Embed Size (px)
Citation preview

a PATTERNS & PHENOTYPES
Notch and Delta mRNAs in Early-Stage andMid-Stage Drosophila Embryos ExhibitComplementary Patterns ofProtein-Producing PotentialsAndrew Shepherd,1 Uma Wesley,2 and Cedric Wesley2*
Notch and Delta proteins generate Notch signaling that specifies cell fates during animal development.There is an intriguing phenomenon in Drosophila embryogenesis that has not received much attentionand whose significance to embryogenesis is unknown. Notch and Delta mRNAs expressed in early-stageembryos are shorter than their counterparts in mid-stage embryos. We show here that the difference insizes is due to mRNA 30 processing at alternate polyadenylation sites. While the early-stage Notch mRNAhas a lower protein-producing potential than the mid-stage Notch mRNA, the early-stage Delta mRNA hasa higher protein-producing potential than the mid-stage Delta mRNA. Our data can explain the comple-mentary patterns of Notch and Delta protein levels in early- and mid-stage embryos. Our data also raisethe possibility that the manner and regulation of Notch signaling change in the course of embryogenesisand that this change is effected by 30 UTR and mRNA 30 processing factors. Developmental Dynamics239:1220–1233, 2010. VC 2010 Wiley-Liss, Inc.
Key words: Notch; Delta; 30 UTR; mRNA; polyadenylation
Accepted 25 January 2010
INTRODUCTION
During oogenesis in Drosophila mela-
nogaster, the mother packs her eggs
with mRNA and protein products
required for early events in embryo-
genesis. These events include, in
order of occurrence, specification of
germ cells and somatic cells, estab-
lishment of the primary embryonic
axes and coordinates, gastrulation
and the establishment of the three ba-
sic germ layers, initiation of segmen-
tation, and the formation of proneural
clusters from which the nervous sys-
tem arises (Nakao and Campos-Ortega, 1996; Campos-Ortega andHartenstein, 1997; Cabrera, 1990;Brennan et al., 1997; Rusconi andCorbin, 1998; Morel et al., 2003;LeComte et al., 2006; http://flybase.org). The zygote might also contributeto these events, as zygotic transcrip-tion is known to initiate before 1 hrAEL (Liang et al., 2008; de Renziset al., 2007; Lecuyer et al., 2007)although it becomes the primarysource of gene expression only byabout 4 hr after egg laying (h AEL) atroom temperature (22�C). The last
event that is primarily under mater-nal control, proneural cluster forma-tion, is completed just before 4 hrAEL. Therefore, we will refer to 0–4-hr AEL embryos as early-stageembryos. Lateral inhibition, the pro-cess that specifies neuronal and epi-dermal precursor cells within eachproneural cluster, is initiated at about4 hr AEL. This process and all subse-quent embryogenesis events are pri-marily under the control of zygoticproducts (Nakao and Campos-Ortega,1996; Campos-Ortega, and Harten-stein, 1997; Cabrera, 1990; Brennan
Dev
elop
men
tal D
ynam
ics
Additional Supporting Information may be found in the online version of this article.1Department of Biology, University of Vermont, Burlington, Vermont2Department of Microbiology and Molecular Genetics, University of Vermont, Burlington, VermontGrant sponsor: NIH (NINDS); Grant number: NS43122; Grant sponsor: Neuroscience COBRE; Grant number: P20 RR016435; Grantsponsor: University of Vermont College of Medicine Bridge.*Correspondence to: Cedric Wesley, Department of Microbiology and Molecular Genetics, 322 Stafford Hall, 95 CarriganDrive, The University of Vermont, Burlington, VT 05405. E-mail: [email protected]
DOI 10.1002/dvdy.22262Published online 3 March 2010 in Wiley InterScience (www.interscience.wiley.com).
DEVELOPMENTAL DYNAMICS 239:1220–1233, 2010
VC 2010 Wiley-Liss, Inc.

et al., 1997; Rusconi and Corbin,1998; LeComte et al., 2006; http://fly-base.org). We will refer to embryosolder than 4-hr AEL but youngerthan 16-hr AEL, in which lateral inhi-bition and parallel or subsequentmajor events are taking place, as mid-stage embryos. As some maternalproducts persist well beyond 4-hrAEL, it is possible they also contrib-ute to developmental regulation inmid-stage embryos (Liang et al. 2008;de Renzis et al. 2007; Lecuyer et al.,2007).
Notch signaling is a basic, evolutio-narily conserved developmental path-way. It regulates developmentalevents based on inter-cellular commu-nication. The critical components ofthe Notch signaling pathway are thecell surface receptor Notch (N) and itscell surface–anchored ligand Delta(Dl). When Dl that is expressed onone cell binds N that is expressed onthe neighboring cell, the N intracellu-lar domain (Nintra) is proteolyticallycleaved and translocated to the nu-cleus where, in association with thetranscription factor Suppressor ofHairless, Nintra activates the tran-scription of target genes such as theEnhancer of split Complex (E(spl)C)genes. Cells with a low level of N sig-naling and a high level of Dl activitycommit to one fate or developmentalprogram (often the default choice)while cells with a high level of N sig-naling and a low level of Dl activitycommit to the alternate fate or devel-opmental program. After commit-ment, the cells require N signaling formaintenance of their fate or program.This process is repeated in all differ-entiation events at all stages of devel-opment (Shellenbarger and Mohler,1978; Cabrera, 1990; Heitzler andSimpson, 1991; Fortini et al., 1993;Struhl et al., 1993; Lieber et al., 1993;Rusconi and Corbin, 1998; Artavanis-Tsakonas et al., 1999; Mumm andKopan, 2000; Brou et al., 2000; Lieberet al., 2002; Schweisguth, 2004; Ahi-mou et al., 2004; LeComte et al.,2006).
Many genetic screens and studieshave identified RNA-binding proteinsas important regulators of N signal-ing (Norga et al., 2003; Kankel et al.,2007; Penn and Schedl, 2007; Okabeet al., 2001; Okano et al., 2002). Anumber of these proteins target
mRNA 30 processing or 30 UTRs. Oneexample is the Drosophila poly(A) po-lymerase (PAP) gene hiiragi that isresponsible for the addition of poly(A)tail to mRNAs (Juge et al., 2002; Mur-ata et al., 2001). Another example isthe hephaestus gene encoding Poly-pyrimidine Tract Binding Protein(PTBP), which regulates mRNA 30
processing (Dansereau et al., 2002).In general, proteins and sequencesinvolved in mRNA 30 processing and30 UTR-based regulation determine ifor how much mRNA is processed atthe nearby poly(A) site and how muchprotein is produced from the mRNA.A poly(A) site is composed of the con-sensus hexamer (AAUAAA) or thenon-consensus hexamer (such asUAUAAA, AUUAAA, or AAUAUA),the Downstream Sequence Element(DSE), Upstream Sequence Elements(USEs), and the cleavage site. Whenthese sequences emerge behind thePol II transcription complex, the basiccomponents of mRNA 30 processing(some of which are associated withthe transcription complex) bind theircognate sequences and form the func-tional complex that cleaves themRNA at the cleavage site and addsthe poly(A) tail to the cleaved mRNA.The poly(A) tail is stabilized and themRNA exported to the cytoplasm fortranslation. In general, USEs deter-mine whether the mRNA 30 process-ing complex forms at a poly(A) siteand the DSE determines the effi-ciency of cleavage and polyadenyl-ation at this site. USEs continue toplay important roles in mRNA stabil-ity, nuclear export, cytoplasmic poly-adenylation (if involved), and transla-tion efficiency (Wells et al., 1998;Wilusz et al., 2001a, b; Li et al., 2001;Soller and White, 2001; Lejeune et al.,2002; Reed and Hurt, 2002; Wickenset al., 2002; Kornblihtt et al., 2004;Bentley 2005; Beaudoing et al., 2007).
mRNA 30 processing and poly-adenylation is required for most if notall genes. However, genes like hiiragithat encode the poly(A) polymeraseshow a special relationship to N sig-naling so much so that mutant alleleseven phenocopy classic N-signalingphenotypes (Murata et al., 2001). Theroles of mRNA 30 processing genesrecovered in N signaling–basedgenetic screens are unknown exceptfor mushashi, which is known to
repress translation of Numb, a nega-tive regulator of N signaling (Okabeet al., 2001; Okano et al., 2002).Others are known just as negativeregulators (for example, hephaestusor sex lethal; Dansereau et al., 2002;Penn and Schedl, 2007) or positiveregulators (for example, hiiragi; Mur-ata et al., 2001). It is not knownwhether these genes function inde-pendently or as part of a specific regu-latory mechanism.We report here evidence for an
mRNA 30 processing-based mecha-nism that regulates N signaling atthe earliest stages of development.We show that N and Dl mRNAs inearly-stage embryos and in mid-stageembryos are processed at alternativepoly(A) sites. The resultant shorter orlonger mRNAs have different protein-producing potentials, which mightexplain the pattern of N and Dl pro-tein expression in these embryos. Ourdata raise the possibility that themanner in which N signaling is acti-vated and regulated changes duringembryogenesis and this change islikely effected through 30 UTR-bind-ing and -processing factors that targetalternative poly(A) sites in the N andDl mRNAs.
RESULTS
Differences in N and Dl
Protein Levels Are
Correlated With Differences
in mRNA sizes
N and Dl mRNAs expressed in 0–4-hrAEL embryos are shorter than theircounterparts expressed in 4–12-hrAEL embryos (Fig. 1). The longer Nand Dl mRNAs are the predominantmRNAs at all stages from 4- to 16-hrAEL by which time the major embryo-genesis events are completed (datanot shown). Others have alsoobserved the shorter Dl mRNA inearly-stage embryos, as early as �1-hr AEL (Vassin et al., 1987; Kopczyn-ski et al., 1988; Alton et al., 1989;Haenlin et al., 1990). The shorter NmRNA is also expressed at very earlystages (see Supp. Fig. S1, which isavailable online; please refer to theExperimental Procedures section fordistribution of embryonic stageswithin each of our samples). We willrefer to the longer N mRNA by its
Dev
elop
men
tal D
ynam
ics
NOTCH AND DELTA mRNAs IN EMBRYOGENESIS 1221
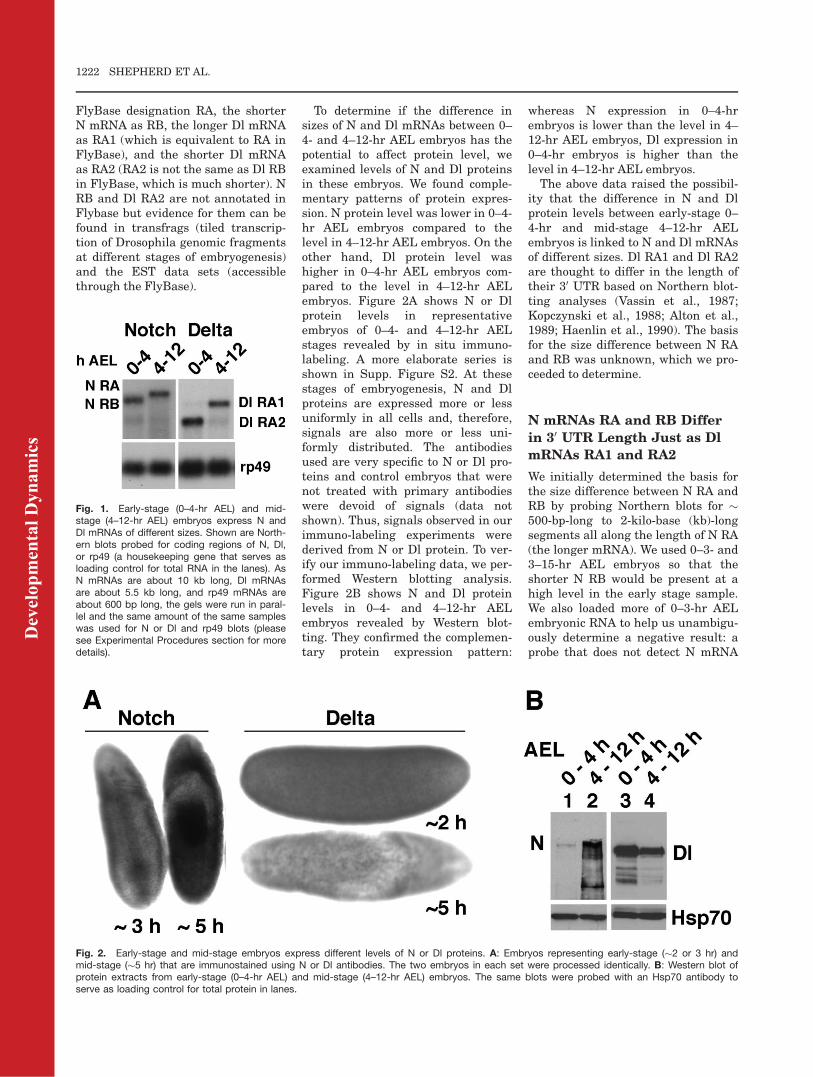
FlyBase designation RA, the shorterN mRNA as RB, the longer Dl mRNAas RA1 (which is equivalent to RA inFlyBase), and the shorter Dl mRNAas RA2 (RA2 is not the same as Dl RBin FlyBase, which is much shorter). NRB and Dl RA2 are not annotated inFlybase but evidence for them can befound in transfrags (tiled transcrip-tion of Drosophila genomic fragmentsat different stages of embryogenesis)and the EST data sets (accessiblethrough the FlyBase).
