Embed Size (px)
Citation preview

eCAM 2007;Page 1 of 13doi:10.1093/ecam/nem138
Review
Novel Diagnosis of Lyme Disease: Potential for CAM Intervention
Aristo Vojdani1, Frank Hebroni2, Yaniv Raphael3, Jonathan Erde4 and Bernard Raxlen5
1Immunosciences Laboratory, Inc., Beverly Hills, CA 90211, 2Department of Psychobiology, University ofCalifornia, 3Department of Biology, University of Southern California, 4Department of Biochemistry,University of California and 5Raxlen Clinic, New York, NY 10024, USA
Lyme disease (LD) is the most common tick-borne disease in the northern hemisphere,producing a wide range of disabling effects on multiple human targets, including the skin, thenervous system, the joints and the heart. Insufficient clinical diagnostic methods, the necessityfor prompt antibiotic treatment along with the pervasive nature of infection impel thedevelopment and establishment of new clinical diagnostic tools with increased accuracy,sensitivity and specificity. The goal of this article is 4-fold: (i) to detail LD infection andpathology, (ii) to review prevalent diagnostic methods, emphasizing inherent problems, (iii) tointroduce the usage of in vivo induced antigen technology (IVIAT) in clinical diagnostics and(iv) to underscore the relevance of a novel comprehensive LD diagnostic approach topractitioners of Complementary and Alternative Medicine (CAM). Utilization of this analyticalmethod will increase the accuracy of the diagnostic process and abridge the time to treatment,with antibiotics, herbal medicines and nutritional supplements, resulting in improved quality ofcare and disease prognosis.
Keywords: Borrelia burgdorferi – in vivo-induced antigen technology – Lyme disease –multi-peptide ELISA
Introduction
Bacterial infection with specific spirochetes from thegenus Borrelia, usually acquired from the bite of aninfected Ixodes tick, is the cause of Lyme disease (LD).
The species complex B. burgdorferi sensu stricto ispredominantly responsible for LD cases in the United
States while B. burgdorferi sensu lato, B. afzelii andB. garinii are the responsible pathogenic species in
Europe and Asia (1,2).Presentation and destructive effects of LD vary widely
depending on the stage and magnitude of spirochetedissemination. Initial effects may include rashes or
flu-like symptoms, while later stages may be characterizedby arthritic, neurological, psychiatric and cardiac
symptoms.
Due to the varying nature of symptom appearancethere exist a number of DNA technologies and sero-logical methods that may be utilized, in conjunction withclinical examination, to support and confirm a diagnosisof LD; however, the techniques maintain certainpitfalls. Divergent symptoms between infected individualsand flawed clinical diagnostics hinder accurate diagnosisand prompt antibiotic treatment, which are essentialfor a good prognosis and the prevention of chronicinfection (3).The conventional treatment for LD is a course of
antibiotic therapy, such as doxycycline, amoxicillin,penicillin, erythromycin and other antibiotics listedin Table 1. Maximal efficacy of oral antibiotic adminis-tration is rendered in the early stages of infection,typically days or weeks following pathogen introduction.At the chronic stage of the disease, administration ofa combination of these medications orally, intramuscu-larly and intravenously is required for several monthsbefore an attenuation of infection is apparent (4–8).
For reprints and all correspondence: Aristo Vojdani, ImmunosciencesLab., Inc., 8693 Wilshire Blvd., Suite 200, Beverly Hills, CA 90211,USA. Tel: +1-310-657-1077; Fax: +1-310-657-1053;E-mail: [email protected]
� 2007 The Author(s).This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/2.0/uk/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work isproperly cited.
eCAM Advance Access published October 15, 2007

The swiftness in diagnosis is further emphasized by the
characteristic abilities of Borrelia, via antigenic changes
on its surface, to avoid the immune system and
subsequently infiltrate areas of the body maintaining
low levels of drug distribution, such as the joints and
nervous system. Due to this complexity, Complementary
and Alternative Medicine (CAM) holds great promise for
the abrogation of disease pathology through the combi-
nation and utilization of accurate diagnostic methods and
effective treatment modalities (9–30).The CAM treatment for LD, summarized in Table 1,
combines intravenous antibiotic therapy, herbal medicine
administration and nutritional medicine to enhance the
immune system’s ability to fight this intracellular micro-
organism (9–30).Considering the side effects of antibiotics and the
length of this treatment with CAM modalities, an
effective procedure for diagnosis of LD is needed to
justify such a course. A novel diagnostic approach to LD,
utilizing in vivo induced antigen technology (IVIAT) and
enzyme-linked immunosorbent assay (ELISA), may
provide the data necessary to justify CAM treatment of
LD and associated spirochetes. Therefore, it is imperative
to review the merit of this diagnostic approach, which
will expedite the accurate diagnosis of LD, in order to
emphasize its importance to practitioners of CAM in
areas of the world greatly affected by LD (31–34).
Prevalent Diagnostic Techniques:The Benefits and Inherent Drawbacks
A number of diagnostic tests for LD aim to detect thepresence of Borrelia antigens and related immune systemcomponents (35–39). While the theory behind thedevelopment of these techniques was sound, these testssuffer from false results (40); recent research has servedto give an understanding as to the reasons underlyingsuch inaccuracies.Polymerase chain reaction (PCR) for LD attempts to
detect the DNA of the spirochete. In a recent study,urine-based PCR for LD was evaluated and shown to beinsufficient for a diagnosis (35).The Center for Disease Control (CDC) proposes a two-
tiered approach to LD diagnosis (36). This includes aninitial screening test such as ELISA with high sensitivity,which is followed by an immunoblot with high specificity.
ELISA and Initial Screening Test for LD
The Lyme ELISA test is intended for the quantitativedetection of IgG and IgM antibodies to B. burgdorferi inhuman serum. Titers of IgG are generally low during thefirst weeks of illness. They peak within three to six weeksafter onset but are often not detectable in symptomaticpatients (37). This assay is performed based on crude
Table 1. CAM treatment of Lyme disease with a combination of different antibiotics, herbal remedies, and nutritional supplements, with pertinentreferences
Antibiotics used for treatment ofLyme disease (Refs 6–8, 10, 11)
Herbal remedies Ref Nutritional supplements Ref
Doxycycline Samento (24) Omega-3 Fatty Acids (26)
Minocycline Cumada (24) Coenzyme-Q10 (26)
Amoxicillin Burbur (24) S-Adenosylmethionine (8, 24, 25)
Augmentin Allicin (25) Grapeseed Extract (26)
Azithromycin Dragon’s blood (25) Magnesium (24)
Biaxin Cat’s claw (25) L-Carnitine (4, 8)
Erythromycin Devil’s claw (24) a-Lipoic acid (14, 15)
Vaucomycin Echinacea (25–28) Dimethylglycine (27)
Clarithromycin (biaxin) Citriodiol(lemon eucalyptus extract)
(9) Vitamin B-complex (4, 8)
Cefuroxime axetil Astragallus (29) Methylcobalamin (4, 8)
Penicillin Nettle (24) Vitamin C and D (12, 13)
Ceftriaxone Ginkgo biloba (4, 8) Hydrolytic enzymes (16, 17)
Rifampin Curcumin (19–20) Mushroom extracts and Beta-glucan (22)
Flagyl Oregano tea (25) Thymic peptides (29)
Plaquenil Boswellia (26) Pycnogenol (27)
Taurox SBTM Parsley extract (24) N-Acetylcysteine and glutathione (26)
Colloidal silver Red chili pepper (capsaicin) (18) Probiotics (23)
Imipinem Quercetin (21) Dehydroepiandrosterone (12)
Royal Jelly (26)
2 of 13 Novel diagnosis of Lyme disease

preparation of B. burgdorferi in culture and its binding tosolid phase matrix. IgG and IgM antibodies in patientsserum binds specifically to antigen coated wells resultingin color development. The intensity of color is equivalentto the level of antibodies.
Western Immunoblot Assay for Confirmation of LD
The western blot assay has been widely used to detect thepresence of antibodies in human serum and plasma tovarious infectious disease agents. In this procedure,component proteins of purified, inactivated bacteriaare electrophoretically separated by SDS-polyacrylamideelectrophoresis followed by electrotransfer to nitrocellu-lose sheets (38). Each strip served as the solid-phaseantigen for an ELISA test. The western blot assay ismore reliable since the cross-reactive antibodies areexcluded and peptide specific antibodies are observed.However, if antibodies are not present in the blood, falsenegative results will be obtained (39).
