Embed Size (px)
Citation preview

INTRODUCTION
The nuclear mitotic apparatus protein (NuMA) was firstdescribed by Lyderson and Pettijohn (1980) as a predominantlynuclear protein that is present in the interphase nucleus and isconcentrated in the spindle pole of mitotic cells. Recently,several nuclear proteins with an immunofluorescence stainingpattern similar to that of NuMA have been defined. Theseproteins, including centrophilin (Tousson et al., 1991), SPN(Kallajoki et al., 1991), SP-H (Maekawa et al., 1991), 1H1/1F1(Compton et al., 1991) and W1 (Tang et al., 1993b), have beendefined by various monoclonal antibodies or autoimmune sera.According to the amino acid or nucleotide sequences obtained,NuMA (Compton et al., 1992; Yang et al., 1992), SPN(Kallajoki et al., 1993), SP-H (Maekawa and Kuriyama, 1993),1H1/1F1 (Compton et al., 1992) and W1 (Tang et al., 1993b)represent different names for the same protein.
Recently, cDNA clones that cover the entire coding regionof human NuMA have been isolated and sequenced (Comptonet al., 1992; Yang et al., 1992; Tang et al., 1993b; Maekawaand Kuriyama, 1993). Secondary structure predictions revealthat NuMA is composed of a long
α-helical central coreflanked by two globular domains (Compton et al., 1992; Yanget al., 1992; Tang et al., 1993b; Maekewa and Kuriyama,1993). Interestingly, the central α-helical region of NuMAshares a high degree of sequence similarity with myosin,
lamins, intermediate filaments and other coiled-coil proteins.This observation suggests that NuMA belongs to the largecoiled-coil protein family.
On the basis of microinjection and cell-cycle-dependentlocalization experiments, several laboratories have proposedthat NuMA may play a role in maintaining or establishingnuclear structure, possibly as a structural component of thenuclear matrix (Lyderson and Pettijohn, 1980; Price andPettijohn, 1986; Kallajoki et al., 1991; Compton et al., 1992;Yang and Snyder, 1992). Recently, Compton and Cleveland(1993) have shown that expression of human NuMA lackingits globular head or tail domains may induce cells to becomemicronucleated. Moreover, expression of wild-type humanNuMA in tsBN2 cells, in which the endogenous NuMA proteinis normally degraded, resulting in micronuclei at a restrictivetemperature, was sufficient to suppress the micronucleationphenotype. These results lead them to suggest that NuMA isrequired for the proper terminal phases of chromosome sepa-ration and/or nuclear reassembly during mitosis. In addition,microinjection of anti-NuMA antibodies into early mitotic ormetaphase cells prevents the formation or causes the collapseof the mitotic spindle apparatus (Yang and Snyder, 1992), sug-gesting that NuMA may play an important role during mitosis.
We have recently isolated a series of overlapping cDNAclones that cover the entire coding region of NuMA (Tang etal., 1993b). Analysis of various cDNA clones revealed that
1389
We have recently shown that the nuclear mitotic apparatusprotein (NuMA) is composed of at least three isoforms thatdiffer mainly at the carboxy terminus, and are generatedby alternative splicing of a common mRNA precursor froma single NuMA gene (
J. Cell Sci. (1993) 104, 249-260).Transient expression of human NuMA-l isoform(T33/p230) in Chinese hamster ovary polyoma (CHOP)cells showed that NuMA-l was present in interphase nucleiand was concentrated at the polar regions of the spindleapparatus in mitotic cells. However, expression of twoother isoforms (NuMA-m and -s) revealed a distinct sub-cellular localization. NuMA-m (U4/p195) and NuMA-s(U6/p194) were present in the interphase cytosol andappeared to be mainly located at the centrosomal region.When cells entered into mitosis, however, NuMA-m and -s
moved to the mitotic spindle pole. Analysis of a series oflinker scanning-mutants and NuMA/β-galactosidasechimeric proteins showed that residues 1972-2007 ofNuMA-l constitute a novel nuclear localization signal(NLS) and residues 1538-2115 are necessary and sufficientfor spindle association. Further analysis of the NLS by site-specific mutagenesis indicated that Lys1988 is essential fornuclear targeting, whereas Arg1984 is not. These resultshave allowed us tentatively to assign specific biologicalactivities to distinct structural domains of the NuMApolypeptide.
Key words: nuclear protein, NuMA, centrosome, spindle pole,nuclear localization signal
SUMMARY
Nuclear mitotic apparatus protein (NuMA): spindle association, nuclear
targeting and differential subcellular localization of various NuMA isoforms
Tang K. Tang*, Chieh-ju C. Tang, Yu-Jane Chao and Cheng-Wen Wu
Institute of Biomedical Sciences, Academia Sinica, Taipei, Taiwan 115, Republic of China
*Author for correspondence
Journal of Cell Science 107, 1389-1402 (1994)Printed in Great Britain © The Company of Biologists Limited 1994

1390
their sequences were identical, except for six sequence blocks(75 bp, 42 bp, 45 bp, 576 bp, 1012 bp and 212 bp), which wereeither inserted or deleted in individual cDNA clones. Amongthese, two sequence blocks (75 bp and 42 bp), which encodea 25 and a 14 amino acid peptide, respectively, are located atthe middle portion of the NuMA cDNA. In contrast, the otherfour blocks (45 bp, 576 bp, 1012 bp, 212 bp), some of whichcarry a translation termination codon, are located at the 3′ endof the NuMA transcript (Tang et al., 1993b).
Interestingly, combinatorial splicing of four sequence blockslocated at the 3′ end of the transcript may generate at least threedifferent NuMA isoforms that differ at their carboxy termini.These isoforms are generated from a single RNA precursor byalternative splicing (Tang et al., 1993b). The T33 form ofNuMA mRNA codes for 2115 amino acids, corresponding tothe longest polypeptide of ~230 kDa (p230). Two other NuMAisoforms, which use different translation termination codonslocated in different 3′ sequence blocks, would producepolypeptides of lower molecular masses. The U4 form of themRNA would code for 1776 amino acids, corresponding top195, and the U6 form would code for 1763 amino acids, cor-responding to p194. For convenience in describing thesedifferent isoforms, the T33/p230 form is now renamed NuMA-l (large), the U4/p195 form NuMA-m (medium), and theU6/p194 form NuMA-s (small). All the isoforms describedhere contain both the 75 and 42 bp sequence blocks (Tang etal., 1993b) that encode 25 and a 14 amino acid (aa) peptides,respectively. For the isoform that lacks the 14 amino acidpeptide, the name of NuMA-
x (-14 aa) may be used, where xrepresents large (l), medium (m), or small (s). According tothis principle, the NuMA-x (-14 aa) represents an isoform thatlacks the 14 amino acid peptide, and so on.
To find out whether any particular isoform shows a subcel-lular localization identical to that reported for the originalNuMA, we have now expressed these three different isoformsof NuMA in cultured cells. Expression of the largest isoform(NuMA-l) showed a cell-cycle-dependent distribution indis-tinguishable from that of the original NuMA in human cells,i.e. present throughout the interphase nuclei but concentratedat the spindle poles during mitosis. To our surprise, expressionof NuMA-m and -s revealed a distinct subcellular localization.NuMA-m and -s were distributed in the cytoplasm of trans-fected cells and appeared to be present mainly at the centro-somal region. In this study, we have also mapped both thenuclear localization signal (NLS) and the spindle associationdomain of human NuMA-l protein. A model for both the struc-tural and functional domains of human NuMA-l is presented.
MATERIALS AND METHODS
Cell cultureThe Chinese hamster ovary cell line that expresses polyoma large Tantigen (CHOP) was kindly provided by Dr James W. Dennis,Toronto, Canada. CHOP cells were propagated and maintained inαMEM containing 10% fetal calf serum (FCS) and 200 µg/ml ofG418 (Gibco) as described previously (Heffernan and Dennis, 1991).
Construction of deletion and chimeric expressionplasmidsFull-length human NuMA cDNAs were assembled and constructedfrom overlapping cDNA fragments (Tang et al., 1993b) by standard
cloning methods (Ausubel et al., 1989; Sambrook et al., 1989). The3′ end DNA fragments of NuMA-l, -m and -s were derived from T33,U4 and U6 cDNA clones, respectively (Tang et al., 1993b). TheEcoRI fragments containing the entire coding sequence for NuMA-l,-m or -s were then inserted into the unique EcoRI site of a eukaryoticexpression vector, pSG5 (Strategene), which contains an SV40 earlypromoter, intron II of the rabbit β-globin gene, and the SV40polyadenylation signal. These constructs were renamed SV/NuMA-l,-m and -s, respectively (see Fig. 1A). All cDNA constructs containedboth the 75 bp and 42 bp sequence blocks (Tang et al., 1993b) in theircoding sequences. A series of truncated linker scaning mutants (seeFig. 6A) were generated by partial digestion of SV/NuMA-l cDNAwith NarI (L1), XhoI (L12) or StyI (L24, L36, L42), filling the 3′recessed termini with the Klenow fragment of Escherichia coli DNApolymerase I in the presence of the appropriate dNTPs and ligatedwith a phosphorylated XbaI linker (Stratagene). The linker is a shortoligonucleotide (CTAGTCTAGACTAG) that contains an XbaIrestriction site and stop codons in all three reading frames. This con-struction allowed us to generate a series of truncated mutants.
