Embed Size (px)
Citation preview

THE JOURNAL OF COMPAEATIVE NEUROLOGY 3703424.49 (1996)
J
Motor Nuclei of Nerves Innervating the Tongue and Hypoglossal Musculature in a Caecilian (Amphibia: Gymnophiona), as
Revealed by HRP Transport
ANDREA SCHMIDT, DAVID E. WAKE, AND MARVALEE H. WAKE Brain Research Institute, University of Bremen, 28334 Bremen, Germany (A.S.); and Department of Integrative Biology and Museum of Vertebrate Zoology, University of
California, Berkeley, California 94720-3140 (D.B.W., M.H.W.)
ABSTRACT The organization of the motor nuclei of the glosmpharyngeal, vagal, wcipitd, first spinal
and second spinal nerves of Typhlomctm natana (Amphibia: Gpnophiona: CaeuhaicIae Typhlonectinae) w a ~ skudied by using homeradmh peroxidase reaction staining. Each nucleus has M t e patterns of cytoarchitecture and of topography. Nuclei are elongate and aome overlap antempsteriorly. The brainstem is elongate, with no distinct demarcation of brain- stem from spinal cord. The occipital nerve emerges through a %parah foramen from that for the vagus and glolsaopharpgeal nemea in the species studied, is distinct from h t h , and ita nucleus i~ more M a r to spinal nuclei in cytaarchitecture. The occipital nerve h with spinal nerves 1 and 2 to contributa to the hypoglosd trunk. A spinal amassory newe i absent. 0 1996 wiley-~is~ Inc.
Indexing tmtm brainstem, ramus hyp~loasus, occipital n w e
The neuroanatomy of the gymnophione amphibians (cae- cilians) has received little attention, especially in corn@- eon to the extensive work published on damandera and fro@. Wddschmidt (1887) briefly d d b e d the anatomy of the brain and cranial nerves in four species of caecilians; Burckhardt ( 189 1) compared the brains of the salamander Ti.itum~ and the caecilian Ickihyophis. Norris and Hughes (1918) examined the cranial and anterior spinal nerves of several species of caecilians; Kuhlenbeck (1822) desmibed the brain, concentrating on Siphnops annul&-. hub- mann (1927) examined morpho*rmis of the brain of Hypogmphis, and Krabbe (1962) compared groaa brain development in three species of salamanders and three of a, each in Werent familia. While none of these authors considered d d s of brain organization in caecil- ians, their studies are the point of departure for OUT work.
Recent studies on the motor nuclei of frogs (Szekely, 1976 Mates2 and Szekely, 1977,1978; $tuesse et al., 1983, 1984; Nikundiwe et al.. 1982; Nikundiwe and Nieuwen- huys, 1983; Oka et al., 1987) and dmnanders IRoth and Wake, 1985; Wake et al., 1988), and b o t h (Szekely and Matesz, 19931, rwealed differences in the organization of motor nuclei (number of motor columns, segregation of motor nuclei) between members of the two orders. Frogs Mer from aalarnanders in having a well-expressed topo- graphic mgre@ion of motor nuclei in the anterior medulla
oblongata. Motor nuclei of wind m e are canstitukd of medial and lataal motor columns. In aalammhrs, there is extensive overlap of adjacent motor nuclei. Spinal nuclei of bobglossine salamanders are constituted of only a medid motor m h m , (Roth and Wake, 1985; h t h et d., 1988; Wake et al., 1988). caecilians ala0 have a more reduoed brain morphology than do from. However, studies on the tectum mewncephali in caecilians (Schmidt and Wake, 1991) suggest that the^ animals represent an “intermedi- ate” degree of morphological complexity, between that of frogs and salamanders. Modes of feeding and lmmotion in caecilians are markedly different from those of damandem and frogs. However, to date, nothing is known about the organization of the caecilian brainatem and the cervical spinal card and its relation t~ feeding and locomotion in caecilians. Therefore, on strict functionalist p u n & , differ- en- in the neuroanatomy of medullary control regions are expected-
The aim of this study is to examine the topolagy and cytoarchitecture of motm nuclei in one species of caecilian, Typhlonates mans , and to make a general comparison
Amaptad February 23,1998 Address reprint requesb to M a m a l e H. Wake, Department of Integrah
Biolw, 9060 YLSB, University of CaliforOia, Berhley, CA 947209140. E-mail: [email protected]?y.edu
0 1996 WILEY-LIS$, INC.
