Embed Size (px)
Citation preview

o a jmi jS94
The Fate of Methanol in Anaerobic Bioreactors
CENTRALE LAN DBO UWC ATA LOG US
0000 0574 0283

Promotor: dr. ir. G. Lettinga
bijzonder hoogleraar in de anaërobe waterzuivering
Co-promotor: dr. J. A. Field
KNAW onderzoeker aan de Landbouwuniversiteit

tùuoî?c> \l°i(
Lourdinha Florencio
The Fate of Methanol in Anaerobic Bioreactors
Proefschrift ter verkrijging van de graad van
doctor in de landbouw- en milieuwetenschappen,
op gezag van de rector magnificus,
dr. C. M. Karssen,
in het openbaar te verdedigen
op vrijdag 10 juni 1994
des namiddags te vier uur in de aula
van Landbouwuniversiteit te Wageningen
\SD (25180

b tb i . l o l l i l vEÄ EANDBOÜWUNIV ERSIQm
EAGEMINtiES
CIP-DATA KONINKLIJKE BIBLIOTHEEK, DEN HAAG
Florencio, Lourdinha
The fate of methanol in anaerobic bioreactors / Lourdinha Florencio. - [S.l. : s.n.] Thesis Wageningen. - With summary in Dutch and Portuguese. ISBN 90-5485-272-0 Subject headings: methanol / anaerobic bioreactors.
Cover: Methanosarcina sp. isolated from an enrichment culture on methanol at pH 4-5.

^UoP"2o i, rft(
Propositions
1. In a mixed culture degrading methanol, the production of methane does not proceed via the intermediate formation of volatile fatty acids, as was proposed by Bhathi et al.
Bhathi, Z. I., K. Furukawa, and M. Fuji ta (1993) Treatment performance and microbial structure of a granular consortium handling metbanolic waste. 1 Ferment. Bioeng. 76:218-223.
2. Contrary to the findings of Lettinga et al., a stable operation can be guaranteed in a one-step anaerobic bioreactor for the treatment of methanolic wastewater.
Lettinga, G., W. de Zeeuw, and E. Oubotirg (1981) Anaerobic treatment of waste containing methanol and higher alcohols. Water Res. 15:171-182.
3. Methanolic wastewater can be treated without addition of any alkalinity, at pH values as low as 4.2.
Chapter 4, this dissertation.
4. We are more related to methanogens than to acetogens. Woese, C. R., O. Kandier, and M. L. Wheelis (1990) Towards a natural system of organisms: proposal for the domains Archaea, Bacteria, and Eucarya. Proc. Natl. Acad. Sei. 87:4576-4579.
5. Modern pop music has its roots in Africa.
6. High-technology makes man redundant.
7. If we were to apply Dutch environmental laws to agriculture then agricultural waste could be considered as a chemical waste. Therefore, either the agricultural practices or the environmental laws are not credible.
8. In a recent electoral campaign, the leader of the Dutch conservative political party, W D , used the argument that children of illegal immigrants should not be allowed to attend school. The intention of his remark was to attract a subtile racist sentiment of certain voters.
9. Allowance of personal self-defense by means of fire arms indicates that the period of the wild west still prevails in the USA.
10. Based on the number of mountain bikes, The Netherlands can no longer be considered as a "low country" or, alternatively, urgently needs a number of artificial mountains.
11. Network does notwork.
Propositions belonging to the thesis "The fate of methanol in anaerobic bioreactors".
L. Florencio. Wageningen, 10 June 1994.

A meus pais e a Mario

ABSTRACT
Florendo, L. (1994) The fate of methanol in anaerobic bioreactors. Ph.D. Dissertation. Wageningen Agricultural University. Wageningen, The Netherlands.
Methanol is an important component of certain industrial wastewaters. In anaerobic environments, methanol can be utilized by methanogens and acetogens. In wastewater treatment plants, the conversion of methanol into methane is preferred because this conversion is responsible for chemical oxygen demand (COD) removal, whereas with the formation of volatile fatty acids (VFA) little COD removal is achieved. Moreover, the accumulation of VFA can lead to reactor instability due to pH drops, in weakly buffered systems. The undesirable formation of VFA has previously been associated with the presence of trace elements and bicarbonate in the medium.
This thesis investigates the environmental factors that lead to the predominance of acetogens over methanogens during anaerobic wastetewater treatment of methanol. For this purpose, batch and continuous experiment were carried out using a model medium composed of methanol and defined mineral nutrients. The main factors studied were: the effect of trace elements, the reactor pH, the bicarbonate level, and the methanol concentration in the reactor.
In Chapter 1 an introductory review is given about anaerobic methylotrophic metabolism in environmental biotechnology. The natural and anthropogenic sources of methanol and other methylated compounds are outlined. In addition, the anaerobic methylotophic microorganisms and the biochemistry of methanol metabolism in methanogens and acetogens are briefly described. Finally, the previous experience in environmental biotechnology with the anaerobic biodégradation of methylotrophic substrates is reviewed. Included is a summary of the reactor types, efficiencies achieved and organic loadings applied for the treatment of wastewaters containing methanol.
In Chapter 2, the effect of trace elements on the anaerobic conversion of methanol was studied. Cobalt was the only trace element tested which greatly enhanced methanogenesis from methanol. In continuous experiments, less acetate was formed in a cobalt-deprived reactor than in a cobalt supplemented reactor. These results suggested that cobalt levels could be used to prevent acetate formation from methanol. Therefore, in Chapter 3 the effect of cobalt addition for each individual trophic group was evaluated. Using specific inhibitors, specific activity and the kinetic parameters, /tmax and Ks, were determined. Methylotrophic methanogens and acetogens were the only trophic group stimulated by cobalt addition, while the other trophic groups utilizing downstream intermediates, H2/C02 or acetate, were largely unaffected. At a low cobalt concentration, both methylotrophs had similar growth rates, whereas at high cobalt levels, acetogens grew slightly faster. The optimal cobalt concentration for both methylotrophic populations was around 0.05 mg • I"1.
In Chapter 4, the possibility of the anaerobic treatment of methanol without addition of any alkalinity was investigated. Methanol can successfully be converted into methane, regardless of the low pH value of alkalinity-free medium. An astonishing high specific methanogenic activity of 3.57 g COD • g'VSS • d"1 was evident, which enable the conversion of 13 g COD • I"1 • d"1 to methane during continuous operation at a reactor pH 4.2. A Methanosarcina-\\ke organism was the main methanogen responsible for the conversion of methanol to methane.
Chapter 5 presents the effect of inorganic carbon, alkalinity addition and reactor methanol concentration in anaerobic reactors. The results indicated that acetogenesis occurred only when exogenous bicarbonate was added, when unionized VFA accumulated and in the presence of high methanol concentrations, confirming the previously determined 60 times higher substrate affinity of methanogens.
The results of this dissertation are summarized in Chapter 6. Methylotrophic methanogens will predominate over a broad pH range (4.2 to 7.2) if either the reactor methanol concentration, inorganic carbon content, or the cobalt concentration is low. Significant acetogenesis can only be expected to predominate if the reactor methanol concentration is high (>1000 mg COD • 1 "'), exogenous inorganic carbon is supplied, cobalt is available and methanogens are inhibited, e.g. by undissociated VFA. All these four conditions have to be met. Addition of moderate levels of NaHC03
(approximately 10-20 meq • l"1) were found to create such conditions if the reactor was overloaded.

Contents
1. Anaerobic methylotrophic metabolism in environmental biotecnology 1
2. Effect of cobalt on anaerobic degradation of methanol 41
3. Importance of cobalt for individual trophic groups in an anaerobic methanol-degrading consortium 61
4. Acidophilic degradation of methanol by a methanogenic enrichment culture 81
5. Substrate competition between methanogens and acetogens during the anaerobic degradation of methanol in UASB reactors. 91
6. Discussion and Conclusions (English, Dutch and Portuguese) 109
Acknowledgements 135
Curriculum vitae 137

Anaerobic methylotrophic metabolism in environmental biotechnology
1.1 Introduction
Acetate is the most important methane precursor in the anaerobic degradation of common organic constituents such as fats, carbohydrates and proteins.48,59,199 In anaerobic digesters, about 70% of the methane produced is derived from acetate whereas about 30% is generated via C02/H2 intermediates.48,59,153 Consequently, considerable research has been devoted to the behaviour of acetoclastic and hydrogenotrophic microorganisms under anaerobic conditions. Less is known about methanol and other methylated compounds, like methylamine, methoxylated aromatic compounds, halomethanes and methane. In anaerobic environments, methylated compounds are substrates for sulfate-reducing, acetogenic and methanogenic microorganisms, the final trophic groups in the anaerobic mineralization of organic matter.81,178,189

Sources of methanol and other methylated compounds
1.2 Sources of methanol and other methylated compounds
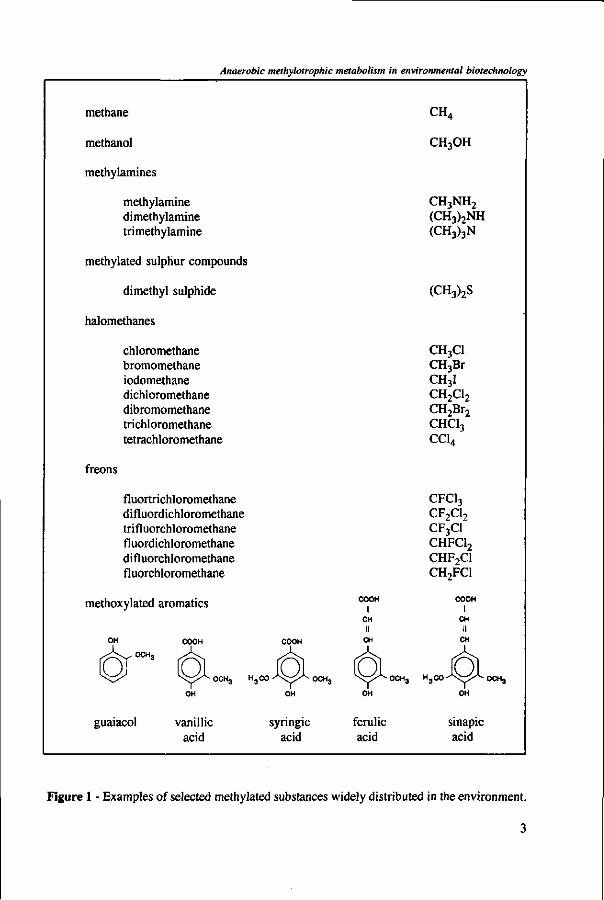
Great is the diversity of methylated substances. They can be simple compounds such as methane, methanol, methylamines, and halomethane, or they can be present in compounds with a more complex structure, like the methyl groups of methoxylated aromatic compounds (Figure 1). Methylated compounds are widely distributed in the environment as natural substances54-60-63-73'100'184 or xenobiotic substances.54-63'100
Methanol
In nature, methanol is derived from methoxylated aromatics and pectin biodégradation. Methanol is released as a fungal biodégradation product of natural methoxylated aromatics,122 which are components of ecologically significant lignin polymers. In vitro studies indicated that methanol is formed from the reaction of fungal peroxidase with 1,4 dimethoxyl benzene.64 Also, Pseudomonas putida is reported to" release methanol from aromatic acids.32 Methanol is also a major product of microbial growth on pectin142 which together with hemicellulose make up the abundant methoxylated polysaccharides in plant tissues. The methanol produced is not further utilized by aerobic, facultative and anaerobic pectinolytic microorganisms, such as Pseudomonas spp., Erwinia spp., and Clostridium spp., respectively.142 Hence, the occurrence of pectin biodégradation in anaerobic ecosystems establishes a niche for methylotrophs.144
Anthropogenic activity is also responsible for the input of methanol into the environment. Methanol is one of the organic constituents in wastewaters from dimethylterephthalate manufacturing, used for production of polyester fibers.20'130 Methanol is produced after thermoacidic precipitation of darkly coloured polyphenols in olive mill wastewaters.102 Methanol was also detected in wastewater from a potato-starch producing factory108 and in landfill leachate.175 Condensation processes in industry are known to generate methanol, such as in coal-gasification plants101 and in kraft pulping mills96'118'196
where methanol is the main organic pollutant of the condensates with concentrations ranging from 1.5 to 24.5 g • l"1.98
Methylamines
Methylated amines are widely distributed throughout the environment. In nature, methylamine and dimethylamine have been detected in vegetable and other foods.7'54
Dimethylamine is also detected as a volatile compound in cattle manure.54 Trimethylamine
Anaerobic methylotrophic metabolism in environmental biotechnology
methane
methanol
methylamines
methylamine dimethylamine trimethylamine
methylated sulphur compounds
dimethyl sulphide
halomethanes
chloromethane bromomethane iodomethane dichloromethane dibromomethane trichloromethane tetrachloromethane
fréons
fluortrichloromethane difluordichloromethane trifluorchloromethane fluordichloromethane difluorchloromethane fluorchloromethane
methoxylated aromatics
OH COOH i i
Ù" $-», . OH
guaiacol vanillic acid
COOH
3 co - x—£•*- ocH3
OH
syringic acid
COOH
1 CH
II
CH 1
ÇU OH
ferulic acid
CH4
CH3OH
CH3NH2
(CH3)2NH (CH3)3N
(CH3)2S
CH3CI CH3Br CH3I CH2C12
C-H^B^ CHC13 CC14
CFC13 CF2C12
CF3C1 CHFC12
CHF2C1 CH2FC1
COOH
1 CH
II
CH 1
H3CoA^OCH3
OH
sinapic acid
Figure 1 - Examples of selected methylated substances widely distributed in the environment.
3

Sources of methanol and other methylated compounds
is formed as a result of microbial breakdown of quartenary amines like choline, glycine-betaine, ß-alanine-betaine and trimethylamine-N-oxide, which are compatible solutes of many marine animal, plants, bacteria and macroalgae providing protection against osmotic tension.25'54'63,197 The consumption of methylated amines by methanogens may play an important role in the nitrogen cycle of marine sediments,65 because it also results in the production of ammonia, which is then available as a nitrogen source for the methanogens and other bacteria.84
The anthropogenic production of methylated amines is also important. Such compounds are utilized in the chemical industry to produce paint removers, pharmaceuticals, surfactants and pesticides.54 Consequently, methylated amines may be released into the environment by the emissions from manufacturing or product use.54
Methoxylated aromatic compounds
Methoxylated aromatic compounds are ubiquitously present in the environment. Their most important source in nature are low molecular plant phenolics, de novo fungal metabolites and from the fungal degradation of lignin. Plant phenolic compounds occur in most fruits and most of them contribute to color and taste.7 Ferulic, sinapic, and vanillic acid are examples of methoxylated plant phenolics widespread in fruits and vegetables.7
Therefore, they are expected to be present in wastewaters in the food processing industry like in the production of fruit juice, syrup, jam, canned fruit, wine, etc. Common wood- and forest litter-degrading fungi are responsible for the de novo biosynthesis of methoxylated anisyl and veratryl metabolites.60 Lignin is an at random methoxylated aromatic polymer, resistant to hydrolysis which comprises from 20 to 30% of wood dry weight.18 Lignin is the most abundant and widely distributed renewable source of aromatics on earth.128 During fungal degradation, lignin yields soluble and low molecular weight compounds, such as the methoxylated aromatic vanillic acid.73 Vanillic acid is the major monomeric intermediate of fungal degradation of lignin.60 In the chemical processing of wood, lignin derivatives are major components in the wastewater streams. 15°
Halomethanes
Halomethanes, such as chloromethane, dibromomethane and bromoform, are largely spread in nature. They can be produced by the action of seaweed and phytoplankton in the oceans,185 or by the action of wood-rotting fungi in terrestrial environments.50 However, the extensive industrial use of synthetic halomethane, such as di- and trichloromethane in

Anaerobic methylotrophic metabolism in environmental biotechnology
solvents, degreasers, pesticides and intermediates in chemical synthesis, may result in localized ground water and drinking water contamination.13'38,54,63'180 Because of their persistent behaviour in the environment, and due to the fact that many halomethanes are carcinogenic or mutagenic, they can consequently be considered as a potential public health hazard.180 Moreover, the increase of halomethane in the atmosphere becomes an important environmental concern due to their ozone depleting potential.184,185
Methane
Methane is the most abundant hydrocarbon in the atmosphere.174 Methane in the atmosphere results from both biological and abiotic sources.76 Biologically derived methane is the end product of the anaerobic degradation of organic matter. It occurs in a wide variety of anaerobic environments where oxygen and sulphate are scarce such as swamps, peat bogs, wetlands, fresh water sediments, rice paddies, animal gastrointestinal tracts, landfills, wet wood of living trees and anaerobic digesters.37,76,84'99'134'171,172,174 Abiotic sources of methane are formed by geothermogenic processes in fossil organic material covered by sediments and also by incomplete burning of wood and fossil fuels.3,174 Fossil methane is released during mining and transportation of coal, lignite, petroleum and natural gas hydrocarbon to the surface, and also during the refining and incomplete combustion of petroleum.3,174 The increase of methane concentration in the atmosphere is of great concern because it plays a direct role in climate changes resulting from the greenhouse effect.76,174
The atmospheric methane budget with its climate consequences has been subject of much research.3,37,76,99,129,174
1.3 Anaerobic methylotrophic microorganisms
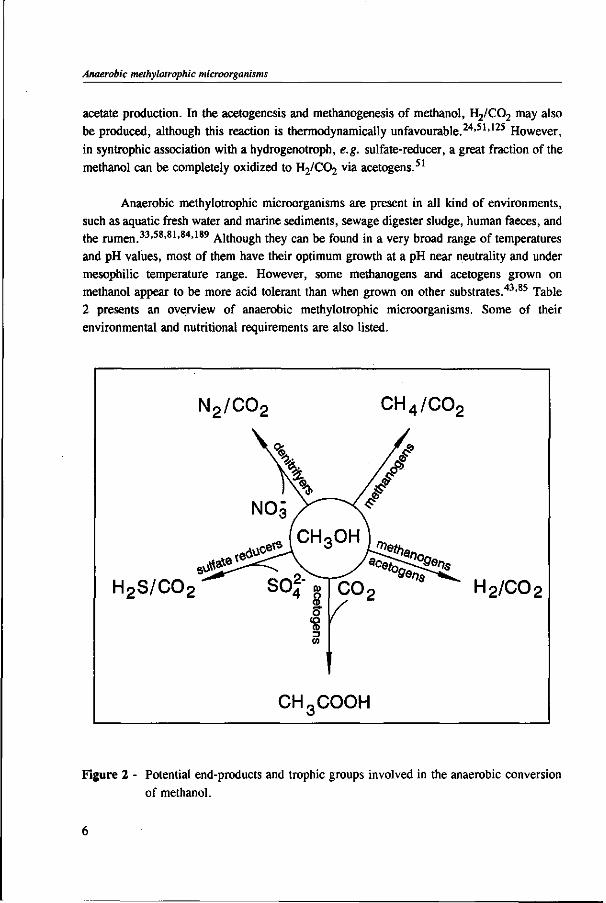
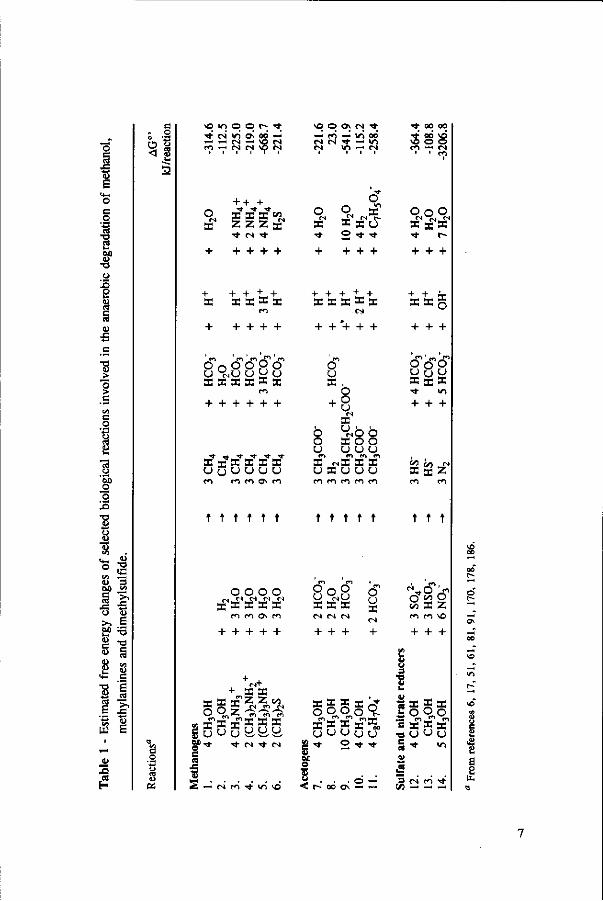
Under anaerobic conditions, methanol can be utilized by several groups of microorganisms. Figure 2 illustrates the potential end products and trophic groups involved in the anaerobic conversion of methanol. When an electron acceptor is present, such as nitrate or sulphate, methanol can be converted to C02 by nitrate- and sulfate-reducing bacteria,91,109 and in the presence of C02, acetogens are able to produce acetate and butyrate from methanol.42,77,81,200,201 Methanogens reduce methanol into methane without requiring any external electron acceptor.160,178 Table 1 presents the estimated free energy changes of selected biological reactions involved in the anaerobic degradation of methanol and some methylated compounds. The reduction of nitrate or sulphate produces more free energy than the formation of methane or acetate from methanol. However, in the absence of nitrate and sulphate, the production of methane is a thermodynamically more favourable process than
Anaerobic methylotrophic microorganisms
acetate production. In the acetogenesis and methanogenesis of methanol, H2/C02 may also be produced, although this reaction is thermodynamically unfavourable.24'51'125 However, in syntrophic association with a hydrogenotroph, e.g. sulfate-reducer, a great fraction of the methanol can be completely oxidized to H2/C02 via acetogens.51
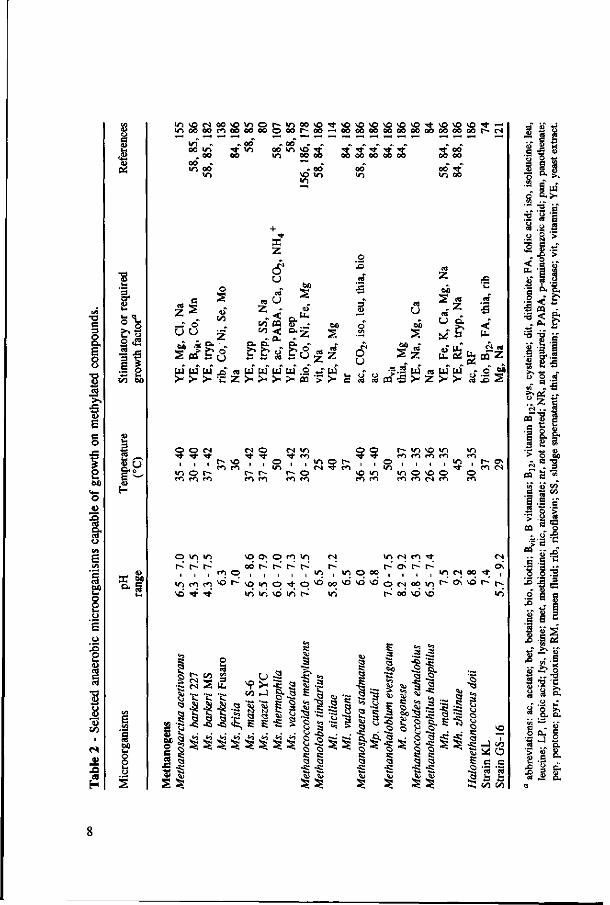
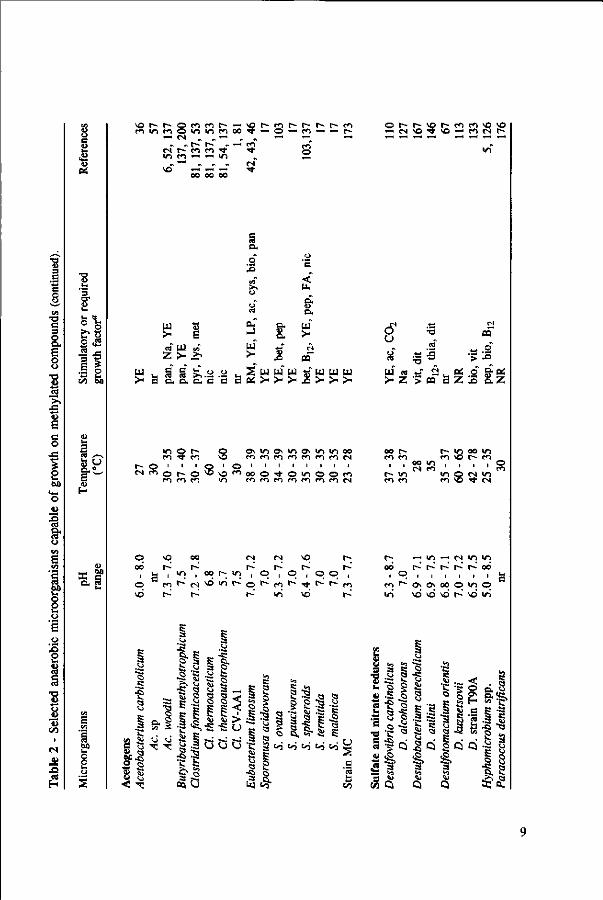
Anaerobic methylotrophic microorganisms are present in all kind of environments, such as aquatic fresh water and marine sediments, sewage digester sludge, human faeces, and the rumen.33'58'81,84*189 Although they can be found in a very broad range of temperatures and pH values, most of them have their optimum growth at a pH near neutrality and under mesophilic temperature range. However, some methanogens and acetogens grown on methanol appear to be more acid tolerant than when grown on other substrates.43'85 Table 2 presents an overview of anaerobic methylotrophic microorganisms. Some of their environmental and nutritional requirements are also listed.
N 2 /C0 2 CH 4 /C0 2
H 2S/C0 2 H 2 /C0 2
CH3COOH
Figure 2 - Potential end-products and trophic groups involved in the anaerobic conversion of methanol.
o
'S Où u
T3
Xi O
1> -C
"8 > o > c
I o '5b o "g IS
CA
V I ^ -<
« 3 °* J2 U <L) >, E £f '•o
ë 'S
I ë "8 1 E J3
• FH -4—»
3 ë pq E ^4
4> £i CS H
c o
öS
<J »
« in q o ^ * >*' es vi o\ od -^ m •" N « c M
i i i i T i
O ac
+ ac
+ + + X X X <«
zzza" •* e s •>*
+ + + +
+ + t - +
*r ac ^ X
+ + + +
(*l Cl 0 \ f t
t t t t t t
O O O O ^ n es <s «s
5È X X X X co co C\ ro
+ + + + +
<s+ + X X
_ ac J z z co
m t_C1 m HM HH hr" «Î *T" X tè ^ •J-1 *J-1
vq o o\ es| •* —' co —' >ri od N N * - « CS VI —' CS
o es
X
+
O
ac x u 2 •* •* + + +
+ + + + + X X X X x
CS
+ +*+ + +
CO CO CO CO CO
t t t t t
O 0 0
X X X
+ + +
o u X cs
+
e •* CS Tt- CS
es ro •* v> ^o
a s * X n * o o o o 9-x ?c x x * u U U (j u T»" O , )• ^
. . O — oo o\ — —
• * 00 00
S ë c S co — CS
i i ro
O O o X x*X
+ + +
X X
+ + +
O o o o p o u ^ u u y u X X X X * X
co
+ + + + + +
• * ^ . -«t - * • * • *
yc ac Ä *™ ™ X U u <-> <~> u u
o u X ,
o + ° (S
X o S^o o O X o o u u u u
r*") f ï <*ï f * l
SC CS HM H C HM
U X u u u
O o O U u u x ac x 't *n + + +
22 '<*> ~F x ac z
î T f
O C3 Q oo X Z m m \ o
+ + +
8
"S
s x x ac i o o o c ac x ac •o o u u § •* "> CU
I 3 O» co Tt
CÄ — ~ - ~ -
3 g. S o o
1
e o •B
o
o
o
e
1 'S
CO
— Xi « H
•8
S .2
.§ B
X »
o o
m NO <s m o o o o
i n » oo m
oo m oo
r n oo oo oo o — ~~ * * -^ — — — — — — — — — — — - •
. 00
00
o o o r ~ - o o * ^ o o o o o o o o o o o o o o o o o o o o r ~ c N
i n oo oo
NO m T l
00 oo OO 00 oo
oo" m
T f OO oo oo
OO Tj-i n oo
Z 2
_ - o~ U U
S o f IÜUJ
o
<u co
Z
«-" CJ W / a >" 'E
PC
z ó4
o CS
« U
co CQ t o <
- O .
& & •
O«
iö. o u
I «
o£
oo
S „ Z
t u
f.2" w" « tu 03 5 > Z >-
l - i < u.
es cd
o f Z o" oo
o 1
i n m
o •
o m
f N
i
r-m
r- NO
I N
i
r-m
©
r~ m
o IT)
f N •* i
r— m
i n
fX o o
? ?? t - m NO m m
m m m m m f~ i i O , , , , IQ , t - ON
NO i n m o NO o o m m m r o r j m m
o i n i n
r-' t ~ r~' C l o
NO ON O m i n od r^ r~ r - t >
CN i n M m 4 t>° ON t-' (-
i f i m n ^ ^ N O K i q ^ O NO ^ ' • * i n i n N O i n ( ^
i n ", i n o o o ' , | ' , " , i n N o o * ,
N O o o v o v o v o ' o f > i o o m ' ^ 0 ^ v ö r ~ ' r - -
i n t—' od NO vb m
V}
o
<>> g •J.
1 1 s-CI
• j :
*! s^
r-rs
co
s
p Sa Vi u.
bark
eri
bark
eri
bark
eri
u •<?>
fris
ia
maz
ei S
m
azei
L
.g «
•s
p a ^ 3 <-> §
â£
£> S
•S §
e « •s
t-> 2
|
s-Ç| •g %) £ §
.3-2
S •«
•1§ ~ - G
• i ^ §
»15 •5
ft c a •5
5 « •S 5 c a
s C
•g
î " -~ 2 3 - s .e;
~ "S 5 •s 2 5
-§
•5 G
NO
< s s < : J l o
• S <s c c
,5N>
^ * I cd ecj
S
S S o •2 g 2
ïi i 1 9 1
s a'g
< | y o a .a
I f a u O ta w - *3
" Z g < t . - S
cö"S J
c Ç il O M
. o ' S
co S
•2 •c
'i 5 5 3 * 3 <â o p e
s 1 e 'S « s £ -s 2
* « s « - 1 a -o -c
U £ M
•U
'> S a.
•SJ 8.
'S
B O
e o o
? ">>
e c o
p
o
e o
i
i 00
JU
S es
H
3 c
'S '3 8" IM
2 <2 co _
* > « r- t - - — en
es r-- m m Tt- TJ-VI c i - H — w->
« - " . . . <s vo —i —' —• • *
0O CO 00
e i i— t— r- i— m o —' e i ~ — r-
U «5 >• E « UÜ ui"
z >- >> g Ü § § £.a >" S o, o. o. e
o IS
o.
> 5
< u.
u
m . S w w* w „ „ „ „ e ai > > - > - * >• > >>
CS o C l
>/i
ei
o m
S o
ei
O en
S s
1
vo IT)
O ci
Cl 1
00 ei
o •
o Cl
Cl i
• *
C l
>/-i C l
o Cl
Cl i
Cl
«1 ci
1
o C l
</-l C l
1
o Cl
00
1
Cl
r-° oo f S f S VO
r-'
O
«3
l/l , » h i f l . O . O , O ©
r~ r- r-' u-i vo i—'
S 3 S
a
S 5 S .?> i? .Ü fc
'S =3 £ •£ C3 G 3 ^_ ï * C\ r* r>*
S i c
_ < 2. S < c
J > I •S u g
tiöl
g
:§ a
•g fc S
t . to to to to to
c o o E
I'S
© t — r ~ v o r - c i c i v o v o — (S »O ^ vo — c i c - ) r -
O U
S 'S
PQ „ o" '> £
>- z >oa c Z x i o . z
oo t— M n o o m c i c i c i vo r- e i
, , oo v i , , , , o <s e i ' Cl
t— </i i n © ts i n c i c i e i vo •<*• <s
- • i n - N m i n r~ i--' t-' r-' r»° oó
ON ov oo o >n o vo vo vo r»° vo i/i
3
"8 1 S •5
• " JN "~> W
c 2 a 'S
f Ci
3 co s : ; fa 3 *>
3 «o
Q
1 s ä 3 »)
3
Q
< .1
fal-ë " § 3

Anaerobic methylotrophic microorganisms
Sulfate- and nitrate-reducing bacteria
Sulfate-reducers are obligately anaerobic microorganisms (bacteria and archaebacteria) that use sulphate and other oxidized sulphur compounds as terminal electron acceptor for the dissimilation of organic compounds.47,189 Aside from methanol,16'109 H2, formate, acetate, propionate, ethanol, acetone, lactate and amino acids are examples of other substrates in the large spectrum of compounds used as electron donors.187'188'189 Sulfate-reducing bacteria are of fundamental importance in the sulphur cycle in aquatic environments. Among sulfate-reducers, members of the genera Desulfovibrio and Desulfobulbus appear to be equally common in fresh water and marine or other saline habitats.188 Desulfovibrio carbinolicum, isolated from an anaerobic wastewater treatment plant, was the first reported sulfate-reducing bacterium capable of growth with methanol as the sole source of energy.109'110
Nitrate reducers are a mixed group of morphologically and nutritionally diverse micro-organisms (bacteria, archaebacteria and the protozoan genus Loxodes) able to reduce nitrate and nitrite to NO, N20 and N2.
132.'76>203 However, not all denitrifying organisms are able to utilize both nitrate and nitrite.176'203 Several organic compounds such as sugars, amino acids, alcohols and organic acids, can be utilized as the electron donor and carbon source. Among them, methanol is the most satisfactory organic chemical used in denitrification processes because it provides high denitrification rates.91 '132 However, several distinct genera of microorganisms, like Thiobacillus, Pseudomonas, Nitrosomonas and Nitrobacter can also oxidize inorganic sulphur compounds, hydrogen, ammonia and nitrite during denitrification.203 Denitrification is generally most efficient under anaerobic conditions, but many bacteria can denitrify at significantly high oxygen concentrations. Denitrifying microorganisms play an important role in the nitrogen cycle, returning fixed nitrogen to the atmosphere. Paracoccus denitrificans, one of the most studied denitrifying bacteria, can utilize a large variety of carbon and energy sources, both under aerobic and anaerobic conditions with nitrate, nitrite or nitrous oxide as terminal electron acceptor.176
Methanogens and acetogens
In the absence of sulphate, nitrate or oxidized metal ions (Fe3"1", Mn4+), methanogens and acetogens are expected to be the predominant group of microorganisms in the anaerobic conversion of methanol.
Methanogens are strictly anaerobic archaebacteria that produce methane for growth from a limited number of substrates, such as CO, H2/C02, formate, methanol, methylamine or acetate.58'178 Some methanogens can also utilize secondary alcohols such as 2-propanol
10

