Embed Size (px)
Citation preview

Neuronal precursor-specific activity of a human doublecortinregulatory sequence
Claudia Karl,*,�,1 Sebastien Couillard-Despres,*,�,1 Peter Prang,� Matthias Munding,*,�,�Werner Kilb,§ Tanja Brigadski,§ Sonja Plotz,*,� Wolfgang Mages,¶ Heiko Luhmann,§Jurgen Winkler,� Ulrich Bogdahn� and Ludwig Aigner*,�
*Volkswagen-Foundation-Research Group, University of Regensburg, Regensburg, Germany
�Department of Neurology, University of Regensburg, Regensburg, Germany
�Institute of Anatomy, University of Regensburg, Regensburg, Germany
§Institute of Physiology and Pathophysiology, Johannes Gutenberg-University, Mainz, Germany
¶Institute of Genetics, University of Regensburg, Regensburg, Germany
Abstract
The doublecortin (DCX) gene encodes a 40-kDa microtubule-
associated protein specifically expressed in neuronal precur-
sors of the developing and adult CNS. Due to its specific
expression pattern, attention was drawn to DCX as a marker
for neuronal precursors and neurogenesis, thereby under-
scoring the importance of its promoter identification and pro-
moter analysis. Here, we analysed the human DCX regulatory
sequence and confined it to a 3.5-kb fragment upstream of the
ATG start codon. We demonstrate by transient transfection
experiments that this fragment is sufficient and specific to
drive expression of reporter genes in embryonic and adult
neuronal precursors. The activity of this regulatory fragment
overlapped with the expression of endogenous DCX and with
the young neuronal markers class III b-tubulin isotype and
microtubule-associated protein Map2ab but not with glial or
oligodendroglial markers. Electrophysiological data further
confirmed the immature neuronal nature of these cells.
Deletions within the 3.5-kb region demonstrated the relevance
of specific regions containing transcription factor-binding sites.
Moreover, application of neurogenesis-related growth factors
in the neuronal precursor cultures suggested the lack of direct
signalling of these factors on the DCX promoter construct.
Keywords: gene regulation, neurogenesis, neuronal migra-
tion, neuronal progenitors, transcription factors.
J. Neurochem. (2005) 92, 264–282.
Neurogenesis in the developing and adult CNS is suppor-ted by a population of neural stem cells that possesses thepotential to proliferate, self-renew and differentiate intoneurones and glia (Alvarez-Buylla and Temple 1998; Gage2000; Alvarez-Buylla and Garcia-Verdugo 2002). Under-standing of molecular mechanisms underlying changes incellular identity during the differentiation of neural stemcells into mature neurones is still superficial. Emergingevidence on the origins of neural stem cells points toradial glia cells and specialized subtypes of glial fibrillaryacidic protein (GFAP)-expressing astrocytes in the embry-onic and adult brain, respectively (Doetsch et al. 1999;Alvarez-Buylla et al. 2001; Seri et al. 2001; Gotz et al.2002; Sanai et al. 2004). In the adult CNS, neural stemcells slowly proliferate to self-renew their population butcan also give rise to fast-dividing transit-amplifyingprogenitors with a limited self-renewal capacity (Seri et al.
2001; Alvarez-Buylla and Garcia-Verdugo 2002; Doetschet al. 2002; Kempermann et al. 2004). Thereafter, molecu-lar and cellular events, such as lineage restriction,cell cycle exit, morphological and functional differentiation
Received April 6, 2004; revised manuscript received August 24, 2004;accepted September 20, 2004.Address correspondence and reprint requests to Ludwig Aigner, PhD,
Volkswagen-Foundation-Research Group, Department of Neurology,University of Regensburg, Franz-Josef-Strauss-Allee 11, D-93053Regensburg, Germany.E-mail: [email protected] authors contributed equally to this article.Abbreviations used: CMV, cytomegalie virus; DCX, doublecortin;
DsRed, Discosoma sp. red fluorescent protein; EGFP, enhanced greenfluorescent protein; FGF, fibroblast growth factor; GFAP, glial fibrillaryacidic protein; MEF, mouse embryonic forebrain; OB, olfactory bulb; rh,recombinant human; SVZ, subventricular zone; VEGF, vascularendothelial growth factor.
Journal of Neurochemistry, 2005, 92, 264–282 doi:10.1111/j.1471-4159.2004.02879.x
264 � 2005 International Society for Neurochemistry, J. Neurochem. (2005) 92, 264–282

and maturation, complete the neurogenic program(Alvarez-Buylla and Temple 1998; Gage 2000; Alvarez-Buylla and Garcia-Verdugo 2002). Due to the limitedavailability of differentiation stage-specific markers, themolecular and cellular mechanisms underlying the distinctneurogenic events are poorly understood.
Neurogenesis is not an immutable event but undergoescontinuous changes. Modulation of neurogenesis arises fromthe regulation of all components of the neurogenic program.A variety of factors and stimuli, such as enriched environ-ment, physical activity, growth factors (e.g. fibroblast growthfactor 2 (FGF-2), insulin-like growth factor 1 (IGF-1), vas-cular endothelial growth factor (VEGF) or erythropoietin) andlesions (e.g. transient global ischaemia, seizure or traumaticbrain injury), have been shown to modulate neurogenesis inthe adult CNS (Kempermann et al. 1997; Kuhn et al. 1997;Parent et al. 1997; Liu et al. 1998; van Praag et al. 1999;Aberg et al. 2000; Dash et al. 2001; Shingo et al. 2001; Jinet al. 2002). Again, due to the scarcity of differentiationstage-specific genes or markers, the cellular targets ofmodulating factors are often unclear. The shortage of markershinders the understanding of several aspects of neurogenesishence, there is a clear need for the identification of additionalcell type-specific markers and genes.
Due to its expression pattern, attention was recentlyfocused on doublecortin (DCX), a 40-kDa microtubule-associated protein. During human cortical development,DCX is required for proper neuronal precursor migration,probably through its interactions with the microtubulenetwork, dynein and/or the mitotic spindle apparatus (Franciset al. 1999; Gleeson et al. 1999a; Corbo et al. 2002; Baiet al. 2003; Couillard-Despres et al. 2004; Tanaka et al.2004b; for review see Couillard-Despres et al. 2001). DCXfunction and its microtubule affinity are regulated by post-translational modifications such as cdk5-dependent phos-phorylation (Schaar et al. 2004; Tanaka et al. 2004a).
During development, DCX is specifically and abundantlyexpressed in migrating neuronal precursors (des Portes et al.1998a; Gleeson et al. 1998; Francis et al. 1999). In the adultbrain, DCX expression is retained within areas of continuousneurogenesis, i.e. the hippocampus and the subventricularzone (SVZ)/rostral-migratory stream/olfactory bulb (OB)axis (Brown et al. 2003) and is rarely detected outside theseregions (Nacher et al. 2001). DCX is transiently expressedduring adult neurogenesis in proliferating neuronal progen-itors and in post-mitotic neuronal precursors (Brown et al.2003). Immunohistological studies revealed that cells expres-sing DCX clearly constitute a cell population distinct fromcells expressing nestin or GFAP, indicating that DCX is notexpressed in multipotent neural stem cells (our own unpub-lished observations). Confusion on this issue has been causedby the use of transgenic mice expressing enhanced greenfluorescent protein (EGFP) under the control of the nestinneural enhancer (Kempermann et al. 2004). In this animal
model, the reporter protein persists in the neuronal precursorsexplaining the reported colocalization of EGFP with DCX.
Expression of DCX is induced as neuroblasts are gener-ated, peaks during the second week after cell birth and isdown-regulated concomitantly with the appearance of mark-ers associated with mature neurones, such as NeuN (Cooper-Kuhn and Kuhn 2002; Brown et al. 2003). Two weeks afterbromo-deoxyuridine labelling, the majority (approximately90%) of newborn cells in the dentate gyrus and the SVZ/rostral-migratory stream/OB axis were found to express DCX(Brown et al. 2003). Similarly, after daily injections ofbromo-deoxyuridine for 12 days, approximately 90% of thecells expressing DCX were found to have incorporatedbromo-deoxyuridine during the labelling period, confirmingthat the DCX-positive cell population is composed of newlygenerated cells (Rao and Shetty 2004). Importantly, DCXexpression was specific to newly generated neuronal precur-sors as virtually all DCX-positive cells express earlyneuronal antigens but lack antigens specific to glia orundifferentiated cells and, conversely, glial cells do notexpress DCX (Brown et al. 2003; Rao and Shetty 2004;Steiner et al. 2004).
The correlation of an increase in DCX expression withlesion- or growth factor-induced up-regulation of neurogen-esis supports the use of DCX as a general neurogenesismarker. For example, induction of cortical neurogenesis afterphototoxic lesion provoked the appearance of DCX-expres-sing neuroblasts in the lesion area (Magavi et al. 2000).Similarly, after transient occlusion of the middle cerebralartery, newly formed neurones expressing DCX can beidentified in the ischaemic neostriatum (Arvidsson et al.2002; Jin et al. 2003a,b). Other examples of enhancedexpression of DCX were described subsequent to stimulationof neurogenesis through VEGF, FGF-2 and heparin-binding-epidermal growth factor (HB-EGF) (Jin et al. 2002,2003b,c). Finally, neural precursor cells were also found toexpress DCX after transplantation (Englund et al. 2002;Tang et al. 2002; Kelly et al. 2004). Taken together, thisevidence suggests that DCX is a very promising and reliablemarker of neurogenesis.
Expression of reporter genes under specific promoters is auseful tool to analyse developmental and neurogenic pro-cesses in the CNS. Examples are the neural stem cell-specificenhancer element of the nestin gene, pan-neuronal tubulin-alpha1 promoter, oligodendroglia precursor 2¢,3¢-cyclic nuc-leotide 3¢-phosphodiesterase promoter and, more recently,post-mitotic neuronal Tis21 locus, the neuronal precursor-specific regulatory element of the pro-opiomelanocortin geneand the neural stem cell-specific locus of the TLX gene(Gloster et al. 1994; Frisen et al. 1995; Gravel et al. 1998;Haubensak et al. 2004; Overstreet et al. 2004; Shi et al.2004). In addition, some of these promoters have beenemployed for the isolation of specific cell types (Roy et al.2000a,b; Wang et al. 2000; Shi et al. 2004). In view of the
DCX promoter identification 265
� 2005 International Society for Neurochemistry, J. Neurochem. (2005) 92, 264–282

