Embed Size (px)
Citation preview

10 INFORMAÇÕES AGRONÔMICAS Nº 147 – SETEMBRO/2014
PAPEL FISIOLÓGICO DO NÍQUEL:ESSENCIALIDADE E TOXIDEZ EM PLANTAS
André Rodrigues dos Reis1
Bruna Wurr Rodak2
1 Professor Assistente Doutor do Curso de Engenharia de Biossistemas, Universidade Estadual Paulista-UNESP, Campus de Tupã, SP; email: [email protected]
2 Doutoranda em Ciências do Solo, Universidade Federal de Lavras, Lavras, MG.3 Doutorando em Irrigação e Drenagem, Faculdade de Ciências Agronômicas, Universidade Estadual Paulista-UNESP, Campus de Botucatu, SP.4 Professor Assistente Doutor do Curso de Agronomia, Universidade Federal do Mato Grosso, Barra do Garças, MT.
Abreviações: ASE = área superficial específica; B = boro; CTC = capacidade de troca catiônica; Ca = cálcio; Co = cobalto; Cu = cobre; Fe = ferro; GSH = glutationa; GSSG = glutationa dissulfeto; Mg = magnésio; Mn = manganês; MOS = matéria orgânica do solo; MS = matéria seca; N = nitrogênio; NADPH = nicotiamida-adenina-dinucleótido-fosfato; Ni = níquel; P = fósforo; SiBCS = Sistema Brasileiro de Classificação dos Solos; Zn = zinco.
Fernando Ferrari Putti3
Milton Ferreira de Moraes4
NÍQUEL NO SOLO
O metal de transição níquel (Ni) é o vigésimo terceiro elemento mais abundante na crostra terrestre, com teor médio de 20 mg kg-1. A concentração total
de Ni nos solos varia de 4 a 2.000 mg kg-1, com valor médio de 29 mg kg-1 (KABATA-PENDIAS e PENDIAS, 2011). Entretanto, solos derivados de rochas ultramáficas, ricas em minerais do grupo serpentina, podem apresentar concentrações de até 7.100 mg kg-1 (BROOKS, 1987). Em pesquisas sobre o teor Ni em solos naturais de várias partes do mundo, Massoura et al. (2006) observaram teores altos, variando de 500 a 12.000 mg kg-1, em amostras de material geológico derivado de serpentina e peridotito, ambos constituintes de rochas ultramáficas.
Levantamentos dos teores totais em solos brasileiros ainda são escassos, sendo quantificados valores variando de < 0,014 a 1.167,52 mg kg-1 de Ni (ROVERS; CAMARGO; VALADARES, 1983; MINEROPAR, 2005a, 2005b; LICHT et al., 2006; MELO et al., 2007; CAIRES, 2009; BIONDI, 2010; PAYE et al., 2010; SANTOS, 2011; BUSCHLE, 2013). Entretanto, Reeves et al. (2007) verificaram, em amostras de solos ultramáficos da região central
de Goiás, principalmente no município de Niquelândia, valores extremamente elevados, de 618 a 14.390 mg kg-1.
Os teores de Ni nos solos apresentam elevada variabilidade e são altamente dependentes da composição do material de origem, com destaque, de maneira geral, para as classes Cambissolos e Calcisols (calcários, sem correspondência no Sistema Brasileiro de Classificação dos Solos – SiBCS).
Os teores disponíveis correspondem a 0,001% dos teores totais (UREN, 1992), na maior parte das vezes, segundo Vanselow (1966), com teores abaixo de 1 mg dm-3 de Ni. Em solos brasileiros, estudos isolados na Bahia, Goiás, Maranhão, Mato Grosso, Rio Grande do Sul e São Paulo mostraram teores disponíveis variando de < 0,013 a 1,4 mg dm-3, extraídos em DTPA, < 0,1 a 0,27 mg dm-3, em Mehlich-1, e teor médio de 0,35 mg dm-3, em Mehlich-3 (ROVERS; CAMARGO; VALADARES, 1983; MELO et al., 2007; RODAK, 2014). O estado no qual os estudos estão mais adiantados em relação à disponibilidade de Ni é o Paraná, onde amostras do horizonte B de toda extensão do estado foram analisadas (Tabela 1), constatando-se teores de < 0,013 a 0,19 e < 0,1 a 0,54 mg dm-3 em DTPA e Mehlich-1, respec-tivamente (RODAK, 2014). Amostras superficiais de solos naturais e
Tabela 1. Teores disponíveis de níquel obtidos com os extratores DTPA e Mehlich-1 em solos naturais e agrícolas do estado do Paraná.
Localização Profundidade(cm)
DTPA Mehlich-1
Solo1Médio Variação Médio VariaçãoMínimo Máximo Mínimo Máximo
- - - - - - - - - - - - - - - - - - - (mg dm-3) - - - - - - - - - - - - - - - - - - - Paraná Horizonte B 0,035 < 0,013 0,19 0,14 < 0,1 0,54 -Ampére 0-20 0,74 - - 1,33 - - Latossolo Vermelho distroférrico típicoDiamante do Sul 0-20 0,65 - - 1,1 - - Neossolo Regolítico eutrófico típicoIporã 0-20 - < 0,013 0,13 - < 0,1 0,24 Latossolo Vermelho distrófico típicoLondrina 0-20 0,45 - - 0,75 - - Latossolo Vermelho distroférrico típicoPalotina 0-20 - 0,15 0,16 - 0,33 0,37 Latossolo Vermelho Eutroférrico típicoParanavaí 0-20 0,11 - - 0,2 - - Argissolo Vermelho distrófico latossólicoPonta Grossa 0-20 0,08 - - 0,11 - - Cambissolo Háplico alumínico típicoUmuarama 0-20 0,09 - - 0,15 - - Argissolo Vermelho distrófico arênico
1 Embrapa Solos (2006).Fonte: Adaptada de Rodak (2014).

INFORMAÇÕES AGRONÔMICAS Nº 147 – SETEMBRO/2014 11
agrícolas em áreas pontuais do Paraná também foram estudadas e os teores variaram de < 0,013 a 0,74 e < 0,1 a 1,33 mg dm-3 em DTPA e Mehlich-1, respectivamente (RODAK, 2014). Ressalta-se que os maiores teores, para ambos os extratores, ocorreram em solos deri-vados de derrames de rochas basálticas, que são rochas ultrabásicas ricas em Ni, evidenciando, mais uma vez, a relação direta com o material de origem.
Diversos fatores podem afetar os teores de Ni disponíveis no solo, como classe textural, matéria orgânica do solo (MOS), óxidos de ferro (Fe) e manganês (Mn), pH, reações de complexa-ção e competições pelos sítios de adsorção (KABATA-PENDIAS e PENDIAS, 2011).
De maneira geral, os horizontes superficiais do solo apre-sentam maior disponibilidade de Ni, fato que ocorre, em parte, devido à presença de MOS, a qual forma complexos estáveis e solúveis com o elemento (UREN, 1992). Entretanto, o conteúdo de Ni na MOS é considerado baixo. Estudos de Cheshire et al. (1977) com metais-traço, entre eles o Ni, constataram que ape-nas 0,59 mg kg-1 e 0,57 mg kg-1 do total de 35 mg kg-1 de Ni no solo encontrava-se na fração de ácidos húmicos e fúlvicos, o que representa apenas 1,7% e 1,6% dos teores totais, respectivamente, ou seja, valores inferiores a 4%. Ressalta-se que o papel da MOS é mais efetivo quando são efetuadas aplicações de resíduos ao solo, como lodo de esgoto – material rico em metais pesados, incluindo Ni (SAUVé; HENDERSHOT; ALLEN, 2000; SMITH, 2009).
Os teores totais de Ni, por outro lado, não apresentam com-portamento definido em profundidade, podendo diminuir, aumentar, manter-se ou mesmo variar. Rovers et al. (1983) sugerem que tais teores acompanham as variações dos teores de Fe no perfil.
A classe textural do solo é fator importante no comportamento do Ni, pois os teores de argila se relacionam positivamente com os de Ni, porém, a relação é negativa com a fração areia. As partículas mais finas têm concentrações mais altas de Ni devido à maior área superficial específica (ASE) e maior quantidade de cargas negativas na superfície, ou seja, maior capacidade de troca catiônica (CTC) (BETTINELLI et al., 2000; SENWO e TAzISONG, 2004; CARIDAD-CANCELA; PAz-GONzÁLEz; ABREU, 2005; RAjAIE et al., 2008). Portanto, o aumento do teor de Ni com a diminuição do diâmetro das frações do solo indica que o comportamento desse nutriente é governado por processos de adsorção, principalmente com os óxidos de Fe e Mn, que apresentam alta capacidade de reter o Ni (MASSOURA et al., 2006; HARMSEN, 2007; QUANTIN et al., 2008; RAJAIE; KARI-MIAN; YASREBI, 2008; ALVES et al., 2011). Desse modo, o Ni no solo encontra-se principalmente na forma mineral, o que explica, provavelmente, sua baixa mobilidade e disponibilidade.
Entretanto, o fator que mais afeta o comportamento do Ni no solo é o pH, pois são variáveis inversamente proporcionais. Tra-balho de Siebielec e Chaney (2006) constatou redução dos teores disponíveis de Ni, ou seja, maior adsorção, com o aumento do pH, com reduções drásticas da disponibilidade do elemento em pH acima de 6,5. Em estudo semelhante, Ponizovsky et al. (2008) analisaram dez solos ácidos da União Européia, com pH na faixa de 3,6 a 6,7, e verificaram que a disponibilidade de Ni foi menor em maiores valores de pH, com deslocamento de cálcio (Ca), magnésio (Mg) e sódio (Na) para a solução em proporções semelhantes à da adsorção de Ni. Ressalta-se que o aumento dos teores disponíveis de Ni em decorrência da redução do pH ou da influência de outros fatores, como por exemplo a adubação, pode levar a uma competição direta pelos sítios de reação do solo com outros cátions, principalmente Fe, cobre (Cu) e zinco (zn), que apresentam similaridades químicas com o Ni, como valência e raio iônico hidratado (UREN, 1992).
NÍQUEL NA PLANTAA primeira constatação da essencialidade do Ni para os seres
vivos foi realizada por Dixon et al. (1975), quando demonstraram que a enzima urease apresentava dois átomos de Ni na sua com-posição estrutural. A essencialidade do Ni nas plantas superiores foi evidenciada por Eskew, Welch e Cary (1983), cultivando soja (Glycine max L. Merrill) em solução nutritiva, a qual apresentou necrose na extremidade dos folíolos devido ao acúmulo de ureia em concentrações tóxicas, consequência da baixa atividade da urease decorrente da deficiência de Ni (Figura 1). Trabalhos posteriores de Eskew, Welch e Norvell (1984) e Brown, Welch e Cary (1987), com feijão (Vigna unguiculata L.) e cevada (Hordeum vulgare L.), confirmaram a essencialidade do Ni, e ele foi inserido na lista de micronutrientes após os pesquisadores verificarem que as sementes das plantas de cevada cultivadas em solução nutritiva com ausência de Ni, após três gerações, eram inviáveis e não germinavam adequa-damente. Posteriormente, outras funções do Ni nas plantas também foram constatadas: na produção de etileno (SMITH e WOOD-BURN, 1984), na resistência das plantas a doenças (GRAHAM, WELCH, WALKER, 1985), na germinação de sementes (BROWN; WELCH; CARY, 1987), como componente estrutural da enzima hidrogenase (EVANS et al., 1987) e na conservação pós-colheita de frutos (ZHENG et al., 2006).
Figura 1. Sintomatologia de deficiência de níquel em trifólios de soja (Glycine max L. Merrill) cultivadas em condições controladas.
Fonte: Eskew et al. (1983).
O Ni é absorvido pelas plantas preferencialmente como cátion divalente (Ni2+), também sendo absorvido na forma de quelatos com compostos orgânicos e metalóforos. Ressalta-se que o mecanismo de contato preferencial desse micronutriente com o sistema radicular ainda não foi elucidado. Após entrar em contato com o sistema radicular, o Ni é absorvido, principalmente, por processo ativo, em canais não específicos, bem como por difusão passiva (YUSUF et al., 2011). A forma e o processo de absorção predominante depende dos teores de Ni no solo e, sobretudo, do efeito do pH sobre sua disponibilidade. Por exemplo, plantas de Lathyrus sativus L. absorveram maior quantidade de Ni até pH 5,0, e à medida que os valores de pH foram aumentando ocorreu redução da absorção, principalmente com valores ≥ 8,0 (PANDAA et al., 2007). Kabata-Pendias e Pendias (2011) mostraram claramente esse comportamento em estudos realizados com diferentes espécies vegetais (Figura 2). Yusuf et al. (2011) observaram que a absorção de Ni também pode ocorrer pelas folhas, quando radioisótopos de 63Ni foram aplicados em folhas de girassol (Helianthus annus L.) e

