Embed Size (px)
Citation preview

NOVEL INSIGHTS ON THE ROLE OF SELENOPROTEIN P IN SPERM VIABILITY
A THESIS SUBMITTED TO THE GRADUATE DIVISION OF THE UNIVERSITY OF HAWAI‘I AT MĀNOA IN PARTIAL FULFILLMENT OF THE
REQUIREMENTS FOR THE DEGREE OF
MASTER OF SCIENCE
IN
CELL AND MOLECULAR BIOLOGY
AUGUST 2012
By
Elizabeth Quynh-Mai Dao Nguyen-Wu
Thesis Committee:
Frederick P. Bellinger, Chairperson
Marla J. Berry
Martin Rayner
Keywords: Selenoprotein, genetic rescue, fertility, sperm, GPx4, Cre-LoxP

ii
ACKNOWLEDGMENTS
I am sincerely thankful to the many people who have been invaluable during my
journey towards this degree. This thesis was made possible by having a great advisor,
Dr. Frederick P. Bellinger, whose scientific prowess, encouragement; supervision and
support have fostered my immense growth as a student of science. Scientific pursuit is
much like many aspects of life, full of challenges and not without adversity. However,
overcoming these challenges makes the successes much sweeter. I am privileged to
have a mentor such as Rick, who is strongly committed to cultivating a richly educational
environment. I am grateful for Dr. Marla Berry’s continued insight, advice, and faith in my
abilities to succeed in my pursuit of a career in science. I am honored to thank Dr. Martin
Rayner for his guidance and persistent belief in my potential during my progression from
a MS and now a PhD student. Dr. Rayner was one of the first professors I met when I
arrived in Hawaii six years ago. His scientific enthusiasm and neuroscience course
helped expand my knowledge of neuroscience. Every member of my committee has
been incredibly supportive of my endeavors throughout my graduate career.
I would like to thank the Bellinger lab for all their assistance, feedback, support and
great lab meetings. I must express my gratitude to Mrs. Ann Hashimoto for her technical
expertise and friendship. I appreciate Ms. Rachel Rueli’s hard work and readiness to
offer assistance whenever needed. I highly appreciate Dr. Marci Reeves for her
invaluable friendship and sharing her wisdom as a graduate student. The camaraderie,
moral support and advice from members of the Berry lab, Ms. Christy “Dr. G” Gilman, Dr.
Lucia Seale, Ms. Mindy McDermott and Mr. Jason Higa brighten everyday at JABSOM.
Thank you to Mr. Nash Witten and Ms. Naghum Alfulaij for getting me out of the lab to
enjoy the beauty of Hawaii on some of those hikes that nearly killed me. Thank you to all
my fellow graduate students, staff and faculty at JABSOM who have been helpful in
many ways and most importantly create an enjoyable and collaborative lab environment.
I would not still be in Hawaii if not for the loving Mrs. Lyn Hamamura who continues
to be absolutely invaluable for her expertise of the graduate program, helping me
navigate the sometimes rough seas of graduate school, and is always looking out for
best interests of the graduate students. I have been fortunate and am humbled by all of
those who supported me in any respect, especially my family for their constant
encouragement and unconditional love through the roller coaster of graduate school and
life.

iii
ABSTRACT
Selenium (Se) is a micronutrient essential for life in many organisms. Selenium is
incorporated into selenoproteins as the twenty-first amino acid, selenocysteine (Sec),
and has antioxidant properties. One member of the family of twenty-five selenoproteins
in humans is Selenoprotein P (Sepp1). This protein is synthesized primarily in liver and
is proposed to transport selenium throughout the body, particularly to the brain and
testes. Sepp1 knockout (KO) mice on (normal) diets without selenium supplementation
have decreased selenoprotein expression in brain and testes. Previous studies have
suggested that Sepp1 male KO mice are infertile due to kinks in the flagellum of
spermatozoa, greatly reducing sperm motility, therefore leading to dramatically
decreased fertility.
In this two-part study, our first objective was to further understand the role of
Sepp1 on sperm viability. We hypothesize that Sepp1 plays a critical role in sperm DNA
viability independent of motility, potentially through modulating glutathione peroxidase 4
(GPx4) biosynthesis. GPx4 is another selenoprotein known to protect cells from
membrane lipid peroxidation and has been implicated in development and fertility [71].
The second objective of this study was to introduce a novel application to rescue Sepp1
global expression in knockout animals (Sepp1r/r CMV+) using Cre recombinase
transgenic mice.
We addressed the role of Sepp1 in sperm DNA viability with intracytoplasmic
spermatozoa injections (ICSI) of Sepp1 KO sperm into wild type oocytes. Surrogate
female mice carrying embryos resulting from injection of Sepp1-/- sperm resulted in a
72.3% reduction in live pups born compared to Sepp1 heterozygous control sperm. Our
results from the ICSI experiments, in which sperm were directly injected into oocytes
without flagella, suggest that Sepp1 is critical for sperm DNA viability independent of
motility. We show through western blot analysis that GPx4 levels are significantly
decreased in the testes and epididymides of Sepp1 KO mice, whereas Sepp1r/r CMV+
rescue mice had restored expression levels comparable to the Sepp1 wild type (Sepp1
WT). Immunohistochemistry studies using an antibody against GPx4 further confirmed
that GPx4 levels were undetectable in fresh Sepp1 KO mouse sperm, while GPx4 levels
in Sepp1r/r CMV+ rescue mice were similar to those of wild type controls.

iv
Cre-Lox recombination is a commonly used genetic tool for site-specific gene
deletion. However, we demonstrate that this system can be used to rescue gene
expression as well, restoring the expression of Sepp1 in KO mice. We show that this
approach produced viable progeny of the systemic Sepp1r/r CMV+ (rescue) mice that
express the CMV-Cre driven Sepp1 gene in all tissues. We confirmed through the Morris
Water Maze (MWM) and other behavior assays that in contrast to Sepp1 KO mice,
Sepp1r/r CMV+ mice had normal neuromotor function and memory compared to
Sepp1+/+. Successful implementation of this method can further be utilized to restrict
gene expression to specific cells.
Our study presents new data showing that Sepp1 is crucial for viability of sperm
DNA, potentially through regulation of GPx4 levels. Furthermore, we demonstrate an
innovative method for restoration of gene expression using the Cre recombinase
transgenic system, which can be applied to restrict gene expression to specific cells.

v
TABLE OF CONTENTS
Acknowledgments..……………………………………………………………………………..ii
Abstract.……………………………………………………………………………………..…..iii
List of Tables…………………………………………………………………………………....vii
List of Figures..…………………………………………………………………………………viii
List of Abbreviations.....…………………………………………………………………………ix
Chapter
I. INTRODUCTION…….…...……………………………….…………………………………
Selenoproteins..…………………………...…….………………………..……1
Synthesis of Selenoproteins...……………….…………………………..……3
Selenoprotein P……………...…………………………………………………5
Selenoprotein P Knockout Mice….………………..……………...………….8
The Role of Selenoprotein P in Male Fertility…...……………...………….12
II. MATERIAL AND METHODS
Animals...……………………………………………………………...……….14
Generation of Sepp1-/- and Sepp1r/r Cre+...……………………...…………14
Animal Behavior...….………………………………………………...……….14
Sperm Morphology...……...………………………………………...………..15
Oocyte and sperm collection………………………………………………..16
Intracytoplasmic Spermatozoa Injections (ICSI)……………….………….16
Immunofluorescence Assay……..…..……………………………...……….17
Western Blot analysis.……..……..….……………………………...……….17
Statistical analysis..…..……..……..………………………………...……….18

vi
III. RESULTS
Selenoprotein P expressing Cre-recombinase
mediated genetic rescue........…………..……………….……...………19
Neuromotor behavior and gait impairments
are recovered in Sepp1r/r CMV+ mice...……………….……....………19
Restoration of spatial learning and memory………………………...……..20
Sepp1r/r CMV+ mice have normal sperm
morphology and motility compared to Sepp1-/- mice...……...………...21
Sepp1 is required for viable sperm DNA,
independent of sperm morphology…...……...……………...………….21
GPx4 protein expression is deficient in
Sepp1 KO testes and epididymides..…………………………………..22
GPx4 immunoreactivity levels are decreased
in Sepp1 KO epididymal sperm………………………………………...22
IV. DISCUSSION & CONCLUSION
Discussion and Conclusion.………………………………………...……….33
Concluding Remarks and Future Studies..…………………………………38
REFERENCES…………………………………………………………………………………39

vii
LIST OF TABLES
Chapter III
Table 1: Comparison of Sepp1 KO and Sepp1-/+ fertility.………………………………...30

viii
LIST OF FIGURES
Chapter I
Figure1: Selenoprotein P biosynthesis pathway.…………………………………………….7
Figure 2: Sepp1 KO rescue strategy...…………………………………………………........10
Chapter III
Figure 3: Generation of Sepp1r/r CMV+ mice………………………………………………..24
Figure 4: Sepp1r/r CMV+ mice have normal motor coordination.……….……………..…..25
Figure 5: Abormal gait exhibited by Sepp1 KO mice are restored in Sepp1 Rescue mice
………………………………………………………………………………………...26
Figure 6: Sepp1r/r CMV+ mice have restored spatial learning and memory...………..27-28
Figure 7: Sperm morphology and motility restored in Sepp1r/r CMV+……………………29
Figure 8: GPx4 protein expression in testes and epididymides are decreased in Sepp1
but restored in Sepp1r/r CMV+ ………………………………………….………..31
Figure 9: GPx4 immunoreactivity is significantly decreased in epididymal sperm of
Sepp1-/- mice………………………………………………………………………..32

ix
LIST OF ABBREVIATIONS
AD Alzheimer’s disease
AP-1 activator protein-1
ApoER2 apolipoprotein E Receptor 2 (aka LRPR8)
BSA bovine serum albumin
CD-1 general multipurpose mouse strain
CZB modified BMOC-2 (Brinster’s medium for ovum culture) medium named
after Chatot, Ziomek, and Bavister
DIOs deiodinases
Dio1 iodothyronine deiodinase 1
DAPI 4',6-diamidino-2-phenylindole
DTT dithiothreitol
ER endoplasmic reticulum
GPx glutathione peroxidase
GSH glutathione
H2O2 hydrogen peroxide
hCG human chorionic gonadotropin
ICSI, intracytoplasmic spermatozoa injection
NGS, normal goat serum
PFA paraformaldehyde
PVP polyvinylpyrrolidone
RBC red blood cells
ROS reactive oxygen species
SBP2 selenocysteine insertion sequence binding protein 2
Se selenium
Sec selenocysteine
SECIS selenocysteine insertion sequence

x
SecS selenocysteine tRNA Synthase
Sel I selenoprotein I
SelH selenoprotein H
SelK selenoprotein K
SelM selenoprotein M
SelN selenoprotein N
SelO selenoprotein O
SelR selenoprotein R
SelS selenoprotein S
SelT selenoprotein T
SelV selenoprotein V
SelW selenoprotein W
SeMet, selenomethionine;
Sepp1 selenoprotein P
SPS2 selenophosphate synthetase 2
TBS tris-buffered saline
tRNA[Ser]Sec selenocysteine tRNA
TRxR thioredoxin reductase
TRx thioredoxin
TYH Toyoda, Yokojama and Hoshi (ICSI media)
U Sec

1
CHAPTER I
INTRODUCTION
Selenium is a nutritional trace mineral essential for human health that is acquired
primarily through dietary intake. Selenium (Se) is primarily found in soil which provides
nutrients for crops and livestock. Although soil and food in the United States have
adequate selenium, soil in other parts of the world, such as parts of China and New
Zealand are selenium deficient. Selenium is incorporated into selenoproteins as the 21st
amino acid, selenocysteine. Many of the twenty-five known selenoproteins in humans, of
which 24 are also found in rodents [19], have antioxidant properties. Selenium deficiency
has been implicated in many diseases and conditions including neurodegeneration [88],
infertility [12,61], cardiovascular disease, immune disorders, and cancer [25,37,43] in
humans, and white muscle disease in livestock [64]. While Se is a vital micronutrient,
excessive Se intake can lead to toxicity, a condition known as selenosis. In certain Se
deficient areas of China, Keshan’s disease is a prevalent but preventable
cardiomyopathy that can be treated with Se supplementation [99,101]. Despite much
progress in describing members of this unique family of proteins, the properties and
mechanisms of many of the selenoproteins are still not clearly understood. Vertebrate
selenoproteins are highly conserved through evolution. Along with the fact that Se is
required for life and implicated in a host of diseases, further elucidating the diverse
functions and mechanisms of the family of selenoproteins may make critical
contributions to treating many diseases and conditions. Here, I provide an overview of
the current understanding of Se and members of the selenoprotein family, including the
two that are central to this study, selenoprotein P (Sepp1) and the antioxidant enzyme
glutathione peroxidase 4 (GPx4).
Selenoproteins
Selenium (Se) is a requisite micronutrient that is biochemically processed by
selenoproteins that incorporate Se through the 21st amino acid, selenocysteine (Sec)
[11]. Sec has a structure that is nearly identical with that of cysteine, with the exception
of a single atom, selenium, in place of sulfur, giving Sec a lower pKa (5.2 versus 8.3)
and higher reactivity [9]. The general selenocysteine biosynthesis pathway has been