For generation of NuMA/β-galactosidase expression plasmids,various lengths of NuMA-l DNA fragments were PCR-amplified fromthe tail region of NuMA-l cDNA by several primer sets. The primersselected for generation of a series of pc NuMA/β-galactosidase con-structs (see Figs 7 and 9) were as follows: pc11 (4611-4629/6497-6517), pc6 (4611-4629/5906-5925), pc18 (5514-5533/6268-6287),pc20 (5913-5932/6268-6287), pc21 (5913-5932/6160-6179), pc23(6087-6106/6160-6179), pc24 (5913-5932/6004-6023), pc25 (6003-6022/6085-6104), pc30 (5940-5960/5964-5983) and pc31 (5940-5960/6004-6023). The numbers in parenthesis indicate the nucleotidepositions of NuMA according to the numbering system of Yang et al.(1992). The PCR amplified fragments were first subcloned into thePCR II vector (Invitrogen) for the convenience of direct cloning andrestriction site exchange. The PCR-amplified products in PCR II werethen released by EcoRI digestion followed by ligation into the uniqueEcoRI site located downstream of the carboxy terminus of the lacZgene in pCH110 vector. Because the primers for PCR amplificationof NuMA-l cDNA were specifically selected, the final PCR amplifiedproducts would be cloned in-frame with the lacZ gene. For genera-tion of the pc7 construct, which encodes a short polypeptide (sevenamino acids) of NuMA-l, two oligonucleotides (N3: 5′-GCGAA-TTCCAGCAGCGCAAACGGGTCTCCGAATTCCA-3′; and N4:5′-CGCTTAAGGTCGTCGCGTTTGCCCAGAGGCTTAAGGT-3′),which each carried an EcoRI recognition site at each end and werecomplementary to each other, were synthesized. Equal amounts ofcomplementary oligonucleotides were then annealed, followed byEcoRI digestion. The EcoRI-released fragment was then subclonedinto the unique EcoRI site of the lacZ gene in the pCH110 vector. Forconstruction of site-directed mutagenesis vectors, pc24 was selectedas a construction backbone for generation of both pc24N1 andpc24N2 mutant constructs. The primers selected for generation ofthese vectors were as follows: pc24N1 (5′-CATCACCACCGGG-CAGCAGCG-3′); pc24N2 (GCAGCAGCGCGAACGGGTCTC).The site-directed mutagenesis clones were generated by using thealtered sites in the in vitro mutagenesis system as described by themanufacturer (Promega). The same mutagenesis primer as describedfor pc24N1 was used to produce two other constructs, pc30 and pc31,which carry a single amino acid substitution at residue 1984 ofNuMA-l. The cDNA sequences in all the constructed mutants wereconfirmed by DNA sequencing.
Cell transfection and immunochemical analysisCHOP cells growing on glass coverslips were transiently transfectedusing Lipofectin (Gibco-BRL) reagent as described by the manufac-turer. (Lipofectin reagent is a 1:1 (w/w) liposome formulation of thecationic lipid N-[1-(2,3-dioleoyloxy)-propyl]-N,N,N-trimethylammo-nium chloride (DOTMA), and dioleoyl phosphatidylethanolamine(DOPE).) Briefly, 2 µg of plasmid DNA diluted in serum-free medium
T. K. Tang and others
1391Nuclear mitotic apparatus protein
was mixed with 7 µl of Lipofectin reagent and incubated at room tem-perature for 10-15 minutes. The solution containing Lipofectin reagent-DNA complexes was layered over cells at 60% confluency andincubated for 6 hours at 37°C. The cells were then washed with PBS(phosphate buffered saline), and grown in complete medium contain-ing 10% FCS for 18-20 hours. The transfected cells were then fixed inmethanol/acetone (1:1, v/v) at −20°C for 5 minutes. Monoclonalantibody W1 (Tang et al., 1993b) was applied to the cells and incubatedfor 60 minutes at 37°C in a humidified chamber. Coverslips werewashed in PBST (PBS containing 0.1% Tween-20) and the bound anti-bodies were detected with fluorescein-conjugated rabbit anti-mouseantibodies (Cappel Laboratories). DNA was detected with propidiumiodide (2.7 µg/ml; Sigma Chemical Co.). For double-labeling experi-ments, transfected cells were fixed and stained with mAb W1 and anti-tubulin antibody as described (Tang et al., 1993b). Coverslips weremounted and observed with a laser scanning confocal microscope(MRC600 model; Bio-Rad Laboratories; for all immunofluorescencephotos described in this paper, except Fig. 8A) or with a Zeiss Axiophotmicroscope equipped with epifluorescence (for Fig. 8A only).
For immunoblot analysis, CHOP cells were transfected with Trans-fectam reagent (Promega), since the transfection efficiency for Trans-fectam in CHOP cells was much higher than that observed for Lipo-fectin. Transfection efficiency was determined by analysis of 500CHOP cells that had been transfected with SV/NuMA-l cDNA anddetected by mAb W1. In general, 10-20% of the inoculated CHOPcells were positively stained by mAb W1 when the Transfectamreagent was used, whereas only 2-5% positive cells were observedusing the Lipofectin reagent. However, CHOP cells transfected withthe Transfectam reagent were not suitable for immunofluorescenceanalysis, because the precipitated DNA on the coverslip increased thebackground staining when we counterstained the nuclear DNA withpropidium iodide.
For immunoblot experiments, 1×106 cells were plated onto 100 mmPetri dishes. After overnight culture, the medium was aspirated andthe solution containing transfectam reagent-DNA complex wasoverlayed on the cells as described by the manufacturer. After 24hours of exposure, the cells were trypsinzied, washed with PBS andthen lysed in 60 µl EBC buffer (Tang et al., 1993b). The cell lysates
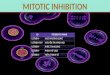
Fig. 1. (A) Schematic representation of various NuMA polypeptides derived from SV/NuMA expression vectors. The numbering system refersto amino acid residues on various NuMA isoforms. The carboxy-terminal 391, 52 and 39 residues are distinct for the NuMA-l, NuMA-m andNuMA-s proteins, respectively. The full-length human NuMA cDNAs that span the entire coding region for NuMA-l, -m and -s weresubcloned into a pSG5 expression vector (see Materials and Methods for details). All cDNA constructs contain both the 75 bp and 42 bpsequence blocks, which encode 25 and 14 amino acids (aa), respectively (black boxes). (B) Immunoblot analysis of various SV/NuMAisoforms after transient transfection of CHOP cells. Lane 1, SV/NuMA-s transfected CHOP cells; lane 2, SV/NuMA-m; lane 3, SV/NuMA-l;lane 4, mock-transfected CHOP cells. The low intensity band of ~194 kDa protein shown in lane 1 reflects the fact that the transfectionefficiency of SV/NuMA-s is ten times lower than that of SV/NuMA-m or SV/NuMA-l due to an unknown reason. (C) Subcellular localizationof human NuMA-l at various cell cycle stages of transfected CHOP cells. CHOP cells were transfected with SV/NuMA-I, fixed, and processedfor immunofluorescence with mAb W1 (a, b, c, d) and a DNA-specific dye, propidium iodide (a′, b′, c′, d′). Interphase (a, a′); metaphase (b, b′);anaphase (c, c′); telophase (d, d′). Bar, 25 µm.
1392
were then clarified by centrifugation at 12,000 g for 15 minutes at4°C. Cell lysates (30 µl) were then mixed with 5× SDS sample bufferand analyzed by 7.5% SDS-PAGE. After electrophoresis, the proteinswere transferred to polyvinylidene difluoride (PVDF) membrane andincubated with mAb W1 (Tang et al., 1993b). The immunoreactivepolypeptides were detected with the Western Exposure Chemilumi-nescent Detection System as described by the manufacturer(Clontech). Briefly, the immunoreactive polypeptides were detectedby the addition of goat anti-mouse IgG-alkaline phosphataseconjugate followed by incubation with chemiluminescent substratesolution for 5 minutes at room temperature. The membranes were thendrained and placed in contact with X-ray film to expose the film.
RESULTS
Expression of SV/NuMA-l reveals a subcellularlocalization pattern that is indistinguishable fromthat of endogeneous NuMA in human cellsTo define the subcellular localization of various NuMA
isoforms, we have developed a transient expression system.Three plasmids (SV/NuMA-l, -m and -s) that carry the entirecoding sequences of human NuMA-l, -m and -s cDNAs,respectively, were constructed and placed under the control ofthe SV40 early promoter (Fig. 1A). Each plasmid was intro-duced via transient transfection into CHOP cells, which werethen subjected to immunoblotting and confocal fluorescencemicroscopic analyses. The basis for detecting the expressedproduct corresponding to each of the constructs is a mono-clonal antibody (mAb W1) originally derived from miceimmunized with the nuclear residue from small cells of thebovine esophageal epithelial (BEE) basal layer (Tang et al.,1993a,b, 1994). This monoclonal antibody recognizes aspecific epitope of NuMA from human, bovine and murinecells (Tang et al., 1993b), but fails to recognize NuMA proteinsof hamster (CHOP) cells on the basis of both immunoblot (Fig.1B, lane 4) and immunofluorescence analyses (compare Fig.1C-a with 1C-a′).