relevance of DCX as a marker for neuronal precursors andneurogenesis, and considering the importance of promoteridentification and analysis for the understanding of DCXgene regulation, we analysed a putative regulatory regionupstream of the human DCX gene. We defined a 3.5-kbregion as an element that is sufficient to provide strong andspecific expression in neuronal precursors and young neur-onal cells.
Materials and methods
Cloning, sequence analysis and expression plasmids
Human sequenceUsing the partial human X chromosome sequence available under
GenBank Accession no. NT025319, two oligonucleotides (5¢primer, AACACCTATTAATGCCCA, location )3523 to )3506relative to the ATG start codon; 3¢ primer, TCAGACACCT
GAGCCTGGGAGAA, location )15 to )37) were designed for
the PCR amplification of a 3509-bp genomic sequence upstream of
the DCX ATG start site. PCR on human genomic DNA derived from
human foetal cortical neuronal precursor cells (Wachs et al. 2003)using an Expand High Fidelity PCR System kit (Roche, Grenzach-
Wyhlen, Germany) amplified a 3.5-kb fragment. This fragment was
blunt ended with the DNA polymerase I Klenow fragment and
phosphorylated using T4 polynucleotide kinase in the presence of
ATP. If not otherwise notified, all enzymes were obtained from New
England Biolabs GmbH (Frankfurt am Main, Germany). The
cytomegalie virus (CMV) promoter from the plasmid pEGFP-N1
(BD/Clontech, Heidelberg, Germany) was deleted using AseI andNheI. The human DCX 3509-bp putative regulatory sequence was
inserted into the SmaI-linearized promoterless pEGFP-N1 vector. In
the resulting construct of approximately 7.7 kb, named phu-
DCX3509-EGFP, the 3509-bp putative human DCX regulatory
fragment controls the expression of the EGFP gene. The integrity of
the 3509-bp fragment was verified by direct sequencing using a
DNA Sequencer 373 (Applied Biosystems, Darmstadt, Germany).
To direct expression of the red fluorescent protein Discosoma sp. redfluorescent protein (DsRed)2 or the luciferase reporter gene from
firefly, the 3509-bp putative human DCX regulatory region was
subcloned into a vector (pDsRed2-1) containing the gene for
DsRed2 (Clontech) and a vector (pGl3-Basic) containing the gene
for firefly luciferase (Promega, Madison, WI, USA). The resulting
vectors were named phuDCX3509-DsRed2 and phuDCX3509-
FFLuci, respectively. Deletion mutants of the 3509-bp fragment
were generated in phuDCX3509-FFLuci by enzymatic digest with
appropriate restriction enzymes or by direct cloning of PCR
products in pGl3-Basic.
Mouse sequenceA PCR-based screening was performed on the library RPCI-23
bacterial artificial chromosome (BAC) II (Resource Center/Primary
Database, RZPD German Resource Center for Genome Research,
Berlin, Germany; library originally from Dr Pieter de Jong,
Kazutoyo Osoegawa, Minako Tateno, Roswell Park Cancer
Institute, Buffalo, NY, USA) using two oligonucleotides (5¢primer, TTGGACATTTTGACGAACGA, location +13 to +32
relative to the ATG start codon; 3¢ primer, CCCTTCTTCCAGTT-
CATCCA, location +365 to +346). Two positive clones
(RPCIB731G16462Q2 and RPCIB731H1192Q2) containing
approximately 25 kb of mouse genomic DNA cloned in the vector
pBACe3.6 were further analysed by direct sequencing using a
DNA Sequencer 373 (Applied Biosystems). The sequence derived
was confirmed by sequencing of mouse genomic DNA derived
from tails of C57Bl/6Ncrl mice and is available under NCBI
nucleotide bank Accession no. AY590498.
Sequence analysis was performed with the following programs:
ALIGN, version 3.2 available at Biology Workbench of the San
Diego Supercomputer Center (http://workbench.sdsc.edu) for align-
ment and MatInspector, version 6.2.1 available at Genomatix
Software GmbH (Munich, Germany; http://www.genomatix.de) for
analyis of transcription factor-binding sites (Quandt et al. 1995).
Cell cultures
If not otherwise cited, solutions and media for cell culture were
obtained from PAN Biotech GmbH (Aidenbach, Germany) and
plastic culture vessels from TPP (Trasadingen, Switzerland).
Ampho293 cells (Clontech), COS7 cells (CRL-1651; ATCC USA,
Manassas, VA, USA), CTX TNA2 (rat astrocyte line, ECACC no.
98102213), D283Med (HTB-185; ATCC USA) and Neuro2A
(CCL-131; ATCC USA) were cultured according to the supplier’s
instructions. N20.1, a mouse oligodendrocyte line (Verity et al.1993), was kindly provided by Anthony T. Campagnoni (University
of California, Los Angeles, CA, USA). For proliferation, N20.1
cells were maintained in HAM’s-F12/Dulbecco’s modified Eagle’s
medium (1 : 1) supplemented with 2.4 g/L glucose, 0.18 g/L L-
glutamine, 100 U/mL penicillin, 0.1 mg/mL streptomycin and
22.5 mg/L gentamicin at 33�C in a 5% CO2-containing humidified
atmosphere. In order to differentiate this line into an oligodendrogl-
ial phenotype, cells were maintained in the same medium but at
39�C for 4 days. All cells were seeded on plastic culture plates at a
density of 2.5 · 104 cells/cm2 1 day before proceeding to the
transfection. Before seeding D283Med cells, culture dishes were
coated with poly-ornithin (250 lg/mL) and laminin (15 lg/mL). At
1 day after transfection, cells were reseeded on glass coverslips
coated with poly-ornithin (250 lg/mL) and laminin (15 lg/mL) at a
density of 7.5 · 104 cells/cm2.
Primary mouse embryonic forebrain (MEF) cultures were pre-
pared as follows. The animal experiments were carried out in
accordance with the European Communities Council Directive
(86/609/EEC) and institutional guidelines. Pregnant C57Bl/6Ncrl
female mice (Charles River Laboratories, Sulzfeld, Germany) were
killed and the uteri promptly removed and immersed in ice-cold
Dulbecco’s phosphate-buffered saline solution. Embryonic day 10.5–
12.5 embryos were released from the uteri and forebrains dissected
and separated from surrounding tissues. Cell dissociation was
performed as described previously (Wachs et al. 2003). Dissociatedcells were collected by 5 min centrifugation at 120 g. The pellet wasresuspended in Neurobasal medium (Invitrogen-Gibco, Karlsruhe,
Germany) and washed three times. Finally, the cell preparation was
resuspended in MEF differentiation medium [Neurobasal medium
supplemented with B27 (Invitrogen-Gibco), 2 mM L-glutamine,
100 U/mL penicillin, 0.1 mg/mL streptomycin, 5% foetal calf
serum], seeded on coverslips coated with poly-ornithin (250 lg/mL)
and laminin (15 lg/mL) at a density of 2.5 · 105 cells/cm2 and
266 C. Karl et al.
� 2005 International Society for Neurochemistry, J. Neurochem. (2005) 92, 264–282

maintained for 4 days at 37�C in a 5% CO2-containing humidified
atmosphere before transfection. In some experiments, additional
factors were added 12 h after transfection at the following concen-
trations: 1 mM all trans-retinoic acid (Sigma-Aldrich, Taufkirchen,
Germany), 20 ng/mL recombinant human brain-derived neuro-
trophic factor (rhBDNF) (Alomone Laboratories, Jerusalem, Israel),
100 ng/mL recombinant mouse vascular endothelial growth factor
(rmVEGF) (R & D Systems, Wiesbaden, Germany), 20 ng/mL
rhEGF (R & D Systems), 20 ng/mL rhFGF basic (R & D Systems)
with 2 lg/mL heparin (Sigma-Aldrich), 20 ng/mL recombinant
human neurotrophin-3 (rhNT-3) (R & D Systems), 20 ng/mL
rhNT-4 (R & D Systems) or 10 ng/mL recombinant human
transforming growth factor beta (rhTGFbeta) (R & D Systems).
Preparation of organotypic slice cultures from post-natal mouse
hippocampus
Slice cultures were prepared from C57Bl/6Ncrl mouse brain at post-
natal days 3–7 according to previously published protocols
(Stoppini et al. 1991; Prang et al. 2001). Briefly, the hippocampus
was dissected and transverse slices of 200 or 400 lm were cut using
a tissue chopper (McIllwain Stoelting, IL, USA) and transferred
onto membranes (Millicell-CM; Millipore, Eschborn, Germany).
Slices were incubated for up to 15 days in a humidified atmosphere
with 5% CO2 at 37�C in 50% minimal essential medium, 25% basal
medium eagle, 25% heat-inactivated horse serum, 0.65% glucose
and glutamax (2 nM) adjusted to pH 7.3. Before ballistic transfec-
tion, slices were incubated for 4 days under cell culture conditions.
Preparation of explant cultures from adult mouse olfactory bulb
Olfactory bulb was prepared from 5-week-old C57Bl/6Ncrl mice,
briefly rinsed in phosphate-buffered saline and cut into pieces using
a razorblade. Tissue was placed on top of 100% Matrigel (Becton
Dickinson, Heidelberg, Germany) to allow migration of cells out of
the explant in MEF differentiation medium and incubated at 37�C in
a 5% CO2-containing humidified atmosphere for up to 7 days before
ballistic transfection.
Cell and tissue transfections
Transfections of cell lines were performed in 10-cm2 culture dishes
(six-well plates) using 3 lg of plasmid per dish in the presence of
5 lL Metafectene (Biontex Laboratories, Munich, Germany)
according to the manufacturer’s protocol. Medium was exchanged
after 12 h. One day after the end of transfection, cells were
trypsinized and reseeded on poly-ornithin/laminin-coated glass
coverslips for another 24 h before immunohistological analysis.
Primary MEF cells were transfected directly on coated 4-cm2 glass
coverslips which they had been seeded on to form a differentiated
culture using 1.2 lg of plasmid and 3 lL Metafectene per well. For
luciferase-reporter transfections, MEF cells had been seeded on
poly-ornithin/laminin-coated 96-well luciferase plates (Greiner bio-
one, Frickenhausen, Germany). Transfections of MEF cells were
performed using 0.1 lg of plasmid per 96-well in the presence of
0.15 lL Metafectene according to the manufacturer’s protocol.
Medium was exchanged after 12 h and cells were incubated for
another 2–4 days under cell culture conditions before immuno-
cytological, electrophysiological or luminescence analysis.
For ballistic transfection of mouse hippocampal slice cultures and
OB explant cultures, helium was used to project spherical gold
particles (1.5–3.0 lm in diameter; Alfa Aesar, Karlsruhe, Germany)
coated with plasmid DNA by ethanol precipitation. Solution (10 lL)containing 1 lg of plasmid DNA coated on 1 lg gold in ethanol
was placed on the screen of a Swinney filter holder (Millipore) that
was connected by tubing to a helium reservoir. Brisk valve opening
for about 0.1 s permitted a rapid flow of helium (exit pressure
400 kPa) which propelled the particles towards the tissue. The
distance between device and targets was 6 cm. Transfected slices
and explant cultures were incubated under cell culture conditions for
another 2–4 days before immunohistochemical analysis.
Western blot analysis
To prepare protein extracts, cells were seeded on 10-cm2 dishes.
Cells were harvested from the culture vessel in phosphate-buffered
saline using a cell scraper and collected by 5 min centrifugation at
200 g. Proteins were extracted, resolved by sodium dodecyl
sulphate–polyacrylamide gel electrophoresis, blotted onto nitrocel-
lulose and incubated with antibodies as described previously
(Brown et al. 2003). To detect DCX or actin, the blocked membrane
was probed with anti-DCX (goat IgG, 1 : 1000 dilution; Santa Cruz
Biotechnology, Santa Cruz, CA, USA) or anti-actin (rabbit IgG,
1 : 5000 dilution; Sigma, Taufkirchen, Germany) antibody over-
night at 4�C. Blots were rinsed and incubated with a horseradish
peroxidase-conjugated anti-goat IgG (1 : 50 000 dilution; Sigma) or
anti-rabbit IgG secondary antibody (1 : 50 000 dilution; Dianova,
Hamburg, Germany) for 2 h at room temperature (20–24�C) and
rinsed before performing ECL detection of protein–antibody
conjugates (ECL-Plus Western Blotting Detection System; Amer-
sham-Pharmacia, Freiburg, Germany) according to the manufac-
turer’s protocol.
Luciferase assays
Luciferase assays were performed using the Dual Glo Luciferase
Assay System (Promega). In this system, a cotransfection with
phuDCX3509-FFLuci and a control vector driving Renilla-lucif-erase under the CMV promoter is performed. The two different
luciferases use different specific substrates and therefore the DCXregulatory region-driven firefly luciferase activity was distinguished
from Renilla-luciferase. Firefly luciferase activity was normalized
against the CMV-driven Renilla-luciferase.
Immunostainings
After 30 min of fixation at room temperature with phosphate-
buffered 4% paraformaldehyde, samples were blocked for a
minimum of 1 h in fish skin gelatin buffer [0.1 M Tris-HCl,
pH 7.5, 0.15 M NaCl, 1% bovine serum albumin, 0.2% Teleostean
gelatin (Sigma-Aldrich), 0.1% Triton X-100] at room temperature.
For staining of slice cultures fish skin gelatin buffer contained 0.5%
of TritonX-100 and 400-lm slices were stained free floating in
solution. The specimens were incubated overnight at 4�C with the
primary antibodies at the following dilutions: goat anti-DCX C18
(1 : 1000; Santa Cruz Biotechnology); rabbit anti-galactocerebro-
side (1 : 250; AB142, Chemicon, Temecula, CA, USA); rabbit anti-
GFAP (1 : 1000; Dako, Glostrup, Denmark); rabbit anti-KI-67
(1 : 500; Novocastra, Newcastle upon Tyne, UK); mouse anti-nestin
(1 : 500; BD Biosciences Pharmingen, San Diego, CA, USA);
mouse anti-bIII-tubulin (1 : 500; clone 5G8, Promega); rabbit anti-
neurofilament 200 (1 : 500; Chemicon); mouse anti-Map2ab
DCX promoter identification 267
� 2005 International Society for Neurochemistry, J. Neurochem. (2005) 92, 264–282