2
conserved in archaea and eukaryotes that have selenoproteins [96]. However, not all
kingdoms and phyla within these domains have the capacity for selenoprotein synthesis
[47]. The bioavailable source of Se for most organisms occurs in the organic form of
selenomethionine which has been implied to be more effectively absorbed and in the
inorganic sodium selenate (i.e. supplements and animal food) [36]. The selenoprotein
family exhibits diverse tissue allocation, from globally to tissue-specific expression.
Subcellular localization also varies greatly, with some selenoproteins solely expressed in
certain organelles or as transmembrane proteins, while others are secreted to
extracellular spaces or plasma [9,74]. Several selenoproteins have been characterized
as antioxidant enzymes that alleviate damage caused by reactive oxygen species (ROS)
[4,22,80]. Selenoproteins may have potential roles as modulators of redox-regulated
signal transduction [4].
Three classes of selenoproteins, the GPxs (glutathione peroxidases), TRxRs
(thioredoxin reductases) and DIOs (deiodinases) were among the first eukaryotic
selenoproteins discovered and are the most extensively studied [9]. The GPxs, the
largest selenoprotein family in vertebrates [54], are integral to antioxidant glutathione
pathways, providing protection from reactive oxygen species (ROS). Five of the GPxs in
humans (four in mice) are selenoenzymes [3]. GPxs are hydroperoxidases that use
glutathione as a cofactor. The TRxRs use NADPH for reduction of TRx (thioredoxin) in
cellular redox pathways [89]. One member of the GPxs which will be further discussed in
this study is the enzyme GPx4. The three known isoforms of GPx4 are cytosolic,
mitochondrial (mGPx4), and nuclear (nGPx4). The last two have been implicated in
recent studies to be involved in spermatogenesis and male fertility. Glutathione
peroxidase reduces hydrogen peroxide and alkyl hydroperoxides at the expense of
glutathione. These selenoenzymes contribute to the antioxidant defense system in
mammalian cells [15,56].
Other selenoproteins include SelW which is highly expressed in skeletal muscle and
found to be involved in white muscle disease in livestock [92]. It is similar to the GPx
family in that it shares the redox motif and binds glutathione [7]. SelH (selenoprotein H)
is a nuclear-localized DNA-binding protein that may act as a transcription factor that is
involved in oxidative cellular stress response [9,63]. SelI (selenoprotein I) was found to
be the mammalian form of the phospholipid-synthesizing enzyme ethanolamine
phosphotransferase [9,40]. SelR (selenoprotein R)/SelX (selenoprotein X) is a member

3
of the methionine sulfoxide reductase family, important for reduction of sulfoxymethyl
groups [9]. SelN (selenoprotein N) is localized to the ER (endoplasmic reticulum)
membrane and may be necessary for proper muscle development [67]. SelS
(selenoprotein S) is also ER-localized and is important for removal of misfolded proteins
from the ER membrane [5]. The 15 kDa selenoprotein, Sep15, and SelK, SelM and SelT
(selenoproteins K, M and T respectively) are small ER proteins with largely unknown
functions [74]. SelM was recently reported to have neuroprotective functions [73]. SelO
and SelV (selenoproteins O and V, respectively) are two of the least known in this family.
SelO is widely distributed, whereas SelV expression is limited to testes [32].
Selenoprotein P (Sepp1 or SelP) is one of the more well studied selenoproteins, and has
been implicated in having a protective function in conditions including neurodegeneration
in Alzheimer’s disease [8,41,88]. Sepp1 is a secretory protein theorized to be the
delivery system that transports Se from liver to other tissues and hence enables
selenoprotein synthesis. Sepp1 has also been suggested to play a critical role during
development and normal male fertility [61]. Numerous studies have shown that selenium
plays an important role in reproductive health [12,20,30,31]. Selenium deficiency has
been reported to be involved in several reproductive and obstetric complications
including male and female infertility, miscarriage, preeclampsia, fetal growth restriction,
preterm labor, and gestational diabetes [57]. As this study focuses on reproductive
biology, we will further discuss our findings involving Sepp1 in male fertility and its
interaction with GPx4 later in this study.
Further investigation is important for elucidating the mechanisms of selenoproteins to
understand how altering levels of Se and different selenoproteins may affect biological
functions and human health.
Synthesis of Selenoproteins
Selenocysteine (Sec) biosynthesis is unique in that it takes place on its tRNA that
recognizes the UGA codon, typically a stop codon [72]. Thus, recoding of the UGA
codon, located in the coding region of selenoprotein mRNAs, from a stop codon to a
selenocysteine-insertion codon is required. During the unique translational sequence of
selenoproteins, cis- and trans-acting factors work synchronously to redirect translational
machinery to insert selenocysteine at UGA codons instead of terminating polypeptide
synthesis. These factors include an element in the 3′-UTR (untranslated region) of

4
eukaryotic selenoprotein mRNAs, termed the Sec insertion sequence (SECIS), which
was discovered following the cloning of another selenoprotein, iodothyronine deiodinase
1 (Dio1) [9,10]. All eukaryotic selenoproteins require a form of the SECIS element for
recoding UGA to the Sec codon [53]. The translational machinery within the cell typically
identifies the UGA codon as a termination signal, thereby releasing the nascent
polypeptide from the ribosome [33]. This biosynthesis of selenocysteine on tRNA[Ser]Sec is
catalyzed by selenocysteine synthase [51,95] and the tRNA[Ser]Sec-modifying enzyme
phosphoseryl-tRNA[Ser]Sec kinase [21]. The translation process also requires a SECIS-
RNA binding protein (SBP2) [26,44] which recruits a specialized elongation factor
[27,91], that delivers selenocysteyl-tRNA[Ser]Sec to the A-site of the ribosome. Another
Sec-tRNA[Ser]Sec binding protein, SecP43, is required for methylation of the 2’-hydroxyl-
ribosyl moiety in the wobble position of the selenocysteyl-tRNA[Ser]Sec. This may regulate
the shuttling of the selenocysteine synthase-selenocysteyl-tRNA[Ser]Sec complex between
the nucleus and cytoplasm [83,97].
Under low dietary selenium conditions, selenocysteine incorporation is inefficient,
resulting in some selenoprotein mRNAs being degraded via nonsense-mediated decay
[58]. Se remains high in testes and brain even in low Se conditions [34], in which Sepp1
may potentially be broken down to recycle Se or remain circulating in blood while
gradually being taken up in brain and testes. Nonsense-mediated decay is a pathway
that targets mRNAs containing premature termination codons for degradation [44]. The
presence of both a UGA codon and an RNA element downstream of the UGA were
shown to be necessary for selenium-dependent regulation of mRNA turnover [58].
Degradation of selenoprotein mRNAs under conditions of low Se is not uniform, with
some transcripts being more sensitive to nonsense-mediated decay than others [86,94].
Several factors may contribute to the sensitivity of selenoprotein mRNAs to nonsense-
mediated decay [84] at different steps of the translation process [21]. This highly
conserved and unique process that is energetically demanding on the cell suggests and
underscores the importance of selenoproteins.

5
Selenoprotein P
Selenoprotein P was first identified through biochemical studies in 1982 [60]. Sepp1
is an unusual selenoprotein, containing ten Sec residues in humans, 16–18 in
amphibians and fish, and 28 in sea urchins [9]. With the exception of selenoprotein P,
most selenoproteins typically have only one selenocysteine residue per polypeptide
chain. Sepp1 is primarily secreted from liver cells to deliver selenium to other tissues
and organs in the body (Fig. 1) [17] . Selenium cycling through Sepp1 in plasma occurs
at a high rate as indicated by the 3-4hr half life in plasma [34]. Roughly 25% of whole-
body selenium passes through rat plasma daily [17]. Plasma Sepp1 most likely supports
homeostatic expression of GPxs, TRxRs and other selenoenzymes [9] through its role in
supplying selenium to cells throughout the body [18]. In addition, albeit at the cost of
oxidizing TRx [9,87], Sepp1 also reduces peroxynitrite-induced protein oxidation and
nitration, as well as lipid and low-density lipoprotein (LDL) peroxidation, indicating a
potential role in oxidant defense [2]. Sepp1 is also synthesized in smaller amounts in the
brain. Sepp1 has been found to be highly expressed in the liver, testes and kidneys.
Under selenium deficient conditions, selenium is better retained in the testes and brain
relative to the other organs, which is likely due to Sepp1 uptake through its known
receptors, ApoER2 and Megalin. When selenium is limited, Sepp1 synthesis has priority
over glutathione peroxidase synthesis [17] and [39,100]
Sepp1 consists of two domains with the smaller C-terminal domain containing nine
selenocyteine residues, while the larger N-terminal domain has only one Sec residue in
a redox motif [34]. Sepp1 binds to at least two receptors of the lipoprotein receptor
family, ApoER2 and Megalin. ApoER2 facilitates the uptake of Sepp1 into the testis and
allows retention in the brain. Megalin facilitates uptake of filtered Sepp1 into the proximal
tubule cells of the kidney [34]. Evidence shows that mutant mice absent of the Sec-rich
C-terminus exhibit severe deficiencies in brain Se if not supplemented in the diet and
have greater susceptibility to infections and morbidity [18,72]. Previous work from
Bellinger et al [8] have also demonstrated that Sepp1 is associated with amyloid plaques
and neurofibrillary tangles in Alzheimer’s disease (AD) in a potentially neuroprotective
capacity and may have neuroprotective properties [88].
The role of Se in fertility has been increasingly studied. Sepp1 has important
implications in normal male reproductive function. Studies have shown that mRNA that
encodes Sepp1 in rats is selectively expressed in Leydig cells, the testosterone

6
producing cells in the testes [61]. It is unknown whether Sepp1 is produced by the
epididymis or whether it may be transported to the tubule lumen to act directly with
sperm. Thus the absence of Sepp1 may contribute to the defective sperm phenotype.
Furthermore, Sepp1 is also suggested to function in oxidant protection [18,59,61] and
has been shown to promote the survival of cells in culture [61,80]. Increased oxidative
stress may occur if epididymal selenoprotein synthesis is compromised [49,61].
Maintaining Sepp1 homeostasis is critical for overall Se balance. Researchers
investigating the role of Se in spermatogenesis have identified a remarkable increase in
Se specifically in late spermatids by X-ray fluorescence microscopy [45]. Se was
primarily observed in the midpiece of the sperm. The study suggests that the enrichment
was due to elevated levels of the mitochondrial form of GPx4 which was completely
reliant on the Se supplied by Sepp1 [45]. Impaired GPx4 biosynthesis, due to selenium
deficiency or to genetic defects in GPx4 or in Sepp1, has been implicated in causing
male infertility [30]. In 2001, the nuclear GPx4 isoform was identified to be exclusively in
the late spermatids and involved in DNA reorganization [68,81]. The mitochondrial
isoform of GPx4 has been suggested as one of the most relevant involved in
spermiogenesis [30]. However, evidence from a recent study demonstrated that absence
of nGPx4 leads to sperm nuclear matrix/chromatin instability that may negatively affect
the embryo development [71]. These and other studies implicate an important
relationship between Sepp1 and GPx4 in reproductive function and other dysfunctions in
human diseases, thus meriting further investigation.

7
Figure 1. Selenoprotein P biosynthesis pathway. Sepp1 is principally synthesized in
the liver, and secondarily in smaller amounts in the brain. Sepp1 is thought to transport
selenium to other tissues in the body. Sepp1 binds to the low-density lipoprotein receptor
ApoER2 in brain and testes and Megalin in kidneys.

