Embed Size (px)
Citation preview

ACTAUNIVERSITATIS
UPSALIENSISUPPSALA
2013
Digital Comprehensive Summaries of Uppsala Dissertationsfrom the Faculty of Medicine 929
Novel Circulating and TissueBiomarkers for Small IntestineNeuroendocrine Tumors andLung Carcinoids
TAO CUI
ISSN 1651-6206ISBN 978-91-554-8735-5urn:nbn:se:uu:diva-205570

Dissertation presented at Uppsala University to be publicly examined in Enghoffsalen,Entrance 50, Uppsala Hospital, Uppsala, Friday, October 18, 2013 at 09:15 for the degree ofDoctor of Philosophy (Faculty of Medicine). The examination will be conducted in English.
AbstractCui, T. 2013. Novel Circulating and Tissue Biomarkers for Small Intestine NeuroendocrineTumors and Lung Carcinoids. Acta Universitatis Upsaliensis. Digital ComprehensiveSummaries of Uppsala Dissertations from the Faculty of Medicine 929. 46 pp. Uppsala.ISBN 978-91-554-8735-5.
Small intestine neuroendocrine tumors (SI-NETs) and lung carcinoids (LCs) are relativelyindolent tumors, which originate from neuroendocrine (NE) cells of the diffuse NE system.Metastases can spread before diagnosis. Thus, potential cures become unavailable, whichentitles new biomarker development. Indeed, we aimed at developing Ma2 autoantibodies andolfactory receptor 51E1 (OR51E1) as potential novel biomarkers and exploring other candidateprotein markers in patients’ serum.
First, we established a sensitive, specific and reliable anti-Ma2 indirect ELISA to distinguishSI-NET patients from healthy controls. We detected longer progression-free and recurrence-free survivals in patients expressing low anti-Ma2 titers. Moreover, a high anti-Ma2 titer wasmore sensitive than chromogranin A for the risk of recurrence after radical operation of SI-NETpatients.
We then investigated OR51E1 expression in SI-NETs and LCs. OR51E1 mRNA expression,analyzed by quantitative real-time PCR, was high in microdissected SI-NET cells, in LC celllines and in frozen LC specimens. Immunohistochemistry (IHC) showed abundant OR51E1protein expression in SI-NETs. OR51E1 co-expressed with vesicular-monoamine-transporter-1in the majority of normal and neoplastic enterochromaffin cells.
Furthermore, the study on LCs revealed that OR51E1, somatostatin receptor (SSTR) 2,SSTR3, and SSTR5 are expressed in 85%, 71%, 25% and 39% of typical carcinoids (TCs),whereas in 86%, 79%, 43% and 36% of atypical carcinoids (ACs). Based on the proposed IHCscoring system, in the LC cases, where all SSTR subtypes were absent, membrane OR51E1expression was detected in 10 out of 17 TCs and 1 out of 2 ACs. Moreover, higher OR51E1scores were detected in 5 out of 6 OctreoScan-negative LC lesions.
In addition, the last presented study used a novel suspension bead array, which targeted124 unique proteins, by using Human Protein Atlas antibodies, to profile biotinylated serumsamples from SI-NET patients and healthy controls. We showed 9 proteins, IGFBP2, IGF1,SHKBP1, ETS1, IL1α, STX2, MAML3, EGR3 and XIAP as significant contributors to tumorclassification.
In conclusion, we proposed Ma2 autoantibodies as a sensitive circulating marker for SI-NETrecurrence; OR51E1 as a candidate therapeutic target for SI-NETs; whereas as a novel diagnosticmarker for LCs and 9 serum proteins as novel potential SI-NET markers.
Keywords: small intestine neuroendocrine tumors, lung carcinoids, Ma2 autoantibodies,immunohistochemistry, olfactory receptor 51E1, blood samples, antibody suspension beadarray
Tao Cui, Uppsala University, Department of Medical Sciences, Endocrine Oncology,Akademiska sjukhuset, SE-751 85 Uppsala, Sweden.
© Tao Cui 2013
ISSN 1651-6206ISBN 978-91-554-8735-5urn:nbn:se:uu:diva-205570 (http://urn.kb.se/resolve?urn=urn:nbn:se:uu:diva-205570)

To my parents, Fang, baby and friends 致爸妈,昉,宝宝及朋友们


List of Papers
This thesis is based on the following papers, which are listed by their Roman numerals.
I Cui T, Hurtig M, Elgue G, Li SC, Veronesi G, Essaghir A, Demoulin JB, Pelosi G, Alimohammadi M, Öberg K, Giandomenico V. Paraneoplastic antigen Ma2 autoantibodies as specific blood biomarkers for detection of early recurrence of small intestine neuroendocrine tumors. PLoS One. 2010 Dec 30;5(12):e16010.
II Cui T*, Tsolakis AV*, Li SC, Cunningham JL, Lind T, Öberg K, Giandomenico V. Olfactory receptor 51E1 protein as a potential novel tissue biomarker for small intestine neuroendocrine carcinomas. Eur J Endocrinol. 2013 Jan 17;168(2):253-61.
III Giandomenico V*, Cui T*, Grimelius L, Öberg K, Pelosi G, Tsolakis AV. Olfactory receptor 51E1 as a novel target for diagnosis in somatostatin receptor negative lung carcinoids. J Mol Endocrinol, Advanced online publication 2013 Aug 22.
IV Darmanis S, Cui T, Drobin K, Li SC, Öberg K, Nilsson P, Schwenk JM, Giandomenico V. Identification of candidate serum proteins for classifying well-differentiated small intestinal neuroendocrine tumors. Submitted Manuscript.
*The authors contributed equally. Reprints were made with permission from the respective publishers.

Other related works
I Li SC, Martijn C, Cui T, Essaghir A, Luque RM, Demoulin JB, Castaño JP, Öberg K, Giandomenico V. The somatostatin analogue octreotide inhibits growth of small intestine neuroendocrine tumour cells. PLoS One. 2012. 7(10):e48411.
II Naboulsi R, Bergström J, Li SC, Cui T, Öberg K, Nystrand M, Giandomenico V. The immunoCAP ISAC technology to develop a novel diagnostic tumor immunoassay for rare small intestinal neuroendocrine tumor patients. Submitted manuscript.
Supervisors
Assoc. Prof. Valeria Giandomenico, PhD, Dept of Medical Sciences, Endocrine Oncology, Uppsala University, Uppsala, Sweden Prof. Kjell Öberg, MD, PhD, Dept of Medical Sciences, Endocrine Oncology, Uppsala University, Uppsala, Sweden Dr. Apostolos V Tsolakis, MD, PhD, Dept of Medical Sciences, Endocrine Oncology, Uppsala University, Uppsala, Sweden Chair
Prof. Magnus Essand, PhD, Dept of Immunology, Genetics and Pathology, Clinical Immunology, Research Group Essand, Rudbeck laboratory, Uppsala University, Uppsala, Sweden Faculty Opponent
Assoc. Prof. Marco Volante, MD, PhD , Dept of Clinical and Biological Sciences, University of Turin, San Luigi Hospital, Turin, Italy Review Board
Prof. Gunnar Westin, PhD, Dept of Surgical Sciences, Endocrine Surgery, Uppsala University Assoc. Prof. Ali Ata Moazzami, PhD, Dept of Chemistry, Organic Chemistry, Swedish Univeristy of Agricultural Sciences Dr. Anders Sundqvist, PhD, Ludwig Institute for Cancer Research, Uppsala Biomedical Center, Uppsala University, Uppsala, Sweden Program Controller
Prof. Eva Tiensuu Janson, MD, PhD, head of Dept of Medical Sciences

Contents
Introduction ................................................................................................... 11 Neuroendocrine cells ................................................................................ 11 General NE markers ................................................................................. 11 Specific markers of NE cells in the GI-tract ............................................ 12 Small intestine neuroendocrine tumors .................................................... 13 Lung carcinoids ........................................................................................ 15 Novel biomarkers in SI-NETs and LCs ................................................... 17 Paraneoplastic antigen Ma2 autoantibodies ............................................. 18 Olfactory receptor 51E1 ........................................................................... 19
Aims of the study .......................................................................................... 22
Materials and Methods .................................................................................. 23 Laser capture microdissection (Paper II) .................................................. 23 Indirect enzyme-linked immunosorbent assay (Paper I) .......................... 23 In vitro transcription-translation (ITT) and sequential immunoprecipitation (IP) (Paper I) .......................................................... 24 Immunohistochemistry (Paper I, II, III) ................................................... 24 Antibody suspension bead array (Paper IV) ............................................. 25 Statistical Analysis (Paper I, II, III, IV) ................................................... 27
Results and Discussion ................................................................................. 28 Paper I. .................................................................................................... 28 Paper II. .................................................................................................... 30 Paper III. ................................................................................................... 31 Paper IV. .................................................................................................. 33
Concluding Remarks and Perspectives ......................................................... 35
Abstract in Chinese (中文摘要) ................................................................... 37
Acknowledgements ....................................................................................... 38
References ..................................................................................................... 41

Abbreviations
5-HIAA 5-hydroxyindoleacetic acid
5-HT 5-hydroxytryptamine, serotonin
5-HTP 5-hydroxytryptophan
AC atypical (lung) carcinoid
Akt protein kinase B
AUC area under (ROC) curve
Bax B-cell lymphoma 2-associated X protein
BGA between group analysis
cAMP cyclic adenosine monophosphate
CD8 cluster of differentiation 8
CgA chromogranin A
CT computed tomography
CTC circulating tumor cell
DOPA 3,4-dihydroxyphenylalanine
DOTA 1,4,7,10-tetraazacyclododecane-1,4,7,10-tetraacetic acid
DTPA diethylene triamine pentaacetic acid
EC enterochromaffin
EGR3 Early growth response protein 3
ENETS European Neuroendocrine Tumor Society
ETS1 protein C-ets-1
FDA (Unite State) Food and Drug Administration
FFPE formalin-fixed paraffin-embedded
GEP-NET gastroenteropancreatic neuroendocrine tumor
GI gastrointestinal
GPCR G protein-coupled receptor
GPER G-protein-coupled estrogen receptor
GPR112 G-protein coupled receptor 112
GRIA2 glutamate receptor, ionotropic, AMPA 2
HPA human protein atlas
HRP horseradish peroxidase
IFNα interferon α

IgG immunoglobulin G
IGF1 insulin growth factor 1
IGFBP2 insulin-like growth factor-binding protein 2
IHC immunohistochemistry
IL1α interleukin 1 alpha
IMP3 insulin-like growth factor II-messenger RNA-binding protein 3
IP immunoprecipitation
IR immunoreactive
ITT in vitro transcription-translation
Ki-67 MKI67, antigen identified by monoclonal antibody Ki-67
LAR long acting release
LC lung carcinoid
LCM laser capture microdissection
LCNEC large-cell neuroendocrine carcinoma
LDCV large dense-core vesicle
LM liver metastasis
LNM lymph node metastasis
MAML3 mastermind-like protein 3
MRI magnetic resonance imaging
mTOR mammalian target of rapamycin
NE neuroendocrine
NEC neuroendocrine carcinoma
NET neuroendocrine tumor
NSE (or ENO2) neuron-speicific enolase (or enolase 2)
OctreoScan
111In-DTPA-D-Phe1 octreotide somatostatin receptor
scintigraphy
OR olfactory receptor
OR51E1 (or PSGR2) olfactory receptor 51 E1 (or prostate-specific G protein-coupled receptor 2)
PAX5 paired box 5
PET positron emission tomography
PI3K phosphoinositide-3-kinase
PFS progression free survival
PNEC pulmonary neuroendocrine cell
PNMA paraneoplastic antigen Ma
PNMA2 paraneoplastic antigen Ma2
PNMA4 (MOAP1) paraneoplastic antigen Ma2 (or modulator of apoptosis-1)

PNS paraneoplastic syndrome
PROMID Placebo-controlled prospective Randomized study on the anti-proliferative efficacy of Octreotide LAR in patients with metastatic neuroendocrine MIDgut tumors
PSGR (or OR51E2) prostate-specific G protein-coupled receptor (or olfactory receptor 51E2)
PT primary tumor
PTEN phosphatase and tensin homologue
QRT-PCR quantitative real-time PCR
RF random forest (analysis)
RFS recurrence free survival
ROC curve receiver operating charateristic curve
RTK receptor tyrosine kinase
SAPE streptavidin, R-phycoerythrin conjugate
SERPINA10 serpin peptidase inhibitor, clade A, member 10
SDS-PAGE sodium dodecyl sulfate-polyacrylamide gel electrophoresis
SHKBP1 SH3KBP1-binding protein 1
SI-NET small intestine neuroendcrine tumor
SPOCK1 sparc/osteonectin, cwcv and kazal-like domains proteoglycan (testican) 1
SRS somatostatin receptor scintigraphy
SSA somatostatin analogue
SSTR somatostatin receptor
SSV small synaptic vesicle
STX2 syntaxin-2
SYN synatophysin
TC typical (lung) carcinoid
TMA tissue microarray analysis
TMB 3,3’,5,5’-tetramethylbenzidine
TNM tumor-node-metastasis
TpH1 tryptophan hydroxylase 1
TTF1 thyroid transcription factor 1
VMAT vesicular monoamine transporter
VMAT1 vesicular monoamine transporter type I (or solute carrier family 18 member 1, SLC18A1)
VMAT2 vesicular monoamine transporter type II (or solute carrier family 18 member 2, SLC18A2)
WHO World Health Organization
XIAP E3 ubiquitin-protein ligase XIAP

11
Introduction
Neuroendocrine cells The neuroendocrine (NE) cells are endocrine cells acquiring NE phenotypes, which are more often disseminated in the diffused neuroendocrine system (DNES) throughout a variety of tissues in our body [1]. NE cells share characteristics with neurons, such as polarized membranes, neurotransmitter-synthesizing enzymes, neural cell adhesion molecules and peptides and amino acid transmitter receptors. They uptake and release neurotransmitters, neuropeptides and amine hormones and store them within cytoplasmic basal membrane-bound granules or vesicles. The highly controlled secretion of specific products into the bloodstream targeting distant organs mirrors the typical endocrine function [2]. Microscopically, NE cells show uniform nuclei and abundant granular or clear cytoplasm. They may form either small insular or trabecular clusters or are properly dispersed among other cells. Immunohistochemical staining of general or specific NE markers enables their exact identification [1].
General NE markers The general NE markers are functionally conserved in the process of NE cell differentiation. They include large dense-core vesicle (LDCV)-associated markers, such as chromogranins; small synaptic vesicle (SSV)-associated markers, such as synatophysin (SYN) and cytosolic markers, such as neuron-specific enolase (NSE) [1], which is called enolase 2 (ENO2) today. In addition, vesicular monoamine transporters (VMATs) are essential markers located in the membrane of both types of vesicles.
The granin family includes eight members. The most important and well known granin is chromogranin A (CgA), which is a heat-stable, hydrophilic and acidic glycoprotein. CgA mRNA and protein are expressed in the majority of normal and neoplastic NE cells. Post-translational cleavage of CgA produces different functional peptides, such as pancreastatin, catestatin, vasostatins, which affect secretion of other hormones and play a role in vasoconstriction and regulate metabolism [3]. SYN is a glycoprotein distributed in a variety of NE cells, including enterochromaffin (EC) cells and the four types of pancreatic cells [4]. SYN is strongly expressed by a variety of neuroendocrine tumors (NETs) arising from different organs.