To determine if the difference insizes of N and Dl mRNAs between 0–4- and 4–12-hr AEL embryos has thepotential to affect protein level, weexamined levels of N and Dl proteinsin these embryos. We found comple-mentary patterns of protein expres-sion. N protein level was lower in 0–4-hr AEL embryos compared to thelevel in 4–12-hr AEL embryos. On theother hand, Dl protein level washigher in 0–4-hr AEL embryos com-pared to the level in 4–12-hr AELembryos. Figure 2A shows N or Dlprotein levels in representativeembryos of 0–4- and 4–12-hr AELstages revealed by in situ immuno-labeling. A more elaborate series isshown in Supp. Figure S2. At thesestages of embryogenesis, N and Dlproteins are expressed more or lessuniformly in all cells and, therefore,signals are also more or less uni-formly distributed. The antibodiesused are very specific to N or Dl pro-teins and control embryos that werenot treated with primary antibodieswere devoid of signals (data notshown). Thus, signals observed in ourimmuno-labeling experiments werederived from N or Dl protein. To ver-ify our immuno-labeling data, we per-formed Western blotting analysis.Figure 2B shows N and Dl proteinlevels in 0–4- and 4–12-hr AELembryos revealed by Western blot-ting. They confirmed the complemen-tary protein expression pattern:
whereas N expression in 0–4-hrembryos is lower than the level in 4–12-hr AEL embryos, Dl expression in0–4-hr embryos is higher than thelevel in 4–12-hr AEL embryos.The above data raised the possibil-
ity that the difference in N and Dlprotein levels between early-stage 0–4-hr and mid-stage 4–12-hr AELembryos is linked to N and Dl mRNAsof different sizes. Dl RA1 and Dl RA2are thought to differ in the length oftheir 30 UTR based on Northern blot-ting analyses (Vassin et al., 1987;Kopczynski et al., 1988; Alton et al.,1989; Haenlin et al., 1990). The basisfor the size difference between N RAand RB was unknown, which we pro-ceeded to determine.
N mRNAs RA and RB Differ
in 30 UTR Length Just as Dl
mRNAs RA1 and RA2
We initially determined the basis forthe size difference between N RA andRB by probing Northern blots for �500-bp-long to 2-kilo-base (kb)-longsegments all along the length of N RA(the longer mRNA). We used 0–3- and3–15-hr AEL embryos so that theshorter N RB would be present at ahigh level in the early stage sample.We also loaded more of 0–3-hr AELembryonic RNA to help us unambigu-ously determine a negative result: aprobe that does not detect N mRNA
Fig. 1. Early-stage (0–4-hr AEL) and mid-stage (4–12-hr AEL) embryos express N andDl mRNAs of different sizes. Shown are North-ern blots probed for coding regions of N, Dl,or rp49 (a housekeeping gene that serves asloading control for total RNA in the lanes). AsN mRNAs are about 10 kb long, Dl mRNAsare about 5.5 kb long, and rp49 mRNAs areabout 600 bp long, the gels were run in paral-lel and the same amount of the same sampleswas used for N or Dl and rp49 blots (pleasesee Experimental Procedures section for moredetails).
Fig. 2. Early-stage and mid-stage embryos express different levels of N or Dl proteins. A: Embryos representing early-stage (�2 or 3 hr) andmid-stage (�5 hr) that are immunostained using N or Dl antibodies. The two embryos in each set were processed identically. B: Western blot ofprotein extracts from early-stage (0–4-hr AEL) and mid-stage (4–12-hr AEL) embryos. The same blots were probed with an Hsp70 antibody toserve as loading control for total protein in lanes.
Dev
elop
men
tal D
ynam
ics
1222 SHEPHERD ET AL.
RB. Figure 3 shows a portion of ouranalyses indicating that N RA andRB differ in the length of the 30 UTR:N RB was detected by probes to the 50
UTR and the coding region but not byprobes to the terminal (term) portionof the 30 UTR. These data suggestedthat N RA and RB differed in thelength of 30 UTR, with N RB lackingmore than 500 bp of the terminal por-tion of the 30 UTR.
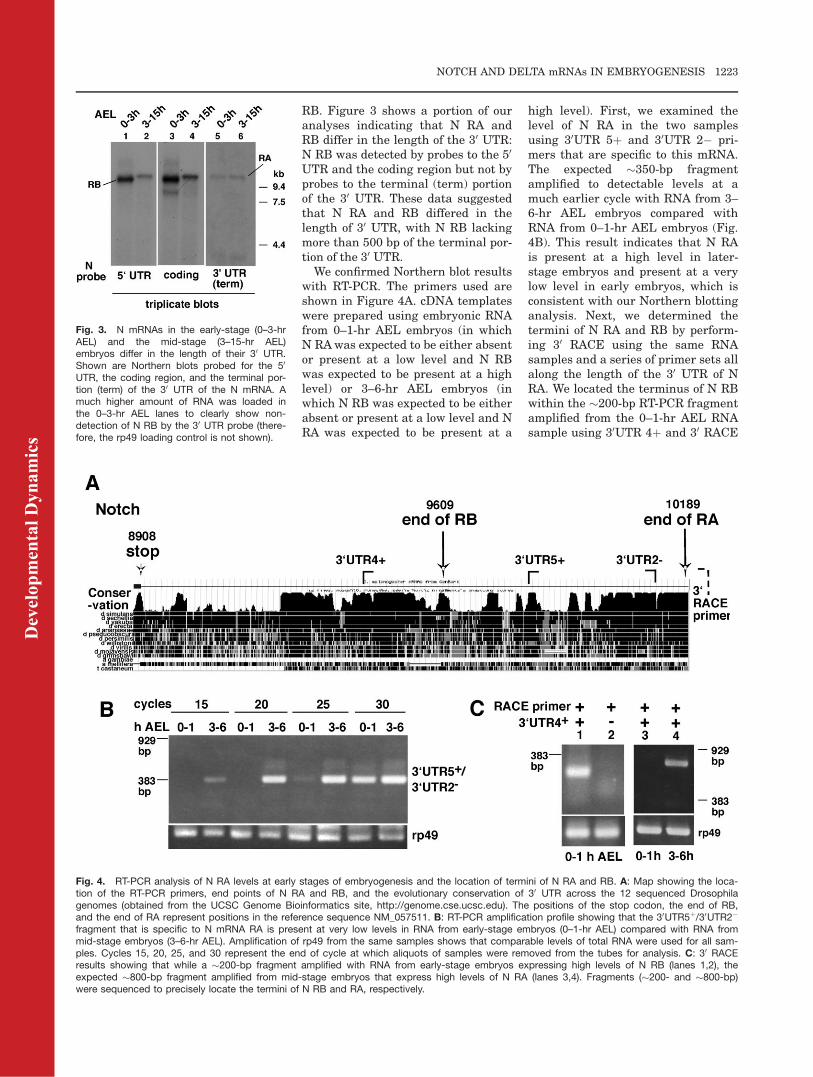
We confirmed Northern blot resultswith RT-PCR. The primers used areshown in Figure 4A. cDNA templateswere prepared using embryonic RNAfrom 0–1-hr AEL embryos (in whichN RAwas expected to be either absentor present at a low level and N RBwas expected to be present at a highlevel) or 3–6-hr AEL embryos (inwhich N RB was expected to be eitherabsent or present at a low level and NRA was expected to be present at a
high level). First, we examined thelevel of N RA in the two samplesusing 30UTR 5þ and 30UTR 2� pri-mers that are specific to this mRNA.The expected �350-bp fragmentamplified to detectable levels at amuch earlier cycle with RNA from 3–6-hr AEL embryos compared withRNA from 0–1-hr AEL embryos (Fig.4B). This result indicates that N RAis present at a high level in later-stage embryos and present at a verylow level in early embryos, which isconsistent with our Northern blottinganalysis. Next, we determined thetermini of N RA and RB by perform-ing 30 RACE using the same RNAsamples and a series of primer sets allalong the length of the 30 UTR of NRA. We located the terminus of N RBwithin the �200-bp RT-PCR fragmentamplified from the 0–1-hr AEL RNAsample using 30UTR 4þ and 30 RACE
Fig. 3. N mRNAs in the early-stage (0–3-hrAEL) and the mid-stage (3–15-hr AEL)embryos differ in the length of their 30 UTR.Shown are Northern blots probed for the 50
UTR, the coding region, and the terminal por-tion (term) of the 30 UTR of the N mRNA. Amuch higher amount of RNA was loaded inthe 0–3-hr AEL lanes to clearly show non-detection of N RB by the 30 UTR probe (there-fore, the rp49 loading control is not shown).
Fig. 4. RT-PCR analysis of N RA levels at early stages of embryogenesis and the location of termini of N RA and RB. A: Map showing the loca-tion of the RT-PCR primers, end points of N RA and RB, and the evolutionary conservation of 30 UTR across the 12 sequenced Drosophilagenomes (obtained from the UCSC Genome Bioinformatics site, http://genome.cse.ucsc.edu). The positions of the stop codon, the end of RB,and the end of RA represent positions in the reference sequence NM_057511. B: RT-PCR amplification profile showing that the 30UTR5þ/30UTR2�
fragment that is specific to N mRNA RA is present at very low levels in RNA from early-stage embryos (0–1-hr AEL) compared with RNA frommid-stage embryos (3–6-hr AEL). Amplification of rp49 from the same samples shows that comparable levels of total RNA were used for all sam-ples. Cycles 15, 20, 25, and 30 represent the end of cycle at which aliquots of samples were removed from the tubes for analysis. C: 30 RACEresults showing that while a �200-bp fragment amplified with RNA from early-stage embryos expressing high levels of N RB (lanes 1,2), theexpected �800-bp fragment amplified from mid-stage embryos that express high levels of N RA (lanes 3,4). Fragments (�200- and �800-bp)were sequenced to precisely locate the termini of N RB and RA, respectively.