MOTOR NUCLEI OF TONGUE NERVES IN CAECILIANS 343
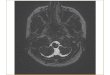
Fig. 1. ~ ~ e w o f t h e b r a i n o f ~ h k " m t e s nutans, illustrating the conformation of the m o r cranial and anterior spinal IWNH (A), and the extent and w e n t of the motm nuclei ofth- nerves in the brain- &m @). E, gImophaymgd nerve; X, va- gus a m ; OCC, oocipid nerve; SPl, fid spinal nerve; SP2, -nd spinal nerve. Scale bar=2mm. B
SP 2
llxl
occ I SPI 1
I SP2
with the condition in salamandem and frogs to determine whether differences in function are correlated with diffm- enoes in neuroanatomical organization. This study estab- M e s a baaeline for further comparative work on the neuroanatomy of caecilians, and elucidates several new questions that we expect to consider through experiments employing a wider range of techniques.
branch-. HRP-reacted brains were also compared to seri- ally sectioned, Palmgren's s t a i d heads in order ta study peripheral nerve patterne and central nervouB system ICNS)tracts.
RESULTS External morphology
MATERIALS AND METHODS The caecilian brainstem cantrash markedv with that of
the occipital nerve, and spinal nmws 1 and 2 by the h0- * peroxidase (HRP) method. Five animals wme prepared as wholemount, cleared and Sudan black 3- stained specimens (according to modihtions of the tech- niques~ of Filipslri and Wileon, 1984 and Nishikawa, 1987; see Wake, 1992).
Under triwjne methane sulfonate (MS 222) anamthesia (1:lOO wlv solution), v a r i m combinationa of the glo;s;sopha- ryngeus (E), vagus Or), occipitalis, spinal 1, and spinal 2 were MVWHI on the left and right sides of the animals . Nerves were identified aoeording to Wake (1992); opera- tional definitions of the occipital and the spinal accessow nemea are provided there and in the Dimssion. HRP tracing and d o n a were performd according to Fritzach et al. (1984). B r a h were then dehydrated in a wries of4 ethanols (76100%) for 30 minutes, then cleared and stored in 100% methyl salicylate. In order to make serial recon- structions of the motor nuclei and neurons, wholemounts were returned to alcohol, embedded in Epon, and cut at 80 pm. Brains so prepared were compared with five specimens macerated in trypsin, bulk-stained with Sudan black B and dizarin red S, cleared in glycerine, and stored in 100% glycerine to which thymol crystals were added to prwent fungal growth (e Wake, 1992, for protocol and compara- tive material). Sudan black ?3 sta in ing of intact animals permits examination of peripheral nerves and their
Motor pathways and nuclei There are two distinct but very
small mtlets that orient rather sharply postmiorly when they entar the brainstem. The motor rootlets of the g l m - phaq-ngeus are well-separated from and cranial ta thaw of the vagus. The mator nucleus of the gIossophaqmgeus is well-separated from the motor nucleus of the vagus (Fig. 1). It is small and compact (Figs. 1, 3B1, and lies entirely in h n t of the obex. Motor neumna of the gl-pharynes are found in the periventricular zone. Vagus (XI. The vagus enters the brain by way of three
motor branches, each composed of two rootlets. The motor nucleus ofthe vagus is relatively large (Figs. 1,2,4A,B) and extends ventrolaterally from a point between the entrances of the glossophargngeus and the vagus in front of the obex posteriorly to a point between the entrances of the &cipihl nerve and the first spinal nerve, well behind the obex (Fig. 4). Cella lying in the dorsal part of the ventrolateral mlumn of motor neurons are characterized by horizontal dendritic arborizations that extend laterally and anhiorly, and canstitUte a lateral neuropil (Fig . 0).
Occipitalis. The oocipital nerve is exclusively m o b and lacks a doreal root. It enters the brain via two branches between the vagus and spinal 1, somewhat nearer the vagus. The antmior branch consists of two rootlets and the posterior branch of four rootlets. The motor nucleus ex-
Gin8mphmpnpeus {ZX).

344 A. SCHMIDT ET AL.
A
B
entrances of the first and second spinal nerves. Dendritic arborizations of motor neurons extend do& and consti- tute a neuropil lateral to the sensory projection of spinal 2 (Fig. 4E).
DISCUSSION Organization of amphibian motor nuclei
There are differences in the organization of motm nuclei among m e m h s of the three orders of amphibians regard- ing the organization of motor columns, cytoarchitecture, and the pattern of segregation of motor nuclei ( M a t a z and Szekdy, 1978; StueBse et al., 1983,1984; Nikundiwe et al., 1982; Nikundiwe and Nieuwenhuys, 1983; O b et al., 1987; Roth and Wake, 1985; Roth et al., 1988; Wake et al., 1988; Szbkely and Matesz, 19993; Wake; 1993). The motor nuclei of from are mow widely separated than are those in damandem. The organization of the braimtem in &a- m a n d ~ L c ~ ~ ~ x e d b y ~ i v e o ~ ~ ~ i n g o f m o t o r nuclei. In caecilians, the internal o m a t i o n ofthe motor nuclei + , is mom similar that of fro@, but fie p H = of mk' more do- resembles that of salamanders (compare Fig. 1 with Fig. 6 in Rath et al., 1990).
