Embed Size (px)
Citation preview

James E. NocekRuminal Dry Matter and Protein Digestion
Evaluation of Specific Variables Affecting In Situ Estimates of
1985, 60:1347-1358.J ANIM SCI
http://jas.fass.org/content/60/5/1347services, is located on the World Wide Web at:
The online version of this article, along with updated information and
www.asas.org
by guest on October 7, 2011jas.fass.orgDownloaded from

EVALUATION OF SPECIFIC VARIABLES AFFECTING IN SITU ESTIMATES OF RUMINAL DRY MATTER AND PROTEIN DIGESTION 1
James E. Nocek
Agway Inc. 2 , Syracuse, NY 13221
Summary
Specific variables associated with the in situ digestion of soybean meal (SBM) were identified and investigated. Soybean meal was incubated in situ for seven periods of time ranging from 1 to 48 h. Bags were introduced into the tureen in two ways: (1) all bags introduced at the same time and (2) bags introduced at different times (all removed at once). The latter procedure resulted in higher rate constants for both dry matter (DM) and nitrogen (N) disappearance. Dry matter disappearance rates of cottonseed meal were not different between four cows in similar stages of lactation. However, differences developed within cows over a 6-wk period. Correction for bacterial N did not significantly alter rate of SBM N disappearance compared with no correction. The amount of SBM DM and N washed from bags before ruminal ex- posure varied with sample grind and weight. Disappearance was higher for bags with poros- ities of 80 and 102/am compared with 6, 20, 40 or 59 /am. Grinding SBM at 1, 2, 5 mm or unground did not influence rate constants for DM or N disappearance from polyester bags. As the ratio of sample weight to surface area increased (2.5, 12.6, 25.3 and 37.9 mg/cm2), DM and N disappearance rates decreased. Bag porosities of 6 and 20 ~m gave lowest and 80 and 102 /am highest rate constants of in situ DM and N disappearance. For SBM, a sample weight to surface area ratio of 12.6 mg/cm 2 and use of bags with porosities of 40, 59, 80 and 102/am resulted in acceptable estimates of ruminal protein availability.
1The author would like to express appreciation to A. Warner, D. G. Braund, R. Ellis, C. Olcott and K. Nash for technical assistance and J. English for assist- ance with statistical analyses.
Dept. of Res. and Dev. 3 Tetko, Inc., Elmsford, NY. Received March 26, 1984. Accepted January 22, 1985.
(Key Words: Polyester Bag, Ruminal Available Protein, Sample Grind, Sample Weight, Bag Porosity.)
Introduction
Factors influencing in situ measurements have been examined; however, recommendations between laboratories are not consistent. Weakley et al. (1977) indicated that dry matter (DM) and nitrogen (N) degradation was less with coarse (2,000/am) than with a fine particle size (500 gm). Uden et al. (1974) suggested a ratio of sample weight to bag surface area of 3.5 mg/cm 2, which is below the maximum of 10 mg/em 2 recommended by Van Hellen and Ellis (1977). Except for barley, Lindberg and Varvikko (1982) reported that DM disap- pearance was greatest from bags with 36-/am pores, followed by 20 and 10 #m.
The in situ digestion technique cannot characterize the digestion rate of solubles and(or) filterable efflux from bags. The possible confounding effect of this fraction may in- fluence digestion rates of insoluble, potentially available materials.
Researchers (Akin and Amos, 1975) have shown that certain ruminal bacteria tenaciously attach to plant particles. The impact of this contamination on N digestion rates has not been investigated.
The objective of the present research was to evaluate specific variables associated with in situ DM and N digestion of SBM. These variables included: bag introduction sequence into the rumen, cow and time variation, bacterial N con- tamination, presoaking, sample weight:bag surface area ratio and bag porosity.
Materials and Methods
Polyester 3 bags measuring 22 x 9 cm with porosities of 59 -+ 2/am were used in all experi- ments, except where otherwise indicated. Seams were double-sewn with nylon thread and
1347 JOURNAL OF ANIMAL SCIENCE, Vol. 60, No. 5, 1985
by guest on October 7, 2011jas.fass.orgDownloaded from

1348 NOCEK
Ingredient
TABLE 1. COMPOSITION OF CONCENTRATE MIX FED TO LACTATING, RUMINAL-CANNULATED COWS a
Percentage b
Hominy feed (IFN 4-02-887) Brewers grains (IFN 5-02-141) Dehydrated alfalfa (IFN 1-00-023) Soybean meal (IFN 5-04-612) Soyhulls (IFN 1-O4-560) Molasses (IFN 4-04-560) Ground limestone (IFN 6-02-632) Dynamate c Trace mineral mixture Vitamin ADE mixture d
48.4 14.7 14.0 12.7 6.8 3.0
.23
.10
.05
.02
aThe total diet consisted of 25% corn silage, 25% hay crop silage (alfalfa) and 50% concentrate mix on a dry matter basis.
bDry matter basis.