Fig. 1B shows that transfected human NuMA-l in CHOP
T. K. Tang and others
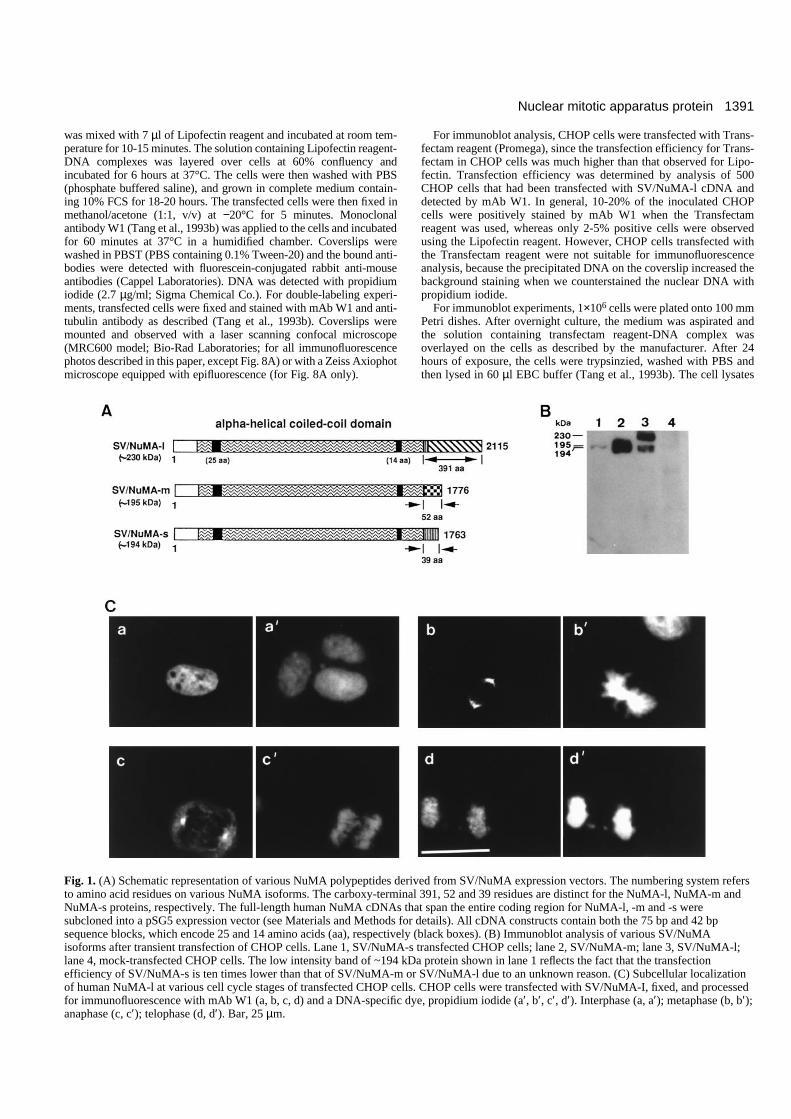
Fig. 2. Subcellular localization of human NuMA-m at various cell cycle stages of transfected CHOP cells. The SV/NuMA-m transfected cellswere double-stained with mAb W1 (left, green) and propidium iodide (center, red). The composite photographs are shown on the right. (a-c)Interphase; (d-f) metaphase; (g-i) telophase. Bar, 25 µm.
1393Nuclear mitotic apparatus protein
Fig
. 3.S
ubce
llula
rlo
caliz
atio
n of
hum
an N
uMA
-s a
tva
riou
s ce
ll cy
cle
stag
es o
f tr
ansf
ecte
dC
HO
P ce
lls. C
HO
Pce
lls w
ere
tran
sfec
ted
with
SV/N
uMA
-s. C
ells
wer
e fix
ed a
ndpr
oces
sed
for
imm
unofl
uore
scen
cew
ith m
Ab
W1
(lef
t,gr
een)
and
prop
idiu
m io
dide
(cen
ter,
red
). T
heco
mpo
site
phot
ogra
phs
of m
Ab
W1
and
prop
idiu
mio
dide
are
sho
wn
onth
e ri
ght.
(a-c
) In
terp
hase
;(d
-f)
met
apha
se;
(g-i
) te
loph
ase.
Bar
,25
µm
.
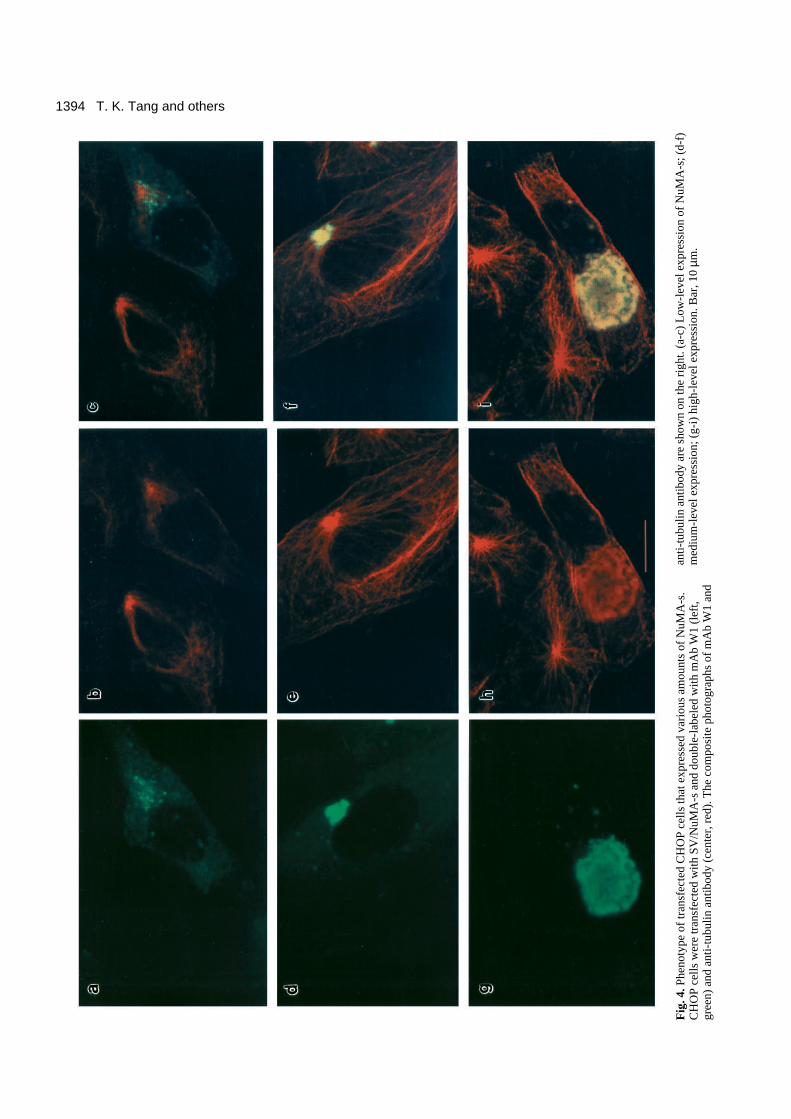
1394 T. K. Tang and others
Fig
. 4. P
heno
type
of
tran
sfec
ted
CH
OP
cells
that
exp
ress
ed v
ario
us a
mou
nts
of N
uMA
-s.
CH
OP
cells
wer
e tr
ansf
ecte
d w
ith S
V/N
uMA
-s a
nd d
oubl
e-la
bele
d w
ith m
Ab
W1
(lef
t,gr
een)
and
ant
i-tu
bulin
ant
ibod
y (c
ente
r, r
ed).
The
com
posi
te p
hoto
grap
hs o
f m
Ab
W1
and
anti-
tubu
lin a
ntib
ody
are
show
n on
the
righ
t. (a
-c)
Low
-lev
el e
xpre
ssio
n of
NuM
A-s
; (d-
f)m
ediu
m-l
evel
exp
ress
ion;
(g-
i) h
igh-
leve
l exp
ress
ion.
Bar
, 10
µm.
1395Nuclear mitotic apparatus protein
cells (lane 3) produced a protein with Mr ~230,000 (p230),which migrated according to the predicted size of the endoge-neous human NuMA (~230,000). The band observed in Fig.1B (lane 3) with a molecular mass lower than p230 wasprobably due to degradation; this band was not detected inextracts derived from mock-transfected CHOP cells (lane 4).Similarly, the predicted polypeptides corresponding to p194and p195 were also observed when CHOP cells were trans-fected with SV/NuMA-s (lane 1) or SV/NuMA-m (lane 2),respectively.
The subcellular localization of human NuMA isoformsexpressed in CHOP cells was further analyzed by fluorescenceconfocal microscopy using mAb W1. For cells at interphase,NuMA-l, like NuMA in human U251 cells (Tang et al., 1993b),accumulated in the nucleus of transfected CHOP cells but wasexcluded from the nucleoli (Fig. 1C-a). Note that severaluntransfected CHOP cells seen by staining with a DNA-specific dye (propidium iodide) (Fig. 1C-a′) did not react withmAb W1 (Fig. 1C-a). When cells entered metaphase, NuMA-l concentrated in the polar regions of the spindle apparatus,forming a characteristic crescent shape (Fig. 1C-b). Frommetaphase to anaphase (Fig. 1C-c), the crescent staining ofhuman NuMA-l diminished gradually but still remained at thespindle poles. Finally, in telophase cells, the human NuMA-lprotein associated with the re-forming nucleus and was subse-quently redistributed throughout the nucleus again (Fig. 1C-d).Similar results have been observed by Compton and Cleveland(1993). Taken together, the cell-cycle-dependent distributionof human NuMA-l in transfected CHOP cells is indistinguish-able from that in human cells.