(1 : 250; Sigma-Aldrich); rabbit anti-calbindin (1: 2000; Swant,
Bellinoza, Switzerland); mouse anti-calretinin (1 : 1000; Swant);
rabbit anti-cholinacetyltransferase (1: 2000; Chemicon); rabbit anti-
GABA (1 : 500; Chemicon) and rabbit anti-serotonin (1 : 500;
Diasorin, Stillwater, MN, USA). The secondary fluorochrome-
conjugated antibodies were diluted 1 : 500 (Rhodamine Red�-X-
conjugated-, Cy�5-conjugated-, FITC-conjugated donkey anti-
mouse, rabbit and goat anti-IgG from Dianova). All antibody
dilutions and washes were performed with the fish skin gelatin
buffer. Nuclear counterstaining was performed with 4¢,6¢-diamidino-
2-phenylindole dihydrochloride hydrate (0.25 lg/lL; Sigma-Ald-
rich) or TO-PRO-3 (Molecular Probes, Eugene, OR, USA). After
the last wash, the samples were briefly rinsed with phosphate-
buffered saline and mounted on slides using Fluoromount (Southern
Biotechnology Associates, Birmingham, AL, USA). In cases where
antigens were sensitive to detergents, Triton X-100 was omitted
from the fish skin gelatin buffer.
Electrophysiological methods
Coverslips with primary MEF cells were transferred 3–4 days after
transfection into a submerged recording chamber (volume 3 mL)
and superfused at a rate of 1–2 mL/min with extracellular solution
containing (in mM): NaCl, 110; KCl, 5; CaCl2, 2; MgCl2, 1; glycine,
0.01; glucose, 10 and HEPES, 15. All experiments were performed
at room temperature (20–24�C). The videomicroscopic setup
consisted of an upright microscope with DIC optics (Axioskop;
Zeiss, Jena, Germany), an infrared filter (KMZ 50-2, I1/2 ¼ 750 nm,
width 57 nm; Schott, Mainz, Germany) and a CCD camera system
(C5405; Hamamatsu, Toyooka, Japan). MEF cells with active
human DCX regulatory region were identified by EGFP expression
visible by fluorescence microscopy. Whole-cell patch-clamp record-
ings were performed according to published procedures (Stuart
et al. 1993). Patch-pipettes (5–12 MW) were pulled from borosil-
icate tubing (2.0 mm outside, 1.16 mm inside diameter; Science
Products, Hofheim, Germany) using a vertical puller (PP-83;
Narishige, Tokyo, Japan), filled with pipette solution containing
(in mM): K-gluconate, 90; NaCl, 5; CaCl2, 0.06; EGTA, 0.6;
HEPES, 15; Na2-ATP, 2 and biocytin, 0.5% (Sigma-Aldrich) and
connected to the headstage of a discontinuous voltage-clamp/
current-clamp amplifier (SEC05L; NPI, Tamm, Germany). Signals
were amplified, low-pass filtered at 3 kHz, digitized online by an
AD/DA board (ITC-16; Heka, Lamprecht, Germany), recorded and
processed with the software WINTIDA 4.11 (Heka) and stored on a
personal computer. All potentials were corrected for a liquid-
junction potential of 10 mV. Resting membrane potential was
measured directly after obtaining whole-cell configuration. All
subsequent experiments were performed from a holding potential of
)60 mV. For the determination of Rinput hyperpolarizing current
pulses ()20 pA) were injected. Action potential amplitude was
measured from the threshold and the action potential duration was
determined at half-maximal amplitude. Tetrodotoxin was applied by
bath application for at least 4 min before recording. GABA was
applied focally via patch pipettes using a pressure application
system (PDES; NPI).
Quantitative analysis
Immunostainings of cultured cells were examined using confocal
laser microscopy. In order to quantify the ratio of cells expressing a
certain marker or reporter protein, five randomly picked visual fields
were examined under a magnification of 200 ·. First, the total cell
number in the field was determined according to a nuclear
counterstain and the number of cells positive for the marker or the
transgene was then determined. All quantification was performed in
three consecutive experiments. Statistical analysis was performed
using the Student’s t-test. In order to detect colabelling of
endogenous markers or colabelling of marker and reporter, cells
expressing the marker of interest were picked randomly and
examined for coexpression. For this purpose, a magnification of
400 · was used and a total number of 120 cells positive for the
marker of interest was examined in each paradigm and in three
consecutive experiments.
Results
Characterization and cloning of the human and mouse
DCX regulatory region
The 9.5-kb transcription product of the human DCX geneoriginates from the alternative splicing of nine exons (desPortes et al. 1998a; Gleeson et al. 1998) (Fig. 1). The firstthree exons can form the 5¢ end of the mRNA. The fourthexon possesses the initial ATG of a 1080-bp long open
Fig. 1 Gene structure and putative regulatory regions of the human,
mouse and rat doublecortin (DCX) gene. The genomic structures and
the putative regulatory sequences upstream of the ATG start codon of
(a) human, (b) mouse and (c) rat DCX gene are shown. Black boxes
indicate exons. Putative binding sites for transcription factors and
CAAT/TATA boxes are shown. Exons upstream of the ATG in the rat
DCX gene are not indicated as complete cDNA sequences are not
available. (d) Regions of homology (see also Table 1).
268 C. Karl et al.
� 2005 International Society for Neurochemistry, J. Neurochem. (2005) 92, 264–282

reading frame, i.e. encoding a polypeptide of 360 aminoacids. Another potential ATG is located in exon 3. The use ofthis codon as translation start point, although not yetreported, would produce a polypeptide bearing an additional42 amino acids (des Portes et al. 1998a). One particularity ofthe DCX transcript is its very long 3¢ untranslated region thatmight influence its stability (des Portes et al. 1998a). Tocharacterize the putative promoter elements, we analysed thegenomic DNA sequence 5¢ to the ATG start codon of thehuman, mouse and rat DCX genes. In total, 30 kb of genomicDNA upstream of the ATG of the human, mouse and ratDCX gene were aligned using sequences available underNCBI gene bank nos NT_025319, NT_039718 andNW_048037. Alignment between sequences of the threespecies revealed sequence identity above 85% only in thefragment between the start codon and the 3.5 kb upstream ofthe latter. Due to this high homology, further experimentsfocused on this 3.5-kb region.
Using genomic DNA derived from human foetal corticalneuronal precursor cells, we amplified by PCR a 3509-bpgenomic sequence located upstream of the ATG start site.This fragment was cloned into a promoterless plasmidencoding the reporter protein EGFP. The resulting vector isreferred to as phuDCX3509-EGFP.
The analysis of the 5¢ region adjacent to the ATG startsites revealed a putative TATA located at )1983 to )1976and a CAAT box at )2079 to )2071 relative to the ATGstart codon of exon 4 in the human DCX gene (Figs 1a and2). In the mouse DCX gene the TATA box is located at)2012 to )2005 and the CAAT box at )2111 to )2103relative to the ATG start codon (Fig. 1b). The rat genomicsequence adjacent to the translation start site contains aTATA box at )2024 to )2018 and a CAAT box at )2125 to)2117 relative to the ATG start codon (Fig. 1c). Anadditional ATG start codon ()359 to )357) has beendescribed in the human exon 3; however, no polypeptidesoriginating from this translational start site have beendescribed (des Portes et al. 1998a). Although the mouseDCX gene sequences include a region homologous to thehuman exon 3, we could not detect transcription of thisputative exon. Using various oligonucleotides designed toamplify by RT-PCR a region spanning the putative mouseexon 3 and exon 4, we could not amplify any transcriptionalproduct (data not shown). In addition, using combinationsof oligonucleotides designed to amplify putative exon 1 toexon 4 or exon 2 to exon 4, the RT-PCR products derivedfrom mouse brains at different developmental stages(embryonic day 12 to adult) did not include sequences
Fig. 2 Putative regulatory sequence of the human doublecortin (DCX) gene. The 3509-bp sequence upstream of the ATG start codon of the
human DCX gene is shown. Putative transcription factor binding sites, CAAT and TATA boxes are underlined, exon sequences are in italics.
DCX promoter identification 269
� 2005 International Society for Neurochemistry, J. Neurochem. (2005) 92, 264–282

from putative exon 3 (data not shown). Therefore, weexcluded the sequences homologous to the human DCXexon 3 in the numbering of the mouse DCX exons. Theproposed regulatory region of the rat DCX gene containsregions highly homologous to sequences coding for exons1–3 on the human and exons 1–2 on the mouse sequence,respectively, although no sequences of rat DCX mRNAcontaining further exons besides protein-encoding exons 3–8 have yet been described.
The result of a homologous/identical regions analysisbetween human, mouse and rat sequences defines severalareas. Of interest are the regions downstream of region 1.These regions, between region 2 and the ATG translationstart site, present high levels of homology across the threespecies, i.e. more than 75% identity, suggesting that
relevant regulatory sequences might be located withinthese regions (Fig. 1d and Table 1). Indeed, a search fortranscription factor consensus sequences 5¢ to the ATG startcodon revealed putative binding sites for a number oftranscription factors, such as Brn-2, Brn-3, NeuroD1,E2F-1, E2F-2, Fast1, Smad3 and Smad4 (Figs 1 and 2,Table 2).
Specific activity of the DCX regulatory region in cells of
neuronal lineage
Activity of the subcloned 3509-bp human DCX regulatoryregion (phuDCX3509-EGFP) was tested by transient trans-fection into different cell types. Established cell lines wereused, such as non-neural cell lines (HEK293 and COS7),glial lineage restricted lines (astrocytic cell line CTX TNA2
Table 1 Homology of putative regulatory regions on human, mouse and rat doublecortin gene
Region
Positions on human
gene relative to
ATG (bp)
Homologous regions
on mouse gene relative
to ATG (bp)
% identity mouse
vs. human
Homologous regions
on rat gene relative
to ATG (bp)
% identity rat
vs. human
Exon 1 )1776 to )1739 )1802 to )1765 100.0 )1814 to )1777 97.4
Exon 2 )1679 to )1571 )1706 to )1592 79.6 )1716 to )1601 80.3
Exon 3 )747 to )355 )744 to )358 87.4 )744 to )353 86.9
Region 1 )2885 to )2358 )2881 to )2354 47.2 )2896 to )2408 47.6
Region 2 )2357 to )1777 )2353 to )1803 79.6 )2407 to )1815 78.4
Region 3 )1738 to )1680 )1764 to )1707 83.1 )1776 to )1717 91.7
Region 4 )1570 to )748 )1591 to )745 75.3 )1600 to )745 74.4
Region 5 )354 to )22 )357 to )23 79.1 )352 to )23 78.3
Table 2 Putative transcription factor bind-
ing sites and their locationTranscription
factor binding
sites
Positions on human
gene relative
to ATG (bp)
Positions on mouse
gene relative
to ATG (bp)
Positions on rat
gene relative
to ATG (bp)
Brn-2 )1538 to )1522 )1559 to )1543 )1568 to )1552)749 to )733 )746 to )730
Brn-3 )1930 to )1914 )1956 to )1940 )2364 to )2348)1968 to )1952
NeuroD1 )1702 to )1690 )1728 to )1716 )1740 to )1728)414 to )402 )1271 to )1259
)1093 to )1081)413 to )401)173 to )161
E2F-1 )1692 to )1678 )2321 to )2307 )1758 to )1744)848 to )834 )1746 to )1732 )600 to )586)762 to )748 )1266 to )1252
)1082 to )1068E2F-2 )1214 to )1200 )304 to )290Smad3 )2126 to )2118Smad4 )2126 to )2118Fast1 )2308 to )2294 )2259 to )2245
)2236 to )2222 )1993–1979
270 C. Karl et al.
� 2005 International Society for Neurochemistry, J. Neurochem. (2005) 92, 264–282