Dev
elop
men
tal D
ynam
ics
NOTCH AND DELTA mRNAs IN EMBRYOGENESIS 1223
primers (Fig. 4C, lanes 1–2). Thesame primers amplified an �800-bpfragment from the 3–6-hr AEL RNAsample, which was expected due tothe high expression of N RA in thissample (Fig. 4C, lanes 3–4). Wesequenced the �200- and �800-bpfragments and found that N RB endsafter a non-consensus poly(A) sitehexamer UAUAAA, about 570 basesupstream of the end of N RA near theconsensus poly(A) site hexamerAAUAAA. Figure 4A shows the loca-tion of the two poly(A) sites andsequence conservation across the 12sequenced Drosophila genomes (Kar-
olchik et al., 2003; http://genome.cse.ucsc.edu). The actual sequencethat is conserved across the two Npoly(A) sites is shown in Supp. Fig-ures S3 and S4. We find that thepoly(A) sites and a large number of 30
UTR sequence elements are highlyevolutionarily conserved suggestingtheir functional significance.
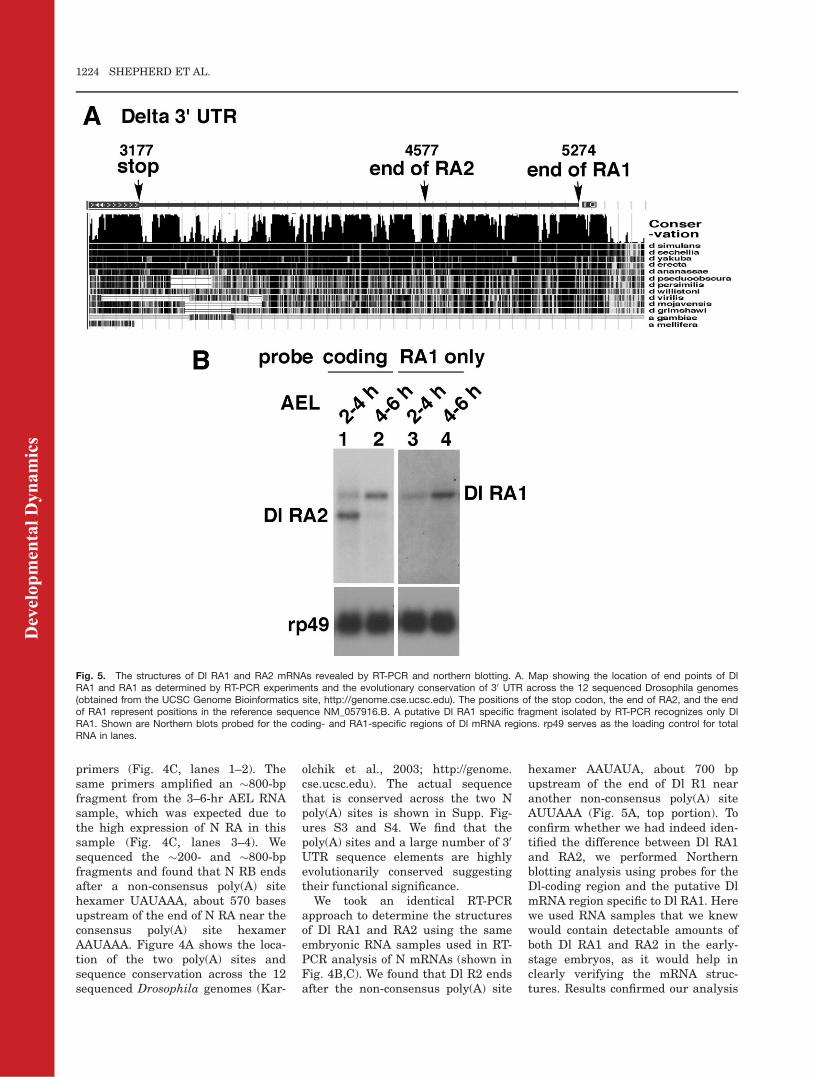
We took an identical RT-PCRapproach to determine the structuresof Dl RA1 and RA2 using the sameembryonic RNA samples used in RT-PCR analysis of N mRNAs (shown inFig. 4B,C). We found that Dl R2 endsafter the non-consensus poly(A) site
hexamer AAUAUA, about 700 bpupstream of the end of Dl R1 nearanother non-consensus poly(A) siteAUUAAA (Fig. 5A, top portion). Toconfirm whether we had indeed iden-tified the difference between Dl RA1and RA2, we performed Northernblotting analysis using probes for theDl-coding region and the putative DlmRNA region specific to Dl RA1. Herewe used RNA samples that we knewwould contain detectable amounts ofboth Dl RA1 and RA2 in the early-stage embryos, as it would help inclearly verifying the mRNA struc-tures. Results confirmed our analysis
Fig. 5. The structures of Dl RA1 and RA2 mRNAs revealed by RT-PCR and northern blotting. A. Map showing the location of end points of DlRA1 and RA1 as determined by RT-PCR experiments and the evolutionary conservation of 30 UTR across the 12 sequenced Drosophila genomes(obtained from the UCSC Genome Bioinformatics site, http://genome.cse.ucsc.edu). The positions of the stop codon, the end of RA2, and the endof RA1 represent positions in the reference sequence NM_057916.B. A putative Dl RA1 specific fragment isolated by RT-PCR recognizes only DlRA1. Shown are Northern blots probed for the coding- and RA1-specific regions of Dl mRNA regions. rp49 serves as the loading control for totalRNA in lanes.
Dev
elop
men
tal D
ynam
ics
1224 SHEPHERD ET AL.

(and those of others) that Dl RA1 andRA2 differ in the length of 30 UTR,with the latter missing about a thirdin the 30 most terminal portion (Fig.5B). Just like N 30 UTR, Dl 30 UTRincludes highly evolutionarily con-served sequence elements spanningthe two poly(A) sites as well as manyregions upstream and downstream ofthese poly(A) sites (Fig. 5A, bottomportion). The actual sequence that isconserved across the two Dl poly(A)sites is shown in Supplementary Fig-ures 5 and 6.
N mRNA RB and Dl mRNA
RA1 Have Lower Protein-
Producing Potential
Our Northern blotting and RT-PCRdata indicated that N mRNA in 0–4-hr AEL embryos is processed at apoly(A) site with a non-consensushexamer to yield N RB and N mRNAin 4–15-hr AEL embryos is processedat a poly(A) site with the consensushexamer to yield N RA. Since the con-sensus hexamer-containing poly(A)site has high affinity for the mRNA30-processing complex, N RA wasexpected to produce more proteinthan N RB. If that were the case, itcould explain the higher level ofNotch protein in 4–15-hr AELembryos compared to the level in 0–4-hr AEL embryos. Since both Dl RA1and RA2 are processed at poly(A) siteswith non-consensus hexamers, therewas no a priori expectation that onewould produce more protein than theother. However, the higher level ofDelta protein in 0–4-hr AEL embryoscompared to the level in 4–15-hr AELembryos suggested that Dl RA2 wouldproduce more protein than Dl RA1.
We directly tested protein-produc-ing potentials of different N and DlmRNAs in cultured Drosophila S2cells. These cells are ideal for the pur-pose because almost all cells in fliesexpress N and Dl and there is strongdevelopmental feedback regulation orcross-regulation by N signaling orother pathways. These regulationswould confound interpretation of ex-perimental results. Drosophila-cul-tured S2 cells do not express endoge-nous N or Dl but can be made toexpress these proteins. Such S2-Nand S2-Dl cells are excellent model
systems for studying the molecularand biochemical aspects of N and Dlfunction as they reproduce all N sig-naling aspects observed in flies. S2cells show minimal, if any, activitiesof other pathways interacting with Nsignaling and minimal feedback regu-lation (Fehon et al., 1990; Klueget al., 1999; Wesley and Saez, 2000;Mishra-Gorur et al., 2002; Wesley andMok, 2003; Ahimou et al., 2004; Bar-dot et al., 2005, Mok et al., 2005 arebut a few examples). The goal of ourexperiments was to determine the ba-sic activity of N RA relative to N RBand of Dl RA1 relative to Dl RA2. Weencountered dominance of the distalN mRNA RA poly(A) site, which wasexpected due to the presence of theconsensus hexamer. To properlyassess the protein-producing potentialof the proximal N RB poly(A) site(with the non-consensus hexamer),we simply deleted the N RA poly(A)site and USEs specific to it. With Dl,we encountered dominance of theproximal RA2 poly(A) site forcing usto resort to mutagenesis of this site’shexamer for assessment of the distalRA1 poly(A) site.
We generated N constructs that con-tained the 50 UTR and the codingsequence with either (1) the complete30UTR corresponding to N RA plus�100 bp of adjacent 30 sequence thatincluded the DSE or (2) the 30 UTR cor-responding to N RB plus �100 bp ofthe adjacent downstream sequence.These constructs were expressedusing the heat-shock promoter inorder to assess expression at differenttime points after a single induction,which would indicate relative stabil-ities of mRNA. The constructs weretransfected into S2 cells. We extractedDNA from these cells, used them totransfect bacteria, and assessed thenumber of bacterial colony-formingunits. We found the transfection effi-ciency to be the same with the differ-ent constructs (data not shown).Experiments with these transfectedS2 cells showed that N RA produces amuch higher amount of the full-lengthN protein than N RB (Fig. 6A, lanes 1–4). This difference was not due to anysignificant difference in the stability ofthe two N mRNAs as their levels weremore or less comparable (Fig. 6A,lanes 5–8, top). Total protein and RNAused in lanes 1–4 and 5–8, respec-
tively, were extracted from the samepopulation of cells and, therefore, canbe directly compared. A probe directedagainst the terminal 30 UTR detectedonly N RA and not N RB, which lacksthe sequence, verifying the expressionof predicted mRNAs in each lane (Fig.6A, lanes 5–8, middle). The lowamount of N protein in NRB lanes wasnot due to ligand-independent N sig-naling–related turnover as the expres-sion of the N-signaling target gene inS2 cells, E(spl)C m3 (Mishra-Goruret al., 2002), was lower in sampleswith N RB compared to samples withN RA (Fig. 6A, lanes 5–8, bottom).E(spl)C m3 expression is generallyabove the background level in N-expressing S2 cells because of the lowlevel of Nintra production even in theabsence of ligands; this level increasesupon treatment with S2-Dl cells (Mis-hra-Gorur et al., 2002; Wesley andMok, 2003; Mok et al., 2005). These S2cell data indicate that N RA has ahigher protein-producing potentialthan N RB, which is consistent withembryo data showing that the level ofN protein is high when N RA is pre-dominant and low when N RB ispredominant.For analysis of the relative activ-
ities of the Dl mRNAs, we mutatedthe proximal RA2 poly(A) siteAAUAUA to AAGAGA, which wasexpected to suppress mRNA 30 proc-essing at this site. We created con-structs with the Dl 50 UTR, the codingregion, and the full-length 30 UTRplus �300-bp downstream region con-taining the DSE that was either (1)wild type or (2) with the mutatedproximal RA2 poly(A) site. We clonedthese constructs downstream of theheat-shock promoter and performedexperiments just as described for N.Northern blot analysis showed thatthe wild type proximal Dl RA2 poly(A)site is dominant as we barely detectedDl transcripts processed at the distalDl RA1 poly(A) site (Fig. 6B lane 1).The construct with the mutated proxi-mal RA2 poly(A) site suppressedmRNA 30processing at this site andpromoted processing at the distalRA1 poly(A) site (Fig. 6B, lane 2). Thesignal from Dl RA2 in lane 1 was com-parable to the combined signals fromDl RA1 and RA2 in lane 2. Westernblotting analysis showed that thesamples in which Dl mRNA was
Dev
elop
men
tal D
ynam
ics
NOTCH AND DELTA mRNAs IN EMBRYOGENESIS 1225

predominantly processed at the proxi-mal RA2 poly(A) site contained ahigher level of Dl protein comparedwith the samples in which the DlmRNA was processed predominantlyat the distal RA1 poly(A) site (Fig. 6B,lanes 3,4). Thus, our S2 cell experi-ments indicate that the early-stage NRB has a lower protein-producingpotential than the mid-stage N RAand that early-stage Dl RA2 has ahigher protein-producing potentialthan the mid-stage Dl RA1.