CDynamate| registered tradename with International Minerals and Chemicals Corporation for feed grade double sulfate of magnesium and potassium.
dprovides 5,200, 2,640 and .5:3 IU of supplemental vitamins A, D and E, respectively, per kg of concentrate mix.
needle holes were sealed with waterproof glue 4. A specific amount of dry feed was weighed into bags, based on calculated dry matter determined on separate aliquots. Bags were closed and tied with 1~ylon string. The rumen suspension device (RSD) consisted of a 9-gauge wire loop (6.8 x 4.2 cm), sandwiched and brazed between four metal washers (4.4 cm diameter), totaling approximately 150 g in weight. At each time interval (1, 2, 4, 8, 12, 24 and 48 h), quadrupli- cate sets of bags were tied to one end of the RSD and .5 m of 13.6-kg-test braided fishline was fastened to the other end. Just before ruminal suspension, bags were soaked in a specific amount of 39 -+ 3 C tap water (1 liter/250 g feed N; Crooker et al., 1978) for 15 rain to remove soluble and(or)59-/~m filterable material. Groups of bags were submersed into the liquid strata of ruminal contents in the mid-ventral region. The fishline was secured to the rumen cannula cover to allow bag movement with contents. An additional bag containing 5 g of cottonseed meal (CSM) ground at 2 mm was used as a control feed to monitor day-to- day variation of lumen function. Cottonseed meal was chosen because it exhibits a linear rate
4Testors; The Testor Corp., Rockford, IL.
of DM disappearance over extended time intervals (100 h), with 15 to 20% residual DM remaining. Test feeds were retested if DM disappearance of CSM varied more than 5% from established standard disappearance curves.
Lactating Holstein cows with ruminal cannulae (10 cm id) were used from 2 to 5 mo after the onset of lactation. Cows were fed a total mixed diet (table 1) consisting of 50:50 forage:concentrate mixture. Animals were fed ad libitum at 500, 1000, 1600 and 2100 h. Bags were rinsed immediately after removal with running tap water and a second time (2 min/set of bags) while manipulating the feed sample residue within bags, until the rinse water was clear. Bags were dried to a constant weight at 55 C. Bags at each time period were emptied, composited and residues were ground (1 mm). Nitrogen analysis was by the macro-Kjeldahl method (AOAC, 1975).
Percentage residual DM and N data were transformed to natural logarithms (log e) and subjected to linear regression vs time. Residues at the 48-h incubation times were used as the undigestible components (DM and N) for each regression and were subtracted from the residue remaining at each ruminal incubation time (Smith et al., 1971). Residual percentages of DM and N at 48 h were less than 3 and 4%, respectively. The following linear regression equation model was used:
by guest on October 7, 2011jas.fass.orgDownloaded from

VARIABLES AFFECTING IN SITU DIGESTION 1349
R = - k t + R0,
where
R = In of percentage residual for DM or Y (Y),
k = digestion rate coefficient (slope), t = time (X),
R0 = residual DM or N at t = 0 (Y inter- cept).
Each regression equation was calculated with and without the zero-hour time point to correct for lag (Mertens and Loften, 1980). Compari- sons for parallelism (Sokal and Rohlf, 1969; Snedecor and Cochran, 1980) revealed no differences (P>.10) in rate constants, so the zero-hour time remained in the regression. Lack-of-fit tests (Sokal and Rohlf, 1969) were- conducted on all regression equations (0 to 24 h) and were nonsignificant for DM and N disappearance, suggesting one linear rate of digestion [Kd -- rate constant (%/h); 1 0 0 x In regression coefficient].
The effects of placing all bags in the rumen at once and removing them at designated time intervals vs introduction of bags in reverse sequence and removing them all at once (i.e., 48-h sample placed in rumen first, 1-h sample last) was investigated. Soybean meal was ground at 2 mm and 5-g (12.6 mg/cm 2) samples were used. Two replications (time series incu- bations) of each procedure were conducted on the same cow to determine the influence on DM and N disappearance rate constants. The latter procedure for bag introduction was used in all subsequent experiments.
Four lactating Holstein cows between 2 and 5 mo postpartum were used to determine the effect of cow and time on CSM DM disappear- ance rates. All cows were fed the same diet for at least 2 wk before, and continuing throughout, the experiment. Duplicate 5-g samples of CSM were incubated (1, 2, 4, 8, 12, 24 and 48 h) for 6 consecutive wk on the same days of each week in each cow.
Soybean meal was used to determine the influence of bacterial N contamination after ruminal exposure and its influence on N disap- pearance rate. Two replications were conducted within the same week with one cow. Sample
s Tetko, Inc., Elmsford, NY.
weights were 5 g/bag with 59-/am porosity bags. Rumen exposure times were those previously described. Diaminopimelic acid (DAPA) analysis (Czerkawski, 1974; el-Shazley and Hungate, 1966) was conducted on composited residues at each fermentation time.
Bacteria were harvested twice (1,000 x g for 10 min followed by centrifugation of the supernatant at 16,000 x g for 20 min) to quantify DAPA. Bacterial N content of the sample residue was calculated from DAPA concentration by the ratio of bacterial N:DAPA in isolated rumen bacteria. Recovery of DAPA was 103.2 -+ 6.1% for 80 /ag added and bac- terial N:DAPA ratio was 34.2 -+ 1.1.
The effect of sample grind, weight and bag porosity on DM and N disappearance were investigated. Samples of soybean meal (SBM) were ground through a 1-, 2- and 5-mm screen in a Wiley Mill, and a fourth size was the unground, commercially processed form. Shown in table 2 are the mean weighted particle sizes of each grind after seiving. Sample weights were to provide 1, 5, 10 and 15 g of DM/bag, with sample DM weight to bag surface area exposures of 2.5, 12.6, 25.3 and 37.9 mg/cm 2.
Bags containing SBM were presoaked as described previously. Upon removal from water, bags were lightly blotted and then dried to a constant weight at 55 C. Three replications of each of the 16 treatments were conducted. Additional bags were prepared to determine the influence of bag porosity on DM and N disap- pearance. Bag porosities were 6, 20, 40, 59, 80 and 102 /~m, according to manufacturers s specifications. Sample weights were 5 g (12.6 mg/cm 2) ground through a 2-mm screen. The presoak procedure was that described pre- viously. Three replications of each of six treatments were conducted.