Drawbacks of Serodiagnosis of LD
The current problems with serodiagnosis of LD include alack of standardization and poor evaluation criteriaregarding sensitivity and specificity. These troubles aredue in part to the broad heterogeneity of the immuno-dominant antigens from disease causing Borrelia strainsand the utilization of a single strain in the whole-celllysate immunoblot assay (36). It is thought that theusage of several strains of different species would increaseassay sensitivity; while the usage of multiple Borreliaspecies grown in culture does provide a broader set ofantigens, it is not sufficient due to antigenic variationsand expression employed by Borrelia to avoid immunedestruction in the mammalian system (40–43). Therefore,it is crucial to combine clinical symptomology withsensitive techniques such as IVIAT Multi-Peptide ELISAthat take into account antigenic variation.
In Vivo Included Antigen Technology:The Key to Enhanced Specificity
IVIAT is a technique that identifies pathogen antigensthat are immunogenic and expressed in vivo duringhuman infection. Regulating gene expression optimizesa pathogen’s ability to proliferate within specific envir-onments, inducing or repressing specific genes to enhancegrowth within the host during infection, thus contributingto overall pathogenicity (41–43).IVIAT identification of these Borrelial gene products,
especially antigens from variable regions, during infectionmay be utilized to develop enhanced diagnostic methods(43). The chronic nature of LD and the antigenicdiversity of the spirochetes suggest the spirochete
undergoes dramatic changes in antigenic composition asit cycles between its arthropod and mammalian hosts andthat this variation plays an important role in immunedetection and destruction. Major proteins or lipoproteinscalled variable major protein (VMP) or VMP-likesequence (VlsE) are responsible for this antigenic varia-tion strategy evolved to avoid immune detection anddestruction (44, 45).Furthermore, different membrane surface protein E
(OSPE) enables the spirochete to evade the immunesystem and to maintain chronic infection (46–48). In thismulti-peptide ELISA assay based on IVIAT (32–34), weselected peptides from different components of Borreliaduring specific life cycles, including:
Outer Surface Proteins and Leukocyte
Function Associated Antigen
A prominent late manifestation of B. burgdorferi infec-tion is Lyme arthritis. Development of antibody reactiv-ity to outer surface protein A (OspA) occurs towardsthe beginning of prolonged arthritic episodes (49).The progression to an autoimmune state begins with across-reactive response between OspA and a self-antigen,e.g. leukocyte function associated antigen (LFA).Identification of a bacterial antigen that displays cross-reactivity with an autoantigen may provide a model fordevelopment of autoimmune disease induced byB. Burgdorferi and provide an important antigen toincorporate into diagnostic methods (50). Outer surfaceprotein C (OspC) is thought to be an important virulencefactor involved in the transmission and establishment ofearly infection (51, 52). Borrelia burgdorferi reduces theexpression of OspC in response to the development ofantibodies, indicating the importance of repressing OspCexpression to avoid clearance and to maintain infection(53–55). Since humans produce highly specific antibodiesagainst OspC, measurement of IgG and IgM antibodiesagainst this highly conserved epitope indicate recentinfection with B. burgdorferi. The Outer surface proteinE (OspE) is expressed in both ticks and mammals, andelicits a strong antibody response. Specificity of antibodyresponse with different OspE variants suggests that thehypervariable regions of OspE are targeted by theantibody response during infection. Consequently, it isimportant to assess the specificity of the antibodyresponse to OspE epitopes during infection and incorpo-rate this information into a testing method to targetantibodies to OspE (56).
Immunodominant Antigens
Two peptides, termed C2 and C6, are conserved acrossB. burgdorferi sensu lato, B. burgdorferi sensu stricto andB. garinii (44–47). Antigenicity of these peptides issupported by the observation that sera from humans,
eCAM 2007 3 of 13

monkeys and mice react with C2 and C6 peptide earlyand persistently through the course of infection, indicat-ing that these peptides contain one or more epitopes thatare broadly antigenic (57).In a study of human serum samples collected in the
northeast and mid-west of the US, from patients withacute or chronic LD, 35 of the 41 samples reacted withand presented significant levels of antibodies against theC6 peptide. The five serum samples that had nodetectable levels of antibodies against the C6 peptidewere obtained from patients who were in the early stageof infection (58).
Variable Major Protein
The spirochetes that cause LD express the surfacelipoprotein VMP or VlsE (45). Infection-inducedsequence changes that alter antigenic properties of VlsEcontribute to immune evasion. In an experimental modela strong VlsE IgG antibody response was apparent bythe fourth week of infection, indicating that VlsE isexpressed during early stage of infection. Serum samplesfrom LD patients also possessed VlsE antibodies thatwere immunoreactive against all species of Borellia (45).This indicates that there are conserved epitopes amongVlsE variants that are antigenic during infections. Basedon these experiments, researchers employed recombinantVlsE in an ELISA test and demonstrated diagnosticsensitivities of 63% for culture-confirmed LD erythemamigrans (EM) cases and 92% for late stage LD infections.
Peptides from Complement Regulator Acquiring
Surface Protein-1 (CRASP-1)
It is well known that the three different genospecies ofB. Burgdorferi exhibit distinct disease manifestations andtissue tropisms during mammalian infection consideringthe ability of pathogenic Borrelia sp. to survive intracel-lular in the mammalian host, these spirochetes must havedeveloped efficient strategies for circumventing the innateimmune response (59–61). Along these lines, it has beensuggested that CRASPs are key mediators that allowB. burgdorferi to evade the innate immune response andcomplement-mediated destruction during the acute stageof infection. Thus CRASP-1 expression is necessary forB. burgdorferi to resist killing by human serum (59, 61).
Peptides from Decorin Binding Protein for Species-Specific
Serodiagnosis of Lyme Arthritis and Neuroborreliosis
Among individual Borrelial proteins from differentspecies, sequence heterogeneity varies up to 40%(62, 64). Decorin binding protein A, a Borrelial outersurface protein, is one of the key proteins. This antigenelicits a strong antibody response during experimentalmurine borreliosis and has been suggested as a potential
vaccine protein (65). Decorin binding protein-peptidefrom B sensu stricto, B. garinii and B. afzelii wereproduced and tested as antigens by ELISA.One hundred percent of patients with neuroborreliosis
(NB) and 93% of patients with Lyme arthritis (LA)reacted positively. Sera from the majority of patients re-acted with one rDbpA only and had no or low cross-reactivity to other variant proteins. In patients withculture-positive EM, the sensitivity of rDbpA immuno-globulin G (IgG) or IgM ELISA was low. The DbpAseems to be a sensitive and specific antigen for theserodiagnosis of LA or NB, but not of EM (65).
Arthritis-Related Protein or Glutathione-S-Transferase
In addition, a 37-kDa arthritis-related protein ofB. burgdorferi outer-surface protein, termed arthritis-related protein (Arp), has been shown to prevent orreduce the severity of arthritis. Antibody responses to thissingle protein resembling glutathione-S-Transference weretested on 124 antibiotic-treated patients with early or latemanifestations of LD (66). Among the 124 antibiotic-treated patients, 53% with culture-proven EM had IgGresponses to recombinant glutathione S-transferase(GST)-Arp, as did 59% of the patients with facial palsyand 68% of those with LA. Therefore, measurement ofantibody against recombinant GST may be additionaltool for detection of B. burgdorferi involvement inpatients with accompanying arthritis during initialepisodes of joint pain or during the maximal period ofarthritis (66).
Antigens from Cross-Reactive Spirochetes and Other
Bacteria
Three genera, Treponema, Borrelia and Leptospira con-tain pathogenic spirochetes that cause disease in humans.All spirochetal disease share remarkable similarities inclinical manifestations including spirochetemia, in theearly stages of the disease, dissemination of spirochetesto body organs, skin lesions, one or more stages ofthe disease are frequently interrupted by a period oflatency, and neurological and cardiac involvement (67).In addition to similarities of their clinical conditions,cross-reactivity in all antibody assays including ELISAand western blot has been observed with treponema andleptospira (67). This cross-reactivity in an antibody assaycould be related to Borrelia hermsii, which is a cause ofrelapsing fever (68), Babesia microti, which causesBabesiosis (69), Ehrlichia or the agent of humangranulocytic ehrlichiosis (Anaplasma phagocytophilia)(69, 70) and Bartonella henselae, which can cause Cat-Scratch disease in a human (71, 72). In addition,serological cross-reactivity between B. burgdorferiand bacteria such as a Yersinia enterocolitica outermembrane protein was shown in a recent study (73).