8
Selenoprotein P Knockout Mice
Sepp1-/- mice were generated by introducing a construct containing a neor cassette
flanked by LoxP sites inserted into the second exon, 9 bases downstream from the
Sepp1 start codon by electroporation into 129S9/SvEvH-derived embryonic stem (ES)
cells [34]. These ES cells were subsequently injected into C57BL/6 blastocysts. The
resulting chimeric males were bred with C57BL/6J females. The stop codons present in
both LoxP sites prevent translation of SEPP1.
In brief, the LoxP sites are part of the Cre-LoxP site-specific recombination system.
LoxP sites consist of two 13 bp inverted repeats separated by an 8 bp asymmetric
spacer region. The Cre-LoxP system was discovered in the 1981 in which Cre is a
recombinase protein that originates from the P1 bacteriophage [85]. The fundamentals
of the system is based on one Cre molecule that binds per 13 bp inverted repeat or two
Cre molecules binding at one LoxP site. Recombination occurs in the asymmetric spacer
region in which directionality of the recombination site is dependent on this 8bp region.
Two arrangements of the LoxP sites can be designed. When two LoxP sites are in
opposite orientation to each other, inversion of the DNA flanked by the sites occurs.
Alternatively, if the two LoxP sites are oriented in the same direction as each other, this
dictates an excision of the flanked DNA, leaving only one remaining LoxP site. The
Sepp1-/- mice were constructed by the latter of the two described LoxP arrangements.
The site-specific excision of a particular piece of DNA can be used to eliminate or
inactive the endogenous gene or a transgene, or activate a transgene [76]. The Cre-
LoxP system is typically used to generate conditional knockout animals. In some cases,
researchers may choose to generate a conditional knockout due to an embryonic
lethality of a complete whole body knockout of a gene of interest. In other cases,
researchers may use this method to study the function of a gene when it is absent or
knocked out in a specific organ or cell population.
In this study, we will demonstrate the novel application of the Cre-LoxP system to
restore the Sepp1 gene in the Sepp1-/- mice using the LoxP site’s own start codon. As
the first objective of this study, based on sequence analysis of the Sepp1-/- construct, we
predicted that introduction of a Cre recombinase transgene to the Sepp1-/- mice would
excise the floxed neor containing construct, however preserving a start codon in the
LoxP site in frame with the Sepp1 gene. Further details regarding the genetic restoration

9
of Sepp1 gene, using this unique application of the Cre-Lox P system, will be discussed
as this is one of the two objectives in this study.
Sepp1 KO mice fed a selenium inadequate diet of 0.1 mg/kg, originally generated by
the Burk lab, exhibited an abundance of deficiencies compared to their wild type
counterparts including reduced weight, smaller body size, poor motor coordination
development, and strikingly reduced fertility [34]. Sepp1 KO mice have very low
selenium concentrations in the brain, the testis, and the fetus, with severe
pathophysiological consequences in each tissue [17]. A study by the Burk lab showed
genetic deletion of Sepp1 did not alter liver selenium levels apart from when Sepp1 KO
mice were on a less than adequate selenium supplemented diet, below 0.1 mg/kg, liver
Se levels were then increased [34]. Selenium levels in brains of mice on the same diet
were reduced by 19% and did not change with selenium supplementation [34]. Kidney
selenium levels were reduced to 76% of normal levels and did increase to normal levels
upon a selenium adequate diet of 0.25 mg/kg diet [34]. Deletion of Sepp1 causes
increased excretion of selenium in the urine and, as a result, decreases whole-body
selenium [16,18]. Sepp1-/- mice on a 0.10 mg/kg selenium diet developed spasticity and
abnormal movements, performing poorly on motor coordination tests such as the rotorod
and pole climb, whereas this diet served as sufficient selenium requirements for wild
type mice [35]. The results of the study implicate the absence of Sepp1 contributing
towards irreversible brain damage. ApoER2 is one of the known receptors in which
Sepp1 binds. In other studies, ApoER2-/- mice on selenium deficient diets, showed
similar neurological deficits to that of the Sepp1-/- [18]. The study suggests that
interruption of selenium supply evidenced by decreased selenium uptake to the brain
leading to neurological deficits were due to impairment to the Sepp1-ApoER2 pathway
[18].

10

11
Figure 2. Sepp1 KO rescue strategy. (A) Selenoprotein P knockout construct design
[34] (B) Sepp1 mice were screened by genotyping and PCR of genomic DNA with
primers spanning the Neor construct and exon 2 (C) Sequence comparison between
Sepp1 wild type and newly recombined Sepp1r/r CMV+: LoxP sequence, signal peptide
sequence of Sepp1 KO after recombination with CMV-Cre gene to produce Sepp1r/r
CMV+ mice.

12
The Role of Selenoprotein P in Male Fertility
Studies have demonstrated that the Sepp1 gene is essential for development of
functional spermatozoa. The testis exports selenium as sperm selenoproteins whereas
the brain is not known to export the element. Severe selenium deficiency has been
found to produce defective spermatozoa resulting in reduced fertility [61]. Sepp1 is
suggested to be an indispensible component of the selenium delivery pathway for
developing germ cells [61].
A previous study found that Sepp1 and GPx4 gene expression were down-regulated
in cadmium-induced testicular pathophysiology in rats [55]. Cadmium is a toxic heavy
metal that is associated with severe testosterone depression and sperm motility defects,
resulting in infertility. Infertile men with oligoasthenozoospermia exhibit decreased sperm
motility and sperm counts. In an early study of infertile men with this condition, it was
revealed that total mitochondrial GPx expression levels were significantly decreased in
sperm of these subjects [42].
Numerous studies in mice have shown that mature spermatozoa of Sepp1−/− males
have specific structural defects in sperm flagella that develop sequentially during
spermiogenesis and after testicular maturation in the epididymis [61]. The study found
development of a truncated mitochondrial sheath, an extrusion of a specific set of
axonemal microtubules and outer dense fibers from the principal piece, and an acute
bend present at the midpiece-principal piece junction [61]. The mitochondrial capsule,
containing the selenoprotein GPx4 as a structural element [52,61], was also implicated
in at least some of the sperm defects [61,69]. GPx4, as a member of the glutathione
peroxidase selenoprotein family, has well-characterized antioxidant activity. This study
determined that these sperm defects found in Sepp1−/− males were similar to those
observed in wild-type (Sepp1+/+) males fed a low selenium diet (<0.25 ppm) [19,61].
Sepp1 has also been reported to be present in vesicle-like structures in the basal region
of Sertoli cells [62], which have been referred to as “nurse" cells since they nourish the
developing sperm cells through the stages of spermatogenesis. Sertoli cells
phagocytose residual cytoplasm during spermatogenesis [62]. Studies in ApoER2-/- mice
suggests that Sertoli cells aid in the uptake of Sepp1 from the interstitial fluid by
hydrolyzing Sepp1 within the lysosomes potentially using a receptor-mediated pathway
[62].

13
ApoER2 has been suggested to mediate endocytosis by which Sepp1 supplies
selenium for spermatogenesis. Researchers have found evidence that Sepp1 functions
in conjunction with ApoER2 and absence of either may interrupt selenium supply, which
could also contribute to the production of defective spermatozoa and some abnormalities
in sperm function. This may be correlated to the signaling function of ApoER2
specifically in the testis, involving Sepp1 in various capacities including selenium
trafficking [62]. The complex interaction of Sepp1 with GPx4 and the ApoER2 receptor
in spermiogenesis and ultimately sperm fertility is an important issue that necessitates
greater understanding in developing effective therapies for male infertility.

14
CHAPTER Il
MATERIALS AND METHODS
Animals
Animals were provided food and water as needed per University of Hawaii veterinary
protocol. All animals in this study were maintained on diets containing adequate Se
(~0.25 ppm). Animals were on kept on a 12-h light cycle and group housed during
breeding and rearing. Prior to and during behavioral testing, animals were individually
housed in polycarbonate cages. Each cage was provided food, water, and a layer of
bed-o-cob (corn cob) bedding (Newco Distributers). All animal protocols were approved
by the University of Hawaii Institutional Animal Care and Use Committee.
Generation of Sepp1-/- and Sepp1r/r CMV+
Sepp1-/- mice were obtained from the laboratory of Dr. Raymond Burk at Vanderbilt
University. Mutant mice were backcrossed to C57BL/6J for at least 10 generations
before arriving in our lab and were bred with our C57BL/6J colony to ensure congenic
strains [38]. As male Sepp1 mice are infertile [34], Sepp1+/− mice were used for breeding
resulting in littermate Sepp1−/− and Sepp1+/+ pups. Sepp1 whole body genetic rescue
mice (Sepp1r/r CMV+) were generated by breeding Sepp1-/+ mice to C57BL/6N-
Hprttm1(CMV-cre)Brd/Mmucd (Jackson Labs). Confirmation of the expected recombination of
Sepp1 was carried by polymerase chain reaction (PCR) of extracted genomic DNA from
mouse tails using specific primers to amplify a 151-bp product in the targeted region
present in the wild type (forward-ACCTCAGCAATGTGGAGAAGCC, reverse-
TGCCCTCTGAGTTTAGCATTG), and a 472-bp and a 224-bp product specific for the
knockout allele and floxed gene, respectively (forward-
ACCTCAGCAATGTGGAGAAGCC, reverse-GATGATCTGGACGAAGAGCATCA).
Products were run on a 1.5% DNA agarose gel, and SYBR® Safe DNA Gel Stain
(InVitrogen) was detected by UV imaging to confirm genotype.
Animal Behavior
Adult Sepp1+/+, Sepp1r/r CMV+, or Sepp1-/- mice 12 to 20 weeks of age (age matched
for each assay) were evaluated for neuromotor and neurobehavior effects using typical
behavior paradigms that included Morris water maze [66], and open field test [70,93].

15
Morris water maze
Hippocampal-dependent spatial learning and memory was assessed using a Morris
Water Maze (MWM) assay [66]. Mice were placed in a large circular pool of opaque
24°C heated water (nontoxic, water based paint was added to achieve opacity). Visual
cues were placed on walls to provide mice with spatial orientation and reference points.
The time required to escape from water onto a hidden platform is measured. The total
time required for mice to swim to a visual platform (60 s maximum) was determined prior
to the training days. Each mouse was given 60 s to find the visible platform or after this
period, the mouse would be removed from water and placed on the platform for 15 s.
During the 8 days of training of 4 trials per day, the platform was submerged and the
total time (60 s maximum) for mice to find the hidden platform was recorded. On the
ninth day, the platform was removed for the probe trial. The total time spent in each
quadrant and the number of platform crossings was monitored over a 60 s period.
Pole Test
Locomotor function was evaluated using a pole test that examines rodent locomotor
function involving the cerebellum, motor cortex, and basal ganglia [29]. Mice were
placed on the top of a pole with heads oriented upward and parallel to the pole. After 2
days of training of 4 trials per day, the time taken to invert to facing downward and the
total time to descend were recorded in 4 trials on the third day. Mice were given 60 s to
perform this task.
Stride test
A stride test, modified from Fernagut et. al, 2002 [28], was used to measure
deficiencies in gait and motor ability. Paw prints were obtained by applying ink to the
hind limb paws of the mice before placing them on graph paper in a narrow runway.
Bright lighting was used to encourage the mice to walk toward a dark enclosure at the
opposite end of the runway. Length of stride for each paw and width of strides were
measured from the resulting footprints.
Sperm morphology
Sperm morphology studies were performed by isolating fresh epididymal sperm from
4-6 month old male Sepp1 WT, Sepp1 KO, and Sepp1r/r Cre+ mice. Mice were

16
euthanized by CO2 prior to harvesting epididymides. In a glass bottomed culture dish,
cauda epididymal sperm were extracted by applying pressure to the cauda epididymis in
Toyoda, Yokojama and Hoshi (TYH) media. Sperm were visualized using an Olympus
IX-71 microscope with DIC optics under 40X and 63X oil objectives and Pictureframe
imaging software.
Oocyte and sperm collection
Mature oocytes at metaphase II were obtained from B6D2F1 females at 2-3 months
old. Equine chorionic gonadotropin (CG) (5 IU) and human CG (hCG) (5 IU) were
administered 48 h apart. About 15-16 h after the injection of hCG, cumulus-oocyte
complexes were collected from the oviducts and treated with 0.1% hyaluronidase to
remove cumulus cells. Sperm were collected from Sepp1+/- and -/- adult males at 4-6
months old, euthanized by cervical dislocation. Cauda epididymal sperm were
suspended in HEPES-CZB medium (20mM HEPES-HCl, 5mM NaHCO3,0.1 mg/ml
polyvinyl alcohol) [98], for 20 min at 37°C before use [79].
Intracytoplasmic spermatozoa injections (ICSI)
ICSI was performed according to Kimura and Yanagimachi [48] with minor
modifications [98]. A drop of sperm suspension was mixed with 12% (w/v)
polyvinylpyrrolidone (PVP) in HEPES-CZB. The head of a single sperm was isolated
from the sperm flagella and injected into each oocyte with a piezo-driven
micromanipulator. We observed pronuclear formation under an inverted Olympus
microscope at 6 h after sperm injection. Fertilized oocytes with two pronuclei were
cultured for 24 h in CZB medium [23] at 37°C under 5% CO2 in air. The next day we
observed 2-cell stage embryo development.
Embryo transfer
Embryos at 2-cell stage were transferred into the oviduct of CD-1 surrogate mothers
(day 0.5) that had been mated with vasectomized CD-1 males the previous night.
Surrogate mothers were euthanized on Day 19.5 after embryo transfer to obtain
newborn pups.