Sample grinds, feed weights to bag surface area and bag porosities were those used to determine water soluble disappearance. Before ruminal suspension, all bags were presoaked. Ruminal suspension times were 1, 2, 4, 8, 12, 24 and 48 h. Three replications of each treat- ment were conducted.
Comparisons for parallelism (Sokal and Rohlf, 1969 and Snedecor and Cochran, 1980) were made between rate constants for the bag introduction procedure and bacterial N con- tamination experiment. Data for each experi- ment (i.e., effects of cow-time variation, water soluble disappearance, sample grind, sample weight and bag porosity on DM and N disap-
by guest on October 7, 2011jas.fass.orgDownloaded from

1350 NOCEK
TABLE 2. PARTICLE SIZE DISTRIBUTION OF SOYBEAN MEAL AFTER GRINDING
Mean Grind, Sieve size, mm particle mm a 2.36 2.00 1.68 1.18 .841 .590 .420 .297 <.297 b size c, mm
%d
Unground 2 15 17 28 12 11 5 5 5 1.23 5 0 3 7 39 20 16 5 5 5 .99 2 0 0 1 13 41 22 9 6 8 .76 1 0 0 0 1 3 48 20 14 14 .52
aUnground = material as received; 5, 2, 1 mm = ground through a Wiley Mill.
bpercentage of material remaining in the bottom collecting pan.
CCalculated from weighted mean, excluding that portion collected in pan.
dpercentage of material that remained on the specific size sieve.
pearance rate constants) were subjected to split-plot analyses of variance techniques. When main effects were significant, mean differences were tested by Duncan ' s mul t iple range test.
Estimated Ruminal Protein Availability. Disappearance rate constants were applied to the predict ion model of Van Soest et al. (1982). Other models uti l izing in situ digestion rates have been proposed and used to est imate ruminal availability (Orskov and McDonald, 1970; Stern and Satter, 1982; Cummins et al., 1983; and Erdman and Vandersall , 1983). Rumina l available protein (RAP) was calculated using the fol lowing basic equat ion:
RAP = 1 0 0 - ( n
e BA i [kdi/(kd i + kPi)]) , i=3
where
e =
A i =
KPi = Kd i =
sum of (1) soluble, (2) insoluble, po- tent ia l ly digestible and (3) undigest- ible fractions, f ract ional protein concen t ra t ion in feed, a m o u n t of feed dry mat te r consti- tu t ing each fract ion, rate of passage for each fract ion, rate of digestion for each fraction.
Because all regression analysis for SBM demons t ra ted nonsignif icant lack-of-fit, it was assumed that SBM has one insoluble, poten- tially digestible componen t .
Estimates of soluble and 59-/~m filterable N digestion ( K d l ) were obta ined by mul t ip lying the insoluble N disappearance rate constant by 3.0 (Nocek et al., 1983). This fraction was assumed to pass with the l iquid phase of rumen contents . Liquid turnover rate (8.8%/h) was obta ined from Hartnell and Satter (1979) for cows be tween 13 and 24 wk of lactat ion. The value used for turnover rate of SBM in the rumen was 4.9%/h as de termined in vivo by Stern et al. (1983). The rate of undigestible passage was assumed to be the same as the insoluble rate. These l i terature estimates were used because they were de te rmined in cows in the stage of lactat ion that closely approximated those used in the present study.
R um en available-protein values obtained f rom these calculations on t r ea tmen t mean N disappearance rate constants were evaluated with in vivo determinat ions ob ta ined by Santos et al. (1984) for lactating dairy cows.
Results and Discussion
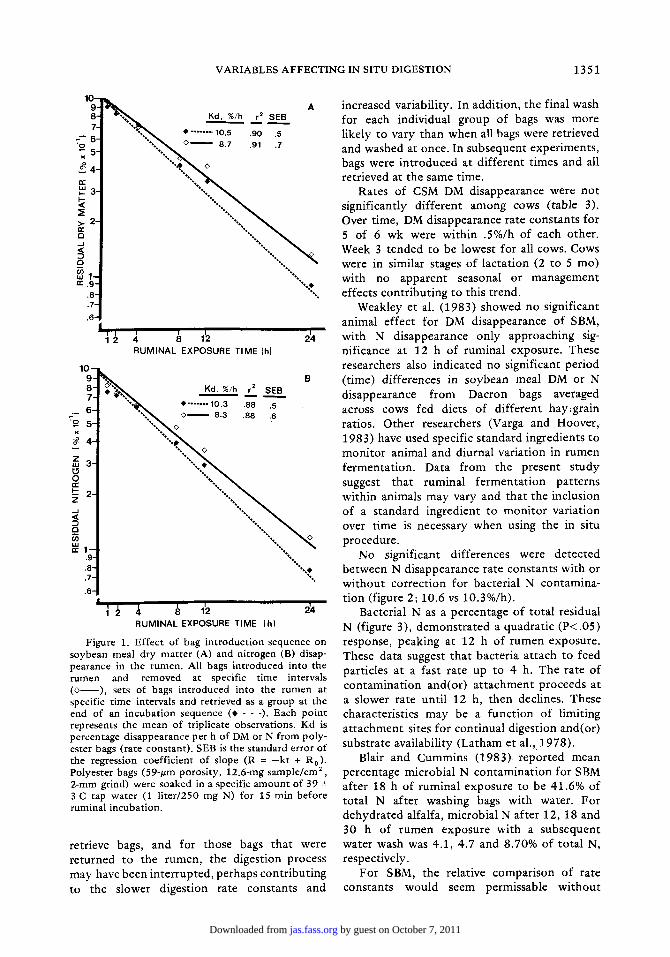
In t roducing bags in the rumen at specific t ime intervals and removing them as a group resulted in a faster (P<.05) rate constant of SBM DM and N disappearance than in t roducing all bags at once and retrieving them at specific subsequent t ime intervals (figures 1A and B). The relative degree of variation wi thin proce- dures was slightly lower for bags retrieved all at once. When bags were in t roduced all at the same time, a t t achment lines became tangled and all bags had to be removed at each incu- ba t ion period. More t ime was required to
by guest on October 7, 2011jas.fass.orgDownloaded from