4 of 13 Novel diagnosis of Lyme disease

Yersinia enterocolitica and Yersinia pseudotuberculosishave been identified as causative organisms of reactivearthritis in humans. Cross-reactivity between Yersinia andB. burgdorferi indicates that molecular mimicry betweenbacteria that does not belong to genus Borrelia with B.burgdorferi is possible (74). This includes other arthro-genic bacteria such Salmonella and Shigella, whichantibodies against them are detected in 67% of patientswith LD (75). Due to the extensive evidence of cross-reactivity between Treponema, Leptospira, Babesia,Ehrlichia, Bartonella and other bacteria that do notbelong to the genus Borrelia, particularly Yersinia with B.Burgdorferi, the diagnosis of LD should be based onclinical findings and complete serological analysis of B.burgdorferi antibodies and the exclusion of its cross-reactive microorganisms (74, 75). Furthermore, antibo-dies against these spirochetes may result in autoimmuneand neurological disorders, which are summarized below:
Autoimmunity Induced by Molecular Mimicry of Peptidesfrom B. burgdorferi and Human Tissue Antigens
Antigenic mimicry of infectious agents and auto-antigensis a proposed patho-mechanism for autoimmune diseases.For example, multiple auto-antigen mimotopes of infec-tion agents (Shigella, Rotavirus and Klebsiella), wereshown to induce autoimmune arthritis and uveitis in rats(73, 75), multiple microbial peptides (76) and Chlamydiapneumonia, specific peptides in multiple sclerosis (77, 78),Campylobacter jejuni in Guillain-Barre Syndrome (79)and Y. enterocolitica and autoimmune thyroid disease(80). In this regard spirochete B. burgdorferi could beanother trigger, which is an amino acid sequencehomology with self antigen that may trigger multi-organ system disorder in genetically predisposed indi-viduals (80). For example, using a basic local alignmentsearch (BLAST), four matched with Y. enterocolitica, andfive matched with B. burgdorferi and thyroid stimulatinghormone receptor (TSH-R), up to 50% identity andsimilarity were detected. Through this, it was concludedthat Borrelia and Yersinia as environmental triggers caninduce autoimmune thyroid disease in a geneticallypredisposed person through molecular mimicry mechan-isms (80). Another target antigen for autoimmunereaction downstream of the original bacterial infectionis cytokeratin-10 (81). These auto-antibodies in synoviallesions of chronic antibiotic treatment-resistant LA aredue to cross-reactivity between cytokratin 10 and aprominent outer surface protein A of B. burgdorferi (81).This cross-reactivity suggests a degree of antigenicmimicry that might contribute toward setting the stagefor autoimmunity, sustaining the response originallyelicited by B. burgdorferi long after elimination of thespirochete.This autoimmune reaction in LA may occur through
different mechanism of action and by utilization of
proteases from its mammalian hosts to degradeextra cellular matrix proteins. Binding of B. burgdorferi
to the host plasminogen and its activator Urokinase, isable to induce the production of the matrix metallo-proteinases (MMPs) and degradation of extra cellularmatrix proteins that allows B. burgdorferi to disseminatein the tissue (82).This up regulation of MMPs not only plays a role in
dissemination of the organism through extra cellularmatrix tissue, but it can also result in destructive pathology(83). Among the MMPs that have been suggestedto participate in cartilage degradation in patients withrheumatoid arthritis are collagenases (MMP-1, -8 and -13),
stromelysins (MMP-3, -10 and -11), and gelatinases(MMP-2 and -9) (84).Borrelia burgdorferi is dependent upon the mammalian
or tick host for the provision of amino acids and otherrequired components. An ABC oligopeptide transportersystem, (OPP) has been well characterized as anaminopeptidase. This and other proteases fulfill impor-
tant roles in both the basic physiology and pathogenesisof many microbial pathogens including the ability toprovide required amino acids. These proteases, byproviding required amino acids, enhance the growth of
spirochetes and their motility through mammalian hosttissues (82–84).
Neurological Manifestations of LD
Neurologic involvement in LD occurs in 10–40% ofpatients within several weeks to months of the tick bite.Both the peripheral and the central nervous system(CNS) can be involved, although disease in the formeroccurs more frequently. Early neuroborreliosis manifests
as lymphocytic meningitis (sometimes meningoencephali-tis), cranial neuropathies and peripheral radiculoneuro-phathies (85). Patients with meningitis have headachesand meningeal signs. Examination of the CSF shows alymphocytic pleocytosis with normal glucose. There is
often evidence of intrathecal antibody reactive withB. burgdorferi antigens, and oligoclonal bands may bepresent. Direct invasion of B. burgdorferi into thesubarachnoid space with a secondary immune responseis believed to be the cause of meningitis. As with other
manifestations of LD, meningitis can revolve sponta-neously despite persistent infection (85–87).Bell’s palsy, due to the involvement of cranial
nerve VII, is the most common cranial neuropathy,may be bilateral, and is often associated with otherperipheral nervous system manifestations. The most
common peripheral nervous system manifestation is amild, patchy, distal sensorimotor (axonal) neuropathy(86). Neurophysiology testing of individuals with radic-ular symptoms is abnormal, and in some cases,lymphocytic pleocytosis in the CSF is demonstrable
eCAM 2007 5 of 13

and associated with inrathecal anti-B. burgdorferi anti-body production (86).CNS involvement is relatively uncommon, usually late,
and occurs in either of two ways: an acute, disseminatedencephalomyelitis and a more indolent, late, slowlyprogressive parenchymal neurologic disorder resemblingmultiple sclerosis. Patients with the acute form presentwith depressed mental status, cognitive deficits andmultifocal abnormalities on neurologic examination.Magnetic resonance imaging (MRI) scans of the brainreveal predominantly white matter lesions, and the CSFexamination can exhibit both lymphocytic pleocytosisand intrathecal anti-B. burgdorferi antibody production.These patients respond well to antibiotic therapy withlittle, if any, residual deficit (85, 86). In contrast, patientswith slowly progressive inflammatory CNS neuroborre-liosis have often been misdiagnosed with multiplesclerosis and have received immunosuppressive treatmentto stabilize their neurologic deficits. These patients havewhite matter lesions on brain MRI scans, which had ledto the speculation that this form of neuroborreliosis is ademyelinating disorder (86).The presence of decorin binding protein B of
B. burgdorferi and antibody against it has been detectedin cerebrospinal fluid of patients with neurologic LD.This indicates that B. burgdorferi may enter the CNSafter infection. To infect the brain, the circulatingspirochetes must first cross the blood-brain barrier(BBB). This barrier is composed of tightly apposedbrain microvascular endothelial cells held together bythat tight junction (87). Several bacteria express theirown proteases that digest the extracellular matrix in orderto invade tissues, but other bacteria, like B. burgdorferi,appear to utilize the fibrinolytic system of the host todisseminate (87, 88). Borrelia burgdorferi and Borreliacrocidurae spirochetes use the fibrinolytic system itdisseminates into host tissues, including the brain. Atthe same time B. burgdorferi induces the expression andsecretion of the urokinase-type plasminogen activator(uPA) and expression of the uPA receptor (uPAR) by avariety of cell types. As the fibrinolytic system candirectly digest components of the extracellular matrix, itcan also activate other proteases, including MMPs. Notonly do these enzymes naturally occur in the bloodcirculation, but they are also expressed as a result of theinteraction between the host cells and B. burgdorferi (89).To prove the transversal of B. burgdorferi across thehuman BBB and systematic endothelial cell barriersin vitro model systems constructed of human brainmicrovascular endothelial cells (BMEC), a human umbili-cal vein endothelial cell (HUVEC) line where used (90).Results showed that B. burgdorferi differentially crosseshuman BMEC and HUVEC and that the human BMECfrom a barrier to traversal. Importantly, traversal of B.burgdorferi across human BMEC induces the expression ofplasminogen activators, activator receptors and MMPs.
This fibrinolytic system linked by an activation cascademay lead to focal and transient degradation of tightjunction proteins that allows B. burgdorferi to invadeCNS (90).In addition to the activation of the fibrinolytic system,
entry of spirochetes into the CNS may result in a severeinflammatory reaction, producing proinflammatory cyto-kines such as interferon-g and TNF-a. The cytokinereaction with neuronal cells may result in a neural cellantigen release and entry in circulation and induction ofboth cellular and humoral immune reaction againstmyelin ganglioside, sulfatides and many other self-antigens. Therefore, the complementary approach to thediagnosis and treatment of LD could not be based onlyon ELISA, western blot or Multi-Peptide ELISA assayfor detection of antibodies against Borrelia-specificantigens (32–34), but against other cross-reactive micro-organisms and different tissue antigens mentioned in thischapter. Measurements of IgG, IgM and IgA antibodiesagainst these important antigens that are related to LDare summarized in Table 2.