Anaerobic methylotrophic metabolism in environmental biotechnology
and 2-butanol, but to a lesser extent because the oxidation is always incomplete.74'202
Pyruvate, a substrate more complex than acetate, was recently reported to support growth of a methanogen as a sole carbon and energy source.12 Methanogens play an important role in the last step of the anaerobic food chain, converting the fermentation products of higher trophic levels into the anaerobically relatively inert products methane and carbon dioxide which are released into the atmosphere. Among methanogens, Methanosarcina barken is physiologically the most versatile methanogen described. Methanosarcina sp. can utilize all methanogenic substrates except formate. Most of the studies on methylated compounds are done with this organism.
Acetogens are obligate anaerobes that synthesize acethyl-CoA for both conservation of energy and growth, and usually form acetate as their main end product.33 Methanol, methyl chloride and trimetylamine are examples of methylated compounds in the large spectrum of substrates used by acetogens, that include H2/C02, CO, various sugars, several methoxylated aromatics, primary aliphatic alcohols and amino acids.29'33'81'103'201 They play a key intermediate role in the bacterial anaerobic degradation of organic matter, producing acetate and H2/C02, which are methanogenic substrates. Among acetogens, Clostridium thermoaceticum has a special historical place because it was with this microorganism that studies were first conducted to elucidate the biochemical pathway of the autotrophic fixation of carbon dioxide by acetogens, the acetyl-CoA (Wood) pathway.33'81
Methanogenesis and acetogenesis from methanol
The methanogenesis71'147'154'160'178'182-201 and acetogenesis6-42'77'78'81'200'201 from methanol is well known. In relation to kinetic parameters, there is information available in the literature about growth rates and cell yields for acetogens and methanogens cultivated on methanol; however, little is known about the affinity for the substrate. Table 3 presents the growth rate and cell yield coefficients for selected acetogens and methanogens grown on methanol and other carbon sources. Methanogens clearly have a higher cell yield and grow faster on methanol compared to acetate, with values equal or higher than when they are grown on H2/C02. Acetogens and methanogens have similar growth rates with methanol. However, acetogens have higher cell yields, which may result in a much lower specific activity in comparison with methanogens.
Besides methanol, the spectrum of methylated compounds that can potentially be used by methylotrophic methanogens and acetogens is large. Table 4 presents some microorganisms involved in the degradation of other methylated compounds than methanol, such as halomethanes and methoxylated aromatic compounds.
11
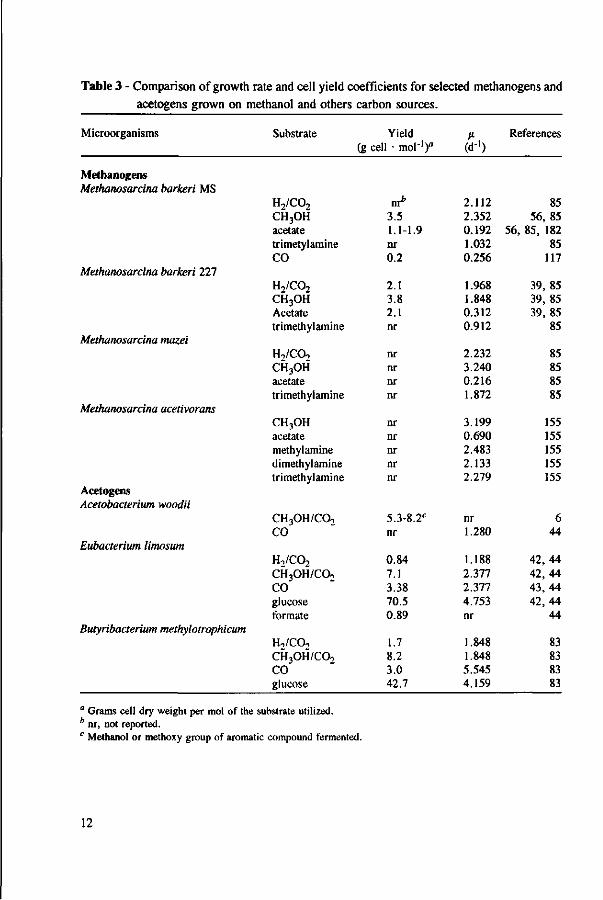
Table 3 - Comparison of growth rate and cell yield coefficients for selected methanogens and
acetogens grown on methanol and others carbon sources.
Microorganisms
Methanogens Methanosarcina barken MS
Methanosarcina barken 227
Methanosarcina mazei
Methanosarcina acetivorans
Acetogens Acetobacterium woodii
Eubacterium limosum
Butyribacterium methylotrophicum
Substrate
H2/C02
CH3OH acetate trimetylamine CO
H2/C02 CH3OH Acetate trimethylamine
H,/C02
CH3OH acetate trimethylamine
CH3OH acetate methylamine dimethylamine trimethylamine
CH3OH/CO-, CO
HVC02
CH3OH/CO., CO glucose formate
rWC02
CH3OH/C02
CO glucose
Yield (g cell • mol-')a
nr* 3.5 1.1-1.9 nr 0.2
2.1 3.8 2.1 nr
nr nr nr nr
nr nr nr nr nr
5.3-8.2c
nr
0.84 7.1 3.38 70.5 0.89
1.7 8.2 3.0 42.7
(d-1)
2.112 2.352 0.192 1.032 0.256
1.968 1.848 0.312 0.912
2.232 3.240 0.216 1.872
3.199 0.690 2.483 2.133 2.279
nr 1.280
1.188 2.377 2.377 4.753 nr
1.848 1.848 5.545 4.159
References
85 56, 85
56, 85, 182 85
117
39, 85 39, 85 39, 85
85
85 85 85 85
155 155 155 155 155
6 44
42,44 42,44 43,44 42,44
44
83 83 83 83
" Grams cell dry weight per mol of the substrate utilized. * nr, not reported. c Methanol or methoxy group of aromatic compound fermented.
12

Table 4 - Methylotrophs and spectrum of methylated substrates or co-substrate metabolized under anaerobic conditions.
Microorganisms
Methanogens Methanosarcina barkeri
Methanosarcina sp. DMC
Methanosarcina mazei
Methanosarcina acetivorans Methanococcoides methylutens strain GS-16
Methanolobus siciliae
Acetogens Acetobacterium carbinolicum Acetobacterium woodii
Eubacterium limosum
Clostidium thermoaceticum
Clostidium thermoautotrophicum
Sporomusa sphaeroides strain MC
Sulfate-reducing Desulfotomaculum orientis
Substrate
tetrachloromethane methylamines trichloromethane tribromomethane methylamines dichloromethane tetrachloromethane trichloromethane tribromomethane tetrachloromethane methylamines methylamines trimethylamine trimethylamine dimethyl sulphide dimethyl sulphide trimethylamine
methoxylated aromatic trichloromethane chloroguaiacols tetrachloromethane vanillic acid syringic acid
compounds
3,4,5-trimethoxybenzoic acid vanillin syringaldehyde 2,4-dimethoxylbenzoic acid 3-methoxycinnamic acid anisol trimethoxybenzoate chloroguaiacols vanillate syringate ferulate trimethoxybenzoate vanillate syringate vanillate syringate trimethylamines methychloride methoxylated aromatic
trimethoxybenzoate
compounds
References
69 53, 112, 179
95 95 95 95 95 95 95 95
53, 179 155 155 121 121 114 114
36 34,35
49 34, 35, 162
6 6, 143
6 6 6 6 6 6
23 49
.23,44 23, 44
44 23 26 26 26 26
103 92, 173
173
67
13

Biochemistry
Anaerobic methane oxidation
The presence of methane in abundant quantities in anaerobic environments raises the question if nature has not devised methods of utilizing this substrate in the absence of oxygen. Although it is a highly controversial topic,145 there is some evidence that biodégradation of methane occurs in anoxic marine sediments.2'181 In fact, some methanogens are reported to be able to oxidize a small fraction of the methane produced,198
and there are indications that sulfate-reducing bacteria may also be able to oxidize a small amount of methane when an additional electron donor is present.28 However, the organisms responsible for anaerobic methane oxidation have not yet been isolated, and little is known about the mechanisms involved in the process.129 The fact that methane is usually a stable end product recovered in high yields during anaerobic digestion and anaerobic treatment of wastewater indicates that significant methane oxidation is not encountered in natural mixed cultures of bioreactors.
1.4 Biochemistry
In recent years, much has been learned about the mechanisms by which methane and acetate are produced by methanogens and acetogens, and the metabolic pathways have been partially resolved. In the elucidation of these pathways, several enzymes, coenzymes, and cofactors, most of them containing transition metals, have been discovered.81,178
Metalloenzymes carry out both electron transfer processes and catalyze essential metabolic reactions in the cell.179 Examples are factor F430 which contains nickel,30 carbon monoxide dehydrogenase containing nickel, zinc and iron81 and corrinoid proteins containing vitamin B12 derivatives in which cobalt is the central ion.164 A corrinoid-dependent methyl transferase catalyses the initial step of methanol conversion in methanogens and acetogens.93'94'165'166 After that, methanol is channelled into the central pathway for C r
compounds for both groups of microorganisms.81'178'201 An overview of the biochemical pathways is illustrated in Figures 3 and 4 and the main features are summarized in the following paragraphs.
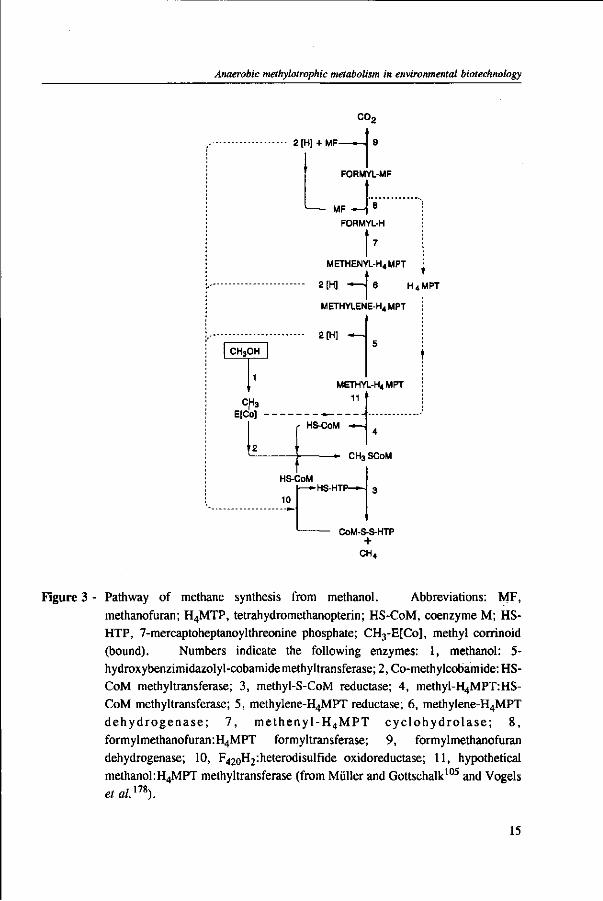
Methanogens
In methanogens, independently of the substrate used for cultivation, several unique coenzymes are involved in the intermediate steps of methane formation, namely coenzyme M, factors F420 and F430, methanofuran, tetrahydromethanopterin and 7-mercaptoheptanoylthreonine phosphate.135 When methanol is used as the sole substrate, one
14
Anaerobic methylotrophic metabolism in environmental biotechnology
CH3OH
CH3
E[Co]
"2
C 0 2
2 [H] + MF - - J 9
FORMYL-MF
MF *—| 8
FORMYL-H
METHENYL-H4MPT
2[H] - ^ 4 e H4MPT
METHYLENE-H4MPT
2[H] -
METHYL-H4 MPT
11 t
HS-CoM
CH3 SCoM
HS-CoM
10 -HS-HTP-«-
CoM-S-S-HTP +
CH4
Figure 3 - Pathway of methane synthesis from methanol. Abbreviations: MF, methanofuran; H4MTP, tetrahydromefhanopterin; HS-CoM, coenzyme M; HS-HTP, 7-mercaptoheptanoylthreonine phosphate; CH3-E[Co], methyl corrinoid (bound). Numbers indicate the following enzymes: 1, methanol: 5-hydroxybenzimidazolyl-cobamidemethyltransferase; 2, Co-methylcobamide: HS-CoM methyltransferase; 3, methyl-S-CoM reductase; 4, methyl-H4MPT:HS-CoM methyltransferase; 5, methylene-H4MPT reductase; 6, methylene-H4MPT dehydrogenase ; 7, methenyl-H 4MPT cyc lohydro lase ; 8, formylmethanofuran:H4MPT formyltransferase; 9, formylmethanofuran dehydrogenase; 10, F42oH2:heterodisulfide oxidoreductase; 11, hypothetical methanol:H4MPT methyltransferase (from Müller and Gottschalk105 and Vogels et al.m).
15

Biochemistry
quarter of the methanol has to be oxidized to C02 in order to provide the organisms with reducing equivalent for reduction of methanol to methane.10 In the reductive route, two methyltransferases containing corrinoid participate in the formation of methylated coenzyme M, the intermediate precursor of methane.94 In the oxidation route, the exact place where methanol enters in the pathway is not yet clear. However, evidence indicates that it might be at the level of tetrahydromethanopterin or coenzyme M as outlined in Figure 3.105>178
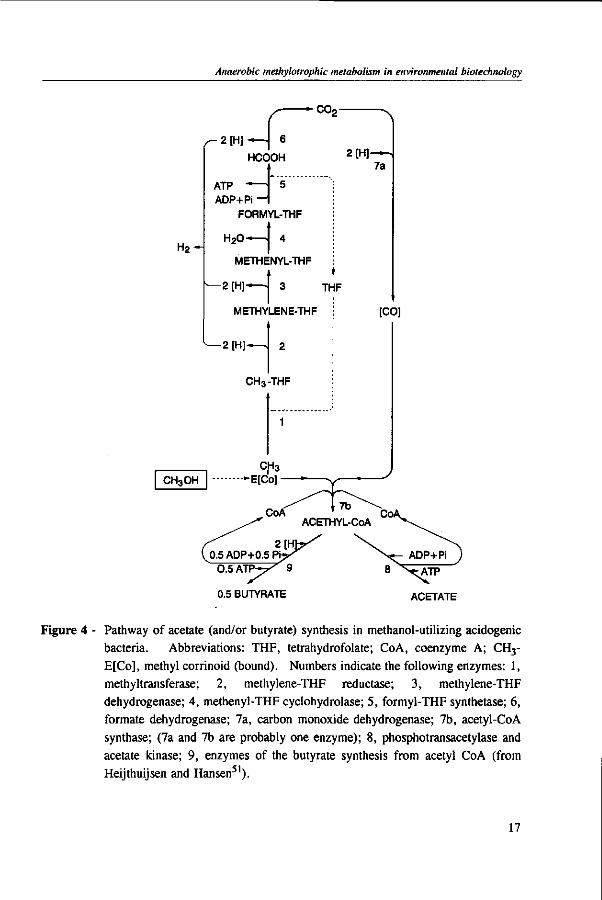
Acetogens
In acetogens, several enzymes are involved in acetate synthesis, such as formate dehydrogenase, corrinoid protein, and carbon monoxide dehydrogenase.81 Carbon monoxide dehydrogenase has a fundamental role in the synthesis of acetyl-CoA, the precursor intermediate of acetate and cell synthesis.193 The formation of acetate from methanol is only possible if other carbon containing compounds more oxidized than methanol such as formate, CO, C02 are present.81,201 Additionally, at low hydrogen partial pressure in coculture with hydrogenotrophic microorganisms, the oxidation of methanol to C02 and H2 may also occur.24,51 The first step of methanol conversion in the acetogen Eubacterium limosum is catalyzed by a methyl-transferase containing corrinoid.93 As in the methanogens, the point where methanol enters the pathway to C02 is still not known, but methyl-tetrahydrofolate is probably one of the first intermediates.51
The corrinoid content
Although methanogens and acetogens phylogenetically belong to distinct domains, a taxonomie division above kingdom level,192 they have in common several features: both microorganisms are strict anaerobes found in similar environments and utilize some of the same substrates.201 Moreover, in many ways the conversion of methanol and other Cx-compounds by these microorganisms have many analogies. These include: sodium dependence,11,52'124'191 acetyl-CoA as a precursor of cell synthesis,55'193 and tetrahydrofolate, the Cl-carrier in acetogens, that is structurally and functionally analogous to tetrahydromethanopterin.82'105 However, it is in the content of corrinoids that methanogens and acetogens grown on methanol share the most similarities. This common characteristic is most likely due to the fact that the initial step in methanol conversion for both groups is catalyzed by similar methyltransferases containing the common corrinoid cofactor.93'94 In addition, there is growing evidence that in methylotrophs, induced-enzymes containing corrinoids are produced by exposure to methanol, such as in Sporomusa ovata.165'166
16
Anaerobic methylotrophic metabolism in environmental biotechnology
CO,
H 2 -
CH3OH
• 2 [ H ] — - I 6
HCOOH 2[H]-7a
ATP - — 5 ADP+Pi -H
FORMYL-THF
H 2 0 ^ — 4
METHENYL-THF
-2[H]-—j 3
METHYLENE-THF
•
THF
-2[H]-
CH3-THF
[CO]
'E[Co]
CoA ' ' " Co, ACETHYL-CoA
2[H , 0.5ADP+0.5PI
0.5 Kï?y/ 9
0.5 BUTYRATE
Figure 4 - Pathway of acetate (and/or butyrate) synthesis in methanol-utilizing acidogenic bacteria. Abbreviations: THF, tetrahydrofolate; CoA, coenzyme A; CH3-E[Co], methyl corrinoid (bound). Numbers indicate the following enzymes: 1, methyltransferase; 2, methylene-THF reductase; 3, methylene-THF dehydrogenase; 4, methenyl-THF cyclohydrolase; 5, formyl-THF synthetase; 6, formate dehydrogenase; 7a, carbon monoxide dehydrogenase; 7b, acetyl-CoA synthase; (7a and 7b are probably one enzyme); 8, phosphotransacetylase and acetate kinase; 9, enzymes of the butyrate synthesis from acetyl CoA (from Heijthuijsen and Hansen51).
17
Biochemistry
Table 5 presents the corrinoid content in methanogens and acetogens growing on methanol and other substrates. The corrinoid contents of methanogens and acetogens vary among species and are dependent on the growth substrate used to cultivate the organisms, but they are always higher when cultivated on methanol.70,200
Due to the large amount of corrinoids produced by methylotroph methanogens and acetogens, it has been proposed that they should be utilized for the industrial production of vitamin Bj2 .90 '200 Moreover, Methanosarcina spp. are reported to release great amounts of extracellular corrinoid into the medium.90,111 Therefore, several attempts have been made to optimize the growth culture conditions for the improvement of vitamin B12
productivity.90,111,116,152
Table 5 - Corrinoid content in methanogens and acetogens.
Microorganism
Methanogens Methanobacterium arbophilicum Methanosarcina barkeri MS Methanosarcina barkeri Fusaro Methanobacterium formicwn Methanobrevibacterium ruminantium PS Methanobacterium thermoautotrophicum
Acetogens Acetobacterium sp. Acetobacterium woodii Butyricumbacterium methylotrophicum Clostridium formicoaceticum Qostridium thermoaceticum Sporomusa ovata
Acetate
Carbon
Methanol (nmoles corrinoids •
nr° 1.6 nr nr nr nr
nr nr nr nr nr nr
nr 4.1 4.3 nr nr nr
8.16 nr
5.6 0.95 nr
3.1
sources
H2/C02
mg"1 cell dry
0.91 2.5 nr
0.6 0.86 0.66
nr 0.65 nr nr 1.20 nr
Glucose weight)
nr nr nr nr nr nr
nr nr
0.25 nr nr nr
Ref.
70 70 90 70 70 70
57 163 200 163 163 163
nr, not reported.
18

Anaerobic methylotrophic metabolism in environmental biotechnology
1.5 Methanol in environmental biotechnology
The anaerobic biodégradation of methylotrophic substrates is of importance to environmental biotechnology for a number of reasons. Firstly, it can provide high chemical oxygen demand (COD) removal efficiency in wastewaters containing high concentrations of methanol, such as kraft pulping condensates. Secondly, methoxy groups can be metabolized, an essential first step in the biodégradation of many aromatic compounds. In addition, the priority halomethane pollutants can be bioremediated by anaerobic processes. Furthermore, methanol is regarded as a suitable and inexpensive cosubstrate for a variety of purposes such as denitrification and dehalogenation.
For wastewaters predominantly containing methanol, several types of high-rate reactor configurations have been reported (Table 6). For a kraft evaporator condensate, efficiencies of 80% of the COD removal were achieved at organic loading rates of 2, 10 and 13 g COD • l"1 • d"1, using an anaerobic filter, an expanded and fluidized bed, respectively, all operated in the mesophilic range.118 At 53°C and with an organic loading of 45.3 g COD • l"1 • d"1, 87% of the COD was removed using a fixed-film bed packed with pumice stone in combination with membrane filtration.98 A COD removal efficiency of 95%, at an organic loading rate ranging from 10 to 15 g COD • l"1 • d"1, was reported for a wastewater containing methanol from the production of formaldehyde.177 For the treatment of synthetic methanolic wastewater, the upflow anaerobic sludge blanket (UASB) reactor, operated in the mesophilic range, achieved up to 99% COD removal at organic loading rates as high as 112 g COD • l"1 • d"1.117 However, several operational problems are reported, such as accumulation of volatile fatty acids and washout of biomass.9'77'78'117'169'195
Several acetogens are able to remove the methoxyl group of methoxylated aromatic phenols to produce acetate in a similar manner as in the acetogenesis of methanol.6,23,26'36'44
The methoxylated compounds such as vanillate, syringate, ferulate are transformed to their corresponding hydroxy derivatives, leaving the aromatic ring intact.6 Trihydroxybenzenes, such as gallic acid and phloroglucinol can be thereafter completely degraded to acetate by the fermentative organisms like Pelobacter acidigallici.I43 However, the ring cleavage seems to be only possible after the removal of methoxy groups,61 since P. acidigallici could not grow on methoxylated aromatics alone, but could grow in coculture with Acetobacterium woodii.143 Such a kind of partnership might have been present in mixed cultures, where the complete degradation of the methoxylated aromatic syringic acid and guaiacol to CH4 and C0 2 was achieved.8'61 In addition, anaerobic treatment processes can lead to conversion of chlorinated methoxybenzenes, such as chloroguaiacol from a kraft mill bleaching effluent, into the corresponding demethylated chlorophenolic compounds such as chlorocatechol.123'194
19
%
60
O
1 c/ï
E
"g
S GÛ
K i
V©
o ^ U 6S
•o
-J — o •
Q O U &
on
WH
I 2 <_>
CD o
I
i - a>
u S
« s1
r- oo o \ t - t— i— r - ^ v o o o r - v o v o o o o o ( r"~ '"•' » - 1 ~ ^ f-* " - 1 O N O N O ^ CN ^ ^ -
en o\ 0\ ö ON ON I - —
, . , « oo ON w> O r- , M i n O O ' ON ON ON ON 00 00 ' O N O N O O O O
O O O 0O M N in r-
-*• ei <s |2 üS^r-.^ N N m r .. .. ™ rO_« —
w ^ ^ i N i i O t i I (N « T). ' ^ ' i i ' ^ ^
oi <N <N "" 5 " a -'S
o 43
8 u-i vi in
S i S S g o o - S o o o o
O O O O TO. TO TO TO •S 'S S 'S B B S ®
E S S E
» n <u n)
O
CU
« CU
« O
S s s s <D
s U S
oo
' .. « « <U c Ö •a g g
2 u- u-
es </ï e <u
• a
eu <u eu
C/ï t/S C/î c e s <u <u a>
- o - o - a e e e o o o o u o
i i « «
«- o o " Ä TOTO'
I s Ö S * « CU « « ~ »- »-E . * .m
o o o O* Cu Cu
cu t o <D
<g * <g M 1_ Ui Wt
^ ^ M M
• S * .
a) d> cu cu
e c c c 2 S 3 2 C/3 C/3 C/3 C/3
E E S E - 2 1 2 5 2 3 3 « O- Cu Cu U t/3 CA
I'S 'S "S "S 'S 'S 'S
O - « 4 3 4 3 . 0 4 3 4 S - a ï a C O C D O a O Q P Q S Q m — i » - i — - - . n i e •— o n t z i t z i t y j c o t z i J ^ ^ s S ö B 5 3 <<<<<<OÜ S S S S *_=. D 3 D 3 P D L Ü U . U - L - U . U - U J 1 1 .
1
o
8. 8 a
Q O U c
I I
ea . 13
'Q 3 O ">
S 3 i ; 1 °
§ 1 i - a .
Il c » 3 Ô
00 | w -S < JJ D S Q -G (J "(3
8.
a. •9
•8
•8 X u
!
+ s
c 'C
1 1 1 M il 2 E
s J •g !
e-o s § o S" .9 S C -S CG a. en a o CU UJ
8. s, ^ j
a a ,-e
11 '1 E S S E » 2 +
ai
20

Anaerobic methylotrophic metabolism in environmental biotechnology
The anaerobic biodegradability of halomethanes is of priority importance for the bioremediation of contaminated groundwaters.13,40'75 Some anaerobic bacteria are able to metabolize halomethanes as a sole substrate for growth. A strictly anaerobic homoacetogenic bacterium isolate, strain MC, utilizes chloromethane as a sole carbon and energy source.92'173 Acetogens in anaerobic mixed cultures are also capable of growth on dichloromethane as a sole carbon and energy source.15'41'161 Other anaerobic bacteria have been shown to cometabolize halomethanes. Dehalogenation of tetrachloromethane, trichloromethane and bromoform in pure cultures of Methanosarcina strains was demonstrated, while generating methane from methanol, methylamines or acetate.95
Tetrachloromethane was transformed to a less chlorinated methane by pure cultures of the acetogens Acetobacterium woodii and Clostridium thermoaceticum, in a fructose-salt and glucose-salt media, respectively,34 and by an anaerobic mixed culture using a cosubstrate cocktail that included methanol.38
When a cheap and efficient organic carbon source is required, environmental biotechnology often relies on the use of methanol as a cosubstrate. In a UASB fed with dairy waste, methanol addition during the start-up aided rapid biomass granulation, and enhanced the settling velocity and specific activity of the sludge.21 Methanol can be used for sulphate reduction processes.97'109 Methanol is already largely used as an electron donor in denitrification processes.22'45'66'72'87'91'132 Furthermore, the use of methanol as a cosubstrate has been shown to significantly improve dehalogenation of a large variety of halogenated compounds.31'40-115'123
1.6 Problems associated with the anaerobic treatment of wastewaters containing methanol
Wastewaters generated by condensating processes are deficient in inorganic nutrients, that are of essential importance for biological treatment.157 Additionally, these kind of wastewaters are poor in alkalinity required for reactor pH stability.136 Thus, macro- and micronutrients as well as alkalinity have to be added in order to achieve a successful anaerobic treatment. However during the anaerobic treatment of methanolic wastewaters, the addition of trace elements and bicarbonate have been reported to cause reactor instability due to unpredictable accumulations of volatile fatty acids.77'78 Undissociated volatile fatty acids are known to cause inhibition to methanogens. For acetoclastic methanogens, the toxic concentration of undissociated acetate ranged between 0.26 to 1 mM.4'68 For thermophilic methylotrophic Methanosarcina species, a complete inhibition was found at about 9 Mm undissociated acetic acid.195
21

Problems of wastewaters containing methanol
Trace elements
Relatively much is known about the trace nutrient requirements for acetoclastic methanogens and several cases are reported in the literature where supplementation of trace elements resulted in an increased treatment efficiency.19,62,106'120'149'190 Speece and collaborators158'159'168 extensively conducted experiments on the stimulation of methane production from acetate by trace nutrients. They suggested trace nutrient supplementation in order to avoid potential process failure related to nutrient deficiency. Among the trace elements, Fe, Ni and Co were essential to achieve a high acetate conversion rate into methane.168
Less is known about the trace element requirements for a mixed population degrading methanol in wastewater. In the few studies previously conducted on nutrient stimulation of methylotrophic methanogenesis during the anaerobic wastewater treatment, no decisive conclusions could be drawn.77'78'118 Some information is available on the trace element requirements for pure cultures of methanogens grown on methanol. Calcium,107
cobalt,79'86,138 iron,79 molybdenum,138,141 nickel,30'79'138 sodium,11'124 selenium,138
vanadium141 and mineral components of yeast extract86 have been identified as essential nutrients for the growth of methanogenic bacteria on methanol. Table 7 summarizes the trace element composition and trace element requirements for Methanosarcina barken grown on methanol.116'140 Except for potassium and iron, there is a close agreement between the trace element content in Methanosarcina barken with the nutrient requirement determined experimentally. Nothing is known in the literature about the trace element requirement of methylotrophic acetogens. Nonetheless, cobalt is expected to play a key role in both trophic groups due to the involvement of cobalt containing corrinoids of methyltransferases in the initial step of methanol conversion.
Bicarbonate
Bicarbonate plays an important role in the anaerobic conversion of methanol, not only as a weak acid in the pH buffering system, but also as a required cosubstrate in the acetogenesis of methanol.81 Because bicarbonate is produced when methane is generated from methanol (reaction 1, Table 1) it will always be available to a limited extent for acetogens. According to the stoichiometry, up to one third of the methanol utilized can be converted to acetate if no exogenous C02 is added. If sufficient buffering capacity is not present, the eventual production of acetic acid by acetogens will cause sharp decreases in the pH.
Bicarbonate plays a central role in the acetogenesis of methanol. In pure cultures of
22
Anaerobic methylotrophic metabolism in environmental biotechnology
Butyribacterium methylotrophicum, the net conversion of methanol to butyrate or acetate was shown to be dependent on the level of bicarbonate.27 However in mixed culture, little is known about the effect of the bicarbonate level on the competition between methanogens and acetogens for methanol.
Table 7 - Trace element composition and nutrient requirement of Methanosarcina barkeri Fusaro grown on methanol.
Methanosarcina barkeri Fusaro
Trace element Cell content0
(mg • g"1 cell)
Experimental nutrient requirement6
Na K S P Ca Mg Fe Ni Co Mo Zn Mn Cu
9200 2500
11,000 12,000
3800 1700 2150 135 60 60
130 5 10
8756 16,131
9245 12,806
3616 1614 230 71 57 nr° nr nr nr
" FromScherer et al.uo
*FromNishio«a/ . 1 1 7
c nr, not reported.
Methanol concentration
Competition for the substrate is a fundamental factor to be considered in the predominance of one microorganism over another. Assuming Monod-type growth kinetics,104
the substrate affinity coefficient, Ks, along with the specific growth rate, iimax are of crucial importance in the prediction of which bacterial population will become predominant. Similar growth rates have been reported for acetogens and methanogens grown on methanol (Table 3). However, little is known about substrate affinity of methanogens and acetogens grown on methanol. Such information is essential for understanding competition for substrate at the low concentrations anticipated inside wastewater treatment reactors. The Ks values found in
23

Scope and structure of the thesis
the literature were 2.7 Mm for the acetogen Eubacterium limosum,44 and 0.18 for the methanogen Methanosarcina barken.*6
1.7 Scope and structure of the thesis
Methanol can be utilized by methanogens and acetogens. In wastewater treatment, the methane production from methanol is highly desired because by this conversion COD is removed, whereas only little COD removal is achieved when organic acids are formed. Moreover, the accumulation of volatile fatty acids can cause reactor instability due to the pH drop in a weakly buffered system. Thus, in view of the limited information about the anaerobic degradation of methanol by mixed cultures, the objective of this thesis was to evaluate which factors determine the predominance of one population over another. For this purpose, batch and continuous experiments were carried out using a model medium composed of methanol and defined mineral nutrients. The main factors studied were: the effect of trace elements, the reactor pH, the bicarbonate level, and the methanol concentration in the reactor.
Chapter 2 describes the effect of trace elements on the anaerobic conversion of methanol. Among the trace elements tested in batch assays, cobalt was the only one which greatly enhanced methanogenesis from methanol. Therefore, in continuous experiments in UASB reactors, the effect of cobalt addition and cobalt-deprivation on the degradation of methanol was evaluated. Chapter 3 deals with the effect of cobalt addition for each individual trophic group in an anaerobic methanol degrading consortium. For this purpose, a sludge was cultivated for one year with methanol in a mineral medium deprived of cobalt. Using specific inhibitors, kinetic parameters for acetogens and methanogens were then determined in batch assays supplemented or not supplemented with cobalt.
The anaerobic treatment of wastewaters containing methanol at low pH can be an attractive option, since the addition of expensive chemicals for supplying alkalinity will not be necessary in that case. Therefore, the possibility of continuous methane production from methanol at low pH was studied in Chapter 4, and the dominant organism responsible for the conversion of methanol to methane was characterized.
In Chapter 5, the effect of bicarbonate level and reactor substrate concentration on the competition for methanol between methanogens and acetogens were evaluated. For this purpose, several levels of external bicarbonate additions were applied to 8 UASB reactors operated continuously. Organic load rates ranged from 7.6 to 22 g COD • l"1 • d"1 to provide different methanol concentrations inside the bioreactor.
24

Anaerobic methylotrophic metabolism in environmental biotechnology
1.8 References
1. Adamse, A. D., and C. T. M. Velzeboer (1982) Features of a Clostridium, strain CV-AA1, an obligatory anaerobic bacterium producing acetic acid from methanol. Antonie van Leeuwenhoek 48:305-313.
2. Alperin, M. J., and W. S. Reeburgh (1985) Inhibition experiments on anaerobic methane oxidation. Appl. Environ. Microbiol. 50:940-945.
3. van Amstel, A. R., R. J. Swart, M. S. Krol, J. P. Beck, A. F. Bouwman, and K. W. van der Hoek (1993) Methane the other greenhouse gas. National Institute of Public Health and Environmental Protection. Report no: 481507001, Bilthoven, The Netherlands.
4. Attal, A., F. Ehlinger, J. M. Audic, and G. M. Faup (1988) pH inhibition mechanisms of acetogenic acetoclastic and hydrogenophilic populations, p. 71-78. In: E. R. Hall and P. N. Hobson (eds). Anaerobic digestion. Pergamon Press, Oxford, England.
5. Attwood, M. M., and W. Harder (1972) A rapid and specific enrichment procedure for Hyphomicrobium spp. Antonie van Leeuwenhoek 38:369-378.
6. Bache, R., and N. Pfennig (1981) Selective isolation of Acetobacterium woodii on methoxylated aromatic acids and determination of growth yields. Arch. Microbiol. 130:255-261.
7. Belitz, H.-D., and W. Grosch (1987) Fruits and fruits products, p. 578-621 In: Food Chemistry. Springer-Verlag, Berlin, Germany.
8. Bhattarai, M. D. (1992) Anaerobic degradation of phenolic compounds in upflow anaerobic sludge bed (UASB) reactors. International Institute for Hydraulic and Environmental Engineering. M.Sc. Thesis E.E.57, Delft, The Netherlands.
9. Bhatti, Z. I., K. Furukawa, and M. Fujita (1993) Treatment performance and microbial structure of a granular consortium handling methanolic waste. J. Ferment. Bioeng. 76:218-223.
10. Blaut, M., and G. Gottschalk (1985) Evidence for a chemiosmotic mechanism of ATP synthesis in methanogenic bacteria. Trends Biochem. Sei. 10:486-489.
11. Blaut, M.,V. Müller, K. Fiebig, and G. Gottschalk (1985) Sodium ions and an energized membrane required by Methanosarcina barken for the oxidation of methanol to the level of formaldehyde. J. Bacteriol. 164:95-101.
12. Bock, A., A. Prieger-Kraft, and P. Schönheit (1994) Pyruvate - a novel substrate for growth and methane formation in Methanosarcina barkeri. Arch. Microbiol. 161:33-46.
13. Bouwer, E. J., and P. L. McCarty (1983) Transformation of 1- and 2-carbon halogenated aliphatic organic compounds under methanogenic conditions. Appl. Environ. Microbiol. 45:1286-1294.
25

References
14. Bouwer, E. J., and J. P. Write (1988) Transformation of trace halogenated aliphatics in anoxic biofilm columns. J. Contam. Hydrol. 2:155-169.
15. Braus-Stromeyer, S. A., R. Haermann, A. M. Cook, and T. Leisinger (1993) Dichloromethane as the sole carbon source for an acetogenic mixed culture and isolation of a fermentative dichloromethane-degrading bacterium. Appl. Environ. Microbiol. 59:3790-3797.
16. Braun, M., and H. Stolp (1985) Degradation of methanol by a sulfate-reducing bacterium. Arch. Microbiol. 142:77-80.
17. Breznak, J. A. (1991) The genus Spowmusa. p. 2014-2021. In: A. Balows, H. G. Triiper, M. Dworkin, W. Harder, and K. Schleifer (eds). The prokaryotes. Springer-Verlag, New York, USA.
18. Cain, R. B. (1980) The uptake and catabolism of lignin-related aromatic compounds and their regulation in microorganisms, p. 21-60 In: T. K. Kirk, T. Higuchi, H. Chang (eds). Lignin biodégradation: microbiology, chemistry and potential applications. CRC Press, Boca Raton, USA.
19. Callander, I. J., and J. P. Barford (1983) Precipitation, chelation and the avaibility of metals as nutrients in anaerobic digestion. II. Application. Biotechnol. Bioeng. 25:1959-1972.
20. Carstens, R. (1992) Untersuchungen zur biologischen Abbaubarkeit von Abwässern der Polyesterproduktion in Thailand. Diplomarbeit Institut für Siedlungswasserwirtschaft und Abfalltechnik, Universität Hannover, Hannover, Germany.
21. Cayless, S. M., D. M. L. da Motta Marques, and J. N. Lester (1990) A study of the effects of methanol in start-up of UASB reactors. Biol. Wastes 31:123-135.
22. Chen, K., and Y. Lin (1993) The relationship between denitrifying bacteria and methanogenic bacteria in a mixed culture system of acclimated sludges. Water Res. 27:1749-1759.
23. Cocaign, M., E. Wilberg, and N. Lindley (1991) Sequential demethoxylation reactions during methylotrophic growth of methoxylated aromatic substrates with Eubacterium limoswn. Arch. Microbiol. 155:496-499.
24. Cord-Ruwisch, R., and B. Ollivier (1986) Interspecific hydrogen transfer during methanol degradation by Spowmusa acidovorans and hydrogenophilic anaerobes. Arch. Microbiol. 144:163-165.
25. Csonka, L. N. (1989) Physiological and genetic response of bacteria to osmotic stress. Microbiol. Rev. 53:121-147.
26. Daniel, S. L., Z. Wu, and H. L. Drake (1988) Growth of thermophilic acetogenic bacteria on methoxylated aromatic acids. FEMS Microbiol. Lett. 52:25-28.
27. Datta, R., and J. Ogeltree (1983) Methanol bioconversion by Butyribacterium methylotrophicum - Batch fermentation yield and kinetics. Biotechnol. Bioeng. 25:991-
26