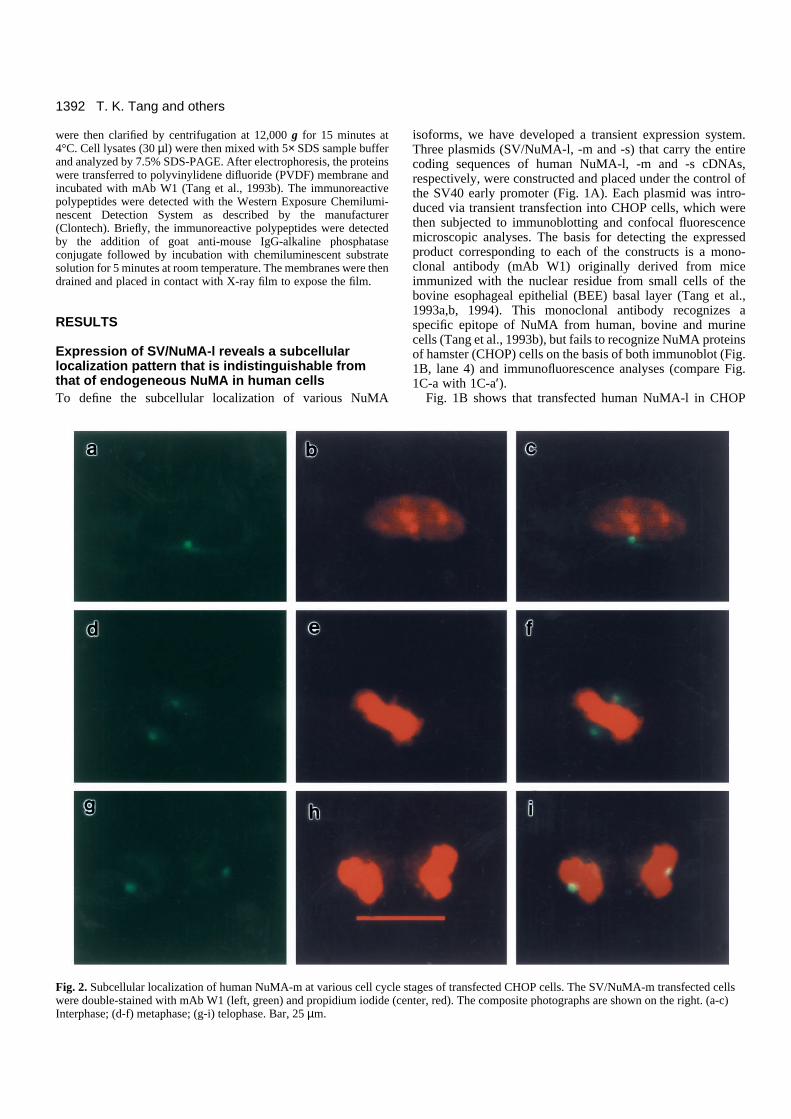
NuMA-m and NuMA-s are present in the interphasecytosol, mainly clustered at the centrosomal regionand move into the polar regions of the mitoticapparatus When we transfected SV/NuMA-m and SV/NuMA-s intoCHOP cells, a quite distinct subcellular localization wasobserved in interphase cells. In contrast to NuMA-l, which wasconcentrated exclusively in the interphase nuclei, NuMA-mand NuMA-s were unevenly distributed in the cytosol. In thepositive-staining interphase cells, NuMA-m and NuMA-s werepresent in the cytoplasm, but were prominently labeled as asmall round dot (Fig. 2a, NuMA-m; Fig. 3a, NuMA-s). Thesedots were most frequently seen in the perinuclear region, andsometimes they were observed above or below the nucleus(data not shown).
When the cells entered metaphase, both NuMA-m (Fig. 2d)and NuMA-s (Fig. 3d) concentrated in the polar regions of thespindle apparatus, a staining pattern similar to that observedfor NuMA-l (Fig. 1C-b). However, unlike the prominentcrescent staining observed for NuMA-l, staining in the shapeof a round dot was commonly found in NuMA-m and NuMA-s transfected cells. These results are illustrated in Fig. 2d-f(NuMA-m) and Fig. 3d-f (NuMA-s), which represent doublefluorescence analyses using mAb W1 (Figs 2d, 3d) and theDNA-specific dye, propidium iodide (Figs 2e, 3e). Composite,double-colored micrographs are shown in Figs 2f and 3f. Asthe cells progress through metaphase to telophase, theseparated daughter chromosomes arrive at the poles and eachgroup of chromosomes starts to form two daughter interphasenuclei. The prominent labeling of the small round dot remained
in the perinuclear position; the most likely interpretation ofthese observations is therefore that the labeled dot-like struc-tures correspond to the centrosomal region.
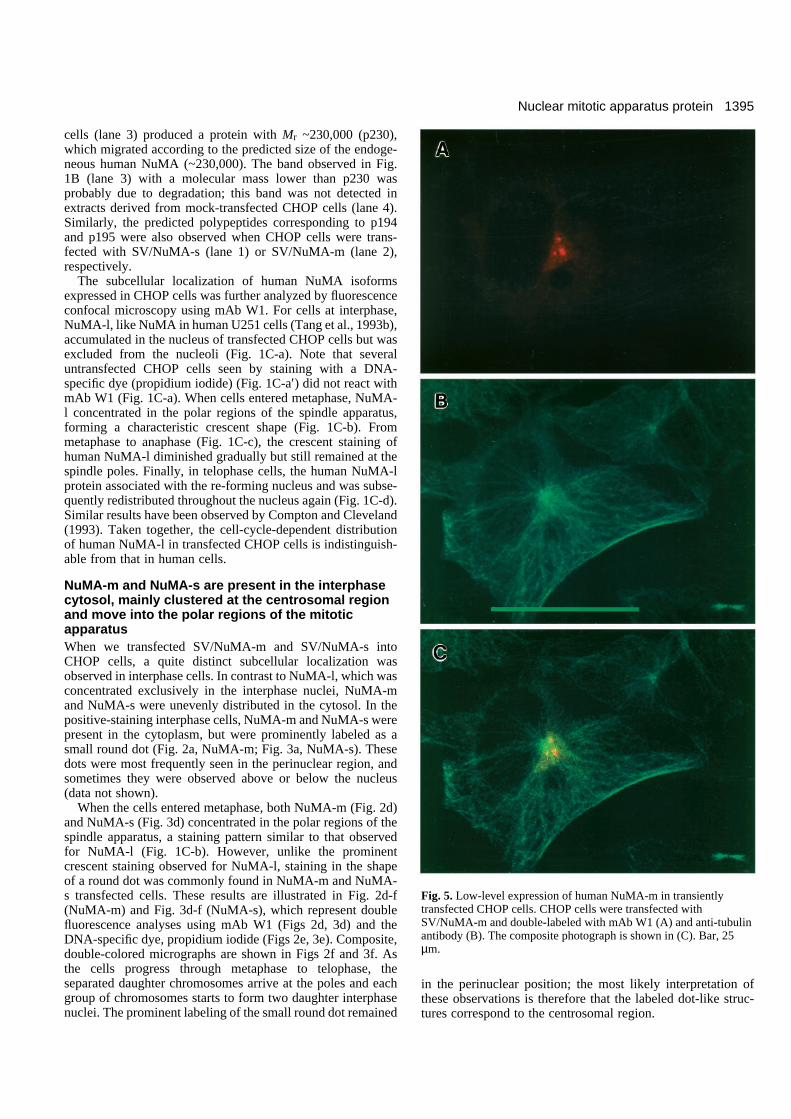
Fig. 5. Low-level expression of human NuMA-m in transientlytransfected CHOP cells. CHOP cells were transfected withSV/NuMA-m and double-labeled with mAb W1 (A) and anti-tubulinantibody (B). The composite photograph is shown in (C). Bar, 25µm.
1396
The phenomenon of NuMA-s association with the centroso-mal region is best illustrated by a comparison of a series ofphotographs that represent a simultaneous analysis of thestaining patterns of transfected CHOP cells that expressedvarious amounts of NuMA-s, after staining with mAbW1 (Fig.4, left) and anti-tubulin antibody (Fig. 4, center). Fig. 4a showsthat in transfected cells that express low levels of NuMA-smany dot-like structures appear to be concentrated at the inter-phase centrosome (microtubule organization center) (Fig. 4a-c). However, in medium-level expression cells, the dot-likestaining gradually aggregated and finally clustered at the regionaround the centrosome (Fig. 4d-f). Interestingly, in high-levelexpression cells, NuMA-s appears to be assembled into a largesheetlike aggregate within the cells (Fig. 4g). The large blurredstaining pattern detected by anti-tubulin antibody (4e and 4h)that was superimposed upon the staining of mAb W1 (4d and4g) could be a cross-reactive signal caused by overproductionof NuMA-s within the cells, since this signal was not detectedin low-level expression cells (Fig. 4b). In general, about 5-10%of transfected cells expressing NuMA-s showed a fluorescence
staining pattern (a small dot-like structure) similar to that inFig. 3a. Of the remaining cells expressing NuMA-s, ~10%showed a low-level expression pattern (as in Fig. 4a-c), ~50%showed a medium-level pattern (as in Fig. 4d-f), and ~30%showed a high-level pattern (as in Fig. 4g-h). A similar stainingpattern was also observed when we transfected SV/NuMA-minto CHOP cells that expressed NuMA-m (Fig. 5).