The brainstem of l'yphlowtes is similar to that of frog, and a clear ~~
P f l ~ o r ~ o f t ~ m ~ ~ ~ ~ ~ ~ b y ~ o ~ b ping of motor nuclei, as oxum in damandem. We found extensive overlap among the motor nuclei of the occipitalis, the vagus, and the rostral half of the first spinal n e m ~ . Overlap of the nuclei of the vagus and spinal 1 also has been described in h a n d e r e (Wake et al., 1988). Neither the occipital motor nucleus nor its nerve occur in frogs or salamanders, or in amniote.
An apparent condensation of sphd 1, sphd 2, and the h y p d m d ~ e m B has omurred in adult anurans. spinal 1, with dorsal and ventral roots and a ganglion, is present in larval frosg (Gaupp, 1899). A oomplex of nerves emerges from the first vertebra in adult fro@, including thome that form the hypoglod ramus (Stuease et al., 1983). However, there is no ganglion, and no clear dorsal root of spinal 1. The rmtral parts of the two dist inct motor columns associ- ated with these nerves overlap the posterior half of the vagal nucleus inBufa (Oh et d., 1987).
Glossopharyngeal and vagd m o w nuclei form a continu- OUB (salamanders: Roth et al., 19881, partially diacontinu- o w (milians), or discontinuous (frogs: Oka et al., 1987) single column of cell b d m , lying at the wntrolated margin of the periventrzcular gray matter. The spinal motor nuclei include two wlumns, one medial and the other ventrohteral, in damandem (except for the derived, impli- fied condition in bolitoglossines; Wake et d., 1988). The occipital motor nucleus in 2'yppAZonec~s is more similar tu the more posterior spinal nuclei than to the more anterior glossopharyngeal and vagal nuclei, in that both medial and ventrolateral motor neurona are preaent. However, either our staining of the medial column is incomplete, or this column is constituted by very few and very small cells in
Fetcho (1986,1987) suggested distinct functions for the spinal motor columns in amniotes. The medial oolumn, which misw early in ontogeny, was thought to innervate axial musculature; the lateral column was considered re-
tSW ~ JgP' R. hypo
Fig. 2. Schematic illustration of the &letal elementi (A) and & P h r d nerve mota (B) h a b d with Posterior Cranial and anterior i d nerves. A is a draHTing ofthe rear o f h e skull, the atlas, and thm f h t spinal (second) vertebra, not drawn to d e , but to acceat the foramina through which the neryei exit. C m d n e w M and X &t together through the lars foramen in the skull, the & p i d through the smaller posterior foramen. spinal 1 &ta through the foramen of the a k the domal and ventral mota of Spinal 2 exit m, the d o 4 through the small anterior foramen, the ventral through the larger, more ventral foramen. B diagrams the n e w and
abbrwiationa ~ d i in Fig. 1). C l o d dark circles indicate do& root piglia. open circles indicate emit foramina. Ventral oomponenta of spinal ne- 1 and 2, and the MW ventra^ d N d nerve, the mu* b w $ b W . A and are wea to extents Of