12 INFORMAÇÕES AGRONÔMICAS Nº 147 – SETEMBRO/2014
Figura 3. Distribuição do níquel nas plantas. Mais de 50% do elemento ficam retidos nas raízes e 80% do total retido na raiz ficam localizados no cilindro vascular. O níquel no caule e nas folhas encontra-se principalmente nos vacúolos, paredes celulares e tricomas epidermais associados a quelantes, tais como: ni-cotianamina (NA), histidina (His), citrato, ácidos orgânicos e proteínas com várias funções importantes, incluindo permeases, metalotioneinas (MT), metalochaperonas e proteínas YS1-like (YSLs).Abreviações na figura: parede celular (CW), cloroplastos (Chl), córtex (Co), citoplasma (Cp), endoderme (En), tricoma epitelial (Ep), epiderme abaxial (LEp), núcleo (N), parênquima paliçádico (PP), floema (P), feixes vasculares (Pi), raízes pelíferas (Rh), parênquima esponjoso (SP), epiderme adaxial (UEp), vacúolos (Va), cilindro vascular (VC), xilema (X).
Fonte: Chen, Huang e Liu (2009).
Ni2+
Quelante
Folha
Raiz
aproximadamente 36% do elemento foi translocado para os demais órgãos. Tendência similar foi observada também em folhas de aveia (Avena sativa L.), soja e tomate (Lycopersicon lycopersicum L.) pulverizadas com solução de sais de Ni.
Após a absorção, o Ni é transportado no xilema na forma iônica, em decorrência da transpiração das plantas, sendo o trans-porte facilitado com a formação de quelatos com ácidos orgânicos, tais como citrato, malato e peptídios (WHITE, 2012), bem como com aminoácidos, com destaque para a histidina (KRAMER et al., 1996), resultando em maior mobilidade do elemento no xilema (WHITE, 2012). Sem a formação de quelatos, o movimento do Ni é severamente retardado devido à elevada CTC na parede celular do xilema. O mecanismo de absorção e redistribuição de Ni nas plantas está ilustrado na Figura 3.
Em estudos iniciais, o Ni foi considerado um elemento imó-vel ou parcialmente móvel no floema, porém, Cataldo, Garland, Wil-dung (1978), estudando o comportamento do Ni em plantas de soja, constataram que o micronutriente apresentava elevada mobilidade ao verificar que, no estádio de senescência, aproximadamente 70% do Ni presente nas folhas tinham sido remobilizados para as semen-tes. Em estudo similar com plantas de trigo (Triticum aestivum L.), Page e Feller (2005) observaram que, após a absorção, o Ni diminuiu rapidamente nas partes mais velhas das raízes, redistribuindo-se para as partes recém-formadas do sistema radicular e, posteriormente, acumulou-se temporariamente nas folhas em desenvolvimento. Estes estudos sugerem que o Ni é redistribuído, ou seja, apresenta alta mobilidade no tecido vegetal, com tendência a acumular-se nos órgãos em desenvolvimento, bem como nas sementes.
Figura 2. Teores de níquel em plantas em função dos teores no solo. (a) folhas de Alyssum sp., acumuladora de Ni; (b) folhas de A. montanum, não acumuladora de Ni; (c) folhas de trigo (Triticum aestivum L.); (d) grãos de cevada (Hordeum vulgare L.); (e) raízes de cenoura (Daucus carota L.); (f) raízes de rabanete (Raphanus sativus L.). Influência do pH do solo sobre os teores de Ni em (g) grãos de aveia (Avena sativa L.). (*Os teores no solo, folhas e raízes são dados em potência de dez).
Fonte: Adaptada de Kabata-Pendias e Pendias (2011).
*
*
Plantas cultivadas em solos não contaminados apresentaram teores foliares que variam entre 0,05 e 5 mg kg-1 de Ni (BROOKS, 1980; WELCH, 1981). Teores adequados para o suprimento das plan-tas variam de 0,01 a 10 mg kg-1 (GERENDÁS et al., 1999; BROWN; WELCH; CARY, 1987), apresentando grande amplitude de variação se comparados aos demais nutrientes essenciais. Essas variações estão relacionadas, principalmente, com as diferenças na absorção, transporte e redistribuição de Ni entre espécies vegetais ou mesmo entre genótipos/cultivares (REBAFKA; SCHULz; MARSCHNER, 1990), bem como com os teores disponíveis no solo. Os teores ade-quados para germinação de sementes de cevada, e que proporcionam melhor desenvolvimento de aveia, cevada, trigo, tomate, arroz (Oryza sativa L.) e abobrinha (Cucurbita pepo L.), são de aproximadamente 100 ng g-1 MS (BROWN; WELCH; CARY, 1987; GERENDAS e SATTELMACHER, 1997). Na Tabela 2 é apresentado um resumo dos estudos sobre o teores de Ni nas plantas cultivadas.
Há espécies, entretanto, que toleram níveis excepcional-mente altos de Ni no substrato e no tecido, sem que ocorra toxidez – as hiperacumuladoras. Estima-se que um grande número de plantas, aproximadamente 317 taxons e 37 famílias, com destaque para os gêneros botânicos Alyssum e Thlaspi (Brassicaceae), pode acumular teores maiores que 1.000 mg kg-1 de Ni na matéria seca (MS) (KUPPER et al., 2001; POLLARD et al., 2002). Entre as plantas estudadas em solos ricos em Ni na região central de Goiás,

INFORMAÇÕES AGRONÔMICAS Nº 147 – SETEMBRO/2014 13
principalmente do município de Niquelândia, verificou-se cerca de 40 espécies hiperacumuladoras, apresentando teores entre 7 e 10.610 mg kg-1 de MS (REEVES et al., 2007). Teores foliares excessivos/tóxicos ocorrem geralmente entre 10 e 50 mg kg-1 de Ni para espécies não acumuladoras (WELCH, 1981).
SINTOMATOLOGIA DA DEFICIÊNCIA E TOXIDEZ DE NÍQUEL EM PLANTAS
Sintomas foliares e dossel da plantaOs sintomas peculiares da deficiência de Ni nas folhas
de pecã (Carya illinoinensis) e de algumas outras plantas são denominados vulgarmente “orelha-de-rato”, pois a ponta das folhas novas apresentam manchas escuras em formato arredondado, o que lhes confere um aspecto parecido com o da orelha de um rato (WOOD; REILLY; NYE-zEPIR, 2004a; 2004b; 2006) (Figura 4), entretanto, a deficiência em condições de cultivo a campo, até o pre-sente momento, foram relatadas apenas para duas espécies vegetais: pecã (WOOD; REILLY; NYEzEPIR, 2006) e Betula nigra (RUTER, 2005). Vários trabalhos descrevem o sintoma de deficiência de Ni como a formação da orelha de rato das folhas, mas outros sintomas morfológicos ainda são desconhecidos (WORLEY, 1979; GRAUKE; MORRIS; KOWALCzUK, 1983; WOOD e REILLY, 2007).
Wood, Reilly e Nyezepir (2004a, 2004b, 2006) iden tificaram pela primeira vez a deficiência de Ni em condições de campo nos pomares de pecã dos Estados Unidos. Em trabalhos posteriores foram verificadas res-postas visíveis das plantas ao fornecimento de Ni, quando os teores disponíveis no solo eram de 0,2 e 0,7 mg dm-3
(Figura 5). Segundo os autores, condições do solo como baixo teor disponível de Ni, excesso de fósforo (P), Cu e zn, estiagens prolongadas, pH do solo acima de 6,5, doses excessivas ou tardias de nitrogênio (N), calagem excessiva e presença de nematoides podem afetar a disponibilidade de Ni e provocar deficiência. Os sintomas em Betula nigra foram semelhantes aos verificados em plantas de pecã, nas quais os caules apresentavam entrenós mais curtos, com folhas pequenas, enrugadas, em alguns casos com coloração verde-escuro, em formato arredondado e com margens necróticas (RUTER, 2005).
Estudando o efeito do Ni sobre o metabolismo do N, em solução nutritiva, Khoshgoftarmanesh, Hosseinib e Afyunia (2011) verificaram necrose nas extremidades das folhas de alface (Lactuca sativa L.). De acordo com os autores, os sintomas são devido ao acúmulo de ureia no tecido vegetal, decorrente da deficiência de Ni.
Basicamente, os principais sintomas visuais da deficiência de Ni são:
1) Clorose O sintoma inicial da deficiência de Ni é a palidez ou amare-
lecimento das folhas durante o crescimento foliar ou crescimento precoce do dossel das plantas. Essa clorose pálida tende a ser uniforme na folha. Esse sintoma visual de clorose não é um bom parâmetro para avaliar a deficiência de Ni, pois a deficiência de outros nutrientes, como enxofre e ferro, também causa clorose foliar.
Figura 5. À direita, sintomatologia da deficiência de níquel em galhos de pecã (Carya illinoinensis); à esquerda, galhos submetidos a uma aplicação de 100 mg L-1 de sulfato de Ni (NiSO4.6H2O), via foliar, sem sintomas de deficiência.
Fonte: Wood, Reilly e Nyezepir (2004a; 2004b).
Tabela 2. Teores de níquel em plantas alimentícias cultivadas.
Planta Variação Média - - - - - (mg kg-1) - - - - -
Soja (Glycine max L. Merrill), folhas 0,08-0,62 -
Soja, grãos 0,39-2,09 -
Trigo (Triticum aestivum L.), grãos 0,17-0,67 0,34
Cevada (Hordeum vulgare L.), grãos 0,10-0,67 0,41
Aveia (Avena sativa L.), grãos 0,20-8,0 1,28
Ervilha (Pisum sativum L.) e feijão (Phaseolus vulgaris L.), sementes
0,2-0,25 -
Couve (Brassica oleracea L.), folhas 0,6-3,3 1,03
Cenoura (Daucus carota L.), raízes 0,26-0,98 -
Cebola (Allium cepa L.), bulbos 0,59-0,4 -
Tomate (Lycopersicon lycopersicum L.), frutos
0,43-0,48 -
Pepino (Cucumis sativus L.), frutos 1,3-2,0 -
Batata (Solanum tuberosum L.), tubérculos 0,29-1,0 -
Maça (Pyrus malus L.), frutos - 0,06
Laranja (Citrus sinensis L. Osbeck), frutos - 0,39
Avelã (Corylus avellana L.) e pecã (Carya illinoinensis), frutos
1,6-1,7 -
Cacau (Theobroma cacao L.), em pó 5-10 -
Fonte: Adaptada de Kabata-Pendias e Pendias (2011).
Figura 4. Sintomatologia da deficiência de níquel em pecã (Carya illinoinensis) cultivada em condições de campo na Costa do Golfo e nas Planícies Costeiras dos estados da Georgia e Flórida (Estados Unidos), denomi-nada, popularmente, orelha-de-rato.
Fonte: Wood, Reilly e Nyezepir (2006).