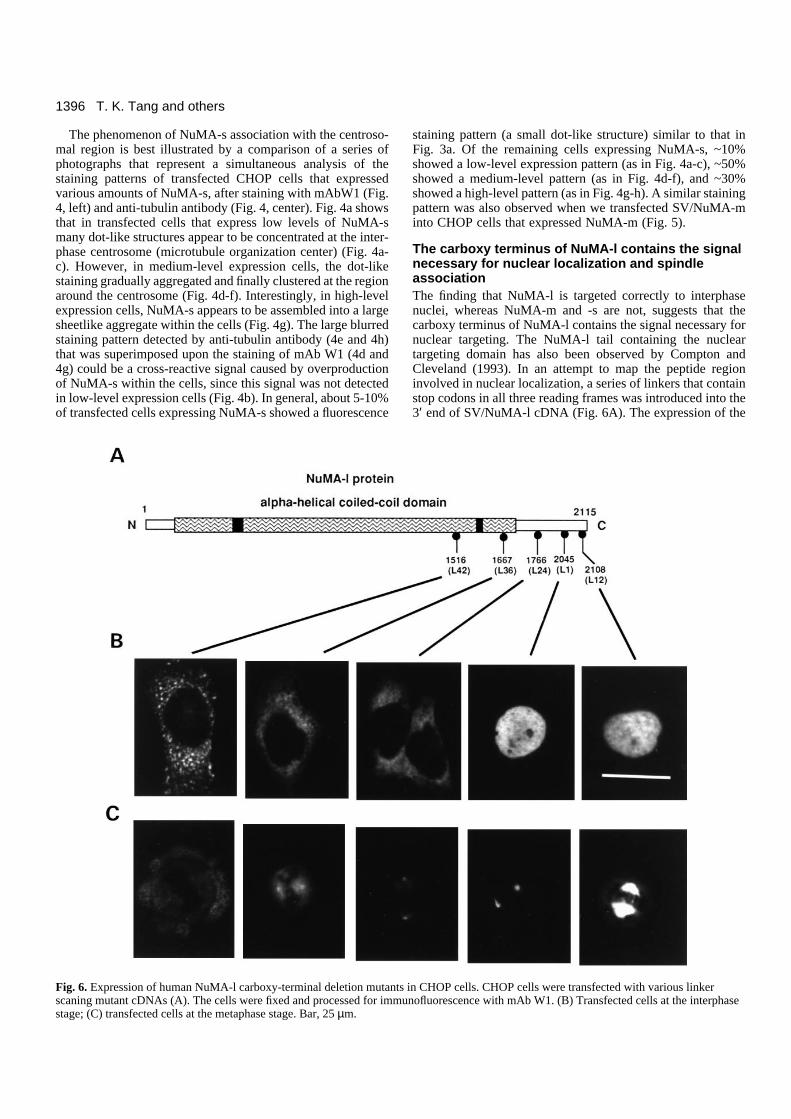
The carboxy terminus of NuMA-l contains the signalnecessary for nuclear localization and spindleassociationThe finding that NuMA-l is targeted correctly to interphasenuclei, whereas NuMA-m and -s are not, suggests that thecarboxy terminus of NuMA-l contains the signal necessary fornuclear targeting. The NuMA-l tail containing the nucleartargeting domain has also been observed by Compton andCleveland (1993). In an attempt to map the peptide regioninvolved in nuclear localization, a series of linkers that containstop codons in all three reading frames was introduced into the3′ end of SV/NuMA-l cDNA (Fig. 6A). The expression of the
T. K. Tang and others
Fig. 6. Expression of human NuMA-l carboxy-terminal deletion mutants in CHOP cells. CHOP cells were transfected with various linkerscaning mutant cDNAs (A). The cells were fixed and processed for immunofluorescence with mAb W1. (B) Transfected cells at the interphasestage; (C) transfected cells at the metaphase stage. Bar, 25 µm.

1397Nuclear mitotic apparatus protein
truncated linker scaning mutants in CHOP cells was analyzedwith mAb W1 by confocal fluorescence microscopy.
Fig. 6 shows the subcellular localization of a series ofcarboxy-terminal deletion mutants of NuMA-l. Linker 12(L12) and linker 1 (L1) mutants both targeted correctly to inter-phase nuclei (Fig. 6B) and mitotic spindle poles (Fig. 6C).However, when the carboxy-terminal end was removed up toor after linker 24 position, the nuclear staining disappeared.Instead, a prominent cytosolic localization was noticed (Fig.6B, L24). Some bright spots distributed in the cytosol werealso observed in L42-transfected cells (Fig. 6B). Interestingly,the spindle association gradually diminished in L36 mutants(Fig. 6C) and was completely lost in L42 mutants (Fig. 6C)and in other mutants after linker 36 (data not shown). Theseresults suggest that the 279 amino acid peptide located betweenL1 and L24 is necessary for nuclear targeting, whereas thepeptide domain (residues 1516-1667) located between L42 andL36 appears to be necessary for spindle association.
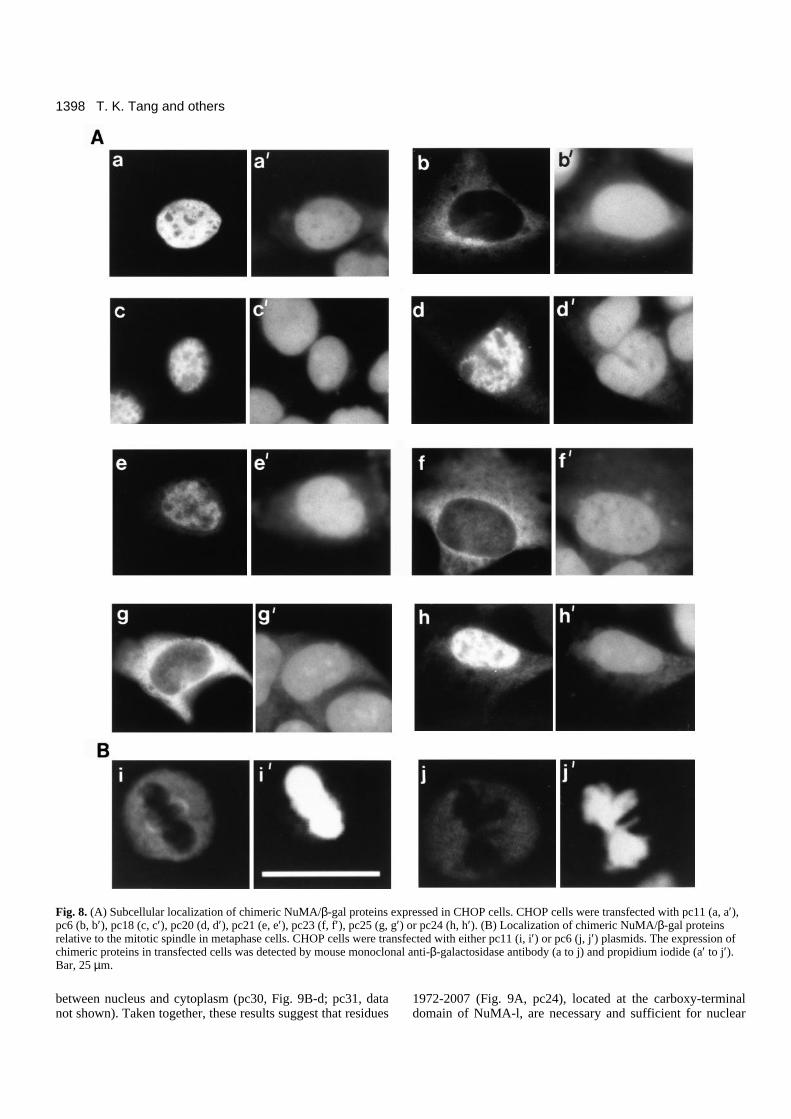
Subcellular localization of the carboxy terminus ofNuMA-l/β-galactosidase chimeric proteinsTo define more precisely the carboxy terminus of NuMA-linvolved in nuclear localization and spindle association, aseries of NuMA-l/β-galactosidase expression vectors wereconstructed (Fig. 7). β-Galactosidase (β-gal), a proteinencoded by the lacZ gene of E. coli was chosen as a cytosollocalization marker. The expression of chimeric proteins thatencode various carboxy-terminal proteins of NuMA-l fused toβ-gal in CHOP cells was analyzed with a monoclonal antibodyspecifically against β-gal. Consistent with earlier reports, wild-type β-gal was detected predominantly in the cytoplasm withno nuclear expression and no spindle association (data notshown). However, pc11 chimeric protein (NuMA1538-2115/β-gal), consisting of the entire carboxy-terminal regionof NuMA-l (Fig. 7), was expressed primarily in the nucleus butnot in the nucleoli (Fig. 8A-a), a pattern similar to thatobserved in NuMA of human cells (Tang et al., 1993b).Analysis of pc6 chimeric protein (NuMA1538-1975/β-gal)displayed a mostly cytoplasmic expression pattern (Fig. 8A-b).Interestingly, pc11 chimeric protein was targeted to the spindlepole in mitotic cells (Fig. 8B-i). However, spindle-pole asso-ciation was not observed in cells transfected with pc6 (Fig. 8B-j), pc20 or pc25 chimeric proteins (data not shown). Takentogether, these results suggest that residues between 1975 and2115 of NuMA-l contain a functional signal for nuclear local-ization, whereas the entire tail region (1538-2115) of NuMA-l is necessary and sufficient for spindle association (Fig. 7).