12
Thus, it has been clinically used as a general marker of normal [5] and neoplastic NE cells [6]. In addition, NSE is the best known cytosolic marker for NE cell recognition. However, the commercial antibodies are not specific, which have detected NSE in a variety of non-neuroendocrine tumor tissues and thus limit the use of this enzymatic marker [7]. VMATs are transporters responsible for the active uptake of monoamine hormones from the cytosol into the storage vesicles. They comprise vesicular monoamine transporters type I (VMAT1), which is called solute carrier family 18, member 1 (SLC18A1) today; and vesicular monoamine transporters type II (VMAT2), which is known as solute carrier family 18, member 2 (SLC18A2). VMAT1 preferentially locates in the LDCVs of different NE cells, such as the chromaffin cells of the adrenal medulla and the EC cells of the gastrointestinal- (GI-) tract, whereas VMAT2 is mainly expressed in the central nervous system and in the histamine containing EC-like cells of the stomach, as well as in the mast cells [8].
Specific markers of NE cells in the GI-tract NE cells of the GI-tract originate from local multipotent GI stem cells located throughout the DNES, where they function as regulators of secretion, absorption, motility, mucosal cell proliferation and potentially control the immune-barrier [4]. They are the largest group of hormone-producing cells, including thirteen [9] or fourteen subtypes [4], according to different pathologists. The EC cells are the most abundant subtype of enteroendocrine cells in the GI mucosa. They are distributed throughout the GI-tract, particularly in the glands of pyloric antrum, duodenum and ileum. The EC cells mainly secrete a monoamine hormone, serotonin (5-hydroxytryptamine; 5-HT) and some neuropeptides, such as substance P and vasoactive intestinal polypeptide. Their secretory granules readily stain with silver (argentaffin stain) [9, 10].
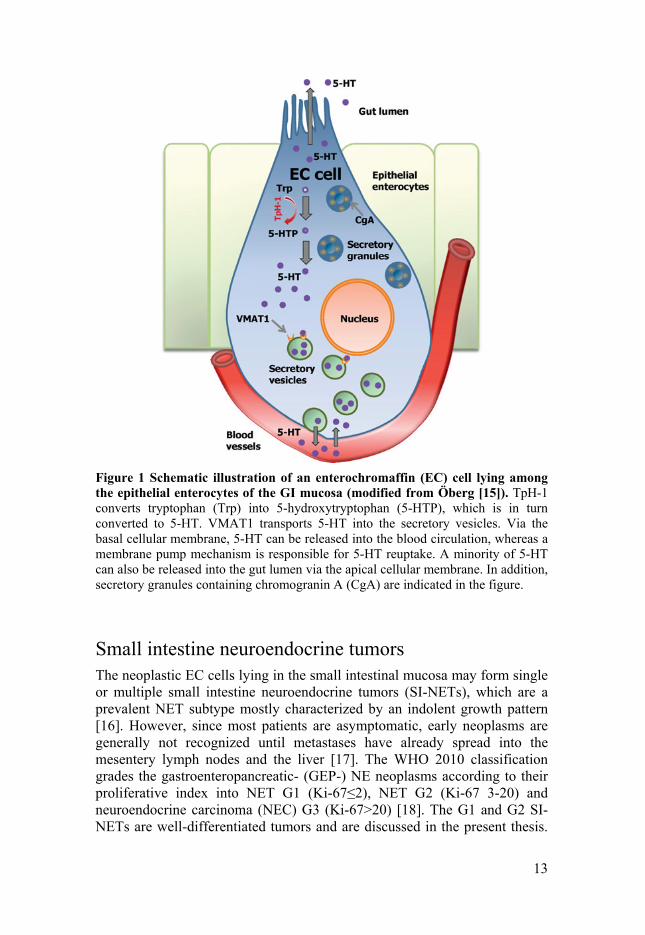
Serotonin is the most important specific marker of the EC cells. Luminal substances and mechanical pressure can activate EC cells by inducing intracellular calcium influx. This triggers the release of pre-stored 5-HT from basal granules into the lumen or lamina propria. Thereafter, serotonin acts on either enterocytes or afferent nerve terminals in the lamina propria, which initiate both secretion and propulsive motor patterns [10, 11]. Cytoplasmic 5-HT is transported via VMAT1 into the secretory vesicles in the EC cells [8], whereas the rate-limiting enzymes of the 5-HT synthesis is the tryptophan hydroxylase-1 (TpH-1) [12], which has been suggested crucial for SI-NET cell recognition [13, 14] (Figure 1).
13
Figure 1 Schematic illustration of an enterochromaffin (EC) cell lying among the epithelial enterocytes of the GI mucosa (modified from Öberg [15]). TpH-1 converts tryptophan (Trp) into 5-hydroxytryptophan (5-HTP), which is in turn converted to 5-HT. VMAT1 transports 5-HT into the secretory vesicles. Via the basal cellular membrane, 5-HT can be released into the blood circulation, whereas a membrane pump mechanism is responsible for 5-HT reuptake. A minority of 5-HT can also be released into the gut lumen via the apical cellular membrane. In addition, secretory granules containing chromogranin A (CgA) are indicated in the figure.
Small intestine neuroendocrine tumors The neoplastic EC cells lying in the small intestinal mucosa may form single or multiple small intestine neuroendocrine tumors (SI-NETs), which are a prevalent NET subtype mostly characterized by an indolent growth pattern [16]. However, since most patients are asymptomatic, early neoplasms are generally not recognized until metastases have already spread into the mesentery lymph nodes and the liver [17]. The WHO 2010 classification grades the gastroenteropancreatic- (GEP-) NE neoplasms according to their proliferative index into NET G1 (Ki-67≤2), NET G2 (Ki-67 3-20) and neuroendocrine carcinoma (NEC) G3 (Ki-67>20) [18]. The G1 and G2 SI-NETs are well-differentiated tumors and are discussed in the present thesis.

14
Furthermore, the European Neuroendocrine Tumor Society (ENETS) proposed the tumor-node-metastasis (TNM) system for staging and both the Ki-67 index and the mitotic count for grading [19, 20]. Both classification systems should be used either with care or combined when it is clinically requested [18, 20, 21].
The SI-NETs are diagnosed mainly by biochemical markers and imaging techniques. Biochemical markers can be detected either in the circulating fluid or in the tumor tissue. CgA and NSE are general circulating NET markers, whereas the most specific SI-NET blood marker, compatible with their EC cell origin, is serotonin [9]. Furthermore, the specific amine or peptide hormones may generate certain hormone-related syndromes in patients suffering from the so-called functioning tumors. Indeed, the serotonin hypersecretion may contribute to the carcinoid syndrome triggering e.g. flushing and/or diarrhea in SI-NET patients. This enables patient diagnosis, by measuring urinary elevation of a 5-HT downstream metabolite, named 5-hydroxyindoleacetic acid (5-HIAA) [22]. Moreover, general tissue NET markers mainly include CgA, SYN and NSE, which characterize the NE nature of the neoplasia. In addition, the Ki-67 index is used in NETs to assess the cell proliferation level.
The SI-NETs are usually imaged via functional approaches, since they highly express hormonal receptors, such as somatostatin receptors (SSTRs). As a consequence, somatostatin receptor scintigraphy (SRS) by using 111In-DTPA-D-Phe1 octreotide (OctreoScan) has become a standard diagnostic approach during the last decades [23]. However, positron emission tomography (PET) by using radiolabeled tracers, such as [68Ga-DOTA]-octreotide/octreotate, has significantly improved imaging sensitivity and specificity, which would clearly replace SRS in the future [24-26]. In addition, since NE cells are able to take up amine and peptide precursors, PET by using [11C]-DOPA [27] and [11C]-5-hydroxy-L-tryptophan (HTP) [28] have also been developed; however, they have limited availability in a minority of hospitals [29]. SRS and PET can hybrid with either computed tomography (CT) or magnetic resonance imaging (MRI) to offer fusion images, which reveal function and anatomy of the malignancies [29, 30].
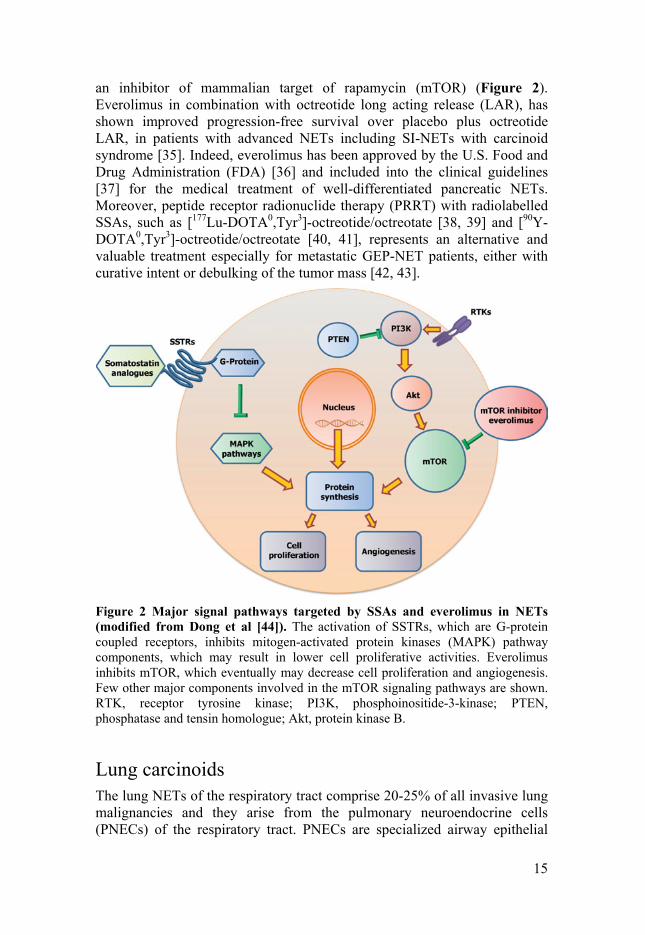
Surgery is the selective cure for localized SI-NETs. However, due to their high metastatic rate and tumor heterogeneity, different therapeutic regimes have been developed, which include the use of somatostatin analogues (SSAs) (Figure 2) and interferon α (IFNα) [31]. SSAs were initially introduced to relieve the symptoms, whereas the PROMID study recently showed their importance in delaying disease progression [32]. IFNα may introduce high symptomatic and biological responses; however, with concomitant side effects. Moreover, the combination of SSAs and IFNα may lower the risk of tumor progression [33]. In the last decade, the improved knowledge of NETs tumor biology has contributed to the novel targeted therapy development [34]. Indeed, everolimus (RAD-001), which is the 40-O-(2-hydroxyethyl) derivative of sirolimus, works similarly to sirolimus as
15
an inhibitor of mammalian target of rapamycin (mTOR) (Figure 2). Everolimus in combination with octreotide long acting release (LAR), has shown improved progression-free survival over placebo plus octreotide LAR, in patients with advanced NETs including SI-NETs with carcinoid syndrome [35]. Indeed, everolimus has been approved by the U.S. Food and Drug Administration (FDA) [36] and included into the clinical guidelines [37] for the medical treatment of well-differentiated pancreatic NETs. Moreover, peptide receptor radionuclide therapy (PRRT) with radiolabelled SSAs, such as [177Lu-DOTA0,Tyr3]-octreotide/octreotate [38, 39] and [90Y-DOTA0,Tyr3]-octreotide/octreotate [40, 41], represents an alternative and valuable treatment especially for metastatic GEP-NET patients, either with curative intent or debulking of the tumor mass [42, 43].
Figure 2 Major signal pathways targeted by SSAs and everolimus in NETs (modified from Dong et al [44]). The activation of SSTRs, which are G-protein coupled receptors, inhibits mitogen-activated protein kinases (MAPK) pathway components, which may result in lower cell proliferative activities. Everolimus inhibits mTOR, which eventually may decrease cell proliferation and angiogenesis. Few other major components involved in the mTOR signaling pathways are shown. RTK, receptor tyrosine kinase; PI3K, phosphoinositide-3-kinase; PTEN, phosphatase and tensin homologue; Akt, protein kinase B.
Lung carcinoids
The lung NETs of the respiratory tract comprise 20-25% of all invasive lung malignancies and they arise from the pulmonary neuroendocrine cells (PNECs) of the respiratory tract. PNECs are specialized airway epithelial