17
Immunofluorescence assay
Sperm preparation for immunofluorescence was performed according to Paul et al
[65]. Sperm were harvested from Sepp1+/+, -/-, and r/r CMV+ 4-6 month old male mice
epididymides in ice cold 1X phosphate buffered saline (PBS). Epididymides were minced
and sperm strained in 70-µm nylon strainer (VWR International), red blood cells (RBCs)
were lysed with 0.45% saline, washed. Epididymis/sperm lysate were sonicated twice on
ice for 30 min in 20 mM phosphate buffer, pH 6 with 1mM EDTA. Samples centrifuged at
500 x g for 5 min at room temperature (RT) and resuspended in 50mM Tris-HCl, pH7.4
containing 1% SDS, incubated for 10 min at RT. After centrifugation (10,000 x g, 1 min)
supernatant was removed, pellet rewashed with 50mM Tris-HCl, pH 7.4. Sperm
resuspended in 25mM Tris-HCl with 0.25M NH4)SO4 and 40mM DTT, and incubated at
RT for 10 min. 30 µl aliquots were placed on glass slides (Fisher Scientific) on ice,
incubated for 40 min. washed in PBS (2 min), fixed in 4% paraformaldehyde (PFA) for
30min at RT. Slides were washed (PBS), air dried and stored at -20°C. Frozen slides
were thawed in PBS for 5 min the following day. Blocking was in 1:4 normal goat serum
(NGS) in 3% bovine serum albumin/tris-buffered saline (BSA/TBS) for 30 min. Samples
were incubated at 4°C with (1:500) rabbit GPx4 monoclonal primary antibody overnight
(AbFrontier). The next day, slides were placed in glass staining containers and gently
rinsed three times with PBS. Alexafluor 546 anti rabbit (InVitrogen) secondary antibody
was applied to samples and incubated for 45 min. at RT in a light protective container.
Sperm samples were washed three times with 1X PBS and mounted using Vectashield
Hardset Mounting Media containing the nuclear stain DAPI (Vector Laboratories).
Samples were viewed with the Zeiss Axioskop Plus 2 and images collected with the
AxioCam MR3 digital camera.
Western Blot analysis
GPx4 protein expression in testes and epididymides of age matched Sepp1 WT, KO,
and rescue mice were measured by western blot. Tissues were lysed with CelLytic MT
(Sigma Aldrich Co) according to the manufacturer’s instructions. Protein lysates were
resolved by SDS-PAGE separated on a 10–20% gradient Tris-HCl Criterion Precast gel
(Bio-Rad Laboratories) and transferred to polyvinylidene difluoride (PVDF). For detection
of GPx4 expression in testes, membranes were incubated in rabbit GPx4 polyclonal
antibody (AbFrontier) diluted 1:2000, and for epididymides, GPx4 polyclonal antibody

18
diluted 1:5000 (Epitomics) in 1:4 Odyssey blocking solution in PBS (LiCore Biosciences)
for 90 min at RT. Following washes in PBS (5 X 5 min), primary antibodies were
detected with LiCor near-infrared fluorescent secondary antibodies in 1:4 blocking
solution in PBS for 45 min followed by washes as described above. Subsequently blots
were developed using β-actin (Sigma Aldrich Co.) as a loading control. Protein was
detected using the Odyssey® Infrared Imaging System (Li-core Biosciences). Scanning
and analysis were performed with Licor Odyssey software.
Statistical analysis.
Statistical analysis was performed with GraphPad Prism software. Interaction
between genotypes and sex was ascertained by Student’s t-test and two-way analysis of
variance (ANOVA) with Bonferroni’s posthoc test for multiple comparisons. To determine
genotype differences between experimental groups in the water maze training and
quadrant entries in the probe trial, repeated-measures ANOVA with Bonferroni’s posthoc
test were used. Statistical significance was defined as having p<0.05 for all statistical
tests.

19
Chapter III
RESULTS
Selenoprotein P expressing Cre-recombinase mediated genetic rescue. Sepp1r/r
CMV+ rescue mice were generated by breeding Sepp1+/- mice to CMV-Cre expressing
mice, C57BL/6N-Hprttm1(CMV-cre)Brd/Mmucd purchased from Jackson Labs. Sepp1 KO
mice, originally generated by the Burk lab, were designed with a reverse-orientation neor
cassette flanked by LoxP sites, inserted into the second exon, 9 bases downstream of
the start codon (Fig 1). The neor containing construct effectively disrupts Sepp1
expression. The genetic deletion of Sepp1 results in a hosts of impairments including
male infertility [61]. Sepp1+/- littermates were used for generating the rescue mice due to
the infertility of Sepp1-/-. Breeding with the CMV-Cre resulted in recombinase expression
in all cells, and the resulting recombination excised the neor construct in the Sepp1-/-
mice. A start codon in the remaining single LoxP site was left in frame with the Sepp1
gene, thus resulting in translation of a form of Sepp1 with a minor mutation in the
Sepp1r/r CMV+ mice (Fig. 3B-C). Sepp1 genetic recombination in the Sepp1r/r CMV+
was confirmed with PCR genotyping, by identifying an amplified 224-bp product specific
for the recombined gene in comparison to the 151-bp product in the targeted region
present in the wild type and a 472-bp for the knockout allele (Fig. 3A). The ability of the
resulting Sepp1r/r CMV+ progeny to sire pups was indicative that the Sepp1 gene was
restored and functional.
Neuromotor behavior and gait impairments are recovered in Sepp1r/r CMV+ mice.
Sepp1 KO mice have severe motor impairments encompassing irregular gait patterns
described to include dragging of the limbs and uneven strides as well as motor
coordination deficits [35,78]. We used the pole and stride tests to determine whether the
Sepp1r/r CMV+ had complete genetic restoration. Motor coordination and general
locomotor function of Sepp1r/r CMV+ mice, assessed with the pole test, were
indistinguishable from those of the wild type group. The rescue mice in comparison to
the wild type group exhibited no significant differences in the total time to descend the
pole, whereas the Sepp1 KO mice had significantly decreased coordination and
locomotion as shown by the greater total amount of time Sepp1 KO mice took to

20
descend to the bottom of the pole (Fig. 4). One-way ANOVA showed an effect of
genotype on the time taken to descend (One-way ANOVA, **P=0.0086 for Sepp1r/r
CMV+, Bonferoni’s post hoc test **P<0.01). For the turn time, One-way ANOVA also
showed effect of genotype (**P=0.0024), with Bonferoni’s posthoc test (**P<0.01 for
Sepp1r/r CMV+ and *P<0.05 for Sepp1+/+).
We subjected the mice to a stride test to determine whether the ataxia seen in
Sepp1-/- mice was recovered in the Sepp1r/r CMV+. The rescue mice had restored gait
patterns similar to wild type mice. Stride length and width were measured for four sets of
paw prints of the mice hind paws. Both Sepp1r/r CMV+ and wild type mice had
significantly different stride length patterns compared to the ataxic irregular stride lengths
of the Sepp1-/- mice. Two-way ANOVA determined there was a significant effect of
genotype (**P= 0.0034) with Bonferroni’s Post hoc analysis (*P<0.05)
Restoration of spatial learning and memory. The Morris Water Maze (MWM) test was
administered to assess whether spatial learning and memory was restored in the
Sepp1r/r CMV+ mice. The visual platform test was given on the first day to introduce
mice to learning to escape the water by climbing onto the platform, prior to beginning the
training period. There were no significant differences between groups in the time (60 sec
maximum) they took to find the visible platform (2-way ANOVA). During the training
period, rate of learning was assessed by daily changes in escape latency times, which
are measured as the time taken to climb onto the hidden platform during each trial (60
sec maximum). Sepp1 rescue mice learned at the same rate as the wild type group, as
shown by lack of differences in escape times between the groups. 2-way repeated
measures ANOVA confirmed that there was no significant difference in escape latency
during the 8 training days between Sepp1r/r CMV+ and Sepp1+/+ mice. Escape latency
times of both wild type and rescue mice were markedly less than those of Sepp1-/- mice.
Sepp1-/- mice MWM data were obtained in a different experiment in which all
experimental procedures and parameters were the same, and kindly provided by Dr.
Matthew Pitts. Sepp1-/- mouse data were included only as an example for comparison of
the behavioral deficits previously reported in Sepp1-/- mice [66,72].
During the probe trial, the time spent in each quadrant was recorded to assess if
mice recalled where the platform was during the training period. The number of platform
crossings was monitored during a 60 second swim. We found no significant differences

21
between wild type and rescue genotypes for time in each quadrant, number of platform
crossings, swim speed or distance (two-way ANOVA, P>0.05). Swim speed and total
distance traveled were recorded to control for factors other than learning and memory
differences that could affect the results, such as motor coordination.
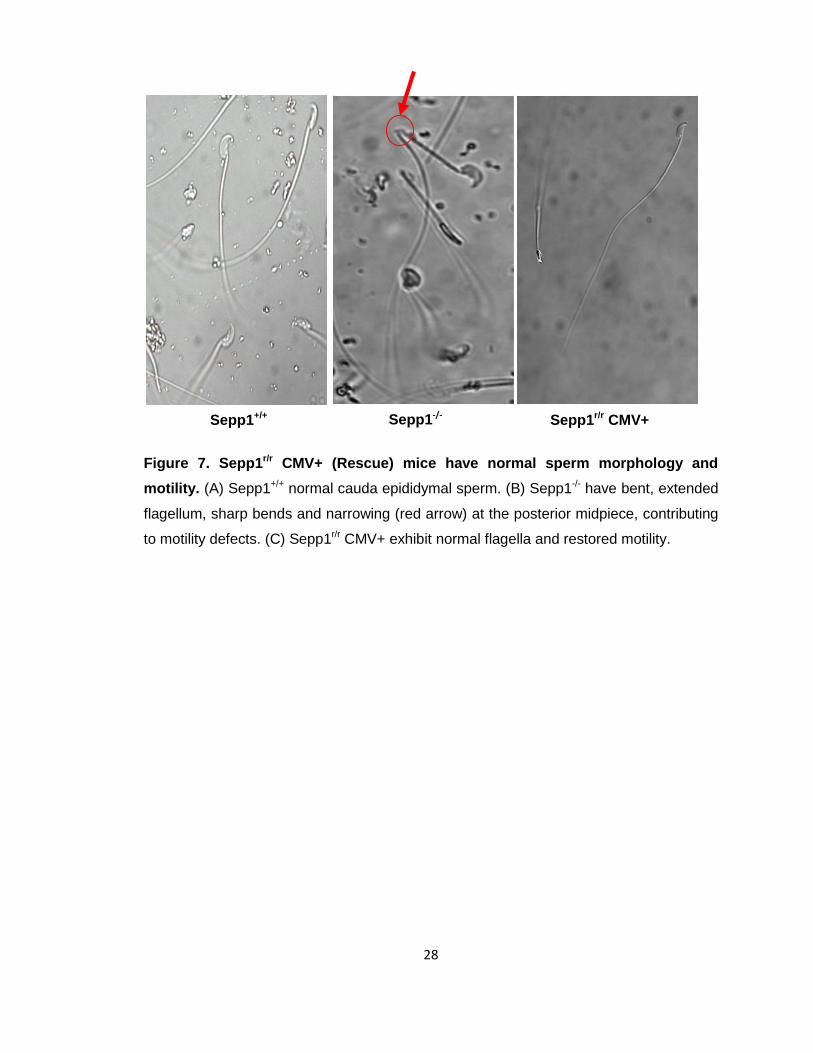
Sepp1r/r CMV+ mice have normal sperm morphology and motility compared to
Sepp1-/- mice. Sepp1 KO mice have been shown to have abnormal sperm morphology.
Structural differences in sperm have been implicated as being central to the infertility of
Sepp1-/- mice [61]. As Sepp1r/r CMV+ male mice were able to sire offspring, we
investigated if they had restored normal sperm morphology. Sperm from cauda
epididymides of Sepp1r/r CMV+ mice as well as those of Sepp1+/+ and Sepp1-/- mice were
harvested for comparison. Microscopy imaging observations confirmed that the sperm
from the rescue mice, Sepp1r/r CMV+, had completely normal sperm morphology similar
to Sepp1+/+ absent of any kinks in the flagellum or narrowing at the posterior midpiece
(Fig. 7). Additionally, Sepp1 rescue mice had normal sperm motility patterns and sperm
numbers as compared to the controls.
Sepp1 is required for viable sperm DNA, independent of sperm morphology.
Previous studies from other groups have implicated the role of Sepp1 in spermiogenesis
and sperm morphology. Current literature shows that Sepp1-/- male mice have defective
sperm morphology contributing to their infertility. In particular, the sharply bent flagella of
Sepp1-/- sperm impede motility and prevent them from reaching oocytes and subsequent
fertilization. However, the possible contribution of other abnormalities such as DNA
aberrations to Sepp1-/- mice infertility has not been explored. Using intracytoplasmic
spermatozoa injection (ICSI), we found that Sepp1 male sperm have reduced viability
that is completely independent of motility. ICSI is a form of invitro fertilization (IVF) in
which sperm heads are isolated and the flagella are discarded. Sperm heads are then
directly injected into the oocyte, effectively excluding flagella morphology and motility as
contributing factors to Sepp1-/- infertility. Our novel ICSI results strongly demonstrate that
Sepp1 is required for sperm viability, independent of sperm flagella morphology (Table
1). The data show reductions in fertilization and development following injection of
Sepp1-/- sperm into oocytes compared to Sepp1+/- sperm. Sepp1-/- mice sperm exhibited
abnormalities reflected in the nearly 50% reduction in injected oocytes reaching the 2-
cell pronuclei (PN) stage compared to heterozygous controls. Despite having a single