To narrow down further the region involved in nuclear local-ization, we then transfected several expression vectors (Fig. 7,pc18, 20, 21, 23, 24, and 25) that each encoded a smallerpeptide of the NuMA-l tail fused to β-gal in CHOP cells.Chimeric proteins consisting of residues 1839-2095 (Fig. 7,pc18; Fig. 8A-c), 1972-2095 (Fig. 7, pc20; Fig. 8A-d), or 1972-2059 (Fig. 7, pc21; Fig. 8A-e) of the NuMA-l tail revealed apredominantly nuclear pattern of localizaion. In contrast,chimeric proteins containing either residues 2030-2059 (Fig. 7,pc23; Fig. 8A-f) or 2002-2034 (Fig. 7, pc25; Fig. 8A-g) fusedto β-gal displayed a mostly cytoplasmic expression pattern.These results suggest that residues 1972-2007 of NuMA-l con-stitute a functional NLS. Indeed, when we transfected pc24,which contains only 36 amino acid residues (1972-2007) of the
NuMA-l tail fused to β-gal, into CHOP cells, we did observeprominent nuclear staining (Fig. 8A-h).
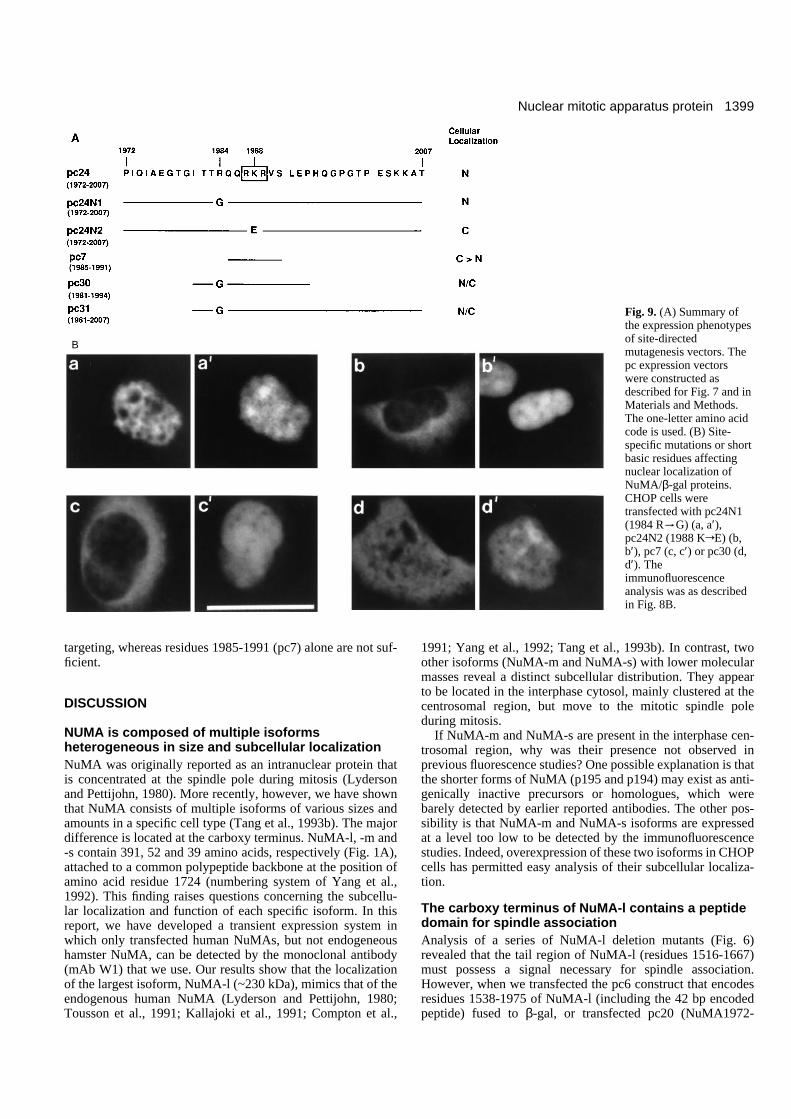
Site-specific mutational analysis of the nuclearlocalization signalThe 36 amino acid peptide (residues 1972-2007) of the NuMA-l protein contains four basic residues clustered within positions1984 to 1989 (Fig. 9A), a feature similar to other known NLS.One mutation with the (K)Lys1988 of NuMA-l substituted byglutamic acid (E) in pc24N2 clone (Fig. 9A) did abolishnuclear localization (Fig. 9B-b). However, another mutation(pc24N1), which has (R)Arg1984 replaced by glycine (G), didnot prevent accumulation of the chimeric protein in the nucleus(Fig. 9B-a). These results suggest that Lys1988 is essential fornuclear targeting, whereas Arg1984 is not. To our surprise, thepc7 chimeric protein (NuMA1985-1991/β-gal) consisting ofonly seven amino acid residues (QQRKRVS) that cover allthree basic residues (RKR), located at amino acid positions1987-1989 (Fig. 9A), was mostly expressed in the cytoplasm(Fig. 9B-c). However, when we transfected with two other con-structs (pc30 and pc31), which encode residues 1981-1994 or1981-2007 of NuMA-l, respectively, fused to β-gal, these twochimeric proteins were homogeneous and distributed equally
alpha-helical coiled-coil domain1 (25 aa) (14 aa)
SV40 ori/early promoter
gpt trps Lac Z
ATG
EcoRI
NuMA-l
P P
N
NuMA-l
C2115
CellularLocalization
SpindleAssociation
pc111538 2115
N
pc61538 1975
C
pc181839 2095
N
pc201972 2095
N
pc211972 2059
N
pc232030 2059
C
pc252002 2034
C
pc241972 2007
N
Fig. 7. Summary of the chimeric NuMA/β-gal expression vectorsand the expression phenotypes of these chimeric polypeptides. Thepc expression vectors were constructed by ligating a series of PCRfragments amplified from NuMA-l cDNA, into the lacZ gene ofpCH110 expression vector (see Materials and Methods for details).P, selected primer sets for PCR amplification. The SV40 earlypromoter of pCH110 directs the high-level expression of a hybridprotein composed of 40 residues of gpt including the ATGtranslation initiation site and 28 residues of trp S that were fused tothe lacZ gene-encoded β-gal protein (Pharmacia). The amino acidresidues of NuMA-l encoded by these chimeric expression vectorsare indicated by numbers below the black boxes, which represent thecoding region of NuMA-l protein. N, nuclear localization; C, cytosollocalization.
1398
between nucleus and cytoplasm (pc30, Fig. 9B-d; pc31, datanot shown). Taken together, these results suggest that residues
1972-2007 (Fig. 9A, pc24), located at the carboxy-terminaldomain of NuMA-l, are necessary and sufficient for nuclear
T. K. Tang and others
Fig. 8. (A) Subcellular localization of chimeric NuMA/β-gal proteins expressed in CHOP cells. CHOP cells were transfected with pc11 (a, a′),pc6 (b, b′), pc18 (c, c′), pc20 (d, d′), pc21 (e, e′), pc23 (f, f′), pc25 (g, g′) or pc24 (h, h′). (B) Localization of chimeric NuMA/β-gal proteinsrelative to the mitotic spindle in metaphase cells. CHOP cells were transfected with either pc11 (i, i′) or pc6 (j, j′) plasmids. The expression ofchimeric proteins in transfected cells was detected by mouse monoclonal anti-β-galactosidase antibody (a to j) and propidium iodide (a′ to j′).Bar, 25 µm.
1399Nuclear mitotic apparatus protein
targeting, whereas residues 1985-1991 (pc7) alone are not suf-ficient.
DISCUSSION
NUMA is composed of multiple isoformsheterogeneous in size and subcellular localizationNuMA was originally reported as an intranuclear protein thatis concentrated at the spindle pole during mitosis (Lydersonand Pettijohn, 1980). More recently, however, we have shownthat NuMA consists of multiple isoforms of various sizes andamounts in a specific cell type (Tang et al., 1993b). The majordifference is located at the carboxy terminus. NuMA-l, -m and-s contain 391, 52 and 39 amino acids, respectively (Fig. 1A),attached to a common polypeptide backbone at the position ofamino acid residue 1724 (numbering system of Yang et al.,1992). This finding raises questions concerning the subcellu-lar localization and function of each specific isoform. In thisreport, we have developed a transient expression system inwhich only transfected human NuMAs, but not endogeneoushamster NuMA, can be detected by the monoclonal antibody(mAb W1) that we use. Our results show that the localizationof the largest isoform, NuMA-l (~230 kDa), mimics that of theendogenous human NuMA (Lyderson and Pettijohn, 1980;Tousson et al., 1991; Kallajoki et al., 1991; Compton et al.,
1991; Yang et al., 1992; Tang et al., 1993b). In contrast, twoother isoforms (NuMA-m and NuMA-s) with lower molecularmasses reveal a distinct subcellular distribution. They appearto be located in the interphase cytosol, mainly clustered at thecentrosomal region, but move to the mitotic spindle poleduring mitosis.