a W e l w n e n b and n- rmt wnliguratione relative to each other.
formation of different motor
t h t of b a n d s r s , in their -ions to form the h g h u s (E. hyp~.; other tion ofthe glO~Pharyngeal a d Vagalnuclei. However? the
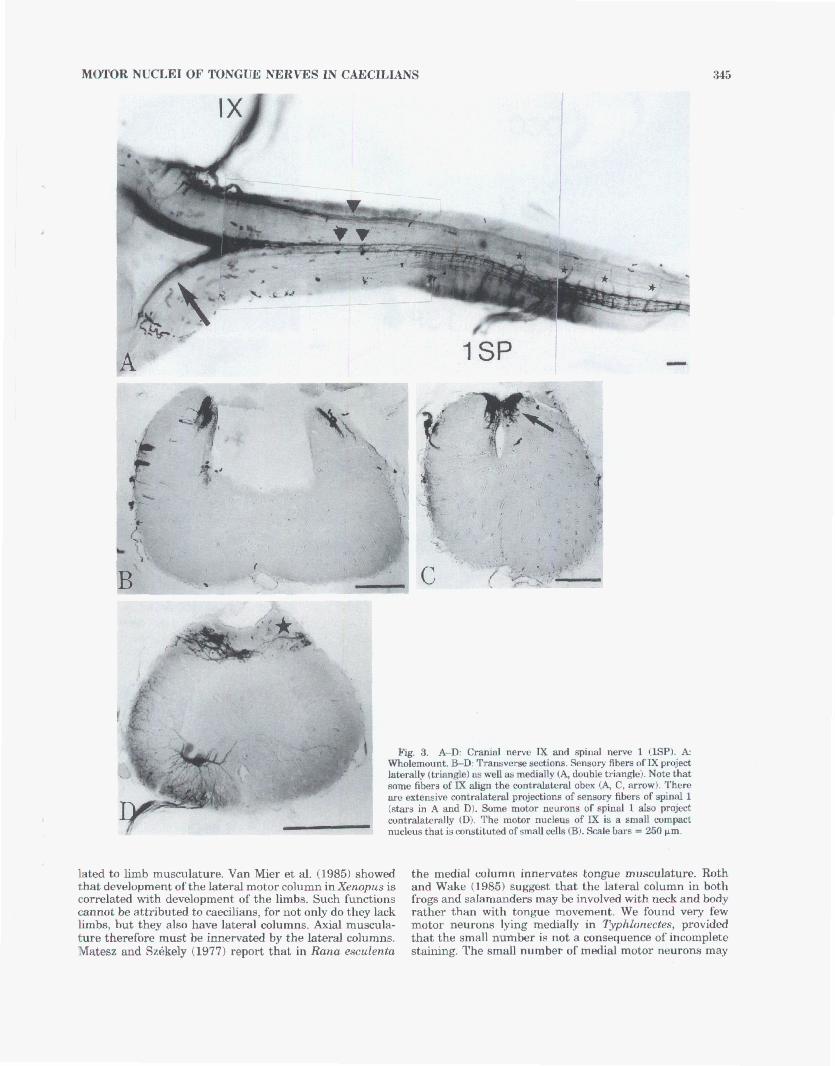
tends forward ta the level between the entrances of the g l ~ ~ ~ ~ p h a r y n g e a l and vagal nerves and posterior to a level btween the entranw of the axipital and first spinal newes (F&. 2,4A,B). There is strong overlap with that of the vagus (Figs. 2,4A). Moat motor neurons of the occipita- lis are found in the ventral p& of the laterd medulla. Dendritic arborizations of these o e h project dorsad and ventrad and appear to contact the lateral neuropil cowti- tuted by neurons of the WE. A few cells lie medially and form a separah column, continuous with that ofthe medial column of more posterior nuclei. These cells are small and weakly stained; we suspect that our staining of these cells may be incomplete. The dendntic arborizations of these cells extend laterad (Fig. 4D).
Spinal 1. The fmt spinal nerve has distinct, large do& and ventral roots, and a large dorsal root gangIion. The dorsal root is widely separated from the ventral root, and is poatdor to it. The motor nucleua of the first spinal nerve lies ventrolaterally. A few small par-shaped cella are found medially, fming a medial column. The ventrolateral col- umn e n d s anteriorly to overlap the pohrior portion of the nuclei of the vagus and occipitalis (Figs. 2, a), and posteriorly to a point anterior to the entrance of the second spinalneme. Ventrolahd motor nemm arborizedomlat- erally. There are few large multipolar d a ; they extend dendrites in all directions. Typhlonectes.
Spinal 2. Motor branches enter the cord via two ante- nor rootlets. A few stained motor murons lie anterior to the entrance of the motor branches (Fig. 4E). The nucleus extenda anteriorly to the level of the midpint between the
)-
I
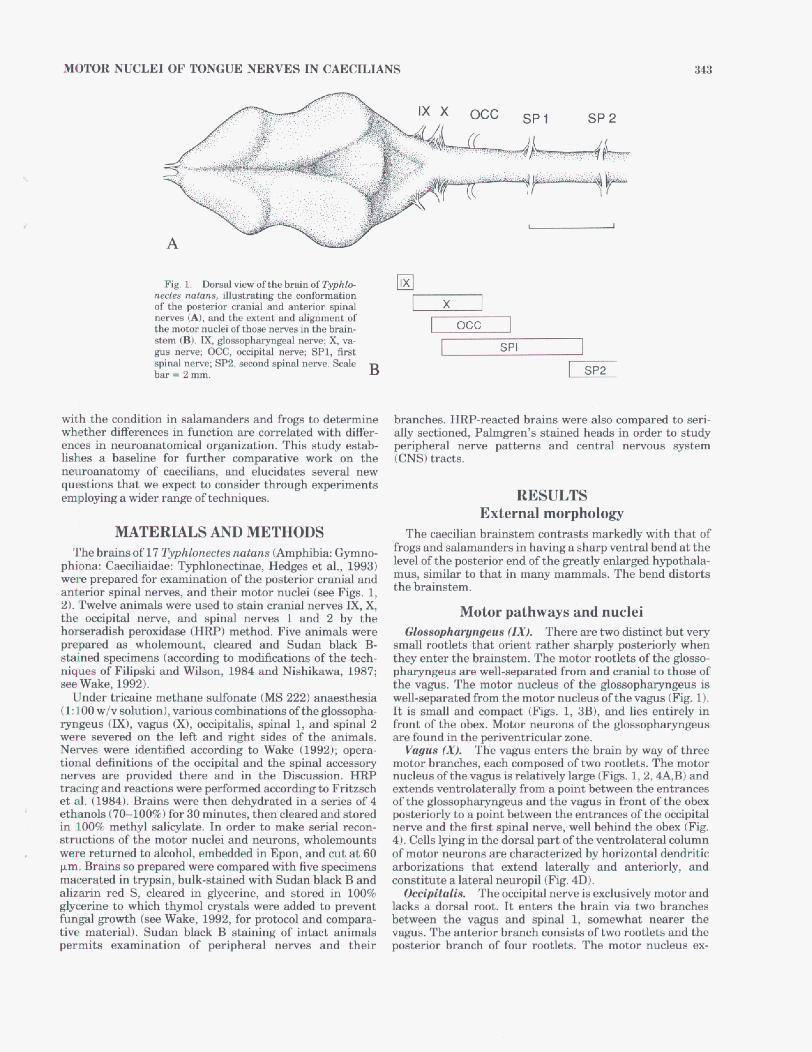
McpToR NUCLEI OF TONGUE NERVES IN CAECILIANS 345
lated to limb musculature. Van Mier et al. (1985) showed that development of the lateral motor column inXerwpw is correlated with development of the limbs. Such functions cannot be attributed to caecilians, for not only do they lack limbs, but they also have lateral columne. Axid muscula- ture therefore muat be innervated by the lateral columns. M a h z and Szbkely (1977) report that in Ram esculenta
the medial oolumn innervates tongue musculature. Roth and Wake (1985) suggest that the lateral column in both frogs and salamanders may be involved with neck and body rather than with tongue movement. We found very few motor neurons lying m d d y in Typhlomectes, provided that the smal l number is not a oomequence of incomplete staining. The s d number of medial motor neurons may