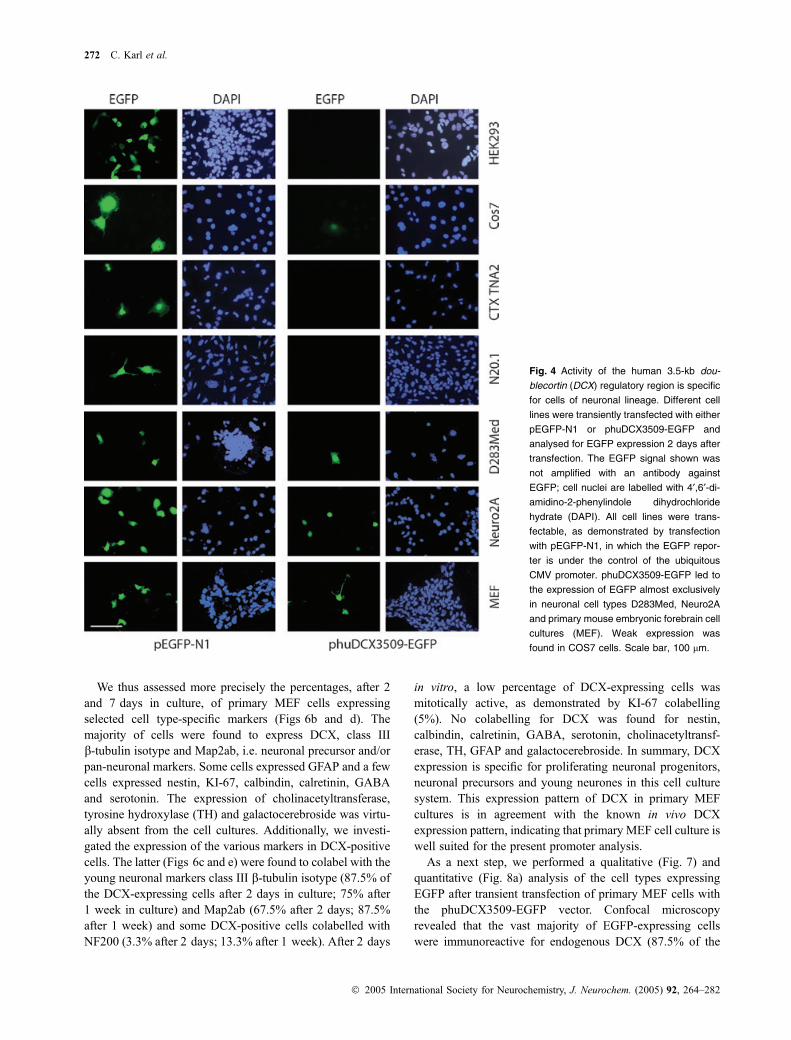
and oligodendroglial line N20.1) and mixed populationneural lines (human medulloblastoma D283Med cells andmouse neuroblastoma cell line Neuro-2a). In addition,primary cell cultures obtained from MEF of embryonic day10.5–12.5 were used. Assessment of DCX expression in thedifferent cell types by western blot analysis revealed noexpression of DCX in the non-neuronal cell lines HEK293and COS7 and no expression in the astrocytic cell line CTXTNA2 or in the oligodendroglial line N20.1 (Fig. 3). Humanmedulloblastoma D283Med cells expressed DCX at lowlevels whereas, in the mouse neuroblastoma cell line Neuro-2a, DCX expression was below the level of detection(Fig. 3). In contrast, strong expression of DCX was found inprimary MEF cell cultures.
In a next step, we compared EGFP reporter expression inthese different cell types after transfection with phu-DCX3509-EGFP or with the control vector pEGFP-N1, inwhich reporter expression is driven by the ubiquitous CMVpromoter. Control transfections with pEGFP-N1 demonstra-ted that all cell types were transfectable and that individualtransfected cells were expressing EGFP at high levels(Fig. 4). After transfection with phuDCX3509-EGFP,expression of EGFP expression was not or barely detectedin the non-neural cells HEK293 and COS7, as well as in theglial CTX TNA2 and the oligodendroglial N20.1 cells.Expression was weakly detectable in the neural cell linesD283Med and Neuro-2a but strong expression was seen inprimary MEF cells (Fig. 4). As compared with the neuronalNeuro-2a and primary MEF cells, the few COS7 cellsexpressing EGFP after transfection of phuDCX3509-EGFP
contained low levels of the reporter (Fig. 4). A comparisonof the western blot data from Fig. 3 concerning EGFPexpression under the control of the DCX regulatory fragment(Fig. 4) revealed a strong correlation between the transfectedpromoter activity and endogenous DCX expression. It isnoteworthy that transfections with phuDCX3509-EGFP orpEGFP-N1 did not influence the level of expression ofendogenous DCX (Fig. 3).
To quantify the transfection and expression efficacy, weassessed the percentage of EGFP-expressing cells aftertransfection with the different vectors. Although all celltypes used above were shown to be transfectable usingpEGFP-N1, the transfection rates varied. Hence, in HEK293,COS7 and Neuro-2a cell cultures, 6–14% of cells expressedEGFP, whereas CTX TNA2, D283Med and primary MEFcells had transfection rates below 3% after transfection withpEGFP-N1 (Fig. 5a). In comparison, the percentage ofEGFP-expressing cells after phuDCX3509-EGFP transfec-tion was much lower and more specific to neuronal cellcultures. We calculated the relative expression efficacy ofphuDCX3509-EGFP using the efficiency of expression ofpEGFP-N1 as a standard (100 · percentage of cells expres-sing phuDCX3509-EGFP/percentage of cells expressingpEGFP-N1). Using this calculation method, we deducedthat up to 30.6% of phuDCX3509-EGFP-transfected primaryMEF cells activated the 3509-bp DCX regulatory fragment(Fig. 5b). Lower expression efficacy was found in Neuro-2aand D283Med cells (approx. 7–12%) and no activity wasfound in the non-neuronal cell lines A293, CTX TNA2 andN20.1. COS7 cells showed little activity (6.3%), indicatingthat the 3509-bp DCX fragment might be ‘leaky’ undercertain cellular environments. Similar results were obtainedusing the neuronal-specific Tau promoter, which alsodisplayed low activity in COS7 cells (Heicklen-Klein andGinzburg 2000). In summary, these experiments suggest thatthe 3509-bp DCX regulatory region is specifically active incells of neuronal lineage.
DCX regulatory region is predominantly active in young
neurones of embryonic brain cultures
Transient transfection experiments were further used toassess the promoter activity of the 3509-bp DCX regulatoryfragment with respect to the endogenous DCX promoter inprimary neuronal precursors. To this end, we made use of themouse embryonic day 10.5–12.5 dissociated forebrain cells,a primary cell culture system physiologically relevant for thisstudy. This culture system comprises the different cell typesof the developing nervous system. Figure 6(a) shows ananalysis of the various cell types arising from the primaryMEF cells after 1 week of culture. Various cell types couldbe revealed using, for example, antibodies directed againstnestin, KI-67, DCX, class III b-tubulin isotype, Map2ab,neurofilament heavy subunit (NF-200), GFAP and galacto-cerebroside, although the latter were less numerous (Fig. 6a).
Fig. 3 Western blot analysis of different cell lines for expression of
doublecortin (DCX). Protein extracts (5 lg of total protein per lane)
from different cell lines probed for DCX and actin as control. HEK293
and COS7 cells, non-neural cell lines; CTX TNA2 cells, astrocytic cell
line; N20.1, oligodendroglial cell line; D283Med, human medullobla-
stoma cell line; Neuro2A, mouse neuroblastoma cell line, primary
mouse embryonic forebrain cells of embryonic day 10.5–12.5 (MEF).
No expression of DCX was found in HEK293, COS7, CTX TNA2 and
N20.1. D283Med cells expressed DCX at low levels but, in Neuro2A,
DCX expression was below level of detection. Note the strong
expression of DCX in primary MEF cell cultures. Transfection with
phuDCX3509-EGFP or pEGFP-N1 did not influence the level of
endogenous DCX expression in primary MEF cell cultures. CMV;
EGFP.
DCX promoter identification 271
� 2005 International Society for Neurochemistry, J. Neurochem. (2005) 92, 264–282
We thus assessed more precisely the percentages, after 2and 7 days in culture, of primary MEF cells expressingselected cell type-specific markers (Figs 6b and d). Themajority of cells were found to express DCX, class IIIb-tubulin isotype and Map2ab, i.e. neuronal precursor and/orpan-neuronal markers. Some cells expressed GFAP and a fewcells expressed nestin, KI-67, calbindin, calretinin, GABAand serotonin. The expression of cholinacetyltransferase,tyrosine hydroxylase (TH) and galactocerebroside was virtu-ally absent from the cell cultures. Additionally, we investi-gated the expression of the various markers in DCX-positivecells. The latter (Figs 6c and e) were found to colabel with theyoung neuronal markers class III b-tubulin isotype (87.5% ofthe DCX-expressing cells after 2 days in culture; 75% after1 week in culture) and Map2ab (67.5% after 2 days; 87.5%after 1 week) and some DCX-positive cells colabelled withNF200 (3.3% after 2 days; 13.3% after 1 week). After 2 days
in vitro, a low percentage of DCX-expressing cells wasmitotically active, as demonstrated by KI-67 colabelling(5%). No colabelling for DCX was found for nestin,calbindin, calretinin, GABA, serotonin, cholinacetyltransf-erase, TH, GFAP and galactocerebroside. In summary, DCXexpression is specific for proliferating neuronal progenitors,neuronal precursors and young neurones in this cell culturesystem. This expression pattern of DCX in primary MEFcultures is in agreement with the known in vivo DCXexpression pattern, indicating that primary MEF cell culture iswell suited for the present promoter analysis.
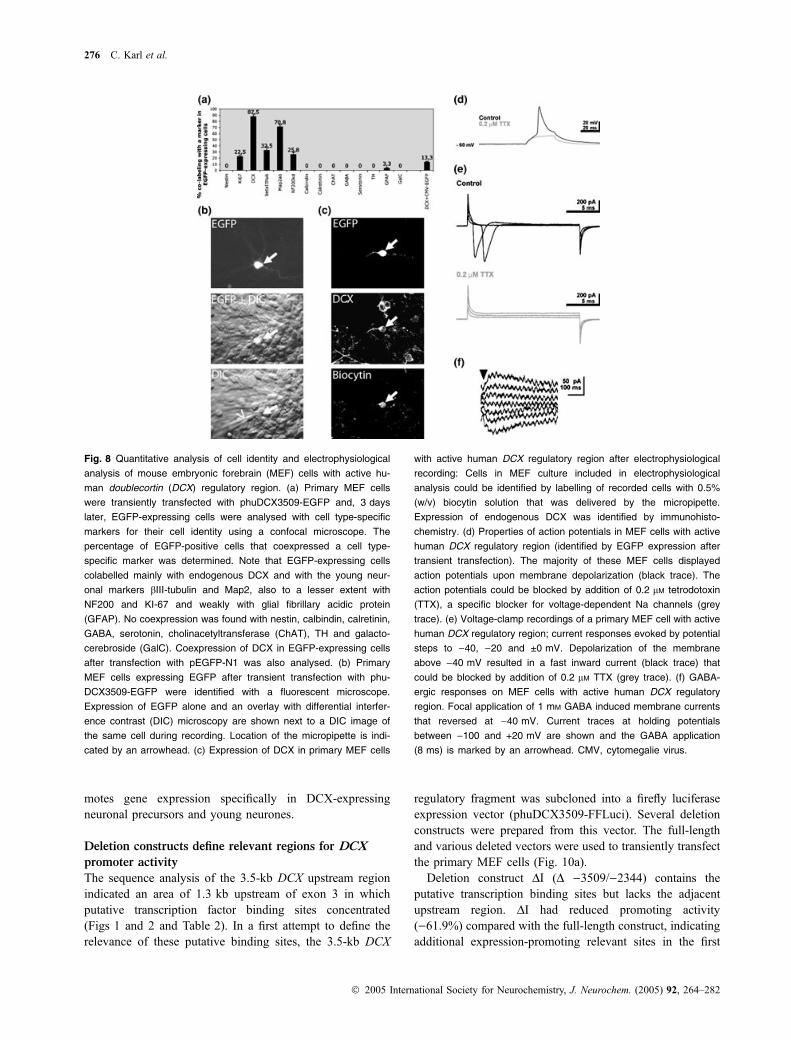
As a next step, we performed a qualitative (Fig. 7) andquantitative (Fig. 8a) analysis of the cell types expressingEGFP after transient transfection of primary MEF cells withthe phuDCX3509-EGFP vector. Confocal microscopyrevealed that the vast majority of EGFP-expressing cellswere immunoreactive for endogenous DCX (87.5% of the
Fig. 4 Activity of the human 3.5-kb dou-
blecortin (DCX) regulatory region is specific
for cells of neuronal lineage. Different cell
lines were transiently transfected with either
pEGFP-N1 or phuDCX3509-EGFP and
analysed for EGFP expression 2 days after
transfection. The EGFP signal shown was
not amplified with an antibody against
EGFP; cell nuclei are labelled with 4¢,6¢-di-amidino-2-phenylindole dihydrochloride
hydrate (DAPI). All cell lines were trans-
fectable, as demonstrated by transfection
with pEGFP-N1, in which the EGFP repor-
ter is under the control of the ubiquitous
CMV promoter. phuDCX3509-EGFP led to
the expression of EGFP almost exclusively
in neuronal cell types D283Med, Neuro2A
and primary mouse embryonic forebrain cell
cultures (MEF). Weak expression was
found in COS7 cells. Scale bar, 100 lm.
272 C. Karl et al.
� 2005 International Society for Neurochemistry, J. Neurochem. (2005) 92, 264–282

