDEAU, M., BIOULAC, B., GROSS, C. E. & BORAUD, T. 2007. Shaping of motor responses by incentive values through the basal ganglia. J Neurosci, 27, 1176-83.
PAXINOS, G. & WATSON, C. 2007. The Rat Brain In Stereotaxic Coordinates. 6th Edition ed.: Academic Press.
PIDOUX, M., MAHON, S., DENIAU, J. M. & CHARPIER, S. 2011. Integration and propagation of somatosensory responses in the corticostriatal pathway: an intracellular study in vivo. J Physiol, 589, 263-81.
PISA, M. & SCHRANZ, J. A. 1988. Dissociable motor roles of the rat's striatum conform to a somatotopic model. Behav Neurosci, 102, 429-40.

102
POTHUIZEN, H. H., JONGEN-RELO, A. L., FELDON, J. & YEE, B. K. 2005. Double dissociation of the effects of selective nucleus accumbens core and shell lesions on impulsive-choice behaviour and salience learning in rats. Eur J Neurosci, 22, 2605-16.
PRASAD, A., XUE, Q., SANKAR, V., NISHIDA, T., SHAW, G., STREIT, W. J. & SANCHEZ, J. C. 2012. Characterization and failure modes of tungsten microwire arrays in chronic neural implants. Journal of Neural Engineering, 9, 1-21.
QUIROGA, R. Q., KREUZ, T. & GRASSBERGER, P. 2002. Event synchronization: a simple and fast method to measure synchronicity and time delay patterns. Physical review. E, Statistical, nonliniar, and soft matter physics, 66, 1-6.
QUIROGA, R. Q., NADASDY, Z. & BEN-SHAUL, Y. 2004. Unsupervised Spike Detection and Sorting with Wavelets and Superparamagnetic Clustering. Neural Computation, 16, 1661-1687.
REDGRAVE, P., RODRIGUEZ, M., SMITH, Y., RODRIGUEZ-OROZ, M. C., LEHERICY, S., BERGMAN, H., AGID, Y., DELONG, M. R. & OBESO, J. A. 2010. Goal-directed and habitual control in the basal ganglia: implications for Parkinson's disease. Nat Rev Neurosci, 11, 760-72.
REDGRAVE, P., VAUTRELLE, N. & REYNOLDS, J. N. 2011. Functional properties of the basal ganglia's re-entrant loop architecture: selection and reinforcement. Neuroscience, 198, 138-51.
RIGA, D., MATOS, M. R., GLAS, A., SMIT, A. B., SPIJKER, S. & VAN DEN OEVER, M. C. 2014. Optogenetic dissection of medial prefrontal cortex circuitry. Front Syst Neurosci, 8, 230.
ROGERS, R. D., BAUNEZ, C., EVERITT, B. J. & ROBBINS, T. W. 2001. Lesions of the medial and lateral striatum in the rat produce differential deficits in attentional performance. Behavioral Neuroscience, 115, 799-811.
ROITMAN, J. D. & LORIAUX, A. L. 2014. Nucleus accumbens responses differentiate execution and restraint in reward-directed behavior. Journal of Neurophysiology, 111, 350-360.
ROITMAN, M. F., WHEELER, R. A. & CARELLI, R. M. 2005. Nucleus accumbens neurons are innately tuned for rewarding and aversive taste stimuli, encode their predictors, and are linked to motor output. Neuron, 45, 587-97.
ROOT, D. H., TANG, C. C., MA, S., PAWLAK, A. P. & WEST, M. O. 2010. Absence of cue-evoked firing in rat dorsolateral striatum neurons. Behav Brain Res, 211, 23-32.
SADAGHIANI, S., HESSELMANN, G., FRISTON, K. J. & KLEINSCHMIDT, A. 2010. The relation of ongoing brain activity, evoked neural responses, and cognition. Front Syst Neurosci, 4, 20.
SAMEJIMA, K., UEDA, M., DOYA, K. & KIMURA, M. 2005. Representation of action-specific reward values in teh striatum. Science, 310, 1337-1340.
SCHALL, J. D. & GODLOVE, D. C. 2012. Current advances and pressing problems in studies of stopping. Curr Opin Neurobiol, 22, 1012-21.
SCHMITZER-TORBERT, N., APOSTOLIDIS, S., AMOA, R., O'REAR, C., KASTER, M., STOWERS, J. & RITZ, R. 2015. Post-training cocaine administration facilitates habit learning and requires the infralimbic cortex and dorsolateral striatum. Neurobiol Learn Mem, 118, 105-12.