Mechanisms by which Tick Salivary Protein-15Facilitates Bacterial Invasion
The causative spirochete of LD not only employsmeans to change antigenic structures, but alsoborrows a practical coat from the saliva of its vector,the Ixodes tick (91, 92). The coating is present on themany active molecules introduced into the mammalianhost during attachment and feeding. The LD bacteriumreplicates and resides in the mid-gut of an infectedtick and moves transiently through the salivary glandsduring tick feeding. This transition into the salivaryglands is accompanied by a major phenotypic switch atthe bacterial surface, from expression of OspA toOspC (91, 92).Binding of OspC to salivary protein-15 (Salp15) in tick
saliva facilitates bacterial invasion of, and binding to,salivary glands. This suggests that one function of OspCis to serve as a ligand for a salivary gland receptor. Thefunction of Salp15 is to inhibit IgG antibody response toforeign antigens by halting CD4+ T-Cell activation, andinducing immune suppression, by binding to a receptoron the CD4+ T-Cell (91, 92). A direct connection occursbetween CD4+ and C-terminal amino acid residues ofSalp15, allowing for Salp15 to act at the very beginningof the signaling cascade initiated by T-Cell receptorbinding (92). This Salp15-mediated immunosuppressivemechanism, along with the activation of the fibrinolyticsystem and proinflammatory cytokine production, allowsfor facilitated invasion of the host’s immune and CNSs,as is shown in Fig. 1. This figure shows that after a tickbite, in addition to B. burgdorferi, tick salivary proteinsenter the host and exert an immunosuppressive effect (A).
6 of 13 Novel diagnosis of Lyme disease
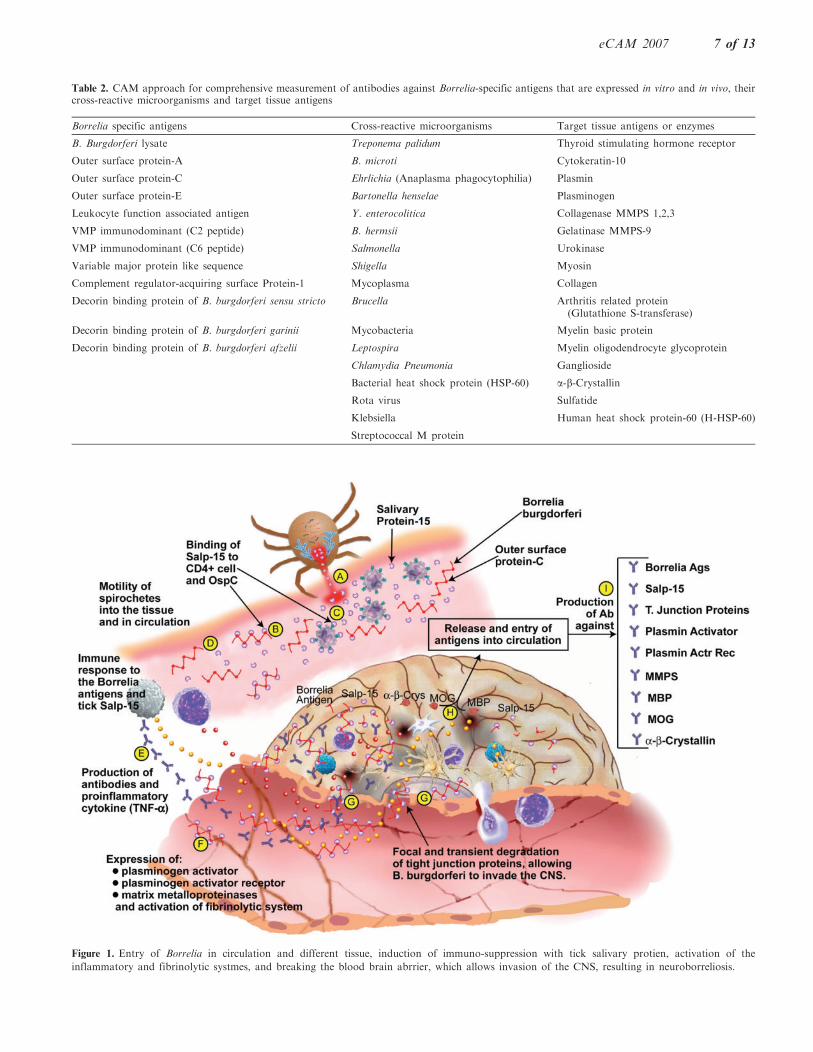
Figure 1. Entry of Borrelia in circulation and different tissue, induction of immuno-suppression with tick salivary protien, activation of the
inflammatory and fibrinolytic systmes, and breaking the blood brain abrrier, which allows invasion of the CNS, resulting in neuroborreliosis.
Table 2. CAM approach for comprehensive measurement of antibodies against Borrelia-specific antigens that are expressed in vitro and in vivo, theircross-reactive microorganisms and target tissue antigens
Borrelia specific antigens Cross-reactive microorganisms Target tissue antigens or enzymes
B. Burgdorferi lysate Treponema palidum Thyroid stimulating hormone receptor
Outer surface protein-A B. microti Cytokeratin-10
Outer surface protein-C Ehrlichia (Anaplasma phagocytophilia) Plasmin
Outer surface protein-E Bartonella henselae Plasminogen
Leukocyte function associated antigen Y. enterocolitica Collagenase MMPS 1,2,3
VMP immunodominant (C2 peptide) B. hermsii Gelatinase MMPS-9
VMP immunodominant (C6 peptide) Salmonella Urokinase
Variable major protein like sequence Shigella Myosin
Complement regulator-acquiring surface Protein-1 Mycoplasma Collagen
Decorin binding protein of B. burgdorferi sensu stricto Brucella Arthritis related protein(Glutathione S-transferase)
Decorin binding protein of B. burgdorferi garinii Mycobacteria Myelin basic protein
Decorin binding protein of B. burgdorferi afzelii Leptospira Myelin oligodendrocyte glycoprotein
Chlamydia Pneumonia Ganglioside
Bacterial heat shock protein (HSP-60) a-b-Crystallin
Rota virus Sulfatide
Klebsiella Human heat shock protein-60 (H-HSP-60)
Streptococcal M protein
eCAM 2007 7 of 13

This is done by tick Salp15 binding to OspC ofB. burgdorferi and protecting it from immune attack(B). Salp15 could also bind to CD4 on helper cells,inhibiting TCR ligation-induced-T-cell signaling andimmunosuppression (C). This way the spirochete canmove freely in the circulation and possibly in differenttissues (D). Eventually, immune response to Borrelia andSalp15 occurs, resulting in antibody as well as proin-flammatory cytokine production (E). Simultaneously, afew spirochetes may make contact with the endothelialcells of the BBB, stimulating expression of plasminogenactivators, plasminogen activator receptors and MMP, allof which contribute to activation of the fibrinolyticsystem (F). This in turn results in focal and transientdegradation of tight junction proteins, allowingB. burgdorferi to invade the CNS (G). This invasion ofthe CNS may result in the destruction of neuronal cells,the release of neural cell antigens and (H). the productionof antibodies against MBP, MOG, a-B-crystallin andother neural cell antigens (I). Another possibility for theproduction of these antibodies against neural cell anti-gens is through molecular mimicry between outer surfaceprotein A (OspA) and neural tissue (93). DNA sequencesof Bb OspA compared with a data bar of DNAsequences of human neural tissue yielded three sequencesthat were identical. The three corresponding Bb peptideswere synthesized, and antibodies were induced againstthem. The antibodies cross-reacted with human neuraltissues, including neurons in the human brain, spinal cordand dorsal root ganglia.These findings imply that antibodies developed by
LD patients against OspA will also bind to their ownneural tissue, representing a form of autoimmune diseasein which a person’s immune system attacks his owntissues (93).