EGFP-expressing cells) (Figs 7 and 8a). EGFP-expressingcells also colabelled for class III b-tubulin isotype (32.5%),Map2ab (70.8%) and NF200 (25.8%). Coexpression ofKI-67 was found in 22.5% of the EGFP-positive cells butnone of the EGFP-positive cells expressed nestin orgalactocerebroside. A small number of EGFP-positive cellswere positive for GFAP (3.3%). This could, however, be dueto some overlap between neuronal markers and GFAP in cellculture conditions. EGFP-expressing cells displayed mor-phology reminiscent of neuronal precursors and youngneurones, indicated by a small cell soma and a bipolar to
oligopolar neurite pattern (Fig. 7). To exclude the possibilitythat the majority of EGFP-positive cells express DCX due toa preferential transfection in these cells, primary MEF cellswere transfected with pEGFP-N1 and EGFP-positive cellswere analysed for DCX expression. Only 13.3% of EGFP-positive cells coexpressed DCX after 1 week in culture. Thisnumber is in accordance with the 18.3% of cells expressingDCX in these cultures (Fig. 6d), indicating that the DCX-expressing cells are not preferentially transfected. In sum-mary, the data indicate that the 3509-bp DCX regulatoryelement used in this study is sufficient to drive specificexpression of the EGFP reporter gene in neuronal precursors.In addition, the data indicate that the activity of the 3509-bpregulatory fragment corresponds to the expression pattern ofthe endogenous DCX gene.
Additional data supporting the young neuronal specificityof the 3509-bp regulatory DCX fragment are derived fromelectrophysiological recordings. To this end, primary MEFcells were transfected with phuDCX3509-EGFP and,3–4 days later, EGFP-expressing cells were analysed elec-trophysiologically using whole-cell patch-clamp recordings(Fig. 8b). After recording, we could detect in some cells theelectrode filling-derived Biocytin signal, the EGFP, togetherwith the endogenous DCX by immunocytochemistry(Fig. 8c). A total of six EGFP-expressing and two non-EGFP-expressing cells were recorded. The resting membranepotential of the EGFP-expressing cells was )42.5 ± 7.7 mV(mean ± SD, n ¼ 6) with an Rinput of 407 ± 173 MW (fornon-EGFP-expressing neurons, )39.0 ± 22.6 mV, 295 ±113.1 MW). In four of the six EGFP-expressing and in thetwo non-EGFP-expressing cells action potentials wereobserved. In EGFP-expressing cells, action potentials had athreshold of )31.2 ± 9.7 mV (vs. )38 ± 0.8 mV for non-EGFP-expressing cells), an amplitude of 29.5 ± 18.4 mV(vs. 30.9 ± 1.4 mV for non-EGFP-expressing cells) and aduration of 5.7 ± 1.1 ms (vs. 3.3 ± 0.9 ms for non-EGFP-expressing cells). The small amplitude and relatively longduration action potentials of EGFP-expressing cells arecharacteristic of immature neurones. Additional electrophys-iological data demonstrate that these action potentials couldbe blocked by low concentrations of the sodium channelblocker tetrodotoxin (0.2 mM) (Fig. 8d). Similarly, thetransient inward current that is induced by membranedepolarization above )40 mV was blocked by low concen-trations of tetrodotoxin (0.2 mM) (Fig. 8e). Focal applicationof 1 mM GABA on two EGFP-expressing cells furtherdemonstrated the presence of GABA receptors. However, thesmall amplitude of the GABAergic membrane currentssuggests low receptor density on the cells (Fig. 8f). Insummary, the electrophysiological data indicate that EGFP-expressing cells are immature young neurones suggestingthat the 3.5-kb DCX regulatory fragment used in this study issufficient and specific for young neurones and neuronalprecursors.
Fig. 5 Quantitative analysis of EGFP expression in different cell lines.
Different cell lines were transiently transfected with pEGFP-N1 or
phuDCX3509-EGFP. Two days after transfection, cells expressing
EGFP were quantified. (a) Percentage of cells expressing EGFP after
transfection with pEGFP-N1 (h) or phuDCX3509-EGFP (j),
respectively. Three individual transfection experiments in which
pEGFP-N1 and phuDCX3509-EGFP transfections performed in par-
allel were analysed. (b) Relative expression efficacy of phuDCX3509-
EGFP [(% cells expressing phuDCX3509-EGFP/% cells expressing
pEGFP-N1) · 100]. Note the relatively high expression efficacy in
primary mouse embryonic forebrain (MEF) cells despite the low
absolute number of EGFP-expressing MEF cells after transfection with
phuDCX3509-EGFP. Statistical analysis using a Student’s t-test
revealed a high level of significance between MEF and COS7 cells
(**p < 0.005, n ¼ 3) and a low level of significance between D283Med
and COS7 cells (*0.1 > p > 0.05, n ¼ 3).
DCX promoter identification 273
� 2005 International Society for Neurochemistry, J. Neurochem. (2005) 92, 264–282

DCX regulatory region is predominantly active
in DCX-expressing neuronal precursors and young
neurones of post-natal and adult neurogenic regions
In the next set of experiments, we qualitatively assessedwhether the 3.5-kb DCX regulatory regions promote aneuronal precursor- and young neuronal-specific activity inpost-natal and adult neurogenic regions. Here, we usedballistic transfection experiments in post-natal hippocampalslice cultures and in adult explant cultures of the SVZ andOB. The hippocampal slice culture system provides aconvenient and relevant model system for the present studyas it allows manipulation and, more importantly, it preserves
many of the topographical features of the in vivo situationsuch as neurogenesis at the subgranular lining, connectivityand wiring (Prang et al. 2001; Raineteau et al. 2004).Similarly, migration of neuronal precursors outward fromOB explants into the surrounding matrigel mimics the in vivomigration away from the SVZ or the dispersion within theadult OB. Moreover, the adult OB contains a high number ofmigrating DCX-expressing cells (Brown et al. 2003) andtherefore it constitutes a good source of cells for studies ofDCX regulatory elements.
Hippocampal slice cultures derived from post-natal mousebrain were transiently transfected using ballistic methods with
Fig. 6 Cell type analysis in primary mouse
embryonic forebrain (MEF) cell cultures. (a)
Primary MEF cells were grown for 7 days in
culture, fixed and processed for immuno-
stainings with antibodies specific for
doublecortin (DCX), KI-67, nestin, NF200,
bIII-tubulin, glial fibrillary acidic protein
(GFAP), Map2, calbindin, calretinin, GABA,
serotonin, cholinacetyltransferase (ChAT),
TH and galactocerebroside (GalC) and
analysed on a confocal microscope. DCX
expression was found in many cells with
elongated processes. Colabelling of DCX-
expressing cells was found with bIII-tubulin
and Map2, some with NF200 and Ki-67 and
none with nestin, GFAP or GalC. Bar,
100 lm. (b–e) Quantitative cell type analy-
sis in primary MEF cells. MEF cells were
grown for 2 days (b and c) and 7 days
(d and e) in vitro, fixed and processed for
immunostainings with antibodies specific
for DCX, KI-67, nestin, NF200, bIII-tubulin,
GFAP, Map2, calbindin, calretinin, GABA,
serotonin, ChAT, TH and GalC. (b and d)
Percentage of cells expressing specific
markers. (c and e) Percentage of DCX-
expressing cells that coexpress a specific
marker. Note that DCX is expressed mainly
in young neurones expressing bIII-tubulin
and Map2.
274 C. Karl et al.
� 2005 International Society for Neurochemistry, J. Neurochem. (2005) 92, 264–282