SCHWARZ, C., HENTSCHKE, H., BUTOVAS, S., HAISS, F., STUTTGEN, M. C., GERDJIKOV, T. V., BERGNER, C. G. & WAIBLINGER, C. 2010. The head-fixed behaving rat--procedures and pitfalls. Somatosens Mot Res, 27, 131-48.
SCLAFANI, A., TOUZANI, K. & BODNAR, R. J. 2011. Dopamine and learned food preferences. Physiol Behav, 104, 64-8.
SHAN, Q., GE, M., CHRISTIE, M. J. & BALLEINE, B. W. 2014. The acquisition of goal-directed actions generates opposing plasticity in direct and indirect pathways in dorsomedial striatum. J Neurosci, 34, 9196-201.
SHAROTT, A., MAGILL, P. J., BOLAM, J. P. & BROWN, P. 2005. Directional analysis of coherent oscillatory field potentials in the cerebral cortex and basal ganglia of the rat. J Physiol, 562, 951-63.
SHIFLETT, M. W., BROWN, R. A. & BALLEINE, B. W. 2010. Acquisition and performance of goal-directed instrumental actions depends on ERK signaling in distinct regions of dorsal striatum in rats. J Neurosci, 30, 2951-9.

103
SHULMAN, G. L., TANSY, A. P., KINCADE, M., PETERSEN, S. E., MCAVOY, M. P. & CORBETTA, M. 2002. Reactivation of Networks Involved in Preparatory States. Cereb Cortex, 12, 590-600.
SMITH, K. S. & GRAYBIEL, A. M. 2013. A dual operator view of habitual behavior reflecting cortical and striatal dynamics. Neuron, 79, 361-74.
SORG, C., MANOLIU, A., NEUFANG, S., MYERS, N., PETERS, H., SCHWERTHOFFER, D., SCHERR, M., MUHLAU, M., ZIMMER, C., DRZEZGA, A., FORSTL, H., BAUML, J., EICHELE, T., WOHLSCHLAGER, A. M. & RIEDL, V. 2012. Increased Intrinsic Brain Activity in the Striatum Reflects Symptom Dimensions in Schizophrenia. Schizophr Bull.
STALNAKER, T. A., CALHOON, G. G., OGAWA, M., ROESCH, M. R. & SCHOENBAUM, G. 2012. Reward prediction error signaling in posterior dorsomedial striatum is action specific. J Neurosci, 32, 10296-305.
STEFANIK, M. T., KUPCHIK, Y. M. & KALIVAS, P. W. 2015. Optogenetic inhibition of cortical afferents in the nucleus accumbens simultaneously prevents cue-induced transient synaptic potentiation and cocaine-seeking behavior. Brain Struct Funct.
STEKETEE, J. 2003. Neurotransmitter systems of the medial prefrontal cortex: potential role in sensitization to psychostimulants. Brain Research Reviews, 41, 203-228.
STUTTGEN, M. C. & SCHWARZ, C. 2010. Integration of vibrotactile signals for whisker-related perception in rats is governed by short time constants: comparison of neurometric and psychometric detection performance. J Neurosci, 30, 2060-9.
SUPER, H., VAN DER TOGT, C., SPEKREIJSE, H. & LAMME, V. A. F. 2003. Internal State of Monkey Primary Visual Cortex (V1) Predicts Figure–Ground Perception. J Neurosci, 23, 3407-3414.
SYED, E. C., GRIMA, L. L., MAGILL, P. J., BOGACZ, R., BROWN, P. & WALTON, M. E. 2016. Action initiation shapes mesolimbic dopamine encoding of future rewards. Nat Neurosci, 19, 34-6.
SYED, E. C., SHAROTT, A., MOLL, C. K., ENGEL, A. K. & KRAL, A. 2011. Effect of sensory stimulation in rat barrel cortex, dorsolateral striatum and on corticostriatal functional connectivity. Eur J Neurosci, 33, 461-70.
TANG, C., PAWLAK, A. P., PROKOPENKO, V. & WEST, M. O. 2007. Changes in activity of the striatum during formation of a motor habit. Eur J Neurosci, 25, 1212-27.
TANG, C. C., ROOT, D. H., DUKE, D. C., ZHU, Y., TEIXERIA, K., MA, S., BARKER, D. J. & WEST, M. O. 2009. Decreased firing of striatal neurons related to licking during acquisition and overtraining of a licking task. J Neurosci, 29, 13952-61.