CAM Diagnosis and Treatment for LD:Examples of Patients’ Results
LD is a complex and devastating disorder requiringefficient means of diagnosis in order to begin treatmentat the onset of infection. We have outlined the method oftransmission and the mechanisms of infection andmaintenance, both of which provide critical informationfor developing new diagnostic techniques. During thevarious life cycles of Borrelia in the body, the expressionof different antigens is tightly regulated such that thespirochete may disseminate and survive for long periodsof time. We have shown that the variety of antigens,identified through IVIAT (43), is integral to accuratelyidentify infection and indicate the stage of infection.Information gauged from clinical diagnostic methodsassessing these two components, most easily implementedinto an ELISA method, provides an infection profilefrom which an appropriate medical treatment may be
determined. In addition to IVIAT and its use in multi-peptide ELISA, work is being done to develop amethod to detect immune response to Ixodes proteins(32–34, 93, 94). Using this novel technology for thediagnosis of LD, examples of four patients with signs andsymptoms of LD are depicted in Fig. 2. Each specimenwas tested for IgG and IgM antibodies simultaneouslyagainst B. burgdorferi lysate and peptides from outersurface protein-A (OspA) and OspC, OspE, LFA,immunodominant peptides (C2+C6), VMP, B. b. sensustricto, B. garinii, B. afzelii, Babesia, Ehrlichia, Bartonella,Treponema palidum, as well as myelin basic protein(MBP), myelin oligodendrocyte glycoprotein (MOG) andcollagen as tissue antigens.Note that in Patient 1, despite presentation of clinical
symptomatology of LD, antibodies were not detectedagainst B. burgdorferi three tested subspecies, antigensand peptides, Babesia, Ehrlichia, Bartonella, Treponemaor tested target tissue antigens. Since symptoms ofhundreds of other diseases can overlap with those ofLD, the classification of disease should not be basedsolely on clinical findings but in conjunction with themost sensitive laboratory examinations available. Forinstance, Patient 2 was classified with Lyme-inducedarthritis, but in this case the diagnosis is confirmed by thepattern of detected antibodies, particularly againstB. burgdorferi lysate and leukocyte function associatedantigen, as well as very high levels of antibodies againstcollagen. In Patient 3, who was classified with neuro-borreliosis, the diagnosis was also confirmed by thepattern of detected antibodies against Borrelia antigensand neuroantigens (MBP and MOG).Finally, Patient 4, who was classified as having multi-
system organ disorder, reacted strongly with all16antigens tested in the assay. While patients of this typewith multi-reactive autoantibodies are called universalreactors, it is plausible that Ig-secreting plasma cells andmemory B cells activated by antigens have undergonesomatic hypermutation and/or class switch recombina-tion, thereby producing these pathogenic autoantibodies(95). Indeed, defects in B cell check points have beenshown to be associated with the development of multi-reactive autoantibodies in patients with systemic lupuserythematosus or rheumatoid arthritis (96). These foursample patients’ test results with four different patterns ofreactivity detected only by our novel comprehensiveapproach to laboratory examinations justifies CAMintervention, as summarized in Table 1 with pertinentreferences (6–29).
Conclusion
Researchers and practitioners of CAM understand thenecessity for an all-inclusive diagnostic approach andtreatment (as shown in Tables 1 and 2) and can bring
8 of 13 Novel diagnosis of Lyme disease
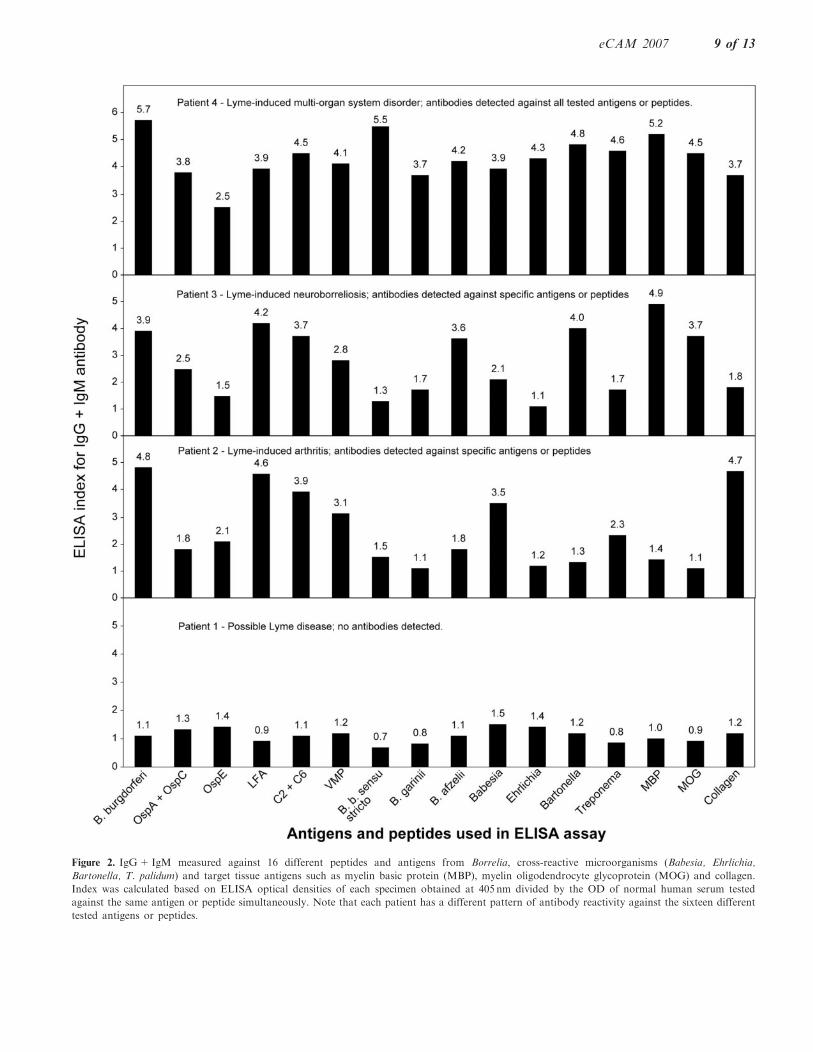
Figure 2. IgG+IgM measured against 16 different peptides and antigens from Borrelia, cross-reactive microorganisms (Babesia, Ehrlichia,
Bartonella, T. palidum) and target tissue antigens such as myelin basic protein (MBP), myelin oligodendrocyte glycoprotein (MOG) and collagen.
Index was calculated based on ELISA optical densities of each specimen obtained at 405 nm divided by the OD of normal human serum tested
against the same antigen or peptide simultaneously. Note that each patient has a different pattern of antibody reactivity against the sixteen different
tested antigens or peptides.
eCAM 2007 9 of 13
new perspectives and ideas to this developmental
endeavor. Antibodies against Borrelia-specific antigens
and their cross-reactive microorganisms and target
tissue antigens shown in Table 1 can be detected in
blood long before people show symptoms of variousautoimmune disorders. We conclude, therefore, that tests
that can detect these predictive autoantibodies could
warn of the need to take preventive action by CAM
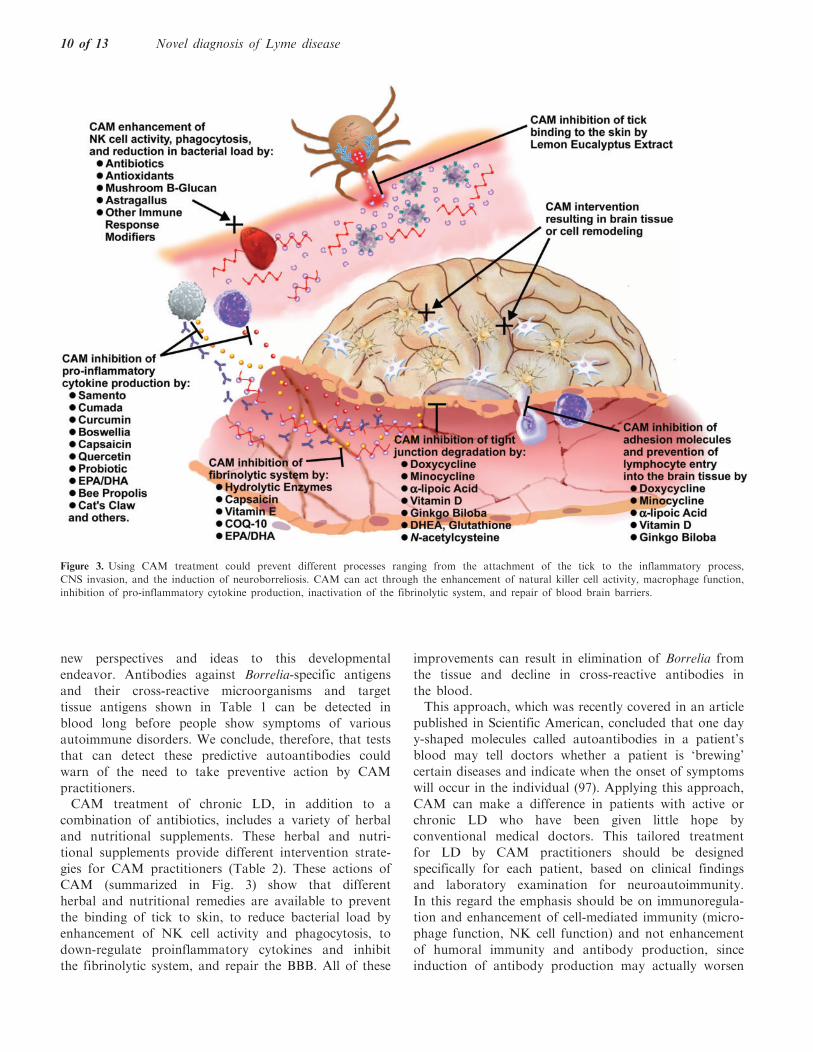
practitioners.CAM treatment of chronic LD, in addition to a
combination of antibiotics, includes a variety of herbal
and nutritional supplements. These herbal and nutri-
tional supplements provide different intervention strate-
gies for CAM practitioners (Table 2). These actions ofCAM (summarized in Fig. 3) show that different
herbal and nutritional remedies are available to prevent
the binding of tick to skin, to reduce bacterial load by
enhancement of NK cell activity and phagocytosis, to
down-regulate proinflammatory cytokines and inhibitthe fibrinolytic system, and repair the BBB. All of these
improvements can result in elimination of Borrelia from
the tissue and decline in cross-reactive antibodies in
the blood.This approach, which was recently covered in an article
published in Scientific American, concluded that one dayy-shaped molecules called autoantibodies in a patient’s
blood may tell doctors whether a patient is ‘brewing’
certain diseases and indicate when the onset of symptoms
will occur in the individual (97). Applying this approach,
CAM can make a difference in patients with active orchronic LD who have been given little hope by
conventional medical doctors. This tailored treatment
for LD by CAM practitioners should be designed
specifically for each patient, based on clinical findingsand laboratory examination for neuroautoimmunity.