The Two N and Dl poly(A)
Sites Are Bona Fide
Alternative Polyadenylation
Sites
With N, the weaker poly(A) site islocated upstream of the strongerpoly(A) site. As a consequence, the
weaker poly(A) site would emergefirst behind the transcription complexand would present the opportunity forprocessing before the emergence ofthe stronger poly(A) site. If the twopoly(A) sites are indeed alternatepolyadenylation sites, we predictedtwo transcripts from constructs con-taining the full-length N 30 UTR andsome downstream sequence (toinclude the DSE). Furthermore, if thetwo poly(A) sites are bona fide alter-nate polyadenylation sites, we pre-dicted that the full-length N 30 UTRand the adjacent downstream regionwould be sufficient for directingmRNA processing at the upstreampoly(A) site to yield RB-like transcriptand the downstream poly(A) site toyield the RA-like transcript.
To test our predictions, we placedthe full-length N 30 UTR and about
100 bases downstream sequence nextto a GFP reporter–coding sequence.This GFP-N30UTR fragment wascloned into a pBluescript vector down-stream of the act5C promoter toobtain the actGFP-N30UTR construct.We sequenced the GFP-N30UTR partand determined there were nosequence errors. The actGFP-N30UTRconstruct was transfected into S2 cellsand Northern analysis performed.The same RNA sample was probed forthe GFP-coding sequence, the full-length N 30 UTR, and the N 30 UTRterminus specific to N RA. Results areshown in Figure 7. Aliquots of totalRNA from the same untransfected S2cells sample were loaded in lanes 1, 4,and 7. Aliquots of total RNA from thesame actGFP-N30UTR sample wereloaded in lanes 2, 5, and 8. Aliquotscontaining a lower amount of totalRNA from another actGFP-N30UTRsample were loaded in lanes 3, 6, and9. Different amounts of two actGFP-N30UTR samples are loaded to shownot only the robustness of transcriptproduction but also properly resolvedbands. Blots from the top of the gel tothe bottom are shown to indicate thestrength of mRNA 30 processing. Asexpected, there is no signal from anyof the probes in lanes with untrans-fected S2 cells RNA (Fig. 7, lanes 1, 4,7). The GFP probe showed a strongband of the size expected from thetranscript ending near the end ofNotch RA, �2,150 bp, and a weakerband of the size expected from thetranscript ending near the end ofNotch RB, �1,550 bp (Fig. 7, lanes 1–3). As expected, both of these bandswere detected by the full-length N30UTR probe but only the larger onewas detected by the N 30UTR probespecific to N RA (Fig. 7, lanes 4–9).Additional smeary bands observedwith the GFP probe (marked withdouble asterisk) are products ofincomplete extension (terminatingbefore the 30UTR) commonly observedwith the over-expression of any genein S2 cells or in flies. The faint �4-kbsignal (marked with an asterisk)derives from the low level of plasmidDNA that is normally recovered fromtotal RNA extractions of transfectedS2 cells (we avoided poly(A) selectionas it might bias recovery if RA andRB transcripts carry poly(A) tails ofdifferent lengths). As is apparent, 30
Fig. 6. N RB and Dl RA1 have a lower protein-producing potential than N RA and Dl RA2,respectively. A: Western blots and Northern blots showing the levels of the N protein (lanes 1–4),N RA and RB (lanes 5–8, top two panels) and E(spl)C m3 mRNA (lanes 5–8, bottom panel) in S2cells transfected with constructs expressing N RA or N RB. The general structure of the plasmidused in these experiments is shown below the blots. Two time points after the heat shock (hs)induction of expression of the constructs are shown (30 min and 3 hr). The bottom panel of lanes1–4 shows the expression of Hsp70 that serves as a loading control for total protein; the bottom-most panel of lanes 5–8 shows the expression of rp49 that serves as a loading control for totalRNA. E(spl)C m3 is an endogenous N-signaling target gene in S2 cells (Mishra-Gorur et al., 2002).Samples in lanes 1 and 5, 2 and 6, 3 and 7, and 4 and 8 were obtained from the same cells. B:Northern and Western blots showing the expression of Dl mRNAs from the Dl gene with the wild-type Dl 30 UTR (wt) or Dl 30 UTR with mutated RA2 poly(A) site hexamer (RA2 mut). The generalstructure of the plasmid used in these experiments is shown below the blots. RA2 mut constructindicate the translation efficiency of Dl mRNA RA1 poly(A) site. rp49 and Hsp70 levels on thesame blots served as loading controls for total RNA and protein, respectively.D
evel
opm
enta
l Dyn
amic
s
1226 SHEPHERD ET AL.
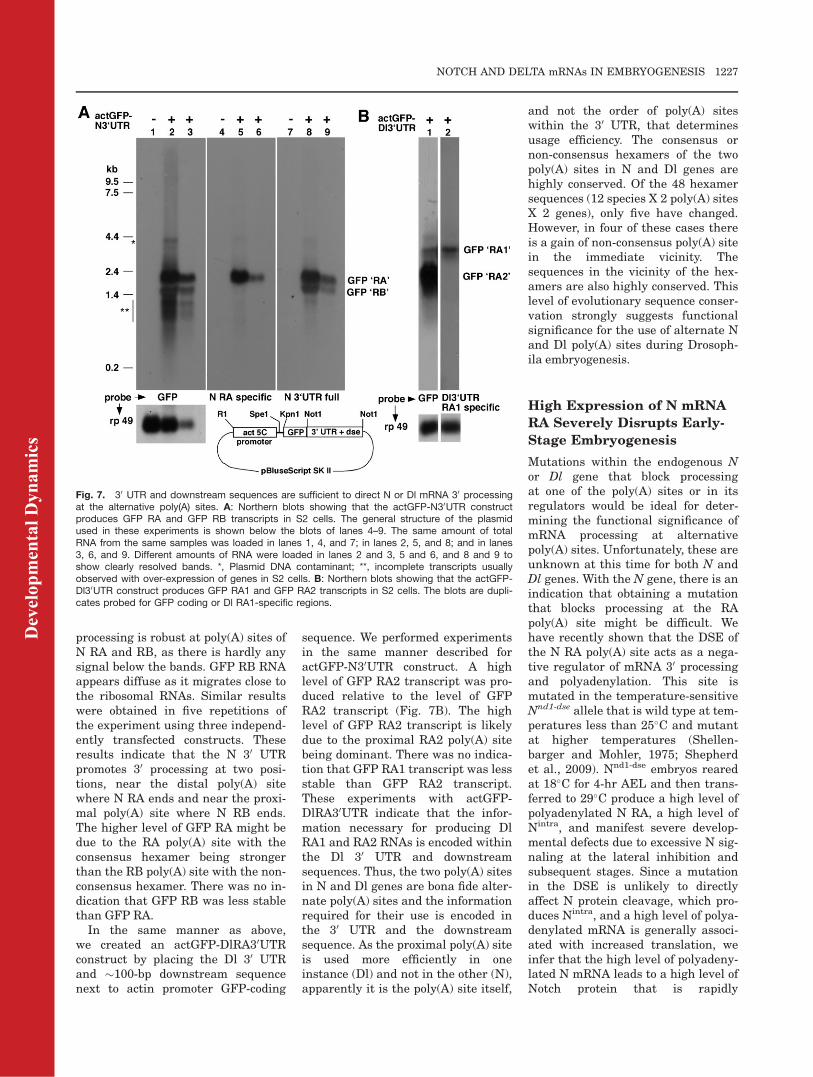
processing is robust at poly(A) sites ofN RA and RB, as there is hardly anysignal below the bands. GFP RB RNAappears diffuse as it migrates close tothe ribosomal RNAs. Similar resultswere obtained in five repetitions ofthe experiment using three independ-ently transfected constructs. Theseresults indicate that the N 30 UTRpromotes 30 processing at two posi-tions, near the distal poly(A) sitewhere N RA ends and near the proxi-mal poly(A) site where N RB ends.The higher level of GFP RA might bedue to the RA poly(A) site with theconsensus hexamer being strongerthan the RB poly(A) site with the non-consensus hexamer. There was no in-dication that GFP RB was less stablethan GFP RA.
In the same manner as above,we created an actGFP-DlRA30UTRconstruct by placing the Dl 30 UTRand �100-bp downstream sequencenext to actin promoter GFP-coding
sequence. We performed experimentsin the same manner described foractGFP-N30UTR construct. A highlevel of GFP RA2 transcript was pro-duced relative to the level of GFPRA2 transcript (Fig. 7B). The highlevel of GFP RA2 transcript is likelydue to the proximal RA2 poly(A) sitebeing dominant. There was no indica-tion that GFP RA1 transcript was lessstable than GFP RA2 transcript.These experiments with actGFP-DlRA30UTR indicate that the infor-mation necessary for producing DlRA1 and RA2 RNAs is encoded withinthe Dl 30 UTR and downstreamsequences. Thus, the two poly(A) sitesin N and Dl genes are bona fide alter-nate poly(A) sites and the informationrequired for their use is encoded inthe 30 UTR and the downstreamsequence. As the proximal poly(A) siteis used more efficiently in oneinstance (Dl) and not in the other (N),apparently it is the poly(A) site itself,
and not the order of poly(A) siteswithin the 30 UTR, that determinesusage efficiency. The consensus ornon-consensus hexamers of the twopoly(A) sites in N and Dl genes arehighly conserved. Of the 48 hexamersequences (12 species X 2 poly(A) sitesX 2 genes), only five have changed.However, in four of these cases thereis a gain of non-consensus poly(A) sitein the immediate vicinity. Thesequences in the vicinity of the hex-amers are also highly conserved. Thislevel of evolutionary sequence conser-vation strongly suggests functionalsignificance for the use of alternate Nand Dl poly(A) sites during Drosoph-ila embryogenesis.