THORN, C. A., ATALLAH, H., HOWE, M. & GRAYBIEL, A. M. 2010. Differential dynamics of activity changes in dorsolateral and dorsomedial striatal loops during learning. Neuron, 66, 781-95.
THORN, C. A. & GRAYBIEL, A. M. 2014. Differential entrainment and learning-related dynamics of spike and local field potential activity in the sensorimotor and associative striatum. J Neurosci, 34, 2845-59.
THRELFELL, S. & CRAGG, S. J. 2011. Dopamine signaling in dorsal versus ventral striatum: the dynamic role of cholinergic interneurons. Front Syst Neurosci, 5, 11.
TOTAH, N. K., JACKSON, M. E. & MOGHADDAM, B. 2013. Preparatory attention relies on dynamic interactions between prelimbic cortex and anterior cingulate cortex. Cereb Cortex, 23, 729-38.
TRAN-TU-YEN, D. A., MARCHAND, A. R., PAPE, J. R., DI SCALA, G. & COUTUREAU, E. 2009. Transient role of the rat prelimbic cortex in goal-directed behaviour. Eur J Neurosci, 30, 464-71.
TRICOMI, E. & LEMPERT, K. M. 2015. Value and probability coding in a feedback-based learning task utilizing food rewards. J Neurophysiol, 113, 4-13.
VAN DIJK, H., SCHOFFELEN, J. M., OOSTENVELD, R. & JENSEN, O. 2008. Prestimulus oscillatory activity in the alpha band predicts visual discrimination ability. J Neurosci, 28, 1816-23.
VAN WAES, V., BEVERLEY, J. A., SIMAN, H., TSENG, K. Y. & STEINER, H. 2012. CB1 Cannabinoid Receptor Expression in the Striatum: Association with Corticostriatal Circuits and Developmental Regulation. Front Pharmacol, 3, 21.

104
VERBRUGGEN, F. & LOGAN, G. D. 2009. Models of response inhibition in the stop-signal and stop-change paradigms. Neurosci Biobehav Rev, 33, 647-61.
VERTES, R. P., HOOVER, W. B. & RODRIGUEZ, J. J. 2012. Projections of the central medial nucleus of the thalamus in the rat: node in cortical, striatal and limbic forebrain circuitry. Neuroscience, 219, 120-36.
VOORN, P., VANDERSCHUREN, L. J., GROENEWEGEN, H. J., ROBBINS, T. W. & PENNARTZ, C. M. 2004. Putting a spin on the dorsal-ventral divide of the striatum. Trends Neurosci, 27, 468-74.
WANG, A. Y., MIURA, K. & UCHIDA, N. 2013. The dorsomedial striatum encodes net expected return, critical for energizing performance vigor. Nat Neurosci, 16, 639-47.
WILLUHN, I., BURGENO, L. M., EVERITT, B. J. & PHILLIPS, P. E. 2012. Hierarchical recruitment of phasic dopamine signaling in the striatum during the progression of cocaine use. Proc Natl Acad Sci U S A, 109, 20703-8.
YAEL, D., ZEEF, D. H., SAND, D., MORAN, A., KATZ, D. B., COHEN, D., TEMEL, Y. & BAR-GAD, I. 2013. Haloperidol-induced changes in neuronal activity in the striatum of the freely moving rat. Front Syst Neurosci, 7, 110.
YIN, H. H., KNOWLTON, B. J. & BALLEINE, B. W. 2006. Inactivation of dorsolateral striatum enhances sensitivity to changes in the action-outcome contingency in instrumental conditioning. Behav Brain Res, 166, 189-96.
YIN, H. H., OSTLUND, S. B. & BALLEINE, B. W. 2008. Reward-guided learning beyond dopamine in the nucleus accumbens: the integrative functions of cortico-basal ganglia networks. Eur J Neurosci, 28, 1437-48.
YIN, H. H., OSTLUND, S. B., KNOWLTON, B. J. & BALLEINE, B. W. 2005. The role of the dorsomedial striatum in instrumental conditioning. Eur J Neurosci, 22, 513-23.
YU, C., FAN, D., LOPEZ, A. & YIN, H. H. 2012. Dynamic changes in single unit activity and gamma oscillations in a thalamocortical circuit during rapid instrumental learning. PLoS One, 7, e50578.
YUAN, Y., MAO, H. & SI, J. 2015. Cortical neural responses to previous trial outcome during learning of a directional choice task. J Neurophysiol, 113, 1963-76.





