Anaerobic methylotrophic metabolism in environmental biotechnology
998. 28. Davis, J. B., and H. F. Yarbrough (1966) Anaerobic oxidation of hydrocarbons by
Desulfovibrio desulfiiricans. Chem. Geol. 1:137-144. 29. Diekert, G. (1991) The acetogenic bacteria, p. 517-533. In: A. Balows, H. G. Triiper,
M. Dworkin, W. Harder, and K. Schleifer (eds). The prokaryotes. Springer-Verlag, New York, USA.
30. Diekert, G., U. Konheiser, K. Piechulla, and R. K. Thauer (1981) Nickel requirement and factor F430 content of methanogenic bacteria. J. Bacteriol. 148:459-464.
31. DiStefano, T. D., J. M. Gossett, and S. H. Zinder (1992) Hydrogen as an electron donor for dechlorination of tetrechloroethene by an anaerobic mixed culture. Appl. Environ. Microbiol. 58:3622-3629.
32. Donnelly, M. I., and S. Dagley (1980) Production of methanol from aromatic acids by Pseudomonas putida. J. Bacteriol. 142:916-924.
33. Drake, H. L. (1992) Acetogenesis and acetogenic bacteria, p. 1-15 In: J. Ledenberg (ed). Encyclopedia of Microbiology. Academic Press, San Diego, USA.
34. Egli, C , T. Tschan, R. Scholtz, A. M. Cook, and T. Leisinger (1988) Transformation of tetrachloromethane to dichloromethane and carbon dioxide by Acetobacterium woodii. Appl. Environ. Microbiol. 54:2819-2824.
35. Egli, C , S. Stromeyer, A. M. Cook, and T. Leisinger (1990) Transformation of tetra- and trichloromethane to C02 by anaerobic bacteria is a non-enzymic process. FEMS Microbiol. Lett. 68:207-212.
36. Eichler, B., and B. Schink (1984) Oxidation of primary aliphatic alcohols by Acetobacterium carbinolicum sp. nov., a homoacetogenic anaerobe. Arch. Microbiol. 140:147-152.
37. Elbert, C , D. Picard, P. Pope, and A. Rosland (1993) Methane emission from oil and gas. p. 41-61. In: A. R. van Amstel (ed). International IPCC Workshop Methane and Nitrous Oxide: methods in national emission inventories and options for control. National Institute of Public Health and Environmental Protection. Report no. 481507003, Bilthoven, The Netherlands
38. Erzmann, M. W., and H. J. Pöpel (1991) Biodegradation of tetrachloromethane under anaerobic conditions. Acta Hydrochim. Hydrobiol. 19:249-255.
39. Ferguson, T. J., and R. A. Mah (1983) Effect of H2-C02 on methanogenesis from acetate or methanol in Methanosarcina spp. Appl. Environ. Microbiol. 46:348-355.
40. Freedman, D. L., and J . M. Gossett (1989) Biological reductive dechlorination of tetrachloroethylene and trichloromethylene to ethylene under methanogenic conditions. Appl. Environ. Microbiol. 55:2144-2151.
41. Freedman, D. L., and J. M. Gossett (1991) Biodegradation of dichloromethane and its utilization as a growth substrate under methanogenic conditions. Appl. Environ.
27

References
Microbiol. 57:2847-2857. 42. Genthner, B. R. S., C. L. Davis, and M. P. Bryant (1981) Features of rumen and
sewage sludge strains of Eubacterium limosum, a methanol- and H2-C02-utilizing species. Appl. Environ. Microbiol. 42:12-19.
43. Genthner, B. R. S., and M. P. Bryant (1982) Growth of Eubacterium limosum with carbon monoxide as the energy source. Appl. Environ. Microbiol. 43:70-74.
44. Genthner, B. R. S., and M. Bryant (1987) Additional characteristics of one-carbon-compound utilization by Eubacterium limosum and Acetobacterium woodii. Appl. Environ. Microbiol. 53:471-476.
45. Germonpre, R., J. Liessens, and W. Verstraete (1991) Fluidised bed denitrification of drinking water with methanol - pilot plant experience, p. 49-52 In: H. Verachten and W. Verstraete (eds). Proceedings of the International Symposium on Environmental Biotechnology, The Royal Flemish Society of Engineers. Oostende, Belgium.
46. Gerritse, J., and J. C. Gottschal (1993) Two-membered mixed cultures of methanogenic and aerobic bacteria in 02-limited chemostats. J. Gen. Microbiol. 139:1853-1860.
47. Gibson, G. R. (1990) Physiology and ecology of the sulfate-reducing bacteria. J. Appl. Bacterid. 69:769-797.
48. Gujer, W., and A. J. B. Zehnder (1983) Conversion processes in anaerobic digestion. Wat. Sei. Tech. 15:127-167.
49. Häggblom, M. M., M. H. Bennan, A. C. Frazer, and L.Y. Young (1993) Anaerobic O-demethylation of chlorinated gauaiacols by Acetobacterium woodii and Eubacterium limosum. Biodegradation 4:107-114.
50. Harper, D. B. (1985) Halomethane from halide ion - a highly efficient fungal conversion of environmental significance. Nature 315:55-57.
51. Heijthuijsen, J. H. F. G., and T. A. Hansen. (1986) Interspecies hydrogen transfer in co-cultures of methanol-utilizing acidogens and sulfate-reducing or methanogenic bacteria. FEMS Microbiol. Lett. 38:57-64.
52. Heise, R., V. Müller, and G. Gottschalk (1989) Sodium dependence of acetate formation by the acetogenic bacterium Acetobacterium woodii. J. Bacteriol. 171:5473-5478.
53. Hippe, H., J. R. Andreesen, and G. Gottschalk (1991) The genus Clostridium (nonmedical aspects), p. 1800-1866. In: A. Balows, H. G. Triiper, M. Dworkin, W. Harder, and K. Schleifer (eds). The prokaryotes. Springer-Verlag, New York, USA.
54. Howard, P. H. (1990) Handbook of environmental fate and exposure data for organic chemicals. Vol. II Solvents. Lewis Publishers, Michigan, USA.
55. Hu, S., E. Pezacka, and H. G. Wood (1984) Acetate synthesis from carbon monoxide by Clostridium thermoaceticum: purification of thecorrinoid protein. J. Biol.
28

Anaerobic methylotrophic metabolism in environmental biotechnology
Chem. 259:8892-8897. 56. Hutten, T. J., H. C. M. Bongaerts, C. van der Drift, and G. D. Vogels (1980)
Acetate, methanol and carbon dioxide as substrate for growth of Methanosarcina barkeri. Antonie van Leeuwenhoek 46:601-610.
57. Inoue, K., S. Kageyama, K. Miki, T. Morinaga, Y Kamagata, K. Nakamura, and E. Mikami (1992) Vitamine B12 production by Acetobacterium sp. and its tetrachloromethane-resistant mutants. J. Ferment. Bioeng. 73:76-78.
58. Jarrel, K. F., and M. L. Kalmokoff (1988) Nutritional requirements of the methanogenic archaebacteria. Can. J. Microbiol. 34:557-576.
59. Jeris, J. S., and P. L. McCarty (1965) The Biochemistry of methane fermentation using C14 tracers. J. Water Pollut. Control Fed. 37:178-192.
60. de Jong, E., J. A. Field, and J. A. M. de Bont (1994) Aryl alcohols in the physiology of ligninolytic fungi. FEMS Microbiol. Rev. (in press)
61. Kaiser, J., and K. W. Hanselmann (1982) Fermentative metabolism of substituted monoaromatic compounds by bacterial community from anaerobic sediments. Arch. Microbiol. 133:185-194.
62. Kelly, C. R., and M. Switzenbaum (1984) Anaerobic treatment: temperature and nutrient effects. Agric. Wastes 10:135-154.
63. Kelly, D. P., G. Malin, and A. P. Wood (1993) Microbial transformations and biogeochemical cycling of one-carbon substrates containing sulphur, nitrogen or halogens, p. 47-63. In: J. C. Murrel and D. P. Kelly (eds). Microbial Growth on C, compounds. Proceedings of the 7th International Symposium. Intercept Ltd, Andover, England.
64. Kersten, P. J., M. Tien, B. Kalyanaraman, and T. K. Kirk (1985) The ligninase of Phanerochaete chysosporium generates cation radicals from methoxybenzenes. J. Biol. Chem. 260:2609-2612.
65. King, G. M. (1984) Utilization of hydrogen, acetate, and "noncompetitive" substrates by methanogenic bacteria in marine sediments. Geomicrobiol. J. 3:275-306.
66. Klapwijk, A., J. C. M. van der Hoeven, and G. Lettinga (1981) Biological denitrification in an upflow sludge blanket reactor. Water Res. 15:1-6.
67. Klemps, R., H. Cypionka, F. Widdel, and N. Pfennig (1985) Growth with hydrogen, and further physiological characteristics of Desulfotomaculum species. Appl. Environ. Microbiol. 143:203-208.
68. Kroeker, E. J., D. D. Sparling, and H. M. Lapp (1979) Anaerobic treatment process stability. J. Water Poll. Control Fed. 51:718-727.
69. Krone, U. E., K. Laufer, R. K. Thauer, and H. P. C. Hogenkamp (1989) Coenzyme F430 as a possible catalyst for the reductive dehalogenetion of chlorinated Cj hydrocarbon in methanogenic bacteria. Biochemistry 28:10061-10065.
70. Krzycki, J., and J. G. Zeikus (1980) Quantification of corrinoids in methanogenic
29

References
bacteria. Curr. Microbiol. 3:243-245. 71. Krzycki, J. A., R. H. Wolkin, and J. G. Zeikus (1982) Comparison of unitrophic
and mixotrophic substrate metabolism by an acetate-adapted strain of Methanosarcina barken. J. Bacterid. 149:247-254.
72. Kuenen, J. G., and L. A. Robertson (1988) Ecology of nitrification and denitrification. p. 161-218. In: J. A. Cole and S. J. Ferguson (eds). The nitrogen and sulphur cycles. Cambridge University Press, Cambridge, England.
73. Kuwahara, M. (1980) Metabolism of lignin-related compounds by bacteria, p. 127-146. In: T. K. Kirk, T. Higuchi, and H. Chang (eds). Lignin biodégradation: microbiology, chemistry and potential applications. CRC Press. Boca Raton, USA.
74. Lalitha K., and S. Krishnan (1991) Isolation of a novel rod shaped methanogen growing on methanol and 2-propanol. Arch. Microbiol. 155:402-408.
75. Leisinger, T., S. La Roche, R. Bader, M. Schmid-Appert, S. Stromeyer, and A. M. Cook (1993) Chlorinated methanes as carbon sources for aerobic and anaerobic bacteria, p. 351-363. In: J. C. Murrel and D. P. Kelly (eds). Microbial Growth on C, compounds. Proceedings of the 7th International Symposium. Intercept Ltd, Andover, England.
76. Lelieveld, J., and P. J. Crutzen (1993) Methane emission into the atmosphere: an overview, p. 17-25. In: A. R. van Amstel (ed). International IPCC Workshop Methane and Nitrous Oxide: methods in national emission inventories and options for control. National Institute of Public Health and Environmental Protection. Report no. 481507003, Bilthoven, The Netherlands
77. Lettinga, G., A. T. van der Geest, S. Hobma, and J. van der Laan (1979) Anaerobic treatment of methanolic wastes. Water Res. 13:725-737.
78. Lettinga, G., W. de Zeeuw, and E. Ouborg (1981) Anaerobic treatment of wastes containing methanol. Water Res. 15:171-182.
79. Lin, D. G., N. Nishio, T. K. Mazumder, and S. Nagai (1989) Infuence of Co2+ , Ni2+ and Fe2+ on the production of tetrapyrroles by Methanosarcina barken. Appl. Microbiol. Biotechnol. 30:196-200.
80. Liu, Y., D. R. Boone, R. Sleat, and R. Mah (1985) •anosarcina mazei LYC, a new methanogenic isolate which produces a disaggregating enzyme. Appl. Environ. Microbiol. 49:608-613.
81. Ljungdahl, L. G. (1986) The autotrophic pathway of acetate synthesis in acetogenic bacteria. Ann. Rev. Microbiol. 40:415-450.
82. Ljungdahl, L. G. (1987) Comparative biochemistry of C,-carrier. p. 121-128. In: H. W. van Verseveld and J. A. Duine (eds). Microbial Growth on Cj compounds. Proceedings of the 5th International Symposium. Martinus Nijhoff Publishers, Dordrecht, The Netherlands.
83. Lynd, L. H., and J. G. Zeikus (1983) Metabolism of H2-C02, methanol, and glucose
30

Anaerobic methylotrophic metabolism in environmental biotechnology
by Butyribacterium methylotrophicwn. J. Bacteriol. 153:1415-1423. 84. Mackie, R. I., B. A. White, and M. P. Bryant (1992) Methanogenesis, Biochemistry
p. 97-109. In: J. Ledenberg (ed). Encyclopedia of Microbiology. Academic Press, San Diego, USA.
85. Maestrojuân, G. M., and D. R. Boone (1991) Characterization of Methanosarcina barkeri MST and 227, Methanosarcina mazei S-6T, and Methanosarcina vacuolata Z-761T. Int. J. Syst. Bacteriol. 41:267-274.
86. Mah, R., M. Smith, and L. Baresi (1978) Studies on an acetate-fermenting strain of Methanosarcina. Appl. Environ. Microbiol. 35:1174-1184.
87. Matëju, V., S. Cizinskâ, J. Krejci, and T. Janoch (1992) Biological water denitrification - A review. Enzyme Microbiol. Technol. 14:170-183.
88. Mathrani, I. M., D. R. Boone, R. A. Mah, G. E. Fox, and P. P. Lau (1988) Methanohalophilus zhilinae sp. nov., an alkaliphilic, halophilic, methylotrophic methanogen. Int. J. Syst. Bacteriol. 38:139-142.
89. Mazumder, T. K., N. Nishio, S. Fukuzaki, and S. Nagai (1986) Effect of sulfur-containing compounds on growth of Methanosarcina barkeri in defined medium. Appl. Environ. Microbiol. 52:617-622.
90. Mazumder, T. K., N. Nishio, S. Fukuzaki, and S. Nagai (1987) Production of extracellular vitamin B12 compounds from methanol by Methanosarcina barkeri. Appl. Microbiol. Biotechnol. 26:511-516.
91. McCarty, P. L., L. Beck, and P. St. Amant (1969) Biological denitrification of wastewaters by addition of organic materials, p. 1271-1285. In: Proceedings of the 24th Industrial Waste Conference. Purdue University, West Lafayette, USA.
92. Meßmer, M., G. Wohlfarth, and G. Diekert (1993) Methyl chloride metabolism of strictly anaerobic, methyl chloride-utilizing homoacetogen strain MC. Arch. Microbiol. 160:383-387.
93. van der Meijden, P., C. van der Drift, and G. D. Vogels (1984) Methanol conversion in Eubacterium limosum. Arch. Microbiol. 138:360-364.
94. van der Meijden, P., H. J. Heythuysen, H. Sliepenbeek, F. P. Houwen, C. van der Drift, and G. D. Vogels (1984) Activation and inactivation of methanol: 2-mercaptoethanesulfonic acid methyltransferase from Methanosarcina barkeri. J. Bacteriol. 153:6-11.
95. Mikesell, M. D., and S. A. Boyd (1990) Dechlorination of chloroform by Methanosarcina strains. App. Environ. Microbiol. 56:1198-1201.
96. Minami, K., T. Horiyama, M. Tasaki, and Y. Tanimoto (1986) Methane production using a bio-reactor packed with pumice stone on an evaporator condensate of a kraft pulp mill. J. Ferment. Technol. 64:523-532.
97. Minami, K., Y. Tanimoto, M. Tasaki, S. Ogawwa, and K. Okamura (1988) Influence of pH on methane and sulfide production from methanol. J. Ferment.
31

References
Technol. 66:117-121. 98. Minami, K., K. Okamura, S. Ogawa, and T. Naritomi (1991) Continuous anaerobic
treatment of wastewater from a kraft pulp mill. J. Ferment. Bioeng. 71:270-274. 99. Minami, K. (1993) Methane production from rice production, p. 143-162. In: A. R.
van Amstel (ed). International IPCC Workshop Methane and Nitrous Oxide: methods in national emission inventories and options for control. National Institute of Public Health and Environmental Protection. Report no. 481507003, Bilthoven, The Netherlands.
100. Mohn, W. W., and J. M. Tiedje (1992) Microbial reductive dehalogenation. Microbiol. Rev. 56:482-507.
101. Mohr, D. H., and J. King (1985) Identification of polar organic compounds in coal-gasification condensate water by gas chromatography-mass spectrometry analysis of high-performance liquid chromatography fractions. Environ. Sei. Technol. 19:929-935.
102. Moktar, H. (1993) Thermoacidic precipitation of darkly coloured polyphenols of olive mill wastewaters. Environ. Technol. 14:495-500.
103. Moller, B., R. Oßmer, B. H. Howard, G. Gottschalk, and H. Hippe (1984) Sporomusa, a new genus of gram-negative anaerobic bacteria including Sporomusa sphaeroides spec. nov. and Sporomusa ovata spec. nov. Arch. Microbiol. 139:388-396.
104. Monod, J. (1949) The growth of bacterial cultures. Ann. Rev. Microbiol. 3:371-394. 105. Müller, V., and G. Gottschalk (1993) Na+ translocation in the course of
methanogenesis from methanol or formaldehyde, p. 155-177 In: E. P. Bakker (ed). Alkali cation transport system in prokaryotes. CRC Press, Boca Raton, USA.
106. Murray, W. D., and L. van den Berg (1981) Effects of nickel, cobalt and molybdenum on performance of methanogenic fixed-film reactors. Appl. Environ. Microbiol. 42:502-505.
107. Murray, P. A., and S. H. Zinder (1985) Nutritional requirements of Methanosarcina sp. Strain Tm-1. Appl. Environ. Microbiol. 50:49-55.
108. Nanninga, H. J., and J. C. Gottschal (1986) Anaerobic purification of waste water from potato-starch producing factory. I. Intermediary compounds and bacterial species involved. Water Res. 20:97-103.
109. Nanninga, H. J., and J. C. Gottschal (1986) Isolation of a sulfate-reducing bacterium growing with methanol. FEMS microbiol. Ecol. 38:125-130.
110. Nanninga, H. J., and J. C. Gottschal (1987) Properties of Desulfovibrio carbinolicus, sp. nov., and other sulfate-reducing bacteria isolated from an anaerobic purification plant. App. Environ. Microbiol. 53:802-809.
111. Naritomi, T., Y. Tanimoto, M. Yamaguchi, K. Okamura, and K. Minami (1993) Effect of glutamate on extracellular corrinoid production by a thermophilic, methanol-utilizing methanogens, Methanosarcina sp CHTI 55. J Ferment. Bioeng. 75:73-75.
32

Anaerobic methylotrophic metabolism in environmental biotechnology
112. Naumann, E., K. Fahlbuusch, and G. Gottschalk (1984) Presence of a trimethylamine: HS-coenzyme M methyltransferase in Methanosarcina barken. Arch. Microbiol. 138:79-83.
113. Nazina, T. N., A. E. Ivanova, L. P. Kanchaveli, and E. P. Rozanova (1988) A new sporeforming thermophilic methylotrophic sulfate-reducing bacterium, Desulfotomaculum kuznetsovii sp. nov. Mikrobiologiva 57:823-827. (Eng. transi.)
114. Ni, S., and D. R. Boone (1991) Isolation and characterization of a dimethyl suphide-degrading methanogen, Methanolobussiciliae HI350, from an oil well, characterization of M. siciliae T4/MT, and emendation of M. siciliae. Int. J. Syst. Bacteriol. 41:410-416.
115. Niels, L., and T. M. Vogel (1990) Effect of organic substrates on dechlorination of Aroclor 1242 in anaerobic sediments. Appl. Environ. Microbiol. 56:2612-2617.
116. Nishio, N., T. Kakizono, R. G. Silveira, S. Takemoto, and S. Nagai (1992) Nutrient control by the gas evolution in methanogenesis of methanol by Methanosarcina barkeri. J. Ferment. Technol. 73:481-485.
117. Nishio, N., R. G. Silveira, K. Hamato, and S. Nagai (1993) High rate methane production in a UASB reactor fed with methanol and acetate. J. Ferment. Technol. 75:309-313.
118. Norrman, J. (1983) Anaerobic treatment of a black liquor evaporator condensate from a kraft mill in three types of fixed-film reactors. Wat. Sei. Tech. 15:247-259.
119. O'Brien, J. M., R. H. Wolkin, T. T. Moench, J. B Morgan, and J. G. Zeikus (1984) Association of hydrogen metabolism with unitrophic or mixotrophic growth of Methanosarcina barkeri on carbon monoxide. J. Bacteriol. 158:373-375.
120. Oleszkiewicz, J . A., and V. K. Sharma (1990) Stimulation and inhibition of anaerobic process by heavy metals - a review. Biol. Wastes 31:45-67.
121. Oremland, R. S., R. P. Kiene, I. Mathrani, M. J. Whiticar, and D. R. Boone (1989) Description of an estuarine methylotrophic methanogen which grows on dimethyl sulphide. Appl. Environ. Microbiol. 55:994-1002.
122. Paice, M. G., I. D. Reid, R. Bourbonnais, F. S. Archibald, and L. Jurasek (1993) Manganese peroxidase, produced by Trametes versicolor during pulp bleaching, demethylates and delignifies kraft pulp. Appl. Environ. Microbiol. 59:260-265.
123. Parker, W. J., G. J. Farquhar, and E. R. Hall (1993) Removal of chlorophenolics and toxicity during high-rate anaerobic treatment of segregated kraft mill bleach plant effluents. Environ. Sei. Tech. 27:1783-1789.
124. Perski, H. J., P. Schönheit, and R. K. Thauer (1982) Sodium dependence of methane formation in methanogenic bacteria. FEBS Lett. 143:323-326.
125. Phelps, T. J., R. Conrad, and J. G. Zeikus (1985) Sulfate-dependent interspecies H2 transfer between Methanosarcina barkeri and Desulfovibrio vulgaris during coculture metabolism of acetate or methanol. Appl. Environ. Microbiol. 50:589-594.
33

References
126. Poindexter, J. S. (1991) Dimorphic prosthecate bacteria: the genera Caulobacter, Asticcacaulis, Hyphomicrobium, Pedomicrobium, Hyphomonas, and Thiodendron. p. 2176-2196. In: A. Balows, H. G. Triiper, M. Dworkin, W. Harder, and K. Schleifer (eds). The prokaryotes. Springer-Verlag, New York, USA.
127. Qatibi, A. I., V. Nivière, and J. L. Garcia (1991) Desulfovibrio alcoholovorans sp. nov., a sulfate-reducing bacterium able to grow on glycerol, 1,2- and 1,3-propanediol. Arch. Microbiol. 155:143-148.
128. Reddy, C. A. (1984) Physiology and biochemistry of lignin degradation, p. 558-571. In: M. J. Klug and C. A. Reddy (eds). Current perspectives in microbial ecology. American Society for Microbiology, Washington, D. C , USA.
129. Reeburgh, W. S., S. C. Whalen, and M. J. Alperin (1993) The role of methylotrophy in the global methane budget, p. 1-14. In: J. C. Murrel and D.P. Kelly (eds). Microbial Growth on Ct compounds. Proceedings of the 7th International Symposium. Intercept Ltd, Andover, England.
130. Reuler, W. (1990) Methan aus Chemieabwasser. Chem. Ind. 113(9):20-24. 131. Rhee, E., and R. E. Speece (1992) Maximal biodégradation rates of chloroform and
trichloroethylene in anaerobic treatment. Wat. Sei. Tech. 25:121-130. 132. Robertson, L. A., and J. G. Kuenen (1992) Nitrogen removal from water and waste.
p. 227-267. In: J. C. Fry, G. M. Gadd, R. A. Herbert, C. W. Jones, and I. A. Watson-Craik (eds). Microbial control of pollution. Cambridge University Press, Cambridge, England.
133. Rosnes, J. T., T. Torsvik, and T. Lien (1991) Spore-forming thermophilic sulfate-reducing bacteria isolated from North Sea oil field waters. Appl. Environ. Microbiol. 57:2302-2307.
134. Roulet, N. T., and E. Matthews (1993) Methane production from natural wetlands, p. 163-170 In: A. R. van Amstel (ed). International IPCC Workshop Methane and Nitrous Oxide: methods in national emission inventories and options for control. National Institute of Public Health and Environmental Protection. Report no. 481507003, Bilthoven, The Netherlands
135. Rouvière, P. E., and R. S. Wolfe (1988) Novel biochemistry of methanogenesis. J. Biol. Chem. 263:7913-7916.
136. Särner, E. (1988) Anaerobic treatment of a mixture of condensate and caustic extraction liquor from a dissolving pulp mill. Wat. Sei. Tech. 20:279-281.
137. Savage, M. D., and H. L. Drake (1986) Adaptation of the acetogen Clostridium thermoautotrophicum to minimal medium. J. Bacteriol. 165:315-318.
138. Scherer, P., and H. Sahni (1981) Effect of trace elements and vitamins on the growth of Methanosarcina barken. Acta Biotechnol. 1:57-65.
139. Scherer, P., and H. Sahm (1981) Influence of sulphur-containing compounds on the growth of Methanosarcina barkeri in a defined medium. European J. Appl. Microbiol.
34

Anaerobic methylotrophic metabolism in environmental biotechnology
Biotechnol. 12:28-35. 140. Scherer, P., H. Lippert, and G. Wolff (1983) Composition of the major elements
and trace elements of 10 methanogenic bacteria determined by inductively coupled plasma emission spectrometry. Biol. Trace Element Res. 5:149-163.
141. Scherer, P. (1989) Vanadium and molybdenum requirement for the fixation of molecular nitrogen by two Methanosarcina strains. Arch. Microbiol. 151:44-48.
142. Schink, B., and J . G. Zeikus (1980) Microbial methanol formation: a major end product of pectin metabolism. Curr. Microbiol. 4:387-390.
143. Schink, B., and N. Pfennig (1982) Fermentation of trihydroxybenzenes by Pelobacter acidigallici gen. nov. sp. nov., a new strictly anaerobic, non-sporeforming bacterium. Arch. Microbiol. 133:195-201.
144. Schink, B., and J. G. Zeikus (1982) Microbial ecology of pectin decomposition in anoxic lake sediments. J. Gen. Microbiol. 128:393-404.
145. Schink, B. (1987) Ecology of Cj-metabolizing anaerobes, p. 81-85. In: H. W. van Verseveld and J. A. Duine (eds). Microbial Growth on C compounds. Proceedings of the 5th International Symposium. Martinus Nijhoff Publishers, Dordrecht, The Netherlands.
146. Schnell, S., F. Bak, and N. Pfennig (1989) Anaerobic degradation of aniline and dehydroxybezenes by newly isolated sulfate-reducing bacteria and description of Desulfobacterium anilini. Arch. Microbiol. 152:556-563.
147. Schnellen, C. G. T. P. (1947) Onderzoekingen over de methaangisting. Ph.D. Thesis. Delft University of Technology. Delft, The Netherlands.
148. Semprini, L., G. D. Hopkins, P. L. McCarty, P. V. Roberts (1992) In-situ transformation of carbon tetrachloride and other halogenated compounds resulting from biostimulation under anoxic conditions. Environ. Sei. Technol. 26:2454-2461.
149. Shen, C. F., N. Kosaric, and R. Blaszczyk (1993) Properties of anaerobic granular sludge as affected by yeast extrade, cobalt and iron supplements. Appl. Microbiol. Biotechnol. 39:132-137.
150. Sierra-Alvarez, R. (1990) The role of natural wood constituents on anaerobic treatment. Ph.D. thesis. Wageningen Agricultural University. Wageningen, The Netherlands.
151. Silveira, R. G., Y. Nishida, N. Nishio, and S. Nagai (1990) Corrinoid production by Methanosarcina barken in a repeated fed-batch reactor with membrane module. Biotechnol. Lett. 12:721-726.
152. Silveira, R. G., T. Kakizono, S. Takemoto, N. Nishio, and S. Nagai (1991) Medium opimization by an orthogonal array design for the growth of Methanosarcina barkeri. J. Ferment. Bioeng. 72:20-25.
153. Smith, P. H., and R. A. Mah (1966) Kinetics of acetate metabolism during sludge digestion. Appl. Microbiol. 14:368-371.
35

References
154. Smith, M. R., and R. A. Mah (1978) Growth and methanogenesis by Methanosarcina strain 277 on acetate and methanol. Appl. Environ. Microbiol. 36:870-879.
155. Sowers, K. R., S. F. Baron, and J. G. Ferry (1984) Methanosarcina acetivorans sp. nov., an acetotrophic methane producing bacterium isolated from marine sediments. Appl. Environ. Microbiol. 47:971-978.
156. Sowers, K. R., and J. G. Ferry (1985) Trace metal and vitamin requirements of Methanococcoides methylutens grown with trimethylamine. Arch. Microbiol. 142:148-151.
157. Speece, R. E (1983) Anaerobic biotechnology for industrial wastewater treatment. Environ. Sei. Tech. 17:416A-427A.
158. Speece, R. E., G. F. Parkin, and D. Gallagher (1983) Nickel stimulation of anaerobic digestion. Wat. Res. 17:677-683.
159. Speece, R. E., G. F. Parkin, S. Bhattacharya, and M. Takashima (1986) Trace nutrient requirements of anaerobic digestion, p. 175-188. In: Proceedings of Water Treatment Conference: Anaerobic treatment, a grown-up technology. Amsterdam, The Netherlands.
160. Stadt man, T. C , and H. A. Barker (1951) Studies on the methane fermentation. IX. The origin of methane in the acetate and methanol fermentations by Methanosarcina. J. Bacteriol. 16:81-86.
161. Stromeyer, S. A., W. Winkenbauer, H. Kohler, A. M. Cook, and T. Leisinger (1991) Dichloromethane utilized by an anaerobic mixed culture: acetogenesis and methanogenesis. Biodegradation 2:129-137.
162. Stromeyer, S. A., K. Stumpf, A. M. Cook, and T. Leisinger (1992) Anaerobic degradation of tetrachloromethane by Acetohacterium woodii: separation of dechlorinative activities in cell extracts and roles for vitamin B12 and other factors. Biodegradation 3:113-123.
163. Stupperich, E., H. Eisinger, and B. Kräutler (1988) Diversity of corrinoids in acetogenic bacteria. J. Biochem. 172:459-464.
164. Stupperich, E., H. Eisinger, and S. Schurr (1990) Corrinoids in anaerobic bacteria. FEMS Microbiol. Rev. 87:355-360.
165. Stupperich, E., P. Aulkemeyer, and C. Eckerskorn (1992) Purification and characterization of a methanol-induced cobamide-containing protein from Sporomusa ovata. Arch. Microbiol. 158:370-373.
166. Stupperich, E., and R. Konle (1993) Corrinoid-dependent methyl transfer reactions are involved in methanol and 3,4-dimethyoxybenzoatemetabolism by Sporomusa ovata. Appl. Environ. Microbiol. 59:3110-3116.
167. Szewzyk, R., and N. Pfennig (1987) Complete oxidation of catechol by the strictly anaerobic sul fate-reducing Desulfohacteirum catecholicum sp. nov. Arch. Microbiol.
36

Anaerobic methylotrophic metabolism in environmental biotechnology
147:163-168. 168. Takashima, M., and R. E. Speece (1989) Mineral requirements for high-rate methane
fermentation of acetate at low SRT. J. Water Pollut. Control Fed. 61:1645-1650. 169. Tanimoto, Y., M. Tasaki, K. Okamura, M. Yamaguchi, and K. Minami (1989)
Screening growth inhibitors of sulfate-reducing bacteria and their effects on methane fermentation. J. Ferment. Bioeng. 68:353-359.
170. Thauer, R. K., K. Jungermann, and K. Decker (1977) Energy conservation in chemotrophic anaerobic bacteria. Bacteriol. Rev. 41:100-180.
171. Thorneloe S. A. (1993) Methane from landfills and open dumps, p. 93-113. In: A. R. van Amstel (ed). International IPCC Workshop Methane and Nitrous Oxide: methods in national emission inventories and options for control. National Institute of Public Health and Environmental Protection. Report no. 481507003, Bilthoven, The Netherlands
172. Thorneloe S. A. (1993) Methane from waste water treatment and septic sewage systems, p. 115-130. In: A. R. van Amstel (ed). International IPCC Workshop Methane and Nitrous Oxide: methods in national emission inventories and options for control. National Institute of Public Health and Enviromental Protection. Report no. 481507003, Bilthoven, The Netherlands
173. Traunecker, J., A. Preuß, and G. Diekert (1991) Isolation and characterization of a methyl chloride utilizing, strictly anaerobic bacterium. Arch. Microbiol. 156:416-421.
174. Tyler, S. C. (1991) The global methane budget, p. 7-38. In: J. E. Rogers and W. B. Whitman (eds). Microbial production and consumption of greehouse gases: Methane, nitrogen oxides, and halomethanes. American Society for Microbiology, Washington, D.C., USA.
175. Venkataramani, E. S., R. C. Ahlert, and P. Corbo (1984) Biological treatment of landfill leachates. CRC Crit. Rev. Environ. Control 14:333-376.
176. van Verseveld, H. W., and A. H. Stouthamer (1991) The genus Paracoccus. p. 2321-2334. In: A. Balows, H. G. Triiper, M. Dworkin, W. Harder, and K. Schleifer (eds). The prokaryotes. Springer-Verlag, New York, USA.
177. Versprille, A. I., R. J. Frankin, and G. R. Zoutberg (1994) BIOBEDR, A succesfull cross-breed between UASB and fluidized-bed. Wat. Sei. Tech. (in press).
178. Vogels, G. D., J. T. Keltjens, and C. van der Drift (1988) Biochemistry of methane production, p. 707-770. In: A. J. B. Zehnder (ed.). Biology of anaerobic bacteria. John Wiley, New York, USA.
179. Wackett, L. P., and C. A. Schanke (1992) Mechanisms of reductive dehalogenation by transition metal cofactors found in anaerobic bacteria, p. 329-356. In: H. Siegel, and A. Siegel (eds). Metal ions in biological systems. Vol. 28. Marcel Dekker, New York, USA.
37

References
180. Wackett, L. P., M. S. P. Logan, F. A. Blocki, and C. Bao-li (1992) A mechanism perspective on bacterial metabolism of chlorinated methanes. Biodegradation 3:19-36.
181. Ward, B. B., K. A. Kilpatrick, P. C. Novelli, and M. I. Scranton (1987) Methane oxidation and methane fluxes in the ocean surface layer and deep anoxic waters. Nature 327:226-229.
182. Weimer, P. J., and J. G. Zeikus (1978) One carbon metabolism in methanogenic bacteria. Arch. Microbiol. 119:49-57.
183. Weimer, P. J., and J. G. Zeikus (1978) Acetate metabolism in Methanosarcina barken. Arch. Microbiol. 119:175-182.
184. Wever, R. (1991) Formation of halogenated gases by natural sources, p. 277-285. In: J. E. Rogers and W. B. Whitman (eds). Microbial Production and consumption of greenhouse gases: methane, nitrogen oxides, and halomethanes. American Society for Microbiology, Washington, D.C., USA.
185. Wever, R. (1993) Source and sinks of halogenated methanes in nature, p. 35-45. In: J. C. Murrel and D. P. Kelly (eds). Microbial Growth on C, compounds. Proceedings of the 7th International Symposium. Intercept Ltd, Andover, England.
186. Whitman, W. B., T. L. Bowen, and D. R. Boone (1991) The methanogenic bacteria. p. 719-767. In: A. Balows, H. G. Triiper, M. Dworkin, W. Harder, and K. Schleifer (eds). The prokaryotes. Springer-Verlag, New York, USA.
187. Widdel, F. (1991) The genus Desulfotomaculum. p. 1792-1799. In: A. Balows, H. G. Triiper, M. Dworkin, W. Harder, and K. Schleifer (eds). The prokaryotes. Springer-Verlag, New York, USA.
188. Widdel, F., and F. Bak (1991) Gram-negative mesophilic sulfate-reducing bacteria. p. 3352-3378. In: A. Balows, H. G. Triiper, M. Dworkin, W. Harder, and K. Schleifer (eds). The prokaryotes. Springer-Verlag, New York, USA.
189. Widdel, F., and T. A. Hansen (1991) The dissimilatory sulfate- and sulphur-reducing bacteria, p. 583-624. In: A. Balows, H. G. Triiper, M. Dworkin, W. Harder, and K. Schleifer (eds). The prokaryotes. Springer-Verlag, New York, USA.
190. Williams, C. M., J. C. H. Shih, and J. W. Spears (1986) Effect of nickel on biological methane generation from a laboratory poultry waste digester. Biotechnol. Bioeng. 28:1608-1610.
191. Winner, C , and G. Gottschalk (1989) H2 and C02 production from methanol and formaldehyde by methanogenic bacterium strain Göl treated with 2-bromoethanesulfonic acid. FEMS Microbiol. Lett. 65:259-264.
192. Woese, C. R., O. Kandier, and M. L. Wheelis (1990) Towards a natural system of organisms: proposal for the domains Archaea, Bacteria and Eucarya. Proc. Natl. Acad. Sei. 87:4576-4579.
193. Wood, H. G., W. Ragsdale, and E. Pezacka (1986) The acetyl-CoA pathway of autotrophic growth. FEMS Microbiol. Rev. 39:345-362.
38