16
cells, which are either solitary cells or clusters called neuroepithelial bodies in the lung. These cells are mainly located in the nasal respiratory epithelium, laryngeal mucosa and throughout the entire respiratory tract from the trachea to the terminal airways. However, PNECs have not been clearly studied as the enterochromaffin cells [45-47].
The lung NETs are divided into well-differentiated NETs, which include only lung carcinoids (LCs); and poorly differentiated NECs, which are large cell neuroendocrine carcinomas (LCNECs) and small cell lung carcinomas (SCLCs). In turn, LCs are subdivided into typical carcinoids (TCs) and atypical lung carcinoids (ACs), the former being low-grade malignant tumors with <2 mitoses/2mm2, absence of necrosis and good prognosis (about 88%, 5-year survival) and the latter being intermediate-grade malignant tumors with up to 10 mitoses/2mm2 and/or presence of punctate necrosis and poorer prognosis (about 50%, 5-year survival). The different metastatic potential to regional lymph nodes, liver and bone accounts for major differences in behavior of TCs and ACs [48].
The established LC biochemical markers include CgA, SYN and NSE. CgA [49] and NSE [50] are expressed in all LCs. SYN is found in 97% of TCs and 89% of ACs [49]. The thyroid transcription factor 1 (TTF1) is an important marker to determine several tumors of lung origin. However, the expression of TTF1 in TCs and ACs widely varies with either all negatives [51] or frequently positive immunostaining [52, 53]. Furthermore, traditional chest radiographs are less sensitive than CT, MRI and SRS for the diagnosis of LCs. Since SSTRs can be expressed in LCs [54], OctreoScan has been used to locate these tumors [55]. Octreoscan mainly targets SSTR2 and has less affinity for SSTR3 and SSTR5 [56]. Usually small biopsies and/or cytological specimens are used as an initial work up to reveal the NE differentiation of these tumors. Surgical material is mainly required to confirm the mitotic count and characterize the punctuate necrosis, which thus further categorizes the tumors in TCs and ACs [49].
Early radical surgery, which is not feasible for peripheral tumors, is the only ideal curative treatment for LC patients [57]. Moreover, clinicians do not have an optimal therapy to rely on to cure inoperable metastatic lung carcinoids at the moment. Metastatic LCs susceptibility to chemotherapy agents may be inadequate, due to low response rates and serious side effects. Thus, multimodality strategies have been developed without achieving a proper curative capacity [58]. Indeed, the SSA-based biotherapy has been introduced mainly to control symptoms; and may also improve long-term survival of LC patients, mainly for metastatic ACs [59]. Moreover, PRRT has been shown to cause either LC tumor response or stabilization during the last years [38]. However, NET patients with negative or low-level uptake in SRS are not optimal candidates either for SSA-based diagnosis or therapies [60]. This specifically underlines the need of novel tumor targets clinical development to diagnose and treat LC patients.

17
Novel biomarkers in SI-NETs and LCs
Although sensitive biomarkers such as CgA and SYN have been fundamental for the clinical diagnosis of NETs, the detection of markers for certain subtypes of tumor, which lack of novel specific biomarkers, is an unmet challenge. Moreover, novel sensitive and specific biomarkers, which consider NET anatomical depiction, need to be established to achieve early diagnosis, metastases prediction and surveillance of early recurrence of NETs. This novel findings may allow early surgical and pharmacotherapeutic intervention to prolong patients survival and improve life’s quality. Indeed, although SSTRs have been widely used, thanks to the continuous development of SSAs, which aimed at controlling symptoms and tumor progression, differentially expressed novel therapeutic targets in tumor and normal cells, are required mainly for the tumors, which lack adequate SSTR expression. During the recent decades, various types of potential biomarkers have been suggested for SI-NETs and LCs. For instance, the identification of circulating tumor cells (CTCs) as prognostic markers in SI-NETs and LCs has been of major importance [61, 62]. Global transcriptomic analysis also provided valuable information about the expression of circulating mRNA and tissue micro-RNA in the neoplastic SI-NE cells [63, 64]. Furthermore, different immunohistochemical analyses confirmed that the connective tissue growth factor (CTGF) is specifically expressed in SI-NETs [65]; whereas insulin growth factor 1 (IGF1) is involved in the early tumorigenesis of SI-NE cells [66]; and new proteins such as paired box 5 (PAX5) [67] and insulin-like growth factor II-messenger RNA-binding protein 3 (IMP3) [68] appear to be useful markers to discriminate LCs from high-grade lung NETs. Moreover, Leja et al have profiled normal small ileal mucosa, SI-NE primary tumors and liver metastases, by using Affymetrix microarrays analysis. The main findings reported six differentially and specifically expressed novel genes in the neuroendocrine tumor cells, which are named GPR112, GRIA2, PNMA2, OR51E1, SPOCK1 and SERPINA10. The transcripts of these genes were analyzed by quantitative real-time PCR (QRT-PCR) on normal intestine mucosa and SI-NET tissues; and on microdissected normal EC cells and SI-NET cells [69]. These results prompted us to study whether some of these genes mRNAs and encoded proteins might have been developed as new diagnostic, prognostic biomarkers or therapeutic targets for SI-NETs and LCs. Indeed, we have mainly focused on PNMA2 and OR51E1. Our in-depth studies led to novel findings in SI-NETs and LCs, throughout our scientific work, which I summarized in my PhD thesis.

18
Paraneoplastic antigen Ma2 autoantibodies Human PNMA2 belongs to a gene family, which include six PNMA genes [70]. The expression of paraneoplastic antigen Ma2, encoded by the PNMA2 gene, is normally restricted to the normal brain tissue [71]. However, it can be detected in tumors located outside the nervous system during carcinogenesis. This has been clearly shown from the scientific studies on testicular cancer [71], cholangiocarcinoma [72] and SI-NETs [69]. Today, the functions of PNMA proteins are not clear. However, the PNMA4 (also named modulator of apoptosis-1, MOAP1) was identified as a Bax-associating protein, which controls apoptosis induction in mammalian cells [73]. Thus, the significant homology of PNMA4 and the other PNMA proteins suggested a potential involvement of PNMA proteins in the apoptosis signaling [70].
Antitumor immune responses to neuronal antigens expressed by tumor cells may lead to detectable levels of autoantibodies in patients’ serum and plasma [74-76]. However, only a few onconeuronal autoantibodies have been characterized in patients suffering from neurological paraneoplastic syndrome (PNS) up to now [77]. The term PNS describes a variety of specific symptoms caused by tumor cells, which either systematically produce a large amount of hormones, growth factors and cytokines, or produce autoantibodies against ectopically expressed neuronal antigens in the tumor cells. The former one triggers hormonal PNS and the latter one the neurological PNS [74]. Because PNS symptoms may appear before tumor diagnosis; potential cases of PNS may suggest early antitumor therapy and immunotherapy to prevent progressive neuronal death [74, 75, 78]. However, to understand whether either the autoantibodies are associated to specific neurological symptoms or they are anticancer immune reaction markers is rather difficult [75].
The Ma2 autoantibodies were first identified in patients suffering from testicular cancer with either paraneoplastic limbic or brain-stem encephalitis by Voltz et al [71]. The exact mechanism why Ma2 autoantibodies start being expressed is not elucidated. However, cytotoxic T-cell activity may potentially participate to this process [79, 80]. Indeed, scientific data have shown that anti-Ma2 positive sera sometimes appear during diagnosis of various tumor types [77, 81-83]. The most common cancer types expressing anti-Ma2 are either the testicular cancer, in young males with associated PNS, or the non-SCLC [75, 77, 78, 81]. SI-NETs and LCs, which originate from specific and independent neuroendocrine cells, synthesize and secrete biologically active compounds, which are able to produce either hormonal PNS or less commonly neurological PNS [74]. However, the overall prevalence of PNS in NETs is rather low and about 15%; and it is mainly caused by NET cells in the GI-tract, the lungs and the breast [74]. Although the Ma2 antigen expression both in primary and metastatic SI-NET cells [84] suggested the exploration of Ma2 autoantibodies in the circulating blood of

19
these patients, the clinicians at Uppsala University Hospital and the European Institute of Oncology have never identified PNS or neurological symptoms in the patients included in our studies.
Olfactory receptor 51E1 The olfactory receptors (ORs) were first discovered by Buck and Axel in 1991 [85]. The OR genes belong to the largest superfamily of the human genome [86]. The size of the OR repertoire is a feature that differs across species. The proportion of intact and pseudo genes are significantly different. The decline of the OR gene family in some primates coincided with the acquisition of trichromatic vision, which suggests that better visual capability may let olfaction redundant [87]. Human beings almost have 400 functional olfactory receptors [86]. They are a G-protein coupled receptors (GPCRs) subclass, which have been considered pivotal as therapeutic targets for a variety of diseases, including cancer [88]. These chemosensory receptors are predominantly expressed in the olfactory sensory neurons; and play a pivotal role in the specific recognition of diverse stimuli [89]. In vertebrates, an intracellular signaling cascade was proposed to be involved in olfaction chemosensory transduction in the olfactory neurons. This cascade, via type III adenylyl cyclase upon odor binding, increases intracellular cAMP and leads to the opening of a nonselective olfactory-specific cyclic-nucleotide gated ion channel to allow calcium influx [89]. The human OR51E1 gene encodes the olfactory receptor, family 51, subfamily E, member 1 (OR51E1). OR51E1 mRNA expression has been detected in normal brain [90] and normal prostate tissue [91]. Furthermore, prostate carcinomas highly express OR51E1 [91, 92]. Moreover, OR51E1 has been suggested as mRNA marker for prostate carcinomas [93, 94] and then for SI-NETs at different stages of disease [69, 95]. The pathophysiological role of OR51E1 during tumorigenesis and tumor progression has not been elucidated. However, OR51E1 has been suggested as a novel diagnostic marker complementary to other traditional markers in prostate cancers [93] and can significantly distinguish tumors from normal tissue and benign hyperplasia in the prostate [94]. Although OR51E1 does not associate with the aggressiveness of prostate cancer, a lower OR51E1 expression correlates with an earlier recurrence [94].
In addition, in the normal lung tissue, the OR51E1 mRNA was detected by sensitive RNA-Seq analysis. The authors also unveiled internal splicing variants of OR51E1 in normal breast, colon, prostate and testis tissue, which may lead to a truncated protein containing only the first transmembrane domain of the receptor [96].
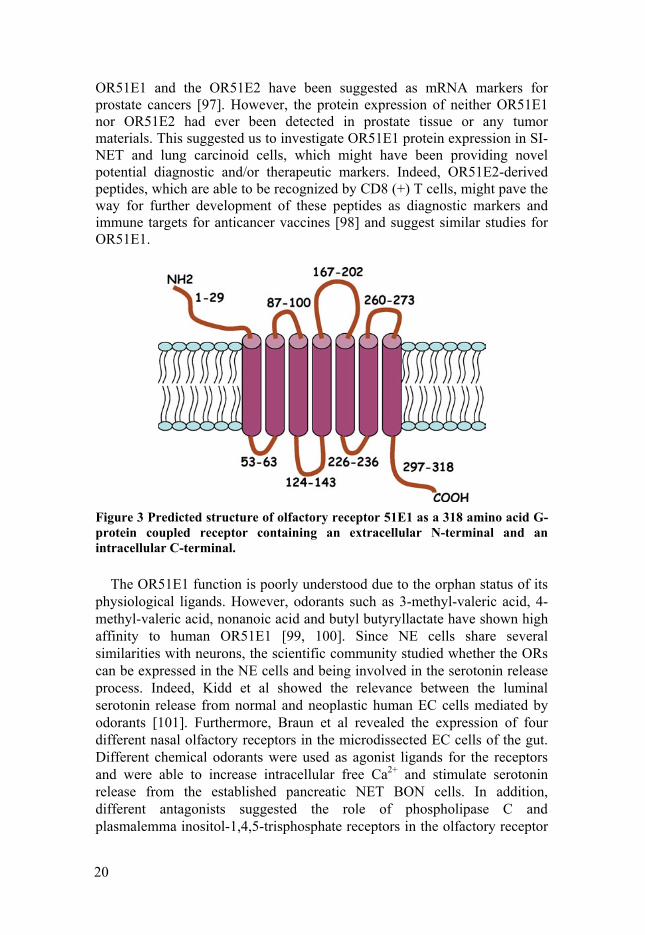
A predicted structure of the full-length OR51E1 protein is shown in Figure 3. OR51E1 shares 57% identity at the amino acid level with the prostate-specific G protein-coupled receptor (PSGR or OR51E2). Both the
20
OR51E1 and the OR51E2 have been suggested as mRNA markers for prostate cancers [97]. However, the protein expression of neither OR51E1 nor OR51E2 had ever been detected in prostate tissue or any tumor materials. This suggested us to investigate OR51E1 protein expression in SI-NET and lung carcinoid cells, which might have been providing novel potential diagnostic and/or therapeutic markers. Indeed, OR51E2-derived peptides, which are able to be recognized by CD8 (+) T cells, might pave the way for further development of these peptides as diagnostic markers and immune targets for anticancer vaccines [98] and suggest similar studies for OR51E1.
Figure 3 Predicted structure of olfactory receptor 51E1 as a 318 amino acid G-protein coupled receptor containing an extracellular N-terminal and an intracellular C-terminal.
The OR51E1 function is poorly understood due to the orphan status of its physiological ligands. However, odorants such as 3-methyl-valeric acid, 4-methyl-valeric acid, nonanoic acid and butyl butyryllactate have shown high affinity to human OR51E1 [99, 100]. Since NE cells share several similarities with neurons, the scientific community studied whether the ORs can be expressed in the NE cells and being involved in the serotonin release process. Indeed, Kidd et al showed the relevance between the luminal serotonin release from normal and neoplastic human EC cells mediated by odorants [101]. Furthermore, Braun et al revealed the expression of four different nasal olfactory receptors in the microdissected EC cells of the gut. Different chemical odorants were used as agonist ligands for the receptors and were able to increase intracellular free Ca2+ and stimulate serotonin release from the established pancreatic NET BON cells. In addition, different antagonists suggested the role of phospholipase C and plasmalemma inositol-1,4,5-trisphosphate receptors in the olfactory receptor

21
downstream signaling pathways [102]. However, the functions of olfactory receptors in either the GI-tract or the lung are still largely unknown. Pluznick et al investigated the mouse olfactory receptor 78, which is the ortholog of human OR51E2, as a responder to short chain fatty acids (SCFAs) in the kidney. This receptor mediates renin secretion thanks to these SCFAs, which are end products of fermentation by the gut microbiota; and are absorbed into the circulation [103]. A main hypothesis suggests that certain SCFAs in the human GI-mucosa are ligands to human OR51E2, which shares significant sequence similarity to OR51E1. Primeaux et al have identified elevated levels of different olfactory receptors in the duodenum of rats, which were fed with high fat diet. The authors suggested the role of gut olfactory receptors in the sensing and regulation of dietary fat [104]. Furthermore, although no current investigation has been focused on olfactory receptor function in the lung tissues, this essential organ of the respiratory tract does express a variety of olfactory receptors, which also include OR51E1 [96]. Moreover, the NE cells in the lung have been shown to express different olfactory receptors in mice, which might imply their potential role in sensing the environmental changes of the air chemical composition [105].