22
copy of the Sepp1 gene, Sepp1+/- sperm injected into oocytes resulted in normal
fertilization and development at all stages observed. Only 45.3% of Sepp1-/- injected
oocytes developed into 2-cell pronuclei zygotes, while 80.7% of oocytes injected with
Sepp1+/- sperm progressed to 2-cell pronuclei zygotes. We also found that the rate of
Sepp1-/- 2-cell pronuclei zygotes that survived (33.7%) until transfer to surrogate mothers
was dramatically reduced compared to that of Sepp1+/- (76%). Finally, the rate of
oocytes injected resulting in live pups born was also greatly reduced. Sepp1+/- control
sperm injections resulted in 31 heterozygous pups, or 37.3% of the number of injected
oocytes, whereas oocytes injected with Sepp1-/- sperm produced only 5 heterozygous
pups, or 5.8% of the initially injected oocytes. Overall, Sepp1-/- sperm injected oocytes
resulted in a more than 6-fold reduction in live born pups in comparison to controls.
GPx4 protein expression is deficient in Sepp1 KO testes and epididymides. Sepp1
maintains Se homeostasis by transporting Se from the liver to other organs. To further
confirm our immunofluorescence microscopy results, we examined GPx4 expression by
western blot. Protein was extracted from testes and epididymides were isolated from
Sepp1+/+, Sepp1-/-, and Sepp1r/r CMV+. Western blot results show that GPx4 protein
expression is dramatically decreased in both testes (one-way ANOVA, **P=0.0023) and
epididymides of Sepp1-/- mice compared to Sepp1+/+ as well as Sepp1r/r CMV+ (Fig. 8).
As further evidence of the restoration of Sepp1 gene expression in Sepp1-/- mice by our
unique application of the Cre-LoxP system, GPx4 protein expression in Sepp1r/r CMV+
testes and epididymides were similar to that of wild type.
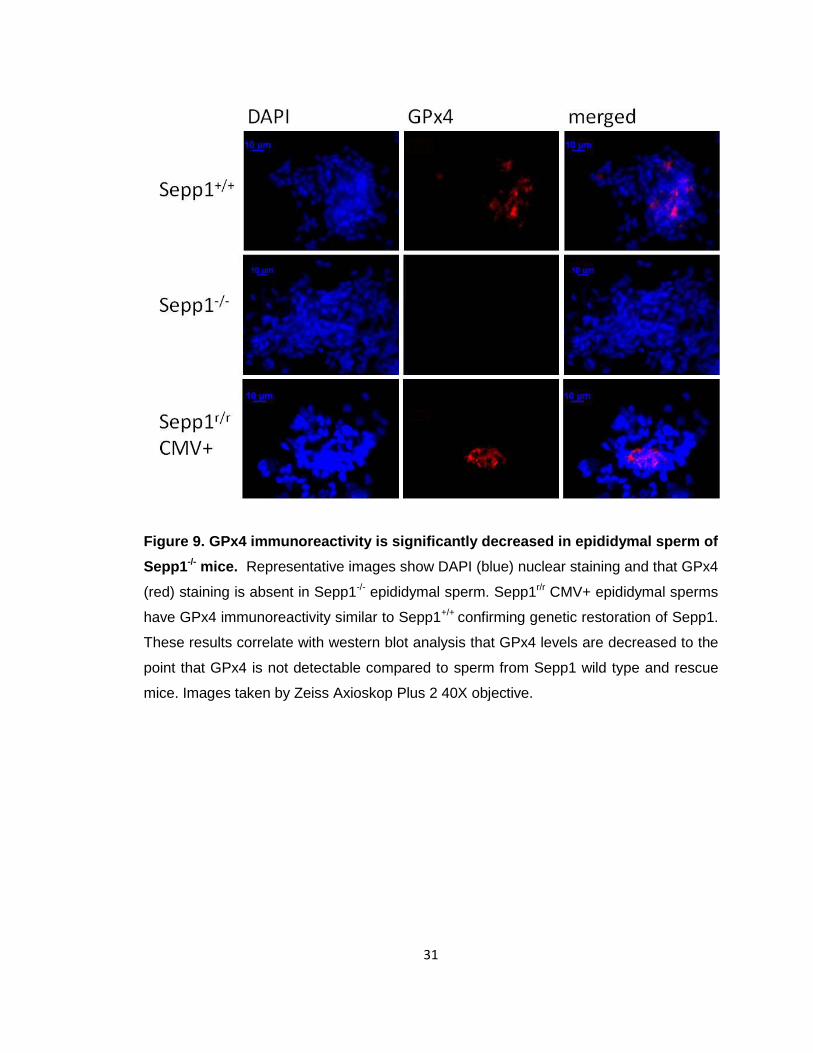
GPx4 immunoreactivity levels are decreased in Sepp1 KO epididymal sperm. The
importance of GPx4 in spermiogenesis is becoming increasingly clear. GPx4, a
phospholipid hydroperoxidase, exists in three isoforms [71], two of which have been
found to be important in spermiogenesis, sperm structure and function. Recent findings
indicate that nuclear GPx4 (nGPx4) is central to sperm chromatin organization [12].
Using immunofluorescence imaging, we demonstrate that nuclear GPx4
immunoreactivity is significantly decreased in Sepp1-/- epididymal sperm compared to
Sepp1+/+ sperm, as shown by the decreased co-localization of GPx4 signal with the
nuclear stain DAPI (Fig. 7). Furthermore, Sepp1r/r CMV+ rescue sperm GPx4 levels were
restored and were similar to that of wild type controls.

23
Figure 3. Generation of Sepp1r/r CMV+ rescue mice. (A) Polymerase chain reaction
(PCR) amplification of mouse tail genomic DNA was used to confirm the genotypes of all
animals. Specific primers were used to detect the 224-bp, 151-bp, and 472-bp product in
the Sepp1r/r CMV+ whole body rescue, Sepp1+/+, and Sepp1-/-, respectively. (B)
Schematic of post recombination of Sepp1 KO with CMV-Cre mice in which the Neor
containing construct flanked by Lox P was excised, using the Lox P site start codon (C)
that remained in frame with Sepp1 gene to restore Sepp1 translation.
A. B. A.
C.

24
Figure 4. Sepp1r/r CMV+ mice have normal motor coordination. (A) Total time taken
for Sepp1r/r CMV+ rescue mice to descend the pole on the pole test was similar to wild
type (WT) and significantly faster than Sepp1 KO mice as reflected in one way ANOVA
(P=0.0086) results. (B) Time taken for mice to coordinate and turn on the pole before
descending was significantly less for the Sepp1 rescue mice than Sepp1 KO mice, and
similar to normal WT, one way ANOVA (P=0.0024) with Bonferroni’s Post hoc analysis
performed for both (*P<0.05, **P<0.01). Values are expressed as mean ± SEM.
A.
B.

25
Figure 5. Abormal gait exhibited by Sepp1 KO mice are restored in Sepp1 Rescue
mice. (A) Representative paw print showing gait patterns of Sepp1 wildtype, KO, and
rescue mice. (B) Sepp1r/r CMV+ mice exhibited more uniform stride length which were
statistically significant compared to Sepp1 KO mice gait in the stride test. 2-way ANOVA
of stride length (P=0.0034) with Bonferroni post hoc analysis showing the difference
between genotype are significant (P<0.05). Length and width of hindpaw prints were
measured from the center of each paw for each set of prints as indicated by the red
lines.
A. B.

26
#
A.
B. C.
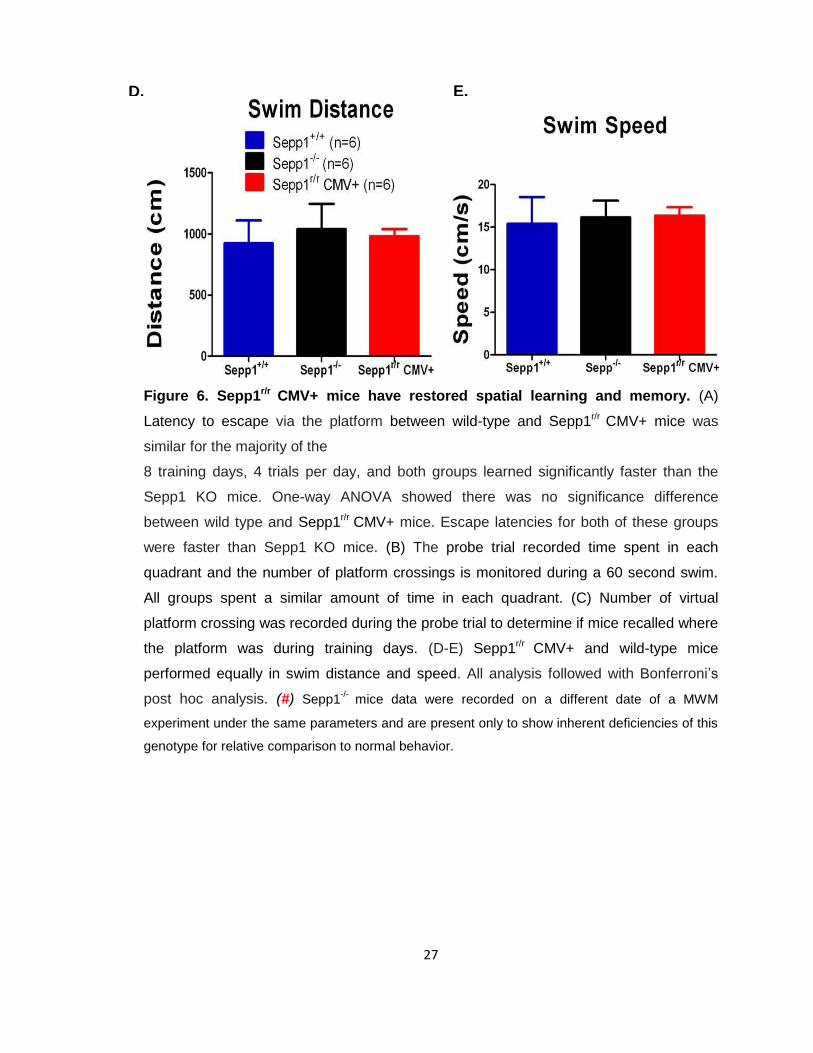
27
Figure 6. Sepp1r/r CMV+ mice have restored spatial learning and memory. (A)
Latency to escape via the platform between wild-type and Sepp1r/r CMV+ mice was
similar for the majority of the
8 training days, 4 trials per day, and both groups learned significantly faster than the
Sepp1 KO mice. One-way ANOVA showed there was no significance difference
between wild type and Sepp1r/r CMV+ mice. Escape latencies for both of these groups
were faster than Sepp1 KO mice. (B) The probe trial recorded time spent in each
quadrant and the number of platform crossings is monitored during a 60 second swim.
All groups spent a similar amount of time in each quadrant. (C) Number of virtual
platform crossing was recorded during the probe trial to determine if mice recalled where
the platform was during training days. (D-E) Sepp1r/r CMV+ and wild-type mice
performed equally in swim distance and speed. All analysis followed with Bonferroni’s
post hoc analysis. (#) Sepp1-/-
mice data were recorded on a different date of a MWM
experiment under the same parameters and are present only to show inherent deficiencies of this
genotype for relative comparison to normal behavior.
D. E.
28
Figure 7. Sepp1r/r CMV+ (Rescue) mice have normal sperm morphology and
motility. (A) Sepp1+/+ normal cauda epididymal sperm. (B) Sepp1-/- have bent, extended
flagellum, sharp bends and narrowing (red arrow) at the posterior midpiece, contributing
to motility defects. (C) Sepp1r/r CMV+ exhibit normal flagella and restored motility.
Sepp1+/+ Sepp1-/- Sepp1r/r CMV+

29
Table 1. Comparison of Sepp1 KO and Sepp1+/- male fertility. Sepp1 is critical in
sperm DNA viability, independent of flagella motility. Intracytoplasmic spermatozoa
injection (ICSI) shows that Sepp1 is required for normal sperm DNA viability,
independent of flagella morphology or motility. Sepp1-/- sperm injected oocytes resulted
in a more than 6-fold reduction in sperm viability in comparison to controls. Sepp1+/-
sperm injected oocytes resulted in 31 pups, 37.3% of the number of injected oocytes.
Oocytes injected with Sepp1-/- sperm produced only 5 heterozygous pups, 5.8% of the
initially injected oocytes.