Anaerobic methylotrophic metabolism in environmental biotechnology
194. Woods, S. L., J. F. Ferguson, and M. M. Benjamin (1989) Characterization of chlorophenol and chloromethoxybenzene biodégradation during anaerobic treatment. Environ. Sei. Technol. 23:62-68.
195. Yamaguchi, M. K. Minami, Y. Tanimoto, and K. Okamura (1989) Effects of volatile fatty acids on methanogenesis of methanol and of pregrowth with methanol on acetate utilization by methanogens. J. Ferment. Bioeng. 68:428-432.
196. Yamaguchi, M., J. Hake, Y. Tanimoto, T. Naritomi, K. Okamura, and K. Minami (1991) Enzyme activity for monitoring the stability in a thermophilic anaerobic digestion of wastewater containg methanol. J. Ferment. Bioeng. 71:264-269.
197. Yancey, P. H., M. E. Clark, S. C. Hand, R. D. Bowlus, and G. N. Somero (1982) Living with water stress: evolution of osmolyte systems. Science 217:1214-1222.
198. Zehnder, A. J. B., and T. D. Brock (1979) Methane formation and methane oxidation by methanogenic bacteria. J. Bacteriol. 137:420-432.
199. Zehnder, A. J. B, K. Ingvorsen, and T. Marti (1982) Microbiology of methane bacteria, p. 45-68. In: Hughes et al. Anaerobic Digestion 1981. Elsevier Biomedical, Amsterdam, The Netherlands.
200. Zeikus, J. G., L. H. Lynd, T. E. Thompson, J. A. Krzycki, P. J. Weimer, and P. W. Hegge (1980) Isolation and characterization of a new, methylotrophic, acidogenic anaerobe, the Marburg strain. Curr. Microbiol. 3:381-386.
201. Zeikus, J. G., R. Kerby, and J. A. Krzycki (1985) Single-carbon chemistry of acetogenic and methanogenic bacteria. Science 227:1167-1173.
202. Zellner, G., K. Bleicher, E. Braun, H. Kneifel, B. J. Tindall, E. Conway de Macario, and J. Winter (1989) Characterization of a new mesophilic, secondary alcohol-utilizing methanogen, Methanobacterium palustre spec. nov. from a peat bog. Arch. Microbiol 151:1-9.
203. Zumft, W. G. (1991) The denitrifying prokaryotes. p. 554-582. In: A. Balows, H. G. Triiper, M. Dworkin, W. Harder, and K. Schleifer (eds). The prokaryotes. Springer-Verlag, New York, USA.
39

Effect of cobalt on the anaerobic degradation of methanol
Abstract
The effect of trace elements on the methanogenesis from methanol and acetate was studied utilizing granular sludge obtained from an anaerobic wastewater treatment plant. The methanogenic activity from methanol was dramatically stimulated by the addition of a cocktail of trace elements in the basal medium. When trace elements were supplied individually, cobalt greatly stimulated methanogenesis which equalled the stimulation observed with the complete trace element mixture. No remarkable influence of any trace element was observed when acetate was used as the substrate. Two UASB (Upflow Anaerobic Sludge Blanket) reactors were operated with and without supplementation of cobalt. Cobalt greatly stimulated both acetogenesis in the initial operational phase and later methanogenesis. The cobalt sufficient column provided almost 3 times the methane productivity compared to the cobalt deprived column. At an organic loading rate of 8 g COD • l"1 • d"1, 87% of the COD was converted to methane in the cobalt sufficient column. Under low cobalt concentration, methanogens compete better for cobalt than acetogens.
Published as: Florencio, L., P. Jenicek, J. A. Field and G. Lettinga (1993) Effect of cobalt on the anaerobic degradation of methanol. J. Ferment. Bioeng. 75:368-374.
41

Introduction
2.1 Introduction
Methanol is an important component of certain chemical industry wastewaters and can be the main pollutant in pulp and paper mill kraft black liquor evaporator condensates.20'24
In kraft condensates, the methanol may account for up 87% of the total chemical oxygen demand (COD), with concentration of methanol in the range of 1.5 to 24.5 g • l'1.21 These kind of wastewater originating from condensate process are lacking in essential inorganic nutrients indispensable for biological treatment. In order to achieve an adequate level of treatment, not only nitrogen and phosphate but trace nutrients will also be required.29
The microorganisms involved in methanol degradation are not the same as those encountered in the degradation of most industrial wastewaters which contain volatile fatty acids, carbohydrates, proteins or fats, where acetoclastic methanogens play a critical role at the end of the food chain. The anaerobic degradation of methanol is unique. Methanol can be converted by several pathways, as is shown in Figure 1. It can be directly converted into methane by methylotrophic methanogens.23'28'33 The transformation of methanol to acetate by acetogens is also possible,1'19'37 and this acetate can be converted into methane by acetoclastic methanogens.6'7'17 Another possibility is the conversion of methanol to H2 and C02 .4 '5 The H2/C02 can be converted to methane and acetate by autotrophic methanogens and acetogens, respectively.3,34 The formation of acetate is limited by the availability of inorganic carbon. In an anaerobic mixed culture, the maximum acetate production will be equal to 50% of the methane production if no exogenous inorganic carbon (e.g. HC03") is added.
Many trace metals are essential for the growth of microorganisms. They are important components of the enzymes that drive numerous anaerobic reactions.9'15 Growth of Methanosarcina barken on methanol is dependent on the trace elements such as cobalt, nickel, molybdenum and selenium.25 Mineral components of yeast extract were also found to be stimulatory17 and a medium optimization method was developed to enhance methane production rate or the cell growth rate in a fed-batch culture.23 For growth of Methanobacterium thermoautotrophicum on H2 and C02 as sole energy and carbon source, the requirement for nickel was found to be higher than for cobalt and molybdenum.27 The growth of Methanococcus vannielii on formate was markedly stimulated by selenium and tungsten.10 For growth of acetogens, several trace elements are essential, namely iron, cobalt, molybdenum, nickel, selenium, and tungsten.15
Little is known about the effect of trace elements on the stimulation of anaerobic treatment of wastewater which predominantly contains methanol. Norrman24 could not detect any obvious improvement of the treatment efficiency of kraft evaporator condensate
42

Anaerobic degradation of methanol
CH3OH
CO
H2/co2
CH 4 /C0 2
CH3COOH
Figure 1 - Possible pathways of methane formation from methanol.
wastewater with addition of nutrients, including trace elements. Lettinga et a/.,13'14 studying the effect of environmental factors on the anaerobic treatment of methanol, observed that the addition of trace nutrients was often accompanied by reactor instability due to accumulation of volatile fatty acids (VFA). The purpose of this paper was to determine which trace nutrient, if any, would be of critical importance for the anaerobic degradation of methanolic wastewaters. A model medium composed of methanol and defined mineral nutrients was used.
2.2 Materials and methods
Biomass
The methanogenic granular sludge used in these experiments was obtained from a full scale UASB reactor treating alcohol distillery wastewater of Nedalco at Bergen op Zoom, The Netherlands. The sludge was elutriated to remove the fines and stored at 4°C under nitrogen gas. Unless otherwise stated, the sludge was pre-acclimated during one week to methanol in order to minimize the lag phase. Prior to the activity assays, the sludge was rinsed with demineralized water. The density of the sludge was 1059 kg • l"1.
Total suspended solids (TSS) and the volatile suspended solids (VSS) content of the sludge were 10.5% and 9.14%, respectively, determined according to Standard Methods.2
43
Materials and methods
The maximum specific methanogenic activities of Nedalco sludge for methanol, acetate and VFA mixture (100:100:100 g acetate:propionate:butyrate per kilogram) was 808, 603 and 542 g COD-CH4 • g"1 VSS • d"1, respectively, after acclimatization. Acetate and VFA mixture were neutralized with NaOH at pH 6.8 prior to utilization.
Basal Medium and Chemicals
The inorganic macronutrients used in all experiments contained (in milligrams per litre of basal medium) NH4C1 (280), K2HP04 • 3H20 (330), MgS04 • 7H20 (100) and CaCl2 • 2H20 (10). The medium was made up in demineralized water. Unless otherwise stated, one millilitre of trace nutrient (cocktail) solution, presented in Table 1, was added per liter of medium. To ensure pH stability, when methanol was used as a substrate, 2.52 g NaHC03 was added per liter of the basal medium.
Table 1 - Composition of trace nutrient solution.
Compound added* Nutrient Concentration11 (mg • 1"')
as compound as nutrient
FeCl2 • 4H20 H3BO3 ZnCl2
MnCl2 • 4H20 CuCl2 • 2H20 (NH4)6Mo7024 • 4H20 AICI3 • 6H20 CoCl2 • 6H20 NiCl2 • 6H20 Na^eOj • 5H20
Fe B Zn Mn Cu Mo Al Co Ni Se
2000 50 50
500 38 50 90
2000 142 164
562 9
24 139 14 4
10 495 35 49
* The trace nutrient solution also contained: EDTA (1000 mg), resazurin (200 mg), and 36% HCl (1 ml) was added per liter solution. 1 ml of trace nutrient solution was supplied per liter of medium.
All chemicals were of analytical grade and were supplied by E. Merck AG, Darmstadt, Germany, except for resazurin, which was purchased from Fluka Chemie AG, Buchs, Switzerland. The gases were supplied by Hoekloos, Schiedam, The Netherlands.
44

Anaerobic degradation of methanol
Anaerobic Activity Test Assay
The maximum specific methanogenic activity tests were performed in 0.6 1 glass serum bottles sealed with a 4 mm rubber septum kept in place by a screw-cap. Before closing, the serum bottles were flushed with 70% nitrogen and 30% carbon dioxide mixture. Each serum bottle contained 1.5 g VSS • l"1 of methanogenic granular sludge, 500 ml of the basal medium and 4.0 g COD • l"1 (methanol or neutralized VFA or acetate). Incubation was done in a temperature controlled room at 30 ± 2 °C. Methane production was monitored during the assays by using a sodium hydroxide solution displacement system. The strength of the solution (3 % w/w) was sufficient to remove the carbon dioxide from the biogas. All experiments were conducted in duplicate or triplicate.
Experimental Set-up
In order to evaluate the importance of trace elements on the specific methanogenic activity, three kinds of experiments were carried out with varying levels of trace elements. In all variants, inorganic macronutrients and sodium bicarbonate were present, as outlined previously. A control, where all trace elements were present (Table 1), was included in all experiments. The experiments are outlined as follows:
i) The effect of trace element cocktail concentration. All trace elements were present in the same proportion as outlined in Table 1. The concentration is expressed relative to the concentration in the basal medium. Methanol was used as substrate.
ii) Presence of individual trace elements. Individual trace elements were supplied at the concentration present in the basal medium. All other trace nutrients were excluded from the medium. Methanol and acetate were used as substrates in different experiments.
iii) Absence of individual trace elements. All trace elements were added at the concentration in the basal medium except for one individual trace element which was excluded. Methanol was used as substrate.
Continuous Experiments
The experiments were performed in a temperature controlled room at 30 ± 2 °C. The Plexiglass cylindrical UASB reactors had a working volume of 2.5 1. No mechanical mixing was applied in the reactors. The reactors were inoculated with 20 g VSS • l"1
45

Materials and methods
anaerobic Nedalco granular sludge. The reactors were started-up with methanol concentrations ranging from 4.8 to 5.2 g COD • l"1. The influent flow rate was adjusted to provide a loading ranging from 8 to 9 g COD • l"1 • d"1. The influent solution contained sufficient macronutrients (N, P, and S) and trace elements required for bacterial growth as outlined in Basal Media and Chemicals with the exception of cobalt. Reactor A was deprived of cobalt addition while reactor B was supplied with cobalt at the same concentration present in the basal medium. The biogas was passed through a concentrated sodium hydroxide solution and then through a column filled with soda lime pellets to remove carbon dioxide. Methane production was monitored with a wet gasmeter. Figure 2 illustrates the experimental set-up.
Figure 2 - Schematic diagram of the UASB reactor used in this study, a: COD influent; b: nutrients; c: nitrogen; d: sampling point; e: sludge bed; f: effluent; g: gas sampling point; h: NaOH; i: lime soda pellets; j : gas meter.
46

Anaerobic degradation of methanol
Analyses
The pH was determined immediately after sampling with a Knick 511 pH-meter (Berlin, Germany) and a Schott Geräte N61 double electrode (Hofheim, Germany).
Samples for methanol and VFA were analyzed after centrifugation at 17000 g for 5 min. When immediate analyse was not possible, the samples were stored at 4°C with the addition of 3% formic acid. Methanol was determined by gas chromatography using a Packard Becker model 417 (Delft, The Netherlands) equipped with a 6 m x 2 mm glass column packed with Supelco port (Bellefont, USA), 100-120 mesh, coated with 10% Fluorad FC 431 (3M, St. Paul, USA). Flow rate of the carrier gas (nitrogen saturated with formic acid) was 30 ml • min"1, and column pressure was ± 3 bar. The column temperature was 80 °C, the injection port and the detector were at 220 °C and 280 °C, respectively. VFA were analyzed in the same way as methanol except that the temperatures of the column, the injector port and the flame ionization detector were 130 °C, 210 °C, and 230 °C, respectively.
The biogas composition (CH4 and C02) was determined in 100 /il gas samples removed from the gas sampling port immediately after sampling, using a Packard Becker gas Chromatograph model 433 (Delft, The Netherlands). The gas Chromatograph was equipped with two columns connected in parallel (split 1:1) - 1.5 m x 1/8" teflon, packed with chromosorb 108, 60-80 mesh (Johns Manville, USA) and a 1.2 m x 1/8" stainless steel, packed with molecular sieve 5A (60-80 mesh) (Chrompack, Bergen op Zoom, The Netherlands). Helium was used as a carrier gas (45 ml • min"1). The oven, detector and injection temperatures were 40 °C, 125 °C and 110 °C, respectively.
The trace element content of the sludge was determinated by flame or graphite tube atomizer in an Atomic Absorption Spectrometer (AAS), Varian model SpectrAA 300 (Springvale, Australia). The burning gas for the flame was a mixture of ainacetylene (2:1). Argon was the flow gas for the furnace. The extraction of trace elements from the sludge was done according to Lustenhouwer and Hin.16
2.3 Results
Effect of combined trace element cocktail
The specific methanogenic activity was greatly stimulated by adding increasing levels of basal trace element cocktail (data not shown). The relative activity without addition of
47
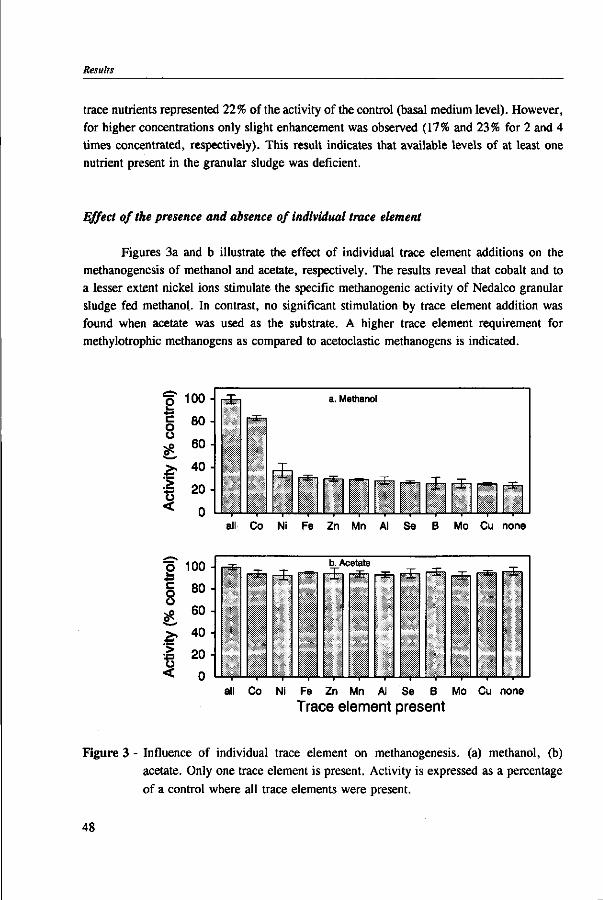
Results
trace nutrients represented 22% of the activity of the control (basal medium level). However, for higher concentrations only slight enhancement was observed (17% and 23% for 2 and 4 times concentrated, respectively). This result indicates that available levels of at least one nutrient present in the granular sludge was deficient.
Effect of the presence and absence of individual trace element
Figures 3a and b illustrate the effect of individual trace element additions on the methanogenesis of methanol and acetate, respectively. The results reveal that cobalt and to a lesser extent nickel ions stimulate the specific methanogenic activity of Nedalco granular sludge fed methanol. In contrast, no significant stimulation by trace element addition was found when acetate was used as the substrate. A higher trace element requirement for methylotrophic methanogens as compared to acetoclastic methanogens is indicated.
Ö 100 c o o #
£• > '•s <
o i—
+* c o ü <£ * > 'T) <
80 -
60-
40-
20-
0
100-
80-
60-
40 -
20 -
n
a. Methanol
ï -5E-1 r f Sir^i
" ' i ' " " i ' '—i ' ' r ' ' i ' ' i ' ' i"" ' i' ' ' i ' ' i all Co Ni Fe Zn Mn Al Se B Mo Cu none
i b. Acetate i l f - i i r * i r ^ i F * ] E-S- * " *
all Co NI Fe Zn Mn AI Se B Mo Cu none
Trace element present
Figure 3 - Influence of individual trace element on methanogenesis. (a) methanol, (b) acetate. Only one trace element is present. Activity is expressed as a percentage of a control where all trace elements were present.
48
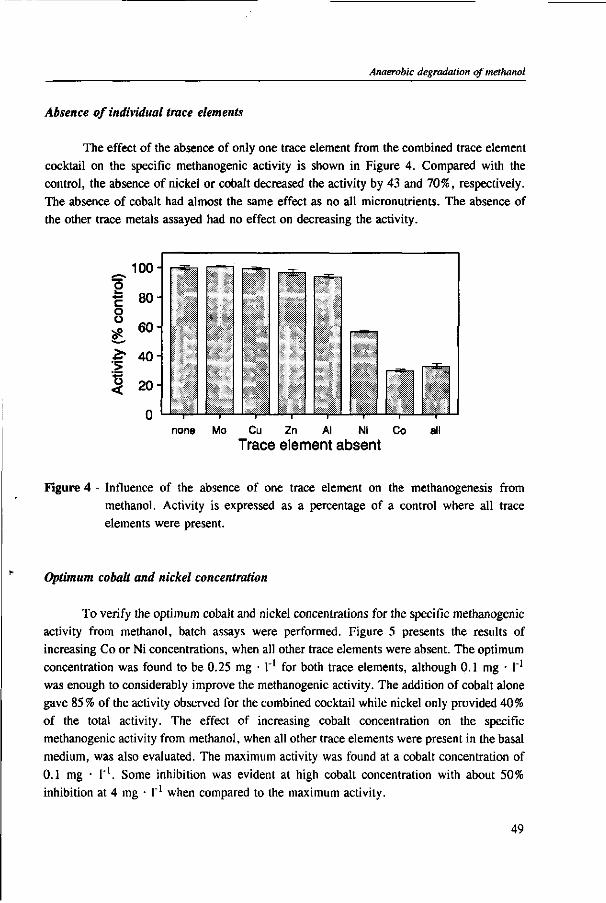
Anaerobic degradation of methanol
Absence of individual trace elements
The effect of the absence of only one trace element from the combined trace element cocktail on the specific methanogenic activity is shown in Figure 4. Compared with the control, the absence of nickel or cobalt decreased the activity by 43 and 70%, respectively. The absence of cobalt had almost the same effect as no all micronutrients. The absence of the other trace metals assayed had no effect on decreasing the activity.
'"I ' — l — I ' ' I—•—'—'l'"" I ' — r
none Mo Cu Zn AI Ni Co Trace element absent
Figure 4 - Influence of the absence of one trace element on the methanogenesis from methanol. Activity is expressed as a percentage of a control where all trace elements were present.
Optimum cobalt and nickel concentration
To verify the optimum cobalt and nickel concentrations for the specific methanogenic activity from methanol, batch assays were performed. Figure 5 presents the results of increasing Co or Ni concentrations, when all other trace elements were absent. The optimum concentration was found to be 0.25 mg • l"1 for both trace elements, although 0.1 mg • l"1
was enough to considerably improve the methanogenic activity. The addition of cobalt alone gave 85% of the activity observed for the combined cocktail while nickel only provided 40% of the total activity. The effect of increasing cobalt concentration on the specific methanogenic activity from methanol, when all other trace elements were present in the basal medium, was also evaluated. The maximum activity was found at a cobalt concentration of 0.1 mg • l"1. Some inhibition was evident at high cobalt concentration with about 50% inhibition at 4 mg • l"1 when compared to the maximum activity.
49
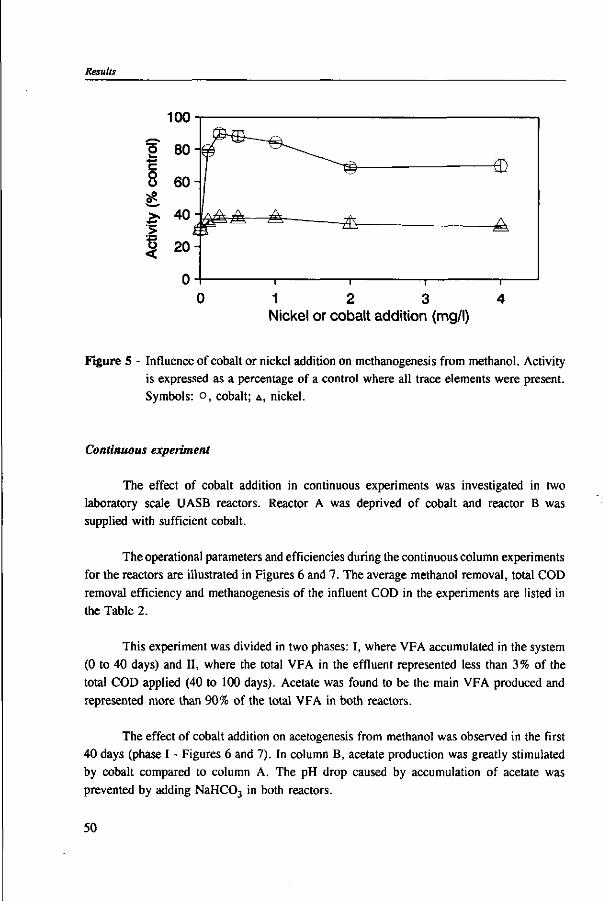
Results
100
T r 1 2 3 Nickel or cobalt addition (mg/l)
Figure 5 - Influence of cobalt or nickel addition on methanogenesis from methanol. Activity is expressed as a percentage of a control where all trace elements were present. Symbols: o, cobalt; A, nickel.
Continuous experiment
The effect of cobalt addition in continuous experiments was investigated in two laboratory scale UASB reactors. Reactor A was deprived of cobalt and reactor B was supplied with sufficient cobalt.
The operational parameters and efficiencies during the continuous column experiments for the reactors are illustrated in Figures 6 and 7. The average methanol removal, total COD removal efficiency and methanogenesis of the influent COD in the experiments are listed in the Table 2.
This experiment was divided in two phases: I, where VFA accumulated in the system (0 to 40 days) and II, where the total VFA in the effluent represented less than 3% of the total COD applied (40 to 100 days). Acetate was found to be the main VFA produced and represented more than 90% of the total VFA in both reactors.
The effect of cobalt addition on acetogenesis from methanol was observed in the first 40 days (phase I - Figures 6 and 7). In column B, acetate production was greatly stimulated by cobalt compared to column A. The pH drop caused by accumulation of acetate was prevented by adding NaHC03 in both reactors.
50
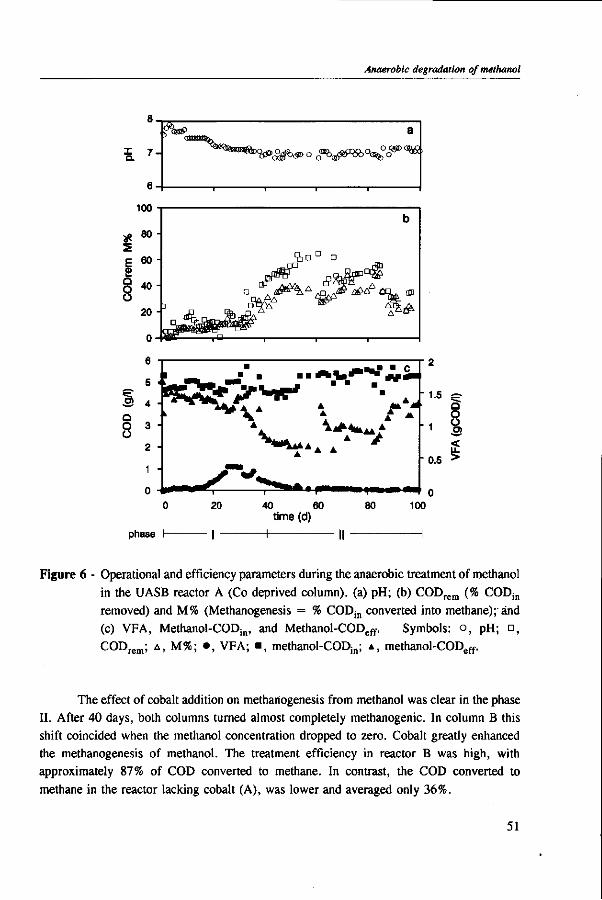
Anaerobic degradation of methanol
ac î. 7^
ossaas?».
100
! ^ ™ ^ P ( ^ > ^ o vbrfPtbo^s*m •
time (d)
phase +
Figure 6 - Operational and efficiency parameters during the anaerobic treatment of methanol in the UASB reactor A (Co deprived column), (a) pH; (b) CODrem (% CODin
removed) and M% (Methanogenesis = % CODin converted into methane); and (c) VFA, Methanol-CODin, and Methanol-CODeff. Symbols: o, pH; • , CODrem; A, M%; • , VFA; • , methanol-CODin; A, methanol-CODeff.
The effect of cobalt addition on methanogenesis from methanol was clear in the phase II. After 40 days, both columns turned almost completely methanogenic. In column B this shift coincided when the methanol concentration dropped to zero. Cobalt greatly enhanced the methanogenesis of methanol. The treatment efficiency in reactor B was high, with approximately 87% of COD converted to methane. In contrast, the COD converted to methane in the reactor lacking cobalt (A), was lower and averaged only 36%.
51
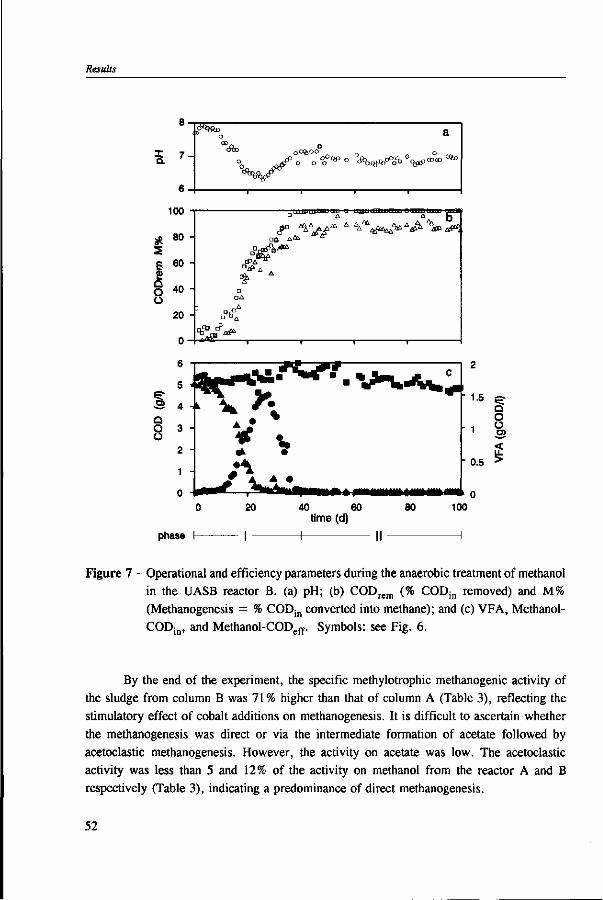
Results
I a. 7-
"%> a o o
6
100
80
60
40
20
0
6
5
4
3
2
1
0
?^~ oOèOO
o o °o<«' ° > hctftr-o £°oV
•F ^V /" ^ » " i A ' V ^
tPA °^A
1 o <
A A g
: ¥» • A,.
1.5 ç O O
1 ü
1 O)
phase h
20
I
40 60
time (d)
H II
80 100
Figure 7 - Operational and efficiency parameters during the anaerobic treatment of methanol in the UASB reactor B. (a) pH; (b) CODrem (% CODin removed) and M% (Methanogenesis = % CODin converted into methane); and (c) VFA, Methanol-CODin, and Methanol-CODeff. Symbols: see Fig. 6.
By the end of the experiment, the specific methylotrophic methanogenic activity of the sludge from column B was 71% higher than that of column A (Table 3), reflecting the stimulatory effect of cobalt additions on methanogenesis. It is difficult to ascertain whether the methanogenesis was direct or via the intermediate formation of acetate followed by acetoclastic methanogenesis. However, the activity on acetate was low. The acetoclastic activity was less than 5 and 12% of the activity on methanol from the reactor A and B respectively (Table 3), indicating a predominance of direct methanogenesis.
52
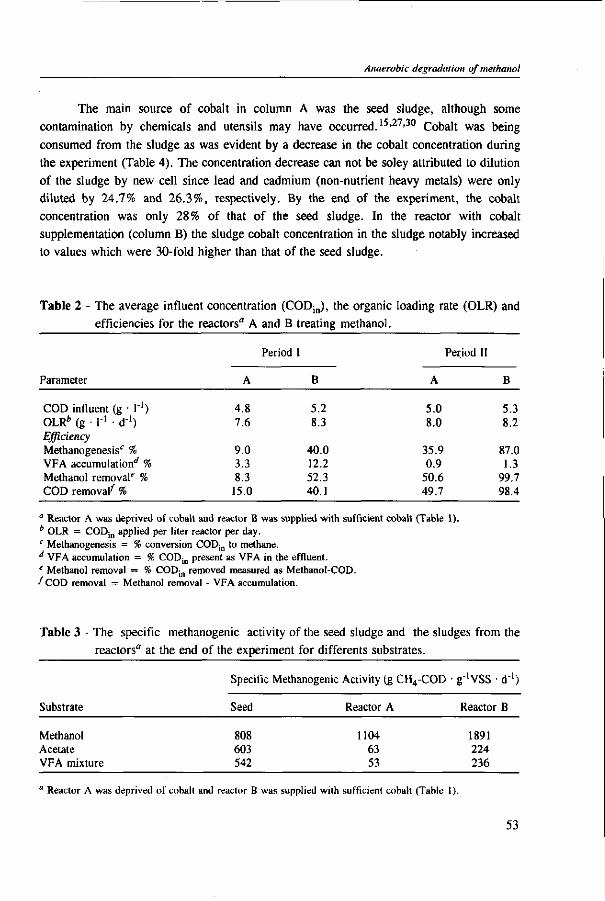
Anaerobic degradation of methanol
The main source of cobalt in column A was the seed sludge, although some contamination by chemicals and utensils may have occurred.15,27'30 Cobalt was being consumed from the sludge as was evident by a decrease in the cobalt concentration during the experiment (Table 4). The concentration decrease can not be soley attributed to dilution of the sludge by new cell since lead and cadmium (non-nutrient heavy metals) were only diluted by 24.7% and 26.3%, respectively. By the end of the experiment, the cobalt concentration was only 28% of that of the seed sludge. In the reactor with cobalt supplementation (column B) the sludge cobalt concentration in the sludge notably increased to values which were 30-fold higher than that of the seed sludge.
Table 2 - The average influent concentration (CODin), the organic loading rate (OLR) and efficiencies for the reactors" A and B treating methanol.
Parameter
COD influent (g • l ' ) OLR* (g • l"1 • d1) Efficiency Methanogenesis'' % VFA accumulation^ % Methanol removal^ % COD removal %
A
4.8 7.6
9.0 3.3 8.3 15.0
Period I
B
5.2 8.3
40.0 12.2 52.3 40.1
Period II
A
5.0 8.0
35.9 0.9
50.6 49.7
B
5.3 8.2
87.0 1.3
99.7 98.4
" Reactor A was deprived of cobalt and reactor B was supplied with sufficient cobalt (Table 1). * OLR = CODin applied per liter reactor per day. c Methanogenesis = % conversion CODin to methane. d VFA accumulation = % CODin present as VFA in the effluent. e Methanol removal = % CODin removed measured as Methanol-COD. J COD removal = Methanol removal - VFA accumulation.
Table 3 - The specific methanogenic activity of the seed sludge and the sludges from the reactors" at the end of the experiment for différents substrates.
Specific Methanogenic Activity (g CH4-COD • g'VSS • d1)
Substrate Seed Reactor A Reactor B
Methanol 808 1104 1891 Acetate 603 63 224 VFA mixture 542 53 236
" Reactor A was deprived of cobalt and reactor B was supplied with sufficient cobalt (Table 1).
53
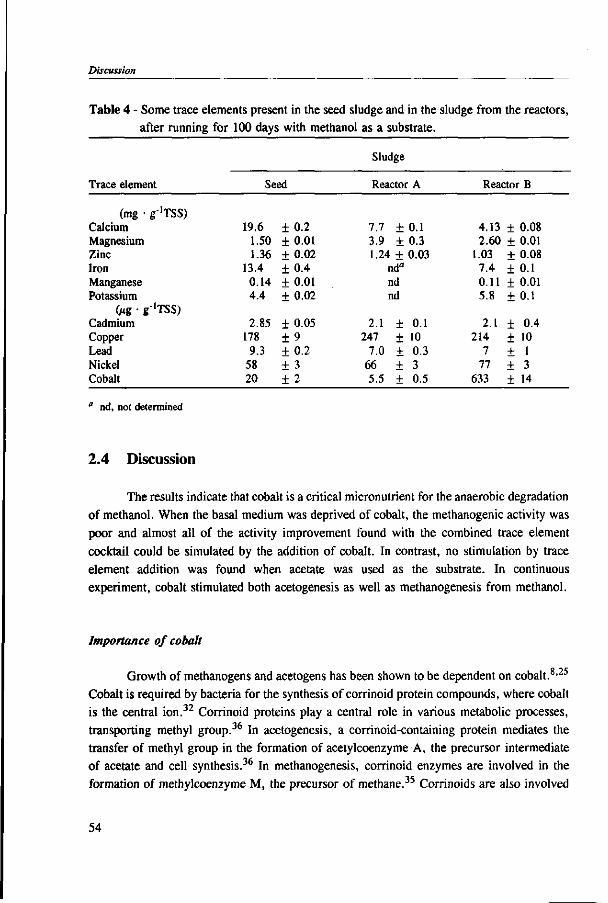
Discussion
Table 4 - Some trace elements present in the seed sludge and in the sludge from the reactors, after running for 100 days with methanol as a substrate.
Trace element
(mg • g~ Calcium Magnesium Zinc Iron Manganese Potassium
(Mg • g"' Cadmium Copper Lead Nickel Cobalt
'TSS)
rss)
Seed
19.6 1.50 1.36
13.4 0.14 4.4
2.85 178 9.3
58 20
± 0.2 ± 0.01 + 0.02 + 0.4 ± 0.01 + 0.02
+ 0.05 ± 9 + 0.2 + 3 ± 2
Sludge
Reactor A
7.7 ± 0.1 3.9 ± 0.3 1.24 ± 0.03
nd° nd nd
2.1 + 0.1 247 ± 10
7.0 ± 0.3 6 6 + 3 5.5 ± 0.5
Reactor B
4.13 + 0.08 2.60 ± 0.01
1.03 + 0.08 7.4 + 0.1 0.11 ± 0.01 5.8 + 0.1
2.1 ± 0.4 214 ± 10
7 + 1 7 7 + 3
633 ± 14
" nd, not determined
2.4 Discussion
The results indicate that cobalt is a critical micronutrient for the anaerobic degradation of methanol. When the basal medium was deprived of cobalt, the methanogenic activity was poor and almost all of the activity improvement found with the combined trace element cocktail could be simulated by the addition of cobalt. In contrast, no stimulation by trace element addition was found when acetate was used as the substrate. In continuous experiment, cobalt stimulated both acetogenesis as well as methanogenesis from methanol.
Importance of cobalt
Growth of methanogens and acetogens has been shown to be dependent on cobalt.8'25
Cobalt is required by bacteria for the synthesis of corrinoid protein compounds, where cobalt is the central ion.32 Corrinoid proteins play a central role in various metabolic processes, transporting methyl group.36 In acetogenesis, a corrinoid-containing protein mediates the transfer of methyl group in the formation of acetylcoenzyme A, the precursor intermediate of acetate and cell synthesis.36 In methanogenesis, corrinoid enzymes are involved in the formation of methylcoenzyme M, the precursor of methane.35 Corrinoids are also involved
54