In this regard the emphasis should be on immunoregula-
tion and enhancement of cell-mediated immunity (micro-
phage function, NK cell function) and not enhancement
of humoral immunity and antibody production, sinceinduction of antibody production may actually worsen
Figure 3. Using CAM treatment could prevent different processes ranging from the attachment of the tick to the inflammatory process,
CNS invasion, and the induction of neuroborreliosis. CAM can act through the enhancement of natural killer cell activity, macrophage function,
inhibition of pro-inflammatory cytokine production, inactivation of the fibrinolytic system, and repair of blood brain barriers.
10 of 13 Novel diagnosis of Lyme disease

the autoimmune reaction or autoimmune disease exhib-ited in many patients with LD.
Acknowledgments
We gratefully acknowledge the valuable insight andsuggestions that were contributed by Professor EdwinL. Cooper towards reviewing this manuscript, as well asthe wonderful illustrations that were created by Mr JoelBautista.
Conflict of Interest:
Aristo Vodjani is co-owner and CEO of ImmunosciencesLab, Inc. He declares no conflict of interest.
References1. Steere AC, Grodzicki RL, Kornblatt AN, Craft JE, Barbour AG,
Burgdorfer W, et al. The spirochetal etiology of Lyme disease.N. Engl. J. Med. 1983;308:733–40.
2. Steere AC. Lyme disease. N. Engl. J. Med. 1989;321:586–96.3. Kantor FS. Disarming Lyme disease. Scientific American
1994;271:34–9.4. Bock SJ. The integrative treatment of Lyme disease. Int. J. Integ.
Med. 1999;1:19–23.5. Howenstine J. New ideas about the cause, spread and therapy of
Lyme disease. Townsend Letter for Doctors 2004;252:64–7.6. Dattwyler RJ, Volkman DJ, Coanty SM, Platkin SP, Luft BJ.
Amoxicillin plus probenecid versus doxycycline for treatment oferythema migrans borreliosis. Lancet 1990;336:1404–6.
7. Luger SW, Paparone P, Wormser GP, Nadelman RB, Grunwaldt E,Gomez G, et al. Comparison of cefuroxime axetil anddoxycycline in treatment of patients with early Lyme diseaseassociated with erythema migrans. Antimi crob. Agents Chemother.1995;39:661–7.
8. Nicolson GL. Diagnosis and therapy of chronic systemic co-infec-tions in Lyme disease and other tick-borne infectious diseases.Townsend Letter 2007;93–8.
9. Gardulf A, Wolfhart I, Gustafson R. A prospective crossover fieldtrial shows protection of lemon eucalyptus extract against tick bites.J. Med. Entemol. 2004;41:1064–7.
10. Tikka TM, Koistinaho JE. Minocycline provides neuroprotectionagainst N-methyl-D-aspartate neurotoxicity by inhibiting microglia.J. Immunol. 2001;166:7527–33.
11. Hoyt JC, Ballering J, Numanami H, Hayden JM, Robbins RA.Doxycycline moderates nitric oxide production in murine lungepithelial cells. J. Immunol. 2006;176:562–72.
12. Du C, Khalil MW, Sriram S. Administration of dehydroepiandros-terone suppresses experimental allergic encephalomyelitis in SJL/Jmice. J. Immunol. 2001;167:7094–101.
13. Spach KM, Nashold FE, Dittel BN, Hayes CE. IL-10 signaling isessential for 1, 25-dihydrooxyvitamin D3-mediated inhibition ofexperimental autoimmune encephalomyelitis. J. Immunol.2006;177:6030–7.
14. Ha H, Lee JH, Kim HM, Kwak HB, Lee S, Kim HH, et al.a-Lipoic acid inhibits inflammatory bone resorption by suppressingprostaglandia E2 synthesis. J. Immunol. 2006;176:111–7.
15. Schreibert G, Musters RJP, Reijerkerk A, de Groot LR, van derPol Hendricx EML, Dopp ED, et al. Lipoic acid affects cellularmigration into the central nervous system and stabilizes blood-brainbarrier integrity. J. Immunol. 2006;177:2630–7.
16. Targoni OS, Tary-Lehmann M, Lehman PV. Prevention of murineEAE by oral hydrolytic enzymes. J. Autoimm. 1999;12:191–8.
17. Mynott TL, Ladhams A, Scarmato P, Engwerda CR. Bromelainfrom pineapple stems proteolytically blocks activation ofextracellular regulated kinase-2 in T cells. J. Immunol.1999;163:2568–75.
18. Beltran J, Ghosh AK, Basu S. Immunotherapy of tumors withneuroimmune ligand capsaicin. J. Immunol. 2007;177:3260–4.
19. Kim HY, Park EJ, Joe EH, Jou I. Curcumin suppresses Januskinase-stat inflammatory signaling through activation of Srchomology 2 domain-containing tyrosine phospatase 2 in brainmicroglia. J. Immunol. 2003;171:6072–9.
20. Gururajan M, Dasu T, Shahidain S, Jenning CD, Robertson DA,Rangnekar VM, et al. Spleen tyrosine kinase, a novel target ofcurcumin is required for B lymphoma growth. J. Immunol.2007;178:111–21.
21. Nair MP, Mahajan S, Reynolds JL, Aalinkeel R, Nair H,Schwartz SA, et al. The flavonoid Quercetin inhibits proinflamma-tory cytokine TNF-a gene expression in normal peripheral bloodmononuclear cells via modulation of NF-KB system. Clin. VaccineImmunol. 2006;13:319–28.
22. Lin H, Cheung SWY, Nesin M, Cassileth BR, CunninghamRundles S. Enhancement of umbilical cord blood cell hematopoiesisby mitake beta-glucan is mediated by granulocyte colony-stimulat-ing factor production. Clin. Vaccine Immunol. 2007;14:21–7.
23. Di Giacinto C, Marinaro M, Sanchez M, Strober W, Boirivant M.Probiotics ameliorate recurrent TH1-mediated murine colitis byinducing IL-10 and IL-10-dependent TGF-b-bearing regulatorycells. J. Immunol. 2005;174:3237–46.
24. Arthur S. The effectiveness of Samento, Cumada, Burbur and DrlLee Cowden’s protocol in the treatment of chronic lyme disease.Townsend Letter 2007;101–6.
25. Duke JA. Herbs with anti-Lyme potential. Townsend Letter2007;114–7.
26. Vojdani A, Erde J. Regulatory T cells, a potent immunoregulatorytarget for CAM researchers: The ultimate antagonist (I). eCAM2006;3:209–15.
27. Vojdani A, Erde J. Regulatory T cells, a potent immunoregulatorytarget for CAM researchers: Modulating allergic and infectiousdisease pathology (II). eCAM 2006;3:1–6.