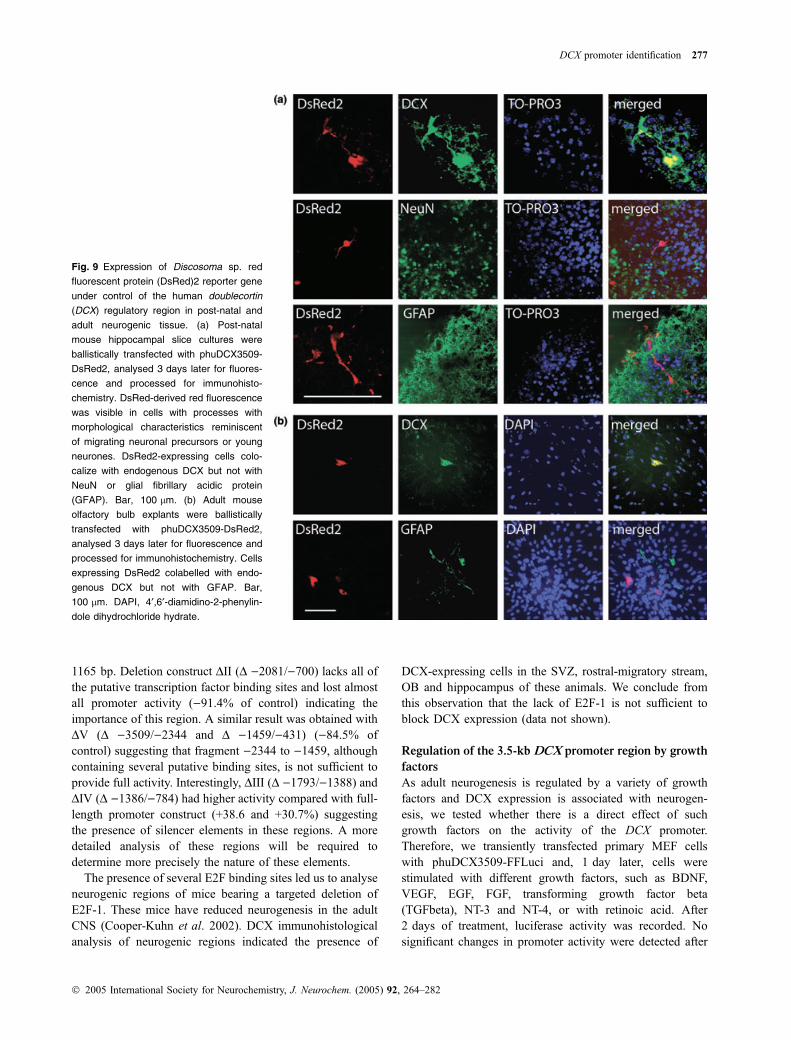
phuDCX3509-DsRed2 or the control vector pDsRed2-N1.Two to four days after transfection, expression of thefluorescent reporter was readily visible in up to 30 cells perslice when the control vector pDsRed2-N1 was employedand in up to five cells per slice when phuDCX3509-DsRed2 was used for transfection. Cells transfected withthe control vector displayed a variety of morphologicalfeatures reminiscent of astrocytes, neuronal precursors andmature neurones. The diversity suggests that different celltypes were transfected and able to express the reportertransgene (data not shown). Slices transfected with phu-DCX3509-DsRed2 were analysed more extensively. In themajority of cases (40 cells analysed), DsRed2-expressing
cells were bipolar and had morphological characteristicsreminiscent of migrating neuronal precursors or youngneurones (Fig. 9a). DsRed2-expressing cells always colo-calized with endogenous DCX but not with NeuN or GFAP(Fig. 9a).
Ballistic transfection of adult mouse OB explants yieldedsimilar results as obtained with the hippocampal slicecultures. After transfection with phuDCX3509-DsRed2, cellsexpressing the fluorescent reporter colabelled with endo-genous DCX (21 of 25 reporter-expressing cells), but notwith GFAP (Fig. 9b). In summary, the ballistic transfectionexperiments in both hippocampal and OB cultures furthersubstantiate that the 3.5-kb DCX regulatory element pro-
Fig. 7 Analysis of cell identity of mouse
embryonic forebrain (MEF) cells with active
human doublecortin (DCX) regulatory
region. Primary MEF cells were transiently
transfected with phuDCX3509-EGFP and,
3 days later, EGFP-expressing cells were
analysed with cell type-specific markers for
their cell identity using a confocal micro-
scope. Left lane, EGFP-expressing cells;
second left lane, expression of cell type-
specific markers; third left lane, cell nuclei
stained with To-pro-3; right lane, merged
illustration. EGFP-positive cells coex-
pressed DCX, Map2 and bIII-tubulin. Some
EGFP cells coexpressed KI-67, very few
EGFP cells expressed glial fibrillary acidic
protein (GFAP) and none of the EGFP cells
expressed nestin. Bar, 100 lm.
DCX promoter identification 275
� 2005 International Society for Neurochemistry, J. Neurochem. (2005) 92, 264–282
motes gene expression specifically in DCX-expressingneuronal precursors and young neurones.
Deletion constructs define relevant regions for DCX
promoter activity
The sequence analysis of the 3.5-kb DCX upstream regionindicated an area of 1.3 kb upstream of exon 3 in whichputative transcription factor binding sites concentrated(Figs 1 and 2 and Table 2). In a first attempt to define therelevance of these putative binding sites, the 3.5-kb DCX
regulatory fragment was subcloned into a firefly luciferaseexpression vector (phuDCX3509-FFLuci). Several deletionconstructs were prepared from this vector. The full-lengthand various deleted vectors were used to transiently transfectthe primary MEF cells (Fig. 10a).
Deletion construct DI (D )3509/)2344) contains theputative transcription binding sites but lacks the adjacentupstream region. DI had reduced promoting activity()61.9%) compared with the full-length construct, indicatingadditional expression-promoting relevant sites in the first
Fig. 8 Quantitative analysis of cell identity and electrophysiological
analysis of mouse embryonic forebrain (MEF) cells with active hu-
man doublecortin (DCX) regulatory region. (a) Primary MEF cells
were transiently transfected with phuDCX3509-EGFP and, 3 days
later, EGFP-expressing cells were analysed with cell type-specific
markers for their cell identity using a confocal microscope. The
percentage of EGFP-positive cells that coexpressed a cell type-
specific marker was determined. Note that EGFP-expressing cells
colabelled mainly with endogenous DCX and with the young neur-
onal markers bIII-tubulin and Map2, also to a lesser extent with
NF200 and KI-67 and weakly with glial fibrillary acidic protein
(GFAP). No coexpression was found with nestin, calbindin, calretinin,
GABA, serotonin, cholinacetyltransferase (ChAT), TH and galacto-
cerebroside (GalC). Coexpression of DCX in EGFP-expressing cells
after transfection with pEGFP-N1 was also analysed. (b) Primary
MEF cells expressing EGFP after transient transfection with phu-
DCX3509-EGFP were identified with a fluorescent microscope.
Expression of EGFP alone and an overlay with differential interfer-
ence contrast (DIC) microscopy are shown next to a DIC image of
the same cell during recording. Location of the micropipette is indi-
cated by an arrowhead. (c) Expression of DCX in primary MEF cells
with active human DCX regulatory region after electrophysiological
recording: Cells in MEF culture included in electrophysiological
analysis could be identified by labelling of recorded cells with 0.5%
(w/v) biocytin solution that was delivered by the micropipette.
Expression of endogenous DCX was identified by immunohisto-
chemistry. (d) Properties of action potentials in MEF cells with active
human DCX regulatory region (identified by EGFP expression after
transient transfection). The majority of these MEF cells displayed
action potentials upon membrane depolarization (black trace). The
action potentials could be blocked by addition of 0.2 lM tetrodotoxin
(TTX), a specific blocker for voltage-dependent Na channels (grey
trace). (e) Voltage-clamp recordings of a primary MEF cell with active
human DCX regulatory region; current responses evoked by potential
steps to )40, )20 and ±0 mV. Depolarization of the membrane
above )40 mV resulted in a fast inward current (black trace) that
could be blocked by addition of 0.2 lM TTX (grey trace). (f) GABA-
ergic responses on MEF cells with active human DCX regulatory
region. Focal application of 1 mM GABA induced membrane currents
that reversed at )40 mV. Current traces at holding potentials
between )100 and +20 mV are shown and the GABA application
(8 ms) is marked by an arrowhead. CMV, cytomegalie virus.
276 C. Karl et al.
� 2005 International Society for Neurochemistry, J. Neurochem. (2005) 92, 264–282
1165 bp. Deletion construct DII (D )2081/)700) lacks all ofthe putative transcription factor binding sites and lost almostall promoter activity ()91.4% of control) indicating theimportance of this region. A similar result was obtained withDV (D )3509/)2344 and D )1459/)431) ()84.5% ofcontrol) suggesting that fragment )2344 to )1459, althoughcontaining several putative binding sites, is not sufficient toprovide full activity. Interestingly, DIII (D )1793/)1388) andDIV (D )1386/)784) had higher activity compared with full-length promoter construct (+38.6 and +30.7%) suggestingthe presence of silencer elements in these regions. A moredetailed analysis of these regions will be required todetermine more precisely the nature of these elements.
The presence of several E2F binding sites led us to analyseneurogenic regions of mice bearing a targeted deletion ofE2F-1. These mice have reduced neurogenesis in the adultCNS (Cooper-Kuhn et al. 2002). DCX immunohistologicalanalysis of neurogenic regions indicated the presence of
DCX-expressing cells in the SVZ, rostral-migratory stream,OB and hippocampus of these animals. We conclude fromthis observation that the lack of E2F-1 is not sufficient toblock DCX expression (data not shown).
Regulation of the 3.5-kb DCX promoter region by growth
factors
As adult neurogenesis is regulated by a variety of growthfactors and DCX expression is associated with neurogen-esis, we tested whether there is a direct effect of suchgrowth factors on the activity of the DCX promoter.Therefore, we transiently transfected primary MEF cellswith phuDCX3509-FFLuci and, 1 day later, cells werestimulated with different growth factors, such as BDNF,VEGF, EGF, FGF, transforming growth factor beta(TGFbeta), NT-3 and NT-4, or with retinoic acid. After2 days of treatment, luciferase activity was recorded. Nosignificant changes in promoter activity were detected after
Fig. 9 Expression of Discosoma sp. red
fluorescent protein (DsRed)2 reporter gene
under control of the human doublecortin
(DCX) regulatory region in post-natal and
adult neurogenic tissue. (a) Post-natal
mouse hippocampal slice cultures were
ballistically transfected with phuDCX3509-
DsRed2, analysed 3 days later for fluores-
cence and processed for immunohisto-
chemistry. DsRed-derived red fluorescence
was visible in cells with processes with
morphological characteristics reminiscent
of migrating neuronal precursors or young
neurones. DsRed2-expressing cells colo-
calize with endogenous DCX but not with
NeuN or glial fibrillary acidic protein
(GFAP). Bar, 100 lm. (b) Adult mouse
olfactory bulb explants were ballistically
transfected with phuDCX3509-DsRed2,
analysed 3 days later for fluorescence and
processed for immunohistochemistry. Cells
expressing DsRed2 colabelled with endo-
genous DCX but not with GFAP. Bar,
100 lm. DAPI, 4¢,6¢-diamidino-2-phenylin-
dole dihydrochloride hydrate.
DCX promoter identification 277
� 2005 International Society for Neurochemistry, J. Neurochem. (2005) 92, 264–282

the various growth factor treatments (Fig. 10b). Therelatively short period of growth factor stimulation waschosen to put direct signalling events in evidence and toavoid artefacts caused by alterations in the cellularcomposition of the cultures. The growth factors used areknown to influence proliferation, survival and maturationof neural cells and could thereby influence the long-termcomposition of the cultures. The slight decrease in activityassociated with retinoic acid, VEGF and FGF treatmentmight be due to enhanced neuronal maturation of the cells.The decrease in activity induced by EGF might relate tothe proliferative effect of EGF on neural stem cells, whichdo not express DCX.
Discussion
The present work demonstrates that a 3.5-kb fragmentupstream of the DCX ATG start codon is sufficient andspecific to drive expression of reporter genes in primaryneuronal precursors and young neurones of dissociated celland organotypic slice culture systems of neurogenic regionsas determined by immunohistological analysis. Moreover,the activity of the 3.5-kb DCX regulatory fragment over-lapped with the expression of the endogenous DCX,suggesting that relevant sequences for regulating DCXexpression are located within this fragment. Minor expres-sion was detected in two types of non-neuronal cells, COS7cells and a few GFAP-expressing embryonic cells. This couldbe due to some leakage of the 3.5-kb fragment, as it was alsodemonstrated for the Tau promoter (Heicklen-Klein andGinzburg 2000), or due to in vitro artefacts of transienttransfection experiments, such as integration events or highcopy number of transgene.
Electrophysiological data confirmed the immature neur-onal character of cells having the capacity to activate theDCX promoter. This was revealed, for example, by theirsmall amplitude and relatively long duration action poten-
tials. Similar immature neuronal characteristics have beendescribed in hippocampal slices of mice expressing EGFPunder the pro-opiomelanocortin promoter (Overstreet et al.2004). In these mice, EGFP expression is confined to youngneurones in the hippocampal dentate gyrus.
Several studies have established the expression pattern ofDCX in adult neurogenic regions and indicated that DCXwas specific for neuronal precursors and young immatureneurones (Brown et al. 2003; Rao and Shetty 2004). In vivo,DCX-expressing cells do not constitute a uniform andhomogeneous population but rather a mixture of newlygenerated neuroblasts and up to 4-week-old neuronalprecursors. Therefore, some neuronal precursors are stillmitotically active whereas others induce the expression ofNeuN (Brown et al. 2003; Kempermann et al. 2004).
On the other hand, in vitro, the DCX expression pattern isless known and only few data are available concerning theuse of DCX as a marker for in vitro neurogenesis study. Forexample, DCX was shown to be expressed in 30–40% ofhippocampal-derived differentiating neural stem cell cultures(Monje et al. 2003) and DCX was localized in differenti-ating retinal progenitors in vitro (Engelhardt et al. 2004).The present work demonstrates colocalization of DCX, orEGFP driven by the 3.5-kb DCX regulatory region, withyoung neuronal markers such as class III b-tubulin isotypeand Map2ab, seldom with the mature neuronal markerNF200 and not with glial or stem cell markers. Thesecolocalization analyses suggest a clear demarcation ofDCX-expressing neuronal precursors from stem cells, gliallineages and mature neurones. In organotypic hippocampalslice cultures, the absence of DCX promoter-driven DsRed2in NeuN-expressing neurones further confirms this findingin a culture model reproducing even more closely thephysiological situation. Moreover, primary MEF cellsdemonstrated that cells in which the DCX regulatorysequence was active were immature in terms of neurotrans-mitter specification.
Fig. 10 Relevance of distinct positions on
the 3509-bp doublecortin (DCX) regulatory
region for reporter gene expression and
regulation by growth factors. Deletion con-
structs were transiently transfected in pri-
mary mouse embryonic forebrain (MEF)
cells and, 2 days later, DCX promoter-dri-
ven firefly luciferase activity was analysed
and normalized for CMV-driven Renilla-lu-
ciferase and for full-length 3.5-kb DCX
promoter fragment. (b) Primary MEF cells
were transiently transfected with phu-
DCX3509-FFLuci and, 1 day later, cells
were stimulated with different factors.
Luciferase activity was determined 2 days
later as described in (a). RA, retinoic acid.
278 C. Karl et al.
� 2005 International Society for Neurochemistry, J. Neurochem. (2005) 92, 264–282