Anaerobic degradation of methanol
in methanogenic cell synthesis.36
The corrinoid content in anaerobic bacteria varies greatly among species and it is also dependent on the substrate utilized.12 Among methanogens, Methanosarcina barkeri contains the highest concentration, and when cultivated on methanol, they contain about three times as much corrinoids as acetate grown cells.12 Considering that 98% of the total cobalt content in the cells is found in corrinoids,31 this may very well be the reason that cobalt dramatically stimulated methylotrophic methanogens while no apparent effect is evident for acetoclastic methanogenesis. The corrinoid content of Methanosarcina barkeri cells ranges from 2.17 to 5.8 mg • g"1 dry cell.12'18 Among acetogens, Butyribacterium methylotrophicum, when cultivated on methanol, contains around 8.0 mg corrinoid • g"1 dry cell, that is more than 22-fold higher than when cultivated on glucose.37 Recently, a remarkable high content of corrinoid, 11 mg • g"1 dry cell, was reported for Acetobacterium sp.8
Role of cobalt on population dynamics
The results show that cobalt plays an important role on the population dynamics. In medium rich or deprived of cobalt, competition for cobalt between acetogens and methylotrophic methanogens is evident.
The results from the continuous experiment show that cobalt deprivation decreased the development of acetogenic bacteria as compared to cobalt sufficient conditions. Therefore, it appears that under limited cobalt, methylotrophic methanogens are better scavengers for cobalt or alternatively they have a lower cobalt optimum.
Even in the cobalt sufficient conditions, there was eventually a shift in population from acetogenic to mostly direct methylotrophic methanogenesis. In both reactors, methanogenesis was mostly carried out by methylotrophic bacteria, thus in the final consortia which developed, acetogens and acetoclastic bacteria only played a minor role. If methanogenesis from methanol were via the intermediate formation of acetate, the effect of cobalt would be different, as shown in Figure 3. The shift in the predominant population, from acetogenic to methanogenic in cobalt rich conditions occured when the reactor methanol concentration decreased to nearly zero (0 to 27 mg COD • l"1 after day 27, Figure 7). Perhaps indicating the moment when methanogens were able to outcompete acetogens for methanol, due to a higher affinity for methanol. In relation to methanol competition between trophic groups, little information is available in the literature on the nmax and Ks values for acetogens and methanogens grown on methanol. Thus, kinetics studies are still needed to better understand the competition between these trophic groups at low methanol concentrations.
55

Discussion
The initial dominance of acetogens could also reflect a higher initial biomass concentration of acetogens in the seed sludge. It is well known that many acetogens are spore formers, like Butyribacterium methylotrophicum,31 Sporomusa ssp22 and Clostridium CV-AAI,1 which indicates an initial advantage over methanogens. Thus, the possibility that there were plenty of acetogen spores in anaerobic sludges must be considered.
Cobalt as a regulating agent in anaerobic wastewater treatment
Cobalt can be used to regulate the anaerobic wastewater treatment of methanolic wastewaters. The reactor instability, caused by a build-up of acetate produced by acetogens, can be prevented using low levels of cobalt in the reactor feed during start-up. Lettinga et a/.13 observed that the addition of trace elements in the feed solution caused acetate accumulation in the reactor. Alternatively, high levels of NaHC03 may be applied to prevent pH drop by the initial acetate accumulation. Even under cobalt sufficient conditions, a shift from acetogenesis to direct methanogenesis occurs if neutral pH conditions are maintained. Once direct methanogenesis has dominated, then cobalt does in fact dramatically stimulate the formation of methane. The cobalt sufficient column provided almost 3 times the methane productivity compared to the cobalt deprived column. The specific methanogenic activity of the sludge was about 2 times higher than that of the cobalt deprived column.
The cobalt required by methanogenic bacteria can be estimated from the cobalt content of Methanosarcina barkeri cells26 and the specific cell yield during the methanogenic phase of the cobalt sufficient column experiment (0.0925 g VSS • g"1 COD-CH4). According to this calculation, 2.7 to 8.5 mg Co will be required per kg COD converted to methane. This agrees closely with the experimentally determined value of 5.1 mg Co • kg"1 COD methanol consumed.23
2.5 Acknowledgements
We would like to thank Adrie Veeken and Richard Tichy for their help with atomic absorption spectrophotometry, and Ronaldo Petruy for conducting some of the experimental work. We also wish to thank Johannes van der Laan and Martin de Wit for the technical assistance. Financial aid from the Brazilian Government, Ministry of Education-CAPES, is gratefully acknowledged.
56

Anaerobic degradation of methanol
2.6 References
1. Adamse, A.,D., and C. T. M. Velzeboer (1982) Features of a Clostridium, strain CV-AA1, an obligatory anaerobic bacterium producing acetic acid from methanol. Antonie van Leeuwenhoek 48:305-313.
2. American Public Health Association (1985) Standard methods for examination of water and wastewater, 16th ed. American Public Health Association. Washington, D.C., USA.
3. Brawn, M., F. Mayer, and G. Gottschalk (1981) Clostridium aceticum (Wieringa), a microorganism producing acetic acid from molecular hydrogen and carbon dioxide. Arch. Microbiol. 128:288-293.
4. Cord-Ruwisch, R., and B. Ollivier (1986) Interspecific hydrogen transfer during methanol degradation by Sporomusa acidovorans and hydrogenophilic anaerobes. Arch. Microbiol. 144:163-165.
5. Cord-Ruwisch, R., H. Seitz, and R. Conrad (1988) The capacity of hydrogenotrophic anaerobic bacteria to compete for trace of hydrogen depends on the redox potential of the terminal electron acceptor. Arch. Microbiol. 149:350-357.
6. Fathepure, B. Z. (1987) Factors affecting the methanogenic activity of Methanothrix soenhgenii VNBF. Appl. Environ. Microbiol. 53:2978-2982.
7. Huser, B. A., K. Wuhrmann, and A. J. B. Zehnder (1982) Methanothrix soehngenii gen. nov. sp. nov., a new acetotrophic non-hydrogen oxidizing methane bacterium. Arch. Microbiol. 132:1-9.
8. Inoue, K., S. Kageyama, K. Miki, T. Morinaga, Y. Kamagata, K. Nakamura, and E. Mikami (1992) Vitamine B12 production by Acetobacterium sp. and its tetrachloromethane-resistant mutants. J. Ferment. Bioeng. 73:76-78.
9. Jarrel, K. F., and M. L. Kalmokoff (1988) Nutritional requirements of the methanogenic archaebacteria. Can. J. Microbiol. 34:557-576.
10. Jones, J. B., and T. C. Stadtman (1977) Methanococcus vannielii: effects of selenium and tungsten on growth. J. of Bacteriol. 130:1404-1406
11. Koesnandar, N. Nishio, and S. Nagai (1991) Effects of trace metal ions on the growth, homoacetogenesis and corrinoid production by Clostridium thermoaceticum. J. Ferment. Bioeng. 71:181-185.
12. Krzycki, J., and J. G. Zeikus (1980) Quantification of corrinoids in methanogenic bacteria. Curr. Microbiol. 3:243-245.
13. Lettinga, G., A. T. van der Geest, S. Hobma, and J. van der Laan (1980) Anaerobic treatment of methanolic wastes. Water Res. 13:725-737.
14. Lettinga, G., W. de Zeeuw, and E. Ouborg (1981) Anaerobic treatment of wastes containing methanol. Water Res. 15:171-182.
15. Ljungdahl, L. G. (1986) The autotrophic pathway of acetate synthesis in acetogenic
57

References
bacteria. Ann. Rev. Microbiol. 40:415-450. 16. Lustenhouwer, J. W. A., and J. A. Hin (1990) Sample digestion and ICP-AES
analysis. Intern. J. Env. Anal. Chem. 39:209-214. 17. Mah, R. A., M. R. Smith, and L. Baresi (1978) Studies on acetate-fermenting strain
of Methanosarcina. Appl. Environ. Microbiol. 35:1174-1184. 18. Mazumder, T. K., N. Nishio, S. Fukuzaki, and S. Nagai (1987) Production of
extracellular vitamin B-12 compounds from methanol by Methanosarcina barken. Appl. Microbiol. Biotechnol. 26:511-516.
19. van der Meijden, P., C. van der Drift, and G. D. Vogels (1984) Methanol conversion in Eubacterium limosum. Arch Microbiol. 138:360-364.
20. Minami, K., T. Horiyama, M. Tasakiet, and Y. Tanimoto (1986) Methane production using a bio-reactor packed with pumice stone on an evaporator condensate of a kraft pulp mill. J. Ferment. Technol. 64:523-532.
21. Minami, K., K. Okamura, S. Ogawa, and T. Naritomi (1991) Continuous anaerobic treatment of wastewater from a kraft pulp mill. J. Ferment. Bioeng. 71:270-274.
22. Moller, B., R. Oßmer, B. H. Howard, G. Gottschalk, and H. Hippe (1984) Sporomusa, a new genus of gram-negative anaerobic bacteria including Sporomusa sphaeroides spec. nov. and Sporomusa ovata spec. nov. Arch. Microbiol. 139:388-396.
23. Nishio, N., T. Kakizono, R. G. Silveira, S. Takemoto, and S. Nagai (1992) Nutrient control by the gas evolution in methanogenesis of methanol by Methanosarcina barken. J. Ferment. Technol. 73:481-485.
24. Norrman, J. (1983) Anaerobic treatment of a black liquor evaporator condensate from a Kraft mill in three types of fixed-film reactors. Wat. Sei. Tech. 15:247-259.
25. Scherer, P., and H. Sahm (1981) Effect of trace elements and vitamines on the growth of Methanosarcina barkeri. Acta Biotechnol. 1:57-65.
26. Scherer, P., H. Lippert, and G. Wolff (1983) Composition of the major elements and trace elements of 10 methanogenic bacteria determinated by inductively coupled plasma emission spectrometry. Biol. Trace Element Res. 5:149-163.
27. Schönheit, P., J. Moll, and R. K. Thauer (1979) Nickel cobalt and molybidenum requirement for growth of Methanobacterium thermoautotrophicum. Arch Microbiol. 123:105-107.
28. Smith, M. R., and R. A. Mah (1978) Growth and methanogenesis by Methanosarcina strain 277 on acetate and methanol. Appl. Environ. Microbiol. 36:870-879.
29. Speece, R. E. (1983) Anaerobic biotechnology for industrial wastewater treatment. Environ. Sei. Technol. 17:416A-427A.
30. Speece, R. E., G. F. Parkin, and D. Gallagher (1983) Nickel stimulation of anaerobic digestion. Water Res. 17:677-683.
31. Stupperich, E., I. Steiner, and J. Eisinger (1987) Substitution of Coa-(5-hydroxybenzimidazolyl) cobamide (Factor III) by vitamin B12 in Methanobacterium
58

Anaerobic degradation of methanol
thermoautotrophicum. J. Bacteriol. 169:3076-3081. 32. Stupperich, E., H. Eisinger, and S. Schurr (1990) Corrinoids in anaerobic bacteria.
FEMS Microbiol. Rev. 87:355-360. 33. Weimer, P. J., and J. G. Zeikus (1978) One carbon metabolism in methanogenic
bacteria. Arch. Microbiol. 119:49-57. 34. Whitmann, W., E. Ankwanda, and R. S. Wolfe (1982) Nutrition and carbon
metabolism of Methanococcus voltae. J. Bacteriol. 149:852-863. 35. van de Wijngaard, W. M. H., C. van der Drift, C , and G. D. Vogels (1988)
Involvement of a corrinoid enzyme in methanogenesis from acetate in Methanosarcina barkeri. FEMS Microbl. Letters 52:165-172.
36. Wood, H. G., W. Ragsdale, and E. Pezacka (1986) The acetyl-CoA pathway of autotrophic growth. FEMS Microbiol. Rev. 39:345-362.
37. Zeikus, J. G., L. H. Lynd, T. E. Thompson, J. A. Krzycki, P. J. Weimer, and P. W. Hegge (1980) Isolation and characterization of a new, methylotrophic, acidogenic anaerobe, the Marburg strain. Curr. Microbiol. 3:381-386.
59

The importance of cobalt for individual trophic groups in an anaerobic methanol
degrading consortium
Abstract
Methanol is an important anaerobic substrate in industrial wastewater treatment and the natural environment. Previous studies indicate that cobalt greatly stimulates methane formation during anaerobic treatment of methanolic wastewaters. To evaluate the effect of cobalt in a mixed culture, a sludge with low background levels of cobalt was cultivated in an upflow anaerobic sludge blanket reactor. Specific inhibitors in batch assays were then utilized to study the effect of cobalt on the growth rate and activity of different microorganisms involved in the anaerobic degradation of methanol. Only methylotrophic methanogens and acetogens were stimulated by cobalt additions, while the other trophic groups utilizing downstream intermediates, H2/C02 or acetate, were largely unaffected. The optimal concentration of cobalt for the growth and activity of methanol utilizing methanogens and acetogens was 0.05 mg • l"1. The higher requirement of cobalt is presumably due to the previously reported production of unique corrinoid-containing enzymes (or coenzymes) by direct utilizers of methanol. This distinctly high requirement of cobalt by methylotrophs should be considered during methanolic wastewater treatment. Methylotroph methanogens presented a 60-fold-higher affinity for methanol than acetogens. This result in combination with the fact that acetogens grow slightly faster than methanogens under optimal cobalt conditions indicates that acetogens can outcompete methanogens only when reactor methanol and cobalt concentrations are high, provided enough inorganic carbon is available.
Published as: Florencio, L., J. A. Field and G. Lettinga (1994) The importance of cobalt for individual trophic groups in an anaerobic methanol degrading consortium. Appl. Environ. Microbiol. 60:227-234.
61

Introduction
3.1 Introduction
Acetate and H2/C02 intermediates during anaerobic degradation of organic matter are recognized as important methanogenic substrates. Consequently, considerable research has been dedicated to the behaviour of acetoclastic and hydrogenotrophic methanogens during anaerobic wastewater treatment. Less studied is the role of methylotrophic methanogens which are involved in the metabolism of Cj substrates in wastewaters, wastes, and the natural environment.
Methanol is utilized in several chemical industries, such as in the production of formaldehyde41 and esters.47 Methanol is also generated in condensation processes at pulp mills25'31 and coal gasification installations.28 In kraft pulp mill evaporator condensates, methanol is the main organic pollutant, with concentrations ranging from 1.5 to 24.5 g l"1.26
Methanol is also present in natural anaerobic environments, such as anoxic lake sediments and rumen fluid.33'35 Methanol is released from the anaerobic decomposition of pectin34 or similarly the equivalent of methanol is metabolized from methoxylated phenols.2'7'13'22'30
In marine sediments, methanol may also be a methane precursor, although methylamines are more important methanogenic substrates.18 The same microorganisms involved in the anaerobic degradation of methanol are also involved in the anaerobic degradation of halomethane compounds.3'8'24'27'38 Halogenated Cj compounds are present in the environment as natural48 or xenobiotic16 substances.
Methanol, a simple Cl compound, potentially supports a complex food chain composed of a variety of possible trophic groups. Table 1 presents the main reactions involved in the anaerobic degradation of methanol and summarizes their stoichiometry and Gibbs free-energy changes under standard conditions. Several species of methanogens17 and acetogens22 are able to directly utilize methanol as a carbon and energy source for growth. Nitrate- and sulfate-reducing bacteria which can utilize methanol are also known.4'23'29
Acetogens produce acetate or H2/C02 from methanol.5 Thus, methanol can indirectly support hydrogenotrophic14'17'22 and acetoclastic organisms16'17 when acetate or H2/C02 is generated.
Previously, we have studied the effect of trace elements on the anaerobic degradation of methanol by anaerobic sludge.10 Cobalt was the only trace element tested which greatly enhanced methanogenesis from methanol. Cobalt had no remarkable influence on methanogenesis when acetate was used as the substrate. Aside from methanogenesis, acetogenesis of methanol was also stimulated by cobalt, which can result in reactor upsets due to a pH drop from the accumulation of acetate.20'21 In continuous experiments, less acetate was formed in a cobalt-deprived reactor than in a cobalt-supplemented reactor. These
62

Cobalt in anaerobic methanol degrading consortium
results suggested that cobalt levels could be used to prevent acetate formation from methanol. Of particular interest is the possibility that the cobalt concentration could be used as a parameter for controlling substrate flow during anaerobic treatment of methanolic wastewaters. Since cobalt may play a crucial role in the anaerobic degradation of methanol, the objective of this study was to determine which trophic groups in a natural mixed culture respond to cobalt addition. For this purpose, a methanol-degrading consortium with low background levels of cobalt was cultivated in a continuous anaerobic reactor deprived of cobalt supplementation.10 Methanogenic and acetogenic activity batch assays utilizing specific inhibitors were used to study the composition of the population. The effect of cobalt on each of the trophic groups was also evaluated by measuring their growth rate.
3.2 Materials and methods
Biomass
All experiments were carried out with freshly collected methanogenic granular sludge taken from a laboratory-scale upflow anaerobic sludge blanket (UASB) reactor. The sludge was cultivated on methanol as the only substrate in a mineral medium deprived of cobalt for one year.10 The sludge was rinsed with demineralized water before being used in the batch assays.
The content of total suspended solids (TSS) and the volatile suspended solids (VSS) of the sludge were 7.7 and 7.0%, respectively, as determined by standard methods.1 The total cobalt concentration of the sludge after four months of cultivation was 5.5 mg Co • kg"1
TSS,10 four times lower than the seed sludge and more than 100 times lower than a control sludge cultivated on methanol in cobalt-sufficient medium.
Culture medium, inhibitors and chemicals.
Unless otherwise stated, the inorganic macronutrients used in all experiments contained the following (in milligrams per liter of basal medium): NH4C1 (280), K2HP04 • 3H20 (330), MgS04 • 7H20 (100), CaCl2 • 2H20 (10), FeCl2 • 4H20 (2), H3B03 (0.050), ZnCl2 (0.050), MnCl2 • 4H20 (0.5), CuCl2 • 2H20 (0.038), (NH4)6Mo7024 • 4H20 (0.050), A1C13 • 6H20 (0.090), NiCl2 • 6H20 (0.142), Na2Se03 • 5H20 (0.164), CoCl2 • 6H20 (2), EDTA (1), resazurin (0.2), and 36% HCl (0.001 ml • l"1). The medium was made up in demineralized water. To ensure pH stability, 6.72 g NaHC03 (80 mM) was added per liter of the basal medium in combination with 30% C02 in the head space. When the substrate
63

Materials and methods
was methanol plus pure hydrogen in the head space, phosphate buffers were utilized instead.11 ThepH values were set around 7.2. When required, the inhibitors vancomycin and 2-bromoethanesulfonic acid (BESA) were utilized at concentrations of 100 mg • l"1 and 6.33 g • l"1 (30mM), respectively.
All chemicals were of analytical grade and were supplied by E. Merck AG, Darmstadt, Germany, except for resazurin, which was purchased from Fluka Chemie AG, Buchs, Switzerland, and vancomycin and BESA which were obtained from Sigma, St Louis, USA, and Janssen, Tilburg, The Netherlands, respectively. The gases were supplied by Hoekloos, Schiedam, The Netherlands.
Analyses
The pH was determined potentiometrically. Methanol and volatile fatty acids (VFA) were determined by gas chromatography by using a flame ionization detector.10 Samples for measuring hydrogen (500 jtl) and methane (100 1) were obtained by using gas-tight syringes equipped with Pressure-Lok valves (Dynatech, Baton Rouge, La.). Methane was analyzed by using a flame ionization gas Chromatograph model 438/S (Packard-Becker, Delft, The Netherlands). The gas Chromatograph was equipped with a steel column (2 m by 2 mm) packed with Porapak Q (80/100 mesh, Millipore Corp., Bedford, Mass.). The temperatures of the column, injector port, and the flame ionization detector were 60, 200, and 220°C, respectively. Nitrogen was used as a carrier gas at a flow rate of 20 ml • min"1. Hydrogen was detected by thermal conductivity in a model 5890 gas Chromatograph (Hewlett Packard, Palo Alto, Calif.). The gas Chromatograph was equipped with a steel column (1.5 m by 2 mm) packed with molecular sieve 0.5 nm (60/80 mesh; Chrompack, Bergen op Zoom, The Netherlands). Argon was used as a carrier gas (45 ml • min"1). The column, the injection port and the thermal conductivity detector temperatures were 40 °C, 110 °C and 125 °C, respectively.
Experimental set-up
Specific inhibitors were utilized to study the composition of the population by activities and apparent growth rate measurements. Figure 1 and Table 1 represent the general strategy utilized for blocking the competitive reactions and the substrates utilized for evaluating the activity or growth of individual pathways for the anaerobic degradation of methanol. To minimize cobalt contamination, all glassware was washed with 5 M HN03
solution and rinsed with demineralized water. All experiments were conducted by using
64
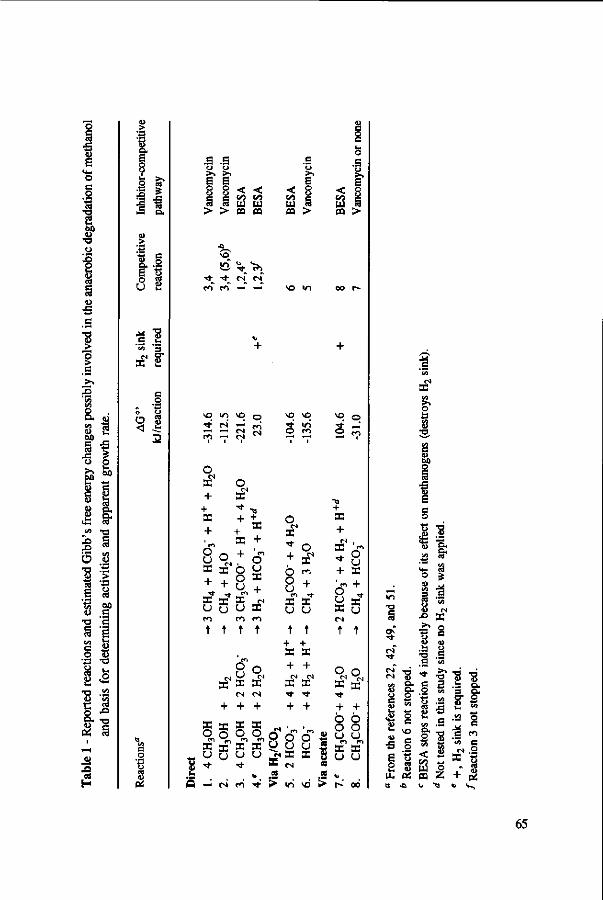
S
î 60
•8 •8
•8 _> >
_>> 3 'S CO
60 _
S I S. s
£5 M §
p <4-H
1/5
Ü
"8 n3 e •a <i> •o S c o
! U r a c* ^H V
A te
H
% 1 C0
•o 5 C/ï
*>
a 0 0 c e Fi S <u •o
5 c/ï
'co
rt X ) T )
§
a. S
II
S. .2
•a ?
x 2
U <
04
E o E S < <
co c/j § rt tu tu > > OQ BQ
•* •* e i c i
< co U m
S o
>
.s
< 8 co g ca >
!
s
o 't'
•
O ^çs x +
+ X
+ o u X + X* u t
ac o ac u •* —i
«-> CN
1
°o> X
+
ac u t
CS
ac
+
X o i u
ts
vq
CS CN
1
O ac • *
+ + X + b o u
u c"i t
O U X CS
+ X O
m X u ' t
ei
O ei
• «
+ X + o u X +
CS X
t
o X CN
+ X o X u *> • * '
u
>
o
3 1
o CS
X ' t + ö o u
m
X u t
+ X +
CS
X
+
o u X CS
wi
vq u-i en
•
O CS
X C l
+ X u T
+ X +
CS
X
+
o u X
vd
Ol
«
1 C9
ca
o
3
• «
+ X
+ CS
X ' t + o u X CN
t
O CS
X
+ ó o u a u *> ^ r-'
o
<*1 1
o u X + X u t
o ac"
+ ö o o
m X CJ
0 0
1/1
• o
9
CN t
CN CN Vi
o c Ui
1-1
•s g o !_i
O
'S C L O .
O
e C
_o
co CO
os «
1 1
V
3
to
I « 0>
e o t3 0)
se a3
.2 > * -o
CO S/3
O
•3
c
o CO
V3 O . O co
< CO UJ
m o
1 & cS
1 CA
IX
g <u .e ' C A
>. •o S c/l
c/l
'•S c
i 4->
O Z
•o
requ
ired
, t s
topp
ed.
.52 O
'S5 c es .2
S « „ ca
+ & « "
•a
65
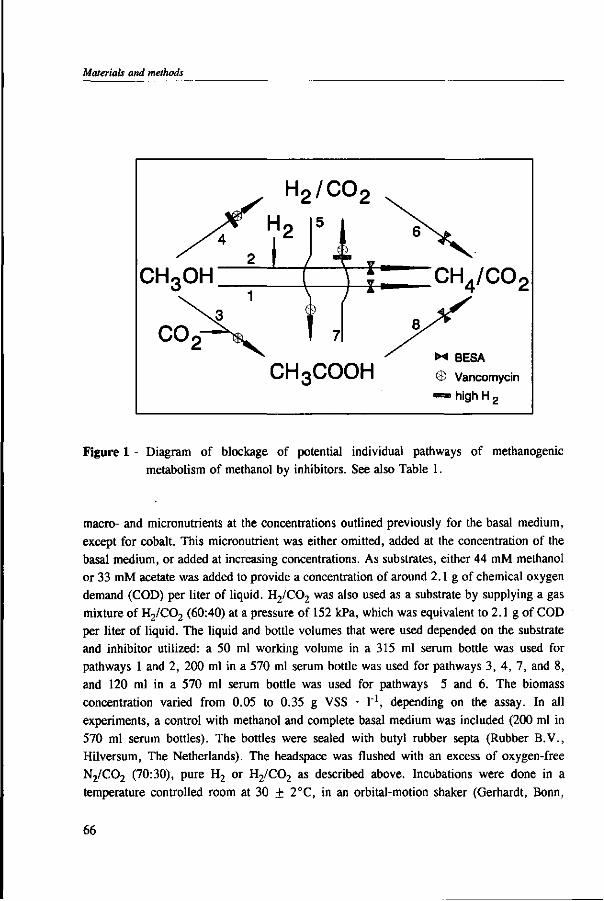
Materials and methods
H 2 / C 0 2 N
x.ï'j A CHoOH \ t
V 1 i CH3COOH
V
^ v - CH,/COo
M BESA
© Vancomycin
= high H 2
Figure 1 - Diagram of blockage of potential individual pathways of methanogenic metabolism of methanol by inhibitors. See also Table 1.
macro- and micronutrients at the concentrations outlined previously for the basal medium, except for cobalt. This micronutrient was either omitted, added at the concentration of the basal medium, or added at increasing concentrations. As substrates, either 44 mM methanol or 33 mM acetate was added to provide a concentration of around 2.1 g of chemical oxygen demand (COD) per liter of liquid. H2/C02 was also used as a substrate by supplying a gas mixture of H2/C02 (60:40) at a pressure of 152 kPa, which was equivalent to 2.1 g of COD per liter of liquid. The liquid and bottle volumes that were used depended on the substrate and inhibitor utilized: a 50 ml working volume in a 315 ml serum bottle was used for pathways 1 and 2, 200 ml in a 570 ml serum bottle was used for pathways 3, 4, 7, and 8, and 120 ml in a 570 ml serum bottle was used for pathways 5 and 6. The biomass concentration varied from 0.05 to 0.35 g VSS - l"1, depending on the assay. In all experiments, a control with methanol and complete basal medium was included (200 ml in 570 ml serum bottles). The bottles were sealed with butyl rubber septa (Rubber B.V., Hilversum, The Netherlands). The headspace was flushed with an excess of oxygen-free N2/C02 (70:30), pure H2 or H2/C02 as described above. Incubations were done in a temperature controlled room at 30 ± 2°C, in an orbital-motion shaker (Gerhardt, Bonn,
66