If NuMA-m and NuMA-s are present in the interphase cen-trosomal region, why was their presence not observed inprevious fluorescence studies? One possible explanation is thatthe shorter forms of NuMA (p195 and p194) may exist as anti-genically inactive precursors or homologues, which werebarely detected by earlier reported antibodies. The other pos-sibility is that NuMA-m and NuMA-s isoforms are expressedat a level too low to be detected by the immunofluorescencestudies. Indeed, overexpression of these two isoforms in CHOPcells has permitted easy analysis of their subcellular localiza-tion.
The carboxy terminus of NuMA-l contains a peptidedomain for spindle associationAnalysis of a series of NuMA-l deletion mutants (Fig. 6)revealed that the tail region of NuMA-l (residues 1516-1667)must possess a signal necessary for spindle association.However, when we transfected the pc6 construct that encodesresidues 1538-1975 of NuMA-l (including the 42 bp encodedpeptide) fused to β-gal, or transfected pc20 (NuMA1972-
Fig. 9. (A) Summary ofthe expression phenotypesof site-directedmutagenesis vectors. Thepc expression vectorswere constructed asdescribed for Fig. 7 and inMaterials and Methods.The one-letter amino acidcode is used. (B) Site-specific mutations or shortbasic residues affectingnuclear localization ofNuMA/β-gal proteins.CHOP cells weretransfected with pc24N1(1984 R
tG) (a, a′),pc24N2 (1988 KrE) (b,b′), pc7 (c, c′) or pc30 (d,d′). Theimmunofluorescenceanalysis was as describedin Fig. 8B.
B

1400
2095/β-gal) into CHOP cells (Fig. 7), no spindle associationwas observed. From these results, it is reasonable to speculatethat the spindle-association signal is located between residues1516 and 1538. Unfortunately, this appears not to be the case,since Compton and Cleveland (1993) have shown that a taillesshuman NuMA protein (CMV/NuMA1-1545), consisting ofresidues from 1 to 1559 based on our and Yang’s numberingsystem (Yang et al., 1992), is not targetted to the spindle pole.(The Compton construct, CMV/NuMA1-1545, lacks the 42 bpencoded peptide, therefore residue 1545 is equivalent to 1559,based on our and Yang’s numbering system.) Their observa-tion excludes the possibility that residues 1516-1538 ofNuMA-l contain the signal for spindle association. To oursurprise, if we transfected the pc11 construct (Fig. 7) encodingthe entire tail region of NuMA-l (residues 1538-2115) fused toβ-gal, it did reveal a spindle association in mitotic cells (Fig.8B-i), although the intensity of the signal was weaker than thatobserved in wild-type SV/NuMA-l (Fig. 1C-b). Altogether,these results suggest that the pc11 chimeric protein contains apeptide domain that is necessary and sufficient for spindleassociation, whereas other sequences shorter than this regionare insufficient for spindle targeting.
It is interesting to note that both NuMA-m and -s also targetto the mitotic spindle. By comparison of the sequences of thespindle association domain (residues 1538-2115) defined inNuMA-l with the entire coding region of NuMA-m (residues1-1776) and -s (residues 1-1763), a short segment located atresidues 1538-1725 is identical in all three NuMA isoforms.However, the pc6 chimeric protein (NuMA1538-1975/β-gal)that includes this region is insufficient to drive the chimericprotein into the spindle (Fig. 7, pc6). These results suggest thata particular amino acid sequence may not be directly involvedin targeting of NuMA-m and -s to the mitotic spindle pole.Instead, the overall protein tertiary structure or post-transla-tional modification of some specific residues in NuMApolypeptides may play a role. In addition, the presence of anegative regulatory sequence located within residues 1538-1975, which inhibits the spindle association, cannot beexcluded.
Furthermore, we have shown that the truncated L36 (1-1667)and L24 (1-1766) polypeptides of NuMA-l isoform are presentat the spindle pole, but not at the interphase centrosomal region.In contrast, both NuMA-m and NuMA-s isoforms are localizedto the centrosomal region in interphase and mitotic cells. Thisobserved differential subcellular localization might be partiallydue to the presence of the unique polypeptide located at thecarboxy termini of NuMA-m and NuMA-s.
Possible roles for shorter NuMA isoforms (NuMA-mand NuMA-s) during mitosisThe targeting of NuMA-m and -s to the interphase centroso-mal region is rather unexpected, since NuMA (renamed asNuMA-l) was originally reported to be present in the inter-phase nuclei, not the centrosomal region. It is interesting tonote that all three isoforms contain a common long α-helicalcoiled-coil domain (Tang et al., 1993b) that has been predictedto serve a function in the dimerization of two NuMA moleculesthrough hydrophobic interactions. Therefore, it is possible thatpre-existing NuMA-m and -s at the interphase centrosomalregion might act as a seed for NuMA-l assembly at a latermitotic stage. In addition, we have shown that the NuMA-l tailpossesses the peptide signal necessary for spindle association.Therefore, NuMA-l might be first brought to the spindle poleregion by its spindle association domain and then become asso-ciated with two other NuMA isoforms (NuMA-m and -s)through a common long α-helical coiled-coil dimerizationdomain (Fig. 10). The observations that NuMA-l associateswith the centrosome immediately after it is released from thenucleus upon nuclear envelope breakdown (Lyderson andPettijohn, 1980; Tousson et al., 1991; Compton et al., 1992;Yang et al., 1992) and that all three isoforms are present in themitotic spindle at a later mitotic stage (this report) are consis-tent with this hypothesis. Furthermore, the primary sequenceof NuMA contains five hypothetical leucine zipper repeatslocated in both the N-terminal and the long α-helical centraldomains (Fig. 10; Tang et al., 1993b; Maekawa and Kuriyama,1993). Although the various NuMA isoforms might form ahomo- or hetero-dimer along its large central coiled-coildomain, other proteins with similar leucine zipper repeatsmight also interact with NuMAs through the leucine residuesof the zipper motif.
Recently, several centrosomal components have been iden-tified and classified into four categories on the basis of theirdistribution during the cell cycle (reviewed by Kalt andSchliwa, 1993). Among these proteins, p34cdc2 has drawnmuch attention, due to its partial association with the centro-some (Bailly et al., 1989) and its mitotic kinase activity(Draetta and Beach, 1988; Labbe et al., 1988). Interestingly,four cdc2 kinase recognition sequence motifs (TPRD, TPKK,SPRI and TPRA at positions 2015-2018, 2055-2058, 2087-2090 and 2106-2109) and one such motif (TPRD at position1732-1735) (Shenoy et al., 1989) have been identified at thecarboxy terminus of NuMA-l (Yang et al., 1992; Tang et al.,1993b) and NuMA-m (Tang et al., 1993b), respectively. Thesefindings, and the presence of all these isoforms at the inter-
T. K. Tang and others
1
(25 aa) (14 aa)
N C2115
"dimerization domain"
spindle association domain
nuclear localization signal
216 1700
1538 2115
1972 2007
L L L LL
globulardomain
globulardomain
alpha-helical coiled-coil domain
Fig. 10. Schematic representation of thestructural (upper) and functional (lower)domains of human NuMA-l. The wavy-line boxrepresents the α-helical coiled-coil domain. Thecross-hatched box (25aa and 14aa) representsan alternative-spliced sequence block. L,leucine zipper motif.

1401Nuclear mitotic apparatus protein
phase centrosomal region (cdc2 and NuMA-m) and the mitoticspindle pole region (cdc2, NuMA-l and NuMA-m) suggest thatsome NuMA isoforms could potentially be phosphorylated andregulated by p34cdc2 kinase. In addition, γ-tubulin has beenreported to be a centrosomal protein (Zheng et al., 1991;Stearns et al., 1991) required for cell-cycle-dependent micro-tubule nucleation (Joshi et al., 1992). The coexistence of theγ-tubulin with shorter NuMA isoforms (NuMA-m and -s) inthe interphase centrosomal region and the mitotic spindle polesuggests a possible connection between these proteins. Furtheranalysis of the protein-protein interactions among various cen-trosome-associated proteins may shed some light on theirfunctions.
NuMA-l tail contains a novel nuclear localizationsignalOur results show that residues 1972-2007 of NuMA-l (pc24)contain a signal that is sufficient for nuclear targeting. Whenwe analyzed this 36 amino acid peptide sequence using theGCG computer program MOTIFS described by Dr AmosBairoch (University of Geneva), no specific peptide motifswere identified. However, a short basic peptide sequence(RQQRKR, residues 1984-1989) containing a high proportionof positively charged amino acids (lysine and arginine) wasnoted within this region, which has been described to be oneof the important characters for NLS (reviewed by Garcia-Bustos et al., 1991; Whiteside and Goodbourn, 1993). Substi-tution of Lys1988 in this region (Fig. 9, pc24N2) with an acidicglutamic acid residue did abolish nuclear localization, whereasreplacement of Arg1984 (Fig. 9, pc24N1) with a non-basic,non-hydrophobic glycine residue did not. These results suggestthat three basic amino acids (RKR) located at residues 1987-1989 may be important for nuclear targeting. To our surprise,the pc7 (NuMA1985-1991/β-gal) chimeric protein, whichcomprises only seven residues (QQRKRVS) including RKR,was found to have a mostly cytosolic staining pattern. Theseresults suggest that Lys1988 is essential. However, the basicresidues (RKR) alone are not sufficient for targeting thechimeric protein into the nucleus.