30
Figure 8. GPx4 protein expression is significantly decreased in Sepp1-/- mice but
restored in Sepp1r/r CMV+ mice. GPx4 protein levels are significantly decreased in
Sepp1-/- mice testes (one-way ANOVA, **P= 0.0023) and epididymides. Sepp1r/r CMV+
testes and epididymides have similar GPx4 protein expression to Sepp1+/+ confirming
genetic restoration of Sepp1. These results show that Sepp1 is necessary for normal
GPx4 protein expression and that Sepp1r/r CMV+ mice have normal GPx4 protein levels
thus complete restoration of a functional Sepp1 gene.
31
Figure 9. GPx4 immunoreactivity is significantly decreased in epididymal sperm of
Sepp1-/- mice. Representative images show DAPI (blue) nuclear staining and that GPx4
(red) staining is absent in Sepp1-/- epididymal sperm. Sepp1r/r CMV+ epididymal sperms
have GPx4 immunoreactivity similar to Sepp1+/+ confirming genetic restoration of Sepp1.
These results correlate with western blot analysis that GPx4 levels are decreased to the
point that GPx4 is not detectable compared to sperm from Sepp1 wild type and rescue
mice. Images taken by Zeiss Axioskop Plus 2 40X objective.

32
CHAPTER IV
DISCUSSION and CONCLUSION
Sepp1 plays an important role in spermatogenesis and has been shown to be
essential to normal sperm morphology. Surprisingly, the issue of sperm DNA viability
due to the absence of Sepp1 has not been addressed. The results of this study
demonstrate that the infertility of male Sepp1-/- mice is partly due to sperm viability. Our
ICSI data clearly demonstrates for the first time that sperm heads, consisting mostly of
nuclei, are defective and result in impaired fertilization. Further, the developmental
defects exhibited in early stages of the fertilization process ultimately lead to a 72.3%
reduction in progeny number from oocytes injected with KO sperm compared to oocytes
injected with heterozygous control sperm. In addition, we show that GPx4 protein levels
in Sepp1 KO testes and epididymides are dramatically decreased overall and particularly
in sperm nuclei. Western blot shows decreased GPx4 immunoreactivity in Sepp1-/-
epididymal sperm compared to both Sepp1+/+ and Sepp1r/r CMV+ mice, and
immunofluorescence indicates that GPx4 colocalizes with the DAPI stained nucleus. We
also show that our unique strategy of using the existing Cre-LoxP sites in the Sepp1-/-
mice to restore the Sepp1 gene was completely successful as the Sepp1r/r CMV+ mice
had restored neuromotor function, spatial learning and memory, and fertility in males.
Furthermore, GPx4 expression was restored to normal levels similar to those of Sepp1+/+
mice.
Sepp1 is a plasma protein thought to mainly function to transport Se from the liver to
testes and other organs [18]. Studies show that Se is preferentially retained in brain and
testes when dietary Se intake is low. Reduced Se levels and glutathione peroxidase
(GPx) activity in reproductive organs of Se deficient male mice have been reported [81].
During spermatogenesis, Se content is noticeably increased in testes [82]. This requires
increased uptake of Se to testes. As a transporter of Se, Sepp1 may be responsible for
Se distribution during the reproductive process. Se deficiency is associated with
abnormal testicular mass, sperm morphology, and is essential for biosynthesis of
testosterone.
Sepp1 KO males are infertile and several studies have indicated that one of the main
contributing factors to this condition is defective sperm morphology [61]. Sepp1 KO

33
males have acute kinks in their sperm flagella rendering them unable to swim to the
oocyte, leading to infertility. Impairments from selenium deficiency can be restored in
wild type animals by dietary supplementation; however this is not the case for Sepp1-/-
mice. With dietary supplementation, neurological function of
Sepp1-/- mice is improved, but other deficits remain. Sepp1-/- mice have abnormal sperm
morphology regardless of supplementation. Sepp1 has direct antioxidant properties as
well as facilitating synthesis of antioxidant selenoproteins such as GPx4.
To test our first hypothesis whether other factors aside from defective sperm
morphology were involved in the infertility of male Sepp1-/- animals, we investigated
whether isolation of the sperm head from the defective flagella followed by ICSI would
produce normal fertilized oocytes and resulting litter size. As evident by our ICSI results,
genetic deletion of Sepp1 reduces viability of the sperm. We observed reduced
fertilization rates and impairments of early cell division stages in oocytes injected with
Sepp1-/- sperm compared to the oocytes injected with Sepp1+/- control sperm. Our data
show that Sepp1-/- male infertility is not entirely due to morphological sperm flagella
defects as suggested by other studies, as absence of flagella in the ICSI procedure did
not result in restored litter size.
It is not surprising that deletion of the Se transporter results in reduction in the levels
of other selenoproteins, such as GPx4. Previous studies have shown that GPx4 levels
are reduced in Sepp1-/- [75]. A 2009 study suggests that of the three GPx4 isoforms
(cytosolic GPx4, nGPx4, mGPx4), the mitochondrially expressed form of the GPx4 gene
is the most relevant one in spermiogenesis [77] while the nuclear form, nGPx4, is not
important for fertility [74,77]. However, a more recent study by Puglisi et. al, shows
convincing evidence that nGPx4 is associated with the sperm nuclear matrix and is
essential for sperm chromatin decondensation [71]. The nuclear isoform of GPx4 is
reportedly expressed in male germ cells. GPX4 contains a selenocysteine as well as
several cysteines with the ability to reduce protein thiols [71]. Our study supports
previous findings that impaired GPx4 biosynthesis, due to selenium deficiency or to
genetic defects in GPx4 itself or in proteins involved in Se distribution and selenoprotein
biosynthesis, results in male infertility [80]. We examined the GPx4 levels in whole
epididymis, epididymal sperm, and testes of the Sepp1 rescue and KO mice compared
to wild type controls to confirm the complete restoration of Sepp1 in our rescue model
and to confirm that the Sepp1-/- mice exhibited alterations in GPx4 levels as reported by

34
other groups. Our results confirmed reduced GPx4 protein expression in testes, where
testosterone and spermatozoa are synthesized. Moreover, we also showed that GPx4
protein expression was reduced notably in the epididymis, where sperm undergo a
maturation process and mature active sperm are stored in the cauda epididymis. To
further confirm the correlation of GPx4 levels, we used immunofluorescence microscopy
to assess GPx4 immunoreactivity. In support of our western blot data, we observed
drastically diminished GPx4 signal in Sepp1-/- epididymal sperm and restored levels of
GPx4 in our rescue mice. Our findings suggest that rendering the Sepp1-/- gene inactive
leads to decreased levels of GPx4. Sperm viability is clearly dependent on Sepp1 to
deliver Se and regulate biosynthesis of GPx4.
The latter part of our study was aimed to genetically restore Sepp1 gene expression
in Sepp1 knockout mice. We describe a strategy that takes advantage of the Cre-LoxP
system, using the start codon in the LoxP site, to restore gene expression in the Sepp1
KO mouse model. Previously the Schweizer lab generated a hepatically targeted Sepp1
transgene to rescue Sepp1-/- mice [75]. However, our approach is novel in that we use
the start codon in the existing LoxP site to genetically restore Sepp1 function, and can
therefore rescue Sepp1 in any tissues with the appropriate Cre and without additional
transgenic manipulations. One of our primary interests in generating this Sepp1 rescue
model is to apply this method to restrict gene expression to specific cells or tissues in
order to study the gene’s function within those cells or tissue. This unique application
can allow researchers to study any gene of interest in highly specific cell populations. A
researcher interested in such an approach to elucidate a specific function of a gene can
specifically design the knockout mouse to have LoxP sites in which the LoxP start codon
will remain in frame with the gene of interest following Cre recombination. Subsequent
mating of mice expressing Cre in specific cell types,, such as a subset of
cardiomyocytes or neurons, will excise the KO construct, leaving behind a LoxP site with
an in frame start codon. Following the successful generation of the Sepp1r/r CMV+ mice,
in which we restored global expression of Sepp1, we have used this unique strategy to
generate a brain specific Sepp1 rescue mouse model. Using a Cre expressing mouse
with a promoter that drives expression in forebrain neurons, this mouse model
expresses Sepp1 restricted to neurons in the forebrain, while Sepp1 gene expression
remains inactive in all other tissues. We confirmed the complete restoration of the Sepp1
gene by performing a battery of behavior tests which resulted in Sepp1r/r CMV+ mice

35
having restored neuromotor function as observed in the pole and stride tests.
Furthermore, Seppr/r CMV+ mice had normal spatial learning and memory and did not
exhibit any of the deficits seen in Sepp1-/- mice when administered the Morris Water
Maze test. Ultimately, the ability of Sepp1 rescue mice to sire pups that were used in the
behavior tests was indication that infertility seen in male KO mice was restored.
Furthermore, the sperm morphology studies confirmed that the rescue mice had normal
sperm morphology and motility.
Further research is essential to understanding the mechanisms involved in Se
function and the roles of various selenoproteins, such as Sepp1 and GPx4, in male
reproductive health. Some studies have suggested that Se deficient Balb/c mice show
decreased mRNA and protein expression patterns for both cJun and cFos (components
of transcription factor AP-1, activator protein-1) [82]. These factors regulate cellular
growth and differentiation and also have regulatory roles in spermatogenesis and
steroidogenesis [82]. Jun has been detected during specific stages of testes
differentiation [1,82]. cFos activity has also been observed in premeiotic germ cells of
mammalian testes during mouse spermatogenesis [46,82]. Studies show that Jun D
knockout mice have impaired spermatogenesis as well as reduced reproductive aptitude
[82,90]. cJun expression has been observed in Leydig cells, potentially enhancing
testosterone secretion, whereas in Se deficient mice, these cells as well as the
seminiferous tubules showed abnormal morphology [82]. Alteration in Se supply may
lead to these altered cJun and cFos patterns in the testicular germ cells, which might be
responsible for decreased germ cell number, differentiation and reduced fertility [82].
The study suggests that this may be an explanation for the mechanism of Se action in
regulating spermatogenesis [82].
As previously mentioned, Sepp1 and especially GPx4 have been reported to have
antioxidant properties protecting cells from oxidative stress and similar insults.
Investigation in sperm damage suggests a link between DNA fragmentation and
oxidative base damage. Lipid peroxidation also induces sperm damage [101]. There is
evidence showing that a significant proportion of the free-radical induced DNA damage
observed in human spermatozoa is due to oxidative processes [102]. Antioxidants such
as Sepp1 and GPx4 may have beneficial effects in alleviating sperm damage and may
be instrumental as a treatment. A study by Shalini et. al showed that sperm from dietary
Se deficient mice exhibited incomplete chromatin decondensation and increased

36
incidence of DNA breaks [81]. Sperm chromatin condensation in spermatogenesis is a
complex process that involves sequential replacement of the majority of histones by
transition proteins and protamines in testis [13,14,24]. During spermatozoa transit
through the epididymis from caput (head) to cauda (tail), protamine thiol oxidation is
completed and intra- and intermolecular cross-links are formed. Subsequently, a
transcriptionally inactive and tightly packed haploid genome results and renders sperm
nuclei more resistant to mechanical and chemical insults and mature [24]. There are
many potential molecular mechanisms underlying this sperm damage, but certainly
taking all these findings into consideration is an important factor.
The exciting results of this study increase our knowledge of the role of Sepp1 in
relation to GPx4 and male fertility. Significantly, our ICSI results highlight that Sepp1 KO
male mice are infertile due to sperm viability issues other than abnormal flagella
morphology. Our findings contribute to the further understanding of the function of Sepp1
in mammalian sperm infertility issues.