Cobalt in anaerobic methanol degrading consortium
Germany) at 120 strokes • min"1. Liquid and gas samples were taken periodically to analyze substrate consumption and product formation. The flasks were shaken vigorously before sampling. All results are reported as the mean value of triplicate cultures. The standard deviations were less than 5% of the triplicate mean values. The final pH values of the cultures were determined at the end of each series and were less than 0.35 pH unit below the initial pH of 7.2.
Determination of specific activities, apparent specific growth rate (n) and substrate qffinity (K)
The specific methanogenic and acetogenic activities were determined from the linear increase of products (CH4 and VF A) in the beginning of the experiment, when no lag phase was observed. In this period, net growth compared with the initial population size was considered negligible; when necessary, higher sludge concentrations were utilized. For determination of growth rates, sludge concentrations that were low enough to ensure an exponential increase in product formation were utilized. Assuming that product formation and growth remained coupled, the apparent specific growth rates were calculated from the slope of the linear portion of the plot of the natural logarithm of the product yield versus time by performing a least-squares analysis. The apparent substrate (methanol) affinity coefficients for methanogens and acetogens were estimated from a Lineweaver-Burk plot by comparing the growth rates as a function of the substrate concentration at different times during substrate depletion.
3.3 Results
The effect of BESA and cobalt on methanol utilization by the cobalt-deprived enrichment culture for a typical experiment is illustrated in Figure 2. Methane production occurred without a lag phase (Fig. 2A), independent of cobalt addition. However, the addition of cobalt greatly stimulated the rate of methane production from methanol, which was totally consumed in less than 4 days in contrast to 14 days required for total substrate consumption in cobalt-deprived bottles. The period at the start of the assay (between 0 and 2 days) was used to determine the microbial activities. In this period, the rate of methane formation was constant, and at least 70% of the substrate applied was converted to methane (Fig. 2A) indicating a large initial population of methylotrophic methanogens. BESA, at a concentration of 30 mM, halted methanogenesis from methanol (Fig. 2B). Instead, VFA were formed in an exponentially increasing pattern which indicated the growth of acetogenic microorganisms during the assay, although the initial activity was low. Acetate was the main
67
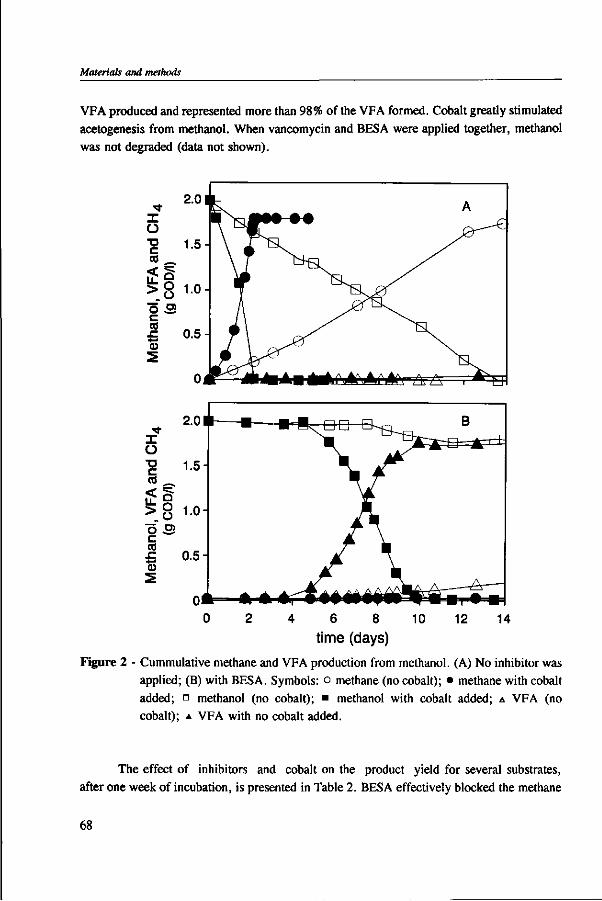
Materials and methods
VF A produced and represented more than 98% of the VF A formed. Cobalt greatly stimulated acetogenesis from methanol. When vancomycin and BESA were applied together, methanol was not degraded (data not shown).
10 12 14 0 2 4 6 8 time (days)
Figure 2 - Cummulative methane and VF A production from methanol. (A) No inhibitor was applied; (B) with BESA. Symbols: o methane (no cobalt); • methane with cobalt added; n methanol (no cobalt); • methanol with cobalt added; A VFA (no cobalt); A VFA with no cobalt added.
The effect of inhibitors and cobalt on the product yield for several substrates, after one week of incubation, is presented in Table 2. BESA effectively blocked the methane
68
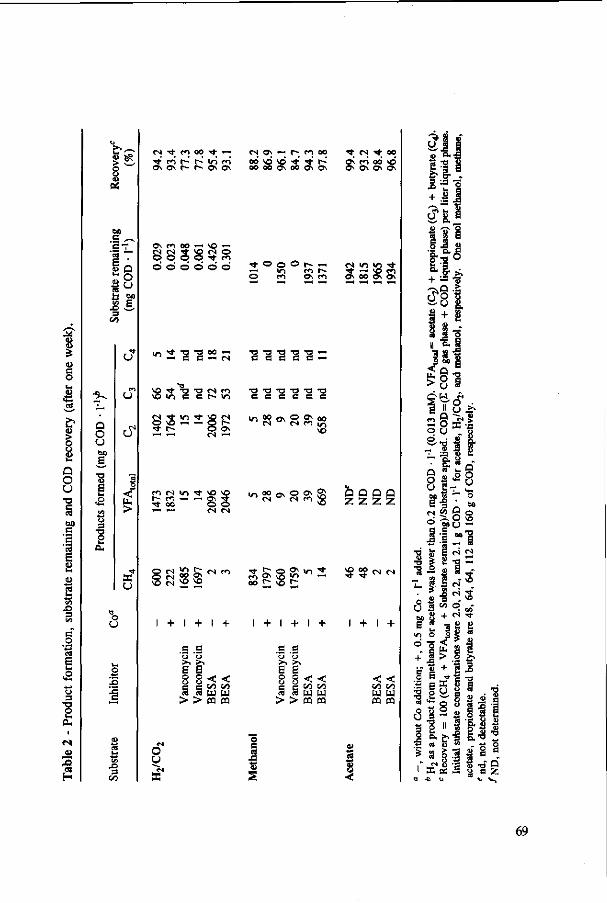
c o
e
3
1 P-,
1
0!
H
s 1 -
is 13 u
i? SP
£ <u c 0 11
£ * * -< cd
fr 0)
> 5 & e Q O U T3
5 ÖD
c 'S •a E u v* <L>
f*
•*s
" Q O U 6 0
E
'S e .2 a 0 3 •O
s £
u
Cs
u
• ^
< u. >
*S X u
o U
15
D CO
P I ^ n « t « t V i P-' |v ui tri ON ON I— r ^ O N ON
CN ON — r - r»> 00 od vd N O • * ' ' t ' r-' OO 0 0 ON 0 0 ON ON
Tf { S • * 0 0
ON C l o ó NO ON ON ON ON
ON C l OO — NO — N N * > O N O © © O © T* o Ö Ö O Ö Ö Ö Tt O o O 1» _
—; m o r-O Cl 0 \ f l
N m i n * • * ~< 0? o ON 0 0 ON ON
NO • * T3 T3 <S C l N O w i e e t - ir,
o < N u-i • * N O NO r~ C l —c - H ON • * Tj- O0 O O —' —< CN| ( S
csi >r> r -r o 00 O N
£ 3 NO NO
I + I + I +
o
S S 8 8 < < S c co co
« U U > > oa oa
•O -U TJ T3
c c c e
""> S <* s S5 °° CN CS C l | /^
•* Kr <=> S " »*> 2 1 ^ !C 00 r- NS f~
I + I + I +
8 8 < < q g co co
> > DD OD
e e CS
x: 'S S
Q Q Q Q
z z z z
> O ? = N < N
I + I +
< < co co tü tU 09 OQ
8 <
e £
« <,
69
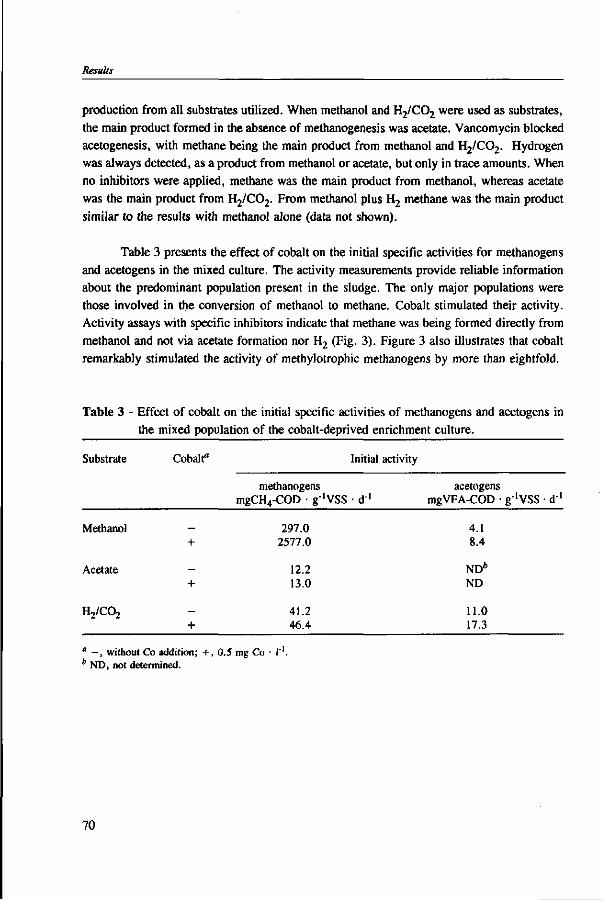
Results
production from all substrates utilized. When methanol and H2/C02 were used as substrates, the main product formed in the absence of methanogenesis was acetate. Vancomycin blocked acetogenesis, with methane being the main product from methanol and H2/C02. Hydrogen was always detected, as a product from methanol or acetate, but only in trace amounts. When no inhibitors were applied, methane was the main product from methanol, whereas acetate was the main product from H2/C02. From methanol plus H2 methane was the main product similar to the results with methanol alone (data not shown).
Table 3 presents the effect of cobalt on the initial specific activities for methanogens and acetogens in the mixed culture. The activity measurements provide reliable information about the predominant population present in the sludge. The only major populations were those involved in the conversion of methanol to methane. Cobalt stimulated their activity. Activity assays with specific inhibitors indicate that methane was being formed directly from methanol and not via acetate formation nor H2 (Fig. 3). Figure 3 also illustrates that cobalt remarkably stimulated the activity of methylotrophic methanogens by more than eightfold.
Table 3 - Effect of cobalt on the initial specific activities of methanogens and acetogens in the mixed population of the cobalt-deprived enrichment culture.
Substrate
Methanol
Acetate
H2/C02
Cobalt0
+
+
+
me mgCH4
thanogens -COD • g"
297.0 2577.0
12.2 13.0
41.2 46.4
VSS
Initial
•d-1
activity
acetogens mgVFA-COD • g'VSS • d 1
4.1 8.4
NDfc
ND
11.0 17.3
" - , without Co addition; +, 0.5 mg Co • I"1. * ND. not determined.
70
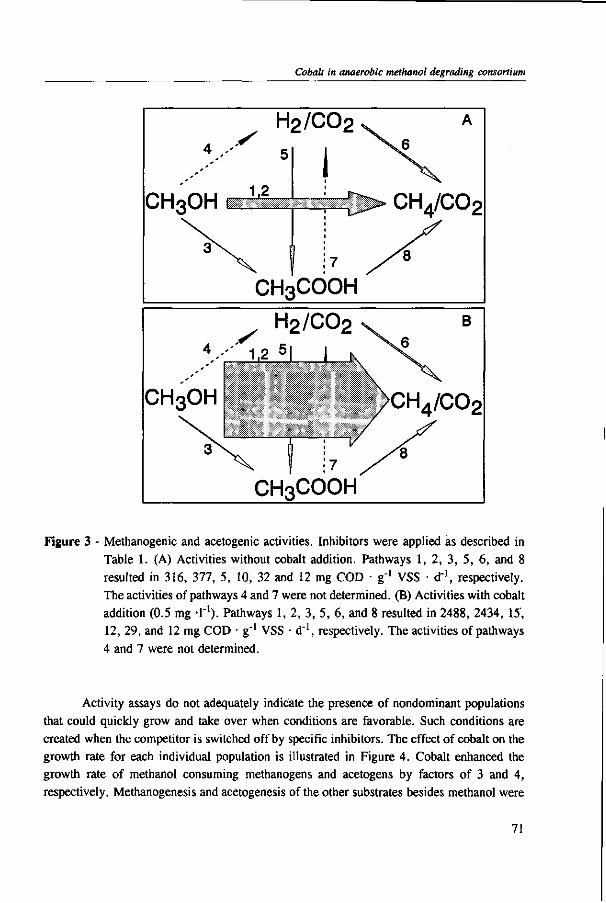
Cobalt in anaerobic methanol degrading consortium
CH3OH 1,2 ^ > CH4/C02
I i7
CH3CÓOH
Figure 3 - Methanogenic and acetogenic activities. Inhibitors were applied as described in Table 1. (A) Activities without cobalt addition. Pathways 1, 2, 3, 5, 6, and 8 resulted in 316, 377, 5, 10, 32 and 12 mg COD • g'1 VSS • d"1, respectively. The activities of pathways 4 and 7 were not determined. (B) Activities with cobalt addition (0.5 mg -l"1). Pathways 1, 2, 3, 5, 6, and 8 resulted in 2488, 2434, 15, 12, 29, and 12 mg COD • g"1 VSS • d"1, respectively. The activities of pathways 4 and 7 were not determined.
Activity assays do not adequately indicate the presence of nondominant populations that could quickly grow and take over when conditions are favorable. Such conditions are created when the competitor is switched off by specific inhibitors. The effect of cobalt on the growth rate for each individual population is illustrated in Figure 4. Cobalt enhanced the growth rate of methanol consuming methanogens and acetogens by factors of 3 and 4, respectively. Methanogenesis and acetogenesis of the other substrates besides methanol were
71
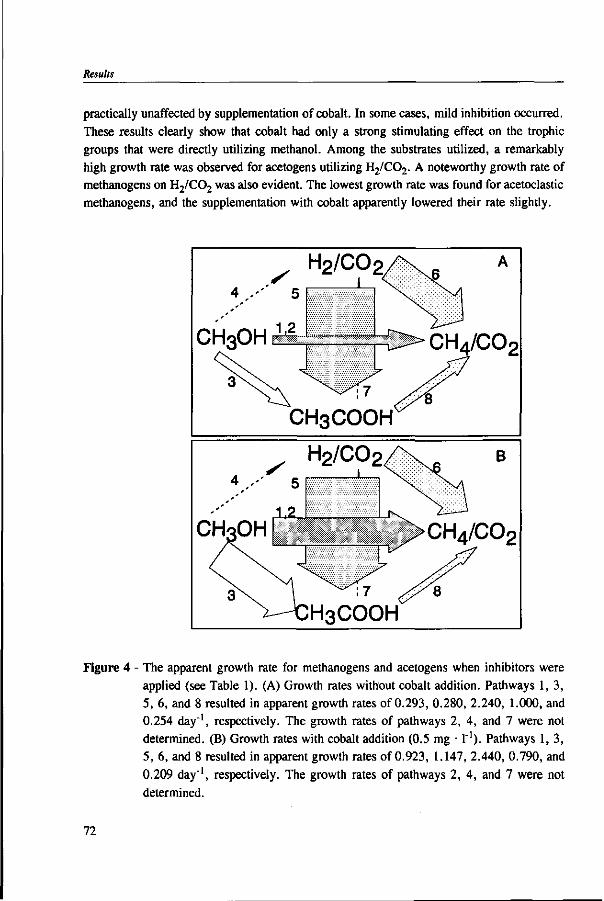
Results
practically unaffected by supplementation of cobalt. In some cases, mild inhibition occurred. These results clearly show that cobalt had only a strong stimulating effect on the trophic groups that were directly utilizing methanol. Among the substrates utilized, a remarkably high growth rate was observed for acetogens utilizing H2/C02. A noteworthy growth rate of methanogens on H2/C02 was also evident. The lowest growth rate was found for acetoclastic methanogens, and the supplementation with cobalt apparently lowered their rate slightly.
^ H2 /C02
4 X ' 5 — J
ChhOH
H3COOH
B
CH4/CO2
Figure 4 - The apparent growth rate for methanogens and acetogens when inhibitors were applied (see Table 1). (A) Growth rates without cobalt addition. Pathways 1,3, 5, 6, and 8 resulted in apparent growth rates of 0.293, 0.280, 2.240, 1.000, and 0.254 day"1, respectively. The growth rates of pathways 2, 4, and 7 were not determined. (B) Growth rates with cobalt addition (0.5 mg • l"1). Pathways 1, 3, 5, 6, and 8 resulted in apparent growth rates of 0.923, 1.147, 2.440, 0.790, and 0.209 day"1, respectively. The growth rates of pathways 2, 4, and 7 were not determined.
72
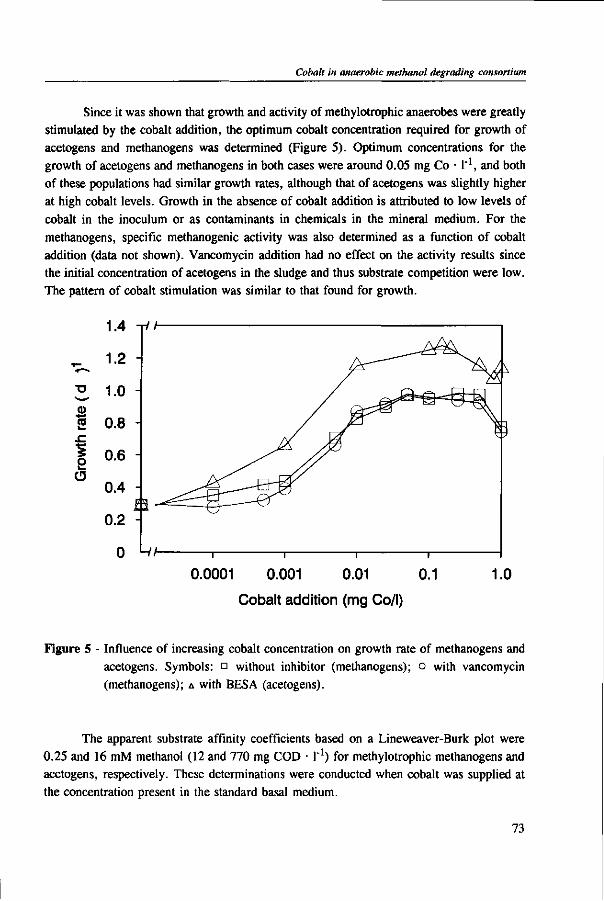
Cobalt in anaerobic methanol degrading consortium
Since it was shown that growth and activity of methylotrophic anaerobes were greatly stimulated by the cobalt addition, the optimum cobalt concentration required for growth of acetogens and methanogens was determined (Figure 5). Optimum concentrations for the growth of acetogens and methanogens in both cases were around 0.05 mg Co • l"1, and both of these populations had similar growth rates, although that of acetogens was slightly higher at high cobalt levels. Growth in the absence of cobalt addition is attributed to low levels of cobalt in the inoculum or as contaminants in chemicals in the mineral medium. For the methanogens, specific methanogenic activity was also determined as a function of cobalt addition (data not shown). Vancomycin addition had no effect on the activity results since the initial concentration of acetogens in the sludge and thus substrate competition were low. The pattern of cobalt stimulation was similar to that found for growth.
^ ,
• D
2
1.4
1.2
1.0
0.8
0.6
0.4
0.2
0 0.0001 0.001 0.01 0.1
Cobalt addition (mg Co/I)
1.0
Figure 5 - Influence of increasing cobalt concentration on growth rate of methanogens and acetogens. Symbols: a without inhibitor (methanogens); o with vancomycin (methanogens); A with BESA (acetogens).
The apparent substrate affinity coefficients based on a Lineweaver-Burk plot were 0.25 and 16 mM methanol (12 and 770 mg COD • l"1) for methylotrophic methanogens and acetogens, respectively. These determinations were conducted when cobalt was supplied at the concentration present in the standard basal medium.
73

Discussion
3.4 Discussion
Influence of cobalt
Cobalt is the central ion in corrinoids present in all methanogens and acetogens.40 A number of corrinoid-dependent reactions are known to take place in the intermediary metabolism of substrates by methanogens and acetogens.49 In methanogens, corrinoids are involved in methyl transfer from methanol to methyl coenzyme M,43,45 the common precursor of methane from all substrates. In acetogens, corrinoids participate in the formation of acetyl coenzyme A, the precursor intermediate of acetate and cell synthesis.49 However, the content of corrinoids in anaerobic bacteria varies greatly among species and substrate utilized but is always higher when cells are grown on methanol.19 The initial step of methanol conversion in methanol consuming anaerobes, such as the methanogen Methanosarcina barken and the acetogen Eubacterium limosum, proceeds in a similar way and is catalyzed by an additional corrinoid-containing enzyme known as methyltransferase.43'44 Recently, an induced corrinoid-containing protein was reported to occur only in methanol-grown cells of an acetogenic bacterium Sporomusa ovata?9 Thus, the high requirement for cobalt found in our studies is presumably due to the production of unique corrinoid-containing enzymes or coenzymes that are only present in methylotrophs.
The addition of cobalt stimulated the activity and growth of both methanogens and acetogens, which directly utilized methanol in the cobalt-deprived enrichment culture. The optimum cobalt concentration in our methylotrophic mixed culture is in the range reported for the marine isolated methanogen Methanococcoides methylutens grown on trimethylamines.37 A much lower requirement for cobalt has been reported for the thermophilic hydrogenotroph methanogen Methanobacterium thermoautotrophicum.36 In our consortium, methanogenesis of acetate and H2/C02 as well as acetogenesis of H2/C02 was not seriously affected by cobalt addition, despite low cobalt levels. It is impossible to ascertain whether cobalt is an essential nutrient for the latter microorganisms under the experimental conditions utilized, because the amount required could be so low that contamination (of chemicals or utensils) and the background level in the seed inoculum could have been adequate. Nonetheless, this study clearly demonstrates that methylotrophic methanogens and acetogens have a distinctly higher requirement for cobalt, which should be considered during wastewater treatment.
Ecology of anaerobic methanol degradation
Although methanol is a simple compound with only one carbon, it can support a very
74

Cobalt in anaerobic methanol degrading consortium
complex food chain under anaerobic conditions. The methylotrophic food web is outlined in Figure 1. Methane can be directly produced from methanol by the action of methylotrophic methanogens or indirectly by action of acetogens in conjunction with acetoclastic or hydrogenotrophic methanogens. Acetate is also directly formed from methanol, but its formation is limited by the availability of inorganic carbon. If no exogenous inorganic carbon (e.g. HC03") is added, the maximum acetate production will be equal to 33% of the consumed methanol, when methanogenesis occurs. The complexity of this food web is even greater when inorganic electron acceptors such as sulfate are also available.14'32 Syntrophy and competition among organisms may occur in such environments.6,32 The formation of H2/C02 from methanol is dependent on the removal of H2 by hydrogenotrophic anaerobes5'14
since hydrogen accumulation would be thermodynamically unfavorable.32'42 Thus, it is necessary that the syntrophic organism keeps the hydrogen partial pressure extremely low.42
Competition for methanol may occur between methylotrophic acetogens and methanogens, and also competition for H2/C02 between hydrogenotrophs is possible. '3 H2 is also produced in minor amounts by direct methanogenesis of methanol.32
Direct methylotrophic methanogenesis was the main pathway in our enrichment culture. The activities of methanogens were almost the same in the presence or absence of vancomycin, indicating that methylotrophic acetogens were not participating in the degradation of methanol. In addition, almost no acetoclastic methanogenic or hydrogenotrophic activities were observed. These populations would be expected if methanol was degraded via H2 or acetate, respectively.
Dominance of methylotrophic methanogens
Several factors might have contributed to the dominance of methylotrophic methanogens over their acetogenic competitors in this food web with an excess of inorganic carbon. These could include competition for cobalt, the size of the initial population and competition for methanol. In relation to cobalt, both populations had similar optimum concentration of cobalt for growth, and their growth rates were almost the same. The growth rate of acetogens was slightly higher at optimal cobalt concentrations. The cobalt optimum can only be determined when the competitor is switched off by specific inhibitors. In coculture, however, the methanogens may have been better scavengers for cobalt. Additionally, an initial advantage in population size would ensure that most of the cobalt would be captured by the methanogens by virtue of sheer numbers. The initial sludge used to seed the cobalt-deprived enrichment culture did in fact have methylotrophic methanogenic activity, while little acetogenic activity was evident. Finally, we have shown that methylotrophic methanogens have approximately 60-fold-higher affinity for methanol than
75
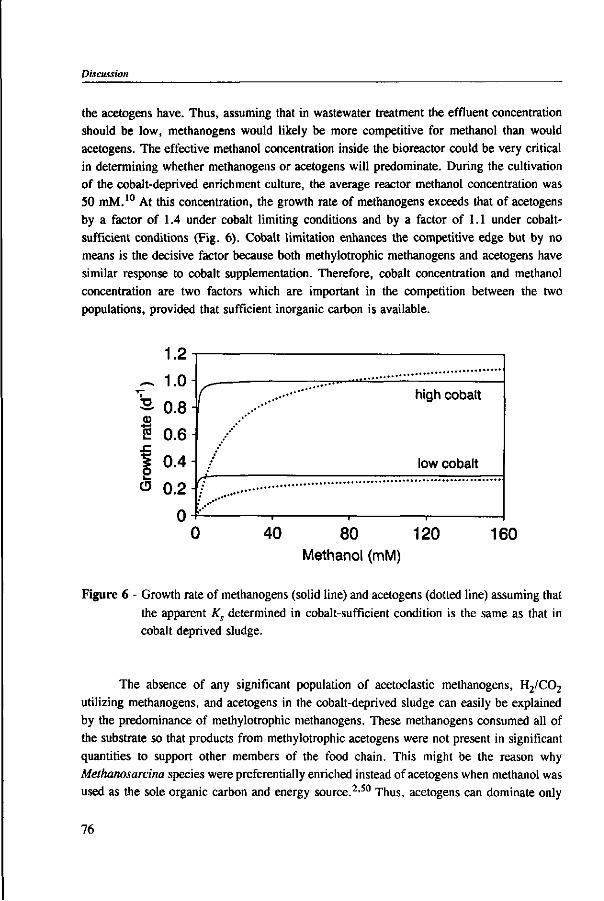
Discussion
the acetogens have. Thus, assuming that in wastewater treatment the effluent concentration should be low, methanogens would likely be more competitive for methanol than would acetogens. The effective methanol concentration inside the bioreactor could be very critical in determining whether methanogens or acetogens will predominate. During the cultivation of the cobalt-deprived enrichment culture, the average reactor methanol concentration was 50 mM.10 At this concentration, the growth rate of methanogens exceeds that of acetogens by a factor of 1.4 under cobalt limiting conditions and by a factor of 1.1 under cobalt-sufficient conditions (Fig. 6). Cobalt limitation enhances the competitive edge but by no means is the decisive factor because both methylotrophic methanogens and acetogens have similar response to cobalt supplementation. Therefore, cobalt concentration and methanol concentration are two factors which are important in the competition between the two populations, provided that sufficient inorganic carbon is available.
1.2 ~ 1.0
2- 0.8 o £5 0.6
% 0.4
o 0.2-7
high cobalt
low cobalt
40 80
Methanol (mM) 120 160
Figure 6 - Growth rate of methanogens (solid line) and acetogens (dotted line) assuming that the apparent Ks determined in cobalt-sufficient condition is the same as that in cobalt deprived sludge.
The absence of any significant population of acetoclastic methanogens, H2/C02
utilizing methanogens, and acetogens in the cobalt-deprived sludge can easily be explained by the predominance of methylotrophic methanogens. These methanogens consumed all of the substrate so that products from methylotrophic acetogens were not present in significant quantities to support other members of the food chain. This might be the reason why Methanosarcina species were preferentially enriched instead of acetogens when methanol was used as the sole organic carbon and energy source.2'50 Thus, acetogens can dominate only
76

Cobalt in anaerobic methanol degrading consortium
when special conditions are created by the specific methanogenic inhibitor BESA or by specific toxicity of methanogens such as that caused by dichloromethane12 and chloroform.15
Also, toxicity due to undissociated acetic acid, resulting from the accumulation of acetic acid exceeding the buffer capacity, selectively inhibited the methanogens and caused the predominance of acetogens.9
3.5 Acknowledgments
We thank A. J. M. Stams for reviewing the manuscript and for valuable suggestions and Sonja Viester for conducting part of the experimental work. We are also grateful to the Brazilian Ministry of Education for providing the scholarship to L. Florencio (CAPES project 909/88-12) that made this work possible.
3.6 References
1. American Public Health Association (1985) Standard methods for examination of water and wastewater, 16th ed. American Public Health Association, Washington, D.C., USA.
2. Bache, R., and N. Pfennig (1981) Selective isolation of Acetobacterium woodii on methoxylated aromatic acids and determination of growth yields. Arch. Microbiol. 130:255-261
3. Belay, N., and L. Daniels (1987) Production of ethane, ethylene, and acetylene from halogenated hydrocarbons by methanogenic bacteria. Appl. Environ. Microbiol. 53:1604-1610.
4. Braum, M., and H. Stolp (1985) Degradation of methanol by a sulfate reducing bacterium. Arch. Microbiol. 142:77-80.
5. Cord-Ruwisch, R., and B. Ollivier (1986) Interspecific hydrogen transfer during methanol degradation by Sporomusa acidovorans and hydrogenophilic anaerobes. Arch. Microbiol. 144:163-165.
6. Cord-Ruwisch, R., H. Seitz, and R. Conrad (1988) The capacity of hydrogenotrophic anaerobic bacteria to compete for trace of hydrogen depends on the redox potential of the terminal electron acceptor. Arch. Microbiol. 149:350-357.
7. Daniel, S. L., E. S. Keith, H. Yang, Y. Lin, and H. L. Drake (1991) Utilization of methoxylated aromatic compounds by the acetogen Clostridium thermoaceticum: expression and specificity of the CO-dependent O-demethylating activity. Biochem. Biophys. Res. Commun. 180:416-422.
77

References
8. Egli, C , T. Tschan, R. Scholtz, A. M. Cook, and T. Leisinger (1988) Transformation of tetrachloromethane to dichloromethane and carbon dioxide by Acetobacterium woodii. Appl. Environ. Microbiol. 54:2819-2824.
9. Florencio, L., J. A. Field, and G. Lettinga Unpublished data. 10. Florencio, L., P. Jenicek, J. A. Field, and G. Lettinga (1993) Effect of cobalt on
anaerobic degradation of methanol. J. Ferment. Bioeng. 75:368-374. 11. Florencio, L., A. Nozhevnikova, A. van Langerak, A. J. M. Stains, J. A. Field,
and G. Lettinga (1993) Acidophilic degradation of methanol by a methanogenic enrichment culture. FEMS Microbiol. Lett. 109:1-6.
12. Freedman, D. L., and J. M. Gossett (1991) Biodegradation of dichloromethane and its utilization as a growth substrate under methanogenic conditions. Appl. Environ. Microbiol. 57:2847-2857.
13. Genthner, B. R. S., and M. Bryant (1987) Additional characteristics of one-carbon-compound utilization by Eubacterium limosum and Acetobacterium woodii. Appl. Environ. Microbiol. 53:471-476.
14. Heijthuijsen, J. H. F. G., and T. A. Hansen (1986) Interspecies hydrogen transfer in co-cultures of methanol-utilizing acidogens and sulfate-reducing or methanogenic bacteria. FEMS Microbiol. Lett. 38:57-64.
15. Hickey, R. F., J. Vanderwielen, and M. S. Switzenbaum (1987) The effect of organic toxicants on methane production and hydrogen gas levels during the anaerobic digestion of waste activated sludge. Water Res. 21:1417-1427.
16. Howard, P. H. (1990) Handbook of environmental fate and exposure data for organic chemicals, vol. II. Solvents. Lewis Publishers, Chelsea, Mich.
17. Jarrel, K. F., and M. L. Kalmokoff (1988) Nutritional requirements of the methanogenic archaebacteria. Can. J. Microbiol. 34:557-576.
18. King, G. M. (1984) Utilization of hydrogen, acetate, and "noncompetitive" substrates by methanogenic bacteria in marine sediments. Geomicrobiol. J. 3:275-306.
19. Krzycki, J., and J. G. Zeikus (1980) Quantification of corrinoids in methanogenic bacteria. Curr. Microbiol. 3:243-245.
20. Lettinga, G., W. de Zeeuw, and E. Ouborg (1981) Anaerobic treatment of wastes containing methanol. Water Res. 15:171-182.
21. Lettinga, G., A. van der Geest, T., S. Hobiiia, and J. van der Laan (1979) Anaerobic treatment of methanolic wastes. Water Res. 13:725-737.
22. Ljungdahl, L. G. (1986) The autotrophic pathway of acetate synthesis in acetogenic bacteria. Annu. Rev. Microbiol. 40:415-450.
23. McCarty, P. L., L. Beck, and P. St. Amant (1969) Biological denitrification of wastewaters by addition of organic materials, p. 1271-1285. In: Proceedings of the 24th Purdue Industrial Waste Conference, Lafayette, Ind. Purdue University, Lafayette, Ind., USA.
78

Cobalt in anaerobic methanol degrading consortium
24. Mikesell, M. D., and S. A. Boyd (1990) Dechlorination of chloroform by Methanosarcina strains. Appl. Environ. Microbiol. 56:1198-1201.
25. Minami, K., T. Horiyama, M. Tasakiet, and Y. Tanimoto (1986) Methane production using a bio-reactor packed with pumice stone on an evaporator condensate of a kraft pulp mill. J. Ferment. Technol. 64:523-532.
26. Minami, K., K. Okamura, S. Ogawa, and T. Naritomi (1991) Continuous anaerobic treatment of wastewater from a kraft pulp mill. J. Ferment. Bioeng. 71:270-274.
27. Mohn, W. W., and J. M. Tiedje (1992) Microbial reductive dehalogenation. Microbiol. Rev. 56:482-507.
28. Mohr, D. H., and J. King (1985) Identification of polar organic compounds in coal-gasification condensate water by gas chromatography-mass spectrometry analysis of high-performance liquid chromatography fractions. Environ. Sei. Technol. 19:929-935.
29. Nanninga, H. J., and J. C. Gottschal (1986) Isolation of a sulfate-reducing bacterium growing with methanol. FEMS Microbiol. Ecol. 38:125-130.
30. Neilson, A. H., A. Allard, C. Lindgren, and M. Remberger (1987) Transformations of chloroguaiacols, chloroveratroles, and chlorocatechols by stable consortia of anaerobic bacteria. Appl. Environ. Microbiol. 53:2511-2519.
31. Norrman, J. (1983) Anaerobic treatment of a black liquor evaporator condensate from a kraft mill in three types of fixed-film reactors. Water Sei. Tech. 15:247-259.
32. Phelps, T. J., R. Conrad, and J. G. Zeikus (1985) Sulfate-dependent interspecies H2
transfer between Methanosarcina barken and Desulfovibrio vulgaris during coculture metabolism of acetate or methanol. Appl. Environ. Microbiol. 50:589-594.
33. Pol, A., and D. I. Demeyer (1988) Fermentation of methanol in the sheep rumen. Appl. Environ. Microbiol. 54:832-834.
34. Schink, B., and J. G. Zeikus (1980) Microbial methanol formation: a major end product of pectin metabolism. Curr. Microbiol. 4:387-390.
35. Schink, B., and J. G. Zeikus. (1982) Microbial ecology of pectin decomposition in anoxic lake sediments. J. Gen. Microbiol. 128:393-404.
36. Schönheit, P., J. Moll, and R. K. Thauer (1979) Nickel, cobalt and molybdenum requirement for growth of Methanobacterium thermoautotrophicum. Arch. Microbiol. 123:105-107.
37. Sowers, K. R., and G. Ferry (1985) Trace metal and vitamin requirements of Methanococcoides methylutens grown with trimethylamine. Arch. Microbiol. 142:148-151.
38. Stromeyer, S. A., W. Winkenbauer, H. Kohler, A. M. Cook, and T. Leisinger (1991) Dichloromethane utilized by an anaerobic mixed culture: acetogenesis and methanogenesis. Biodegradation 2:129-137.
40. Stupperich, E., H. Eisinger, and S. Schurr (1990) Corrinoids in anaerobic bacteria. FEMS Microbiol. Rev. 87:355-360.
79

References
39. Stupperich, E., P. Aulkemeyer, and C. Eckerskorn (1992) Purification and characterization of a methanol-induced cobamide-containing protein from Sporomusa ovata. Arch. Microbiol. 158:370-373.
41. Tani, Y., T. Yonehara, Y. Sakai, and B. D. Yoon (1987) Microbiological synthesis from C^-compounds: application of some methylotrophic functions to synthesis of useful chemicals, p. 282-288. In: H. W. van Verseveld and J.A. Duine (ed). Microbial Growth on C{ compounds. Proceedings of the 5th International Symposium. Martinus Nijhoff Publishers, Dordrecht, The Netherlands.
42. Thauer, R. K., K. Jungermann, and K. Decker (1977) Energy conservation in chemotrpphic anaerobic bacteria. Bacteriol. Rev. 41:100-180.
43. van der Meijden, P., H. J. Heythuysen, H. Sliepenbeek, F. P. Houwen, C. van der Drift, and G. D. Vogels (1984) Activation and inactivation of methanol: 2-mercaptoethanesulfonic acid methyltransferase from Methanosarcina barken. J. Bacteriol. 153:6-11.
44. van der Meijden, P., C. van der Drift, and G. D. Vogels (1984) Methanol conversion in Eubacterium limosum. Arch. Microbiol. 138:360-364.
45. van de Wijngaard, W. M. H., C. van der Drift, and G. D. Vogels (1988) Involvement of a corrinoid enzyme in methanogenesis from acetate in Methanosarcina barkeri. FEMS Microbl. Lett. 52:165-172.
46. Vogels, G. D., J. T. Kelt Jens, and C. van der Drift (1988) Biochemistry of methane production, p. 707-771. In A. J. B. Zehnder (ed.), Biology of anaerobic bacteria. John Wiley, New York.
47. Wesselingh, J. A., G. H. Lameris, P. J. van den Berg, and A. G. Montfoort (1992) Van aardgas naar methanol. Delftse Uitgevers Maatschappij, Delft, The Netherlands.
48. Wever, R. (1991) Formation of halogenated gases by natural sources, p. 277-285. In J. E. Rogers and W. B. Whitman (ed.), Microbial production and consumption of greenhouse gases: methane, nitrogen oxides, and halomethanes. American Society for Microbiology, Washington, D.C.
49. Wood, H. G., W. Ragsdale, and E. Pezacka (1986) The acetyl-CoA pathway of autotrophic growth. FEMS Microbiol. Rev. 39:345-362.
50. Zeikus, J. G., L. H. Lynd, T. E. Thompson, J. A. Krzycki, P. J. Weimer, and P. W. Hegge (1980) Isolation and characterization of a new, methylotrophic, acidogenic anaerobe, the Marburg strain. Curr. Microbiol. 3:381-386.
51. Zinder, S. H., and M. Koch (1984) Non-aceticlastic methanogenesis from acetate: acetate oxidation by a thermophilic syntrophic coculture. Arch. Microbiol. 138:263-272.
80

4
Acidophilic degradation of methanol by a methanogenic enrichment culture
Abstract
An acidophilic methanogenic enrichment culture was obtained in a continuous up-flow anaerobic sludge blanket reactor operated at pH 4.2 with methanol as the sole carbon source. The specific methylotrophic methanogenic activity of the enriched reactor sludge at pH 5 was 3.57 g COD • g_1VSS • d"1 and the apparent doubling time of the biomass was 15.8 h. Acidic conditions were obligatory, since the enrichment culture was not able to produce methane nor grow at pH 7. Based on morphological characteristics, the dominant methanogenic species in the enrichment culture was a Methanosarcina.
Published as: Florencio, L., A. Nozhevnikova, A. van Langerak, A. J. M. Slams, J. A. Field and G. Lettinga (1993) Acidophilic degradation of methanol by a methanogenic enrichment culture. FEMS Microbiol. Lett. 109:1-6.
81

Materials and Methods
4.1 Introduction
Methane production in natural acidic environments, such as in peatlands, is well known.14 However, little is known about the microorganisms involved in methanogenesis at low pH, below 5. So far, a strain of Methanobacteriaceae15 and Methanobacterium espanolae10 have been isolated under acidic conditions. In both cases, isolations were obtained with H2/C02 as substrates. Generally, methanogens in wastewater treatment systems are most active in the neutral pH range. This is probably due to the predominance of aceticlastic methanogens with pH optima of around 7.6,8 When Methanosarcina barken strains were grown on methanol and methanol/H2 mixture instead of acetate they displayed robust growth at pH values as low as 4.3.8
Methanol is the main pollutant in evaporator condensates from the kraft pulping industry.9 These wastewaters do not contain any trace elements nor alkalinity.12 Their anaerobic treatment at low pH could be an interesting option, as addition of alkalinity would not be needed in that case. The aim of this study was to investigate the anaerobic degradation of methanol at a low pH and to characterize the dominant species responsible for the conversion of methanol to methane.
4.2 Materials and methods
Continuous Experiment
The continuous experiment was performed in a temperature controlled room at 30 + 2 °C in small cylindrical glass upward-flow anaerobic sludge blanket reactor (UASB) with a working volume of 0.1 l.4 The reactor was inoculated with 2.0 g of volatile suspended solids (VSS) anaerobic granular sludge obtained from a full-scale UASB reactor treating alcohol distillery wastewater of Nedalco at Bergen op Zoom, The Netherlands. Total suspended solids (TSS) and the VSS content of the sludge were 10.5% and 9.14%, respectively, as determined according to Standard Methods.1 The influent contained methanol as the sole carbon source at concentrations ranging from 4.8 to 5.2 g COD • l"1 (100 to 108 mM). The hydraulic retention time was 5.3 h, and the organic load rate applied averaged 22 g COD • l"1 • d"1. Macronutrients (N, P, and S) and trace elements required for bacterial growth were supplied with a basal mineral medium in milligrams per liter: NH4C1 (280), K2HP04 • 3H20 (330), MgS04 • 7H20 (100), CaCl2 • 2H20 (10), FeCl2 • 4H20 (2), H3B03
(0.050), ZnCl2 (0,050), MnCl2 • 4H20 (0.5), CuCl2 • 2H20 (0.038), (NH4)6Mo7024 • 4H20 (0.050), A1C13 • 6H20 (0.090), CoCl2 • 6H20 (2), NiCl2 • 6H20 (0.142), Na2Se03 • 5H20 (0.164), EDTA (1), resazurin (0.2), and 36% HCl (0.001 ml.!"1). No alkalinity was added
82