22
Aims of the study
The overall aim of my project was to study novel circulating and tissue biomarkers in well-differentiated SI-NETs and LCs to eventually improve their diagnosis, prognosis and therapy. The specific aims of the study were elucidated in the included papers: Paper I. To evaluate whether Ma2 autoantibodies can be detected in the blood of SI-NET patients and developed as a novel circulating biomarker for clinical diagnosis and recurrence of the malignancy. Paper II. To explore whether OR51E1 protein can be detected in SI-NETs and developed as a new clinical tissue biomarker for these tumors. Paper III. To investigate OR51E1 mRNA and protein expression in LCs and evaluate whether OR51E1 can be developed as a novel target to diagnose SSTR negative LCs. Paper IV. To discover novel candidate biomarker protein profiles for SI-NETs by investigating proteomic signatures in serum of SI-NET patients and healthy individuals.

23
Materials and Methods
The majority of the materials and methods included in this thesis are fully described in the listed papers. However, the most important methods used during the experimental work are briefly described as follows.
Laser capture microdissection (Paper II) Laser capture microdissection (LCM) was used to collect SI-NET and tumor adjacent microenvironment cells to investigate their potential OR51E1 expression. Snap-frozen tissue specimens were sliced into ~10 μm sections using a microtome cryostat (Leica) and adhered to polyethylenenaphthalate membrane frame slides (Life Technologies). Tumor cells and cells of adjacent tumor microenvironment, from each section, were extracted by using the ArcturusXT microdissection system (Life Technologies) according to the manufacturer’s instructions.
Indirect enzyme-linked immunosorbent assay (Paper I) A novel indirect ELISA was developed to detect Ma2 autoantibody titers in sera and plasma of NET patients and healthy donors. GST-PNMA2 recombinant protein (Abnova) was coated on Maxisorp strips (Nunc). Strips were blocked and incubated with blood samples diluted 1:400 in neutral phosphate buffer saline (PBS, pH 7.4). Horseradish peroxidase (HRP)-conjugated rabbit anti-human IgG (Dako) was used as secondary detection antibody. The end-point color was developed by using 3,3’,5,5’-tetramethylbenzidine (TMB) (Dako) as a substrate and H2SO4 as a stop solution. Strips were properly washed between steps. Absorbance was detected by using Multiskan Ascent microplates photometer (Thermo Fisher) and Ascent software v2.6 (Thermo Elelectron). Blank absorbance was subtracted and a 4-parameter logistic fitting standard curve was established by using serial dilutions of the serum sample from a SI-NET patient expressing high anti-Ma2 titer. Autoantibody titers for each individual were expressed as arbitrary units and a proper cut-off was chosen to distinguish patients from healthy controls. The precision of the novel indirect ELISA was expressed in intra- and inter-assay percent coefficients of variation (CV %), which were always below 10%.

24
In vitro transcription-translation (ITT) and sequential immunoprecipitation (IP) (Paper I) ITT and sequential IP were performed to verify the specificity of serum anti-Ma2. 35S-Met–radiolabeled human PNMA2 protein was produced by ITT from the full-length cDNA clone for human PNMA2 (BioScience Geneservice) by using 35S-methionine and TnT SP6 Quick Coupled Transcription-Translation System (Promega). During the first immunoprecipitation, the serum Ma2 autoantibodies at different titers, from 800 up to 4500 arbitrary units, which belonged to different healthy donors and SI-NET patients, were allowed to bind 35S-labeled PNMA2 at 4°C overnight. Then, incubation with protein G-Sepharose beads (GE Lifescience) was performed at 4°C for 2 hours at room temperature. Beads were collected by centrifugation and supernatants were removed. Efficient washings were introduced to minimize background. The 35S-labeled PNMA2 was then released by heating at 80°C for 5 minutes and was collected in the supernatant after centrifugation. During the second immunoprecipitation, a commercial goat anti-Ma2 antibody (Santa Cruz) bound to different amounts of 35S-labeled PNMA2, which corresponded to different titers of anti-Ma2 in the human serum samples previously used. Thereafter, we added protein G-Sepharose beads to each sample and continued the incubation at 4°C for 2 hours. The samples were centrifuged and the beads were washed 5 times. The beads were then resuspended in 4× sodium dodecyl sulfate-polyacrylamide gel electrophoresis (SDS-PAGE) loading buffer containing 5% β-mercaptoethanol. The immunocomplexes, which contained 35S-labeled PNMA2 proteins, were released and denatured from the beads by heating at 95°C for 5 minutes. After centrifugation, 20 µl of each sample were resolved by 10% SDS-PAGE. The gel was dried, subjected to autoradiography by using phosphorimager 425S (Molecular Dynamics) and analyzed by using ImageQuant software.
Immunohistochemistry (Paper I, II, III) Immunohistochemistry analysis detected PNMA2, OR51E1 and SSTRs protein expression on formalin-fixed paraffin-embedded (FFPE) tissue specimens. FFPE material with histopathologically confirmed diagnosis of SI-NETs and LCs were obtained as 4-μm sections from the Pathology Biobank, Uppsala University Hospital and the European Institute of Oncology in Milan. Deparaffinization and antigen retrieval were performed and the primary antibodies were applied to the sections. The polymer detection system (DakoCytomation, EnVision System-HRP, Dako) was used for IHC. Diaminobenzidine was used as chromogen and nuclear counterstain was performed by using Meyer’s hematoxylin (Histolab Product). The antibodies were diluted in the Dako’s antibody diluents before use. They

25
included polyclonal rabbit anti-PNMA2 (1:350, HPA001936, Atlas Antibodies) (Paper I), HRP-conjugated rabbit anti-human IgG (1:500, A0423, Dako) (Paper I), polyclonal rabbit anti-OR51E1 antibody (1:500, A1854, LSBio) (Paper II, III), polyclonal rabbit anti -SSTR2A (1:5000, SS-800), -SSTR3 (1:5000, SS-850), -SSTR5 (1:5000, SS-890) (Gramsch Laboratories) and monoclonal rabbit anti-SSTR2A (1:200, UMB-1, Epitomics) (Paper III). The sections were evaluated under Axiophot light microscope (Carl Zeiss) and microphotographs were obtained by using AxioVision Rel. 4.5 Software. The percentage of the immunoreactive (IR) cells was estimated by a light microscope at a magnification of ×400 by using a square grid in one of the oculars. Four randomly selected areas were examined on tissue specimens, whereas the entire neoplastic tissue was examined for the smaller lesions. The specificity of all immunoreactions was verified by substituting the primary antibody with nonimmune IgG; and in some cases by neutralizing the immunostaining via pre-absorption of primary antibodies with their respective blocking peptides. Appropriate positive control tissue specimens were also included and immunostained by using different antibodies. The immunohistochemical results of OR51E1 took into account both the cellular compartmentalization (membrane vs. cytoplasm) and the percentage of tumor cells (<50% vs. >50%). The results were semi-quantified for the relevant antibodies on a scale from 0 to 3+, by using a similar approach to a previously elaborated SSTR2 scoring system [106]. Our scoring system is explained in details in the Materials and Methods section of Paper III.
Furthermore, the co-localization study was performed by using double immunofluorescence to detect co-expression of OR51E1 and VMAT1 in normal and malignant EC cells. The specimens were microwave treated, blocked with normal donkey serum (017-000-121, Jackson ImmunoResearch) and incubated with a mixture diluted polyclonal goat anti-VMAT1 (1:400, C-19, sc-7718, Santa Cruz) and polyclonal rabbit anti-OR51E1 (1:100, A1854, LSBio). Then the sections were incubated with a mixture of fluorescence-labeled secondary antibodies (Jackson ImmunoResearch), mounted and evaluated under Axioplan2 imaging microscope (Carl Zeiss). Images were acquired with a ×630 magnification and analyzed with AxioVision Rel. 4.8 Software, as described in Paper II.
Antibody suspension bead array (Paper IV) An antibody suspension bead array (SBA) is a powerful single-binder immunoassay to identify target proteins in a complex solution such as serum and plasma [107], by using a panel of antibodies that are routinely validated for specificity on planar protein microarrays against their antigens [108]. The assay system is able to detect proteins down to picomolar levels with dynamic ranges over three orders of magnitude [107]; and it has been

26
successfully used to identify candidate biomarkers in different types of malignancies, such as prostate cancer [109]. In our study, an antibody SBA was established to explore new potential serum protein markers, which might have been able to distinguish well-differentiated SI-NET patients at different stages from healthy donors. We prepared a list of protein targets by using information from the literature and data from our published [84, 110] and unpublished SI-NET microarray analyses. Protein profiles were generated by using a set of 184 HPA antibodies, which targeted 124 unique proteins. Furthermore, we collected serum samples from two independent individual cohorts, cohort 1 and cohort 2. Both cohorts included healthy controls and untreated SI-NET patients at different stages of disease.
First, serum samples from cohort 1, which included 77 individuals, were used at the discovery phase of our analyses. Briefly, proteins in the serum samples were labeled with biotin. Antibodies were coupled to beads as previously described [111]. All different color-coded bead IDs, carrying different capture antibodies, were mixed to develop a novel SBA. Then, the serum samples were heat-treated and incubated with the SBA overnight. Diluted fluorescent streptavidin, R-phycoerythrin conjugate (SAPE, Invitrogen), as a reporter dye, was added to bind biotinylated captured proteins. Proper washing steps were included. The experimental readout is subsequently facilitated by the co-occurrence of the coupled reporter dye and the detected beads color-code (Figure 4). The array was analyzed by a Luminex FlexMap3D instrument. Median signal intensities of each bead ID were used for subsequent analyses.
Next, a variety of promising target proteins were selected from our results at the discovery phase of our study, by using univariate and multivariate statistical analyses. We thus proceeded with a second marker validation phase, by using a similar experimental procedure as described above, on an independent sample cohort, cohort 2, which included 132 individuals. Based on further statistical analyses, this second phase of our analyses generated a narrower panel of candidate serum protein markers for well-differentiated SI-NETs diagnosis.

27
Figure 4 Workflow of antibody suspension bead arrays by using serum or plasma samples. A, sample distribution into microtiter plates; B, biotin-label proteins in diluted samples; C, array setup by coupling antibodies onto beads with distinct color codes; D, heat-treat samples and incubate with beads; E, remove unbound proteins, add fluorescent streptavidin and measure the beads.
Statistical Analysis (Paper I, II, III, IV) The statistical analyses were performed to evaluate the significance of our results. The analyses were two-tailed and performed by using either GraphPad Prism 5 (GraphPad Software, La Jolla, CA) (Paper I, II, III) or the R statistical software [112] (survival analysis in Paper I and all the statistical analyses in Paper IV). All p values <0.05 (Paper I, II, III) or <0.01 (Paper IV) were considered significant. Since we used a variety of these analyses, they are described in detail in each paper.