High Expression of N mRNA
RA Severely Disrupts Early-
Stage Embryogenesis
Mutations within the endogenous Nor Dl gene that block processingat one of the poly(A) sites or in itsregulators would be ideal for deter-mining the functional significance ofmRNA processing at alternativepoly(A) sites. Unfortunately, these areunknown at this time for both N andDl genes. With the N gene, there is anindication that obtaining a mutationthat blocks processing at the RApoly(A) site might be difficult. Wehave recently shown that the DSE ofthe N RA poly(A) site acts as a nega-tive regulator of mRNA 30 processingand polyadenylation. This site ismutated in the temperature-sensitiveNnd1-dse allele that is wild type at tem-peratures less than 25�C and mutantat higher temperatures (Shellen-barger and Mohler, 1975; Shepherdet al., 2009). Nnd1-dse embryos rearedat 18�C for 4-hr AEL and then trans-ferred to 29�C produce a high level ofpolyadenylated N RA, a high level ofNintra, and manifest severe develop-mental defects due to excessive N sig-naling at the lateral inhibition andsubsequent stages. Since a mutationin the DSE is unlikely to directlyaffect N protein cleavage, which pro-duces Nintra, and a high level of polya-denylated mRNA is generally associ-ated with increased translation, weinfer that the high level of polyadeny-lated N mRNA leads to a high level ofNotch protein that is rapidly
Fig. 7. 30 UTR and downstream sequences are sufficient to direct N or Dl mRNA 30 processingat the alternative poly(A) sites. A: Northern blots showing that the actGFP-N30UTR constructproduces GFP RA and GFP RB transcripts in S2 cells. The general structure of the plasmidused in these experiments is shown below the blots of lanes 4–9. The same amount of totalRNA from the same samples was loaded in lanes 1, 4, and 7; in lanes 2, 5, and 8; and in lanes3, 6, and 9. Different amounts of RNA were loaded in lanes 2 and 3, 5 and 6, and 8 and 9 toshow clearly resolved bands. *, Plasmid DNA contaminant; **, incomplete transcripts usuallyobserved with over-expression of genes in S2 cells. B: Northern blots showing that the actGFP-Dl30UTR construct produces GFP RA1 and GFP RA2 transcripts in S2 cells. The blots are dupli-cates probed for GFP coding or Dl RA1-specific regions.
Dev
elop
men
tal D
ynam
ics
NOTCH AND DELTA mRNAs IN EMBRYOGENESIS 1227
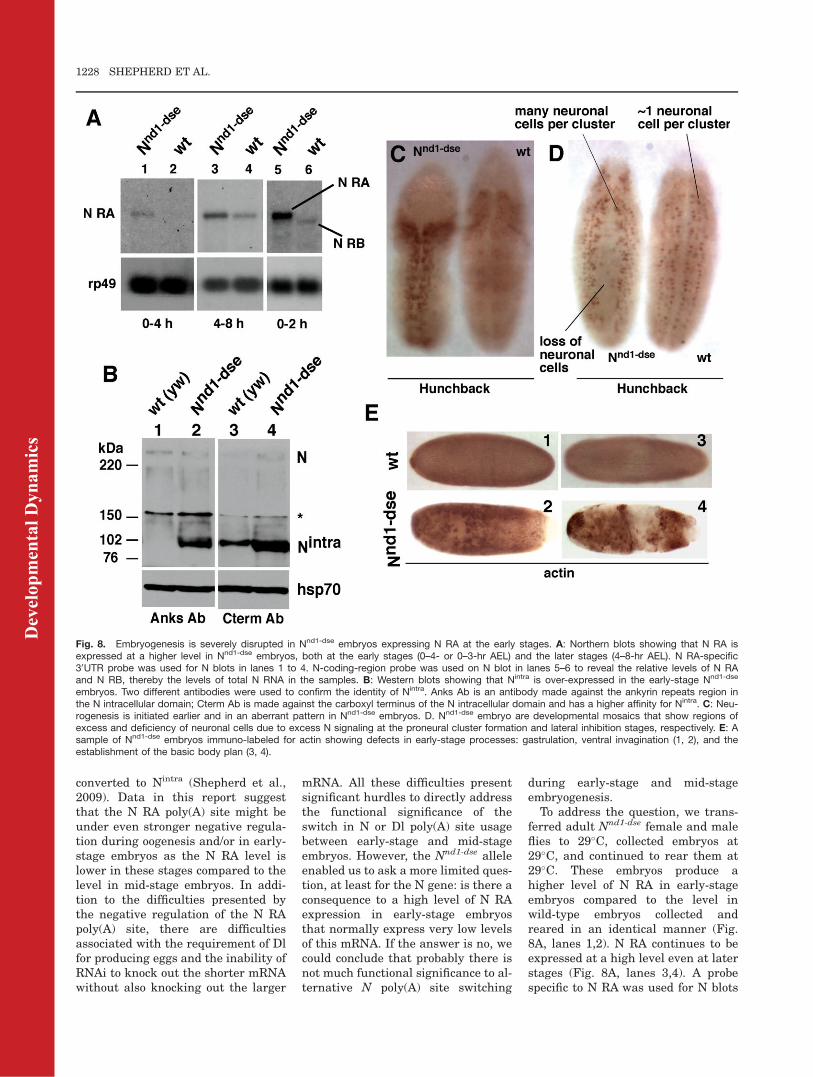
converted to Nintra (Shepherd et al.,2009). Data in this report suggestthat the N RA poly(A) site might beunder even stronger negative regula-tion during oogenesis and/or in early-stage embryos as the N RA level islower in these stages compared to thelevel in mid-stage embryos. In addi-tion to the difficulties presented bythe negative regulation of the N RApoly(A) site, there are difficultiesassociated with the requirement of Dlfor producing eggs and the inability ofRNAi to knock out the shorter mRNAwithout also knocking out the larger
mRNA. All these difficulties presentsignificant hurdles to directly addressthe functional significance of theswitch in N or Dl poly(A) site usagebetween early-stage and mid-stageembryos. However, the Nnd1-dse alleleenabled us to ask a more limited ques-tion, at least for the N gene: is there aconsequence to a high level of N RAexpression in early-stage embryosthat normally express very low levelsof this mRNA. If the answer is no, wecould conclude that probably there isnot much functional significance to al-ternative N poly(A) site switching
during early-stage and mid-stageembryogenesis.To address the question, we trans-
ferred adult Nnd1-dse female and maleflies to 29�C, collected embryos at29�C, and continued to rear them at29�C. These embryos produce ahigher level of N RA in early-stageembryos compared to the level inwild-type embryos collected andreared in an identical manner (Fig.8A, lanes 1,2). N RA continues to beexpressed at a high level even at laterstages (Fig. 8A, lanes 3,4). A probespecific to N RA was used for N blots
Fig. 8. Embryogenesis is severely disrupted in Nnd1-dse embryos expressing N RA at the early stages. A: Northern blots showing that N RA isexpressed at a higher level in Nnd1-dse embryos, both at the early stages (0–4- or 0–3-hr AEL) and the later stages (4–8-hr AEL). N RA-specific30UTR probe was used for N blots in lanes 1 to 4. N-coding-region probe was used on N blot in lanes 5–6 to reveal the relative levels of N RAand N RB, thereby the levels of total N RNA in the samples. B: Western blots showing that Nintra is over-expressed in the early-stage Nnd1-dse
embryos. Two different antibodies were used to confirm the identity of Nintra. Anks Ab is an antibody made against the ankyrin repeats region inthe N intracellular domain; Cterm Ab is made against the carboxyl terminus of the N intracellular domain and has a higher affinity for Nintra. C: Neu-rogenesis is initiated earlier and in an aberrant pattern in Nnd1-dse embryos. D. Nnd1-dse embryo are developmental mosaics that show regions ofexcess and deficiency of neuronal cells due to excess N signaling at the proneural cluster formation and lateral inhibition stages, respectively. E: Asample of Nnd1-dse embryos immuno-labeled for actin showing defects in early-stage processes: gastrulation, ventral invagination (1, 2), and theestablishment of the basic body plan (3, 4).
Dev
elop
men
tal D
ynam
ics
1228 SHEPHERD ET AL.

in lanes 1–4 (Fig. 8). N-coding region-specific probe showed that the over-expressed N mRNA in Nnd1-dse
embryos is primarily N RA (Fig. 8A,lanes 5,6). Western blotting analysisusing antibodies specific to theankyrin repeats (Anks Ab; Lieberet al., 1993) and the carboxyl termi-nus (Cterm Ab; LeComte et al., 2006)of the N-intracellular domain showedthat Nintra accumulates to a high levelin 0–4-hr AEL Nnd1-dse embryos (Fig.8B). The Cterm Ab has a higher affin-ity for Nintra and it clearly reveals therelative levels of Nintra in wild-typeand Nnd1-dse embryos. We do notobserve a significant accumulation ofthe full-length N protein, possibly dueits rapid conversion to Nintra (Shep-herd et al., 2009). Nintra continues tobe expressed at high levels even inmid-stage Nnd1-dse embryos (Shepherdet al., 2009).
When we examined developmentalevents taking place in 0–4-hr AELNnd1-dse embryos, we found many ofthem to be severely disrupted. Forexample, immuno-staining with theneuronal cell marker Hunchbackshowed that in Nnd1-dse embryos, neu-rogenesis is precociously initiated, ex-cessive, and aberrantly patterned(Fig. 8C). This is the stage at whichthe proneural clusters are just begin-ning to form in wild-type embryos(Cabrera, 1990; Rusconi and Corbin,1998). Soon after, we find develop-mental mosaics with some regionsmanifesting excess neuronal cells andother regions manifesting loss (Fig.8D). Other early-stage processes suchas germ-cell specification and gastru-lation were also disrupted (data notshown). Over all, early-stage events(taking place in 0–4-hr AEL embryos)were severely disrupted in 57.02% ofNnd1-dse embryos compared with9.02% of wild-type embryos at 29�C (n¼ 1,000). In a sizeable fraction ofthese embryos, it was impossible toeven characterize the defects. A sam-ple of such Nnd1-dse embryos and com-parable-stage wild-type embryos isshown in Figure 8E.
All early-stage (0–4-hr AEL) mu-tant phenotypes described above areinferred to be due to the Nnd1-dse allelebased on the following observations.One, Nnd1-dse allele was recently re-isolated and the procedure employedwould have randomized �80% of the
genomic (autosomal) background(Shepherd et al., 2009). Two, Nnd1-dse
is a temperature-sensitive allele thatis wild type at the permissive temper-ature (<25�C) and mutant at the re-strictive temperatures (>27�C).Accordingly, we did not observe early-stage mutant phenotypes when Nnd1-
dse parental flies and embryos werereared at the permissive temperature.Three, we did not observe early-stagemutant phenotypes when Nnd1-dse pa-rental flies and embryos under 3-hrAEL were reared at the permissivetemperature. When the embryos wereshifted to the restrictive temperatureafter 3-hr AEL, mutant phenotypesrelated to lateral inhibition and sub-sequent processes were observed inNnd1-dse embryos (Shepherd et al.,2009). Four, when Nnd1-dse embryoswere reared continuously at the re-strictive temperature, the few adultNnd1-dse flies that emerged displayedall the classic N phenotypes, such aswing notching and thick veins, andnone recognizable as belonging toother pathways. While the classic Nphenotypes might appear to be due tothe loss of N function, they might bedue to the opposite N function at anearlier stage, comparable to the mosaicphenotypes observed in embryos(shown in Fig. 8C,D). Mosaic pheno-types appearing to be consistent withboth gain and loss of N signaling havealso been reported during wing devel-opment at larval and pupal stages ofNnd1-dse flies (Royet et al., 1998). Five,the basis for the mutant phenotypeslinked to the Nnd1-dse allele has beengenetically mapped to the N locus andlocalized to the 30 UTR and down-stream sequence of the N gene (pleasesee extended discussion of this matterin Shepherd et al., 2009). These fiveobservations strongly support our in-ference that the phenotypes weobserve in early-stage (0–4-hr AEL)embryos are due to theNnd1-dse allele.