V A R I A B L E S A F F E C T I N G IN SITU D I G E S T I O N 13 51
1 0 - 9- 8- 7-
~"- 6- 'o x 5-
_~4-
>. 2- a _J
e,-
.8-
.7- .6-
Kd, %/h r 2 SE.~.B A
�9 ........ ~o.s .9o .s
.o.o.o~
' 4 ~, ~ 1'2 2~. RUMINAL EXPOSURE TIME Ihl
= !; .8- .7- .6-
lOq . s 8 t ~ " ~ . Kd, %/h r 2 SEB 7 �9 . . . . ' ~ ~ " ' ~ ....... 10.3 .88 .5
61 "....~ C. 8.3 .88 .6
4- "~
~ %%~176
"..... *~176176
RUMINAL EXPOSURE TIME Ihl
Figure 1. Effect of bag introduction sequence on soybean meal dry matter (A) and nitrogen (B) disap- pearance in the rumen. All bags introduced into the rumen and removed at specific time intervals ( ~ - - ) , sets of bags introduced into the rumen at specific time intervals and retrieved as a group at the end of an incubation sequence (* - - -). Each point represents the mean of triplicate observations. Kd is percentage disappearance per h of DM or N from poly- ester bags (rate constant). SEB is the standard error of the regression coefficient of slope (R = --kt + R0). Polyester bags (59-t~m porosity, 12.6-rag sample/era 2, 2-mm grind) were soaked in a specific amount of 39 -+ 3 C tap water (1 liter/250 mg N) for 15 rain before ruminal incubation.
retr ieve bags, and for those bags that were re turned to the rumen, the digestion process may have been in terrupted, perhaps cont r ibut ing to the slower digest ion rate constants and
increased variabili ty. In addi t ion, the final wash for each individual group of bags was more l ikely to vary than when all bags were retr ieved and washed at once. In subsequent exper iments , bags were in t roduced at d i f fe rent t imes and all retr ieved at the same time.
Rates of CSM DM disappearance were no t significantly d i f ferent among cows (table 3). Over t ime, DM disappearance rate constants for 5 of 6 wk were wi th in .5%/h of each other. Week 3 tended to be lowest for all cows. Cows were in similar stages of lacta t ion (2 to 5 too) wi th no apparent seasonal or managemen t effects cont r ibut ing to this t rend.
Weakley et al. (1983) showed no significant animal ef fec t for DM disappearance of SBM, with N disappearance only approaching sig- nificance at 12 h of ruminal exposure. These researchers also indicated no significant per iod ( t ime) differences in soybean meal DM or N disappearance f rom Dacron bags averaged across cows fed diets o f d i f ferent hay:grain ratios. Other researchers (Varga and Hoover , 1983) have used specific s tandard ingredients to mon i to r animal and diurnal variat ion in rumen fe rmenta t ion . Data f rom the present s tudy suggest that ruminal f e rmen ta t i on pat terns wi th in animals may vary and that the inclusion of a s tandard ingredient to mon i to r var ia t ion over t ime is necessary when using the in situ procedure .
No significant differences were de tec ted be tween N disappearance rate constants wi th or w i thou t correc t ion for bacter ia l N contamina- t ion (figure 2; 10.6 vs 10.3%/h).
Bacterial N as a percentage o f to ta l residual N (figure 3), demons t ra t ed a quadrat ic (P<.05) response, peaking at 12 h o f rumen exposure . These data suggest tha t bacter ia a t tach to feed particles at a fast rate up to 4 h. The rate of con tamina t ion and(or) a t t a chmen t proceeds at a slower rate unt i l 12 h, then declines. These characterist ics may be a func t ion of l imiting a t t achmen t sites for cont inual digest ion and(or) substrate availabili ty (La tham et al . , 1978).
Blair and Cummins (1983). r epor ted mean percentage microbial N con tamina t ion for SBM after 18 h of ruminal exposure to be 41.6% of to ta l N after washing bags wi th water . For dehydra ted alfalfa, microbial N after 12, 18 and 30 h of r umen exposure wi th a subsequent water wash was 4.1, 4.7 and 8.70% of to ta l N, respectively.
For SBM, the relative compar ison of rate constants wou ld seem permissable w i thou t
by guest on October 7, 2011jas.fass.orgDownloaded from

13 5 2 NOCEK
< e~
0
0
r ~ [-,
<
<
Z o
Z o
eo
~4
M
. . . . ~
0
....,
I - I v
+ I
0
.~ V
i .