28
Results and Discussion
Paper I. Ma2 autoantibodies are a more sensitive biomarker than CgA for SI-NET progression and recurrence after radical surgery with curative intent A novel indirect ELISA was set up to screen the anti-Ma2 titers in serum or plasma samples. We confirmed the specificity of serum Ma2 autoantibodies by using western blot and sequential immunoprecipitation analyses. Major findings of this investigation underlined that anti-Ma2 titers significantly discriminated a large portion of SI-NET patients from healthy controls; either considering the diverse stages of disease or all the diseased groups. This may imply that anti-Ma2 is likely produced during SI-NET development. Furthermore, the blood levels were constantly kept during tumor progression. In addition, the sensitivity of the assay was up to 50%, whereas the specificity was 98%. Receiving operating characteristic (ROC) curve analyses, which are of major importance to establish the reliability of a novel biomarker, showed ideal values. Indeed, the areas under the curves (AUCs) had values between 0.734 and 0.816, which indicated a good accuracy of anti-Ma2 titer as a potential reliable candidate diagnostic marker.
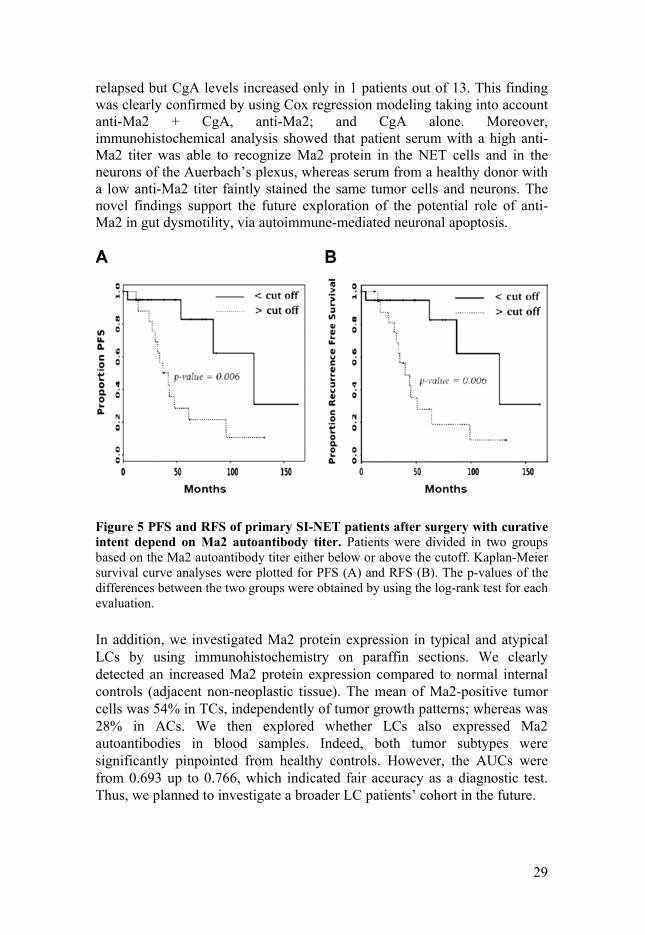
Two of the most essential clinical problems in managing SI-NETs are tumor progression and early recurrence detection. We showed that 19 patients expressing Ma2 autoantibody titer < cutoff had a longer free survival (PFS) and recurrence free survival (RFS), compared to another 17 patients expressing Ma2 autoantibody titer > cutoff. The significance of the analyses, was clearly expressed by p-values = 0.006 in both patient cohorts, as shown in Figure 5. Furthermore, this highlighted that patients with Ma2 autoantibody titer > cutoff reached an estimated median survival time of about 40 months, whereas the patients with levels < cutoff reached one of about 125 months. Moreover, the hazard ratios were 4.31 (p-value = 0.011) for PFS and 4.24 (p-value = 0.012) for RFS, which confirmed this innovative findings. Although, an increased circulating CgA level is considered important in indicating tumor recurrence [113], this parameter was not a pivotal indication in our patient cohorts. Indeed, the group of 19 patients with anti-Ma2 < cutoff, showed that only 4 patients had tumor recurrence during follow-up; and 2 out of 4 had increased CgA concentration. Unexpectedly, in the group of 17 patients with anti-Ma2 > cutoff, 13 patients
29
relapsed but CgA levels increased only in 1 patients out of 13. This finding was clearly confirmed by using Cox regression modeling taking into account anti-Ma2 + CgA, anti-Ma2; and CgA alone. Moreover, immunohistochemical analysis showed that patient serum with a high anti-Ma2 titer was able to recognize Ma2 protein in the NET cells and in the neurons of the Auerbach’s plexus, whereas serum from a healthy donor with a low anti-Ma2 titer faintly stained the same tumor cells and neurons. The novel findings support the future exploration of the potential role of anti-Ma2 in gut dysmotility, via autoimmune-mediated neuronal apoptosis.
Figure 5 PFS and RFS of primary SI-NET patients after surgery with curative intent depend on Ma2 autoantibody titer. Patients were divided in two groups based on the Ma2 autoantibody titer either below or above the cutoff. Kaplan-Meier survival curve analyses were plotted for PFS (A) and RFS (B). The p-values of the differences between the two groups were obtained by using the log-rank test for each evaluation.
In addition, we investigated Ma2 protein expression in typical and atypical LCs by using immunohistochemistry on paraffin sections. We clearly detected an increased Ma2 protein expression compared to normal internal controls (adjacent non-neoplastic tissue). The mean of Ma2-positive tumor cells was 54% in TCs, independently of tumor growth patterns; whereas was 28% in ACs. We then explored whether LCs also expressed Ma2 autoantibodies in blood samples. Indeed, both tumor subtypes were significantly pinpointed from healthy controls. However, the AUCs were from 0.693 up to 0.766, which indicated fair accuracy as a diagnostic test. Thus, we planned to investigate a broader LC patients’ cohort in the future.

30
Paper II. OR51E1 is highly expressed in SI-NET cells and co-localizes with VMAT1 in the majority of normal and neoplastic EC cells The OR51E1 coding sequence was wild-type in SI-NETs at different stages of disease. Higher OR51E1 transcript expression was detected in the microdissected tumor cells, from all investigated SI-NET patients at different stages, compared to the matched microdissected adjacent microenvironment cells (p<0.01). Indeed, Leja et al previously suggested OR51E1 overexpression in SI-NETs. However, microdissection was not performed to narrow the cell type, which might specifically express OR51E1 in the SI-tumor blocks used in the previous analyses [69] .
OR51E1 protein expression in SI-NETs was investigated in FFPE primary tumors and metastases. Prostate carcinoma slides were used as an external positive control, since it is well known that prostate carcinomas express OR51E1 mRNA [97, 114-116]. Indeed, we detected 90% prostate carcinoma cells immunoreactive (IR) for OR51E1. In the SI-NET cells, 31 out of 89 specimens showed over 50% OR51E1-IR tumor cells. They included 18 out of 43 primary tumors, 7 out of 28 mesentery metastases and 6 out of 18 liver metastases. The cytoplasmic pattern was detected in 80% of the OR51E1-IR tumor cells, whereas the membranous immunostaining was present in 20% of the cells. Indeed, it is known that the cytoplasmic portion of GPCRs may be responsible for receptor internalization [117]; however, to address this concept clearly requires a broader investigation on the OR51E1 internalization. The strongest immunostaining was mainly detected in the tumor cell clusters facing the fibrovascular stroma. This finding might imply a potential role of OR51E1 in the interaction with the surrounding cellular microenvironment [118]. Furthermore, perinuclear immunostaining was detected in about 20% of the tumor cells. Although this pattern is rare, the ectopically expressed recombinant G-protein-coupled estrogen receptor (GPER) has been found with perinuclear accumulation shortly after ligand addition [119]. Thus, it is possible to hypothesize that OR51E1 perinuclear localization might reflect an unusual process, similar to the one used by GPER, in which delayed sorting and accumulation of OR51E1 may occur in the perinuclear compartment after endocytosis. Moreover, no immunoreactivity was detected, either after replacement of the antibody by nonimmune serum or after the neutralization test in SI-NETs. Since the hepatic cell staining in the evaluated liver metastatic specimen persisted during the neutralization test, this suggested the presence of a nonspecific immunostaining in the liver. Our unpublished quantitative real-time PCR analysis data on normal human hepatic cells showed absence of OR51E1 mRNA expression. Thus, either the undetectable levels or absence of OR51E1 in the normal liver, which is a major metastatic organ for SI-NET

31
patients, keeps the possibility to potentially develop OR51E1 protein as a therapeutic target in the future.
We next investigated the potential co-localization of OR51E1 and VMAT1. Our findings indicated that 60% of EC cells lying in the mucosa adjacent to the tumor expressed OR51E1, whereas 40% of the EC cells were non-IR for OR51E1. We assumed that there may be two subpopulations of EC cells, which are characterized by either the presence or the lack of OR51E1 receptor expression. Indeed, similar results were obtained from the co-localization studies in the tumor specimens, where 64% of the VMAT1-IR tumor cells express OR51E1, whereas 91% of OR51E1-IR tumor cells co-localize with VMAT1. We suggested that the neoplastic cells, which express VMAT1 and OR51E1 may derive from the EC cells, which show OR51E1 immunoreactivity, whereas the tumor cells, which uniquely express VMAT1, may originate from different non-IR OR51E1 EC cells.
Paper III. OR51E1 is highly expressed in somatostatin receptor negative typical and atypical lung carcinoids We confirmed the wild-type OR51E1 coding sequence in established human TC-NCI-H727 cells and AC-NCI-H720 cells. Furthermore, QRT-PCR analysis revealed OR51E1 transcript expression in the two cell lines and in 9 out of 12 TCs and 7 out of 9 ACs. Higher OR51E1 expression was clearly detected in TC and AC tumor cells compared to the normal surrounding tissue.
Our immunohistochemical results were evaluated based on the scoring system, which is described in the Materials and Methods section of Paper III. OR51E1 protein was highly expressed mainly in the tumor cells membrane of primary LCs and metastases. OR51E1, SSTR2, SSTR3 and SSTR5 were detected in 85%, 71%, 25% and 39% of primary TCs and in 86%, 79%, 43% and 36% of primary ACs. In summary, the Score 0 (S0) group mainly expressed the SSTRs, whereas the Score 3 (S3) group OR51E1. The Score 1 (S1) and Score 2 (S2) groups included intermediate distributions of both OR51E1 and SSTRs. Furthermore, 79% primary TCs had OR51E1 higher or equal scores than each SSTR2, SSTR3 and SSTR5 (p=0.0083) score, whereas primary ACs percentage was 86%. No statistical difference in the OR51E1 scores and in each SSTR score, was detected between primary TCs and ACs or between primary LCs and LC metastases or between smaller (< 3cm) and larger (≥ 3cm) primary LCs. Thus, OR51E1 and each SSTR were distributed uniformly across the entire spectrum of LCs.
Furthermore, the analysis of 22 SSTR-non-IR (S0) lesions revealed that membrane localization (S2 or S3) of OR51E1 was detected in the tumor cells in 10 out of 17 primary TCs, 2 out of 3 TC lymph node metastases and 1 out of 2 primary ACs. In the 14 LC lesions where SSTR2 was predominantly localized in the cytoplasm (S1), 7 out of 9 primary TCs, 2 out

32
of 3 TC lymph node metastases; and 2 out of 2 primary ACs showed membrane OR51E1 immunohistochemical pattern (S2 or S3).
Moreover, we analyzed the LC lesions for which OctreoScan data were available. We showed that OR51E1 scores (S2-S3) were higher than SSTR2, SSTR3 and SSTR5 (S0-S1) in 5 out of 6 OctreoScan-negative TC lesions (5 primary tumors and 1 lymph node metastasis). OR51E1 also showed high scores (S2-S3) in 5 out of 6 OctreoScan grade 1 lesions and in 7 out of 7 OctreoScan grade 2-4 lesions, which were graded according to Kwekkeboom et al [120] (p=0.042 for OR51E1 vs. SSTRs, in all the above-mentioned 19 lesions). These findings clearly revealed that OR51E1 immunoreactivity was detected throughout all the tumors regardless the tracer uptake levels.
The OR51E1 membrane immunoreactivity may be essential in NET diagnosis, which is indicated mainly by the clinical relevance of membrane localization of other G-protein coupled receptors, such as SSTRs. Indeed, a standardized and reliable clinical report of SSTRs expression should consider their immunoreactivity in the tumor cellular membrane, rather than in the cytoplasm [106]. In our study, OR51E1 has shown more extensive membrane localization compared to SSTR2, SSTR3 and SSTR5 (Figure 6), which are targeted by most of the clinically available SSAs. These findings indicated that OR51E1 might offer a higher uptake in LC cells, compared to the SSA-based diagnostics. Indeed, OR51E1 has shown an advantage over SSTRs since it has prevalence across the entire spectrum of LCs, even in OctreoScan-negative and/or SSTR non-IR LCs. This increases the opportunity to use OR51E1 as an additional marker to SSTRs in diagnosing these tumors. However, it depends on the potential development of high-affinity OR51E1 monoclonal antibodies, which are required to develop a novel immuno-PET [121].
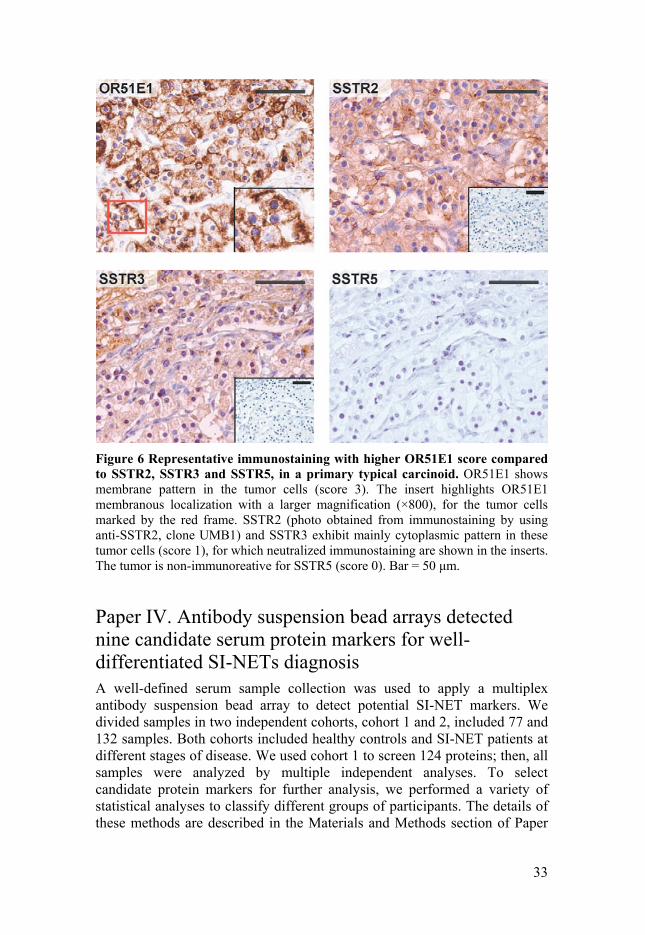
33
Figure 6 Representative immunostaining with higher OR51E1 score compared to SSTR2, SSTR3 and SSTR5, in a primary typical carcinoid. OR51E1 shows membrane pattern in the tumor cells (score 3). The insert highlights OR51E1 membranous localization with a larger magnification (×800), for the tumor cells marked by the red frame. SSTR2 (photo obtained from immunostaining by using anti-SSTR2, clone UMB1) and SSTR3 exhibit mainly cytoplasmic pattern in these tumor cells (score 1), for which neutralized immunostaining are shown in the inserts. The tumor is non-immunoreative for SSTR5 (score 0). Bar = 50 μm.
Paper IV. Antibody suspension bead arrays detected nine candidate serum protein markers for well-differentiated SI-NETs diagnosis A well-defined serum sample collection was used to apply a multiplex antibody suspension bead array to detect potential SI-NET markers. We divided samples in two independent cohorts, cohort 1 and 2, included 77 and 132 samples. Both cohorts included healthy controls and SI-NET patients at different stages of disease. We used cohort 1 to screen 124 proteins; then, all samples were analyzed by multiple independent analyses. To select candidate protein markers for further analysis, we performed a variety of statistical analyses to classify different groups of participants. The details of these methods are described in the Materials and Methods section of Paper

34
IV. We then selected 20 proteins, which were further investigated in the larger cohort 2. After an independent analysis on the second cohort, we identified 6 proteins, IGFBP2, SHKBP1, ETS1, IGF1, IL1α and STX2, which correctly classified all SI-NETs from healthy controls and exhibited a classification accuracy of 86% (sensitivity=92%, specificity=72%).
Since biomarker signatures should ideally be disease-stage specific, we compared the data from individuals at different stages (primary tumor, PT; lymph node metastasis, LNM and liver metastasis, LM) to healthy controls. These analyses showed that different targets such as, IGF1, IL1α, SHKBP1, and EGR3 are pivotal to classify SI-NETs at the stage of PT; IL1α, XIAP, STX2 and SHKBP1 classify LNM patients, whereas IGF1, IL1α, IGFBP2, MAML3 and SHKBP1 classify LM patients.
Although the antibody suspension bead array used in this study cannot exclude potential off-target binding events, dependent on weak affinity interactions between an antibody and other abundant proteins, we increased our confidence in on-target binding through (i) multiple independent analyses of the same sample cohort, (ii) analysis of additional sample cohort and (iii) using several antibodies per target protein.
In addition, since the sensitivity of an assay is highly dependent on the antibody characteristics (e.g. target affinity, functionality as capture reagent) as well as on the antigen (e.g. accessibility, stability, modification), we verified our findings by analyzing a subset of patients and healthy controls from cohort 2 using commercially available sandwich assays for IGF1 and IGFBP2. In conclusion, we identified, by using a novel antibody suspension bead array, nine candidate serum protein markers, namely IGFBP2, IGF1, SHKBP1, ETS1, IL1α, STX2, MAML3, EGR3 and XIAP, which significantly distinguished healthy donors from well-differentiated SI-NET patients at different stages of disease.