14 INFORMAÇÕES AGRONÔMICAS Nº 147 – SETEMBRO/2014
2) Redução do tamanho e formato alterado da folha O indicador visual mais claro e apurado para detectar a
deficiência de Ni nas folhas são a redução do tamanho foliar e o embotamento das pontas das folhas ou folíolos (Figura 4). A intensidade do embotamento da ponta das folhas ou folíolos varia de acordo com a intensidade da severidade da deficiência de Ni.
O ápice foliar apresenta característica morfológica obtusa. Em folhas de pecã, a região apical de folhas normais com formato agudo torna-se obtusa (Figura 4). Esse embotamento abrupto da região apical de folhas ou folíolos (geralmente folhas fisiologi-camente mais velhas) em certos ramos pode ocorrer de maneira uniforme no dossel das plantas. Em casos claros de deficiência severa de Ni, em condições de campo, o aparecimento da orelha- de-rato ocorre mais frequentemente no topo do dossel. No campo experimental do Departamento de Agronomia, nos Estados Uni-dos, teores foliares de pecã abaixo de 0,85 mg kg-1 promoveram disfunção morfológica e acarretaram na formação da orelha-de-rato (NYCzEPIR; WOOD; REILLY, 2006).
A redução da superfície foliar diminui proporcionalmente com o aumento da deficiência foliar de Ni. Essa redução varia entre 10% e 75% em árvores adultas. Essas folhas deficientes tendem a aumentar no topo do dossel das plantas (WOOD e REILLY, 2007).
3) Região verde-escuro nas pontas foliaresFolhas deficientes em Ni apresentam região verde-escuro
na parte apical da folha, como demonstrado na Figura 4. Essa característica é muito comum aparecer após poucas semanas da emissão das folhas ou folíolos, o que torna muito claro o sintoma visual característico da deficiência de Ni.
4) Necrose apical foliarA deficiência severa de Ni provoca necrose na região apical
das folhas, como ilustrado na Figura 4. A região necrosada da folha tende a aumentar com a idade e a proporção da deficiência. Aparen-temente, a região das células mortas é consequência do acúmulo de ureia (ácidos láctico e oxálicos), devido à baixa atividade da enzima urease. A deficiência de Ni é mais pronunciada na região meriste-mática ou nas regiões de crescimento da folha. Essas características são excelentes indicadoras da deficiência de Ni nas plantas.
5) Curvatura e enrugamento da região apical foliarA região de expansão celular nas margens foliares é redu-
zida ao ponto de curvar e aparecer o enrugamento da região apical (Figura 4). Folhas afetadas são mais finas e flexíveis e tendem ser quebradiças. Esse sintoma também é um bom indicador da defi-ciência de Ni em plantas lenhosas.
6) Ausência do desenvolvimento laminarEm folhas ou folíolos severamente deficientes, o desenvol-
vimento laminar é completamente interrompido e apresenta arranjo vascular reduzido e desprovido de lâminas internervais. Em casos extremos, a área foliar pode ser extremamente reduzida ou nula e isso ocorre principalmente nos primeiros anos de implantação da cultura. Essa redução foliar ocorre concomitantemente com o alon-gamento anormal e afinamento das gemas. Ambas as características são bons indicadores da deficiência de Ni.
A HISTÓRIA DA ORELHA-DE-RATO E A RELAÇÃO COM A DEFICIÊNCIA DE NÍQUEL EM PLANTAS
O primeiro relato da orelha-de-rato foi feito por Marz (1918). Inicialmente, foi encontrada em árvores de quintais e viveiros na Flórida, Mississipi e na Georgia. Na época, a desordem morfoló-gica foi atribuída a injúrias do frio e alguns anos depois atribuída a
doenças provocadas por patógenos (DEMARE, 1926). Na década de 1930, a orelha-de-rato foi observada em pomares de pecã e Gammon e Sharpe (1956) suspeitavam que o problema era causado por deficiência de Mn, no entanto, aplicações via solo ou foliar de elemento não erradicaram a orelha-de-rato.
Durante a formação de pomares de pecã na Costa do Golfo na Georgia, a severidade da orelha-de-rato era tão grande que muitas plantas morreram (WOOD; REILLY; NYEzEPIR, 2004a). Muitas plantas foram replantadas e mesmo assim morreram, o que provocou um problema econômico na época. A sintomatologia descrita por Wood, Reilly e Nyezepir (2004a) indicava a deficiência de algum micronutriente ou o desbalanceamento entre os micronutrientes como a causa principal da orelha-de-rato. Aplicações de Cu durante o transplantio corrigiam os sintomas de orelha-de-rato de um ano agrícola para o outro, mas os mesmos sintomas apareciam no ano seguinte. Aplicações foliares de Cu não tinha nenhum efeito sobre os sintomas da orelha-de-rato (WOOD; REILLY; NYEzEPIR, 2004b). Wood, Reilly e Nyezepir (2004a) também observaram uma forte correlação entre Cu e zn na incidência da severidade da orelha-de-rato. Cátions divalentes, como zn, Cu e Ni, presentes na solução do solo, usam o mesmo canal de íons para entrada no sistema radicular de plantas (KOCHIAN, 1991), o que acarretou a ideia da possibilidade da orelha-de-rato estar relacionada ao Ni. Finalmente, em 2004, a equipe do Dr. Bruce Wood relatou que a principal causa da orelha-de-rato em pecã era a deficiência de Ni (WOOD; REILLY; NYEzEPIR, 2004c). Pulverizações foliares com 100 mg L-1 (na forma de NiSO4.6H2O) foram suficientes para erradicar a orelha-de-rato (Figura 5). Após a aplicação de Ni, as folhas de pecã voltaram a se desenvolver normalmente. De acordo com os autores, a aplicação foliar de Ni não apresentou fitotoxici-dade visual nas folhas. O Ni absorvido pelas folhas foi acumulado em regiões de reserva da planta e posteriormente translocado para regiões dormentes, brotos e meristemas apicais. A quantidade de Ni translocado na planta foi suficiente para corrigir os sintomas de deficiência e eliminar a orelha-de-rato.
Wood, Reilly e Nyezepir (2004a) submeteram uma planta de pecã com sintomas de deficiência de Ni nos galhos a uma aplicação de 100 mg L-1 de sulfato de Ni (NiSO4.6H2O) via foliar, em condições de campo. Nas folhas que receberam pulverização com solução de Ni, o teor do elemento era de aproximadamente 7 mg kg-1, compa-rado a 0,5 mg kg-1 nas folhas controle, as quais exibiam o sintoma de deficiência de Ni ou orelha-de-rato (Tabela 3). Em outras árvores do pomar que não apresentavam orelha-de-rato o teor foliar médio de Ni era de aproximadamente 4 mg kg-1. Aparentemente, houve o efeito da época de aplicação do Ni nas folhas de pecã. O teor foliar de Ni nas folhas tratadas durante a primavera (julho) era quase quatro vezes maior (26 mg kg-1) em relação ao das plantas que receberam pulverização de Ni no outono.
TOXIDEZ DE NÍQUEL: SINTOMAS FOLIARES E NOS GRÃOSSintomas de deficiência de Ni dificilmente são detectados
em plantas nas condições de campo, porém, o efeito fitotóxico do Ni é conhecido já há muito tempo. Uren (1992) cita trabalhos nos quais são relatados que teores de 2 mg kg-1 de Ni foram suficientes para inibir o crescimento de raízes. Também relatam que foram observados sintomas de toxicidade em trigo quando o teor de Ni no solo era de 8 mg kg-1, enquanto a aveia não apresentou qual-quer sintoma quando o teor de solo era de 147 mg kg-1. Com base nessas observações, conclui-se que a toxicidade de Ni é função da espécie vegetal, sendo esta toxicidade normalmente associada à deficiência de cobalto (Co) (McGRATH e SMITH, 1990). Poulik

INFORMAÇÕES AGRONÔMICAS Nº 147 – SETEMBRO/2014 15
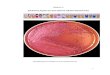
Figura 7. Sintomatologia de toxidez de níquel em soja (Glycine max L. Merrill) cultivar BRS 360 RR cultivada em um Latossolo Ver-melho eutroférrico típico submetido à aplicação de 5,0 mg dm-3 de Ni, sob condições controladas.
Fonte: Rodak (2014).
(1997) observou que plantas de aveia com teor de 168 mg kg-1 de Ni apresentaram sintomas de toxicidade e morreram. Por outro lado, o teor de Ni no solo menor do que 56 mg kg-1 aumentou a produção de espigas. O sintoma visual da toxicidade de Ni pode ser confundido com o da deficiência de outros elementos essenciais (PAIVA et al., 2003), tal como a clorose causada pela deficiência de Mn ou Fe.
De maneira geral, a sintomatologia da toxidez inclui clorose, devido à menor absorção de Fe, crescimento reduzido das raízes e da parte aérea (Figura 6), em casos mais severos, deformação de várias partes da planta e manchas peculiares nas folhas (CHEN; HUANG; LIU, 2009). Entretanto, ressalta-se que em estudo recente de Rodak (2014), plantas de soja submetidas a 5,0 mg dm-3 de Ni, em condições controladas, apresentaram sintomas de toxidez nas folhas, com teores em torno de 2 mg kg-1 de Ni a partir do estádio de desenvolvimento R5 (Figura 7). No estádio de desenvolvimento R1 observou-se comportamento semelhante ao verificado para a massa seca total, no qual as plantas de soja cultivadas em solo muito argiloso, na dose de 5 mg dm-3 de Ni, apresentaram menor desenvolvimento.
Sintomas de toxidez foram visíveis nas folhas a partir do estádio de desenvolvimento R5 (Figura 7), fato não observado nas plantas cultivadas no solo franco-arenoso (RODAK, 2014). Esses sintomas de toxidez de Ni são semelhantes aos relatados na literatura: as plantas apresentam amarelecimento entre as nervuras, semelhante à deficiência de Mn, crescimento reduzido das raízes e da parte aérea, podendo, em casos mais severos, apresentar deformação de várias partes da planta, e manchas peculiares nas folhas (MISHRA e KAR, 1974). Os estudos de Rodak (2014) ainda revelaram que os teores de Ni nos grãos de soja apresentaram comportamento inverso ao observado nas folhas, ou seja, maiores
Tabela 3. Teor foliar de níquel em plantas de pecan que apresentavam severo sintoma de orelha-de-rato e plantas com a deficiência corrigida após aplicação foliar de solução de níquel.
Tratamento1 Severidade da orelha-de-rato2
Teor foliar de Ni3
- - - - - - - - - (mg kg-1) - - - - - - - - - Plantas normais - controle 1,0 b4 4,0 bOrelha-de-rato - controle 7,4 a 0,5 cOrelha-de-rato + Ni (outono) 1,0 b 7,0 aPlantas normais - controle 1,0 b 5,0 bOrelha-de-rato - controle 7,7 a 0,4 cOrelha-de-rato - controle + Ni (primavera) 1,0 b 26,0 a
1 Tratado com sal de sulfato de Ni utilizando ureia e surfactante não iônicos. Os tratamentos foram aplicados nas folhas em outubro (outono) e em abril, aproximadamente 10 dias após a abertura dos brotos (primavera).
2 A severidade da orelha-de-rato foi baseada na seguinte escala: 1 = sem sintomas; 2 = entre 1% e 25% de folhas ou folíolos que apresentaram embotamento; 3 = 26% a 50% de folhas ou folíolos que apresentaram embotamento; 4 = mais de 50% de folhas ou folíolos que apresentaram embotamento; 5 = engruvinhamento da parte embotoada da região apical foliar; 6 = necrose das pontas foliares; 7 = região verde-escuro próximo à região apical foliar; 8 = atrofiamento da parte aérea da planta; 9 = apare-cimento de múltiplas novas partes aéreas (envassouramento); 10 = morte do dossel. Nota-se que a severidade da orelha-de-rato + tratamento de Ni na primavera foi contada após os tratamento das folhas.
3 O teor foliar de Ni foi determinado na massa seca de plantas coletadas em julho, após o tratamento de Ni entre outubro e abril.
4 Médias seguidas por letras diferentes são estatisticamente significativas a 5% de probabilidade pelo teste de Tukey-Kramer.
Fonte: Wood, Reilly, Nyezepir (2004a).
teores de Ni foram verificados nos grãos de soja cultivada no solo muito argiloso, que variaram de 0,42 a 13,56 mg kg-1, enquanto na soja cultivada no solo franco-arenoso a variação foi de 0,39 a 4,28 mg kg-1. A maior translocação de Ni observada nas plantas cultivadas no solo muito argiloso se deve ao maior crescimento das plantas nesse solo, resultando em maiores valores de acúmulo de Ni: de 3 a 20 μg/planta, enquanto nas plantas cultivadas no solo franco-arenoso variaram de 1,3 a 10 μg/planta. Com base nesses resultados, sugere-se que ocorra translocação do Ni para os grãos, corroborando com os dados de Cataldo, Garland e Wildung (1978), os quais constataram que, a partir do início da senescência, as plantas de soja translocam aproximadamente 70% do Ni para os grãos. Rodak (2014) ressalta que o teor médio de 13,56 mg dm-3 de Ni nos grãos das plantas cultivadas no solo muito argiloso, com fornecimento de 5,0 mg dm-3 de Ni, excede os valores comumente relatados na literatura como adequados, podendo ser a causa dos sintomas de toxidez verificados nos grãos de soja, ou seja, menor desenvolvimento e enrugamento, como ilustrado na Figura 8.
Figura 6. Plantas de soja no início do florescimento (R1), cultivadas em solo (a) franco arenoso e (b) muito argiloso, submetidos a doses de Ni.
Fonte: Rodak (2014).
(a)
(b) 0 0,2 0,4 0,8 1,0 5,0 mg dm-3 Ni
0 0,2 0,4 0,8 1,0 5,0 mg dm-3 Ni