The demarcation between the DCX-expressing cells andthe neural stem cells was recently occulted by resultsobtained using a transgenic mouse model in which theneuronal element of the nestin enhancer controls theexpression of the reporter protein EGFP (Kronenberg et al.2003). In these animals, colocalization of the EGFP reporterwith DCX was documented. Expression of reporter proteinsunder a specific promoter, however, does not necessarilyreproduce exactly the expression patterns of the correspond-ing endogenous gene. This could be due to the influence oftransgene insertion site, lack of some genomic elements (e.g.introns) as well as differences in stability of the mRNA andpolypeptide. For example, it was demonstrated that injectedEGFP mRNAs in Xenopus embryos led to a fluorescencesignal that was detectable for more than 21 days (Verkhushaet al. 2003). Therefore, in nestin-EGFP transgenic mice, alonger persistence of the reporter protein in differentiatingneural stem cells could be the cause of the documentedcolocalization of DCX with the EGFP reporter. Similarly, inprimary MEF-transfected cells, longer persistence of theEGFP reporter might be at the origin of the small populationof EGFP-expressing cells that also express the matureneuronal marker NF200.
Unfortunately, the transient transfection techniqueemployed in the present study does not allow for long-termanalysis. Therefore, it is impossible to follow dynamicchanges in promoter activity such as the expected down-regulation during neuronal maturation or the expectedinduction during neuronal determination. Only circumstantialevidence based on expression profiles obtained with the DCXpromoter fragment and the ubiquitous CMV promotersuggests that the DCX promoter activity might be down-regulated during neuronal maturation and not be active inmature neurones. More detailed analysis of the dynamicbehaviour of the DCX promoter fragment is desirable butsuch studies require either stable transfections, retroviralinsertions of the construct or transgenic mice.
Several putative binding sites for transcription factorsinvolved in neuronal development, including Brn-2,NeuroD1, E2F-1 and E2F-2, are localized within the3.5-kb DCX regulatory element. Transfection studies usingdeletion constructs indicated the relevance of regions con-taining these binding sites. However, additional work isrequired to identify specifically the role of these transcriptionfactors in the global activity of the DCX promoter. Previousstudies on various knockout mouse lines substantiate theexistence of a link between some of these transcriptionfactors and DCX promoter activity. For example, Brn-2, aclass III POU (Pit, Oct, Unc) domain transcription factor,might be a candidate for regulating DCX gene activation.Brn-2 is expressed in migrating neuronal precursors, where itis required for cell-autonomous control of radial glia-guidedmigration (McEvilly et al. 2002). Moreover, mice bearing atargeted deletion of the Brn-2 gene have neuronal migration
defects reminiscent of the Reelin/Cdk5/DCX signallingpathway (McEvilly et al. 2002). NeuroD is also a promisingtranscription factor candidate for the regulation of DCXexpression as it regulates several cell autonomous neuronaldetermination and differentiation mechanisms. Targeteddeletion of NeuroD in mice led to hippocampal malforma-tions similar to those described in DCX knockout mice (Liuet al. 2000; Corbo et al. 2002). The cell cycle regulatorsE2F-1 and E2F-2 might also regulate DCX expression.Although mice with a targeted deletion of the E2F-1 gene didnot display overt brain malformations, the rate of adultneurogenesis was reduced compared with wild-type mice(Cooper-Kuhn et al. 2002). However, our preliminary resultsdemonstrate that the lack of E2F-1 in these mice did notabolish DCX expression in adult neurogenic regions. Wecannot, at present, exclude the possibility that regions outsidethe one described above influence the DCX promoter activity.
Analysis of the DCX promoter is also of clinical relevance.Linkage analyses in pedigrees comprising both X-linkedlissencephaly and subcortical band heterotopia cases led tothe identification of the DCX gene (des Portes et al. 1998a;Gleeson et al. 1998; Couillard-Despres et al. 2001). Sincethen, mis-sense, nonsense, deletion and insertion mutationsin the DCX gene have been found to be associated withneuronal migration disorders (des Portes et al. 1998a,b;Gleeson et al. 1998, 1999b, 2000a,b; Aigner et al. 2000,2003). Nevertheless, at least 20% of patients with subcorticalband heterotopia do not have mutations in the DCX codingregion nor other genes associated with migration disorderssuch as LIS1 (Leventer et al. 2000). It is conceivable thatsome of these patients may have mutations in key regions ofthe DCX promoter, resulting in lower expression of DCX andin neuronal migration defects.
In addition to the diagnostic potential of the DCXregulatory sequence, a therapeutic application could beenvisaged. Up-regulation of DCX expression by acting onthe regulatory sequences in order to influence the migration ofneuronal precursor cells during development or during adultneurogenesis could be part of a therapeutic intervention. Thefeasibility of such an approach has been supported by cellculture experiments in which the migration of neuronalprecursors was enhanced by over-expression of DCX(Tanaka et al. 2004a).
The present work provides a first molecular and cellbiological description of a DCX regulatory element. A regioncorresponding to the 3.5 kb upstream of the DCX translationstart site was shown to contain the regulatory elementspromoting specific expression in neuronal precursors. This3.5-kb region contained clusters of binding sites fortranscription factor, e.g. Brn-2 and NeuroD, which wererequired for efficient activity of the fragment’s expression-promoting activity. Transgenic mice bearing the constructwill help to further determine cell specificity of the regulatoryelement and its dynamics.
DCX promoter identification 279
� 2005 International Society for Neurochemistry, J. Neurochem. (2005) 92, 264–282

Acknowledgements
The authors would like to thank the Volkswagen-Foundation
(Hannover, Germany) and Fritz-Thyssen-Foundation (Koln, Ger-
many) for funding. Thanks to Anthony T. Campagnoni (University
of California, Los Angeles, CA, USA) for the N20.1 oligodendro-
cyte line.
References
Aberg M. A., Aberg N. D., Hedbacker H., Oscarsson J. and Eriksson P.S. (2000) Peripheral infusion of IGF-I selectively induces neur-ogenesis in the adult rat hippocampus. J. Neurosci. 20, 2896–2903.
Aigner L., Fluegel D., Dietrich J., Ploetz S. and Winkler J. (2000) Iso-lated lissencephaly sequence and double-cortex syndrome in aGerman family with a novel doublecortin mutation. Neuropediat-rics 31, 195–198.
Aigner L., Uyanik G., Couillard-Despres S. et al. (2003) Somaticmosaicism and variable penetrance in doublecortin-associatedmigration disorders. Neurology 60, 329–332.
Alvarez-Buylla A. and Garcia-Verdugo J. M. (2002) Neurogenesis inadult subventricular zone. J. Neurosci. 22, 629–634.
Alvarez-Buylla A. and Temple S. (1998) Stem cells in the developingand adult nervous system. J. Neurobiol. 36, 105–110.
Alvarez-Buylla A., Garcia-Verdugo J. M. and Tramontin A. D. (2001) Aunified hypothesis on the lineage of neural stem cells. Nat. Rev.Neurosci. 2, 287–293.
Arvidsson A., Collin T., Kirik D., Kokaia Z. and Lindvall O. (2002)Neuronal replacement from endogenous precursors in the adultbrain after stroke. Nat. Med. 8, 963–970.
Bai J., Ramos R. L., Ackman J. B., Thomas A. M., Lee R. V. andLoTurco J. J. (2003) RNAi reveals doublecortin is required forradial migration in rat neocortex. Nat. Neurosci. 6, 1277–1283.
Brown J. P., Couillard-Despres S., Cooper-Kuhn C. M., Winkler J.,Aigner L. and Kuhn H. G. (2003) Transient expression of dou-blecortin during adult neurogenesis. J. Comp. Neurol. 467, 1–10.
Cooper-Kuhn C. M. and Kuhn H. G. (2002) Is it all DNA repair?Methodological considerations for detecting neurogenesis in theadult brain. Brain Res. Dev. Brain Res. 134, 13–21.
Cooper-Kuhn C. M., Vroemen M., Brown J., Ye H., Thompson M. A.,Winkler J. and Kuhn H. G. (2002) Impaired adult neurogenesis inmice lacking the transcription factor E2F1. Mol. Cell Neurosci. 21,312–323.
Corbo J. C., Deuel T. A., Long J. M., LaPorte P., Tsai E., Wynshaw-Boris A. and Walsh C. A. (2002) Doublecortin is required in micefor lamination of the hippocampus but not the neocortex. J. Neu-rosci. 22, 7548–7557.
Couillard-Despres S., Winkler J., Uyanik G. and Aigner L. (2001)Molecular mechanisms of neuronal migration disorders, Quo Va-dis? Curr. Mol. Med. 1, 677–688.
Couillard-Despres S., Uyanik G., Ploetz S., Karl C., Koch H., Winkler J.and Aigner L. (2004) Mitotic impairment by doublecortin isdiminished by doublecortin mutations found in patients. Neuro-genetics 5, 83–93.
Dash P. K., Mach S. A. and Moore A. N. (2001) Enhanced neurogenesisin the rodent hippocampus following traumatic brain injury.J. Neurosci. Res. 63, 313–319.
Doetsch F., Caille I., Lim D. A., Garcia-Verdugo J. M. and Alvarez-Buylla A. (1999) Subventricular zone astrocytes are neural stemcells in the adult mammalian brain. Cell 97, 703–716.
Doetsch F., Petreanu L., Caille I., Garcia-Verdugo J. M. and Alvarez-Buylla A. (2002) EGF converts transit-amplifying neurogenic
precursors in the adult brain into multipotent stem cells. Neuron 36,1021–1034.
Engelhardt M., Wachs F. P., Couillard-Despres S. and Aigner L. (2004)The neurogenic competence of progenitors from the postnatal ratretina in vitro. Exp. Eye Res. 78, 1025–1036.
Englund U., Bjorklund A. and Wictorin K. (2002) Migration patternsand phenotypic differentiation of long-term expanded humanneural progenitor cells after transplantation into the adult rat brain.Brain Res. Dev. Brain Res. 134, 123–141.
Francis F., Koulakoff A., Boucher D. et al. (1999) Doublecortin is adevelopmentally regulated, microtubule-associated proteinexpressed in migrating and differentiating neurons. Neuron 23,247–256.
Frisen J., Johansson C. B., Torok C., Risling M. and Lendahl U. (1995)Rapid, widespread, and longlasting induction of nestin contributesto the generation of glial scar tissue after CNS injury. J. Cell Biol.131, 453–464.
Gage F. H. (2000) Mammalian neural stem cells. Science 287, 1433–1438.
Gleeson J. G., Allen K. M., Fox J. W. et al. (1998) Doublecortin, abrain-specific gene mutated in human X-linked lissencephaly anddouble cortex syndrome, encodes a putative signaling protein. Cell92, 63–72.
Gleeson J. G., Lin P. T., Flanagan L. A. and Walsh C. A. (1999a)Doublecortin is a microtubule-associated protein and is expressedwidely by migrating neurons. Neuron 23, 257–271.
Gleeson J. G., Minnerath S. R., Fox J. W. et al. (1999b) Characterizationof mutations in the gene doublecortin in patients withdouble cortex syndrome [see comments]. Ann. Neurol. 45, 146–153.
Gleeson J. G., Minnerath S., Kuzniecky R. I., Dobyns W. B., Young I.D., Ross M. E. and Walsh C. A. (2000a) Somatic and germlinemosaic mutations in the doublecortin gene are associated withvariable phenotypes. Am. J. Hum. Genet. 67, 574–581.
Gleeson J. G., Luo R. F., Grant P. E. et al. (2000b) Genetic and neu-roradiological heterogeneity of double cortex syndrome. Ann.Neurol. 47, 265–269.
Gloster A., Wu W., Speelman A., Weiss S., Causing C., Pozniak C.,Reynolds B., Chang E., Toma J. G. and Miller F. D. (1994) The Talpha 1 alpha-tubulin promoter specifies gene expression as afunction of neuronal growth and regeneration in transgenic mice.J. Neurosci. 14, 7319–7330.
Gotz M., Hartfuss E. and Malatesta P. (2002) Radial glial cells asneuronal precursors: a new perspective on the correlation of mor-phology and lineage restriction in the developing cerebral cortex ofmice. Brain Res. Bull. 57, 777–788.
Gravel M., Di Polo A., Valera P. B. and Braun P. E. (1998) Four-kilobasesequence of the mouse CNP gene directs spatial and temporalexpression of lacZ in transgenic mice. J. Neurosci. Res. 53, 393–404.
Haubensak W., Attardo A., Denk W. and Huttner W. B. (2004) Neuronsarise in the basal neuroepithelium of the early mammalian telen-cephalon: a major site of neurogenesis. Proc. Natl Acad. Sci. USA101, 3196–3201.
Heicklen-Klein A. and Ginzburg I. (2000) Tau promoter confers neur-onal specificity and binds Sp1 and AP-2. J. Neurochem. 75, 1408–1418.
Jin K., Zhu Y., Sun Y., Mao X. O., Xie L. and Greenberg D. A. (2002)Vascular endothelial growth factor (VEGF) stimulates neurogene-sis in vitro and in vivo. Proc. Natl Acad. Sci. USA 99, 11 946–11 950.
Jin K., Sun Y., Xie L., Peel A., Mao X. O., Batteur S. and Greenberg D. A.(2003a) Directed migration of neuronal precursors into the ischemiccerebral cortex and striatum. Mol. Cell Neurosci. 24, 171–189.
280 C. Karl et al.
� 2005 International Society for Neurochemistry, J. Neurochem. (2005) 92, 264–282