28. Vojdani A, Erde J. Regulatory T cells, a potent immunoregul-atory target for CAM researchers: Modulating tumor immunity,autoimmunity and alloreactive immunity (III). eCAM2006;3:309–16.
29. Rosenbaum M, Vojdani A, Susser M, Watson CM. Improvedimmune activation markers in chronic fatigue and immunedysfunction syndrome (CFIDS) patients treated with thymic proteinA. J. Nutrit. Environ. Med. 2001;11:241–7.
30. Chu D, Wong WL, Mavligit GM. Immunotherapy with Chinesemedical herbs I. Immune restoration of local xenogenic graft-versus-host reaction in cancer patients by fractionated astragalus membra-naceous invitro. J. Clin. Lab. Immunol. 1988;25:119.
31. Schmidt BL. PCR in laboratory diagnosis of human Borreliaburgdorferi infections. Clin. Microbiol. Rev. 1997;10:185–201.
32. Vojdani A, Raxlen BT. Simultaneous measurement of antibodiesagainst Borrelia, Babesia and Ehrlichia by Western Blot and multi-peptide ELISA. FASEB J. 2005;19:95, Abstract.
33. Vojdani A. Part 1: In vivo-induced antigen technology: the mostsensitive method of detection for Lyme disease and other tick-bornediseases. Townsend Letter 2007;285:107–13.
34. Vojdani A. Part 2: The use of lymphocyte proliferation assay andcytokine production in seronegative patients with Lyme arthritis orneuroborreliosis. Townsend Letter 2007;286:104–11.
35. Rauter C, Mueller M, Diterich I, Zeller S, Hassler D, Meegans T,et al. Critical evaluation of urine-based PCR assay fordiagnosis of Lyme borreliosis. Clin. Diag. Lab. Immunol.2005;12:910–7.
36. Centers for Disease Control and Prevention. Recommendation fortest performance and interpretation from the Second NationalConference on Serologic Diagnosis of Lyme Disease. Morb. MortalWkly. Rep. 1995;44:590.
37. Luger SW, Krauss E. Serologic tests for Lyme disease: interlabora-tory variability. Arch. Intern. Med. 1990;150:761–816.
38. Dressler F, Whalen JA, Reinhardt BN, Steere AC. Western blottingin the serodiagnosis of Lyme disease. J. Infect. Dis.1993;167:392–400.
39. Engstrom SM, Shoop E, Johnson RC. Immunoblot interpretationcriteria for serodiagnosis of early Lyme disease. J. Clin. Microbiol.1995;33:419–27.
eCAM 2007 11 of 13

40. Saah AJ, Hoover DR. ‘‘Sensitivity’’ and ‘‘specificity’’ reconsidered:the meaning of these terms in analytical and diagnostic settings.Ann. Intern. Med. 1997;126:91–4.
41. Mahan MJ, Slauch JM, Mekalanos JJ. Selection of bacterialvirulence genes that are specifically induced in host tissues.Science 1993;259:686–8.
42. Mahan MJ, Heithoff DM, Sinsheimer RL, Low DA. Assessment ofbacterial pathogenesis by analysis of gene expression in the host.Annu. Rev. Genet. 2000;34:139–64.
43. Rollins SM, Peppercorn A, Hang L, Hillman JD, Calderwood SB,Handfield M, et al. In vivo induced antigen technology (IVIAT).Cellular Microbiol. 2005;7:1–9.
44. Sung SY, McDowell JV, Marconi RT. Evidence for the contribu-tion of point mutations to VlsE variation and for apparentconstraints on the net accumulation of sequence changes in VlsEduring infection with Lyme disease spirochetes. J. Bacteriol.2001;168:5855–61.
45. McDowell JV, Sung S, Hu LT. Evidence that the variable regions ofthe central domain of VlsE are antigenic during infection with Lymedisease spirochetes infection and immunity. Infect. Immun.2002;70:4196–203.
46. Magnarelli LA, Ijdo JW, Padula SJ, Flavell RA, Fikrig E. Serologicdiagnosis if Lyme borreliosis by using enzyme-linked immunosor-bent assays with recombinant antigens. J. Clin. Microbiol.2000;38:1735–9.
47. Alitalo A, Meri T, Lankinen H, Seppala I, Lahdenne P, Hefty PS,et al. Complement inhibitor factor H binding to Lyme diseasespirochetes is mediated by inducible expression of multiple plasmidencoded outer surface protein E paralogs. J. Irnmunol.2002;169:3847–53.
48. Metts MS, McDowell JV, Theisen M, Hansen PR, Marconi RT.Analysis of the OspE determinants involved in binding of Factor Hand OspE targeting antibodies elicited during Borrelia burgdorferiinfection in mice. Infect. Immun. 2003;71:3587–96.
49. Crowley C, Huber BT. Host-adapted Borrelia burgdorferi in miceexpresses OspA during inflammation. Infect. Immun.2005;71:4003–10.
50. Gross DM, Forsthubert T, Tary-Lehmann M, Etling C, Ito K,Nagy ZA, et al. Identification of LFA-1 candidate autoantigen intreatment resistant Lyme arthritis. Science 1998;281:703–5.
51. Earnhart CG, Buckles EL, Dumler JS, Marconi RT. Demonstrationof OspC type diversity in invasive human Lyme disease isolates andidentification of previously uncharacterized epitope that defines thespecificity of the OspC murine antibody response. Infect. Immun.2005;73:7869–77.
52. Earnhart CG, Marconi RT. OspC phylogenetic analyses support thefeasibility of a broadly protective polyvalent chimeric Lyme diseasevaccine. Clin. Vaccine Immunol. 2007;14:628–34.
53. Fingerle V, Goettner G, Gern L, Wilske B, Schulte-Spechtel U.Complementation of a Borrelia afzelii OspC mutant highlights thecrucial role of OspC for dissemination of Borrelia afzelii in Ixodesricinus. Int. J. Med. Microbiol. 2007;297:97–107.
54. Xu Q, McShan K, Liang FT. Identification of an OspC operatorcritical for immune evasion of Borrelia burgdorferi. Mol. Microbiol.2007;64:220–31.
55. Xu Q, Seemanapali SV, McShan K, Liang FT. Constitutiveespression of outer surface protein C diminishes the ability ofBorrelia burgdorferi to evade specific humoral immunity. Infect.Immun. 2006;74:5177–84.
56. Metts MS, McDowell JV, Theisen M, Hansen PR, Marconi RT.Analysis of the OspE determinants involved in binding of factor Hand OspE-targeting antibodies elicited during Borrelia burgdorferiinfection in mice. Infect. Immun. 2003;71:3587–96.
57. Crother TR, Champion CI, Whitelegg JP, Aguilera R, Wu XY,Blanco DR, et al. Temporal analysis of the antigenic composition ofBorrelia burgdorferi during infection in rabbit skin. Inf. Immun2004;72:5063–5072.
58. Philipp MT, Wormser GP, Marques AR, Bitker S, Martin DS,Nowakowski J, et al. A decline in C6 antibody titers occurs insuccessfully treated patients with culture-confirmed early localizedor early disseminated Lyme borreliosis. Clin. Diag. Lab. Immunol.2005;12:1069–74.
59. Wallich R, Pattathu J, Kitiratschky V, Brenner C, Zipfel PF,Brade V, et al. Identification and functional characterization of
complement regulator-acquiring surface protein-1 of the Lymedisease spirochete Borrelia afzelii and Borrelia garinii. Infect.Immun. 2005;73:2351–9.
60. Haupt K, Kraiczy P, Wallich R, Brade V, Skerka C, Zipfel PF.Binding of human factor H-related protein 1 to serum-resistantBorrelia burgdorferi is mediated by Borrelial complement regulator-acquiring surface proteins. J. Infect. Dis. 2007;196:59–61.
61. Kraiczy P, Rossman E, Brade V, Simon MM, Skerka C, Zipfel PF,et al. Bidning of human complement regulators FHL-1 and factor Hto CRASP-1 orthologs of Borrelia burgdorferi. Wien. Klin.Wochenschr. 2006;118:669–76.
62. Hanson MS, Cassat DR, Quo BP, Patel NK, McCarthy MP,Dorward DW, et al. Active and passive immunity against Borreliaburgdorferi decorin binding protein A (DbpA) protects againstinfection. Infect. Immun. 1998;6:2143–53.
63. Leuba-Garcia S, Martinez R, Gem L. Expression of outer surfaceproteins A and C of Borrelia afzelii in Ixodes ricinus ticks and in theskin of mice. Zentbl. Bakteriol. 1998;287:475–84.