16 INFORMAÇÕES AGRONÔMICAS Nº 147 – SETEMBRO/2014
Ni via foliar e solo, respectivamente. Entretanto, aspectos fisioló-gicos das plantas foram potencializados com a aplicação da dose em torno de 0,5 mg dm-3 de Ni no solo. A princípio, teores foliares de 0,3 a 0,8 mg kg-1, teores disponíveis de 0,2 a 0,6 mg dm-3 em DTPA e 0,4 a 0,8 mg dm-3 em Mehlich-1 podem ser tomados como sugestão aproximada de níveis de referência para Ni em solos e folhas-diagnóstico de soja (RODAK, 2014).
A falta de resposta dos componentes de produtividade à adu-bação com Ni em condições de campo, mesmo em solos com teores disponíveis iniciais baixos, provavelmente se deve ao teor inicial de Ni na semente. Brown, Welch e Cary (1987), cultivando cevada em solução nutritiva, verificaram que apenas o conteúdo de Ni da semente foi suficiente para suprir até três gerações precedentes sem que houvesse deficiência do mesmo. Em trabalho de Rodak (2014), os teores na semente supriram de 0,75% a 18,88% da quantidade total de Ni acumulado em plantas de soja, sendo esse um dos principais fatores que justificam a ausência de resposta em produtividade.
Em trabalhos recentes, Kutman, Kutman e Kakmak (2012, 2014) estudaram as reservas de Ni em sementes de soja, juntamente com o fornecimento foliar desse micronutriente e de fertilizantes nitrogenados em solução nutritiva. Constataram respostas à adu-bação com Ni e ureia quando os teores iniciais nas sementes eram baixos e médios, principalmente no metabolismo do N, porém, sem acréscimos de produtividade. Foram verificados redução dos danos foliares decorrentes da pulverização com uréia bem como aumento da biomassa aérea e do conteúdo de clorofila em função dos teores iniciais de Ni nas sementes e de seu fornecimento via foliar (Figura 9, Figura 10 e Figura 11), evidenciando, portanto, que as reservas na semente exercem influência direta sobre a resposta das plantas à adubação com Ni.
Figura 9. Plantas de soja (Glycine max cv. Nova), aos 29 dias, cultivadas a partir de sementes com baixo, médio e alto teor de níquel e submetidas à aplicação foliar de ureia e de 0 e 0,2 μM de Ni (-Ni e +Ni, respectivamente), em solução nutritiva.
Fonte: Kutman, Kutman e Kakmak (2012).
Sem ureia foliar Com ureia foliar
São escassas as informações referentes aos teores de Ni nas sementes de soja, logo, com o intuito de compreender o com-portamento dos teores nas sementes, Rodak (2014) efetuou um breve estudo exploratório no qual os principais genótipos de soja atualmente cultivados no Brasil foram avaliados quanto ao teor de Ni. Notou que os genótipos de soja diferiram grandemente quanto aos teores do elemento na semente, variando de 0,5 a 2,09 mg kg-1. O local de cultivo, em razão do teor disponível no solo, influencia grandemente os teores nas sementes; por exemplo, o cultivar BRS 360 RR apresentou maiores teores de Ni nas sementes quando os teores disponíveis no solo eram mais elevados.
LEIS BRASILEIRAS E CRITÉRIOS DE ADUBAÇÃO COM NÍQUEL
O Ni foi inserido na legislação brasileira de fertilizantes na Instrução Normativa N° 05, de 23 de fevereiro de 2007 (MAPA, 2014) e os estudos a respeito de sua exigência pelas culturas em condições de campo e forma de utilização na agricultura se encontram em fase inicial. A fonte de Ni mais empregada e estudada na agricultura é o sulfato de níquel (NiSO4.6H2O), entretanto, insumos como os termofosfatos não são considerados fontes, embora contenham quan-tidade significativa desse micronutriente, com teores médios de até 330 mg kg-1 (GABE, 1998). Outras fontes, além das mencionadas, vêm sendo comercializadas, como quelados e fertilizantes foliares, entretanto, ainda são escassos os estudos científicos sobre as mesmas.
Excluindo-se os trabalhos em soluções nutritivas, os critérios de adubação foram definidos, basicamente, para as culturas de pecã e soja. A aplicação foliar de 100 mg L-1 de sulfato de Ni (NiSO4.6H2O) foi eficiente para minimizar os sintomas de deficiência em pecã (WOOD; REILLY; NYEzEPIR, 2006). Para soja, a adição de até 0,02 mg dm-3 de Ni proporcionou incrementos de produtividade (BERTRAND, 1973). Martins (comunicação pessoal), citado por Moraes et al. (2010), cultivando soja a campo, em condições brasi-leiras, verificou que o fornecimento de 50 g ha-1 de Ni proporcionou aumento de até 6,2 sc ha-1, aventando que o Ni estaria atuando no controle de doenças fúngicas e, assim, aumentando a produtividade da soja de forma indireta. Entretanto, ressalta-se que, para soja, trabalhos de Alovisi et al. (2011) e Rodak (2014) não verificaram incremento nos componentes de produtividade em função do fornecimento de
Figura 8. Aspecto visual de grãos de soja cultivada em solo (a) franco- arenoso e (b) muito argiloso, submetidos a doses de níquel.
Fonte: Rodak (2014).
Alto
teor
de
M
édio
teor
de
B
aixo
teor
de
Ni n
a se
men
te
N
i na
sem
ente
Ni n
a se
men
te
0 0,2 0,4 0,8 1,0 5,0 mg dm-3 Ni
0 0,2 0,4 0,8 1,0 5,0 mg dm-3 Ni
(a)
(b)

INFORMAÇÕES AGRONÔMICAS Nº 147 – SETEMBRO/2014 17
As primeiras observações sobre a possível necessidade de Ni no cafeeiro durante o ciclo de desenvolvimento dos frutos foram descritas por Reis et al. (2009). Mesmo sem aplicar Ni no solo, os teores de Ni aumentaram gradativamente nas folhas e nos frutos. Ressalta-se que a área experimental foi adubada com termofosfatos, os quais são fontes de Ni para as plantas. Reis et al. (2011) verificaram que os teores de Ni nas folhas e nos grãos diminuem em função da adubação nitrogenada na forma de ureia. Isso provavelmente ocorre devido ao fator de diluição, consequência do crescimento do dossel e dos frutos na presença de N. Contudo, os autores afirmam que os teores adequados de Ni para o cafeeiro são de aproximadamente 2 a 3 mg kg-1 em frutos verdes e de 5 a 10 mg kg-1 em frutos cereja. O aumento crescente do teor de Ni no fruto em função do desen-volvimento da planta ainda não está esclarecido. Mais pesquisas devem ser conduzidas para explicar o fenômeno. Provavelmente, o cafeeiro adubado com ureia como fonte nitrogenada pode neces-sitar de uma pulverização com Ni durante o desenvolvimento dos frutos para formar sementes de boa qualidade, como já observado em algumas culturas e descrito por Brown, Welch e Cary (1987).
O PAPEL DO NÍQUEL NA FISIOLOGIA E NA BIOQUÍMICA DAS PLANTAS
Brown, Welch e Cary (1987) consideram o Ni um micronutriente essencial em certas espécies vegetais, especialmente quando cultivadas com ureia, pois é parte da enzima urease (DIXON et al., 1975), e sua deficiência leva à redução da atividade enzimática em tecidos de soja [Glycine max (L.) Merr.], arroz (Oryza sativa L.) e fumo (Nicotiana tabacum L.), resultando em acúmulo excessivo de ureia, tornando-a fitotóxica (POLACCO, 1977; POLACCO e HOLLANDO, 1993).
A urease (EC 3.5.1.5; urea amidohidroxilase) é uma metaloenzima que catalisa a hidrólise da ureia para formar NH3 e CO2 (DIXON et al., 1975), como ilus-trado na Figura 12. é encontrada em vários organismos, incluindo plantas, fungos e bactérias (HIRAYAMA et al., 2002). A função principal da urease é permitir aos organismos o uso da ureia gerada externa ou internamente como fonte de N (MOBLEY e HAUSINGER, 1989; MOBLEY; ISLAND; HAUSINGER, 1995; ANDREWS; BLAKELEY; zERNER, 1984; TEzOTTO et al., 2012).
Nas plantas, uma quantidade significativa do fluxo do N ocorre através da ureia (47% N), o qual é reciclado somente pela ação da urease (POLLACCO e HOLLAND, 1993). A ureia pode também ser gerada pelo catabolismo de ureídeos (alantoato e alantoína). Tem sido demonstrado que o ureídeo-glicolato, um produto da degradação do alantoato, é um precursor da ureia (TODD e POLLACCO, 2004; MUÑOz et al., 2006).
Bloqueios genéticos e químicos da atividade da urease nas folhas causam um tipo de necrose foliar, asso-ciado ao acúmulo de ureia, o que promove alterações no ciclo da ornitina (KROGMEIER, 1989; POLLACCO e HOLLAND, 1993). Nas plantas, a urease tem função na assimilação da ureia derivada de ureídeos ou arginina (POLLACCO e HOLLAND, 1993). A arginina é um dos aminoácidos livres predominantes no caule de plantas perenes, especialmente durante o inverno, e é considerada o principal composto de armazenamento de N. De fato, a arginina se acumula na casca durante o outono e diminui gradualmente durante a primavera seguinte (SUzUKI,
Figura 11. Efeito dos teores de níquel na semente (alto e baixo) em (a) folhas primárias e (b) trifólios de soja (Glycine max cv. Nova) aos 22 dias, em função do fornecimento de 2×10-7 M de Ni e de 2×10-3 M de N como ureia e nitrato, em solução nutritiva.
Fonte: Kutman, Kutman e Kakmak (2014).
Figura 10. Sintomatologia de toxidez nas folhas jovens de soja (Glycine max cv. Nova) aos 28 dias após a aplicação foliar de ureia. As plantas foram cultivadas a partir de sementes com baixo, médio e alto teor de Ni e submetidas à aplicação foliar de 0 e 0,2 μM de níquel, -Ni e +Ni, respectivamente. (†1 teor médio de Ni nas folhas; †2 n.d. = abaixo do limite de detecção, < 1 mg kg-1).
Fonte: Kutman, Kutman e Kakmak (2012).
+ N
i
- N
i
Baixo teor de Ni Médio teor de Ni Alto teor de Ni na semente na semente na semente

18 INFORMAÇÕES AGRONÔMICAS Nº 147 – SETEMBRO/2014
1984). No entanto, a relevância fisiológica da urease bem como a variação sazonal de aminoácidos do ciclo da ornitina não estão esclarecidas.
Quando a atividade da urease é baixa, devido ao inadequado suprimento de Ni, a ureia pode se acumular em níveis conside-ráveis, especialmente em plantas tratadas com ureia, o que pode eventualmente levar a alterações nos compostos intermediários do ciclo da ornitina (GERENDÁS e SATTELMACHER, 1997). Os autores verificaram, ainda, que o Ni aumentou significativa-mente a atividade da urease em diversas espécies. Além disso, a deficiência de Ni afetou o metabolismo dos aminoácidos em feijão- caupi (WALKER et al., 1985), reduziu a atividade da urease, induziu a deficiência no metabolismo do N e afetou os aminoácidos (glicina, asparagina, arginina, ornitina e citrulina) em diversas espécies (GERENDÁS e SATTELMACHER, 1997). Folhas deficientes em Ni exibem relativamente alto nível de xantina, ácido alantóico e ureídeo-glicolato – três intermediários da cadeia do catabolismo de ureídeos. O acúmulo desses intermediários indica reduzida ativi-dade enzimática nas três últimas fases do catabolismo dos ureídeos (BAI; REILLY; WOOD, 2006).
A principal rota geradora de ureia nas plantas é a reação da arginase (Figura 12). Quando a atividade da urease é reduzida por um suprimento inadequado de Ni, uma perturbação no metabolismo do N pode ser antecipada sempre que for adicionada ureia externa ou ocorrer a liberação de ureia pela reação da arginase (BAI; REILLY; WOOD, 2006). Como a demanda de N pelas plantas é altamente dependente da hidrólise da ureia, a deficiência de Ni resulta em paralisação do metabo-lismo do N, o que provoca acúmulo de ureia e diminuição na síntese de aminoácido (GERENDÁS e SATTELMACHER, 1997; 1999).
A essencialidade do Ni somente ganhou aspecto prático após a constatação, e posterior correção, da deficiência de Ni em poma-res de pecã nos Estados Unidos (WOOD; REILLY; NYEzEPIR, 2004a, 2004b, 2004c). A literatura recente sobre nutrição mineral de plantas já trata o Ni como micronutriente (MARSCHNER, 2012). Dentre outros aspectos positivos do Ni, destacam-se a sua influência no complexo enzimático hidrogenase, que aumenta a eficiência da fixação do N por leguminosas (KLUCAS, 1983), e a sua participação na síntese de fitoalexinas, aumentando a resistência das plantas às doenças (WALKER et al., 1985).
O Ni possui alta mobilidade na planta, encontrando-se em todos os tecidos e, princi-palmente, nas sementes (MALAN e FARRANT, 1998). O acúmulo ocorre de modo diferencial entre os tecidos e ao longo do ciclo vital da planta, sendo maior nos grãos, nas folhas e nas partes jovens. O Ni é capaz de modificar a absorção e o transporte do nitrato, alterando a atividade das enzimas nitrato redutase e glutamina sintetase, essenciais para a síntese de aminoácidos (PALACIOS e MATAIX, 1999).
FLORESCIMENTO DO CAFEEIRO E A IMPORTÂNCIA DO NÍQUEL NESSE PROCESSO
O processo de florescimento do cafeeiro compreende várias etapas, como: indução floral; iniciação floral; diferenciação, crescimento e desen-volvimento das peças florais do botão; dormência do botão floral e florada (abertura da flor ou antese). Cada uma dessas fases é afetada por fatores exóge-
nos e endógenos particulares que determinam diferentes padrões de crescimento e desenvolvimento dos órgãos florais, de acordo com a variedade/cultivar e as condições ambientais predominantes (ALVES e GUIMARãES, 2010).
A maior parte das pesquisas sobre floração do cafeeiro restringe-se a eventos que ocorreram na pré-florada ou na flo-rada. Entretanto, informações reunidas nas áreas de fisiologia do florescimento ainda são escassas. Grande parte dos estudos nessa fase da floração se restringe aos efeitos negativos da seca e da alta temperatura. Em recentes estudos envolvendo o efeito do déficit hídrico sobre o abortamento de flores e frutos em café arábica foi constatado um aumento de 50% para 83% na queda total de flores/frutos (ALVES e GUIMARãES, 2010). Esse abortamento foi elevado para 93% em rosetas com flores anormais (estrelinhas), em resposta à temperatura elevada.
Uma vez que em condições de campo normalmente ocorrem longos veranicos na fase de florescimento do cafeeiro, alguns pesquisadores, além do uso de irrigação, estudam meios para evitar o abortamento das flores, principalmente pela pulverização de caldas contendo Ca e boro (B). Considerando que os resultados encontrados até agora são contraditórios, novas alternativas têm sido pesquisadas, entre elas o uso de Ni como forma de garantir o pegamento das flores.
Malavolta et al. (2006) observaram que metade do total do Ni presente nos citros estava nas flores e destacaram que possi-velmente esta alta demanda do micronutriente pelas flores se deve ao seu papel de ativador da urease (DIXON et al., 1975) que, ao agir sobre a ureia, libera amônia (NH3) e dióxido de carbono (CO2). Há algum tempo se utiliza a pulverização de citros com ureia como meio de aumentar o teor de NH3 nas folhas. Lovatt et al. (1988) propuseram que o aumento dos níveis de NH3 nas folhas durante o estádio de indução floral leva a incrementos na iniciação floral via síntese de várias poliaminas, como putrescina, espermidina e espermina (Figura 12). Desse modo, Malavolta et al. (2006) supõem que o alto conteúdo de Ni nas flores poderia aumentar a atividade de urease e, portanto, a produção de amônia endógena. Entretanto, não se sabe, ainda, se a aplicação de Ni, via solo ou folha, em cafeeiros bem nutridos com N aumenta o florescimento e, proporcionalmente, o pegamento das flores.
Figura 12. Origem e destino da ureia na planta.