A c
P
A- E
-=F- I 1 SP
Fig, 4. A-E: Cranial nerve X Q), mpital (OCC), qind 1 and spinal 2 (lSP, 2SP). I/B,C,E Wholemwnts. D Tranaverae section of the wholemount in A at the level of X and &pital. The anterior part of the nucleus ofspinal 1 and the posterior pwb ofthe X and mipitd nuclei -lap (B). In D, motm neurons are found in three @tione: medial Mar), venbolabral Itmangle), and in the dorsal part of the ventral horn
(arrow). We consider the latter cella part of the Xth n-8, and the ventrolateral and medial motor neurom apeoccipihl. Motor neurons of s p i d 1 and of spinal 2 project dmuohkdy (thin -8). S e n s ~ y projectionnl of spinal 1 extend forward, beyond the obem (thick arm^). S c a l e h = 250 pm.

MOTOR NUCLEI OF TONGUE NERVES IN CAECILIANS 347
be related to a reduction of tongue cornonenti in these animals (Wake, 1992).
Occipital and/or epinal accessory nerves The presence of an exclusively motor cwipital nerve and
its large nucleuus in mme species of caecilians long has been known (Marcus, 1910; Norris and Hughes, 1918). Wake (1992) reviewed the lihature on occipital nerves, and concluded that the nerve in caecilians is appropriately 80 d&gnated for w r a l reasons (see below). The wcipitalis EPparently L involved ody in the ConBtitution of the ramus hypoglossus. After leaving the brain and proceeding drs- tally, either through a common foramen with the glowpha- ryngeus and vagus (several other taxa; see Wake, 19921, or though its own cranial foramen (BB in the s w reported herein), the occipitalis fuses with spinal 1, which then joins s~nal2 (Norris and Hughes, 1418; Wake, 1992). Wedo not yet hem anterograde Staining that would identify the innervation targets of the specific mmponents of the ramus hypogloBsm.
We do not find a spinal accessory nerve in Typhlonectes, mr in other species prepared for study Wake, unpublished data), nor has one h e n reported in the literature for caecilians. Some have questioned the identification of the mcipitalis as such (permnal communication to M. Wake), rather than as B spinal BCO~BSOIY, since the latter now has been found in aalamanders (Roth et al., 1984; Roth and Wake, 1986, Wake et al., 1988) and frogs (Mataz and Bbkely, 1977; O h et al., 1987; S&kely and Matesz, 1993, as well a8 many 0 t h vertebrate *. Location of its motor neurons auggwts that the caecilian oocipitalis might re- semble a spinal nerve; however, a~ Wdce (1992, 1993a,c) discussed, the dp i ta l i s of caecilians is not homologous to the spinal accessmy nerve of salamanders. Therefore, caecil- ians are dihnguished from their sister taxa by the presence of an occipital nerve in derived species, and by the apparent absence of a spinal accessom nerve. Consequently, the general conclusion that a spinal m r y nem is present in all amphibians (Szkkely andMatesz, 1993) does not apply to caediam, b a e d on current information.