37
Concluding Remarks and Future Studies
Selenoprotein synthesis is dependent primarily on dietary intake of the trace element
Se. Selenoproteins have a unique energetically costly expenditure for biosynthesis. The
25 human (24 in mice) selenoproteins have highly diverse tissue and subcellular
localization with each exerting specific functions in the subcompartments in which they
reside [9,74]. The unique aspect of Sepp1 having 10 Sec residues suggests its
importance for synthesis of other selenoproteins and its role in preventing loss of Se
levels in brain and testes during low dietary Se intake [6,32,50]. Future experiments that
may help further elucidate the mechanism of Sepp1 in sperm and male infertility include
examining sperm chromatin decondensation and sperm fragmentation in the Sepp1-/-
mouse model. Elucidating the functional roles and significance of individual
selenoproteins, such as Sepp1 and GPx4, will eventually provide crucial insight into how
dietary Se and selenoproteins affect human health.

38
REFERENCES
[1] Alcivar AA, Hake LE, Hardy MP, Hecht NB. Increased levels of junB and c-jun mRNAs in male germ cells following testicular cell dissociation. Maximal stimulation in prepuberal animals. J. Biol. Chem. 1990 Nov 25;265(33):20160–20165.
[2] Arteel GE, Mostert V, Oubrahim H, Briviba K, Abel J, Sies H. Protection by selenoprotein P in human plasma against peroxynitrite-mediated oxidation and nitration. Biol. Chem. 1998 Sep;379(8-9):1201–1205.
[3] Arthur JR. The glutathione peroxidases. Cellular and Molecular Life Sciences. 2001 Feb;57(13):1825–1835.
[4] Bao L, Avshalumov MV, Patel JC, Lee CR, Miller EW, Chang CJ, et al. Mitochondria are the source of hydrogen peroxide for dynamic brain-cell signaling. J. Neurosci. 2009 Jul 15;29(28):9002–9010.
[5] Bar-Nun S. The Role of p97/Cdc48p in Endoplasmic Reticulum-Associated Degradation: From the Immune System to Yeast [Internet]. In: Wiertz E, Kikkert M, editors. Dislocation and Degradation of Proteins from the Endoplasmic Reticulum. Berlin/Heidelberg: Springer-Verlag; [cited 2012 Jul 1]. p. 95–125.Available from: http://www.springerlink.com/index/10.1007/3-540-28007-3_5
[6] Behne D, Hilmert H, Scheid S, Gessner H, Elger W. Evidence for specific selenium target tissues and new biologically important selenoproteins. Biochim Biophys Acta. 1988;966:12–21.
[7] Beilstein MA, Vendeland SC, Barofsky E, Jensen ON, Whanger PD. Selenoprotein W of rat muscle binds glutathione and an unknown small molecular weight moiety. Journal of Inorganic Biochemistry. 1996 Feb;61(2):117–124.
[8] Bellinger FP, He QP, Bellinger MT, Lin Y, Raman AV, White LR, et al. Association of selenoprotein P with Alzheimer’s pathology in human cortex. J Alzheimers Dis. 2008;15:465–472.
[9] Bellinger FP, Raman AV, Reeves MA, Berry MJ. Regulation and function of selenoproteins in human disease. Biochem. J. 2009 Aug 15;422(1):11–22.
[10] Berry MJ, Banu L, Harney JW, Larsen PR. Functional characterization of the eukaryotic SECIS elements which direct selenocysteine insertion at UGA codons. EMBO J. 1993 Aug;12(8):3315–3322.
[11] Berry MJ, Tujebajeva RM, Copeland PR, Xu X-M, Carlson BA, Martin GW, et al. Selenocysteine incorporation directed from the 3′UTR: Characterization of eukaryotic EFsec and mechanistic implications. BioFactors. 2001;14(1-4):17–24.

39
[12] Boitani C, Puglisi R. Selenium, a key element in spermatogenesis and male fertility. Adv. Exp. Med. Biol. 2008;636:65–73.
[13] Brewer L, Corzett M, Balhorn R. Condensation of DNA by spermatid basic nuclear proteins. J. Biol. Chem. 2002 Oct 11;277(41):38895–38900.
[14] Brewer LR, Corzett M, Balhorn R. Protamine-induced condensation and decondensation of the same DNA molecule. Science. 1999 Oct 1;286(5437):120–123.
[15] Brigelius-Flohe R. Selenium compounds and selenoproteins in cancer. Chem Biodivers. 2008;5:389–395.
[16] Burk RF, Hill KE, Motley AK, Austin LM, Norsworthy BK. Deletion of selenoprotein P upregulates urinary selenium excretion and depresses whole-body selenium content. Biochim. Biophys. Acta. 2006 Dec;1760(12):1789–1793.
[17] Burk RF, Hill KE. SELENOPROTEIN P: An Extracellular Protein with Unique Physical Characteristics and a Role in Selenium Homeostasis. Annual Review of Nutrition. 2005 Aug 21;25(1):215–235.
[18] Burk RF, Hill KE. Selenoprotein P-expression, functions, and roles in mammals. Biochim. Biophys. Acta. 2009 Nov;1790(11):1441–1447.
[19] Burk RF. Production of selenium deficiency in the rat. Meth. Enzymol. 1987;143:307–313.
[20] Carlson BA, Xu X-M, Gladyshev VN, Hatfield DL. Selective rescue of selenoprotein expression in mice lacking a highly specialized methyl group in selenocysteine tRNA. J. Biol. Chem. 2005 Feb 18;280(7):5542–5548.
[21] Carlson BA, Xu X-M, Kryukov GV, Rao M, Berry MJ, Gladyshev VN, et al. Identification and characterization of phosphoseryl-tRNA[Ser]Sec kinase. Proc. Natl. Acad. Sci. U.S.A. 2004 Aug 31;101(35):12848–12853.
[22] Chabory E, Damon C, Lenoir A, Kauselmann G, Kern H, Zevnik B, et al. Epididymis seleno-independent glutathione peroxidase 5 maintains sperm DNA integrity in mice. J. Clin. Invest. 2009 Jul;119(7):2074–2085.
[23] Chatot CL, Ziomek CA, Bavister BD, Lewis JL, Torres I. An improved culture medium supports development of random-bred 1-cell mouse embryos in vitro. J. Reprod. Fertil. 1989 Jul;86(2):679–688.
[24] Conrad M, Moreno SG, Sinowatz F, Ursini F, Kölle S, Roveri A, et al. The nuclear form of phospholipid hydroperoxide glutathione peroxidase is a protein thiol peroxidase contributing to sperm chromatin stability. Mol. Cell. Biol. 2005 Sep;25(17):7637–7644.
[25] Cooper ML, Adami HO, Gronberg H, Wiklund F, Green FR, Rayman MP. Interaction between single nucleotide polymorphisms in selenoprotein P and mitochondrial

40
superoxide dismutase determines prostate cancer risk. Cancer Res. 2008;68:10171–10177.
[26] Copeland PR, Fletcher JE, Carlson BA, Hatfield DL, Driscoll DM. A novel RNA binding protein, SBP2, is required for the translation of mammalian selenoprotein mRNAs. EMBO J. 2000 Jan 17;19(2):306–314.
[27] Fagegaltier D, Hubert N, Yamada K, Mizutani T, Carbon P, Krol A. Characterization of mSelB, a novel mammalian elongation factor for selenoprotein translation. EMBO J. 2000 Sep 1;19(17):4796–4805.
[28] Fernagut PO, Diguet E, Labattu B, Tison F. A simple method to measure stride length as an index of nigrostriatal dysfunction in mice. J. Neurosci. Methods. 2002 Jan 30;113(2):123–130.
[29] Fleming SM, Salcedo J, Fernagut P-O, Rockenstein E, Masliah E, Levine MS, et al. Early and progressive sensorimotor anomalies in mice overexpressing wild-type human alpha-synuclein. J. Neurosci. 2004 Oct 20;24(42):9434–9440.
[30] Flohé L. Selenium in mammalian spermiogenesis. Biol. Chem. 2007 Oct;388(10):987–995.
[31] Foresta C, Flohé L, Garolla A, Roveri A, Ursini F, Maiorino M. Male fertility is linked to the selenoprotein phospholipid hydroperoxide glutathione peroxidase. Biol. Reprod. 2002 Sep;67(3):967–971.
[32] Gromer S, Eubel JK, Lee BL, Jacob J. Human selenoproteins at a glance. Cell. Mol. Life Sci. 2005 Nov;62(21):2414–2437.
[33] Hill KE, Lloyd RS, Burk RF. Conserved nucleotide sequences in the open reading frame and 3’ untranslated region of selenoprotein P mRNA. Proc. Natl. Acad. Sci. U.S.A. 1993 Jan 15;90(2):537–541.
[34] Hill KE, Zhou J, McMahan WJ, Motley AK, Atkins JF, Gesteland RF, et al. Deletion of selenoprotein P alters distribution of selenium in the mouse. J. Biol. Chem. 2003 Apr 18;278(16):13640–13646.
[35] Hill KE, Zhou J, McMahan WJ, Motley AK, Burk RF. Neurological dysfunction occurs in mice with targeted deletion of the selenoprotein P gene. J. Nutr. 2004 Jan;134(1):157–161.
[36] Hoefig CS, Renko K, Köhrle J, Birringer M, Schomburg L. Comparison of different selenocompounds with respect to nutritional value vs. toxicity using liver cells in culture. J. Nutr. Biochem. 2011 Oct;22(10):945–955.
[37] Hoffmann PR, Berry MJ. The influence of selenium on immune responses. Mol Nutr Food Res. 2008 Nov;52(11):1273–1280.

41
[38] Hoffmann PR. Mechanisms by which selenium influences immune responses. Arch Immunol Ther Exp (Warsz. 2007;55:289–297.
[39] Holben DH, Smith AM. The diverse role of selenium within selenoproteins: a review. J Am Diet Assoc. 1999 Jul;99(7):836–843.
[40] Horibata Y, Hirabayashi Y. Identification and characterization of human ethanolaminephosphotransferase1. The Journal of Lipid Research. 2006 Dec 5;48(3):503–508.
[41] Hwang DY, Cho JS, Oh JH, Shim SB, Jee SW, Lee SH, et al. Differentially expressed genes in transgenic mice carrying human mutant presenilin-2 (N141I): correlation of selenoprotein M with Alzheimer’s disease. Neurochem Res. 2005;30:1009–1019.
[42] Imai H, Suzuki K, Ishizaka K, Ichinose S, Oshima H, Okayasu I, et al. Failure of the expression of phospholipid hydroperoxide glutathione peroxidase in the spermatozoa of human infertile males. Biol. Reprod. 2001 Feb;64(2):674–683.
[43] Jablonska E, Gromadzinska J, Sobala W, Reszka E, Wasowicz W. Lung cancer risk associated with selenium status is modified in smoking individuals by Sep15 polymorphism. Eur J Nutr. 2008;47:47–54.
[44] de Jesus LA, Hoffmann PR, Michaud T, Forry EP, Small-Howard A, Stillwell RJ, et al. Nuclear Assembly of UGA Decoding Complexes on Selenoprotein mRNAs: a Mechanism for Eluding Nonsense-Mediated Decay? Molecular and Cellular Biology. 2006 Feb 14;26(5):1795–1805.
[45] Kehr S, Malinouski M, Finney L, Vogt S, Labunskyy VM, Kasaikina MV, et al. X-ray fluorescence microscopy reveals the role of selenium in spermatogenesis. J. Mol. Biol. 2009 Jun 26;389(5):808–818.
[46] Kierszenbaum AL. Mammalian spermatogenesis in vivo and in vitro: a partnership of spermatogenic and somatic cell lineages. Endocr. Rev. 1994 Feb;15(1):116–134.
[47] Kim H-Y, Gladyshev VN. Different catalytic mechanisms in mammalian selenocysteine- and cysteine-containing methionine-R-sulfoxide reductases. PLoS Biol. 2005 Dec;3(12):e375.
[48] Kimura Y, Yanagimachi R. Intracytoplasmic sperm injection in the mouse. Biol. Reprod. 1995 Apr;52(4):709–720.
[49] Koga M, Tanaka H, Yomogida K, Tsuchida J, Uchida K, Kitamura M, et al. Expression of selenoprotein-P messenger ribonucleic acid in the rat testis. Biol. Reprod. 1998 Jan;58(1):261–265.
[50] Korotkov KV, Novoselov SV, Hatfield DL, Gladyshev VN. Mammalian selenoprotein in which selenocysteine (Sec) incorporation is supported by a new form of Sec insertion sequence element. Mol. Cell. Biol. 2002 Mar;22(5):1402–1411.