INFORMAÇÕES AGRONÔMICAS Nº 147 – SETEMBRO/2014 19
RESPOSTAS ANTIOXIDATIVAS À APLICAÇÃO DE NÍQUEL
Estudos sobre os efeitos do Ni no metabolismo das plantas demonstraram que o metal pode induzir à produção de espécies rea-tivas de oxigênio - EROs (O2•
–) (FOYER et al., 1997). As espécies reativas de oxigênio demonstraram induzir a peroxidação lipídica de membranas celulares, e o nível de peroxidação lipídica, medido pela quantidade de substâncias reativas ao ácido tiobarbitúrico (TBARS), tem sido utilizado como um índice do efeito tóxico do estresse oxidativo (DEUNER et al., 2011).
As espécies reativas de oxigênio mostraram ser produzidas como uma resposta geral a distintas condições abióticas estressantes (ANZA; RIGA; GARBISU, 2005; FIDALGO et al., 2004; ALVES et al., 2012). A destruição eficiente das espécies reativas de oxigênio requer a ação de várias enzimas antioxidantes, como superóxido dismutase, catalase, ascorbato peroxidase, glutationa redutase e guaiacol peroxidase. As superóxido dismutases são consideradas a primeira linha de defesa contra as espécies reativas de oxigênio, sendo responsáveis pela dismutação do O2•
–, gerando H2O2 e O2. A catalase, a ascorbato peroxidase e a guaiacol peroxidase são enzimas que catalisam a conversão do H2O2 em água e O2 (GRATãO et al., 2005). A glutationa redutase catalisa a redução dependente de nicotiamida- adenina-dinucleótido-fosfato (NADPH) da glutationa dissulfeto (GSSG), forma oxidada, para glutationa (GSH), a forma reduzida
(GRATãO et al., 2005). A ascorbato peroxidase, a glutationa redu-tase e a GSH são componentes importantes do ciclo ascorbato- glutationa, responsável pela remoção do H2O2 em diferentes com-partimentos celulares (FOYER e NOCTOR, 2005, GRATãO et al., 2005). A GSH é, também, o substrato para a biossíntese de fitoquelatinas, que estão envolvidas na desintoxicação de metais pesados (INOUHE, 2005). Vários resultados de pesquisa referentes ao efeito no Ni na atividade enzimática da rota fotossintética, na rota do N e sobre as enzimas antioxidativas estão compilados na Tabela 4. Dependendo da concentração de Ni no solo ou na solução nutritiva, a atividade enzimática de diferentes culturas pode ser aumentada ou diminuída de acordo com a fisiologia de cada planta.
Além disso, o Ni induz a produção de fitoquelatinas, mas a ligação de metais com fitoquelatinas foi apenas demonstrada para Cu, Pb e Cd (KUKKOLA, 2000). Além disso, plantas naturalmente tole-rantes a metais não elevam a produção de fitoquelatinas como parte de sua defesa contra o Ni (FREEMAN et al., 2004). Entretanto, o Ni pode formar complexo estável com a GSH (RAO e SRESTY, 2000).
Apesar das informações da relação entre metais pesados e estresse oxidativo disponíveis em anos recentes, continua difícil estabelecer uma conclusão geral sobre as concentrações críticas de metais tóxicos nos solos (ROMERO-PUERTAS et al., 2004). O O2 molecular é relativamente não reativo em seu estado natural, mas
Tabela 4. Efeito do Ni nas atividades enzimáticas.
Enzima Processo [Ni], mM AT1 Espécie Referência
Rubisco Fixação de CO2 0,5 - 1,0 ↓ Cajanus cajan Sheoran et al. (1990)Gliceraldeído 3-fosfato dehidrogenase
Ciclo de Calvin 0,5 - 1,0 ↓ Cajanus cajan Sheoran et al. (1990)
3-fosfoglicerol quinase Ciclo de Calvin 0,5 - 1,0 ↓ Cajanuscajan Sheoran et al. (1990)Aldolase Ciclo de Calvin 0,5 - 1,0 ↓ Cajanus cajan Sheoran et al. (1990)Frutose 1,6-bisfosfatase Ciclo de Calvin 0,5 - 1,0 ↓ Cajanus cajan Sheoran et al. (1990)NADP e NAD dependente de fosfogliceraldeído dehidrogenase
Ciclo de Calvin 0,5 - 1,0 ↓ Cajanus cajan Sheoran et al. (1990)
Redutase do nitrato Redução do nitrato 1 ↓ Beta vulgaris Kevresan et al. (1998)Redutase do nitrato Redução do nitrato < 0,4 (mg dm-3) ↑ Glycine max Rodak (2014)Redutase do nitrato Redução do nitrato > 0,4 (mg dm-3) ↓ Glycine max Rodak (2014)Redutase do nitrato Redução do nitrato 0,1 ↓ Triticum aestivum Gajewska e Sklodowska (2009)H+-ATPase Transporte de íons 0,5 ↑ Oryza sativa ROS et al. (1990)Glutamina sintetase Síntese de glutamina 1 ↓ Beta vulgaris Kevresan et al. (1998)Glutamina sintetase Síntese de glutamina 0,1 ↓ Triticum aestivum Gajewska e Sklodowska (2009)Fd-GOGAT Síntese de glutamato 0,1 ↓ Triticum aestivum Gajewska e Sklodowska (2009)NAD-GOGAT Síntese de glutamato > 0,1 ↑ Triticum aestivum Gajewska e Sklodowska (2009)Alanina aminotransferase Transformação de alanina
em piruvato0,2 ↓ Glycine max El-Shintinawy e El-Ansary
(2000)IAA oxidase Oxidação do IAA < 0,05 ↑ Oryza sativa DAS et al. (1978)IAA oxidase Oxidação do IAA > 0,05 ↓ Oryza sativa DAS et al. (1978)Glutationa redutase Redução da glutationa 0,01-1 ↑ Alyssum maritimum Schicker e Caspi (1999)Ascorbato oxidase Oxidação do ascorbato < 0,05 ↑ Oryza sativa DAS et al. (1978)Ascorbato oxidase Oxidação do ascorbato > 0,05 ↓ Oryza sativa DAS et al. (1978)Superóxido dismutase Deativação do O2.
– 0,01 ↓ Alyssum maritimum Schicker e Caspi (1999)Superóxido dismutase Deativação do O2.
– 0,1 ↑ Alyssum maritimum Schicker e Caspi (1999)Catalase Degradação do H2O2 < 0,05 ↑ Oryza sativa DAS et al. (1978)Catalase Degradação do H2O2 > 0,05 ↓ Oryza sativa DAS et al. (1978)Peroxidase Oxidação de polifenóis 1-40 ↑ Triticum aestivum Pandolfini et al. (1992)
1 AT = atividade enzimática, ↓ = diminui, ↑ = aumenta.

20 INFORMAÇÕES AGRONÔMICAS Nº 147 – SETEMBRO/2014
a produção de espécies reativas de oxigênio, como o superóxido, peróxido de hidrogênio, radical hidroxil e o oxigênio “singlet”, é uma consequência inevitável do metabolismo aeróbico. Sob con-dições normais de crescimento, a produção de espécies reativas de oxigênio nas células ocorre em baixa taxa, entretanto, fatores ambientais adversos que perturbam a homeostase celular aumentam a produção de espécies reativas de oxigênio, levando ao estresse oxidativo (GRATãO et al., 2005).
APLICAÇÃO DO NÍQUEL E RELAÇÕES COM DOENÇAS E PATÓGENOS
O Ni tem efeito direto ou indireto sobre microrganismos que promovem doenças em plantas, incluindo viroses (PENNAzIO e ROGGERO, 1988), bactérias (WANG; zENG; KANG, 2000) e fungos (DUBEY e DWINVEDI, 1988; SINGH; SHUKLA; DWIN-VEDI, 1992). Sais de Ni são especialmente efetivos nas doenças causadas por fungos (KISHORE; PANDE; RAO, 2001; SUGHA e SINGH, 1990) e nematoides (KAHN e SALAM, 1990). A inibição direta de micróbios por pulverizações com Ni ocorre na concentra-ção aproximada de 200 mg L-1, porém, essa concentração pode ser tóxica para algumas plantas.
A resistência induzida, ou efeito indireto, ocorre pela produ-ção de fitoalexinas nas plantas hospedeiras susceptíveis ao patógeno (SINHA, 1995). A aplicação de sais de Ni em baixas concentrações reduz a indicidência de doenças causadas por Helminthosporium oryzae no arroz, Pyricularia oryzae em plântulas de arroz e ferrugem no amendoim (WOOD e REILLY, 2007). Essa indução de resistência é caracterizada pela aplicação de Ni em concentrações não tóxicas para a planta hospedeira. Estudo realizado por Reilly, Crawford e Buck (2005) mostrou a resposta da incidência de Puccinia hemoro-callidis em Hemerocallis em função da aplicação de Ni. A ferrugem é a principal doença em Hemorocallis e traz muitos danos econômicos. Hemerocallis é uma planta ornamental também conhecida como lírio-de-um-dia. Apesar do nome, não são lírios verdadeiros (Lilium, Liliaceae). As flores da maior parte das espécies desabrocham pela manhã e murcham à noite, sendo estas substituídas por uma ou mais na mesma inflorescência no dia seguinte. Observou-se que uma apli-cação de Ni foi suficiente para eliminar a ferrugem, como ilustrado na Figura 13. A ferrugem desapareceu completamente 15 dias após a aplicação de sulfato de Ni, porém, ocorreu fitotoxidade na dose de 400 mg L-1. Nesse estudo, a germinação dos urediósporos em placas de Petri não foi afetada em dose de até 200 mg L-1. Essa dose não afetou a germinação do urediósporo do fungo, o que permite afirmar que o Ni aumentou a resistência da planta.
A ferrugem causada pelo fungo Fusicladosporium caryige-num é a principal doença da cultura da pecã, provocando perdas na produtividade e danos econômicos. A aplicação de Ni, além de corri-gir os sintomas de orelha-de-rato, promove resistência contra o fungo, como ilustrado na Figura 14. A germinação dos esporos do fungo Fusicladosporium caryigenum foi significativamente afetada pelas doses de Ni utilizadas no estudo. Isso sugere que o Ni, além de atuar como indutor de resistência nas plantas, apresenta também a função de fungicida. No entanto, são necessárias pesquisas relacionando a aplicação de Ni em culturas tropicais e a indução de resistência a microrganismos, patógenos e pragas nas condições brasileiras.
As plantas, em geral, possuem quatro estratégias de defesa contra os patógenos: 1) alta produção de cutina, suberina e cera; 2) produção de produtos secundários contendo N; 3) produção de compostos fenólicos e 4) produção de terpenos. A produção de cutinas, compostos secundários, compostos fenólicos e terpenos
Figura 13. Efeito do níquel na incidência da ferrugem (Puccinia hemero-callidis) em Hemerocallis.
Fonte: Cortesia do Dr. Bruce Wood (USDA - Department of Agriculture, Agricultural Research Service, USA).
200 ppm Ni2+ 100 ppm Ni2+ 0 ppm Ni2+
pela via do ácido mevalônico é altamente dependente da acetil CoA oriunda do piruvato. Bai et al. (2006) demonstraram em seus estudos que a deficiência de Ni afeta o metabolismo do carbono nas plantas. Plantas deficientes em Ni acumulam ácido lático nas folhas devido à baixa atividade da acetil CoA sintase e, consequentemente, acumulam altos teores de triptofano, valina, leucina e glicina. A correta pulverização de Ni nas plantas, além de ativar a síntese de fitoalexinas, aumenta a atividade da acetil CoA sintase, o que proporciona maior produção de compostos secundários e, conse-quentemente, maior tolerância das plantas ao ataque de patógenos.
REFERÊNCIAS AHMADA, M. S. A.; ASHRAF, M.; HUSSAINA, M. Phytotoxic effects of nickel on yield and concentration of macro-and micro-nutrients in sunflower (Helianthus annuus L.) achenes. Journal of Hazardous Materials, v. 185, p. 1295-1303, 2011.ALI, M. A.; ASHRAF, M.; ATHAR, H. R. Influence of nickel stress on growth and some important physiological/biochemical attributes in some diverse
Figura 14. Efeito da concentração de níquel na incidência da ferrugem em pecã (Fusicladosporium caryigenum).
Fonte: Cortesia do Dr. Bruce Wood (USDA - Department of Agriculture, Agricultural Research Service, USA).
0 ppm Ni2+ 25 ppm Ni2+ 50 ppm Ni2+
100 ppm Ni2+ 150 ppm Ni2+ 200 ppm Ni2+