Jin K., Xie L., Childs J., Sun Y., Mao X. O., Logvinova A. andGreenberg D. A. (2003b) Cerebral neurogenesis is induced byintranasal administration of growth factors. Ann. Neurol. 53, 405–409.
Jin K., Sun Y., Xie L., Batteur S., Mao X. O., Smelick C., Logvinova A.and Greenberg D. A. (2003c) Neurogenesis and aging: FGF-2 andHB-EGF restore neurogenesis in hippocampus and subventricularzone of aged mice. Aging Cell 2, 175–183.
Kelly S., Bliss T. M., Shah A. K. et al. (2004) Transplanted human fetalneural stem cells survive, migrate, and differentiate in ischemic ratcerebral cortex. Proc. Natl Acad. Sci. USA 101, 11 839–11 844.
Kempermann G., Kuhn H. G. and Gage F. H. (1997) More hippocampalneurons in adult mice living in an enriched environment. Nature386, 493–495.
Kempermann G., Jessberger S., Steiner B. and Kronenberg G. (2004)Milestones of neuronal development in the adult hippocampus.Trends Neurosci. 27, 447–452.
Kronenberg G., Reuter K., Steiner B., Brandt M. D., Jessberger S.,Yamaguchi M. and Kempermann G. (2003) Subpopulations ofproliferating cells of the adult hippocampus respond differently tophysiologic neurogenic stimuli. J. Comp. Neurol. 467, 455–463.
Kuhn H. G., Winkler J., Kempermann G., Thal L. J. and Gage F. H.(1997) Epidermal growth factor and fibroblast growth factor-2have different effects on neural progenitors in the adult rat brain.J. Neurosci. 17, 5820–5829.
Leventer R. J., Pilz D. T., Matsumoto N., Ledbetter D. H. and DobynsW. B. (2000) Lissencephaly and subcortical band heterotopia:molecular basis and diagnosis. Mol. Med. Today 6, 277–284.
Liu J., Solway K., Messing R. O. and Sharp F. R. (1998) Increasedneurogenesis in the dentate gyrus after transient global ischemia ingerbils. J. Neurosci. 18, 7768–7778.
Liu M., Pleasure S. J., Collins A. E., Noebels J. L., Naya F. J., Tsai M. J.and Lowenstein D. H. (2000) Loss of BETA2/NeuroD leads tomalformation of the dentate gyrus and epilepsy. Proc. Natl Acad.Sci. USA 97, 865–870.
Magavi S. S., Leavitt B. R. and Macklis J. D. (2000) Induction ofneurogenesis in the neocortex of adult mice. Nature 405, 951–955.
McEvilly R. J., de Diaz M. O., Schonemann M. D., Hooshmand F. andRosenfeld M. G. (2002) Transcriptional regulation of corticalneuron migration by POU domain factors. Science 295, 1528–1532.
Monje M. L., Toda H. and Palmer T. D. (2003) Inflammatory blockaderestores adult hippocampal neurogenesis. Science 302, 1760–1765.
Nacher J., Crespo C. and McEwen B. S. (2001) Doublecortin expressionin the adult rat telencephalon. Eur. J. Neurosci. 14, 629–644.
Overstreet L. S., Hentges S. T., Bumaschny V. F., de Souza F. S., SmartJ. L., Santangelo A. M., Low M. J., Westbrook G. L. and Rubin-stein M. (2004) A transgenic marker for newly born granule cellsin dentate gyrus. J. Neurosci. 24, 3251–3259.
Parent J. M., Yu T. W., Leibowitz R. T., Geschwind D. H., Sloviter R. S.and Lowenstein D. H. (1997) Dentate granule cell neurogenesis isincreased by seizures and contributes to aberrant network reor-ganization in the adult rat hippocampus. J. Neurosci. 17, 3727–3738.
des Portes V., Pinard J. M., Billuart P. et al. (1998a) A novel CNS generequired for neuronal migration and involved in X-linked subcor-tical laminar heterotopia and lissencephaly syndrome. Cell 92,51–61.
des Portes V., Francis F., Pinard J. M. et al. (1998b) Doublecortin is themajor gene causing X-linked subcortical laminar heterotopia(SCLH). Hum. Mol. Genet. 7, 1063–1070.
van Praag H., Christie B. R., Sejnowski T. J. and Gage F. H. (1999)Running enhances neurogenesis, learning, and long-term potenti-ation in mice. Proc. Natl Acad. Sci. USA 96, 13 427–13 431.
Prang P., Del Turco D. and Kapfhammer J. P. (2001) Regeneration ofentorhinal fibers in mouse slice cultures is age dependent and canbe stimulated by NT-4, GDNF, and modulators of G-proteins andprotein kinase C. Exp. Neurol. 169, 135–147.
Quandt K., Frech K., Karas H., Wingender E. and Werner T. (1995)MatInd and MatInspector: new fast and versatile tools for detectionof consensus matches in nucleotide sequence data. Nucl. Acids Res.23, 4878–4884.
Raineteau O., Rietschin L., Gradwohl G., Guillemot F. and Gahwiler B.H. (2004) Neurogenesis in hippocampal slice cultures. Mol. CellNeurosci. 26, 241–250.
Rao M. S. and Shetty A. K. (2004) Efficacy of doublecortin as a markerto analyse the absolute number and dendritic growth of newlygenerated neurons in the adult dentate gyrus. Eur. J. Neurosci. 19,234–246.
Roy N. S., Benraiss A., Wang S., Fraser R. A., Goodman R., CouldwellW. T., Nedergaard M., Kawaguchi A., Okano H. and Goldman S.A. (2000a) Promoter-targeted selection and isolation of neuralprogenitor cells from the adult human ventricular zone. J. Neuro-sci. Res. 59, 321–331.
Roy N. S., Wang S., Jiang L. et al. (2000b) In vitro neurogenesis byprogenitor cells isolated from the adult human hippocampus. Nat.Med. 6, 271–277.
Sanai N., Tramontin A. D., Quinones-Hinojosa A. et al. (2004) Uniqueastrocyte ribbon in adult human brain contains neural stem cells butlacks chain migration. Nature 427, 740–744.
Schaar B. T., Kinoshita K. and McConnell S. K. (2004) Doublecortinmicrotubule affinity is regulated by a balance of kinase andphosphatase activity at the leading edge of migrating neurons.Neuron 41, 203–213.
Seri B., Garcia-Verdugo J. M., McEwen B. S. and Alvarez-Buylla A.(2001) Astrocytes give rise to new neurons in the adult mammalianhippocampus. J. Neurosci. 21, 7153–7160.
Shi Y., Chichung Lie D., Taupin P., Nakashima K., Ray J., Yu R. T.,Gage F. H. and Evans R. M. (2004) Expression and function oforphan nuclear receptor TLX in adult neural stem cells. Nature427, 78–83.
Shingo T., Sorokan S. T., Shimazaki T. and Weiss S. (2001) Erythro-poietin regulates the in vitro and in vivo production of neuronalprogenitors by mammalian forebrain neural stem cells. J. Neurosci.21, 9733–9743.
Steiner B., Kronenberg G., Jessberger S., Brandt M. D., Reuter K. andKempermann G. (2004) Differential regulation of gliogenesis inthe context of adult hippocampal neurogenesis in mice. Glia 46,41–52.
Stoppini L., Buchs P. A. and Muller D. (1991) A simple method fororganotypic cultures of nervous tissue. J. Neurosci. Meth. 37, 173–182.
Stuart G. J., Dodt H. U. and Sakmann B. (1993) Patch-clamp recordingsfrom the soma and dendrites of neurons in brain slices usinginfrared video microscopy. Pflugers Arch. 423, 511–518.
Tanaka T., Serneo F. F., Tseng H. C., Kulkarni A. B., Tsai L. H. andGleeson J. G. (2004a) Cdk5 phosphorylation of doublecortinser297 regulates its effect on neuronal migration. Neuron 41, 215–227.
Tanaka T., Serneo F. F., Higgins C., Gambello M. J., Wynshaw-Boris A.and Gleeson J. G. (2004b) Lis1 and doublecortin function withdynein to mediate coupling of the nucleus to the centrosome inneuronal migration. J. Cell Biol. 165, 709–721.
Tang Z., Yu Y., Guo H. and Zhou J. (2002) Induction of tyrosinehydroxylase expression in rat fetal striatal precursor cells followingtransplantation. Neurosci. Lett. 324, 13–16.
Verity A. N., Bredesen D., Vonderscher C., Handley V. W. and Cam-pagnoni A. T. (1993) Expression of myelin protein genes and other
DCX promoter identification 281
� 2005 International Society for Neurochemistry, J. Neurochem. (2005) 92, 264–282

myelin components in an oligodendrocytic cell line conditionallyimmortalized with a temperature-sensitive retrovirus. J. Neuro-chem. 60, 577–587.
Verkhusha V. V., Kuznetsova I. M., Stepanenko O. V., Zaraisky A. G.,Shavlovsky M. M., Turoverov K. K. and Uversky V. N. (2003)High stability of Discosoma DsRed as compared to AequoreaEGFP. Biochemistry 42, 7879–7884.
Wachs F. P., Couillard-Despres S., Engelhardt M. et al. (2003) Highefficacy of clonal growth and expansion of adult neural stem cells.Lab. Invest. 83, 949–962.
Wang S., Roy N. S., Benraiss A. and Goldman S. A. (2000) Promoter-based isolation and fluorescence-activated sorting of mitoticneuronal progenitor cells from the adult mammalian ependymal/subependymal zone. Dev. Neurosci. 22, 167–176.
282 C. Karl et al.
� 2005 International Society for Neurochemistry, J. Neurochem. (2005) 92, 264–282





