~ ~
0 ~ *" 0
by guest on October 7, 2011jas.fass.orgDownloaded from

VARIABLES AFFECT ING IN SITU DIGESTION 13 5 3
1 0 ~ - ~ o 9~%.%.. Kd,%/h r Z SEB
7 ~ ~ " . , , ~ " ...... 10.3 .96 .6
- - 6 4 O ~ . , < > - - 10.6 ,g8 .5 "70 5 "%" " . t
Z "%~ �9 EM ~ 0 3- %..
%~176
2- ~176176 _~ %.. ",,., m %.. ~,= ~
1.7_ ".o%..
14 ~ ~ 1~ 2'4 RUMINAL EXPOSURE TIME Ihl
Figure 2. Effec t of bacterial N con tamina t ion on the rate of soybean meal N disappearance. Ni t rogen disappearance w i t h o u t (e - - -) and wi th (0 ) correction for bacterial N. Kd is percentage disappear- ance of N or N corrected for bacterial N from poly- ester bags. SEB is the standard error of the regression coefficient or slope (R = - k t + R0). Polyester bags (59-/am porosity, 12.6-mg sample/era ~ , 2-ram grind) were soaked in a specific amount of 39 -+ 3 C tap water (1 l i ter/250 mg feed N) for 15 min before ruminal incubation.
correction for bacterial N, because they were not different. However, extrapolation to time zero, for quantification of N associated with a particular rate, may be questionable. More
20-
18-
16"
~ 1 4 -
12- j- Z 10- ,-I
8 - ee Lgl
in 4-
2-
; ".... p e r c e n t a g e o f t o t a l " | ....... residual nitrogen
I "[ ./" ........... § § ~ "~176
i ............ .[i I §
+
f RUMINAL EXPOSURE TIME Ihl
Figure 3. Bacterial nitrogen contamination of soy- bean meal at specific t imes after tureen exposure in polyester bag.
t y p e s of feeds need to be investigated to determine the degree of microbial N c o n - t a m i n a t i o n in assoc ia t ion w i t h t he in si tu procedure.
Presoaking did not result in uniform DM disappearance for sample weight and grind. There were interactions (P<.05) between grind and weight for both DM and N disappearance. With the exception of 10-~and 15-g samples at 1-mm grind, DM disappearance tended to decrease w i th increasing particle size (f igure 4A). Clumping of feed was often observed in l-ram grind bags, which became more n o t i c e a b l e as sample weight increased. Other r e s e a r c h e r s (Figroid et al., 1972; Stern et al., 1983) have observed clumping with different feedstuffs. Ehle et al. (1982) demonstrated no effect of
32] gu
_~ 28- e~ ~ -
~ 26-
~ - n 24 -
2 2 -
~20 Lu z o -
~18-
16- .z 8
14- Z
12-
10-
A +,
%% SE = -+ 24 x
~ §
~ "%% O. ~.s ~'S %'%% %. .." \ s.o~- % Sample
~ "~ . %.%. Weightlgl �9 ~ o %%
, ~ ~ " + 5
. . - .." "... x . "*o
01 0 "~176176176176 ,%. "..~ "e10
"015
~ 5 ung~ound SAMPLE GRIND Imml
%%%% SE = -+ 16 B
%% %%
o %% +
..'�9 '.. %.~ Sample s N,.�9149 ~ % Weight igl
',, " , , . ,o o . . . . . ~ \ - , f t .
"\ No '". / " %.+ 5 \ N - t �9 "\ \ - �9 ...-
�9 ~ ~ �9176
i ~ ung~-ound SAMPLE GRIND Imm]
Figure 4. Effect of sample grind and sample weight on soybean meal dry matter (A) and nitrogen (B) dis- appearance from polyester bags (59-~m porosity) soaked for 15 min in 39 C water (1 l i ter/250 mg feed N). Each point is the mean of triplicate replications.
by guest on October 7, 2011jas.fass.orgDownloaded from

1354 NOCEK
32-
30-
Dry Matter o/~ / SE :-'1.3 /
/ 28-
~ - o o j O
7
~ 20- e ~j - Nitrogen j l "
~18- SE = -* 1.8 ~ SSS /
/ /
16- / I /
14- �9 . . . . e- "~ 'S ' ' 'e
g 2'0 dO 5'9 8'0 162. BAG POROSITY luml
Figure 5. Effect of bag porosity on soybean meM dry" matter and nitrogen disappearance from polyester bags (12.6-mg sample/cm 2 , 2-ram grind) soaked for 15 min in 39 C water (1 liter/250 mg feed N). Each point is the mean of triplicate replication.
SBM particle size on DM disappearance from bags, with no systematic effect on N disap- pearance.
Presoaking produced nonuniform N disap- pearance values (figure 4B). Bags containing 1 and 5 g of SBM demonstrated a tendency for decreasing N disappearance as particle size increased (except 5 mm). Bags containing 10 and 15 g tended to decrease, then increase in N disappearance as sample grind increased. The 5-g sample showed a consistent trend for both DM and N disappearance (figure 4A, B). All sample weights (except 15 g) decreased in N disappearance from 1- to 2-mm grind.
There were no differences (P>.10) between pore sizes up to 59 /am for DM and N disap- pearance (figure 5), however, disappearance tended to increase with increased bag pore size. Bag porosities of 80 and 102/am demonstrated more (P<.05) DM disappearance than smaller pore sizes and were not different from each other. Nitrogen washout for 6 and 20/am was lower (P<.05) than that for 80 or 102/am, with 4-0 /am not being different from all others. Lindberg and Knutsson (1981) showed increased loss of SBM DM as bag porosity increased from
10 to 36/am, after soaking bags in water for 6 h. They observed disappearance values con- siderably less than the present study (36 /am, 2.6%), using a 1-g sample weight ground to pass a 1-mm screen with agitation.