INFORMAÇÕES AGRONÔMICAS Nº 147 – SETEMBRO/2014 21
canola (Brassica napus L.) cultivars. Journal of Hazardous Materials, v. 172, p. 964-969, 2009.ALOVISI, A. M. T.; MAGRI, J.; DUTRA, J. E.; MAGRI, E.; SANTOS, M. J. G.; ALOVISI, A. A. Adubação foliar com sulfato de níquel na cultura da soja. Ensaios e Ciência: Ciências Biológicas, Agrárias e da Saúde, v. 15, n. 2, p. 25-32, 2011.ALVES, J. D.; GUIMARãES, R. J. Sintomas de desordens fisiológicas em cafeeiro. In: GUIMARãES, R. J.; MENDES, A. N. G.; BALIzA, D. P. (Org.). Semiologia do cafeeiro: sintomas de desordens nutricionais, fitos-sanitárias e fisiológicas. 1. ed. Lavras: Editora UFLA, 2010. p. 169-215.ALVES, S.; TRANCOSO, M. A.; GONÇALVES, M. L. S.; SANTOS, M. M. C. A nickel availability study in serpentinised areas of Portugal. Geoderma, v. 164, p. 155-163, 2011.ALVES, J. D.; zANANDREA, I.; DEUNER, S.; GOULART, P. F. P.; SOUzA, K. R. D.; SANTOS, M. O. Antioxidative responses and morpho- anatomical adaptations to waterlogging in Sesbania virgata. Trees, v. 27, n. 3, p. 717-728, 2012.ANDREEVA, I. V.; GOVORINA, V. V.; YAGODIN, B. A.; DOSIMOVA, O. T. Dynamics of nickel accumulation and distribution in oat plants. Agrokhimiya, v. 4, p. 68-71, 2000.ANDREWS, R. K.; BLAKELEY, R. L.; zERNER, B. Urea and urease. Advances in inorganic biochemistry, v. 6, p. 245-283, 1984.ANzA, M.; RIGA, P.; GARBISU, C. Time course of antioxidant respon-se of Capsicum annuum subjected to progressive magnesium deficiency. Annals of Applied Biology, v. 146, p. 123-134, 2005.BAI, C.; REILLY, C. C.; WOOD, B. W. Nickel deficiency disrupts meta-bolism of ureides, amino acids, and organic acids of young pecan foliage. Plant Physiology, v. 140, n. 2, p. 433-443, 2006.BERTRAND, D. Importance du nickel, comme oligo-élément, pour les Rhizobium des nodosités des legumineuses. Comptes Rendus Hebdo-madaires des Seances de L´Academie des Sciences, v. 276, n. 12, p. 1855-1858, 1973.BETTINELLI, M.; BEONE, G. M.; SPEzIA, S.; BAFFI, C. Determination of heavy metals in soils and sediments by microwave-assisted digestion and inductively coupled plasma optical emission spectrometry analysis. Analytica Chimica Acta, v. 424, p. 289-296, 2000.BIONDI, C. M. Teores naturais de metais pesados nos solos de refe-rência do estado de Pernambuco. 2010. 58 p. Tese (Doutorado) - Uni-versidade Federal Rural de Pernambuco, Recife, 2010.BROOKS, R. R. Accumulation of nickel by terrestrial plants. In: NRIA-GU, J. O. (Ed). Nickel in the environment. New York: John Wiley, 1980. p. 407-430.BROOKS, R. R. Serpentine and its vegetation: a multidisciplinary approach. London: Croom Helm, 1987. 454 p.BROWN, P. H.; WELCH, R. M.; CARY, E. E. Nickel: a micronutrient essential for higher plants. Plant Physiology, v. 85, n. 3, p. 801-803, 1987.BUSCHLE, B. N. Valores de referência de qualidade para elementos--traço em solos da Planície Litorânea do Estado do Paraná. 2013. 49 p. Dissertação (Mestrado) – Universidade Federal do Paraná, Curitiba, 2013.CAIRES, S. M. Determinação dos teores naturais de metais pesados em solos do Estado de Minas Gerais como subsídio ao estabelecimento de Valores de Referência de Qualidade. 2009. 304 p. Tese (Doutorado) – Universidade Federal de Viçosa, Viçosa, 2009.CARIDAD-CANCELA, R.; PAz-GONzÁLEz, A.; ABREU, C. A. Total and extractable nickel and cadmium contents in natural soils. Commu-nications in Soil Science and Plant Analysis, v. 36, p. 241-252, 2005.CATALDO, D. A.; GARLAND, T. R.; WILDUNG, R. E. Nickel in plants: II. Distribution and chemical form in soybean plants. Plant Physiology, v. 62, p. 566-570, 1978.CHEN, C.; HUANG, D.; LIU, J. Functions and toxicity of nickel in plants: recent advances and future prospects. Clean, v. 37, n. 4-5, p. 304-313, 2009.CHESHIRE, M. V.; BERROW, M. L.; GOODMAN, B. A.; MUNDIE, C. M. Metal distribution and nature of some Cu, Mn and V complexes
in humic and fulvic acid fractions of soil organic matter. Geofhimica et Cosmochlmica Acta, v. 41, p. 1131-1138, 1977.DALTON, D. A.; EVANS, H. J.; HANUS, F. J. Stimulation by nickel of soil microbial urease activity and urease and hydrogenase activities in soy-beans growth in a low-nickel soil. Plant and Soil, v. 88, p. 245-258, 1985.DEMAREE, J. B. Little leaf disease of pecans. Phytopathology, v. 16, p. 277-283, 1926.DEUNER, S.; ALVES, J. D.; zANANDREA, I.; GOULART, P. F. P.; SIL-VEIRA, N. M.; HENRIQUE, P. C.; MESQUITA, A. C. Stomatal behavior and components of the antioxidative system in coffee plants under water stress. Scientia Agricola, v. 68, n. 1, p. 77-85, 2011.DIXON, N. E.; GAzzOLA, C.; BLAKELEY, R. L.; zERNER, B. Jack bean urease (EC 3.5.1.5) a metalloenzyme: simple biological role for nickel. Journal of the American Chemical Society, v. 97, n.14, p. 4131-4133, 1975.DUBEY, R. C.; DWINVEDI, R. S. Effect of heavy stress metals on growth and survival of Macrophomina phaseolina (Tassi). Biology and Fertility of Soils, v. 6, p. 311-314, 1988.EL-SHINTINAWY, F.; EL-ANSARY, A. Differential effect of Cd2+ and Ni2+ on amino acid metabolism in soybean seedlings. Biologia Plantarum, v. 43, p. 79-84, 2000.EMBRAPA. EMPRESA BRASILEIRA DE PESQUISA AGROPECUÁ-RIA. Sistema brasileiro de classificação de solos. 2. ed. Rio de Janeiro: Embrapa Solos, 2006. 306 p.ESKEW, D. L.; WELCH, R. M.; CARY, E. E. Nickel: an essential mi-cronutrient for legumes and possibly all higher-plants. Science, v. 222, n. 4624, p. 621-623, 1983. ESKEW, D. L.; WELCH, R. M.; NORVELL, W. A. Nickel in higher plants: further evidence for an essential role. Plant Physiology, v. 76, n. 3, p. 691-693, 1984.EVANS, H. J.; HARKER, A. R.; PAPEN, H.; RUSSELL, S. A.; HA-NUS, F. J.; zUBER, M. Physiology, biochemistry and genetics of the uptake hydrogenase in rhizobia. Annual Review of Microbiology, v. 41, p. 335-361, 1987.EWAIS, E. A. Effects of cadmium, nickel and lead on growth, chlorophyll content and proteins of weeds. Biologia Plantarum, v. 39, p. 403-410, 1997.FIDALGO, F.; SANTOS, A.; SANTOS, I.; SALEMA, R. Effects of long term salt stress on antioxidant defence systems, leaf water relations and chloroplast ultrastructure of potato plants. Annals of Applied Biology, v. 145, p. 185-192, 2004.FOYER, C. H.; NOCTOR, G. Redox homeostasis and antioxidant signa-ling: A metabolic interface between stress perception and physiological responses. The Plant Cell, v. 17, p. 1866-1875, 2005.FOYER, C. H.; LOPEz-DELGADO, H.; DAT, J. F.; SCOTT, I. M. Hydro-gen peroxide and glutathione-associated mechanism of acclimatory stress tolerance and signalling. Physiologia Plantarum, v. 100, p. 241-254, 1997.FREEMAN, J. L.; PERSANS, M. W.; NIEMAN, K.; ALBRECHT, C.; PEER, W.; PICKERING, I. J.; SALT, D. E. Increased glutathione biosynthe-sis plays a role in nickel tolerance in Thlaspi nickel hyperaccumulators. The Plant Cell, v. 16, p. 2176-2191, 2004.FUENTES, I. I.; ESPADAS-GIL, F.; TALAVERA-MAY, C.; FUENTES, G.; SANTAMARÍA, J. M. Capacity of the aquatic fern (Salvinia minima Baker) to accumulatehigh concentrations of nickel in its tissues, and its effect on plantphysiological processes. Aquatic Toxicology, v. 155, p. 142-150, 2014.GAJEWSKA, E.; SKLODOWSKA, M. Nickel-induced changes in nitro-gen metabolism in wheat shoots. Journal of Plant Physiology, v. 166, p. 1034-1044, 2009.GABE, U. Teor e disponibilidade para a soja de micronutrientes e elementos potencialmente tóxicos em fertilizantes minerais e calcários. 1998. 84 p. Tese (Doutorado) – Escola Superior de Agricultura Luiz de Queiroz, Universidade de São Paulo, Piracicaba, 1998.GAMMON, N., SHARPE, R. H. Mouse-ear: A manganese deficiency of pecans. Proceedings of the American Society for Horticultural Science, v. 68, p. 195-200, 1956.