42
[51] Lee BJ, Worland PJ, Davis JN, Stadtman TC, Hatfield DL. Identification of a selenocysteyl-tRNA(Ser) in mammalian cells that recognizes the nonsense codon, UGA. J. Biol. Chem. 1989 Jun 15;264(17):9724–9727.
[52] Leestma JE, Sepsenwol S. Sperm tail axoneme alterations in the Wobbler mouse. J. Reprod. Fertil. 1980 Jan;58(1):267–270.
[53] Low SC, Berry MJ. Knowing when not to stop: selenocysteine incorporation in eukaryotes. Trends Biochem. Sci. 1996 Jun;21(6):203–208.
[54] Mariotti M, Ridge PG, Zhang Y, Lobanov AV, Pringle TH, Guigo R, et al. Composition and evolution of the vertebrate and mammalian selenoproteomes. PLoS ONE. 2012;7(3):e33066.
[55] Messaoudi I, Banni M, Saïd L, Saïd K, Kerkeni A. Involvement of selenoprotein P and GPx4 gene expression in cadmium-induced testicular pathophysiology in rat. Chem. Biol. Interact. 2010 Oct 6;188(1):94–101.
[56] Michiels C, Raes M, Toussaint O, Remacle J. Importance of Se-glutathione peroxidase, catalase, and Cu/Zn-SOD for cell survival against oxidative stress. Free Radic. Biol. Med. 1994 Sep;17(3):235–248.
[57] Mistry HD, Broughton Pipkin F, Redman CWG, Poston L. Selenium in reproductive health. Am. J. Obstet. Gynecol. 2012 Jan;206(1):21–30.
[58] Moriarty PM, Reddy CC, Maquat LE. Selenium deficiency reduces the abundance of mRNA for Se-dependent glutathione peroxidase 1 by a UGA-dependent mechanism likely to be nonsense codon-mediated decay of cytoplasmic mRNA. Mol. Cell. Biol. 1998 May;18(5):2932–2939.
[59] Moschos MP. Selenoprotein P. Cell. Mol. Life Sci. 2000 Dec;57(13-14):1836–1845.
[60] Motsenbocker MA, Tappel AL. A selenocysteine-containing selenium-transport protein in rat plasma. Biochimica et Biophysica Acta (BBA) - General Subjects. 1982 Oct;719(1):147–153.
[61] Olson GE, Winfrey VP, Nagdas SK, Hill KE, Burk RF. Selenoprotein P is required for mouse sperm development. Biol. Reprod. 2005 Jul;73(1):201–211.
[62] Olson GE, Winfrey VP, Nagdas SK, Hill KE, Burk RF. Apolipoprotein E receptor-2 (ApoER2) mediates selenium uptake from selenoprotein P by the mouse testis. J. Biol. Chem. 2007 Apr 20;282(16):12290–12297.
[63] Panee J, Stoytcheva ZR, Liu W, Berry MJ. Selenoprotein H Is a Redox-sensing High Mobility Group Family DNA-binding Protein That Up-regulates Genes Involved in Glutathione Synthesis and Phase II Detoxification. Journal of Biological Chemistry. 2007 Jun 21;282(33):23759–23765.

43
[64] Papp LV, Lu J, Holmgren A, Khanna KK. From Selenium to Selenoproteins: Synthesis, Identity, and Their Role in Human Health. Antioxidants & Redox Signaling. 2007 Jul;9(7):775–806.
[65] Paul C, Nagano M, Robaire B. Aging results in differential regulation of DNA repair pathways in pachytene spermatocytes in the Brown Norway rat. Biol. Reprod. 2011 Dec;85(6):1269–1278.
[66] Peters MM, Hill KE, Burk RF, Weeber EJ. Altered hippocampus synaptic function in selenoprotein P deficient mice. Mol Neurodegener. 2006;1:12.
[67] Petit N. Selenoprotein N: an endoplasmic reticulum glycoprotein with an early developmental expression pattern. Human Molecular Genetics. 2003 May 1;12(9):1045–1053.
[68] Pfeifer H, Conrad M, Roethlein D, Kyriakopoulos A, Brielmeier M, Bornkamm GW, et al. Identification of a specific sperm nuclei selenoenzyme necessary for protamine thiol cross-linking during sperm maturation. FASEB J. 2001 May;15(7):1236–1238.
[69] Pilder SH, Olds-Clarke P, Orth JM, Jester WF, Dugan L. Hst7: a male sterility mutation perturbing sperm motility, flagellar assembly, and mitochondrial sheath differentiation. J. Androl. 1997 Dec;18(6):663–671.
[70] Prut L, Belzung C. The open field as a paradigm to measure the effects of drugs on anxiety-like behaviors: a review. Eur. J. Pharmacol. 2003 Feb 28;463(1-3):3–33.
[71] Puglisi R, Maccari I, Pipolo S, Conrad M, Mangia F, Boitani C. The nuclear form of glutathione peroxidase 4 is associated with sperm nuclear matrix and is required for proper paternal chromatin decondensation at fertilization. J. Cell. Physiol. 2012 Apr;227(4):1420–1427.
[72] Raman AV, Pitts MW, Seyedali A, Hashimoto AC, Seale LA, Bellinger FP, et al. Absence of selenoprotein P but not selenocysteine lyase results in severe neurological dysfunction. Genes Brain Behav. 2012 Jul;11(5):601–613.
[73] Reeves MA, Bellinger FP, Berry MJ. The neuroprotective functions of selenoprotein M and its role in cytosolic calcium regulation. Antioxid. Redox Signal. 2010 Apr 1;12(7):809–818.
[74] Reeves MA, Hoffmann PR. The human selenoproteome: recent insights into functions and regulation. Cellular and Molecular Life Sciences. 2009 Apr 28;66(15):2457–2478.
[75] Renko K, Werner M, Renner-Müller I, Cooper TG, Yeung CH, Hollenbach B, et al. Hepatic selenoprotein P (SePP) expression restores selenium transport and prevents infertility and motor-incoordination in Sepp-knockout mice. Biochem. J. 2008 Feb 1;409(3):741–749.

44
[76] Sauer B, Henderson N. Site-specific DNA recombination in mammalian cells by the Cre recombinase of bacteriophage P1. Proc. Natl. Acad. Sci. U.S.A. 1988 Jul;85(14):5166–5170.
[77] Schneider M, Forster H, Boersma A, Seiler A, Wehnes H, Sinowatz F, et al. Mitochondrial glutathione peroxidase 4 disruption causes male infertility. The FASEB Journal. 2009 May 5;23(9):3233–3242.
[78] Schomburg L, Schweizer U, Holtmann B, Flohe L, Sendtner M, Kohrle J. Gene disruption discloses role of Selenoprotein P in selenium delivery to target tissues. Biochem J. 2003;370:397–402.
[79] Scott KA, Yamazaki Y, Yamamoto M, Lin Y, Melhorn SJ, Krause EG, et al. Glucose parameters are altered in mouse offspring produced by assisted reproductive technologies and somatic cell nuclear transfer. Biol. Reprod. 2010 Aug 1;83(2):220–227.
[80] Shalini S, Bansal MP. Co-operative effect of glutathione depletion and selenium induced oxidative stress on API and NFkB expression in testicular cells in vitro: insights to regulation of spermatogenesis. Biol. Res. 2007;40(3):307–317.
[81] Shalini S, Bansal MP. Dietary selenium deficiency as well as excess supplementation induces multiple defects in mouse epididymal spermatozoa: understanding the role of selenium in male fertility. Int. J. Androl. 2008 Aug;31(4):438–449.
[82] Shalini S, Bansal MP. Role of selenium in spermatogenesis: differential expression of cjun and cfos in tubular cells of mice testis. Mol. Cell. Biochem. 2006 Nov;292(1-2):27–38.
[83] Small-Howard A, Morozova N, Stoytcheva Z, Forry EP, Mansell JB, Harney JW, et al. Supramolecular complexes mediate selenocysteine incorporation in vivo. Mol. Cell. Biol. 2006 Mar;26(6):2337–2346.
[84] Squires JE, Stoytchev I, Forry EP, Berry MJ. SBP2 Binding Affinity Is a Major Determinant in Differential Selenoprotein mRNA Translation and Sensitivity to Nonsense-Mediated Decay. Molecular and Cellular Biology. 2007 Sep 10;27(22):7848–7855.
[85] Sternberg N, Hamilton D, Hoess R. Bacteriophage P1 site-specific recombination. II. Recombination between loxP and the bacterial chromosome. J. Mol. Biol. 1981 Aug 25;150(4):487–507.
[86] Sunde RA, Raines AM, Barnes KM, Evenson JK. Selenium status highly-regulates selenoprotein mRNA levels for only a subset of the selenoproteins in the selenoproteome. Biosci Rep. 2008;
[87] Takebe G, Yarimizu J, Saito Y, Hayashi T, Nakamura H, Yodoi J, et al. A comparative study on the hydroperoxide and thiol specificity of the glutathione peroxidase family and selenoprotein P. J. Biol. Chem. 2002 Oct 25;277(43):41254–41258.

45
[88] Takemoto AS, Berry MJ, Bellinger FP. Role of selenoprotein P in Alzheimer’s disease. Ethn Dis. 2010;20(1 Suppl 1):S1–92–5.
[89] Tamura T, Stadtman T. Mammalian thioredoxin reductases [Internet]. In: Methods in Enzymology. Elsevier; 2002 [cited 2012 Jul 1]. p. 297–306.Available from: http://linkinghub.elsevier.com/retrieve/pii/S0076687902470292
[90] Thépot D, Weitzman JB, Barra J, Segretain D, Stinnakre MG, Babinet C, et al. Targeted disruption of the murine junD gene results in multiple defects in male reproductive function. Development. 2000 Jan;127(1):143–153.
[91] Tujebajeva RM, Copeland PR, Xu XM, Carlson BA, Harney JW, Driscoll DM, et al. Decoding apparatus for eukaryotic selenocysteine insertion. EMBO Rep. 2000 Aug;1(2):158–163.
[92] Vendeland SC. Rat Skeletal Muscle Selenoprotein W: cDNA Clone and mRNA Modulation by Dietary Selenium. Proceedings of the National Academy of Sciences. 1995 Sep 12;92(19):8749–8753.
[93] Walsh RN, Cummins RA. The Open-Field Test: a critical review. Psychol Bull. 1976 May;83(3):482–504.
[94] Weiss Sachdev S, Sunde RA. Selenium regulation of transcript abundance and translational efficiency of glutathione peroxidase-1 and -4 in rat liver. Biochem. J. 2001 Aug 1;357(Pt 3):851–858.
[95] Xu X-M, Carlson BA, Mix H, Zhang Y, Saira K, Glass RS, et al. Biosynthesis of selenocysteine on its tRNA in eukaryotes. PLoS Biol. 2007 Jan;5(1):e4.
[96] Xu X-M, Carlson BA, Zhang Y, Mix H, Kryukov GV, Glass RS, et al. New developments in selenium biochemistry: selenocysteine biosynthesis in eukaryotes and archaea. Biol Trace Elem Res. 2007 Dec;119(3):234–241.
[97] Xu X-M, Mix H, Carlson BA, Grabowski PJ, Gladyshev VN, Berry MJ, et al. Evidence for direct roles of two additional factors, SECp43 and soluble liver antigen, in the selenoprotein synthesis machinery. J. Biol. Chem. 2005 Dec 16;280(50):41568–41575.
[98] Yamazaki Y, Wakayama T, Yanagimachi R. Contribution of cumulus cells and serum to the maturation of oocyte cytoplasm as revealed by intracytoplasmic sperm injection (ICSI). Zygote. 2001 Nov;9(4):277–282.
[99] Yang GQ, Ge KY, Chen JS, Chen XS. Selenium-related endemic diseases and the daily selenium requirement of humans. World Rev Nutr Diet. 1988;55:98–152.
[100] Yang JG, Hill KE, Burk RF. Dietary selenium intake controls rat plasma selenoprotein P concentration. J. Nutr. 1989 Jul;119(7):1010–1012.

46
[101] Zhou BF, Stamler J, Dennis B, Moag-Stahlberg A, Okuda N, Robertson C, et al. Nutrient intakes of middle-aged men and women in China, Japan, United Kingdom, and United States in the late 1990s: the INTERMAP study. J Hum Hypertens. 2003 Sep;17(9):623–630.
[102] Zribi N, Chakroun NF, Elleuch H, Abdallah FB, Ben Hamida AS, Gargouri J, et al. Sperm DNA fragmentation and oxidation are independent of malondialdheyde. Reprod. Biol. Endocrinol. 2011;9:47.





