35
Concluding Remarks and Perspectives
SI-NET and LC patients’ clinical workup has been significantly improved during the last few decades. However, these malignancies have usually metastasized at diagnosis. This lowers the curative surgical intervention to rare events. Thus, the heterogeneity of these tumors, which results in various clinical presentations, established the unmet need of detecting novel diagnostic, prognostic and therapeutic tools. Indeed, our scientific work aimed at exploring several novel circulating and tissue biomarkers, to consider their potential clinical development; and it was performed in collaboration with several national and international contributors.
We first showed that high Ma2 autoantibody titer in the blood of SI-NET patients was a sensitive and specific biomarker, superior to CgA for the risk of tumor progression and tumor recurrence detection, after radical operation with a curative intent, as reported in Paper I. We then showed that OR51E1 protein was highly expressed in SI-NETs at different stages of disease, which suggested its potential therapeutic molecular target development, as reported in Paper II. Furthermore, we extended our findings on OR51E1, by showing that OR51E1 was highly cell membrane expressed across the entire spectrum of LCs, irrespectively from the SSTR expression status and the tumor size. This was of major importance and suggested that OR51E1 may become a potential novel diagnostic target in somatostatin receptor negative LC tumors, as discussed in Paper III. Moreover, we proposed nine novel potential serum protein markers, namely IL1α, SHKBP1, IGF1, IGFBP2, STX2, ETS1, MAML3, EGR3 and XIAP, to classify well-differentiated SI-NETs at different stages, by using an innovative multiplex approach, as reported in Paper IV. Looking at the potential breakthrough in the field of NETs, which came out from our experimental work, some novel research projects can be addressed as follows.
NET tissue biomarkers, such as enzymes and cell surface receptors, might be overexpressed in the tumor cells compared to the normal epithelial cells during NET tumorigenesis and tumor progression. We are aware of different diagnostic and therapeutic regimens, which have been developed by using the uptake of a variety of radiolabelled peptide analogues or chemical reagents. Since we showed that OR51E1 was a highly expressed protein in SI-NET and LC cells, this protein may be further developed as a candidate target i) for immuno-PET [121] to improve early NET diagnosis; ii) to establish patient stratification and to follow the progression of SI-NETs. This

36
clearly represents a long term project, which requires a strong financial support. However, the initial steps may be briefly summarized in two needed aspects to start with. First, specific recombinant monoclonal OR51E1 antibody development and second, the establishment of xenograft models to perform in vitro and in vivo studies by using established NET cell lines and/or primary NET cell cultures. The success of these initial steps may support the interest in proceeding as suggested. Hopefully, the eventual OR51E1 development for novel targeted solid tumor radioimmunotherapy [122], in combination with other radiopharmaceuticals, cytotoxic drugs or radio sensitizers [123], may improve the management of SI-NET patients. Although it is essential to investigate whether OR51E1 has a significant role in LC cell growth, OR51E1 development may hopefully be pivotal for the inoperable SSTR negative LCs, which urgently request novel effective therapeutic options, following the same approach suggested above for SI-NET patients.
NET circulating markers, which are different biochemical entities such as the circulating DNA, RNA, proteins and autoantibodies, may result from natural responses to several phenomena. I would like briefly reminding of their importance during tumorigenesis, tumor progression, recurrence, therapeutic response or other pathogenesis process, which develop inside a whole-body. The significant advantages of this type of markers are that they may be measured via a minimally invasive approach and may reflect an early event undetectable in the tissue. Indeed, our finding that Ma2 autoantibody titers are a more sensitive marker than circulating CgA, during SI-NET progression and recurrence, has significantly added a possibility to develop prognostic tests and improve the surveillance and stratification of patients during follow-ups. However, broader analyses in different patient cohorts and the use of different methods to repeat the experimental results are requested before clinical development.
In addition, the novel detection of a panel of nine candidate serum protein markers, may unveil potential malignancy pathogenesis processes, by further studying different molecular signaling pathways. However, our study has offered a great opportunity to use modern high-throughput proteomic approaches, such as a novel antibody suspension bead array to discover and validate circulating NET markers. Since the combination of different biomarkers as a panel may improve diagnostic sensitivity, specificity and accuracy, the application of multiplex proteomic approaches would hopefully improve NET patients’ clinical management.

37
Abstract in Chinese (中文摘要)
小肠神经内分泌肿瘤(SI-NET)和肺类癌(LC)是起源于位于弥散神经内分泌系统
中的神经内分泌细胞的相对生长缓慢的肿瘤。由于肿瘤可于诊断之前转移,当
前有效的治疗方法仍然匮乏,这同时也使得对于新的生物标记物的研发变得有
意义。因此,我们在本论文中分别研究了Ma2自身抗体(抗Ma2),以及潜在的
新型生物标记物嗅觉受体51E1(OR51E1)。我们还探讨了患者血清中的其他候
选蛋白标记物。 首先,我们建立了一个灵敏特异而可靠的抗Ma2间接酶联免疫吸附试验,用
以区分SI-NET患者组和健康对照组。在表达低滴度抗Ma2的患者中,我们检测
到了较长的病情无恶化存活率以及肿瘤无复发存活率。此外,高滴度抗Ma2比嗜铬粒蛋白A更为灵敏地检测到了SI-NET患者根治手术后复发的风险。 接下来,我们研究了SI-NET和LC患者肿瘤中的OR51E1受体蛋白的表达。我
们用实时定量PCR技术检测到了OR51E1信使核糖核酸在显微切除的SI-NET肿
瘤细胞中,以及在LC细胞系和冷冻LC标本中的高度表达。免疫组化结果显示出
OR51E1蛋白在SI-NET肿瘤组织中的高度表达。OR51E1与囊泡单胺转运蛋白1在大多数正常和肿瘤的肠嗜铬细胞中可共表达。
另外,我们针对LC患者的研究显示,OR51E1受体蛋白以及促生长素抑制素
受体(SSTR)2,SSTR3和SSTR5分别在85%,71%,25%和39%的典型性肺类
癌(TC),以及86%,79%,43%和36的非典型性肺类癌(AC)中表达。基于我们
我提出的免疫组化结果得分系统,在无SSTR表达的LC中,OR51E1蛋白在17个TC中的10个以及2个AC中的1个中呈细胞膜表达。而且,在6个OctreoScan显象呈阴性的LC中,有5个OR51E1免疫组化得分很高。 此外,在本论文最后的一项研究中,我们采用了一种新型的悬浮磁珠阵列
技术,通过使用来自于人类蛋白质图谱项目的针对124种独特蛋白质的抗体,
对SI-NET患者和健康对照组的用生物素标记过的血清样本进行了分析。结果
显示,通过利用9种蛋白,即IGFBP2,IGF1,SHKBP1,ETS1,STX2,IL1α,MAML3,EGR3和XIAP,我们可以显著的对肿瘤进行分类。 综上所述,我们提出Ma2自身抗体可作为一个体液中灵敏的生物标记物用以
暗示SI-NET肿瘤的复发; OR51E1受体蛋白可作为一个在SI-NET治疗中所能用
及的候选生物靶分子,并在LC中作为一种新型的潜在生物标记物。此外,我
们在SI-NET患者血清中检测到了9种新的候选标记物蛋白。

38
Acknowledgements
The work included in my thesis was performed mainly at the Department of Medical Sciences, Endocrine Oncology, Uppsala University, Uppsala; and partly at the Science for Life Laboratory, School of Biotechnology, KTH - Royal Institute of Technology, Stockholm. The work was funded mainly by Lions Cancerforskningsfond, Tore Nilssons Stiftelse för Medicinsk Forskning and Erik, Karin och Gösta Selander Stiftelse; and partly by Science for Life Laboratory Stockholm, ProNova VINN Excellence Centre for Protein Technology and the Knut and Alice Wallenberg Foundation.
I would like sincerely appreciating all the people that directly or indirectly contributed to this work and supported me through the years:
Assoc. Prof. Valeria Giandomenico, my main supervisor. You are definitely much more than a supervisor. You are a precise scientist, an inspiring captain, a strategic, humorous and “sweet” mentor, a default word corrector, a warmhearted friend, a talkative philosopher and a “queen of cats”. You have been supporting me really everywhere at your best, since a normal sunshine afternoon in March, 2008. No words can simply express my thanks; however, I will say anyway - “thank you and grazie”!
Prof. Kjell Öberg, my co-supervisor. To me, you are a mountain in the field of neuroendocrine tumors. However, you are not just a piece of hard rock. You have magnificent and meandering ridges that gently embrace any small creatures in your arms, with your typical Swedish smiles. You show them what a peak looks like and how to climb there to get closer to the sun!
Dr. Apostolos V. Tsolakis, my co-supervisor. Till the end of life a silkworm keeps spinning silk, till burning itself out a candle goes on lighting us. Yes, you are such a hard worker. Being a medical doctor and a scientist gives one almost no time to supervise students in the meantime; however, nothing is impossible for you!
Prof. Eva Tiensuu Janson, the head of Medical Science Department. Thanks to you and your predecessors that have established not only a renowned clinical center but a wonderful working place!
Su-Chen Li, my closest colleague and friend since master program. I enjoy our daily scientific discussions. Thanks for your delicious and delicate foods and your company at work during the last five years! …and of course, also Shangfeng ge (brother), thanks for creating a nice social environment in Uppsala. I wish your beloved 家和 (Carl) grow happily and healthily!
Åsa Forsberg, our previous technician. Thanks for taking daily care of my technical questions and ordering requests during the first three years of my PhD program!
Rakan Naboulsi, my lab mate and friend. Bro, I enjoyed the never-ending discussions with you in science and jokes in life. Moreover, we absolutely need to go to Camp Nou together one day to watch a real match!

39
Dr. Spyros Darmanis, my wonderful collaborator, coauthor and friend. Thank you for sharing your expertise on novel technologies, statistical analyses, experimental design and biomarker development. Moreover, thanks for the significant contribution to my thesis with our nice collaborative work!
Monica Hurtig, our research nurse. You are one of the most efficient workers that I have ever seen. Thanks for the always on-time blood sample collections and the precise clinical information!
Dr. Graciela Elgue, my ELISA teacher. Your humor offered me a relaxed working environment when I was still an early bird at labs in Sweden. You are such an optimistic scientist who is always enjoying lab works and music; and look forward!
Prof. Giuseppe Pelosi, our fantastic collaborator and coauthor in Milan. It was a nice talk with you when we were in Barcelona! Thanks for the precious plasma samples from lung carcinoid patients!
Prof. Jean-Beaptiste Demoulin and Dr, Ahmed Essaghir, our great collaborators and coauthors in Belgium. Your expertise in the statistical analyses made the beginning of my PhD program easier, which might be a tough period for every student!
Dr. Mohammad Alimohammadi, my coauthor. You are such a kind and smiling person. Thanks for teaching me immunoprecipitation and sequential immunoprecipitation! ...and Assoc. Prof. Angelica Loskog at IGP, Rudbeck, thanks for the Jurkat cells. Do you still remember?
Dr. Janet L. Cunningham, my coauthor. Thanks for the time that we spent in front of the microscope together!
Assoc. Prof. Thomas Lind, my coauthor and friend. I enjoyed discussing any scientific question with you. Thanks for your company during lunch times, especially with your nice Swedish smile!
Prof. Lars Grimelius, my coauthor. I deeply appreciate the precious experience to work with a respectable pathologist like you!
Prof. Jochen M. Schwenk, Prof. Peter Nilsson and Dr. Kimi Drobin, my coauthors. Thanks for sharing your expertise during our collaboration!
Prof. Magnus Essand, our best national collaborator. I met you when I just started to work at Rudbeck and since then you have generously helped me in uncountable occasions wherever. Beyond science, you are the best civilian musician that I have ever seen!
Dr. Di Yu, my best friend in Magnus’s group, who never said no to me. You were about to call me “Shixiong” (senior male student in lab) but you did not. Luck you! ...and of course also Chuan Jin; it is weird to write in the reverse way of a familiar Chinese name like yours. Let’s keep this friendship no matter in Sweden or China!
Dr. Justyna Leja, Dr. Berith Nilsson and all the other previous colleagues at IGP Department, Rudbeck. Thanks for all your previous experimental supports!
Dr. Jan Grawé, our collaborator at Rudbeck. Thanks for teaching me laser capture microdissection!