22 INFORMAÇÕES AGRONÔMICAS Nº 147 – SETEMBRO/2014
GERENDÁS, J., SATTELMACHER, B. Influence of Ni supply on growth, urease activity and nitrogen metabolites of Brassica napus grown with NH4NO3 or urea as N source. Annals of Botany, v. 83, p. 65-71, 1999.GERENDÁS, J.; SATTELMACHER, B. Significance of Ni supply for gro-wth, urease activity and the concentrations of urea, amino acids and mineral nutrients of urea-grown plants. Plant and Soil, v. 190, p. 153-162, 1997.GERENDÁS, J.; zHU, z.; SATTELMACHER, B. Influence of N and Ni supply on nitrogen metabolism and urease activity in rice (Oryza sativa L.). Journal of Experimental Botany, v. 49, n. 326, p. 1545-1544, 1998.GERENDÁS, J.; POLACCO, J. C.; FREYERMUTH, S. K.; SATTEL-MACHER, B. Significance of nickel for plant growth and metabolism. Journal of Plant Nutrition and Soil Science, v. 162, p. 241-256, 1999.GRAHAM, R. D.; WELCH, R. M.; WALKER, C. D. A role for nickel in the resistance of plants to rust. In: AUSTRALIAN AGRONOMY CONFERENCE, 3., 1985, Hobart. Annals... Hobart: Australian Society of Agronomy, 1985. 337 p.GRATãO, P. L.; POLLE, A.; LEA, P. J.; AzEVEDO, R.A. Making the life of heavy metalstressed plants a little easier. Functional Plant Biology, v. 32, p. 481-494, 2005.GRAUKE, L. J.; MORRIS, H. J.; KOWALCzUK, J. G. Mouse-ear: A symptom of nutrient imbalance. Proc. Southeastern Pecan Growers Association, v. 76, p. 141-147, 1983.HARMSEN, J. Measuring bioavailability: from a scientific approach to stan-dard methods. Journal of Environmental Quality, v. 36, p. 1420-1428, 2007.HIRAYAMA, C.; SUGIMURA, M.; SAITO, H.; NAKAMURA, M. Pu-rification and properties of urease from leaf of mulberry. Morus alba, Phytochemistry, v. 53, p. 325-330, 2002.INOUHE, M. Phytochelatins. Brazilian Journal of Plant Physiology, v. 17, p. 65-78, 2005.KABATA-PENDIAS, A.; PENDIAS, H. Trace elements from soil and plant. 4. ed. New York: CRC Press, Taylor and Francis Group, 2011. 576 p.KEVRESAN, S.; PETROVIC, N.; POPOVIX, M.; KANDRAC, J. Effect of heavy metals on nitrate and protein metabolism in sugar beet. Biologia Plantarum, v. 41, p. 235-240, 1998.KHAN, M. W.; SALAM, M. A. Interactions of Meloidogyne javanica, Fu-sarium udum and Rihizobium on pigeon pea in the presence of nickel and cobalto as pollutants. Annals of Applied Biology, v. 116, p. 549-556, 1990.KHOSHGOFTARMANESHA, A. H.; HOSSEINIB, F.; AFYUNIA, M. Nickel supplementation effect on the growth, urease activity and urea and nitrate concentrations in lettuce supplied with different nitrogen sources. Scientia Horticulturae, v. 130, p. 381-385, 2011.KISHORE, G. K.; PANDE, S.; RAO, J. N. Control of foliar diseases of groundnut using inorganic and metal salts. International Arachis News-letter, v. 21, p. 33-35, 2001.KLUCAS, R. V. Nickel, a micronutrient for hydrogen dependent growth of Rhizobium japonicum and for expression of urease activity in soybean leaves. Proceedings of the National Academy of Sciences of uSA, v. 80, p. 2253-2257, 1983.KOCHIAN, L. V. Mechanisms of micronutrient uptake and translocation in plants. In: LUXMOORE, R. J. (Ed.). Micronutrients in agriculture. 2. ed. Madison, Wis.: Ed. Soil Sci. Soc. Amer. Inc., 1991. p. 229-296.KOVACEVIC, G.; KASTORI, R.; MERKULOC, L. J. Dry matter and leaf structure in young wheat plants as affected by cadmium, lead and nickel. Biologia Plantarum, v. 42, p. 119-123, 1999.KRAMER, U.; COTTER-HOWELLS, J. D.; CHARNOCK, J. M.; BAKER, A. J. M.; SMITH, A. C. Free histidine as a metal chelator in plants that accumulate nickel. Lett. Nature, v. 379, p. 635-638, 1996.KRAMER, U.; SMITH, R. D.; WENzEL, W. W.; RASKIN, I.; SALT, D. E. The role of metal transport and tolerance in nickel hyperaccumulation by Thlaspi goesingense. Plant Physiology, v. 115, p. 1641-1650, 1997.KROGMEIER, M. J.; McCARTY, G. W.; BREMNER, J. M. Phytotoxicity of foliar-applied urea. Proceedings of National Academy of Science, v. 86, p. 8189, 1989.
KUKKOLA, E.; RAUTIO, P.; HUTTUNEN, S. Stress indications in copper- and nickel-exposed Scots pine seedlings. Environmental and Experimental Botany, v. 43, p. 197-210, 2000.KUPPER, H.; LOMBI, E.; zHAO, F. J.; WIESHAMMER, G.; McGRA-TH, S. P. Cellular compartmentation of nickel in the hyperaccumulators Alyssum lesbiacum, Alyssum bertolonii and Thlaspi goesingense. Journal of Experimental Botany, v. 52, p. 2291-3000, 2001.KUTMAN, B. Y.; KUTMAN, U. B.; CAKMAK, I. Effects of seed nickel reserves or externally supplied nickel on the growth, nitrogenmetabolites and nitrogen use efficiency of urea- or nitrate-fed soybean. Plant and Soil, v. 376, p. 261-276, 2014.KUTMAN, B. Y.; KUTMAN, U. B.; CAKMAK, I. Nickel-enriched seed and externally supplied nickel improve growth and alleviate foliar urea damage in soybean. Plant and Soil, v. 1007, p. 1-15, 2012.LICHT, O. A. B.; XUEJING, X.; QIN, z.; MIYAzAWA, M.; FERREIRA, F. J. F.; PLAWIAK, R. A. B. Average reference values of geochemical and geophysical variables in stream sediments and soils, state of Paraná, Brazil. Boletim Paranaense de Geociências, v. 58, p. 59-87, 2006.LOVATT, C. J.; zHENG, Y.; HAKE, K.D. Demonstration of a change in nitrogen metabolism influencing flower initiation in Citrus. Israel Journal of Botany, v. 37, p. 181-188, 1988.MALAN, H. L.; FARRANT, J. M. Effects of the metal pollutants cadmium and nickel on soybean seed development. Seed Science Research, v. 8, p. 445-453, 1998.MALAVOLTA, E.; LEãO, H. C.; OLIVEIRA, S. .; LAVRES JÚNIOR, L.; MORAES, M. F.; CABRAL, C. P.; MALAVOLTA, M. Repartição de nutrientes nas flores, folhas e ramos da laranjeira cultivar Natal. Revista Brasileira de Fruticultura, v. 2, n. 3, p. 506-511, 2006.MAPA. MINISTéRIO DA AGRICULTURA, PECUÁRIA E ABASTE-CIMENTO. Instrução Normativa MAPA 5/2007 (D.O.u. 01/03/2007). Disponível em: <http://www.agricultura.gov.br/vegetal/sanidade-vegetal/legislacao>. Acesso em: 31 julho de 2014.MARz, J. Diseases and insect pests of pecans. Fla. Agr. Expt. Sta. Bul., v. 147, p. 135-163, 1918.MARSCHNER, P. Mineral nutrition of higher plants. 3. ed. London: Academic Press, 2012. 64351 p.MASSOURA, S. T.; ECHEVARRIA, G.; BECQUER, T.; GHANBAJA, J.; LECLERC-CESSAC, E.; MOREL, J. L. Control of nickel availability by nickel bearing minerals in natural and anthropogenic soils. Geoderma, v. 136, p. 28-37, 2006.McGRATH, S. P.; SMITH, S. Nickel. In: ALLOWAY. B. J. Heavy metals in soils. New York: John Wiley, 1990. p. 125-50.MELO, W. J.; AGUIAR, P. S.; MELO, G. M. P.; MELO, V. P. Nickel in a tropical soil treated with sewage sludge and cropped with maize in a long- term field study. Soil Biology and Biochemistry, v. 39, p. 1341-1347, 2007.MILOSEVIC, N.; GOVEDARICA, M.; KASTORI, R.; PETROVIC, N. Effect of nickel on wheat plants, soil microorganisms and enzymes. Studia universitatis Babes-Bolyai Biologia, v. 47, n. 1, p. 79-84, 2002.MINEROPAR. MINERAIS DO PARANÁ S. A. Geoquímica de solo - horizonte B: relatório final de projeto. Curitiba, v. 1, 2005a. 408 p. MINEROPAR. MINERAIS DO PARANÁ S. A. Geoquímica de solo - horizonte B: relatório final de projeto. Curitiba, v. 2, 2005b. 449 p.MISHRA, D., KAR, M. Nickel in plant growth and metabolism. The Botanical Review, v. 40, p. 395-452, 1974.MOBLEY, H. L. T.; HAUSINGER, R. P. Microbial ureases: significance, regulation, and molecular characterization. Microbiological Reviews, v. 53, n. 1, p. 85-108, 1989.MOBLEY, H. L. T.; ISLAND, M. D.; HAUSINGER, R. P. Molecular biology of microbial ureases. Microbiological Reviews, v. 59, n. 3, p. 451-480, 1995.MONTARGÈS-PELLETIER, E.; CHARDOT, V.; ECHEVARRIA, G.; MI-CHOT, L. J.; BAUER, A.; MOREL, J. L. Identification of nickel chelators in three hyperaccumulating plants: an X-ray spectroscopic study. Phytoche-mistry, v. 69, p. 1695-1709, 2008.

INFORMAÇÕES AGRONÔMICAS Nº 147 – SETEMBRO/2014 23
MORAES, M. F.; ABREU JUNIOR, C. H.; LAVRES JUNIOR, J. Micro-nutrientes. In: PROCHNOW, L. I.; CASARN, V.; STIPP, S. R. (Ed.). Boas práticas para uso eficiente de fertilizantes. Piracicaba: International Plant Nutrition Institute, v. 2., 2010. p. 205-278.MUÑOz, A.; RASO, M. J.; PINEDA, M.; PIEDRAS, P. Degradation of ureidoglycolate in French bean (Phaseolus vulgaris) is catalysed by a ubi-quitous ureidoglycolate urea-lyase. Planta, v. 224, n. 1, p. 175-184, 2006.NETO, A. P. Metabolismo do nitrogênio e concentração de nutrientes no cafeeiro irrigado em razão da dose de nitrogênio. 94 p. 2009. Dissertação (Mestrado) – Escola Superior de Agricultura “Luiz de Quei-roz”, Universidade de São Paulo, Piracicaba, 2009.NYCzEPIR, A. P.; WOOD, B. W.; REILLY, C. C. Association of Meloi-dogyne partityla with mouse-ear disorders of pecan in Georgia. Journal of Nematology, v. 4, p. 272, 2006.PALACIOS, G.; MATAIX, J. The influence of organic amendment and ni-ckel pollution on tomato fruit yield and quality. Journal of Environmental Science and Health, Part B, v. 34, p. 133-150, 1999.PAGE, V.; FELLER, U. Selective transport of zinc, manganese, nickel, cobalt and cadmium in the root system and transfer to the leaves in young wheat. Annals of Botany, v. 96, p. 425-434, 2005.PANDAA, G. C.; DASA, S. K.; BANDOPADHYAYB, T. S.; GUHA, A. K. Adsorption of nickel on husk of Lathyrus sativus: behavior and binding mechanism. Colloids Surf B: Biointerfaces, v. 57, p. 135-142, 2007. PANDOLFINI, T.; GABBRIELI, R.; COMPARINI, C. Nickel toxicity and peroxidase activity in seedlings of Triticum aestivum L. Plant Cell Enviroment, v. 15, p. 719-725, 1992.PAIVA, H. N.; CARVALHO, J. G.; SIQUEIRA, J. O.; FERNANDES, A. R.; MIRANDA, J. R. P. Efeito da aplicação de doses crescentes de níquel sobre o teor e o conteúdo de nutrientes em mudas de ipê-roxo (Tabebuia impetiginosa (Mart) Standley). Scientia Forestalis, n. 63, p. 158-166, 2003.PAYE, H. S.; MELLO, J. W. V.; ABRAHãO, W. A. P.; FERNANDES FI-LHO, E. I.; DIAS, L. C. P.; CASTRO, M. L. O.; MELO, S. B.; FRANÇA, M. M. Valores de referência de qualidade para metais pesados em solos no Estado do Espírito Santo. Revista Brasileira de Ciência do Solo, v. 34, p. 2041-2051, 2010.PENNAzIO, S., ROGGERO, P. Effects of trace elements on the natural resistance of asparagus bean to tobacco necrosis virus and ethylene pro-duction. Advances in Horticultural Science, v. 2., 23-26, 1988.PONIEDzIAŁEK, M.; SĘKARA, A.; CIURA, J.; JĘDRSzCzYK, E. Nickel and manganese accumulation and distribution in organs of nine crops. Folia Horticulturae, v. 17, n. 1, p. 11-22, 2005.PONIzOVSKY, A. A.; THAKALI, S.; ALLEN, H. E.; DI TORO, D. M.; ACKERMAN, A. J.; METzLER, D. M. Nickel partitioning in acid soils at low moisture content. Geoderma, v. 145, p. 69-76, 2008.POLACCO, J. C. Nitrogen metabolism in soybean tissue culture. Plant Physiology, v. 59, p. 827-830, 1977.POLLACCO, J. C.; HOLLAND, M. A. Roles of urease in plant cells. International Review of Cytology, v. 145, p. 65-103, 1993.POLLARD, A. J.; POWELL, K. D.; HARPER, H. A.; SMITH, J. Á. C. The genetic basis of metal hyperaccumulation in plants. Critical Reviews in Plant Sciences, v. 21, p. 539-566, 2002.POULIK, z. The danger of accumulation of nickel in cereals oncontamina-ted soil. Agriculture, Ecosystems and Environment, v. 63, p. 25-29, 1997.QUANTIN, C.; ETTLER, V.; GARNIER, J.; SEBEK, O. Sources and extractibility of chromium and nickel in soil profiles developed on Czech serpentinites. Comptes Rendus Geoscience, v. 340, p. 872-882, 2008.RAJAIE, M.; KARIMIAN, N.; YASREBI, J. Nickel transformation in two calcareous soil textural classes as affected by applied nickel sulfate. Geoderma, v. 144, p. 344-351, 2008.RAO, K. V. M.; SRESTY, T. V. S. Antioxidative parameters in seedling of pige on pea (Cajanus cajan L.) Millspaugh to zn and Ni stress. Plant Science, v. 157, v. 1, p. 113-128, 2000.
RHAMANM, H.; SABREEN, S.; ALAM, S.; KAWAI, S. Effects of nickel on growth and composition of metal micronutrients in barley plants grown in nutriente solution. Journal of Plant Nutrition, v. 28, p. 393-404, 2005.REBAFKA, F. P.; SCHULz, R.; MARSCHNER, H. Erhebungsunter-suchungen zur pflanzenverfügbarkeit von nickel auf böden mit hohen geogenen nickelgehalten. Angewandte Botanik, v. 64, p. 317-328, 1990.REEVES, R. D.; BROOKS, R. R.; MacFARLANE, R. M. Nickel uptake by californian Streptanthus and Caulanthus with particular reference to the hyperaccumulator S. polygaloides Gray (Brassicacea). American Journal of Botany, v. 68, p. 708-712, 1981.REEVES, R. D.; BAKER, A. J. M.; BECQUER, T.; ECHEVARRIA, G.; MIRANDA, z. J. G. The Xora and biogeochemistry of the ultramafic soils of Goiás state, Brazil. Plant and Soil, v. 293, p. 107-119, 2007.REILLY, C.; CRAWFORD, M.; BUCK, J. Nickel suppresses daylily rust, Puccinia hemerocallidis on susceptible daylilys, Hemerocallis in greenhouse and field trials. (Abstr.). Phytopathology, v. 95, p. S88- S88, 2005.REIS, A.R.; FAVARIN, J. L.; GALLO, L. A.; MALAVOLTA, E.; MORAES, M. F.; LAVRES JUNIOR, J. Nitrate reductase and glutamine synthetase activity in coffee leaves during fruit development. Revista Brasileira de Ciência do Solo, v. 33, p. 315-324, 2009.REIS, A. R.; FAVARIN, J. L.; GALLO, L. A.; MORAES, M. F.; TEzOT-TO, T.; LAVRES JUNIOR, J. Influence of nitrogen fertilization on nickel accumulation and chemical composition of coffee plants during fruit development. Journal of Plant Nutrition, v. 34, p. 1853-1866, 2011.REVOREDO, M. D.; MELO, W. J. Disponibilidade de níquel em solo tratado com lodo de esgoto e cultivado com sorgo. Bragantia, v. 65, n. 4, p. 679-685, 2006.RIESEN, O.; FELLER, U. Redistribution of nickel, cobalt, manganese, zinc, and cadmium via the phloem in young and maturing wheat. Journal of Plant Nutrition, v. 28, p. 421-430, 2005.RODAK, B. W. Níquel em solos e na cultura de soja. 2014. 101 p. Dissertação (Mestrado) – Universidade Federal do Paraná, Curitiba, 2014.ROMERO-PUERTAS, M. C.; RODRIGUEz-SERRANO, M.,; CORPAS, F. J.; GÓMEz, M.; DEL RIO, L. A.; SANDALIO, L. M. Cadmium-induced subcellular accumulation of O2•- and H2O2 in pea leaves. Plant, Cell and Environment, v. 27, p. 1122-1134, 2004.ROVERS, H.; CAMARGO, O. A.; VALADARES, J. M. A. S. Níquel total e solúvel em DTPA em solos do estado de São Paulo. Revista Brasileira de Ciência do Solo, v. 7, n. 3, p. 217-220, 1983.ROS, R.; COOKE, D. T.; BURDEN, R. S.; JAMES, C. S. Effects of the herbicide MCPA, and the heavy metals cadmium and nickel on lipids composition, Mg2+- ATPase activity and fluidity of plasma membrane from rice, Oryza sativa (cv. Bahia) shoots. Journal of Experimental Botany, v. 41, p. 457-462, 1990.RUBIO, M. I.; ESCRIG, I.; MARTINEz-CORTINA, C.; LOPEz-BENET, F. J.; SPANz, A. Cadmium and nickel accumulation in rice plants. Effects on mineral nutrition and possible interaction of abscisic and gibberellic acids. Plant Growth Regulations, v. 14, p. 151-157, 1994. RUTER, J. M. Effect of nickel applications for the control of mouse ear disorder on river birch. Journal of Environmental Horticulture, v. 23, n. 1, p. 17-20, 2005.SABIR, M.; GHAFOOR, A.; SAIFULLAH, M.; REHMAN, z. U.; AH-MAD, H. R.; AzIz, T. Growth and metal ionic composition of Zea mays as affected by nickel supplementation in the nutrient solution. International Journal of Agriculture and Biology, v. 13, n. 2, p. 186-190, 2011.SAGNER, S.; KNEER, R.; WANNER, G.; COSSON, J. P.; DEUS- NEUMANN, B.; zENK, M. H. Hyperaccumulator, complexation and distribution of nickel in Sebertia acuminata. Phytochemistry, v. 47, n. 3, p. 339-343, 1998.SAJWAN, K. S.; ORNES, W. H.; YOUNGBLOOD, T. V.; ALVA, A. K. Uptake of soil applied cadmium, nickel and selenium by bush beans. Water, Air, Soil Pollution, v. 91, p. 209-217, 1996.