64. Schwan TG, Piesman J. Temporal changes in outer surface proteinsA and C of the Lyme disease-associated spirochete, Borreliaburgdorferi, during the chain of infection in ticks and mice.J. Clin. Microbiol. 2000;38:382–8.
65. Heikkila T, Seppala I, Saxen H, Panelius J, Yrjanainen H,Lahdenne P. Species specific serodiagnosis of Lyme arthritis andNeuroborreliosis due to Borrelia burgdorferi stricto, B. afzelii,B. gorinii by using Decorin binding protein A. J. Clin. Microbiol.2002;40:453–60.
66. Salazar CA, Rothemich M, Drouin EE, Glickstein L, Steere AC.Human Lyme arthritis and the immunoglobulin G antibodyresponse to the 37-kilodalton arthritis-related protein of Borreliaburgdorferi. Infect. Immun. 2005;73:2951–7.
67. Schmid GP. Epidemiology and clinical similarities of humanspirochetal diseases. Rev. Infect. Dis. 1989;2:51460–9.
68. Guyard C, Chester ME, Raffel SJ, Schrumpf ME, Policastro PF,Porcella SF, et al. Relapsing fever spirochetes contain chromosomalgenes with unique direct tandemly repeated sequences. Infect.Immun. 2005;73:3035–7.
69. Homer MJ, Lodes MJ, Reynolds LD, Zhang Y, Douglass JF,McNeill PD, et al. Identification and characterization of putativesecreted antigens from Babesia microti. J. Clin. Microbiol.2003;41:723–9.
70. Brown WC, McGuire TC, Zhu D, Lewin HA, Sosnow J,Palmer GH. Highly conserved regions of the immunodominantmajor surface protein 2 of the genogroup II Ehrlichial pathogenAnaplasma marginale are rich in naturally derived CD4+ Tlymphocyte epitopes that elicit strong responses. J. Immunol.2001;166:1114–24.
71. Gilmore RD, Bellville TM, Sviat SL, France M. The Bartonellavinsonii subsp. arupensis immunodominant surface antigen BrpAgene, encoding a 382-kilodalton protein composed of repetitivesequence, is a member of a multigene family conserved amongBartonella species. Infect. Immun. 2005;73:3128–36.
72. Werner JA, Feng S, Kasten RW, Hodzec E, Chomel BB,Barthold SW. Cloning, characterization and expression ofBartonella henselaeP26. Clin. Vaccine. Immunol. 2006;13:830–6.
73. Rawlins ML, Gerstner C, Hill HR, Litwin CM. Evaluation of awestern blot method for detection of Yersinia antibodies: evidenceof serological cross-reactivity between Yersinia outer membraneproteins and Borrelia burgdorferi. Clin. Diag. Lab. Immunol.2005;12:1269–74.
74. Bruckbauer HR, Preac-Mursic V, Fuchs R, Wilske B. Cross-reactive proteins of Borrelia burgdorferi. Eur. J. Clin. Microbiol.Infect. Dis. 1992;11:224–32.
75. Sobieszczanska B, Prozondo-Mordanska A. Cross-reactivity betweenBorrelia burgdorferi and arthrogenic bacteria in sera from patientswith reactive arthritis. Rocz. Akad. Med. Bialmyst. 1996;41:90–95.
76. Grogan JL, Kramer A, Nogai A, Dong L, Ohde M, Schneider-Mergener J, et al. Cross-reactivity of myelin basic protein-specificT-cells with multiple microbial peptides: Experimental autoimmuneencephalomyelitis induction in TCR transgenic mice. J. Immunol.1999;163:3764–70.
77. Vojdani A, Campbell AW, Anyanwu E, Kashanian A, Bock K,Vojdani E. Antibodies to neuron-specific antigens in children withautism: possible cross-reaction with encephalitogenic proteins
12 of 13 Novel diagnosis of Lyme disease

from milk. Chlamydia pneumoniae and Streptococcus group A.J. Neuroimmunol. 2002;129:168–77.
78. Vojdani A, Vojdani E, Cooper EL. Antibodies to myelin basicprotein, myelin oligodendrocytes peptides, a-b-crystallin, lympho-cyte activation and cytokine production in patients with multiplesclerosis. J. Internal Med. 2003;254:363–74.
79. Greunewald R, Ropper AH, Lior H. Serologic evidence ofCampylobacter jejuni coli enteritis in patients with Guillain-Barresyndrome. Arch. Neurol. 1991;48:1080–2.
80. Benvenga S, Guarneri F, Vaccaro M, Santarpia L, Trimarchi F.Homologies between proteins of Borrelia burgdorferi and thyroidautoantigens. Thyroid 2004;14:964–6.
81. Ghosh S, Seward R, Costello CE, Stollar BD, Huber BT.Autoantibodies from synovial lesions in chronic, antibiotictreatment-resistant Lyme arthritis bind cytokeratin-10. J. Immunol.2006;177:2486–94.
82. Hu LT, Perides G, Noring R, Klempner MS. Binding of humanplasminogen to Borrelia burgdorferi. Infect. Immun 1995;63:3491–6.
83. Ahrens D, Koch AE, Pope RM, Stein.Picarella M, Niedbala MJ.Expression of matrix metalloproteinase 9 (96-kd gelatinase B) inhuman rheumatoid arthritis. Arthritis Rheum. 1996;39:1576–87.
84. Behera AK, Hilderbrand E, Scagliotti J, Steere AC, Hu LT.Induction of host matrix metalloproteinases by Borrelia burgdorferidiffers in human and murine Lyme arthritis. Infect. Immun.2005;73:126–34.
85. Halperin JJ, Volkman DJ, Wu P. Central nervous systemabnormalities in Lyme neuroborreliosis. Neurology 1991;41:1571.
86. Schmutzhard E, Pohl P, Stanek P. Borrelia burgdorferi antibodiesin patients with relapsing-remitting form and chronic progressiveform of multiple sclerosis. J. Neurol. Neurosur. Psychiatry1988;51:1215.
87. Garcia-Monco JC, Villar BF, Alen JC, Benach JL. Borreliaburgdorferi in the central nervous system: experimental and clinicalevidence for early invasion. J. Infect. Dis. 1990;161:1187–93.
88. Kim KS. Pathogenesis of bacterial meningitis from bacteraemia toneuronal injury. Nat. Rev. Neurosci. 2004;4:376–85.
89. Coleman JL, Benach JL. The urokinase receptor can be induced byBorrelia burgdorferi through receptor of the innate immune system.Infect. Immun. 2003;71:5556–64.
90. Grab DJ, Perides G, Dumler JS, Kim KJ, Park J, Kim YV, et al.Borrelia burgdorferi, host-derived proteases and the blood-brainbarrier. Infect. Immun. 2005;73:1014–22.
91. Alarcon-Chaidez F, Ryan R, Wikel S, Dardick K, Lawler C,Foppa IM, et al. Confirmation of tick bite by detection of antibodyto Ixodes calreticulin salivary protein. Clin. Vaccine Immunol.2006;13:1217–22.
92. Garg R, Juncadella IJ, Ramamoorthi N, Ananthnarayanan SK,Thomas V, Rincon M, et al. Cutting edge: CD4 is the receptor forthe tick saliva immunosuppressor, Salp15. J. Immunol.2006;177:6579–83.
93. Alaedini A, Latov N. Antibodies against OspA epitopes of Borreliaburgdorferi cross-react with neural tissue. J. Neuroimmunol.2005;159:192–5.
94. Riberiro JMC, Alarcon-chaidez F, Francischetti IMB, Mans B,Mather TN, Valenzuela JG, et al. An annotated catalog of salivarygland transcripts from Ixodes scapularis ticks. Insect Biochem. Mol.Biol. 2006;36:111–29.
95. Hsu HC, Wu Y, Yang P, Wu Q, Job G, Chen J, et al.Overexpression of activation-induced cytidine deaminase in B cellsis associated with production of highly pathogenic autoantibodies.J. Immunol. 2007;178:5357–65.
96. Yurasov S, Wardemann H, Hammersen J, Tsuiji M, Meffre E,Pascual V, et al. Defective B cell tolerance checkpoints in systemiclupus erythematosus. J. Exp. Med. 2005;201:703–11.
97. Notkins AL. New predictors of disease. Scientific American2007;296:72–9.
Received May 16, 2007; accepted August 15, 2007
eCAM 2007 13 of 13





