The constitution of spinal nerve 1 in amphibians
The first spinal nerve of adult salamanders typically is camposed of several ventral rootlets, and lacks a dorsal root and ganglion. When the latter are p-nt in embryos, they are lost in adults (Roth and Wake, 1986). Spinal 1 is strictly motor and innervates tongue musEulature via the ramus hypoglossus. The second spinal nerve has d o d and ven- tral roots, and a dorsal ga@lion; it inmwates varioue throat, tongue, and neck mu&q 8s well as contributing a branch to the brachial plexus. In fmga, spinal nerve 1 is present in tadpoles but b p p a m after metrunorphmis (Ram: Gaupp, 18991, and the contribution to the ramus hypgloasuus emerges from the ventral ramuB of the second e p i d nerve (Stuesse et al., 1983j. The second spinal nerve also has a domal ramus and dorsal root ganghon. A third s t a t e of the ankrirmmt s p i d nerve ocrmrs in caeciliancl. The fmt spinal nerve not only is present, but is composed of dorsal and ventral roots and a dorsal root ganglion. The main branch of the ventral root wntributes to the m u s hypoglossus, together with a branch of the second apinal, d in some species the occipitalis, and/or B branch of the vagus, andlor a branch of qinal3 (Wake, 1992). Retention of the presumed ancestral stab of the fmt spinal nerve is
unexpeckl, given the profound emndary modification from a limbed to B limbless predator, and the extensive asmchtion of antmior spinal nervm With the feeding mechanism.
Caecilian VB. anuran and urodelan brainstem gtructure
Ph&wnetic hiatorg. All living members of the Gymno- phiona lack limbs, girdles, and the associated musculature and innervation, including the absence of brachial and spinal plexuses. Conoomitant to limb loss, the b d y of medians has h o m e very elongate (90-285 vertebrae; Taylor, 1968), though the tail is lost, or nearly go, in all taxa. Limb loss and its comelab present a pattern of morphology distinctly different from that of salamanders, even the elongate taxa, and of frogs. No rnembsrs of the Anum or the Caudata have lost all of their limbs, though they may be reduced (and hindlimbs lost) in salamanders; dl retain the appropriate musculature and its innemtion. Limb loss and secondary ~mplifimtion of the tongue have been conjectured by Wake (1992) to provide conditionrs that permit morphological innovation, such as incorporation of additional nerves (Oocipihlis, rami of the vagus and of spinal 3) into the ramus hypoglossus.
The brainstem of salamanders and frogs is not limited by the end of the skull, but is confluent with the anterior spinal cord (summarized by Ebth et al., 1990). A similar situation exists in caecilians, in which the brainstem ex- tends well posterior to the foramen magnum. In fact, the caecilian brainatem is mare elongate than that of elongate damandem We rspeculate that the combination of reduc- tion of morphological structure and oyerall body elongation has permitted the elongation of the brainstem as well. We cannot explain the p m n m of the occipital nerve, since it may be either a retention of the ancestral condition, or a reacquidtion. However, the constitution of spinal nerve 1 by dorsal and ventral roots and a dorsal root ganghon is apparently a retention of the ancestral vertebrate condi- tion, found in all c a d i a m examined, but lost in &- mandera and frogs. It is po&bIe that reduction of both limb and tongue components otherwise innervated by spinal nerve 1 might “permit” retention of the ancestral condition without specialization.
Ontogenetic his-. Little is known about d a n devdopment, e-y that of the brain, The length of the developmental period is known for only a few species. However, all indications are that time to metamorphosis or birth is pmtrackd in c~ecilians, for the free-living larval period is approximately one year (lehthyophls glutinosus: Breckenria and Jayashghe, 1979; Breckenridge et al., 1987; Philippine Ichthyophis spp.: Taylor, 1960). The gesta- tion p a i d in viviparous spck is also long in the only taxa for which there are data, 11 months in Demophis mexicu- nus and Gymnopis rnultipZica.ta, and 7-9 months in m h l o - nectes compressicuudu (summarized in Wake, 1993b).
helopment in CBeCiliana is highly oepkalized, with head morphology much a d v a n d over that of the posterior part of the body early in gestation (see Wake and Hanken, 1982). Comparative ontogenetic studies of amphibians suggest that heterochmnic prooesws during development have a major hfluence on the degree of lamination, formation of nuclei, and number of migrated cells within the brain (Schmidt and Wake, 1991; Roth et al., 1993; Schmidt and Roth, 1993). During brain development, the lateral motor column is constituted by d s that migrate from the

348 k SCHMIDT ET AL.
periventridar ependymal layer to the peripheral neuropil. Studies of fmgs Wan Mier et ai., 1985) and salamanders (Nishikawa et al., 1991) show that this occurs after the medial motor column has been constituted. The abaence of the lateral motor column in hlitaglossine damanders (Wake et al., 19881 is thought to be a eonsequence of paedomorphosis, in that later ontogenetic events (Le., the migration of cells into the periphery to form the lateral column) do not occur. Paedomorphosis hrls effected what Roth et al. (1993) d a ‘‘gecOndary aimplification” of brain morphology. The brain of caecilians is b r a c h i z e d by a similar simplifiation. There is little or no lamination, and there are only a few migrakd cell6 in the superficial neuropil in moat brain regions (Schmidt and Wake, 1991). However, in general, medians have more migrated 4 1 s within the brain than do salamanders. Typhlorsectes has neurons that form a lateral motor column. Cornparatiw ontagemtic studies on cellular migration within the tectum mesencephali of all three amphibian orders (Schmidt and Rot4 1993; Schmidt and Wake, unpubbhed data) indicate that in mdiam, late onhgenetic pmcessss are not re- tarded to the degree that tbey m? in salamanders. The complexity of the brainstem in m e d i a n s , more d a r to that of frogs than ta that of salamandem, may be a comequence of the early development of the medulla oblongata, aince retardation largely af€& later-developing brain regiom.