24 INFORMAÇÕES AGRONÔMICAS Nº 147 – SETEMBRO/2014
SANTOS, S. N. Valores de referência de metais pesados em solos de Mato Grosso e Rondônia. 2011. 101 p. Dissertação (Mestrado) – Escola Superior de Agricultura “Luiz de Queiroz”, Universidade de São Paulo, Piracicaba, 2011.SAUVé, S.; HENDERSHOT, W.; ALLEN, H. E. Solid-solution parti-tioning of metals in contaminated soils: dependence on pH, total metal burden and organic matter. Environmental Science and Technology, v. 34, p. 1125-1131, 2000.SCHICKLER, H.; CASPI, H. Response of antioxidative enzymes to ni-ckel and cadmium stress in hyperaccumulator plants of Genus, Alyssum. Physiologia Plantarum, v. 105, p. 39-44, 1999.SENWO, z. N.; TAzISONG, I. A. Metal contents in soils of Alabama. Communications in Soil Science and Plant Analysis, v. 35, p. 2837-2848, 2004.SERENGIN, I. V.; KOzHEVNIKOVA, A. Physiological role of nickel and its toxic effects on higher plants. Russian Journal of Plant Physiology, v. 53, n. 2, p. 257-277, 2006.SERVENE, B. C. Nickel accumulation by Hybanthus floribundus. Nature, v. 248, p. 807-808, 1974.SHEORAN, I. S.; SINGAL, H. R.; SINGH, R. Effect of cadmium and nickel on photosynthesis and the enzymes of the photosynthetic car-bon reduction cycle in pigeonpea (Cajanus cajan L.). Photosynhtesis Research, v. 23, p. 345-351, 1990.SIEBIELEC, G,; CHANEY, R. L. Manganese fertilizer requirement to prevent manganese defi ciency when liming to remediate Ni-phytotoxic soils. Communications in Soil Science and Plant Analysis, v. 37, p. 1-17, 2006.SINHA, A. K. Possible role of phytoalexins inducers chemicals in plant disease control. In: DANIEL, M.; PURKAYASTHA, R. P. (Ed.). Han-book of phytoalexin metabolism and action. New York: Marcel Dekker, 1995. p. 555-591.SINGH, R. K.; SHUKLA, R. R.; DWINVEDI, R. S. Effect of cádmium, cobalt and nickel salts on the survivability of sclerotia and plant infection by Sclerotium rolfsii causing root-rot disease of barley. Tropical Science, v. 332, p. 269-274, 1992.SINGH, S.; KAYASTHA, A. M.; ASTHANA, R. K.; SINGH, S. P. Res-ponse of garden pea to nickel toxicity. Journal of Plant Nutrition, v. 27, p. 1543-1560, 2004.SMITH, S. R. A critical review of the bioavailability and impacts of heavy metals in municipal solid waste composts compared to sewage sludge. Environment International, v. 35, p. 142-156, 2009.SMITH, N. G.; WOODBURN, J. Nickel and ethylene involvement in the senescence of leaves and flowers. Naturwissenschaften, v. 71, n. 4, p. 210-211, 1984.SRESTY, T. V. S.; MADHAVA RAO, K. V. Ultrastructural alterations in response to zinc and nickel stress in the root cells of Pigeonpea. Environ-mental and Experimental Botany, v. 41, p. 3-13, 1999.STANKOVIC, z.; PAJEVIC, S.; VUCKOVIC, M.; STOJANOVIX, S. Concentrations of trace metals in dominat aquatic plants of the lake Provala. Biologia Plantarum, v. 43, p. 583-585, 2000.SUGHA, S. K.; SING, B. M. Comparative efficacy of fungicides against blister blight of tea (Exobasidium vexans Masee). Indian Journal of Mycology and Plant Pathology, v. 20, p. 211-219, 1990.SUzUKI, T. Total nitrogen and free amino acids in Morus alba stems from autumn through spring. Physiologia Plantarum, v. 60, p. 473, 1984.TABATABAEI, S. J. Supplements of nickel affect yield, quality, and nitrogen metabolism when urea or nitrate is the sole nitrogen source for cucumber. Journal of Plant Nutrition, v. 32, p. 713-724, 2009.TAN, X. W.; IKEDA, H.; ODA, M. Effects of nickel concentration in the nutrient solution on the nitrogen assimilation and growth of tomato seedlings in hydroponic culture supplied with urea or nitrate as the sole nitrogen source. Scientia Horticulturae, v. 84, p. 265-273, 2000.
TAYLOR, G. J.; CROWDER, A. A. Copper and nickel tolerance in Typha latifolia clones from contaminated and uncontaminated environments. Canadian Journal of Botany, v. 62, p. 1304-1308, 1984.
TEzOTTO, T.; FAVARIN, J. L.; AzEVEDO, R. A.; ALLEONI, L. R. F.; MAzzAFERA, P. Coffee is highly tolerant to cadmium, nickel and zinc: plant and soil nutritional status, metal distribution and bean yield. Field Crops Research, v. 125, p. 24-34, 2012.
TODD, C. D.; POLLACCO, J. C. Soybean cultivars ‘Williams 82’ and ‘Maple Arrow’ produce both urea and ammonia during ureide degradation. Journal of Experimental Botany, v. 55, n. 398, p. 867-877, 2004.
UREN, N. C. Forms, reaction and availability of nickel in soils. Advances in Agronomy, v. 48, p. 141-203, 1992.
VANSELOW, A. P. Nickel. In: CHAPMAN, H. D. (Ed.). Diagnostic criteria for plants and soils. Riverside: University of California, 1966. p. 302-309.
VELIKOVA, V.; TSONEV, T.; LORETO, F.; CENTRITTO, M. Changes in photosynthesis, mesophyll conductance to CO2, and isoprenoid emissions in Populus nigra plants exposed to excess nickel. Environmental Pollution, v. 159, p. 1058-1066, 2011.
WALKER, C. D.; GRAHAM, R. D.; MADISON, J. T.; CARY, E. E.; WEL-CH, R. M. Effects of Ni deficiency on some nitrogen metabolites in cowpeas (Vigna unguiculata L. Walp.). Plant Physiology, v. 79, p. 474-479, 1985.
WANG, H.; zENG, F.; KANG, J. Resistance to bacterial blight induced by nickel nitrate in rice seedling. Journal of Hunan Agriculture university, v. 26, p. 119-121, 2000.
WANG, S.; NAN, z.; LIU, X.; LI, Y.; QIN, S.; DING, H. Accumulation and bioavailability of copper and nickel in wheat plants grown in contaminated soils from the oasis, northwest China. Geoderma, v. 152, p. 290-295, 2009.
WELCH, R. M. The biological significance of nickel. Journal of Plant Nutrition, v. 3, p. 345-356, 1981.
WHITE, P. J. Long-distance transport in the xylem and phloem. In: MARS-CHNER, P. (Ed.). Mineral nutrition of higher plants. 3. ed. United States of America: Elsevier, 2012. p. 49-70.
WOOD, B. W.; REILLY, C. C. Nickel and plant disease. In: DATNO-FF, L. E.; ELMER, W. H.; HUBER, D. M. (Ed.). Mineral nutrition and plant disease. The American Phytopathological Society, 2007. p. 215-231.WOOD, B. W.; REILLY, C. C.; NYCzEPIR, A. P. Field deficiency of ni-ckel in trees: symptoms and causes. Acta Horticulturae, v. 721, p. 83-97, 2006.WOOD, B. W.; REILLY, C. C.; NYCzEPIR, A. P. Mouse-ear of pecan: I. Symptomatology and occurrence. HortScience, v. 39, n. 1, p. 87-94, 2004a. WOOD, B. W.; REILLY, C. C.; NYCzEPIR, A. P. Mouse-ear of pecan: II. Influence of nutrient applications. HortScience, v. 39, n. 1, p. 95-100, 2004b. WOOD, B. W.; REILLY, C. C.; NYEzEPIR, A. P. Mouse-ear of pecan: A nickel deficiency. HortScience, v. 39, n. 6, p. 1238-1242, 2004c.WORLEY, R. E. Pecan cultivars differ in leaf elemental concentration of normal and mouse-ear leaf tissue. HortScience, v. 14, p. 51-52, 1979.
YANG, X. E.; BALIGAR, V. C.; FOSTER, J. C.; MARTENS, D. C. Accumulation and transport of nickel in relation to organic acids in rye-grass and maize grown with different nickel level. Plant and Soil, v. 196, p. 271-276, 1997.YUSUF, M.; FARIDUDDIN, Q.; HAYAT, S.; AHMAD, A. Nickel: an overview of uptake, essentiality and toxicit in plants. Bulletin of Environ-mental Contamination and Toxicology, v. 86, p. 1-17, 2011.
zHENG, Q. L.; NAKATSUKA, A.; MATSUMOTO, T.; ITAMURA, H. Pre-harvest nickel application to the calyx of ‘Saijo’ persimmon fruit prolongs postharvest shelf-life. Postharvest Biology and Technology, v. 42, p.98-103, 2006.