Our data showing that a high levelof N RA expression in early-stageembryos is associated with severe dis-ruption of embryogenesis suggeststhat there might be functional signifi-cance to why the N RA poly(A) site isstrongly suppressed in early-stageembryos. We emphasize that this ismerely a suggestion, as there are cav-eats regarding the Nnd1-dse allele. Asmentioned above, we have inferred
that the increased level of N RA inNnd1-dse embryos leads to the increasedlevel of Nintra based on current knowl-edge of mRNA 30 processing and trans-lation. We cannot rule out the effect ofNintra, which is independent of NRA orvice versa. In addition,Nnd1-dse/wt het-erozygote embryos are morphologi-cally wild type (data not shown), indi-cating that Nnd1-dse behaves as arecessive allele when paired with thewild-type allele. We do not know whybut it is possible that the wild type al-lele in Nnd1-dse/þ embryos facilitatescompensatory negative regulation asit contains the wild-type DSE requiredfor negative regulation of N RAexpression and activity (Shepherdet al., 2009). Although the high level ofNintra in Nnd1-dse embryos (at the re-strictive temperature) argues in favorof this allele being a gain-of-functionallele when on its own, the early-stageembryogenesis phenotypes are noteasily classifiable as gain or lossof function. Thus, the genetic behaviorof Nnd1-dse is complex, which makesinterpretations difficult. Complex be-havior is not unusual for an N allelebut Nnd1-dse is different than otherwell-known complex alleles such assome Abruptex alleles and the N60g11
allele. The latter alleles behave asdominant gain-of-function alleles inthe presence of the wild-type N allelebut as loss-of-function alleles whenhomozygous (Xu et al., 1990; Brennanet al., 1997).
DISCUSSION
Our data show that during Drosoph-ila embryogenesis N and Dl mRNAsare processed at evolutionarily con-served alternative poly(A) sites thatcontain either the consensus hexamer(N RA) or one of the non-consensushexamers (N RB, Dl RA1, or Dl RA2).Early-stage embryos (0–4-hr AEL)and mid-stage embryos (>4-hr AEL)prefer one site or the other. N and DlmRNA processed at the two poly(A)sites have different protein-producingpotentials. The early-stage N RB andthe mid-stage Dl RA1 produce lessprotein than the mid-stage N RA andthe early-stage Dl RA2, respectively.The expression pattern of thesemRNAs and their protein-producingpotentials could explain the differencein N and Dl protein levels between
Dev
elop
men
tal D
ynam
ics
NOTCH AND DELTA mRNAs IN EMBRYOGENESIS 1229

early-stage (0–4-hr AEL) and mid-stage (>4-hr AEL) embryos.
The core components of N signalingsuch as N and Dl are both tightly andfinely regulated. Many mechanismshave been previously described thatfunction at the level of the protein ortranscription (Heitzler and Simpson,1991; Kooh et al., 1993; Irvine andWeischaus, 1994; Doherty et al., 1996;Heitzler et al., 1996; de Celis et al.,1997; Huppert et al., 1997; Qi et al.,1999; Rothwell et al., 1999; zur Lageand Jarman, 1999; Bruckner et al.,2000; Moloney et al., 2000; Parkset al., 2000; Culi et al., 2001; Laiet al., 2001; Pavlopoulos et al. 2001;Mishra-Gorur et al., 2002; Paninet al., 2002; Tsuda et al., 2002; Blandet al., 2003; Ikeuchi and Sisodia,2003; La Voie and Selkoe, 2003; Morelet al., 2003; Six et al., 2003; Wesleyand Mok, 2003; Sakata et al., 2004;Wang and Struhl, 2004, 2005; Wilkinet al., 2004; zur Lage et al., 2004; Em-ery et al., 2005; Le Borgne et al.,2005; Sapir et al., 2005; LeComteet al., 2006). Our data indicate that Nsignaling might also be regulated at amore primary level, the level ofmRNA 30 processing (at alternativepoly(A) sites). This regulation couldbe important for genes such as Nwithout a strong promoter (the N pro-moter remains undefined despiteefforts by many labs). Regulation atthe level of mRNA 30 processing mightbe the reason why many RNA-bindingproteins are recovered in geneticscreens based on N signaling (Norgaet al., 2003; Kankel et al., 2007).Among these proteins, only Sex Le-thal (Sxl) is known to affect N mRNAtranslation, in a sex-specific manner(Penn and Schedl, 2007). However,Sxl is unlikely to be involved in the al-ternative polyadenylation phenom-enon described in this report as most(if not all) embryos produce N RB inthe early-stage embryos and N RA inthe mid-stage-embryos. If alternativepolyadenylation were sex specific, wewould have observed a 50:50 ratiobetween N RA and N RB or betweenDl RA1 and Dl RA2. Our data providea useful framework for examining thefunctions of the other RNA-bindingproteins recovered in genetic screensbased on N signaling.
It has been known for a long timethat in early-stage and mid-stage
embryos, the mRNA products of anumber of genes differ in the lengthof their 30 UTR, Dl being one of them.Over the years, for one reason or theother, we have examined more than100 genes related to our studies byNorthern blotting. Almost a third ofthem show transcripts in the early-stage and mid-stage embryos that dif-fer in the length of their 30 UTR. It ispossible that many of these genes alsomanifest the N/Dl phenomenon wehave described in this report. N andDl are both required for N signaling,which is known to regulate embryo-genesis events in both early-stage andmid-stage embryos. Our data suggestthat the pattern of protein-producingcapacities of N mRNAs would be com-plementary to those of Dl mRNAs,which raises some interesting possi-ble scenarios for the regulation ofembryogenesis. As the Dl mRNA pro-duces more protein in early-stageembryos (0–4-hr AEL), it is possiblethat in these embryos the Dl proteinis the primary target of fine-scale pro-tein level regulation. On the otherhand, the N protein might be the pri-mary target in mid-stage embryos (af-ter 4-hr AEL) where the N mRNA pro-duces more protein. Our Nnd1-dse datasuggest that N might not be subjectedto fine-scale regulation when its levelis high. Therefore, the opposite sce-nario is also possible wherein the fine-scale regulation requires a low level ofthe protein. Another possible scenariois that processing at the alternativepoly(A) sites of a gene might functionto adjust protein levels based on theamounts of mRNA contributed by themother and the zygote. Recently, it hasbeen shown that for a large fraction ofgenes expressed in early-stageembryos both the mother and the zy-gote contribute mRNA (e.g., de Renziset al., 2007). All these possible scenar-ios might depend on the presence orabsence of specific mRNA 30-process-ing regulatory factors in early-stageand mid-stage embryos.
Sequence elements in the immedi-ate neighborhood of a poly(A) site arenecessary and sufficient to directmRNA 30-processing and polyadenyl-ation. However, these processes canbe regulated by sequences that are faraway or events that took place earlyon. For example, some of the proteinsthat are part of the mRNA 30-process-
ing complex are thought to associatewith the Pol II transcription complexsoon after transcription is initiated(Wells et al., 1998). Thus, in vivo anal-ysis of the regulation N or Dl expres-sion through mRNA 30 processingcould be quite challenging becausethis regulation might not be effectivewith exogenous promoters. In vivoregulation might also depend on theinteraction between mRNA 30-proc-essing regulators present in the early-stage and mid-stage embryos aboutwhich we know very little. Our datawould provide an excellent frameworkfor future studies aimed at systemati-cally analyzing all the conserved Nand Dl 30 sequences, identifying RNA-binding factors involved in the regula-tion of mRNA 30 processing at alterna-tive poly(A) sites in the N and Dlgenes, and determining the basis forthe different translation efficiencies ofthe early-stage and zygotic stage Nand Dl mRNAs. N RA, N RB, Dl RA1,and Dl RA2 mRNAs expressed pre-dominantly in either the early-stageor the mid-stage embryos would serveas excellent readouts.
EXPERIMENTAL
PROCEDURES
Fly Procedures
Fly stocks were obtained from theDrosophila Stock Center, Blooming-ton, Indiana. Nnd1-dse stock is de-scribed in Shepherd et al. (2009). Pro-cedures described in Ashburner et al.(2004), Campos-Ortega and Harten-stein. (1997), and Weischaus andNusslein-Volhard (1998) were followedfor rearing flies, collecting timedembryos, and processing embryos formolecular analyses. All fly stocks wereentrained to the circadian cycle so thatthey lay most of their eggs within a 4-hr period around dusk when reared at22�C (Ashburner et al., 2004). Thisprocedure minimizes age variationwithin embryo samples. Experimentalsamples were collected at 22�C (allexperiments except the Nnd1-dse experi-ments) or at 29�C (the Nnd1-dse experi-ments). As an environmental chamberwas used for rearing experimentalstocks, temperature could be adjustedindependent of the circadian cycle (thecircadian clock is temperature com-pensated and, therefore, not affectedby changes in temperature). At 22�C,
Dev
elop
men
tal D
ynam
ics
1230 SHEPHERD ET AL.

the mother primarily control embryo-genesis in 0–4-hr AEL and the zygoteprimarily controls embryogenesis atthe later stages (after 4-hr AEL). Nnd1-
dse stock was maintained at 18�C.Nnd1-dse and the control (yw) cageswere maintained at 29�C for embryocollections. In these experiments, thetimes were adjusted to reflect thefaster developmental rate at 29�C(Ashburner et al., 2004). However, forpresentation of results the times areconverted back to reflect developmen-tal rate at 22�C for easy comparisonwith the other data.
In the following description, alltimes correspond to development at22�C. Samples were collected at 1-, 2-, 3-, or 4-hr intervals (depending on theexperiment) and the majority ofembryos in these samples representedthe middle 45, 90, 150, or 180 min,respectively. Following collection, em-bryos were aged to required stages. Forobtaining the 4–12-hr AEL sample, wecombined RNA from a 0–4-hr collectionthat was aged for 4 hr (i.e., the 4–8-hrsample) and RNA from a 0–4-hr samplethat was aged for 8 hr (i.e., the 8–12-hrsample). For obtaining the 3–15-hr AELsample, we combined RNAs from 0–3-hrcollections that were aged for 3 hr (i.e.,the 3–6-hr sample), 6 hr (i.e., the 6–9-hrsample), 9 hr (i.e., the 9–12-hr sample),or 12 hr (i.e., the 12–15-hr sample) werecombined. For obtaining the 3–6-hrsample, 0–3-hr collection was aged for 3hr. For obtaining the 2–4- and the 4–6-hr sample, 0–2-hr collections were agedfor 2 or 4 hr, respectively.
Molecular Procedures
The procedures followed for immuno-staining, Northern blotting, Westernblotting, collection of different embry-onic developmental stages, and cellculture experiments are described inLieber et al. (1992, 1993), Wesley andMok (2003), Bardot et al. (2005), Moket al. (2005), and LeComte et al.(2006). N mRNAs are around 10 kblong, Dl mRNAs are about 5.5 kblong, and rp49 mRNAs are about 600bp long. Furthermore, the two �10-kb-long N mRNAs migrate close to-gether under a wide range of gel elec-trophoresis conditions. Therefore, Nor Dl Northern blots are based on along-run electrophoresis gel (21 hr at35 V) that resolves the larger N
mRNAs fairly well and rp49 Northernblots are based on a short-run electro-phoresis gel (14 hr at 35 V). a466 Nantibody (LeComte et al., 2006),C594.9B Dl antibody (DevelopmentalStudies Hybridoma Bank, Universityof Iowa, Iowa City, Iowa), anti-Hunch-back antibody (a generous gift fromPaul Macdonald), and actin antibody(Abcam, Cambridge, MA) were usedin immunostaining and signals devel-oped with a Horseradish peroxidase(HRP) or Alkaline phosphatase (AP)conjugated secondary antibody. aNI Nantibody (Anks Ab; Lieber et al.,1993), a466 N antibody (Cterm Ab),C594.9B Dl antibody, and anti-Hsp 70antibody (Sigma, St. Louis, MO) wereused in Western blotting.