A major limitation associated with the bag technique is the inability to characterize ruminal availability of soluble and(or) filterable material that may be degraded to a greater or lesser extent than the insoluble, digestible material (Mahadevan et al., 1980; Nocek et al., 1983). These preruminal disappearance data indicate that even before ruminal exposure, there are several physical characteristics of both feeds and bags that must be considered.
Test for homogeneity of regression profiles indicated sample grind had no effect (P>.10) on DM or N disappearance rates from polyester bags (table 4). These data agree with Ehle et al. (1982), who demonstrated no significant effect of SBM particle size (1, 180, 600, 300 and 150 /am) on DM disappearance using 1-g samples (10.7 mg/cm 2). Weakley et al. (1977) indicated that SBM DM and N degradation was less with a coarse particle size (2,000/am) than with a fine size (500 /am). Presoaking could minimize differences between grinds because large particles may become hydrated and swol- len, allowing more accessibility for microbial attachment and digestion.
Rate constants for DM and N were higher (P<.01) for 1-g compared with all other sample weights. There were no mean differences in rate constants for DM and N among 5-, 10- and 15-g samples; however, those for 5-g samples tended to be consistently higher than for 10- or 15-g samples.
The importance of sample surface area ratio has been several researchers (Uden et
weight to bag emphasized by al., 1974; Van
Hellen and Ellis, 1977). More recently, Varga and Hoover (1983) compared DM disappearance from various concentrates and forage with two bag sizes (9 • 17 and 13 x 21 cm) and two sample weights (25 and 59 g). Dry matter disappearance was depressed by 3 and 8 per- centage units for sample weight to bag surface area ratios of 9.2 vs 20.3 and 18.3 vs 32.7 mg/cm 2 .
Generally, as bag pore size increased, rate constants for SBM DM and N disappearance also increased; however, the increases were mainly associated with two porosity changes, 20 to 40 /am and 59 to 80/am (table 5). Rate constants from bags with pororisites of 6 and
by guest on October 7, 2011jas.fass.orgDownloaded from

VARIABLES AFFECTING IN SITU DIGESTION 13 5 5
M
:!
~
. , ~ . -
Z
. . ~
. - ~
g ~ d N
. ~ .
M M M M
II ,,-.,
~ o ~
by guest on October 7, 2011jas.fass.orgDownloaded from

1356 NOCEK
TABLE 5. THE EFFECT OF BAG POROSITY ON SOYBEAN MEAL DRY MATTER AND NITROGEN DISAPPEARANCE RATES FROM POLYESTER BAGS a
DM N
Porosity, tam Kd b SEB c r 2 d Kd SEB r 2
6 6.1 ef .3 .95 5.2 f .4 .91 20 5.0 f .3 .93 4.2 f .3 .90 40 10.8 g .6 .95 10.2g .7 .93 59 11.0g .5 .94 10.4g .9 .92 80 12.9 h .6 .97 12.8 h .7 .95
102 12.6 h .5 .98 12.2 h .5 .97
apolyester bags (12.6-mg sample/era 2 , 2-mm grind) were soaked in a specific amount of (1 liter/250 mg feed N) for 15 min before ruminal incubation.
bKd is percentage disappearance per h of DM or N from polyester bags (rate constant).
CSEB = Standard error of regression coefficient or slope (R = --kt + R 0 ).
dr2 = Coefficient of determination.
eEach value is the mean of triplicate incubation replications in time for the same cow.
f'g'hMean Kd in the same column that do not have a common superscript differ (P<.05).
39 + 3 C tap water
20 # m were s lower ( P < . 0 1 ) t h a n those of o t h e r sizes, and n o t d i f f e r en t b e t w e e n each o the r . Ra te c o n s t a n t s o b t a i n e d wi th bag poros i t i es of 40 and 5 9 / ~ m were s lower ( P < . 0 5 ) t h a n 80 and 102 gtm. O t h e r researchers (Uden et al., 1974 ; L indbe rg and Varv ikko , 1982) have d e m o n - s t r a t ed s imilar f indings.
R a t e of d iges t ion is an impor t an t ' c r i t e r ion for d e t e r m i n i n g o p t i m u m c o m b i n a t i o n s of phys ica l f ac to r s to use for in situ experi- m e n t a t i o n , b u t b o t h depressed and s t i m u l a t e d ra tes m a y be equal ly e r roneous . A m o s t logical
c o n s i d e r a t i o n would be to select cr i ter ia t h a t wou ld p red ic t in vivo rumina l d iges t ion m o s t accura te ly . However , very few in vivo es t imates of R A P fo r SBM in l ac ta t ing cows have b e e n r epor t ed . San tos et al. ( 1 9 8 4 ) indica tes in vivo RAP fo r SBM was 70 .5% for lac ta t ing cows. O t h e r in vivo s tudies for ca t t le show ranges f r o m 63 to 79% (Po t t e r e t al., 1971; Kropp e t al., 1 9 7 7 ; Merchen et at., 1979 ; Loe rch e t al., 1983; Weakley and Owens , 1983) . Calcula ted es t imates of RAP for SBM sub jec ted to d i f f e ren t in si tu var iables are s h o w n in tab le 6.