40
Dr. Malin Grönberg, my colleague. Thanks for your ordering supports and all other supports through the years. I always enjoy talking with you!
Assoc. Prof. Dominic-Luc Webb and Prof. Per Hellström, our colleagues and collaborators. Thanks for providing expert knowledge and great collaboration in the study of gastroenterological disorders!
Dr. Cihan Cetinkaya, my colleague and friend. To work in the cell culture room can be very boring; however, it has never happened to me when you were there – in a shoe box office next to my engine-like ventilation sound from the hood!
Dr. Cécile Martijin, my colleague and friend. I started to miss your endless jokes and “bad” smiles since you left this corridor! ...and also your beloved husband, Hank; thanks to both of you for the sunny and rainy day in your home with your marvelous international foods!
Dr. Chuan Wang, my friend. Thanks for everything that you helped at work and in life during the last years. You were the first Chinese that I saw in Sweden and you lent me your telephone to call my home when I just landed on Aug 22, 2007!
To all other past and present colleagues at the Endocrine Oncology Section, to Dr. Andreea Barbu, Dr. Margareta Halin Lejonklou, Xia Chu, Prof. Gunnar Westin, Katarina Edfeldt and to all the other colleagues and friends working in our corridors that have worked or discussed with me, thanks for creating a nice working environment!
Anders Söderberg, our IT technician. Thanks for helping me with all my computer problems!
My master program classmates and their beloved partners: Di Wu, Yanling Cai, Yang Sun, Yani Zhao, Jinghui Luo, Huixin Yu, Li Li, Yu Sun, Haisha Ma, Bo Xu, Sung-kyu Park and all the others. Thank you all for the time that we spent together through the years!
My “new” friends thanks to my wife: Xia Yang, Shuanglin Hu, Jing Ni, Xin Hu, Yu Zhang, Yuan Tian, Wei Xia, Miao Wu, Xin He, Shuyi Li and all the others. Thank you all for creating a joyful Chinese environment abroad!
Yang Jin and Jingwei Yan, my two earliest Chinese friends in Sweden. I always enjoy the endless jokes and chats among us especially during the dark Swedish winters!
My old friends in China, we did not lose anything through the years. Instead, we gained precious memories as a forever gift in life!
My mum and dad! Without you, I don’t even have myself as a human, far away to mention my scientific growth. You always support me since my childhood and I have never said thanks! 爸妈,没有你们我根本无法成人,何谈成才。你们从小到大一直支持着我,而我从来也
没有表达过感谢!
My parents-in-law, my uncles, aunts, grandparents and cousins. Thanks for your great supports forever! 我其他所有的家人们,感谢你们一直以来伟大的支持!
Fang, my cute and beloved wife! You always support me wherever. I feel weird to say “thank you” to you so I decided to write rather than to say. Will you generously accept it? Take your hand to spend the whole life together with you! 昉, 我的可爱的心爱的老婆, 你无时无刻地支持
着我。我觉得对你说谢谢很奇怪,所以我决定写而不是说。你能大方地接受我的谢意吗?执子之
手,与子偕老!

41
References
1. Klöppel, G., Tumour biology and histopathology of neuroendocrine tumours. Best
Pract Res Clin Endocrinol Metab, 2007. 21: p. 15-31. 2. Wiedenmann, B., et al., Molecular and cell biological aspects of neuroendocrine
tumors of the gastroenteropancreatic system. J Mol Med (Berl), 1998. 76: p. 637-47. 3. Modlin, I.M., et al., Chromogranin A--biological function and clinical utility in
neuro endocrine tumor disease. Ann Surg Oncol, 2010. 17: p. 2427-43. 4. Rindi, G., et al., The "normal" endocrine cell of the gut: changing concepts and new
evidences. Ann N Y Acad Sci, 2004. 1014: p. 1-12. 5. Portela-Gomes, G.M., et al., Co-localization of synaptophysin with different
neuroendocrine hormones in the human gastrointestinal tract. Histochem Cell Biol, 1999. 111: p. 49-54.
6. Wiedenmann, B. and W.B. Huttner, Synaptophysin and chromogranins/secretogranins--widespread constituents of distinct types of neuroendocrine vesicles and new tools in tumor diagnosis. Virchows Arch B Cell Pathol Incl Mol Pathol, 1989. 58: p. 95-121.
7. Manfé, A.Z., et al., Usefulness of chromogranin A, neuron-specific enolase and 5-hydroxyindolacetic acid measurements in patients with malignant carcinoids. In Vivo, 2011. 25: p. 1027-9.
8. Wimalasena, K., Vesicular monoamine transporters: structure-function, pharmacology, and medicinal chemistry. Med Res Rev, 2011. 31: p. 483-519.
9. Modlin, I.M., et al., Gastroenteropancreatic neuroendocrine tumours. Lancet Oncol, 2008. 9: p. 61-72.
10. Racke, K., et al., Regulation of 5-HT release from enterochromaffin cells. Behav Brain Res, 1996. 73: p. 83-7.
11. Bertrand, P.P. and R.L. Bertrand, Serotonin release and uptake in the gastrointestinal tract. Auton Neurosci, 2010. 153: p. 47-57.
12. Rahman, S., I.A. Khan, and P. Thomas, Tryptophan hydroxylase: a target for neuroendocrine disruption. J Toxicol Environ Health B Crit Rev, 2011. 14: p. 473-94.
13. Vikman, S., et al., Gene expression in midgut carcinoid tumors: potential targets for immunotherapy. Acta Oncol, 2005. 44: p. 32-40.
14. Gilbert, J.A., L.A. Bates, and M.M. Ames, Elevated aromatic-L-amino acid decarboxylase in human carcinoid tumors. Biochem Pharmacol, 1995. 50: p. 845-50.
15. Öberg, K., Neuroendocrine Gastrointestinal and Lung Tumors (Carcinoid Tumors), Carcinoid Syndrome, and Related Disorders, in Williams Textbook of Endocrinology, K.S.P. Shlomo Melmed, P. Reed Larsen, Henry M. Kronenberg, Editor 2011, Elsevier: Philadelphia, USA. p. 1813.
16. Lawrence, B., et al., The epidemiology of gastroenteropancreatic neuroendocrine tumors. Endocrinol Metab Clin North Am, 2011. 40: p. 1-18, vii.
17. Scherübl, H., et al., Management of early gastrointestinal neuroendocrine neoplasms. World J Gastrointest Endosc, 2011. 3: p. 133-9.

42
18. Rindi G, A.R., Bosman FT, Capella C, Klimstra DS, Klöppel G, Nomenclature and classification of neuroendocrine neoplasms of the digestive system, in WHO Classification of tumours of the digestive tract, C.F. Bosman FT, Hruban RH, Theise ND, Editor 2010, IARC Press: Lyon, France. p. 13-14.
19. Rindi, G., et al., TNM staging of foregut (neuro)endocrine tumors: a consensus proposal including a grading system. Virchows Arch, 2006. 449: p. 395-401.
20. Rindi, G., et al., TNM staging of midgut and hindgut (neuro) endocrine tumors: a consensus proposal including a grading system. Virchows Arch, 2007. 451: p. 757-62.
21. Oberg, K. and D. Castellano, Current knowledge on diagnosis and staging of neuroendocrine tumors. Cancer Metastasis Rev, 2011. 30 Suppl 1: p. 3-7.
22. Spiller, R., Serotonin and GI clinical disorders. Neuropharmacology, 2008. 55: p. 1072-80.
23. Kwekkeboom, D.J., et al., ENETS Consensus Guidelines for the Standards of Care in Neuroendocrine Tumors: somatostatin receptor imaging with (111)In-pentetreotide. Neuroendocrinology, 2009. 90: p. 184-9.
24. Kroiss, A., et al., 68Ga-DOTA-TOC uptake in neuroendocrine tumour and healthy tissue: differentiation of physiological uptake and pathological processes in PET/CT. Eur J Nucl Med Mol Imaging, 2013. 40: p. 514-23.
25. Putzer, D., et al., Somatostatin receptor PET in neuroendocrine tumours: 68Ga-DOTA0,Tyr3-octreotide versus 68Ga-DOTA0-lanreotide. Eur J Nucl Med Mol Imaging, 2013. 40: p. 364-72.
26. Oberg, K., Molecular Imaging Radiotherapy: Theranostics for Personalized Patient Management of Neuroendocrine Tumors (NETs). Theranostics, 2012. 2: p. 448-58.
27. Ahlstrom, H., et al., Pancreatic neuroendocrine tumors: diagnosis with PET. Radiology, 1995. 195: p. 333-7.
28. Orlefors, H., et al., Positron emission tomography with 5-hydroxytryprophan in neuroendocrine tumors. J Clin Oncol, 1998. 16: p. 2534-41.
29. Sundin, A., Radiological and nuclear medicine imaging of gastroenteropancreatic neuroendocrine tumours. Best Pract Res Clin Gastroenterol, 2012. 26: p. 803-18.
30. Oberg, K., Gallium-68 somatostatin receptor PET/CT: is it time to replace (111)Indium DTPA octreotide for patients with neuroendocrine tumors? Endocrine, 2012. 42: p. 3-4.
31. Yao, J.C., et al., Everolimus for advanced pancreatic neuroendocrine tumors. N Engl J Med, 2011. 364: p. 514-23.
32. Rinke, A., et al., Placebo-controlled, double-blind, prospective, randomized study on the effect of octreotide LAR in the control of tumor growth in patients with metastatic neuroendocrine midgut tumors: a report from the PROMID Study Group. J Clin Oncol, 2009. 27: p. 4656-63.
33. Kölby, L., et al., Randomized clinical trial of the effect of interferon alpha on survival in patients with disseminated midgut carcinoid tumours. Br J Surg, 2003. 90: p. 687-93.
34. Yao, J.C., D.R. Lagunes, and M.H. Kulke, Targeted Therapies in Neuroendocrine Tumors (NET): Clinical Trial Challenges and Lessons Learned. Oncologist, 2013. 18: p. 525-32.
35. Pavel, M.E., et al., Everolimus plus octreotide long-acting repeatable for the treatment of advanced neuroendocrine tumours associated with carcinoid syndrome (RADIANT-2): a randomised, placebo-controlled, phase 3 study. Lancet, 2011. 378: p. 2005-12.
36. National Cancer Institute. FDA approval for everolimus. National Cancer Institute Website 2011. Available at: http://www.cancer.gov/cancertopics/druginfo/fda-everolimus. Accessed Feb 28, 2012.

43
37. Falconi, M., et al., ENETS Consensus Guidelines for the management of patients with digestive neuroendocrine neoplasms of the digestive system: well-differentiated pancreatic non-functioning tumors. Neuroendocrinology, 2012. 95: p. 120-34.
38. van Essen, M., et al., Peptide receptor radionuclide therapy with 177Lu-octreotate in patients with foregut carcinoid tumours of bronchial, gastric and thymic origin. Eur J Nucl Med Mol Imaging, 2007a. 34: p. 1219-27.
39. Kam, B.L., et al., Lutetium-labelled peptides for therapy of neuroendocrine tumours. Eur J Nucl Med Mol Imaging, 2012. 39 Suppl 1: p. S103-12.
40. Vinjamuri, S., et al., Peptide receptor radionuclide therapy with (90)Y-DOTATATE/(90)Y-DOTATOC in patients with progressive metastatic neuroendocrine tumours: assessment of response, survival and toxicity. Br J Cancer, 2013. 108: p. 1440-8.
41. Bodei, L., et al., Yttrium-labelled peptides for therapy of NET. Eur J Nucl Med Mol Imaging, 2012. 39 Suppl 1: p. S93-102.
42. Kwekkeboom, D.J., W.W. de Herder, and E.P. Krenning, Somatostatin receptor-targeted radionuclide therapy in patients with gastroenteropancreatic neuroendocrine tumors. Endocrinol Metab Clin North Am, 2011. 40: p. 173-85, ix.
43. van Vliet, E.I., et al., Treatment of gastroenteropancreatic neuroendocrine tumors with peptide receptor radionuclide therapy. Neuroendocrinology, 2013. 97: p. 74-85.
44. Dong, M., A.T. Phan, and J.C. Yao, New strategies for advanced neuroendocrine tumors in the era of targeted therapy. Clin Cancer Res, 2012. 18: p. 1830-6.
45. Van Lommel, A., Pulmonary neuroendocrine cells (PNEC) and neuroepithelial bodies (NEB): chemoreceptors and regulators of lung development. Paediatr Respir Rev, 2001. 2: p. 171-6.
46. Van Lommel, A., et al., The pulmonary neuroendocrine system: the past decade. Arch Histol Cytol, 1999. 62: p. 1-16.
47. Otto, W.R., Lung epithelial stem cells. J Pathol, 2002. 197: p. 527-35. 48. Beasley, M.B., et al., Carcinoid tumour, in World Health Organization
Classification of Tumours. Pathology and Genetics of Tumours of the Lung, Pleura, Thymus and Heart, W.D. Travis, et al., Editors. 2004, IARC Press: Lyon, France. p. 59-62.
49. Travis, W.D., Lung tumours with neuroendocrine differentiation. Eur J Cancer, 2009. 45 Suppl 1: p. 251-66.
50. Said, J.W., et al., Immunoreactive neuron-specific enolase, bombesin, and chromogranin as markers for neuroendocrine lung tumors. Hum Pathol, 1985. 16: p. 236-40.
51. Sturm, N., et al., Expression of thyroid transcription factor-1 in the spectrum of neuroendocrine cell lung proliferations with special interest in carcinoids. Hum Pathol, 2002. 33: p. 175-82.
52. Oliveira, A.M., et al., Thyroid transcription factor-1 distinguishes metastatic pulmonary from well-differentiated neuroendocrine tumors of other sites. Am J Surg Pathol, 2001. 25: p. 815-9.
53. Du, E.Z., et al., TTF-1 expression is specific for lung primary in typical and atypical carcinoids: TTF-1-positive carcinoids are predominantly in peripheral location. Hum Pathol, 2004. 35: p. 825-31.
54. Righi, L., et al., Somatostatin receptor tissue distribution in lung neuroendocrine tumours: a clinicopathologic and immunohistochemical study of 218 'clinically aggressive' cases. Ann Oncol, 2010. 21: p. 548-55.
55. Pelosi, G., et al., Peptide receptors in neuroendocrine tumors of the lung as potential tools for radionuclide diagnosis and therapy. Q J Nucl Med Mol Imaging, 2006. 50: p. 272-87.