Brainstem or@xation in caedham therefore differs from that of their aster taxa, salamanders and fregs, as a consequence of (1) a combination of phylogenetic con- straints, perhaps ass&t.d with limb loss and simplifica- tion of the tongue, that are espcdly reflected in ontogeny, and (2) particular featursrs of neural ontogeny, such as pattern of cell migration and motor column eskblhhment. This study p-te baseline data for one spies of the Order Gymnophiona, and introduces a series of quwtions, both tachnical (degree of staitllng of aensory pathways, staining of lateral mcipital nucleus components) andlor empirical (projection patterns of nmr01-1~ of distinct cytoar- chitecturd characteristics, anterograde staining to specify hnervation ptbrns by partkular n m rami, comparative and ontogenetic biology). We propose to resolve them &SUM in the near future with a series of t a r H experiments utilizing a braader array of techniques and of taxa.
ACKNOWLEDGMENTS The research perid in Berkelg. in 1990 for A. Schmidt
was faditaid by fun& from the Deutache Forschungage- meinschat%. M.H. Wake’s research on d a n morphology is supported by the National Science Foundation, as is D.B. Walie’s work on salamander bjolom. We thank Karen Klitz of the Museum of Vertebrate zoology, U n i d t y of Califor- nia, Berkeley, for execution of the illustrations.
LITERATURE CITED l3rds-n- W.R., and S. Jayasinghe (1979) O b m t i o n s o n the eggs and
B , W.R, S. Nathanael, mdL. F’erkm(1987) %me aqmta ofthe =d development of l chthmhk glutinmu (Amphibia: Gymno-
phiona). J. ZmL London21 1:437-~9 . Burekhardt, R. 11891) Untemucbungen am Hm und Geruchmrgm yon
Triron undlchthpphis. 2. Wm Zool. 52369-370. Fetcho, J.R (1986) The organisation of the motoneurom innervating the
axial rnuenrlature ofwaaabrates. IC. Florida watm craakm Weds ~ ~ p ~ d r i s ) . J. Comp. Neuml.249:551-683.
larvae of2~htlsyophisgllldd110~~~. Ckyh. J. Sci. 13:187-302. ’
Fetcho, J.R. (1987) A &ew ofthe orgnnbtion m d evolution of m h n e u - mm innarratingthe axillary musculature ofvertebratas. Brain Rea Rev.
F i l i i G.T., and M.V.H. W h n (1984) Sudan Black B a@8 nerve atain for whole c h a r d fishss. Copem 1984:2&208.
Fritrsch, B., A.M. Nikundiwe, and U. Will (1QW proiectiOn patterns of kal- l ine ailbents in WE: a wmparative HRP study. d. &imp. N d . Rz9:461469.
Gaupp, E. 11899) d Ec%er‘s and R. Wietlamhmrn’r Anatinnie dm Frodm. Vol. I-In. B r a u d w e i g : V i und Soh.
Hedges, S.B., RA. Nuwbaum, and L.R h k m n (1993) caedlian phylopny and b g r a p h y inferred from mitochondrial DNA q u e m of the 125 rRNA and 16s rRNA p m (Amphibia: -om). Herp. Manogr. 7:-76.
Krabbe, K.H. (1962) Studies on the Morphogene8is of the Brain in Some Urodeles and Gymnophiom 1“Amp)llbians”) (Morph& of the Vertebrate Brain K). copenhagen: Ejmr M W .
K-nbeck, H. (19.22) Zur Morphologic des ~ U o p h i o n e n g e h i m S . Jen. Med. Nat. rn453-W.
Laubmann, W. (1427) b r die Morphagenese vom Gehin und Gerucham- gan Gymnopluonen. (Beitrug fur Kenntm . der Gymnophionen X). I. Teil. Uber die Enhicklung dm clehkm. &it. f. Anat. u. Entw. 84:697- 637.
Marcua, H. (1910) Beitrap zur k n t n h der Gymnophionen. lV. Zur Entwickluqpg64khM des Kopfea. 11. Teil. Fmtdr. R. Hertwig 2::37M. Jena, Gustav Fkher.
ma43-m.
Matew, C., and G. S&eaely (1977) Thedormmedial nuclear group of cranial n e m in the frog. Acta Bio. Acad. Sci. Hungary 28:461474.
Ma-, C., and G. Mkely (1978) Tbe motor mlumn and wmry projections of the branchial cranial ne- in tha frog. J. Comp. Neuml. 158:47-52.