TABLE 6. ESTIMATION OF IN SITU SOYBEAN MEAL RUMINAL AVAILABLE PROTEIN a AS INFLUENCED BY SAMPLE GRIND, FEED TO BAG SURFACE AREA RATIO
AND BAG POROSITY
m m
Feed weight to bag Grind surface area ratio Porosity
Estimated Estimated Estimated RAP, % mg/cm 2 RAP, % tam RAP, %
1 72 2.5 79 6 53 2 70 12.6 70 20 48 5 71 25.3 65 40 69 Unground 71 37.9 65 59 72
80 73 102 73
n aEstimated by the method of Van 8oest et al. (1982), rumen available protein (RAP) = 100 - ( i_e3_ BA i
[kdi/(kd i + kPi)] ), where e -- sum of (1) soluble, (2) insoluble, potentially digestible and (3) undigestible frac- tions. Santos et al. (1984) in vivo estimate of ruminal protein availability for soybean meal in lactating cows = 70.5%.
by guest on October 7, 2011jas.fass.orgDownloaded from

VARIABLES AFFECTING IN SITU DIGESTION 1357
Estimated RAP for different SBM grinds was similar, and within 1 to 2 percentage units of the in vivo estimate by Santos et al. (1984). A feed to bag surface area ratio of 2.5 mg/cm 2 overestimated and ratios of 25.3 and 37.9 mg/cm 2 underestimated in vivo RAP. A ratio of 12.6 mg/cm 2 resulted in a RAP that most closely approximated in vivo. It is realized that there may be quality differences between the SBM used by Santos et al. (1984) and the present trial. Ultimately, size of bags and sample weight will be dictated by the number and type of analyses one desires to conduct on the residue, and density of the feed ingredient.
Bag porosities of 6 and 20 /am underesti- mated, whereas porosity of 40, 59, 80 and 102 /am compared favorably with in vivo RAP. Weakley et al. (1983) concluded that bag materials of small pore (5 /am) size limit influx of digesting agents, regardless of restriction on efflux of digested residues. Van Hellen and Ellis (1974) tested porosities of 1.2, 5 and 10 /am and reported no significant influx of neutral detergent fiber (NDF) into bags after 48 h of rumen exposure. However, there was a signifi- cant influx of NDF into bags with porosities of 100/am. These researchers recommended a pore size of no larger than 10 /am. It may be that different pore sizes are necessary, depending on the specific ingredient and nutrient being investigated.
Under the conditions and criteria used in this study, the following conclusions can be made for in situ digestion variables and tech- niques involving SBM or like ingredients.
1) Bag Introduction: Ruminal introduction of bags in reverse order (i.e., 100 h first, 1 h last) decreased variation for in situ digestion profiles.
2) Cow and Time Variation: Use of a standard ingredient (CSM) detected time-related variation for in situ diges- t ion profiles.
3) Bacterial N Contamination: Did not affect N digestion rate constant.
4) Presoaking: Disappearance of soluble DM and N are affected by particle size, sample weight to bag surface area and bag porosity.
5) Particle Size: Grinds of 1, 2, 5 mm, or unground, had no influence on DM or N digestion rate constants.
6) Feed Weight to Bag Surface Area Ratio: 12.6 mg/cm 2 compared most favorably
with literature in vivo estimates for ruminal N digestion.
7) Pore Size: 40 to 102 /am were similar and higher in estimated ruminal protein availability than smaller pore sizes, and compared more favorably with in vivo literature estimates.
Literature Cited
Akin, D. E. and H. E. Amos. 1975. Rumen bacterial degradation of forage cell walls investigated by electron microscopy. Appl. Microbiol. 5:692.
AOAC. 1975. Official Methods of Analysis (12th Ed.). Association of Official Analytical Chemists, Washington, DC.
Blair, L. and K. A. Cummins. 1983. Microbial nitrogen contamination of feedstuffs contained in poly- ester bag. J. Dairy Sci. 66(Suppl. 1):189.
Crooker, B. A., C. J. Sniffen, W. H. Hoover and L. L. Johnson. 1978. Solvents for soluble nitrogen measurements in feedstuffs. J. Dairy Sci. 61:437.
Cummins, K. A., J. E. Nocek, C. E. Polan and J. H. Herbein. 1983. Nitrogen degradability and microbial protein synthesis in calves fed diets of varying degradability defined by the bag tech- nique. J. Dairy Sci. 66:2356.
Czerkawski, J. W. 1974. Methods for determining 2-6-diaminopimelic acid and 2-aminoethylphos- phoric acid in gut contents. J. Food Agr. Sci. 23:45.
Ehle, F. R., M. R. Murphy and J. H. Clark. 1982. In situ particle size reduction and the effect of particle size on degradation of crude protein and dry matter in the rumen of dairy steers. J. Dairy Sci. 65:963.
el-Shazly, K. and R. E. Hungate. 1966. Method for measuring diaminopimelic acid in total rumen contents and its application to the estimation of bacterial growth. AppL Microbiol. 14:27.
Erdman, R. A. and H. J. Vandersall. 1983. Effect of rumen protein degradability on milk yield of cows in early lactation. J. Dairy Sci. 66:1873.
Figroid, W., W. H. Hale and B. Theurer. 1972. An evaluation of the nylon bag technique for esti- mating tureen utilization of grains. J. Anim. Sci. 35:113.
HarmeU, G. F. and L. D. Satter. 1979. Determination of rumen fill, retention time and ruminal turn- over rates of ingesta at different stages of lacta- tion in dairy cows. J. Anim. Sci. 48:381.
Kropp, J. R., R. R. Johnson, J. R. Males and F. N. Owens. 1977. Microbial protein synthesis with low quality roughage rations: Isonitrogenous substitution of urea for soybean meal. J. Anim. Sci. 46:837.