First Choice RLM-RACE Kit(Ambion, Austin, TX) was used forRT-PCR. Standard procedures wereused for other PCRs and cloning. Theprimers used were the 30 RACEprimer in the kit, 30UTR5þ (50caccaatggaaacgtataagtc30), 30UTR2� (50agtttcgttttgctgtctggc30), 30UTR4þ (50atcgattaaacgtttgtgggac30). For making theactGFP-N30UTR constructs, theKpnI-NotI GFP coding sequence frag-ment from pEGFP (Clontech, PaloAlto, CA) was inserted after the actin5C promoter in the pBluescript (pBS)plasmid (a pBS plasmid with the�2.7-kb actin promoter EcoRI frag-ment was a generous gift from SimonKidd). The Notch 30UTR sequence,amplified with primers containingNotI sites, was cloned into the NotIsite at the end of the GFP-codingsequence in pEGFP. Plasmids withthe correct orientation were checkedby sequencing and used to transfectS2 cells. For making the hs-N30UTRRA and RB plasmids, an XbaI frag-ment containing the full-length NotchcDNA was cloned into the pBS ks�
plasmid. A larger Not1-StuI fragmentcontaining the Notch XbaI fragmentplus an additional Spe I site down-stream of the 30 XbaI site was excisedand cloned into NotI-StuI sites of apCaSpeR-hs vector without the hsp7030UTR-containing-Bam HI-fragment(DBam) to obtain the Notch RNA RA-producing construct (hsN30UTR RA).The SacII–SpeI fragment containingthe Notch 30UTR sequence from thenatural Sac II site immediately afterthe Notch stop codon to the terminusof Notch RNA RB was PCR amplified,
checked by sequencing, and usedto replace the full-length Notch 30
UTR region in hsN30UTR RA toobtain the construct producing theNotch RNARB (hsN30UTR 0RB0).For mutating the upstream poly(A)
hexamer of Dl, we used the Quick-Change Site-Directed Mutagenesis Kit(Stratagene, La Jolla, CA) with a high-fidelity pfu turbo polymerase enzyme.A Dl cDNA clone in pBluescript wasused as the template. Multiple cloneswere isolated, mutagenesis checked bysequencing, and cloned into the heat-shock pCaSpeR 3 vector. For generat-ing for the actGFP-Dl30UTR construct,we PCR amplified Dl 30 UTR and 150-bp downstream sequence using pri-mers with Not 1 sequence. This Not1fragment was used to replace the N 30
UTR in actGFP-N30UTR construct.Correct orientation was ascertainedusing Kp1 and Nhe1 enzymes. Dl 30
UTR sequence was checked for errors.Images and figures were processed
using the Photoshop (Adobe, SanJose, CA) and Canvas (Deneba) pro-grams. Any adjustment was appliedto whole images.
ACKNOWLEDGMENTSWe thank Jingping Li, Lee-Peng Mok,and Boris Bardot for help in the initialstages of the study; the DrosophilaStock Center and Dr. Spyros Artavanis-Tsakonas (Harvard University) for flystocks; Vermont Cancer Center forsequencing; Matt Rand and GregoryGilmartin for comments on the initialversions of the manuscript; NIH(NINDS) grant NS43122, Departmentof Microbiology and Molecular Geneticsat the University of Vermont (thanks toSusan Wallace), Neuroscience COBRE(P20 RR016435) at the University ofVermont (thanks to Rodney Parsonsand Cindy Forehand), and the Univer-sity of Vermont College of MedicineBridge grant for funds (thanks to Wolf-gang Dostmann andMarkus Thali). Wealso thank anonymous reviewers fortheir helpful comments that haveimproved the quality of ourmanuscript.
REFERENCES
Alton AK, Fechtel K, Kopczynski CC,Shepard SB, Kooh PJ, MuskavitchMAT. 1989. Molecular genetics of Delta,a locus required for ectodermal differen-tiation in Drosophila. Dev Genetics 10:261–272.
Dev
elop
men
tal D
ynam
ics
NOTCH AND DELTA mRNAs IN EMBRYOGENESIS 1231

Ahimou F, Mok L-P, Bardot B, Wesley CS.2004. The adhesion force of Notch withDelta and the rate of Notch signaling. JCell Biol 167:1217–1229.
Artavanis-Tsakonas S, Rand MD, LakeRJ. 1999. Notch signaling: Cell fate con-trol and signal integration in develop-ment. Science 284:770–776.
Ashburner M, Golic KG, Hawley RS.2004. Drosophhila. A laboratory hand-book. 2nd ed. New York: Cold SpringHarbor Press. p 1409.
Bardot B, Mok L-P, Thayer T, Ahimou F,Wesley CS. 2005. The Notch amino ter-minus regulates protein levels andDelta induced clustering of DrosophilaNotch receptors. Exp Cell Res 304:202–223.
Beaudoing E, Freier S, Wyatt JR, Clav-erie J-M, Gautheret D. 2007. Patternsof variant polyadenylation signal usagein human genes. Genome Res 10:1001–1010.
Bentley DL. 2005. Rules of engagement:co-transcriptional recruitment of pre-mRNA processing factors. Curr OpinCell Biol 17:251–256.
Bland CE, Kimberly P, Rand MD. 2003.Notch-induced proteolysis and nuclearlocalization of the Delta ligand. J BiolChem 278:13607–13610.
Brennan K, Tateson R, Lewis K, Marti-nez-Arias A. 1997. A functional analysisof Notch mutations in Drosophila.Genetics 147:177–188.
Brou C, Logeat F, Gupta N, Bessia C,LeBail O, Doedens JR, Cumano A,Roux P, Black RA, Israel A. 2000. Anovel proteolytic cleavage involved inNotch signaling: the role of the disinte-grin-metalloprotease TACE. Mol Cell 5:207–216.
Bruckner K, Perez L, Claussen H, CohenS. 2000. Glycosyltransferase activity ofFringe modulates Notch-Delta interac-tions. Nature 406:411–415.
Cabrera CV. 1990. Lateral inhibition andcell fate during neurogenesis in Drosoph-ila: the interactions between scute, NotchandDelta. Development 110:733–742.
Campos-Ortega JA, Hartenstein V. 1997.The embryonic development of Drosoph-ila melanogaster, 2nd ed. New York:Springer. p405.
Culi J, Martin-Blanco M, Modolell J.2001. The EGF receptor and N signal-ing pathways act antagonistically inDrosophila mesothorax bristle pattern-ing. Development 128:299–308.
Dansereau DA, Lunke MD, FinkielszteinA, Russell MA, Brook WJ. 2002.Hephaestus encodes a polypyrimidinetract binding protein that regulatesNotch signalling during wing develop-ment in Drosophila melanogaster. De-velopment 129:5553–5566.
de Celis J F, Bray S, Garcia-Bellido A.1997. Notch signaling regulates veinletexpression and establishes boundariesbetween veins and interveins in theDrosophila veins. Development 124:1919–1928.
De Renzis S, Elemento O, Tavazoie S,Wieschaus EF. 2007. Unmasking activa-
tion of the zygotic genome using chro-mosomal deletions in the Drosophilaembryo. PLoS Biol 5:e117
Doherty D, Feger G, Younger-Shepherd S,Jan LY, Jan YN. 1996. Delta is a ventralto dorsal signal complementary to Ser-rate, another Notch ligand, in Drosophilawing formation. Genes Dev 10:421–434.
Emery G, Hutterer A, Berdnik D, MayerB, Wirtz-Peitz F, Gaitan MG, KnoblichJA. 2005. Asymmetric Rab11 Endo-somes regulate Delta recycling andspecify cell fate in the Drosophila nerv-ous system. Cell 122:1–11
Fehon RG, Kooh PJ, Rebay I, Regan CL,Xu T, Muskavitch M, Artavanis-Tsako-nas S. 1990. Molecular interactionbetween the protein products of theneurogenic loci Notch and Delta, twoEGF-homologous genes in Drosophila.Cell 61:523–534.
Fortini ME, Rebay I, Caron LA, Artava-nis-Tsakonas S. 1993. An activatedNotch receptor blocks cell-fate commit-ment in the developing Drosophila eye.Nature 365:555–557.
Haenlin M, Kramatschek B, Campos-Ortega JA. 1990. The pattern of tran-scription of the neurogenic gene Deltaof Drosophila melanogaster. Develop-ment 110:905–914.
Heitzler P, Simpson P. 1991. The choice ofcell fate in the epidermis of Drosophila.Cell 64:1083–1092.
Heitzler P, Bourouis M, Ruel L, CarteretC, Simpson P. 1996. Genes of theEnhancer of split and achaete-scutecomplexes are required for a regulatoryloop between Notch and Delta duringlateral signaling in Drosophila. Devel-opment 122:161–171.
Huppert SS, Jacobsen TL, MuskavitchMAT. 1997. Feedback regulation is cen-tral to Delta-Notch signaling requiredfor Drosophila wing vein morphogene-sis. Development 124:3283–3291.
Ikeuchi T, Sisodia SS. 2003. The Notchligands, Delta1 and Jagged2, are sub-strates for presenilin-dependent ‘‘gamma-secretase’’ cleavage. J Biol Chem 278:7751–7754.
Irvine KD, Wieschaus E. 1994. fringe, aboundary-specific signaling molecule,mediates interactions between dorsaland ventral cells during Drosophilawing development. Cell 79:595–606
Juge F, Zaessinger S, Temme C, Wahle E,Simonelig M. 2002. Control of poly(A)polymerase level is essential to cyto-plasmic polyadenylation and early de-velopment in Drosophila. EMBO J 21:6603–6613.
Kankel MW, Hurlbut GD, Upadhyay G,Yajnik V, Yedvobnick B, Artavanis-Tsa-konas S. 2007. Investigating the geneticcircuitry of Mastermind in Drosophila,a Notch signal effector. Genetics 177:2493–2505.
Karolchik D, Baertsch R, Diekhans M,Furey TS, Hinrichs A, Lu YT, RoskinKM, Schwartz M, Sugnet CW, ThomasDJ, Weber RJ, Haussler D, Kent WJ.2003. The UCSC Genome BrowserDatabase. Nucleic Acids Res 31:51–54.
Klueg KM, Muskavitch MAT. 1999.Ligand-receptor interactions and trans-endocytosis of Delta, Serrate and Notch:members of the Notch signaling path-way in Drosophila. Development 112:3289–3297.
Kooh PJ, Fehon RG, Muskavitch MAT.1993. Implications of dynamic patternsof Delta and Notch expression for cellu-lar interactions during Drosophila de-velopment. Development 117:493–507.
Kopczynski CC, Alton AK, Fechtel K,Kooh PJ, Muskavitch MAT. 1988. Delta,a Drosophila neurogenic gene, is tran-scriptionally complex and encodes aprotein related to blood coagulation fac-tors and epidermal grpwth factor of ver-tebrates. Genes Dev 2:1723–1735.
Kornblihtt AR, de la Mata M, Fededa JP,Munoz MJ, Nogues G. 2004. Multiplelinks between transcription and splic-ing. RNA 10:1489–1498.
Lai EC, Deblandre GA, Kintner C, RubinGM. 2001. Drosophila neuralized is aubiquitin ligase that promotes theinternalization and degradation ofdelta. Dev Cell 1:783–794.