Nikundiwe, AM., d LL Nieuwenhuys (1983) The cell m a w s in the brain @tern of the South A f r h u clawed W Xkqvua lmuds: A tqmgraphd and topological analysis. J. Comp. Neurol. 213:1W219.
Nhudiwe, AM., R. de Bornvan Hiuren, and HJ. t%n Donkelaar (1982) Dorsal root pmjectionsl in the clawed toad rXeMpua laeuiB) as demon-
e m 7:20i3M108.
Black B: Pr0-w VB. regreeaive methods.
mkd by m b m h labeling i&h h m d h p ~ r o x i d a ~ . N d -
Nishhw& K (1987) 8taining wphibian peripheral m with Sudan
Nishikawq KC., G. Ftoth, and U. Dicke (1091) Motor m m m aad motor 00lumns of the anterior apinal cord of -dem Pdatching d e d o p e n t and phylopnetic &hibution. Brain. Beh. Ewl. 37.368- 382.
Norria, H.W., and S.P. Hughea (1918) The uanial and anterior ~piinal mnw of the CaeEilian amphibm. J. Morphol. 31:4-.
Oh, Y., H. Tahrmchi, M. Satm, and I1 Ueda (1987) CoMtic I d a 8tudy of the morphology and diatribution of the cranial nerve efferent neuron8 Imotonerupns and pregangliollic paranympthetic neuronal and rostral apind motoneuron8 in the J a w toad. J. Comp. NeuroL 269~400- 483.
Roth, G., and D.B. Wake {IsSS) ThB 8 t rw tww of the brainstem and c e d spinal cord in lunglem damandm (Feu& Plethodontidse) and its relation to f d i n g . J. Gamp. Neuml. 241:9!%110.
Roth, G., D.B. Wake, M.H. Wake, and G. Rettig (1984) Distribution of acceamryand hypoglosd nervea in the hindbrain and spinal card of lunglew salamanders. Fmdy F‘lalhDdontrdae. Neunmci. Lett. 4.45247.
Roth, G., KC. Nishilrawa, U. Dicke, and D.B. Wake (1988) To~mgraphy and #um-chit&ure of the motor nuclei in the brainatem of salamandera. J.
Roth, G.. K.C. Niahikawa, D.B. Wake, U. Dicke, and T. Matmahima (1990)
1 9 8 7 : W 9 1 .
C m p . Neur01.27k181-194.
Mechanicsand mummorphology of feeding in amphibians. Neth. J. Zool. 1 40:IZ6-195.
Roth, G., KC. Nshikaws, C. NaqjobMantwffel, A. Schmidt, and D.B. Wake (1993) Pdomorphoais and simplification in the nervous BYBW of #ahmandam. Brain Behave Evol. 42137-170.
Schmidt, A, and G. Roth (1983) Patterns of d u l a r proliferation and migration in the developing tectum maaencepbeli of the fmg Ram tempwDria and the &ander Pkui-deks wdt l . Cell %sue Rea. a 7 2 : w ~ w .
Schmidt, k, and MH. Wake (L991) P h y 1 - m in the morph~l~gi- ad diffarentiation of the d i m tectum rnemncqhak. In N. Elmer and H. Penelin (eda.): SynapTmnamhianMduhtbm Pmc. 19th Gwttiu- e n Neumbiologsr Conferenca. Stuttgart: Georg “hieme Verlag, p. 336.

MOTOR NUCLEI OF TONGUE NERVES IN CAECILIANS 349
' 4
i
Wake, D.B. (1993) Brainatem orfpuization and branchiomuric n m . &a Anat. 148:124-131.
Wake, D.B., K.C. Nishikawa, U. M e , and G. Roth (1988) &gmhatiOn o f the motm nuclei in the cerpical spinal coni of &andera J. a m p . NmmL 278: 196208.
h y p b w Morphol. 212:37-85.
ian phylogenetic mlation&ip. Hmp. Mow. F42-56.
Wake, M.H. (1992) patterns of p r i p h d h m y ~ of the tonerue in caecilian^ (Amph~bia: Gymnophional. J.
Wake, M.H. I19D3a) Non-traditiod chanrcters in the aqtwsment ofcaecil-
Wake, M.H. (la4sb) The evohltion of oviductal wbhn in amphibians. J.
Wake, M.H. (19%~) Evolutionary d i v e m i h t i m of rranial and apinal nerve^ and their tar& in the gymnophiope amphhhs. Acta Anat. 14&.16& 188.
Wake, M.H., aud J. €hiken (1982) The dwehptnent of the skull of Dennophis maimnus (Amphibia: Ggmnophional, with mmments on skull kinesia and amphibian relationshipa. J. Morphol. 173.%%!223.
Waldwhmidt, J. 118871 Zur Am- &E N- dm -hi- onen. Jen. 2. Med. Nat. ZOr461-475.
~ x p . zool. mmwia.