Latharn, M. J., B. E. Brooker, G. L. Pettipher and P. J. Harris. 1978. Ruminococcus flavefaciens cell coat and adhesion to cotton cellulose and to cell walls in leaves of perennial ryegrass (Lolium perenne). Appl. Environ. Microbiol. 1:156.
Lindberg, J. E. and P. G. Knutsson. 1981. The effect of bag pore size on the loss of particulate matter and on degradation of cell wall fibre. Agr. En- vironm. 6:171.
Lindberg, J. E. and T. Varvikko. 1982. The effect of
by guest on October 7, 2011jas.fass.orgDownloaded from

1358 NOCEK
bag pore size on ruminal degradation of dry matter, nitrogenous compounds and cell walls in nylon bags. Swedish J. Agr. Res. 12:163.
Loerch, S. C., L. L. Berger, S. D. Plegge and G. R. Fahey, Jr. 1983. Digestibility and rumen escape of soybean meal, blood meal, meat and bone meal and dehydrated alfalfa nitrogen. J. Anim. Sci. 57:1037.
Mahadevan, S., J. D. Erfle and F. D. Sauer. 1980. Degradation of soluble and insoluble proteins by Bacteroides amylophilus protease and by rumen microorganisms. J. Anim. Sci. 50:723.
Merchen, N., T. Hanson and T. Klopfenstein. 1979. Ruminal bypass of brewers dried grains protein. J. Anita. Sci. 49:192.
Mertens, D. R. and J. R. Loften. 1980. The effect of starch on forage fiber digestion kinetics in vitro. J. Dairy Sci. 63:1437.
Nocek, J. E., C. E. Polan and J. H. Herbein. 1983. Total amino acid release rates of soluble and insoluble protein fractions of concentrate feed- stuffs by Streptomyces griseus. J. Dairy Sci. 66:1663.
Orskov, E. R. and I. McDonald. 1970. The estimation of protein degradability in the rumen from incubation measurements weighed according to rate of passage. J. Agr. Sci. (Camb.) 92:499.
Potter, C. D., C. O. Little, N. W. Bradley and G. E. Mitchell. 1971. Abomasal nitrogen in steers fed soybean meal, urea, or urea and two levels of molasses. J. Anim. Sci. 32:531.
Santos, K. A., M. D. Stern and L. D. Satter. 1984. Protein degradation in the rumen and amino acid absorption in the small intestine of lactating dairy cattle fed various protein sources. J. Anim. Sci. 58:244.
Smith, L. W., H.K. Goering, D. R. Waldo and D. H. Gordon. 1971. In vitro digestion rate of forage cell wall components. J. Dairy Sci. 54:71.
Snedecor, G. W. and W. G. Cochran. 1980. Analysis of covariance, comparison of regression lines. In:
Statistical Methods (7th Ed.). pp 385--388. Iowa State Univ. Press, Ames.
Sokal, R. F. and F. J. Rohlf. 1969. Comparisons of regression lines. In: Biometry. pp 448-458. W. H. Freeman and Co., San Francisco, CA.
Stern, M. D., M. E. Ortega and L. D. Satter. 1983. Retention time in rumen and degradation of protein supplements fed to lactating dairy cattle. J. Dairy Sci. 66:1264.
Stern, M. D. and L. D. Satter. 1982. In vivo estimation of protein degradability in the rumen. F. N. Owens (Ed.) Protein Requirements for Cattle: Symposium p 57. MP-109, Oklahoma State Univ., Stillwater.
Uden, P., R. Parra and P. J. Van Soest. 1974. Factors influencing reliability of the nylon bag technique. J. Dairy Sci. 57:622.
Van Hellen, R. W. and W. C. Ellis. 1977. Sample container porosities for rumen in situ studies. J. Anita. Sci. 44:141.
Van Soest, P. J., C. J. Sniffen, D. R. Mertens, D. G. Fox, P. H. Robenson and V. Krishnamoorthy. 1982. A net protein system for cattle: The rumen sub- model for nitrogen. In: F. N. Owens (Ed.) Protein Requirements for Cattle: Symposium. pp 265- 279. MP-109, Oklahoma State Univ., Stillwater.
Varga, G. A. and W. H. Hoover. 1983. Rate and extent of neutral detergent fiber degradation of feed- stuffs in situ. J. Dairy Sci. 66:2109.
Weakley, D. C. and F. N. Owens. 1983. Influence of roughage level on soybean meal degradation and microbial protein synthesis in the rumen. J. Anim. Sei. 57(Suppl. 1):474.
Weakley, D. C., F. N. Owens, D. G. Heath and B. J. Shockey. 1977. Particle size and soybean meal value for ruminants. J. Anita. Sci. 45(Suppl. 1):268.
Weakley, D. C., M. D. Stern and L. D. Satter. 1983. Factors affecting disappearance of feedtusffs from bags suspended in the rumen. J. Anim. Sci. 56:493.
by guest on October 7, 2011jas.fass.orgDownloaded from

Citations
eshttp://jas.fass.org/content/60/5/1347#otherarticlHighWire-hosted articles: This article has been cited by 3
by guest on October 7, 2011jas.fass.orgDownloaded from




![البوابة الإجتماعية : استقبال · 2019. 5. 21. · 2019 30 2019 37 j + 1985 5 1985 12 1985 5 1985 12 30 2019 37 12 2019 30 5 1985 2019 2019 37 (1 €3.1£'.4]1985](https://img.pdfslide.net/doc/110x75/5fedd4702235b618ec02783f/-2019-5-21-2019-30-2019.jpg)