44
56. Papotti, M., et al., Expression of somatostatin receptor types 2, 3 and 5 in biopsies and surgical specimens of human lung tumours. Correlation with preoperative octreotide scintigraphy. Virchows Arch, 2001. 439: p. 787-97.
57. Filosso, P.L., et al., Bronchial carcinoid tumors: surgical management and long-term outcome. J Thorac Cardiovasc Surg, 2002a. 123: p. 303-9.
58. Gustafsson, B.I., et al., Bronchopulmonary neuroendocrine tumors. Cancer, 2008. 113: p. 5-21.
59. Filosso, P.L., et al., Long-term survival of atypical bronchial carcinoids with liver metastases, treated with octreotide. Eur J Cardiothorac Surg, 2002b. 21: p. 913-7.
60. van Essen, M., et al., Peptide Receptor Radionuclide Therapy with radiolabelled somatostatin analogues in patients with somatostatin receptor positive tumours. Acta Oncol, 2007b. 46: p. 723-34.
61. Khan, M.S., et al., Circulating tumor cells and EpCAM expression in neuroendocrine tumors. Clin Cancer Res, 2011. 17: p. 337-45.
62. Khan, M.S., et al., Circulating tumor cells as prognostic markers in neuroendocrine tumors. J Clin Oncol, 2013. 31: p. 365-72.
63. Modlin, I.M., et al., Principal component analysis, hierarchical clustering, and decision tree assessment of plasma mRNA and hormone levels as an early detection strategy for small intestinal neuroendocrine (carcinoid) tumors. Ann Surg Oncol, 2009. 16: p. 487-98.
64. Li, S.C., et al., Global microRNA profiling of well-differentiated small intestinal neuroendocrine tumors. Mod Pathol, 2013. 26: p. 685-96.
65. Cleland, J.G., The renin-angiotensin system in heart failure. Herz, 1991. 16: p. 68-81.
66. Kaltsas, G.A., et al., Expression of connective tissue growth factor and IGF1 in normal and neoplastic gastrointestinal neuroendocrine cells and their clinico-pathological significance. Endocr Relat Cancer, 2011. 18: p. 61-71.
67. Sica, G., et al., PAX-5 expression in pulmonary neuroendocrine neoplasms: its usefulness in surgical and fine-needle aspiration biopsy specimens. Am J Clin Pathol, 2008. 129: p. 556-62.
68. Findeis-Hosey, J.J. and H. Xu, Insulin-like growth factor II-messenger RNA-binding protein-3 and lung cancer. Biotech Histochem, 2012. 87: p. 24-9.
69. Leja, J., et al., Novel markers for enterochromaffin cells and gastrointestinal neuroendocrine carcinomas. Mod Pathol, 2009. 22: p. 261-72.
70. Schüller, M., D. Jenne, and R. Voltz, The human PNMA family: novel neuronal proteins implicated in paraneoplastic neurological disease. J Neuroimmunol, 2005. 169: p. 172-6.
71. Voltz, R., et al., A serologic marker of paraneoplastic limbic and brain-stem encephalitis in patients with testicular cancer. N Engl J Med, 1999. 340: p. 1788-95.
72. Chapman, M.H., et al., Whole genome RNA expression profiling of endoscopic biliary brushings provides data suitable for biomarker discovery in cholangiocarcinoma. J Hepatol, 2012. 56: p. 877-85.
73. Tan, K.O., et al., MAP-1, a novel proapoptotic protein containing a BH3-like motif that associates with Bax through its Bcl-2 homology domains. J Biol Chem, 2001. 276: p. 2802-7.
74. Kaltsas, G., et al., Paraneoplastic syndromes secondary to neuroendocrine tumours. Endocr Relat Cancer, 2010. 17: p. R173-93.
75. Gozzard, P. and P. Maddison, Which antibody and which cancer in which paraneoplastic syndromes? Pract Neurol, 2010. 10: p. 260-70.
76. Lleo, A., et al., Definition of human autoimmunity--autoantibodies versus autoimmune disease. Autoimmun Rev, 2010. 9: p. A259-66.
77. Monstad, S.E., et al., Onconeural antibodies in sera from patients with various types of tumours. Cancer Immunol Immunother, 2009. 58: p. 1795-800.
78. Dalmau, J., et al., Clinical analysis of anti-Ma2-associated encephalitis. Brain, 2004. 127: p. 1831-44.

45
79. Castle, J., et al., Anti-Ma2-associated encephalitis with normal FDG-PET: a case of pseudo-Whipple's disease. Nat Clin Pract Neurol, 2006. 2: p. 566-72; quiz 573.
80. Mathew, R.M., et al., Orchiectomy for suspected microscopic tumor in patients with anti-Ma2-associated encephalitis. Neurology, 2007. 68: p. 900-5.
81. Hoffmann, L.A., et al., Anti-Ma and anti-Ta associated paraneoplastic neurological syndromes: 22 newly diagnosed patients and review of previous cases. J Neurol Neurosurg Psychiatry, 2008. 79: p. 767-73.
82. Maat, P., et al., Multiplex serology of paraneoplastic antineuronal antibodies. J Immunol Methods, 2013. 391: p. 125-32.
83. Archer, H.A., et al., Mesothelioma and anti-Ma paraneoplastic syndrome; heterogeneity in immunogenic tumours increases. Pract Neurol, 2013.
84. Leja, J., et al., Novel markers for enterochromaffin cells and gastrointestinal neuroendocrine carcinomas. Mod Pathol, 2009. 22: p. 261-72.
85. Buck, L. and R. Axel, A novel multigene family may encode odorant receptors: a molecular basis for odor recognition. Cell, 1991. 65: p. 175-87.
86. Olender, T., D. Lancet, and D.W. Nebert, Update on the olfactory receptor (OR) gene superfamily. Hum Genomics, 2008. 3: p. 87-97.
87. Zhang, X. and S. Firestein, Genomics of olfactory receptors. Results Probl Cell Differ, 2009. 47: p. 25-36.
88. Lappano, R. and M. Maggiolini, G protein-coupled receptors: novel targets for drug discovery in cancer. Nat Rev Drug Discov, 2011. 10: p. 47-60.
89. Ache, B.W. and J.M. Young, Olfaction: diverse species, conserved principles. Neuron, 2005. 48: p. 417-30.
90. Vanti, W.B., et al., Novel human G-protein-coupled receptors. Biochem Biophys Res Commun, 2003. 305: p. 67-71.
91. Weigle, B., et al., D-GPCR: a novel putative G protein-coupled receptor overexpressed in prostate cancer and prostate. Biochem Biophys Res Commun, 2004. 322: p. 239-49.
92. Fuessel, S., et al., Transcript quantification of Dresden G protein-coupled receptor (D-GPCR) in primary prostate cancer tissue pairs. Cancer Lett, 2006. 236: p. 95-104.
93. Wang, J., et al., The prostate-specific G-protein coupled receptors PSGR and PSGR2 are prostate cancer biomarkers that are complementary to alpha-methylacyl-CoA racemase. Prostate, 2006. 66: p. 847-57.
94. Weng, J., et al., PSGR2, a novel G-protein coupled receptor, is overexpressed in human prostate cancer. Int J Cancer, 2006. 118: p. 1471-80.
95. Nilsson, O., Profiling of ileal carcinoids. Neuroendocrinology, 2013. 97: p. 7-18. 96. Flegel, C., et al., Expression profile of ectopic olfactory receptors determined by
deep sequencing. PLoS One, 2013. 8: p. e55368. 97. Wang, J., et al., The prostate-specific G-protein coupled receptors PSGR and
PSGR2 are prostate cancer biomarkers that are complementary to alpha-methylacyl-CoA racemase. Prostate, 2006. 66: p. 847-57.
98. Matsueda, S., et al., Identification of prostate-specific G-protein coupled receptor as a tumor antigen recognized by CD8(+) T cells for cancer immunotherapy. PLoS One, 2012. 7: p. e45756.
99. Fujita, Y., et al., Deorphanization of Dresden G protein-coupled receptor for an odorant receptor. J Recept Signal Transduct Res, 2007. 27: p. 323-34.
100. Saito, H., et al., Odor coding by a Mammalian receptor repertoire. Sci Signal, 2009. 2: p. ra9.
101. Kidd, M., et al., Luminal regulation of normal and neoplastic human EC cell serotonin release is mediated by bile salts, amines, tastants, and olfactants. Am J Physiol Gastrointest Liver Physiol, 2008. 295: p. G260-72.
102. Braun, T., et al., Enterochromaffin cells of the human gut: sensors for spices and odorants. Gastroenterology, 2007. 132: p. 1890-901.

46
103. Pluznick, J.L., et al., Olfactory receptor responding to gut microbiota-derived signals plays a role in renin secretion and blood pressure regulation. Proc Natl Acad Sci U S A, 2013. 110: p. 4410-5.
104. Primeaux, S.D., H.D. Braymer, and G.A. Bray, High fat diet differentially regulates the expression of olfactory receptors in the duodenum of obesity-prone and obesity-resistant rats. Dig Dis Sci, 2013. 58: p. 72-6.
105. van Helden, Y.G., et al., Downregulation of Fzd6 and Cthrc1 and upregulation of olfactory receptors and protocadherins by dietary beta-carotene in lungs of Bcmo1-/- mice. Carcinogenesis, 2010. 31: p. 1329-37.
106. Volante, M., et al., Somatostatin receptor type 2A immunohistochemistry in neuroendocrine tumors: a proposal of scoring system correlated with somatostatin receptor scintigraphy. Mod Pathol, 2007. 20: p. 1172-82.
107. Schwenk, J.M., et al., Antibody suspension bead arrays within serum proteomics. J Proteome Res, 2008. 7: p. 3168-79.
108. Sjoberg, R., et al., Validation of affinity reagents using antigen microarrays. New biotechnology, 2011.
109. Schwenk, J.M., et al., Toward next generation plasma profiling via heat-induced epitope retrieval and array-based assays. Mol Cell Proteomics, 2010. 9: p. 2497-507.
110. Li, S.C., et al., The somatostatin analogue octreotide inhibits growth of small intestine neuroendocrine tumour cells. PloS one, 2012. 7: p. e48411.
111. Haggmark, A., et al., Classification of protein profiles from antibody microarrays using heat and detergent treatment. New biotechnology, 2012. 29: p. 564-70.
112. Ihaka, R. and R. Gentleman, R: a language for data analysis and graphics. Journal of Computional and Graphical Statistics, 1996. 5: p. 299-3214.
113. Welin, S., et al., Elevated plasma chromogranin A is the first indication of recurrence in radically operated midgut carcinoid tumors. Neuroendocrinology, 2009. 89: p. 302-7.
114. Fuessel, S., et al., Transcript quantification of Dresden G protein-coupled receptor (D-GPCR) in primary prostate cancer tissue pairs. Cancer Lett, 2006. 236: p. 95-104.
115. Weng, J., et al., PSGR2, a novel G-protein coupled receptor, is overexpressed in human prostate cancer. Int J Cancer, 2006. 118: p. 1471-80.
116. Weigle, B., et al., D-GPCR: a novel putative G protein-coupled receptor overexpressed in prostate cancer and prostate. Biochem Biophys Res Commun, 2004. 322: p. 239-49.
117. Xu, Z.Q., X. Zhang, and L. Scott, Regulation of G protein-coupled receptor trafficking. Acta Physiol (Oxf), 2007. 190: p. 39-45.
118. Tarin, D., Clinical and Biological Implications of the Tumor Microenvironment. Cancer Microenviron, 2012.
119. Cheng, S.B., et al., Retrograde transport of the transmembrane estrogen receptor, G-protein-coupled-receptor-30 (GPR30/GPER) from the plasma membrane towards the nucleus. Steroids, 2011. 76: p. 892-6.
120. Kwekkeboom, D.J., et al., Radiolabeled somatostatin analog [177Lu-DOTA0,Tyr3]octreotate in patients with endocrine gastroenteropancreatic tumors. J Clin Oncol, 2005. 23: p. 2754-62.
121. Knowles, S.M. and A.M. Wu, Advances in immuno-positron emission tomography: antibodies for molecular imaging in oncology. J Clin Oncol, 2012. 30: p. 3884-92.
122. Barbet, J., et al., Radiolabeled antibodies for cancer imaging and therapy. Methods Mol Biol, 2012. 907: p. 681-97.
123. Song, H. and G. Sgouros, Radioimmunotherapy of solid tumors: searching for the right target. Curr Drug Deliv, 2011. 8: p. 26-44.


Acta Universitatis UpsaliensisDigital Comprehensive Summaries of Uppsala Dissertationsfrom the Faculty of Medicine 929
Editor: The Dean of the Faculty of Medicine
A doctoral dissertation from the Faculty of Medicine, UppsalaUniversity, is usually a summary of a number of papers. A fewcopies of the complete dissertation are kept at major Swedishresearch libraries, while the summary alone is distributedinternationally through the series Digital ComprehensiveSummaries of Uppsala Dissertations from the Faculty ofMedicine.
Distribution: publications.uu.seurn:nbn:se:uu:diva-205570
ACTAUNIVERSITATIS
UPSALIENSISUPPSALA
2013












used toselectasubsetofthestrongestpredictorsassociated](https://img.pdfslide.net/doc/110x75/5f9ef868f702b9462033eac2/circulating-biomarkers-may-be-unable-to-detect-infection-1064-modeling-strategy.jpg)
