Acidophilic degradation of methanol
in the basal medium. NaCl (1000 mg • l"1) was added to the basal medium after 98 days of operation. Methane production was monitored by using a sodium hydroxide solution displacement system.4
Batch Experiments
Media and Buffer solution
The basal mineral medium with trace elements and vitamins used for the activity tests and batch enrichments was described before.13 Phosphate buffers were utilized to set the pH at different levels. Solution 1 was made up with 87.1 g • l"1 K2HP04 and 89.0 g • l"1
Na2HP04 • 2H20, and solution 2, with 68.1 g • l"1 KH2P04 and 78.0 g • l"1 NaH2P04 • 2H20. The proportion of the two solutions and C02 composition of the gas phase depended on the pH which was desired.
Biomass
The inoculum was taken from the bottom of the UASB reactor described above after one year of operation.
Anaerobic Activity Assay
The specific methanogenic activity test was performed in triplicate in 120 ml glass serum bottles sealed with butyl rubber septa (Rubber B.V., Hilversum, The Netherlands) and aluminium caps. The headspace was first flushed with an excess of oxygen-free N2/C02 (4/1) and then a pressure of approximately 170 kPa was applied. Methanol was added at a concentration of 5.0 g COD • l"1 (104 mM). The bottles were inoculated and subsequently incubated in a temperature controlled room at 30 + 1 °C. Liquid and gas samples were taken periodically to analyze methanol consumption and methane production, respectively. The flasks were shaken vigorously before sampling.
Batch Enrichment Cultures
Methanogenic enrichment cultures were obtained at different pH values with methanol 0.96 to 4.8 g COD • l"1 (20 - 100 mM) using the same media and phosphate buffers as
83

materials and methods
described for the anaerobic activity assay. In order to purify the methanogenic bacteria, 100 mg • l"1 of vancomycin or alternatively 100 mg • l"1 of benzilpenicillin were added to the media. An enriched culture was obtained by serial dilution in fresh media.
Analyses and Chemicals
The pH was determined immediately after sampling with a Knick 511 pH-meter (Berlin, Germany) and a Schott Geräte N61 double electrode (Hofheim, Germany).
Prior to the analysis of methanol and volatile fatty acids (VFA), samples were centrifugated at 17000 g for 5 min. Methanol was determined by gas chromatography using a Packard Becker model 417 (Delft, The Netherlands) equipped with a 6 m x 2 mm glass column packed with Supelco port (Bellefont, USA), 100-120 mesh, coated with 10% Fluorad FC 431 (3M, St Paul, USA). The flow rate of the carrier gas (nitrogen saturated with formic acid) was 30 ml • min"1, and the column pressure was +3 bar. The column temperature was 80 °C, the injection port was 220 °C and the detector 280 °C. VFA were analyzed in the same way as methanol, but the temperatures of the column, the injector port and the flame ionization detector were 130 °C, 210 °C, and 230 °C, respectively.
The biogas composition of the continuous reactor was determined by sampling with a syringe at a gas sampling port. Immediately after sampling, 100 /tl was injected into a Packard Becker Gas Chromatograph model 433 (Delft, The Netherlands), equipped with two columns connected in parallel (split 1:1) 1.5 m x 1/8" Teflon, packed with chromosorb 108, 60-80 mesh (Johns Manville, USA) and a 1.2 m x 1/8" stainless steel, packed with molecular sieve 5A, 60-80 mesh (Chrompack, Bergen op Zoom, The Netherlands). The two columns allowed the analyse of C02 and CH4 in one sample. Helium was used as a carrier gas (45 ml • min"1). The oven, detector and injection temperatures were 40 CC, 125 °C and 110 °C, respectively. For the gas composition of the headspace in the batch experiments, the carrier gas was argon (30 ml • min"1). The oven temperature was 100 °C and the detector was 160 °C.
Morphology of cultures was studied with phase-contrast and luminescent microscopy (Wild Leitz, Heerbrugg, Switzerland).
All chemicals were of analytical grade and were supplied by E. Merck AG, Darmstadt, Germany, except for resazurin, which was purchased from Fluka Chemie AG, Buchs, Switzerland, vancomycin and bezilpenicillin from Sigma, St Louis, USA, and for the gases which were supplied by Hoekloos, Schiedam, The Netherlands.
84
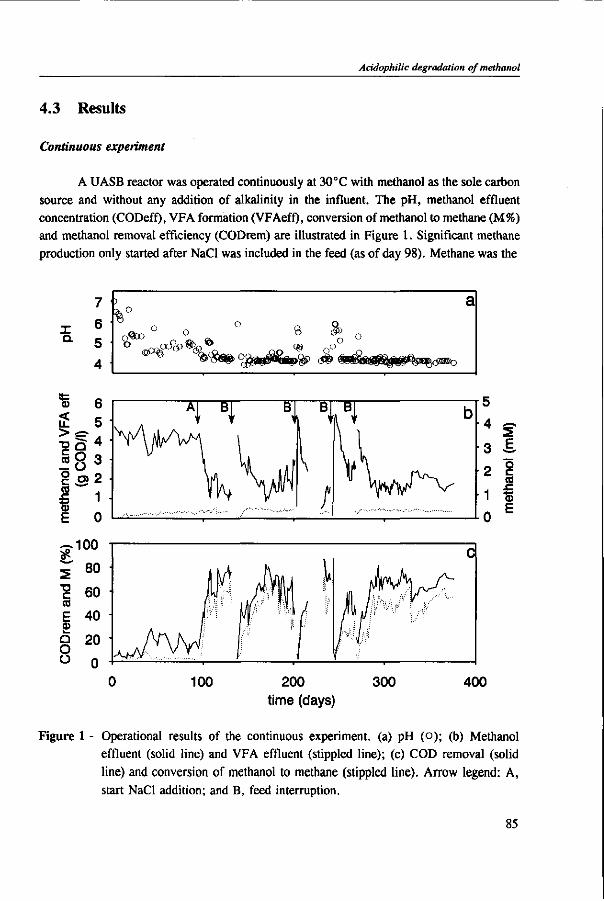
Acidophilic degradation of methanol
4.3 Results
Continuous experiment
A UASB reactor was operated continuously at 30°C with methanol as the sole carbon source and without any addition of alkalinity in the influent. The pH, methanol effluent concentration (CODeff), VF A formation (VFAeff), conversion of methanol to methane (M%) and methanol removal efficiency (CODrem) are illustrated in Figure 1. Significant methane production only started after NaCl was included in the feed (as of day 98). Methane was the
100 200 time (days)
300 400
Figure 1 - Operational results of the continuous experiment, (a) pH (o); (b) Methanol effluent (solid line) and VFA effluent (stippled line); (c) COD removal (solid line) and conversion of methanol to methane (stippled line). Arrow legend: A, start NaCl addition; and B, feed interruption.
85

Results
main product of methanol conversion during the rest of the reactor operation when the pH was quite stable around 4.2. The experimental averaged conversion of methanol to methane (as of day 98) was 54% when the organic loading rate was 24 g COD • 1"' • d"1. About 30% of the methanol applied to the reactor was not converted. Some VF A was formed, accounting for 8% of the methanol COD consumed. The butyrate concentration ranged from 2 to 3 mM and the acetate concentration 0.5 to 1 mM. The biomass yield based on mass balancing (between day 160 and day 190 and between day 285 and day 328) averaged 0.0775 g VSS • g"1 methanol-COD consumed (4.27 g dry cells • mol"1 methanol consumed). For these calculations it was assumed that 1 g VSS was equivalent to 1.15 g dry cells7 and the experimental formula for bacterial dry mass was C5H7N02
5 that gives 1 g VSS being equal to 1.42 g COD. The maximum methanogenic activity of the biomass sampled from the reactor was 3.57 g CH4-COD • g"1 VSS • d"1 (48.6 mmol CH4 • g
_1 cell dry weight • d"1). The doubling time based on the exponential increment of methane production in batch enrichment cultures was 15.8 hours. The reactor methanogenic activity was very sensitive to accidental interruptions in the feed. The time required for recuperation was dependent on the duration of the feed interruption. In the reactor, several morphologically distinct bacterial forms were present. The dominant species were methanosarcina-type clumps. Rod shaped cells of different sizes were also present.
Batch enrichment cultures
Figure 2 presents the results of methanol consumption, methane and VF A formation at pH 5 and pH 7 after inoculation with fresh biomass from the reactor. At pH 5, methanogenesis started immediately. VFA formation also occurred to a lesser extent. Methanosarcina-like cells were the most prevalent microorganisms. In this experiment, cell lysis was observed after substrate depletion. When the sludge was first stored at 4°C, prior to use in the batch assays, methanogens developed very poorly. In contrast, acetogens were not disturbed by storage (data not shown).
At pH 7, methanol conversion was very slow and acetogenesis was more important than methanogenesis. About 1.1 mmol of methanol was consumed in 10 days, but only 0.12 mmol of methane was produced. Acetate and butyrate were the most important VFA formed and amounted to 120 and 25 ^imol, respectively.
Table 1 presents the results of methane and VFA production from methanol at several pH values using an enrichment culture obtained after 4 transfers at pH 4.5. At low pH (4 to 5), methanogenesis proceeded very well, and almost all of the methanol was converted into methane. VFA was also produced but in minor amounts. Microscopical observations
86
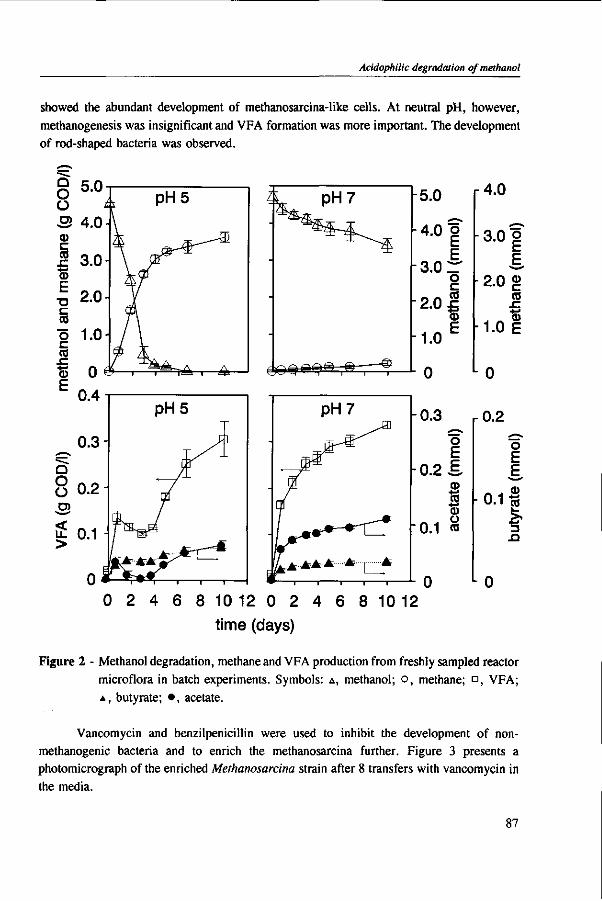
Acidophilic degradation of methanol
showed the abundant development of methanosarcina-like cells. At neutral pH, however, methanogenesis was insignificant and VFA formation was more important. The development of rod-shaped bacteria was observed.
0 2 4 6 8 10 12 0 2 4 6 8 10 12 time (days)
Figure 2 - Methanol degradation, methane and VFA production from freshly sampled reactor microflora in batch experiments. Symbols: A, methanol; o, methane; n, VFA; A, butyrate; • , acetate.
Vancomycin and benzilpenicillin were used to inhibit the development of non-methanogenic bacteria and to enrich the methanosarcina further. Figure 3 presents a photomicrograph of the enriched Methanosarcina strain after 8 transfers with vancomycin in the media.
87
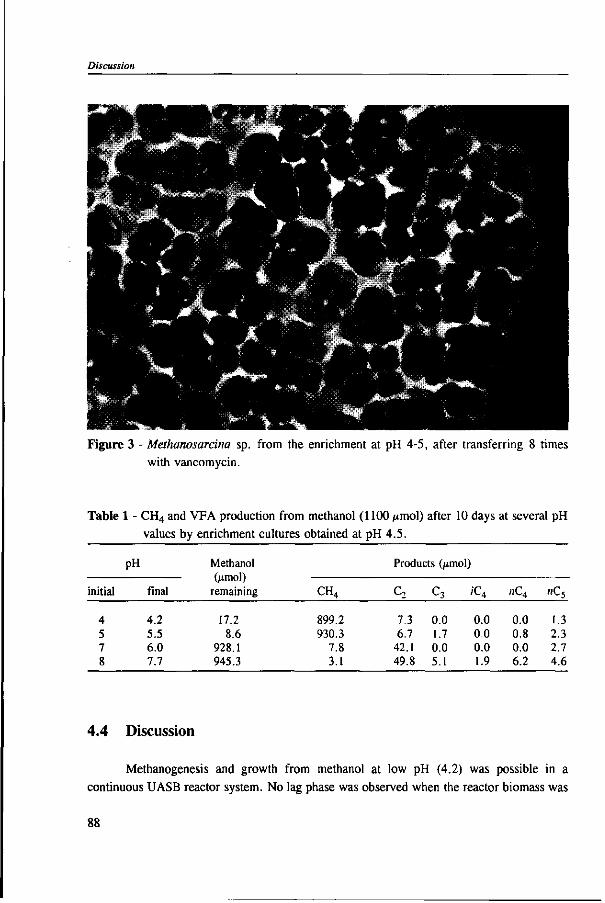
Discussion
Figure 3 - Methanosarcina sp. from the enrichment at pH 4-5, after transferring 8 times with vancomycin.
Table 1 - CH4 and VF A production from methanol (1100 /tmol) after 10 days at several pH values by enrichment cultures obtained at pH 4.5.
initial
4 5 7 8
pH
final
4.2 5.5 6.0 7.7
Methanol (/tmol)
remaining
17.2 8.6
928.1 945.3
CH4
899.2 930.3
7.8 3.1
Products (/iinol)
c2
7.3 6.7
42.1 49.8
c3
0.0 1.7 0.0 5.1
«C4
0.0 0.0 0.0 1.9
«c4
0.0 0.8 0.0 6.2
«C5
1.3 2.3 2.7 4.6
4.4 Discussion
Methanogenesis and growth from methanol at low pH (4.2) was possible in a continuous UASB reactor system. No lag phase was observed when the reactor biomass was

Acidophilic degradation of methanol
immediately transferred from the reactor to enrichment bottles at pH 4 to 5. A bacterium with Metnanosarcina-like morphology was the main methanogen responsible for the conversion of methanol to methane at low pH. The enrichment cultures were obligate acidophiles since they did not produce methane nor grow at neutral pH values. This is distinct from the previously reported acid tolerant Methanosarcina strains which were active over a broad pH range of 4.3-7.8.8 However, the Methanosarcina-like strain of our study had specific growth rates at pH 5 which were similar to those reported for the Methanosarcina barkeri strains.8
Alkalinity free medium did not contain sodium. Sodium is an essential ion for all methanogens.i l Energy conservation occurs by means of a sodium gradient across the cytoplasmic membrane.2 In the present study, sodium was required for the acidophilic methylotrophic methanogens.
Feed interruptions in the continuous system and substrate depletion in batch experiments led to the inactivation of methanogens at low pH. Probably the bacteria growing at low pH values require a high energy level for maintenance, as energy is needed for the regulation of the internal pH. When acidophilic microorganisms are not able to pump out protons, acidification of the cytoplasma takes place, leading to an irreversible inactivation of the bacteria.3
Acidophilic methylotrophic methanogenesis offers an attractive perspective for anaerobic wastewater treatment. Methanolic wastewaters can potentially be treated without any alkalinity additions (e.g. expensive NaHC03). Moreover, the acidophiles have an astonishingly high specific methanogenic activity enabling the conversion of 13 g COD • l"1
• d"1 to methane during continuous operation at pH 4.2. On the other hand, it should be kept in mind that the decay rate of the methanogens involved is apparently high, which means that feed interruptions may lead to system upsets. More research is required to find measures for solving these problems.
4.5 Acknowledgements
We would like to thank Sonja Viester for conducting part of the experimental work. Financial aid from the Brazilian Government, Ministry of Education-CAPES, and The European Environmental Research Organization, The Netherlands, are gratefully acknowledged.
89

References
4.6 References
1. American Public Health Association(1985) Standard methods for examination of water and wastewater, 16th ed. American Public Health Association, Washington, D.C., USA.
2. Blaut, M.,V. Müller, K. Fiebig, and G. Gottschalk (1985) Sodium ions and an energized membrane required by Methanosarcina barken for the oxidation of methanol to the level of formaldehyde. J. Bacteriol. 164:95-101.
3. Booth, I. R. (1985) Regulation of cytoplasmic pH in bacteria. Microbiol. Rev. 49:359-378.
4. Field, J., M. J. H. Leyendeckers, R. Sierra-Alvarez, G. Lettinga, and L. H. A. Habets (1991) Continuous anaerobic treatment of autoxidized bark extractes in laboratory-scale columns. Biotech, and Bioeng. 37:247-255.
5. Hoover, S. R., and N. Porges (1952) Assimilation of dairy wastes by activated sludge. II: The equation of synthesis and oxygen utilization. Sewage and Ind. Wastes 24:306-312.
6. Huser, B. A., K. Wuhrmann, and A. J. B. Zehnder (1982) Methanothrix soenhgenii gen. nov. sp. nov., a new acetotrophic non-hydrogen oxidazing methane bacterium. Arch. Microbiol. 132:1-9.
7. Luria, S. L. (1960) The bacterial protoplasm: composition and organization, p. 1-34. In: Cunsalus, I. C. and R. Y. Stanier (eds). The Bacteria. Vol. 1. Academic Press, New York, USA.
8. Maestrojuân, G. M., and D. R. Boone (1991) Characterization of Methanosarcina barken MST and 227, Methanosarcina mazei S-6T, and Methanosarcina vacuolata Z-761T. Int. J. Syst. Bacteriol. 41:267-274.
9. Minami, K., T. Horiyama, M. Tasaki, and Y. Tanimoto (1986) Methane production using a bio-reactor packed with pumice stone on an evaporator condensate of a kraft pulp mill. J. Ferment. Technol. 64:523-532.
10. Patel, G. B., G. D. Sprott, and J. E. Fein (1990) Isolation and characterization of Methanobacterium espanolae sp. nov., a mesophilic, moderately acidophilic methanogen. Int. J. Syst. Bacteriol. 40:12-18.
11. Perski, H. J., P. Schönheit, and R. K. Thauer (1982) Sodium dependence of methane formation in methanogenic bacteria. FEBS Lett. 143:323-326.
12. Speece, R. E (1983) Anaerobic biotechnology for industrial wastewater treatment. Environ. Sei. Tech. 17:416A-427A.
13. Stams, A.J.M., K. C. F. Grolle, C. T. M. J. Frijters, and J. B. van Lier (1992) Enrichment of thermophilic propionate-oxidizing bacteria in syntrophy with Methanobacterium thermoautotrophicum or Methanobacterium thermofomicicum. Appl. Environ. Microbiol. 58:346-352.
14. Williams, R. T., and R. L. Crawford (1984) Methane production in Minnesota peatlands. Appl. Environ. Microbiol. 47:1266-1271.
15. Williams, R. T., and R. L. Crawford (1985) Methanogenic bacteria, including an acid-tolerant strain, from peatlands. Appl. Environ. Microbiol. 50:1542-1544.
90

Substrate competition between methanogens and acetogens during the anaerobic degradation
of methanol in UASB reactors
Abstract
During the continuous biodégradation of methanol by natural mixed cultures in UASB granular sludge, methylotrophic methanogens and acetogens compete aggressively for the substrate. Volatile fatty acid (VFA) formation by acetogens is undesirable from a wastewater treatment point of view since little chemical oxygen demand (COD) removal is achieved by this conversion. Moreover, VFA accumulation causes reactor instability due to pH decreases in weakly buffered systems. On the other hand, methane formation by methanogens is responsible for COD removal. To evaluate the effect of methanol concentration and inorganic carbon on the competition between acetogens and methanogens for methanol, eight UASB reactors were operated continuously with different levels of sodium bicarbonate at variable methanol loadings. The results indicated that methylotrophic methanogens will predominate in a broad pH range (4.2 to 7.2) if either the reactor methanol concentration or inorganic
Submitted for publication.
91

Introduction
carbon is low. Continuous steady state methanogenesis from methanol resulted in COD removals of 99.8 % to 54% at organic loadings ranging from 7.6 to 22 g COD • l"1 • d"1. On the other hand, significant acetogenesis occurred if exogenous inorganic carbon was supplied and if the reactor methanol concentration was greater than 1000 mg COD • l"1. Inorganic carbon is a cosubstrate required by acetogens in order to carboxylate methanol. The meagre endogenous resources of inorganic carbon released by methanogenesis were not sufficient to support significant acetogenesis. The observation that acetogenesis only occurred in the presence of high reactor methanol concentrations confirms the previously determined 60-times-higher substrate affinity of methanogens. Stable steady state acetogenesis was only observed when unionized VFA, which inhibited the methanogens, were present at high levels for prolonged periods of time. Additions of moderate levels of NaHC03 (15 meq • l"1) were found to create such conditions since VFA accumulation exceeded the buffer capacity. Continuous acetogenesis from methanol at organic loading rates of 21 g COD • l"1 • d"1
resulted in poor COD removal (16.3%). In contrast, high levels of NaHC03 supplementation ( > 25 meq • l"1) favoured methanogenesis since the alkalinity was strong enough to prevent toxicity levels of undissociated acids even in periods when VFA accumulated as a result of organic overloading.
5.1 Introduction
Methanol is an important organic pollutant present in some industrial wastewaters. In anaerobic environments, methanol can be utilized by methanogens and acetogens.10'14 In wastewater treatment plants, the conversion of methanol into methane is a prerequisite because this conversion is responsible for chemical oxygen demand (COD) removal, whereas with the formation of volatile fatty acids (VFA) little COD removal is achieved. Moreover, the accumulation of VFA can lead to reactor instability due to pH drops12'13 in weakly buffered system.
The formation of VFA from methanol has either been associated with the presence of trace elements and bicarbonate in the medium12,13 or with nutrient (nitrogen) deficiency.17
Trace element concentration is one of the environmental factors that may influence the competition between methanogens and acetogens in anaerobic mixed cultures degrading methanol. We have studied the effect of trace elements on the degradation of methanol.6
Among the trace elements, only cobalt was found to greatly stimulate methane and acetate production from methanol.8 In a cobalt-deprived bioreactor, methane production was still evident while less acetate was formed, indicating that methanogens can outcompete acetogens under cobalt limitation. However, other factors that lead to the dominance of methanogens over acetogens still need to be more fully understood.
92

Methanol competition in UASB reactor
The concentration of the available substrate is one of the most important factors that influence the competition between methanogens and acetogens. Thus, the effective methanol concentration inside the reactor may play an important role in the final fate of methanol. In a wastewater treatment plant, the effluent concentration should be kept low. In such an environment, methanogens are favoured over acetogens due to their 60-times-higher substrate affinity for methanol, although acetogens have a slightly higher specific growth rate than methanogens.8
Bicarbonate plays an important role in the anaerobic conversion of methanol, not only as a weak acid in the pH buffering system but also it is a required co-substrate in the acetogenic breakdown of methanol. Bicarbonate is produced when methanol is directly converted into methane. On the other hand, it is consumed when acetate or butyrate are produced by the acetogenesis from methanol (Table 1). Thus, anaerobic methanol conversions involve the production or consumption of bicarbonate. The formation of acids is limited by the availability of inorganic carbon in the system, that will always be present at least to a limited extent when methanogenesis occurs. According to the stoichiometry, for every two moles of methane produced from methanol only one mole of acetic acid can be formed when no exogenous C02 is added. In other words, only up to one third of the methanol can be consumed by acetogens. This interrelationship between inorganic carbon consumption and VFA production directly affects the buffering capacity of the reactor. If insufficient buffering capacity is present, the eventual production of acids will obviously decrease the pH.
In view of the limited information available about the substrate competition between methanogens and acetogens during the anaerobic degradation of methanol, the objective of this study was to evaluate the effect of inorganic carbon, alkalinity addition and reactor methanol concentration on the final fate of methanol in anaerobic reactors. For this purpose, eight UASB reactors were operated continuously with different levels of bicarbonate using a mineral medium with methanol as the only source of organic carbon.
Table 1 - Methane, acetate and butyrate formation from methanol.
Reaction
4 CH3OH - 3 CH4 + HCO3- + H + + H20 4 CH3OH + 2 HCO3- - 3 CH3COO- + H+ + 4 H20 10 CH3OH + 2 HCO3- - 3 C3H7COO- + H + + 10 H20
References
16 9 9
93

Materials and methods
5.2 Materials and methods
Biomass
The methanogenic granular sludge used in these experiments was obtained from a full-scale UASB reactor treating alcohol distillery wastewater of Nedalco at Bergen op Zoom, The Netherlands. The sludge was elutriated to remove the fines. Total suspended solids (TSS) and volatile suspended solids (VSS) content of the sludge were 10.5% and 9.14%, respectively.x
Basal medium and chemicals
Unless otherwise stated, the inorganic macro-nutrients used in all experiments contained (in milligrams per litre of basal medium): NH4C1 (280), K2HP04 • 3H20 (330), MgS04 • 7H20 (100), CaCl2 • 2H20 (10), FeCl2 • 4H20 (2), H3B03 (0.050), ZnCl2
(0.050), MnCl2 • 4H20 (0.5), CuCl2 • 2H20 (0.038), (NH4)6Mo7024 • 4H20 (0.050), A1C13
• 6H20 (0.090), NiCl2 • 6H20 (0.142), Na2Se03 • 5H20 (0.164), CoCl2 • 6H20 (2), EDTA (1), resazurin (0.2), and 36% HCl (0.001 ml • l"1). The medium was made up in demineralized water. Bicarbonate (NaHC03) or phosphate (K2HP04) buffers were added as outlined in Table 2. For the bicarbonate free reactors (C and H), NaCl (1000 mg • l"1) was added to the basal medium.
All chemicals were of analytical grade and were supplied by E. Merck AG, Darmstadt, Germany, except for resazurin, which was purchased from Fluka Chemie AG, Buchs, Switzerland.
Table 2 -
Alkalinity (meq • 1"''
NaHC03
K2HP04c
Bicarbonate and phosphate added to the reactors.
Aa B
30 30 0 0
C
0 0
Reactor D E
15 50 0 0
F
20-50b
0
G
2.5-50b
0
H
0 20
a No cobalt was added in the medium. b See Figures 3 and 4. c Present as nutrient at 1.46 mM.
94

Methanol competition in UASB reactor
Experimental set up
The reactors were operated in a temperature controlled room at 30 ± 2 °C. The Plexiglass and glass cylindrical UASB reactors had a working volume of 2.5 1 and 0.1 1, respectively.7'8 No mechanical mixing was applied in the reactors. The reactors were inoculated with 20 g VSS per liter reactor anaerobic Nedalco granular sludge. The influent contained methanol as the sole carbon source at concentrations ranging from 4.8 to 17.2 g COD • l"1. The influent solution for all reactors contained sufficient macronutrients (N, P, and S) and trace elements required for bacterial growth as outlined in Basal Media and Chemicals with the exception of reactor A, that was deprived of cobalt addition.6 Methane production was monitored with a wet gasmeter6 or by using a sodium hydroxide solution displacement system.5 Bicarbonate and phosphate buffers were applied in the concentrations presented in Table 2.
Analysis
The pH was determined immediately after sampling with a Knick 511 pH-meter (Berlin, Germany) and a Schott Geräte N61 double electrode (Hofheim, Germany). Prior to the analysis of methanol and VF A, samples were centrifuged at 17000 g for 5 min. Methanol, VFA and biogas composition were determined by gas chromatography as described elsewhere.6
5.3 Results
To investigate the final fate of methanol degradation under anaerobic conditions, eight UASB reactors were operated continuously at 30°C with methanol as the only organic carbon source.
The operational parameters and methanol conversion during the continuous experiments for the reactors A, B and C were published elsewhere6'7 and only a brief summary is presented in the following paragraph. The operational results from reactors D, E, F, G and H are illustrated in Figures 1 to 5. The average methanol removal, the total COD removal efficiency and the percentage of methanogenesis of the influent COD for reactors D to H are listed in the Table 3.
Reactor A was operated without supplementation of cobalt in the basal media and reactor B was the control reactor, with cobalt supplementation.6 Both reactors were supplied
95
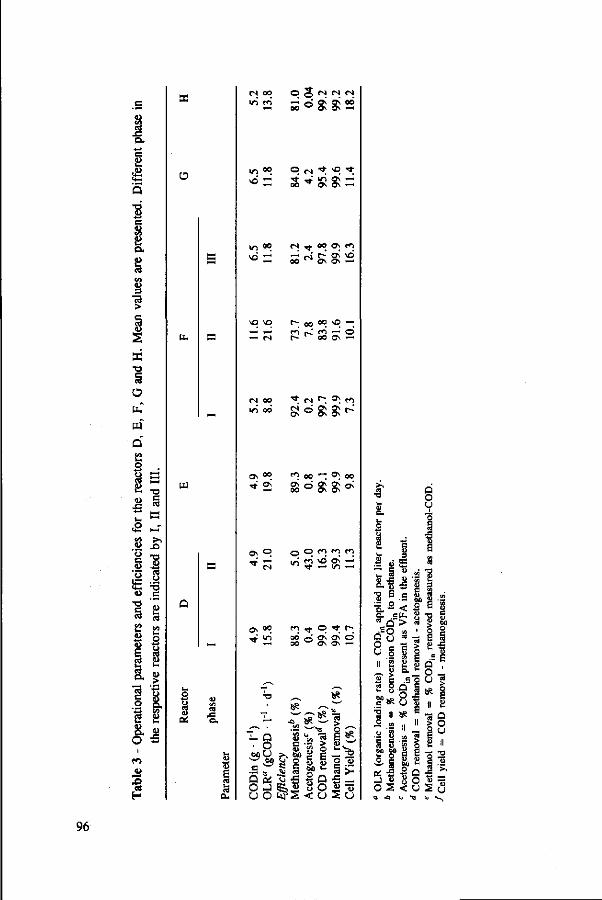
cd J3 O.
4-*
a a ë S i o.
s CO <L> 3
73
•o 5 O
m
9 k i
•* -»
W
o us co O
'S c ' o
!C <L)
T3
S T3
§ • — 4
••H
>* X>
I o
^ G
ti\
CCJ ( 3
S2 ««
E s o .
73 e
_o *+-•
2 * e>
£ S e« H
S !_ 1> >
'•S
S O. co
a <u
o
u.
tu
ÖS
<N 00 O J N N tN — 0O
0 Os Os 00 ° Os Os —
O M • * VO •* Tt-' —: in' os' —' 00 ^ OS Os —
N ^ 00 ON fO ~ M r-' os sd oo °* o\ o\ —
t-. oo m — o oo <ji -
<N 00 in od
os °°
os 9
Os ° °
e s o o s o s I-.'
<^ 00 ~ ^ OO
Q o e i e i e i u-ï e i so o s —' "^ n- — in —•
r-J OS Os O 0 Os Os —
•O ^ « & O ,
-7
^-y—N •
ï - Q • O sou
Q 0 « e 'S
>—' « IA
'vi
c 0) 60 0 e «
0 rf <6>.§
•—s
& 0 Cfl
c <u 00 0
ï8 • s •a >
i 4)
O 3 O < U
73
lg rt >-
•a
o
"I CU "3
•8 e
3 o a ** a c
«a &. u §
« o >- o
O O CJ
•S <U 3
..i - f l».
> e
0 . c
D O U
0
« 1
cd > 0
e « 0
U <u
o _ u « * I f g 1° s s
as 5 0 S
.2 >
il ta) v .
< u
2 75
S
II 2 13
u Q -Q (j t ï
96