LaVoie MJ, Selkoe DJ. 2003. The Notchligands, Jagged and Delta, are sequen-tially processed by alpha-secretase andpresenilin/gamma-secretase and releasesignaling fragments. J Biol Chem 278:34427–34437.
Le Borgne R, Bardin A, Schweisguth F.2005. The roles of receptor and ligandendocytosis in regulating Notch signal-ing. Development 132:1751–1762.
LeComte M, Wesley U, Mok L-P, WesleyCS. 2006. Evidence for involvement ofdominant negative Notch molecules inthe normal course of Drosophila devel-opment. Dev Dyn 235:411–426.
Lecuyer E, Yoshida H, Parthasarathy N,Alm C, Bak T, Cerovina T, Hughes TR,Tomancak P, Krause HM. 2007. Globalanalysis of mRNA localization reveals aprominent role in organizing cellulararchitecture and function. Cell 131:174–187.
Lejeune F, Ishigaki Y, Li X, Maquat LE.2002. The exon junction complex isdetected on CBP80-bound but noteIF4E-bound mRNA in mammaliancells: dynamics of mRNP remodeling.EMBO J 21:3536–3545.
Li Y, Chen ZY, Wang W, Baker CC, KrugRM. 2001. The 30-end-processing factorCPSF is required for the splicing of sin-gle-intron pre-mRNAs in vivo. RNA 7:920–931.
Liang H-L, Nien C-Y, Liu H-Y, MetzsteinMM, Kirov N, Rushlow C. 2008. Thezinc-finger protein Zelda is a key acti-vator of the early zygotic genome inDrosophila. Nature 456:400–403.
Lieber T, Wesley CS, Alcamo E, Hassel B,Krane JF, Campos-Ortega JA, YoungMW. 1992. Single amino acid substitu-tions in EGF-like elements of the Notchand Delta proteins modify Drosophiladevelopment and affect cell adhesion invitro. Neuron 9:847–859.
Lieber T, Kidd S, Alcamo E, Corbin V,Young MW. 1993. Antineurogenic
Dev
elop
men
tal D
ynam
ics
1232 SHEPHERD ET AL.

phenotypes induced by truncated Notchproteins indicate a role in signal trans-duction and may point to a novel func-tion for Notch in nuclei. Genes Dev 7:1949–1965.
Lieber T, Kidd S, Young MW. 2002. Kuz-banian-mediated cleavage of DrosophilaNotch. Genes Dev 16:209–221.
Mishra-Gorur K, Rand MD, Perez-Villa-mil B, Artavanis-Tsakonas S. 2002.Down-regulation of Delta by proteolyticprocessing. J Cell Biol 159:313–324.
Mok L-P, Qin T, Bardot B, LeComte M,Homayouni A, Ahimou F, Wesley C.2005. Delta activity independent of itsactivity as a ligand of Notch. BMC DevBiol 5:6.
Moloney DJ, Panin VM, Johnston JSH,Chen J, Shao L, Wilson R, Wang Y,Stanley P, Irvine KD, Haltiwanger RS,Vogt TF. 2000. Fringe is a glycosyl-transferase that modifies Notch. Nature406:369–375.
Morel V, Le Borgne R, Schweisguth F.2003. Snail is required for Delta endo-cytosis and Notch-dependent activationof single-minded expression. Dev GenesEvol 213:65–72.
Mumm JS, Kopan R. 2000. Notch signal-ing: from the outside in. Dev Biol 228:151–165.
Murata T, Nagaso H, Kashiwabara S,Baba T, Okano H, Yokoyama KK. 2001.The hiiragi gene encodes a poly(A) poly-merase, which controls the formation ofthe wing margin in Drosophila mela-nogaster. Dev Biol 233:137–147.
Nakao K, Campos-Ortega JA. 1996. Per-sistent expression of genes of theenhancer of split complex suppressesneural development in Drosophila. Neu-ron 16:275–286.
Norga KK, Gurganus MC, Dilda CL,Yamamoto A, Lyman RF, Patel PH,Rubin GM, Hoskins RA, Mackay TF,Bellen HJ. 2003. Quantitative analysisof bristle number in Drosophila mela-nogaster mutants identifies genesinvolved in neural development. CurrBiol 13:1388–1397.
Okabe M, Imai T, Kyurusu M, Hiromi Y,Okano H. 2001. Translational repres-sion determining neuronal potential inDrosophila asymmetric cell division.Nature 411:94–98.
Okano H, Imai T, Okabe M. 2002. Musha-shi: a translational regulator of cellfate. J Cell Sci 115:1355–1359.
Panin VM, Shao L, Lei L, Moloney DJ,Irvine KD, Haltiwanger RS. 2002.Notch ligands are substrates for proteinO-Fucosyltransferase-1 and Fringe. JBiol Chem 277:29945–29952.
Parks AL, Kleug KM, Stout JR, Muska-vitch MAT. 2000. Ligand endocytosisdrives receptor dissociation and activa-tion in the Notch pathway. Develop-ment 127:1373–1385.
Pavlopoulos E, Pitsouli C, Kleug KM,Muskavitch MAT, Moschonas NK, Deli-dakis C. 2001. neuralized encodes a pe-ripheral membrane protein involved indelta signaling and endocytosis. DevCell 1:807–816.
Penn JKM, Schedl P. 2007. The masterswitch gene Sex-lethal promotes femaledevelopment by negatively regulatingthe N-signaling pathway. Dev Cell 12:275–286.
Qi H, Rand MD, Wu X, Sestan N, WangW, Rakic P, Xu T, Artavanis-TsakonasS. 1999. Processing of the notch liganddelta by the metalloprotease Kuzba-nian. Science 283:91–94.
Reed R, Hurt E. 2002. A conservedmRNA export machinery coupled topre-mRNA splicing. Cell 108:523–531.
Rothwell WF, Zhang CX, Zelano C, HsiehT-S, Sullivan W. 1999. The Drosophilacentrosomal protein Nuf is required forrecruiting Dah, a membrane associatedprotein, to furrows in the early embryo.J Cell Sci 112:2885–2893.
Royet J, Bouwmeester T, Cohen SM.1998. Notchless encodes a novel WD40-repeat-containing protein that modu-lates Notch signaling activity. EMBO J17:7351–7360.
Rusconi JC, Corbin V. 1998. Evidence fora novel Notch pathway required formuscle precursor selection in Drosoph-ila. Mech Dev 79:39–50.
Sakata T, Sakaguchi H, Tsuda L, Higashi-tani A, Aigaki T, Matsuno K, HayashiS. 2004. Drosophila Nedd4 regulatesendocytosis of notch and suppresses itsligand-independent activation. Curr Biol14:2228–2236.
Sapir A, Assa-Kunik E, Tsruya R,Schejter E, Shilo B. 2005. Unidirec-tional Notch signaling depends on con-tinuous cleavage of Delta. Development132:123–132.
Schweisguth F. 2004. Notch signaling ac-tivity. Curr Biol 14:R129–138.
Shellenbarger DL, Mohler JD. 1975. Tem-perature-sensitive mutations of theNotch locus in Drosophila melanogaster.Genetics 81:143–162.
Shellenbarger DL, Mohler JD. 1978. Tem-perature-sensitive periods and autonomyof pleiotropic effects of l(1)Nts1, a condi-tional Notch lethal in Drosophila. DevBiol 62:432–446.
Shepherd AS, Singh R, Wesley CS. 2009.Notch mRNA expression in Drosophilaembryos is negatively regulated at the levelof mRNA 30 processing. PloS One 4:e8063.
Six E, Ndiaye D, Laabi Y, Brou C, Gupta-Rossi N, Israel A, Logeat F. 2003. TheNotch ligand Delta 1 is sequentiallycleaved by an ADAM protease and h-secretase. Proc Natl Acad Sci USA 100:7638–7643.
Soller M, White K. 2001. ELAV inhibits30-end processing to promote neuralsplicing of ewg pre-mRNA. Genes Dev17:2526–538.
Struhl G, Fitzgerald K, Greenwald I.1993. Intrinsic activity of the Lin-12and Notch intracellular domains invivo. Cell 74:331–345.
Tsuda L, Nagaraj R, Zipursky SL, Bane-rjee U. 2002. An EGFR/Ebi/Sno path-way promotes delta expression byinactivating Su(H)/SMRTER repressionduring inductive notch signaling. Cell110:625–637.
Vassin H, Bremer KA, Knust E, Campos-Ortega JA. 1987. The neurogenic geneDelta of Drosophila melanogaster isexpressed in neurogenic territories andencodes a putative transmembrane pro-tein with EGF-like repeats. EMBO J 6:3431–3440.
Wang W, Struhl G. 2004. DrosophilaEpsin mediates a select endocytic path-way that DSL ligands must enter toactivate Notch. Development 131:5367–5380.
Wang W, Struhl G. 2005. Distinct rolesfor Mind bomb, Neuralized and Epsinin mediating DSL endocytosis and sig-naling in Drosophila. Development 132:2883–2894.
Wells SE, Hillner PE, Vale RD, Sachs AB.1998. Circularization of mRNA by eu-karyotic translation initiation factors.Mol Cell 2:135–140.
Weischaus E, Nusslein-Volhard C. 1998.Looking at embryos. In: Roberts DB, editor.Drosophila. A practical approach, 2nd ed.Ithaca, NY: IRL Press. p179–214.
Wells SE, Hillner PE, Vale RD, Sachs AB.1998. Circularization of mRNA by eu-karyotic translation initiation factors.Mol Cell 2:135–140.
Wesley CS, Mok L-P. 2003. Regulationof Notch signaling by a novel mechanisminvolving Suppressor of Hairless stabilityand carboxyl terminus-truncated Notch.Mol Cell Biol 23:5581–5593.
Wesley CS, Saez L. 2000. Analysis ofNotch lacking the carboxyl terminusidentified in Drosophila embryos. J CellBiol 149:683–696.
Wickens M, Bernstein DS, Kimble J,Parker R. 2002. A PUF family portrait:30UTR regulation as a way of life.Trends Genet 18:150–157.
Wilkin MB, Carbery AM, Fostier M,Aslam H, Mazaleyrat SL, Higgs J,Myat A, Evans DA, Cornell M, BaronM. 2004. Regulation of notch endosomalsorting and signaling by DrosophilaNedd4 family proteins. Curr Biol 14:2237–2244.
Wilusz CJ, Gao M, Jones CL, Wilusz J,Peltz SW. 2001a. Poly(A)-binding pro-teins regulate both mRNA deadenyla-tion and decapping in yeast cytoplasmicextracts. RNA 7:1416–1424.
Wilusz CJ, Wormington M, Peltz SW.2001b. The cap-to-tail guide to mRNAturnover. Nat Rev Mol Cell Biol 2:237–246.
Xu T, Rebay I, Fleming RJ, Scottgale TN,Artavanis-Tsakonas S. 1990. The Notchlocus and the genetic circuitry involvedin early Drosophila neurogenesis.Genes Dev 4:464–475.
zur Lage P, Jarman AP. 1999. Antagonismof EGFR and Notch signaling in thereiterative recruitment of Drosophilaadult chordotonal sense organ precur-sors. Development 126:3149–3157.
zur Lage PI, Powell LM, Prentice DR,McLaughlin P, Jarman AP. 2004. EGFreceptor signaling triggers recruitmentof Drosophila sense organ precursors bystimulating proneural gene autoregula-tion. Dev Cell 7:687–696.
Dev
elop
men
tal D
ynam
ics
NOTCH AND DELTA mRNAs IN EMBRYOGENESIS 1233





