Recently, it has been reported that the rate of nuclear translo-cation of SV40 large T antigen depends on the phosphoryla-tion of the serine residue located at the casein kinase II sitenear the NLS (Rihs et al., 1991). In yeast SW15 protein, phos-phorylation of the serine residue located at the potential cdc2kinase recognition sites within its NLS may also regulate theintracellular localization of SW15 (Moll et al., 1991).However, neither the potential cdc2 nor casein kinase II recog-nition sites are present within the 36 amino acid peptide inNuMA-l. Moreover, no sequence similar to the bipartitenuclear localization domain reported in nucleoplasmin(Robbins et al., 1991) could be identified within this region.Taken together, the above findings suggest that the 36 aminoacid peptide located at the middle portion of the carboxyterminus (1972-2007) of NuMA-l constitutes a novel func-tional NLS. This nuclear localization signal is distinguishedfrom most previously described NLSs by its large size and itsabsence of any known phosphorylation sites for cdc2 andcasein kinase II. In addition, its short basic peptide alone isinsufficient for nuclear targeting. Further analysis of nucleartargeting by this novel NLS will be needed to elucidate thegeneral mechanism of nuclear protein translocation.
In summary, we have shown that NuMA is composed ofmultiple isoforms heterogeneous in size and subcellular local-ization. In this report, we have also defined several functionaldomains of NuMA-l, and presented a structural and functionalmodel for human NuMA-l (Fig. 10), which may provide someimportant insights into our understanding of the function ofvarious NuMA isoforms.
This work was supported by program project grant NSC83-0203-B001-102 from the National Science Council of the Republic of Chinaand a research grant from the Institute of Biomedical Sciences,Academia Sinica. We are grateful to Dr John L. Wang for his thought-ful comments and suggestions. We thank Dr Cathy Fletcher forreading this manuscript.
REFERENCES
Ausubel, F. M., Brent, R., Kingston, R. E., Moore, D. D., Seidman, J. G.,Smith, J. A. and Struhl, K. (1989). In Current Protocols in MolecularBiology. John Wiley and Sons, Inc. New York.
Bailly, E., Doree, M., Nurse, P. and Bornens, M. (1989). p34cdc2 is located inboth nucleus and cytoplasm; part is centrosomally associated at G2/M andenters vesicles at anaphase. EMBO J. 8, 3985-3995.
Compton, D. A., Yen, T. J. and Cleveland, D. W. (1991). Identification ofnovel centromere/kinetochore-associated proteins using monoclonalantibodies generated against human mitotic chromosome scaffolds. J. CellBiol. 112, 1083-1097.
Compton, D. A., Szilak, I. and Cleveland, D. W. (1992). Primary structure ofNuMA, an intracellular protein that defines a novel pathway for segregationof proteins at mitosis. J. Cell Biol. 116, 1395-1408.
Compton, D. A. and Cleveland, D. W. (1993). NuMA is required for theproper completion of mitosis. J. Cell Biol. 120, 947-957.
Draetta, G. and Beach, D. (1988). Activation of cdc2 protein kinase duringmitosis in human cells: Cell cycle-dependent phosphorylation and subunitrearrangement. Cell 54, 17-26.
Garcia-Bustos, J., Heitman, J. and Hall, M. N. (1991). Nuclear proteinlocalization. Biochim. Biophy. Acta 1071, 83-101.
Heffernan, M. and Dennis, J. W. (1991). Polyoma and hamster papovaviruslarge T antigen-mediated replication of expression shuttle vectors in Chinesehamster ovary cells. Nucl. Acids Res. 19, 85-92.
Joshi, H. C., Palacios, M. J., McNamara, L. and Cleveland, D. W. (1992). γ-Tubulin is a centrosomal protien required for cell cycle-dependentmicrotubule nucleation. Nature 356, 80-83.
Kallajoki, M., Weber, K. and Osborn, M. (1991). A 210 kD nuclear matrixprotein is a functional part of the mitotic spindle; a microinjection studyusing SPN monoclonal antibodies. EMBO J. 10, 3351-3362.
Kallajoki, M., Harborth, j., Weber, K. and Osborn, M. (1993).Microinjection of a monoclonal antibody against SPN antigen, nowidentified by peptide sequences as the NuMA protein, induces micronuclei inPtK2 cells. J. Cell Sci. 104, 139-150.
Kalt, A. and Schliwa, M. (1993). Molecular components of the centrome.Trends Cell Biol. 3, 118-128.
Labbe, J. C., Lee, M. G., Nurse, P., Picard, A. and Doree, M. (1988).Activation at M-phase of a protein kinase encoded by a starfish homologue ofthe cell cycle control gene cdc2+. Nature 335, 251-254.
Lyderson, B. and Pettijohn, D. (1980). Human-specific nuclear protein thatassociates with the polar region of the mitotic apparatus: distribution in ahuman/hamster hybrid cell. Cell 22, 489-499.
Maekawa, T. and Kuriyama, R. (1991). Differential pathways of recruitmentfor centrosomal antigens to the mitotic poles during bipolar spindleformation. J. Cell Sci. 100, 533-540.
Maekawa, T. and Kuriyama, R. (1993). Primary structure and microtubule-interacting domain of the SP-H antigen: a mitotic map located at the spindlepole and characterized as a homologous protein to NuMA. J. Cell Sci. 105,589-600.
Moll, T., Tebb, G., Surana, U., Robitsch, H. and Nasmyth, K. (1991). Therole of phosphorylation and the CDC28 protein kinase in cell cycle-regulatednuclear import of the S. cerevisiae transcription factor SE15. Cell, 66, 743-758.
Price, C. M. and Pettijohn, D. E. (1986). Distribution of the nuclear mitotic

1402
apparatus protein (NuMA) during mitosis and nuclear assembly. Exp. CellRes. 166, 295-311.
Rihs, H-P., Jans, D. A., Fan, H. and Peters, R. (1991). The rate of nuclearcytoplasmic protein transport is determined by the casein kinase II siteflanking the nuclear localization sequence of the SV40 T-antigen. EMBO J.10, 633-639.
Robbins, J., Dilworth, S. M., Laskey, R. A. and Dingwall, C. (1991). Twointerdependent basic domains in nucleoplasmin nuclear targeting sequence:Identification of a class of bipartite nuclear targeting sequence. Cell 64, 615-623.
Sambrook, J., Fritsch, E. F. and Maniatis, T. (1989). Molecular Cloning.Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York.
Shenoy, S., Bagrodia, S., Choi, J.-K., Copeland, T., Maller, J., andShalloway, D. (1989). Purified maturation promoting factor phosphorylatespp60 c-src at the sites phosphorylated during frbroblast mitosis. Cell 57, 763-774.
Stearns, T., Evans, L. and Kirschner, M. (1991). γ-Tubulin is a highlyconserved component of the centrosome. Cell 65, 825-836.
Tang, T. K., Hong, T.-M., Lin, C.-Y., Lai, M.-L., Liu, C. H. L., Lo, H.-J.,Wang, M.-E., Chen, L. B., Chen, W.-T., Ip, W., Lin, D. C., Lin, J. J.-C.,Lin, S., Sun, T.-T., Wang, E., Wang, J. L., Wu, R., Wu, C.-W. and Chien,S. (1993a). Nuclear protein of the bovine esophageal epithelium. I.Monoclonal antibody W2 specifically reacts with condensed nuclei ofdifferentiated superficial cells. J. Cell Sci. 104, 237-247.
Tang, T. K., Tang, C.-J. C., Chen, Y.-L. and Wu, C.-W. (1993b). Nuclear
proteins of the bovine esophageal epithelium. II. The NuMA gene gives riseto multiple mRNAs and gene products reactive with monoclonal antibodyW1. J. Cell Sci. 104, 249-260.
Tang, T. K., Tang, C.-J. C., Tsou, T.-C., Hong, T.-M., Hong, R.-L., Chang,C.-J., and Wu, C.-W. (1994). Study of nuclear proteins associated with cellproliferation, differentiation, and mitosis by confocal fluorescencemicroscopy. In Multidimensional Microscopy (ed. P. C. Cheng, T. H. Lin, W.L. Wu, and J. L. Wu), pp. 305-319. Springer-Verlag Inc., New York.
Tousson, A., Zeng, C., Brinkley, B. R. and Valdiria, M. M. (1991).Centrophilin: a novel mitotic spindle protein involved in microtubulenucleation. J. Cell Biol. 112, 427-440.
Whiteside, S. T. and Goodbourn, S. (1993). Signal transduction and nucleartargeting: regulation of transcription factor activity by subcellularlocalisation. J. Cell Sci. 104, 949-955.
Yang, C. H., Lambie, E. J. and Snyder, M. (1992). NuMA: an unusually longcoiled-coil related protein in the mammalian nucleus. J. Cell Biol. 116, 1303-1317.
Yang, C. H. and Snyder, M. (1992). The nuclear-mitotic apparatus protein isimportant in the establishment and maintenance of bipolar mitotic spindleapparatus. Mol. Biol. Cell 3, 1259-1267.
Zheng, Y., Jung, M. K. and Oakley, B. R. (1991). γ-Tubulin is present inDrosophila melanogaster and homo sapiens and is associated with thecentrosome. Cell 65, 817-823.
(Received 6 December 1993 - Accepted 2 February 1994)
T. K. Tang and others