Methanol competition in UASB reactor
with 30 meq NaHC03 • l"1. Cobalt greatly stimulated both acetogenesis and methanogenesis
from methanol. However, the production of VFA from methanol under cobalt deprived medium was rather poor and methanogenesis was the main pathway of methanol degradation. Reactor C was operated without addition of alkalinity in the influent.7 Due to the absence of alkalinity, the pH dropped and was quite stable at 4.2. In this reactor, 54% and 8% of the COD applied was converted into methane and VFA, respectively. About 30% of the methanol applied to the reactor was not converted.
The experimental results of Reactor D, supplied with 15 meq • l"1 of NaHC03, are presented in Figure 1. The results can be divided in two distinct phases: I, the methanogenic phase, where methane was the main product of methanol degradation (15 to 75 days) and II, the acetogenic phase (80 to 180 days), when VFA was the main product. In phase I, methane represented about 88% of the methanol utilization. VFA accumulation was low (below 20 mg COD • l"1). The pH was stable at around 6.6. The COD removal efficiency was high and averaged 99%. Phase II started at day 80, when significant accumulation of VFA began. The pH dropped to near 5 and remained below 6 during the next 60 days (till day 140), when the level of NaHC03 in the feed was increased to 50 meq • l"1. During the period of low pH, the undissociated acetate and butyrate concentration in the reactor reached values up to 346 mg COD • l"1 and 341 mg COD • l"1 (5.4 and 3.9 mM), respectively, and averaged 2 mM for each acid over period II. The increase of the NaHC03 concentration to 50 meq • l"1
restored the pH to above 6 but methanogenesis did not recover. In phase II, VFA formation accounted for about 43 % of the COD applied and the COD removal efficiency decreased significantly (16.3%). The methanol concentration in the reactor always exceeded 1000 mg COD • l"1, and methane production remained very low (5%).
Reactor E, operated with 50 meq NaHC03 per liter in the feed, started with an organic loading rate (OLR) of 10 g COD • l"1 • d"1 (Figure 2). At day 18, a 20-hours shock load of 30 g COD • l"1 • d"1 increased the methanol concentration in the reactor for a short period. The production of VFA, that already had started, was greatly enhanced and VFA accumulated. The methanol conversion to methane and the total COD removal efficiency decreased during the period of VFA accumulation (days 20 to 50). The maximum concentration of undissociated acetate was only 0.55 mM (35 mg COD • l"1) and lower than 0.01 mM (1.6 mg COD • l"1) for butyrate. Due to the high amount of alkalinity, the pH remained near neutrality. At around day 40, the organic load was increased to 20 g COD • l"1 • d"1 but the methanol concentration in the reactor remained low and VFA were no longer detectable after day 50. An apparent steady state started and methane was the main product from methanol, averaging 89% of the COD converted.
The operation results from reactor F (Figure 3) can be divided in three phases: I and
97
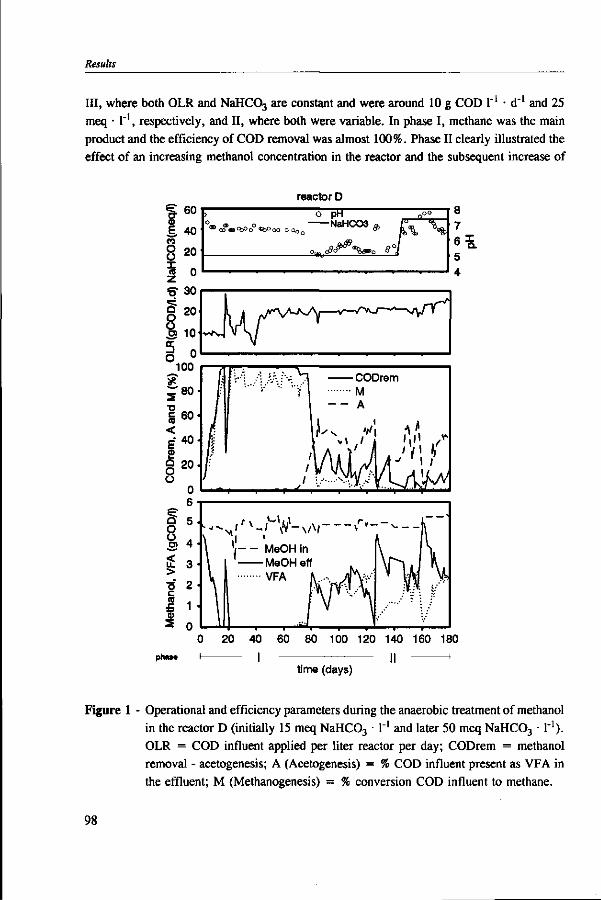
Results
III, where both OLR and NaHC03 are constant and were around 10 g COD l"1 • d"1 and 25 meq • l"1, respectively, and II, where both were variable. In phase I, methane was the main product and the efficiency of COD removal was almost 100%. Phase II clearly illustrated the effect of an increasing methanol concentration in the reactor and the subsequent increase of
reactor D
£. 40
8 20
1 0 W 30
> O pH
° - a?« W«bOoo oo0 o N a H C 0 3 *
o°°
(k \ %
8 7
6£ 5 4
phase
time (days)
Figure 1 - Operational and efficiency parameters during the anaerobic treatment of methanol in the reactor D (initially 15 meq NaHC03 • l"
1 and later 50 meq NaHC03 • l"1).
OLR = COD influent applied per liter reactor per day; CODrem = methanol removal - acetogenesis; A (Acetogenesis) = % COD influent present as VFA in the effluent; M (Methanogenesis) = % conversion COD influent to methane.
98
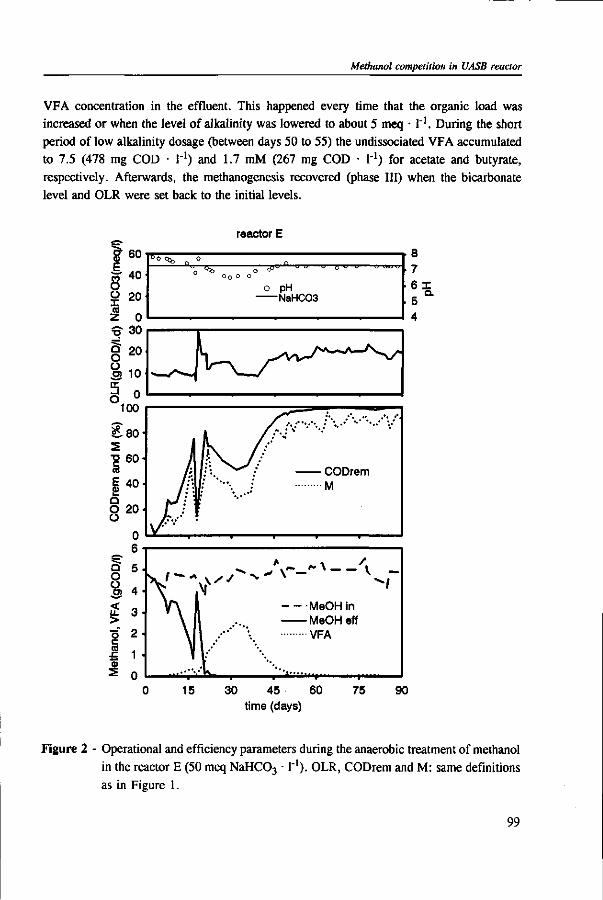
Methanol competition in UASB reactor
VF A concentration in the effluent. This happened every time that the organic load was increased or when the level of alkalinity was lowered to about 5 meq • l"1. During the short period of low alkalinity dosage (between days 50 to 55) the undissociated VFA accumulated to 7.5 (478 mg COD • l"1) and 1.7 mM (267 mg COD • l"1) for acetate and butyrate, respectively. Afterwards, the methanogenesis recovered (phase III) when the bicarbonate level and OLR were set back to the initial levels.
reactor E
30 45 60 time (days)
Figure 2 - Operational and efficiency parameters during the anaerobic treatment of methanol in the reactor E (50 meq NaHC03 • l"
1). OLR, CODrem and M: same definitions as in Figure 1.
99
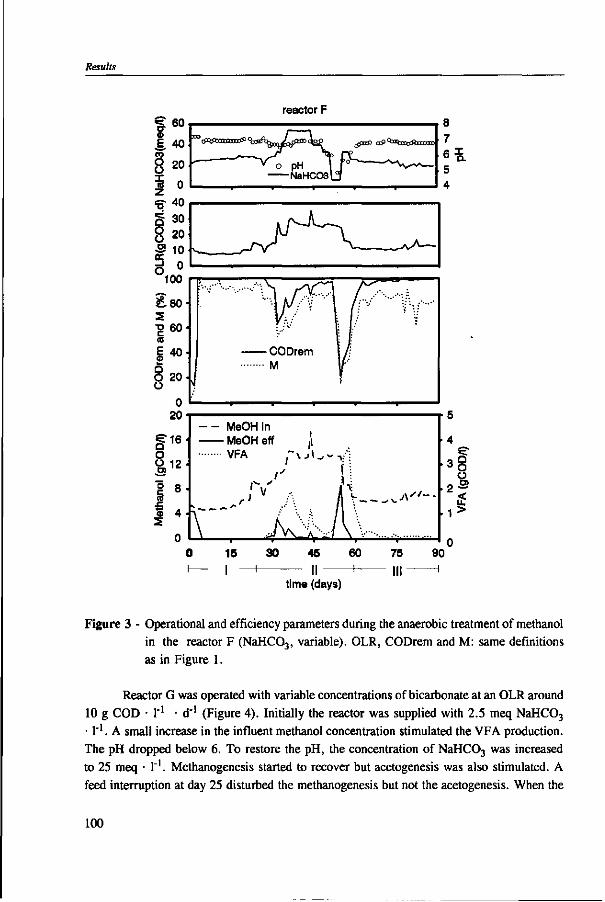
Results
reactor F
time (days)
Figure 3 - Operational and efficiency parameters during the anaerobic treatment of methanol in the reactor F (NaHC03, variable). OLR, CODrem and M: same definitions as in Figure 1.
Reactor G was operated with variable concentrations of bicarbonate at an OLR around 10 g COD • l"1 • d"1 (Figure 4). Initially the reactor was supplied with 2.5 meq NaHC03
• l"1. A small increase in the influent methanol concentration stimulated the VFA production. The pH dropped below 6. To restore the pH, the concentration of NaHC03 was increased to 25 meq • l"1. Methanogenesis started to recover but acetogenesis was also stimulated. A feed interruption at day 25 disturbed the methanogenesis but not the acetogenesis. When the
100
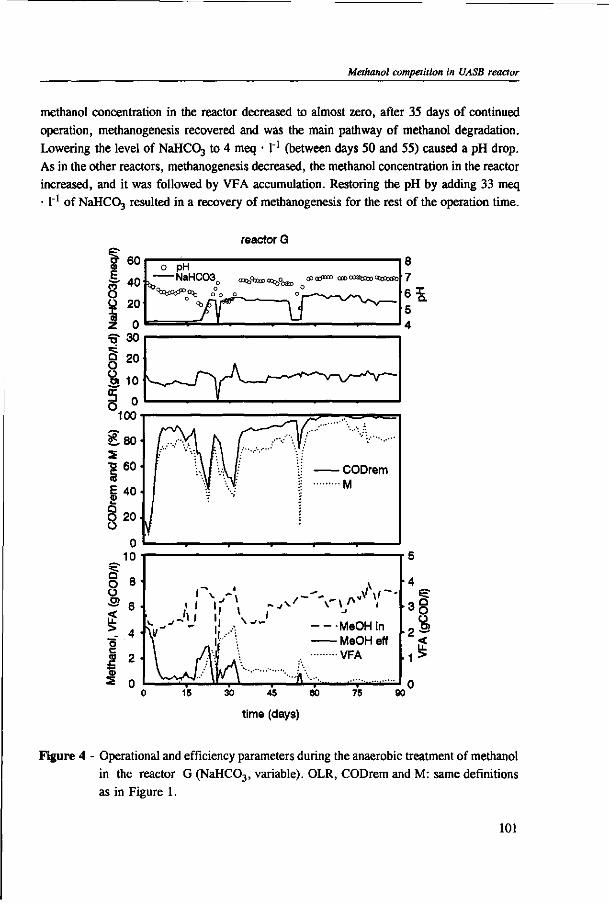
Methanol competition in UASB reactor
methanol concentration in the reactor decreased to almost zero, after 35 days of continued operation, methanogenesis recovered and was the main pathway of methanol degradation. Lowering the level of NaHC03 to 4 meq • l"1 (between days 50 and 55) caused a pH drop. As in the other reactors, methanogenesis decreased, the methanol concentration in the reactor increased, and it was followed by VF A accumulation. Restoring the pH by adding 33 meq • l"1 of NaHC03 resulted in a recovery of methanogenesis for the rest of the operation time.
reactor G
S 8 ü
f6 6 Ô c M
a
4 •
\-\* Ai-
MeOHin MeOH eff VFA
Ä
•4
3 o O
15 30 45 60 75 90
time (days)
Figure 4 - Operational and efficiency parameters during the anaerobic treatment of methanol in the reactor G (NaHC03, variable). OLR, CODrem and M: same definitions as in Figure 1.
101
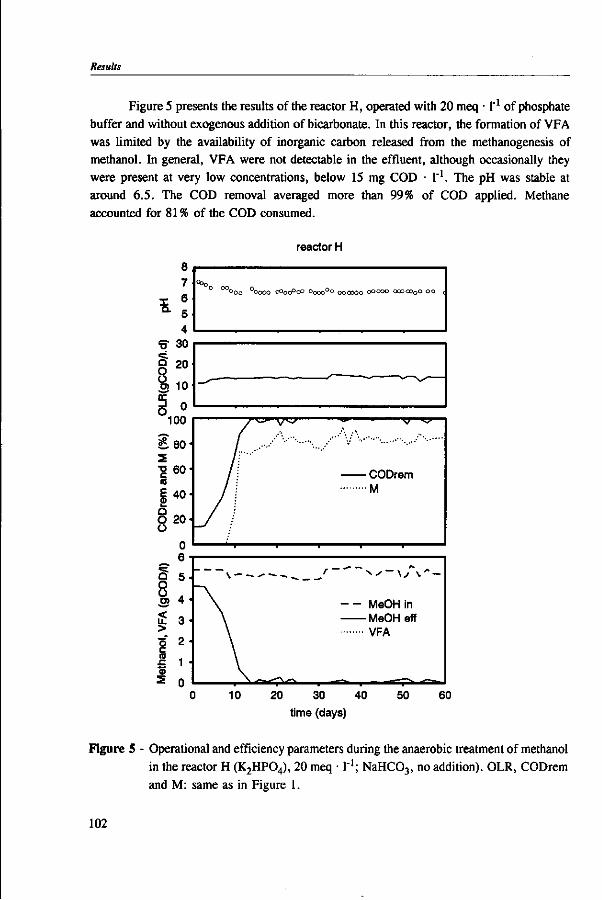
Results
Figure 5 presents the results of the reactor H, operated with 20 meq • l"1 of phosphate buffer and without exogenous addition of bicarbonate. In this reactor, the formation of VFA was limited by the availability of inorganic carbon released from the methanogenesis of methanol. In general, VFA were not detectable in the effluent, although occasionally they were present at very low concentrations, below 15 mg COD • l"1. The pH was stable at around 6.5. The COD removal averaged more than 99% of COD applied. Methane accounted for 81 % of the COD consumed.
reactor H
<*
T3 C « E o Q O Ü
80-
60-
40-
20-
) K-* \J<J
- CODrem •M
MeOH in - MeOH eff
VFA
10 20 30 40 time (days)
50 60
Figure 5 - Operational and efficiency parameters during the anaerobic treatment of methanol in the reactor H (K2HP04), 20 meq • l"1; NaHC03, no addition). OLR, CODrem and M: same as in Figure 1.
102

Methanol competition in UASB reactor
5.4 Discussion
The COD removal efficiency and stability of anaerobic reactors treating wastewaters containing methanol are dependent on which route methanol is degraded. Four important factors may determine the final fate of methanol: methanol concentration in the reactor, the level of exogenous bicarbonate, high concentrations of undissociated VFA and the presence of cobalt in the media.
The effect of methanol concentration
The methanol concentration in the reactor plays an important role in the competition between methylotrophic methanogens and acetogens, provided inorganic carbon is present. Acetogens in our mixed cultures have low substrate affinity for methanol (1^ = 16 mM or 770 mg COD • l"1), compared to methanogens (Ks=0.25 mM or 12 mg COD • l"1).8 The high Ks of acetogens would mean that under the conditions tested, the rate of acetogenesis would have more or less a first order relationship to the reactor methanol concentration. Indeed, when methanol concentration in the reactor was high, the production of organic acids was strongly stimulated whereas, every time that the concentration was low, methane was the main end product (Figure 6), confirming the previously determined 60-fold higher substrate affinity of methanogens.8 Thus, acetogens only have the chance to overcome methanogens when the reactor methanol concentration is high. Values of methanol concentration higher than 1000 mg COD • l"1 seem to be critical for the predominance of acetogens, provided that inorganic carbon and cobalt are supplied to the media, as discussed below.
The effect of exogenous bicarbonate additions and undissociated VFA
Acetogenesis from methanol is only possible if other carbon containing compounds more oxidized than methanol are present, such as acetate, formate, CO and C02 .14 In this study, methanol and bicarbonate were the organic and inorganic carbon sources, respectively. Three levels of exogenous addition of bicarbonate were evaluated: no addition, moderate (15 meq • l"1), and sufficient addition to keep the pH higher than 6 (> 25 meq • l"1).
When no exogenous inorganic carbon was added (reactor C and H), regardless of the pH, methane was the main end product from the anaerobic conversion of methanol. The endogenous inorganic carbon generated by methanogens was not sufficient to support significant acetogenesis, and even so it was not completely used (C02 represented 12% of the biogas). If the endogenous inorganic carbon were all used, the production of acetate or
103

Discussion
butyrate would be around 29 mM (1900 and 4600 mg COD • l"1, for acetate and butyrate, respectively, that depending on the pH, could result in toxicity levels of undissociated VFA.
In reactors where alkalinity was added at moderate levels to the influent (e.g. approximately 15 meq NaHC03 • l"
1), as soon as acetogens started to grow, the eventual production of organic acids caused a pH drop because there was not enough alkalinity to buffer the VFA produced. Low pH itself is not toxic to methylotrophic methanogens.7,15
Indeed, Methanosarcina barken strains have been reported to grow well on methanol in a broad pH range, from 4.3 to 7.8.15 However, the fraction of undissociated VFA increases when the pH decreases. Undissociated VFA has been reported to be toxic to methanogens.2'3,4 For acetoclastic methanogens, the toxic concentration of undissociated acetate lies between 0.26 and 1 mM.3'11 For thermophilic methylotrophic methanogens, complete inhibition was evident at 8.87 mM undissociated acetic acid whereas partial inhibition was found at around 4 mM.18 In reactor D the concentration of undissociated VFA increased up to 5.4 and 3.9 mM for acetate and butyrate, respectively, which might explain why methanogenesis was inhibited. The methanol concentration in the reactor increased. Because bicarbonate and methanol were available, acetogens could grow competitively, and then more VFA were produced. When the concentration of the undissociated VFA in the reactor remained high for prolonged periods (reactor D), methanogens were slowly wiped out. The addition of bicarbonate rose the pH and consequently diminished the fraction of undissociated VFA in the reactor, but recovery back to methanogenesis was no longer possible because by then acetogens had already predominated in the sludge.
In the reactors with enough buffer capacity (NaHC03 >25 meq • l"1), the alkalinity is sufficient to buffer against pH drops when VFA accumulate during occasional overloadings. The pH did not drop and consequently the undissociated VFA fraction was very small. Methanogens were not inhibited by undissociated VFA and when the methanol concentration decreased, methanogenesis readily recovered. Alternatively, acetoclastic methanogens may also develop and consume the produced acetate. However, results of previous studies6'8 indicate that acetoclastic methanogens are not present in significant quantities in the sludge under similar conditions. When the reactor alkalinity dropped for a short period of time (reactor F and G), conditions were temporarily created for acetogens: undissociated VFA accumulated inhibiting methanogens. However, acetogens did not have sufficient time to overtake the population, and as soon as the pH was restored (lowering the undissociated VFA), methanogenesis immediately recovered. Thus, if methanogens are exposed to undissociated VFA for a short period of time (e.g. 5 days in the present study), the reactor will not be seriously upset.
Figure 6 summarizes the relation between methanol concentration in the effluent and
104
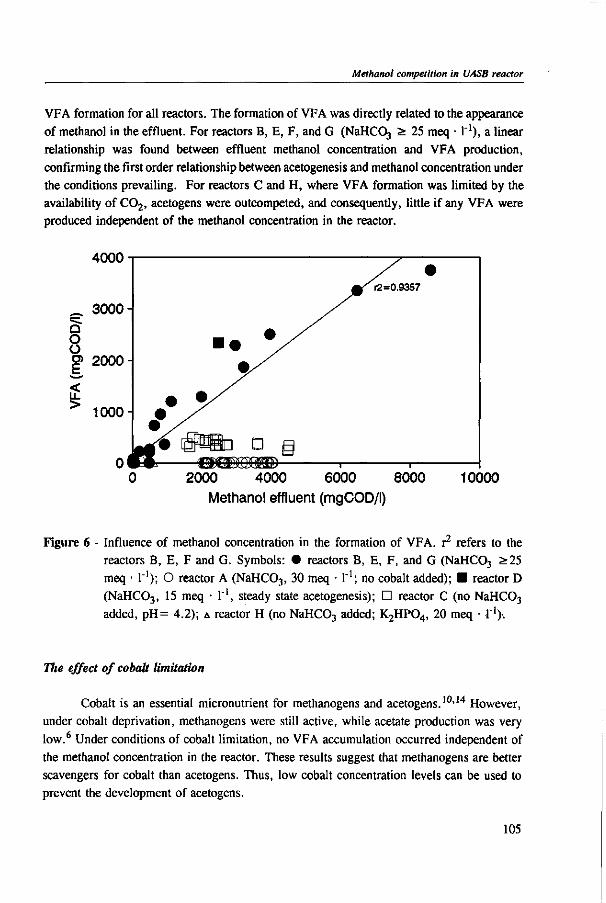
Methanol competition in UASB reactor
VFA formation for all reactors. The formation of VFA was directly related to the appearance of methanol in the effluent. For reactors B, E, F, and G (NaHCO^ > 25 meq • l"1), a linear relationship was found between effluent methanol concentration and VFA production, confirming the first order relationship between acetogenesis and methanol concentration under the conditions prevailing. For reactors C and H, where VFA formation was limited by the availability of C02 , acetogens were outcompeted, and consequently, little if any VFA were produced independent of the methanol concentration in the reactor.
4000
_ 3000
r2=0.9357
2000 4000 6000 8000 Methanol effluent (mgCOD/l)
10000
Figure 6 - Influence of methanol concentration in the formation of VFA. r2 refers to the reactors B, E, F and G. Symbols: • reactors B, E, F, and G (NaHC03 >25 meq • l"1); O reactor A (NaHC03, 30 meq • l"1; no cobalt added); • reactor D (NaHC03, 15 meq • l"1, steady state acetogenesis); • reactor C (no NaHC03
added, pH= 4.2); A reactor H (no NaHC03 added; K2HP04, 20 meq • l"1)-.
The effect of cobalt limitation
Cobalt is an essential micronutrient for methanogens and acetogens.10'14 However, under cobalt deprivation, methanogens were still active, while acetate production was very low.6 Under conditions of cobalt limitation, no VFA accumulation occurred independent of the methanol concentration in the reactor. These results suggest that methanogens are better scavengers for cobalt than acetogens. Thus, low cobalt concentration levels can be used to prevent the development of acetogens.
105

Conclusions
5.5 Conclusions
Table 4 summarizes the final fate of methanol in UASB reactors degrading methanol. Four factors are of importance: the methanol concentration inside the reactor, the cobalt concentration level, the presence of exogenous inorganic carbon and high concentration of undissociated VF A. The results of the present study indicate that methanogens are generally destined to win the competition over acetogens if either the reactor methanol concentration, inorganic carbon content, or the cobalt concentration is low. Significant acetogenesis can only be expected to predominate if the reactor methanol concentration is high, exogenous inorganic carbon is supplied, cobalt is available and methanogens are inhibited, e.g. by undissociated VFA. All these four conditions have to be met. Addition of moderate levels of NaHC03 (approximately 10-20 meq • l"1) were found to create such conditions if the reactor was overloaded.
Table 4 - Influence of the bicarbonate level on the final fate of methanol in the reactors.
NaHC03 added (meq -l"1)
Reactor Final fate of methanol
Remarks
0
15
2:25
30
C, H methane
D VFA
B,C,E,F,G methane
A methane
VFA formation is low and is limited by inorganic carbon. Methanogenesis is not affected by low pH. pH instability due to VFA formation. Toxic level of undissociated VFA for long periods of time wipe out methanogens. Some VFA are formed when reactor is overloaded (e.g. methanol in the effluent) but pH remains high due to high buffer capacity. The undissociated VFA are below toxicity level. Methanogens predominate when reactor is not overloaded. Formation of VFA is low in Co deprived media.
5.6 Acknowledgements
We would like to thank A. van Langerak for her technical assistance. We are also grateful to the Brazilian Ministry of Education for providing a scholarship to L. Florencio (CAPES project 909/88-12) that made this work possible.
106

Methanol competition in UASB reactor
5.7 References
1. American Public Health Association (1985) Standard methods for examination of water and wastewater, 16th ed. American Public Health Association, Washington, D.C., USA.
2. Andrews, J. F. (1969) Dynamic model of the anaerobic digestion process. J. Sanit. Eng. 95:95-116.
3. Attal, A., F. Ehlinger, J. M. Audic, and G. M. Faup (1988) pH inhibition mechanisms of acetogenic acetoclastic and hydrogenophilic populations, p. 71-78. In: E. R. Hall and P. N. Hobson (eds). Anaerobic digestion. Pergamon Press, Oxford, England.
4. Duarte, A. C , and G, K. Anderson (1982) Inhibition modelling in anaerobic digestion. Wat. Sei. Tech. 14:749-763.
5. Field, J. A, M. J. H. Leyendeckers, R. Sierra-Alvarez, G. Lettinga, and L. H. A. Habets (1991) Continuous anaerobic treatment of autoxidized bark extrate in laboratory-scale columns. Biotechnol. Bioeng. 37:247-255.
6. Florencio, L., P. Jenicek, J. A. Field, and G. Lettinga (1993) Effect of cobalt on anaerobic degradation of methanol. J. Ferment. Bioeng. 75:368-374.
7. Florencio, L., A. Nozhevnikova, A. van Langerak, A. J. M. Stams, J. A. Field, and G. Lettinga (1993) Acidophilic degradation of methanol by a methanogenic enrichment culture. FEMS Microbiol. Lett. 109:1-6.
8. Florencio, L., J. A. Field, and G. Lettinga (1994) The importance of cobalt for individual trophic groups in an anaerobic methanol degrading consortium. Appl. Environ. Microbiol. 60:227-234.
9. Heijthuijsen, J. H. F. G., and T. A. Hansen (1986) Interspecies hydrogen transfer in co-cultures of methanol-utilizing acidogens and sulfate-reducing or methanogenic bacteria. FEMS Microbiol. Lett. 38:57-64.
10. Jarrel, K. F., and M. L. Kalmokoff (1988) Nutritional requirements of the methanogenic archeabacteria. Can. J. Microbiol. 34:557-576.
11. Kroeker, E.J., D. D. Schulte, A. B. Sparling, and H. M. Lapp (1979) Anaerobic treatment process stability. J. Water Poll. Control Fed. 51:718-727.
12. Lettinga, G., A. T. van der Geest, S. Hobma, and J. van der Laan (1979) Anaerobic treatment of methanolic wastes. Water Res. 13:725-737.
13. Lettinga, G., W. de Zeeuw, and E. Ouborg (1981) Anaerobic treatment of wastes containing methanol. Wat. Res. 15:171-182.
14. Ljungdahl, L. G. (1986) The autotrophic pathway of acetate synthesis in acetogenic bacteria. Ann. Rev. Microbiol. 40:415-450.
15. Maestrojuân, G. M., and D. R. Boone (1991) Characterization of Methanosarcina barken MST and 227, Methanosarcina mazei S-6T, and Methanosarcina vacuolata Z-
107

References
761T. Int. J. Syst. Bacteriol. 41:267-274. 16. Wood, H. G., W. Ragsdale, and E. Pezacka (1986) The acetyl-CoA pathway of
autotrophic growth. FEMS Microbiol. Rev. 39:345-362. 17. Yamaguchi, M., K. Minami, Y. Tanimoto, and K. Okamura (1989) Effects of
volatile fatty acids on methanogenesis and of pregrowth with methanol on acetate utilization by methanogens. J. Ferment. Bioeng. 68:428-432.
18. Yamaguchi, M., J. Hake, Y. Tanimoto, T. Naritomi, K. Okamura, and K. Minami (1991) Enzyme activity for monitoring the stability in a thermophilic anaerobic digestion of wastewater containing methanol. J. Ferment. Bioeng. 71:264-269.
108

Discussion and Conclusions
6.1 Introduction
Methanol is an important organic pollutant present in several industrial wastewaters and occurs in the environment as a natural compound. In kraft evaporator condensate of pulping mills, methanol is the main organic pollutant and may account for up to 87% of the total chemical oxygen demand (COD).9 Wastewaters originating from condensation processes, do not contain inorganic nutrients and alkalinity. Inorganic nutrients are essential for biological treatment and alkalinity is required for reactor pH stability.
Under anaerobic conditions, methanol potentially supports a complex food chain composed of a variety of possible trophic groups. Methylotrophic methanogens can directly convert methanol into methane. Methylotrophic acetogens produce acetate and butyrate but this conversion is limited by the availability of inorganic carbon. While the conversion of methanol to H2/C02 is usually thermodynamically unfavourable, acetogens are also able to generate H2/C02 from methanol in a syntrophic partnership with hydrogen consumers, e.g. sulfate reducing bacteria2'5 and tetrachloroethene reducing bacteria.3 Thus, methanol can indirectly support hydrogenotrophic and acetoclastic methanogens via acetate or H2/C02
generation by acetogenic bacteria.
109

The effect of cobalt
The COD removal efficiency and stability of anaerobic reactor treating methanolic wastewaters is dependent on which route methanol is degraded. Methane is the target end-product responsible for an effective COD removal, whereas with the production of volatile fatty acids (VFA) little COD removal is achieved. Thus, it is necessary that the acetogenesis is prevented or that eventual VFA accumulations are converted into methane as well. However, the direct conversion of methanol into methane is preferable over the indirect formation via VFA, since methylotrophic methanogens have a 4 to 6 times higher specific methanogenic activity than the acetoclastic methanogens (see Table 3, Chapter 1). Moreover, VFA accumulation may lead to reactor instability in a weakly buffered system.
This dissertation investigates the environmental factors that are of importance in the predominance of methylotrophic methanogens over acetogens in a natural mixed culture during anaerobic wastewater treatment in bioreactors. Technological and microbiological aspect are investigated. Additionally, the route in which methanol is converted into methane is also presented.
Five important factors were found to contribute to the final fate of methanol in anaerobic bioreactors: the presence of cobalt in the media, the reactor methanol concentration, the pH inside reactor, the level of bicarbonate, and high concentrations of undissociated volatile fatty acids.
6.2 The effect of cobalt
In Chapter 2, the effect of trace elements on the methanogenesis of methanol and acetate was studied. Cobalt was the only trace element tested which greatly enhanced methanogenesis from methanol. In contrast, no remarkable influence of any trace element was observed when acetate was used as the substrate. In a continuous experiment, less acetate was formed in a cobalt-deprived reactor than in a cobalt supplemented reactor. These results suggested that methanogens are better scavengers for cobalt than acetogens and that cobalt levels could be used to prevent acetate formation from methanol. It was hypothesized that the cobalt concentration could be used as a parameter for controlling substrate flow during anaerobic treatment of methanolic wastewaters.
The effect of cobalt addition on each individual trophic group potentially involved in the anaerobic conversion of methanol was evaluated in Chapter 3. For this purpose, a sludge was cultivated for one year with methanol in a mineral medium deprived of cobalt. Activity assays with specific inhibitors indicated that methane was being formed directly from methanol and not via the intermediate formation of acetate or H2/C02. The addition of cobalt
110

Discussion and conclusions
stimulated only those trophic groups which directly utilized methanol, while the other trophic groups utilizing downstream intermediates, H2/C02 or acetate, were largely unaffected. Consequently, the influence of increasing cobalt concentrations on the growth rate and specific activity was determined for methylotrophic methanogens and acetogens. At low cobalt concentration, both trophic groups had similar activities and growth rates, whereas at the optimal cobalt concentration, acetogens had a slightly higher values for specific activity and growth. Both trophic groups had similar cobalt optima for growth and activity, with values around 0.05 mg • l"1.
This higher cobalt requirement by methylotrophic microorganisms compared to hydrogenotrophic and acetoclastic methanogens has several technological and microbiological implications. From a technological standpoint, cobalt is an important nutrient that should be considered during the anaerobic treatment of wastewaters with methylotrophic substrates. From a microbiological standpoint, this high requirement for cobalt is presumably due to the production of corrinoids where cobalt is the central ion.10 Methylotrophic methanogens and acetogens are reported to possess a much higher corrinoid (i.e. vitamin B12) content than the same or other microorganisms consuming other substrates (Table 5, Chapter 1). This is probably due to the involvement of unique corrinoid-containing methyltransferases in the initial step of methanol conversion in both methanogens and acetogens.6'7 The metabolism of methanol induces the production of these corrinoid-containing enzymes, such as in the acetogenic bacterium Sporomusa ovata grown on methanol.11'12 In the mixed culture enriched in this study, cobalt addition resulted in a much higher growth rates and specific activities for the methylotrophic microorganisms (Chapters 2 and 3). Presumably, cobalt enhances these rates by stimulating the production of corrinoids. Recently, it has been reported that the addition of vitamin B12 greatly enhanced the biotransformation of tetrachloromethane under anaerobic conditions.4 Therefore, cobalt addition can potentially be used in practice to speed up both methanol and halomethane biodégradation in anaerobic bioreactors.
6.3 The effect of methanol concentration
Since cobalt alone could not explain the predominance of methanogens in our mixed culture, other factors were investigated. The concentration of the available substrate in the bioreactor is another important factor that was considered to possibly influence the competition between methanogens and acetogens. Therefore, substrate affinity coefficients were determined for both of these trophic groups (Chapter 3). Substrate affinity together with growth rate are of importance in the evaluation of substrate competition. Table 1 summarizes the kinetic parameters for the methanogens and acetogens of our mixed culture
111

The effect of methanol concentration
grown on methanol. These results are also illustrated in Figure 6, Chapter 3. Methanogens were found to have a 60-fold-higher affinity for methanol than acetogens. As long as the reactor methanol concentration is lower than about 80 mM, then methanogens will have a faster growth rate than acetogens and, as such, will be expected to outcompete. This prediction was confirmed in continuous experiments where the formation of VFA was directly related to periods when high levels of methanol ( > 1000 mg COD • l"1) occurred in the bioreactors during occasional organic overloadings (Chapter 5). At low cobalt concentrations however (< 0.0001 mg Co • l"1), both populations had similar growth rates indicating that methanogens will easily predominate over acetogens in a much wider methanol concentration range. Therefore, the role of cobalt deprivation is to enhance the competitive edge of methanogens over acetogens.
Table 1 - Growth rate and Ks for methanogens and acetogens grown on methanol and cultivated in a natural mixed culture with specific inhibitors.
Trophic group ixtnax Ks
(mM) + Co
0.3 0.3
(d-1) -Co
1.2 1.0
acetogens 0.3 1.2 16 methanogens 0.3 1.0 0.25
6.4 The effect of exogenous addition of bicarbonate, pH and undissociated volatile fatty acids
The effect of pH inside the reactor was evaluated by applying different levels of alkalinity. In Chapter 4, the anaerobic treatment of methanol without the addition of any alkalinity was investigated. Methanol can successfully be converted into methane, regardless of the low pH value established in the alkalinity-free medium. An astonishing high specific methanogenic activity of 3.57 g COD • g"1 VSS • d"1 was evident, which enabled the conversion of 13 g COD • l*1 • d"1 to methane during continuous operation at pH 4.2. However, significant methanogenesis only occurred after addition of NaCl indicating that sodium was a required nutrient. A Methanosarcina-like organism was the main methanogen responsible for the conversion of methanol to methane. However, the reactor methanogenic activity was very sensitive to accidental interruptions in the feed. Additionally, substrate depletion in batch experiments led to the inactivation of methanogens at low pH. These
112

Discussion and conclusions
results suggested that the decay rate of the acidophilic methanogens involved is apparently high, which means that feed interruption may lead to system upset. Since sodium and substrate are required, methanogens growing at low pH probably require a high energy level for maintenance, as energy is needed for pumping protons out of the cell cytoplasma in order to regulate the intracellular pH. Future research should investigate whether rising the pH during feed interruptions can prevent the die-off of acidophilic methylotrophic methanogens.
Bicarbonate plays an important role in the anaerobic conversion of methanol, not only as a weak acid in the pH buffering system but it is also a required cosubstrate in the acetogenic breakdown of methanol. Bicarbonate is produced when methanol is converted into methane. According to the stoichiometry, up to one third of the methanol can potentially be consumed by acetogens from the endogenous methanogenic supplied bicarbonate. For the complete conversion of methanol to acetate, exogenous bicarbonate must be added. The total requirement of bicarbonate (endogenous plus exogenous) is 0.64 g HC03" • g"1 COD for the complete conversion of methanol into acetate.
Without exogenous addition of bicarbonate, methanogens are the predominant trophic group because the meagre endogenous resources of bicarbonate generated by methanogenesis cannot support significant acetogenesis. When exogenous bicarbonate is applied, more favourable conditions are created for the development of acetogens. If insufficient buffering capacity is present, the eventual production of C02 and VF A decreases the pH. Low pH itself is not toxic to methylotrophic methanogens (Chapter 4). However, the fraction of undissociated VFA increases when the pH decreases due to VFA production by acetogens. Undissociated VFA exerts toxicity to methanogens. When the concentration of undissociated VFA remains high for prolonged periods, methanogens are slowly wiped out and acetogens predominate in the biorectors. If sufficient buffering capacity is present, the eventual production of VFA during occasional overloadings will not decrease the pH and, consequently, the undissociated VFA fraction will be too small to significantly disturb the methanogens.
6.5 Conclusions and Recommendations
Up to date, the anaerobic treatment of methanolic wastewaters has been considered troublesome due to undesirable accumulation of VFA. Four factors are of importance in the predominance of acetogens or methanogens: the methanol concentration inside the reactor, the cobalt concentration level, the presence of exogenous inorganic carbon and high concentration of undissociated VFA.
113

Conclusions and recommendations
Figure 1 presents a simple conceptual model based on the findings of this dissertation. Methanogens will predominate if either the reactor methanol concentration, inorganic carbon content or the cobalt concentration is low. Moreover, methanol is converted directly to methane by methylotrophic methanogens and not via the intermediate formation of VF A. On the other hand, significant acetogenesis can only be expected to predominate if the reactor methanol concentration is high, exogenous inorganic carbon is supplied, cobalt is available and methanogens are inhibited, e.g. by undissociated VFA. All four conditions have to be met.
METHANOL
low [methanol] or
low [cobalt] or
low [C02]
and
low [undissociated VFA]
{ METHANE
high [methanol]
and
high [cobalt]
and
high [COz]
and high [undissociated VFA] for prolonged period
\
ACETATE
Figure 1 - The final fate of methanol in anaerobic bioreactors.
Depending on the target end-product, different measures have to be taken for the predominance of methanogens or acetogens. For methane production, some measures are presented below for the complete predominance of methanogens. During the start-up period, low levels of cobalt should be applied. Additionally, the methanol concentration in the reactor should be kept low by underloading the reactor. Once methanogenesis has predominated, a higher organic load rate can be applied by stimulating the methanogens with
114

Discussion and conclusions
cobalt supplementation. In order to prevent the accumulation of undissociated VFA, high levels of alkalinity should also be applied. Much care should be taken when NaOH is used to increase the pH. In the absence of a weak acid necessary for creation of buffering capacity, the addition of NaOH can break the delicate pH balance and accidently cause severe alkaline pH values. NaHC03 is preferred since it supplies alkalinity together with a weak acid. A cheap source of NaHC03 could be obtained by scrubbing the biogas with NaOH. Alternatively, methane production can also achieved without any addition of alkalinity at low pH. However, this latter possibility has yet to be improved before practical application.
For the predominance of acetogenesis, cobalt concentration should be applied at optimal conditions, the reactor methanol concentration should be maintained high by organic overloading and an excess of exogenous bicarbonate must be supplied. Additionally, methanogens have to be inhibited by ensuring that undissociated VFA levels are high. Addition of moderate levels of NaHC03 (approximately 10-20 meq • l"1) were found to create such conditions if the reactor was overloaded. Since dichloromethane and trichloromethane are specific methanogenic inhibitors, acetogens might be expected to predominate in halomethane contaminated sites subject to anaerobic bioremediation. Sludge pasteurization can be also considered, since many methylotrophic acetogens, such as Sporomusa sp.,9 Clostridium CV-AAI,1 and Butyribacteriwn methylotrophicum13 are spore formers.
6.6 Broader Applications
The results obtained in this thesis on the anaerobic degradation of methylotrophic substrates have a broad range of applications. Some examples of applications in environmental biotechnology include: COD removal from methanolic wastewaters, the biodégradation of methoxylated aromatics and halomethanes, and the use of methanol as a cheap cosubstrate. Furthermore, the results of this thesis provide clues to natural processes occurring in nature such as the formation of biogas in acid peats. Methylotrophic methanogens are active over a broad pH range. Thus, biogas production in acid peats may very well be due to the conversion of methylotrophic substrates derived from methyl ethers/esters in natural plant phenolics, hemicellulose, and pectin.
6.7 References
1. Adamse, A.,D., and C. T. M. Velzeboer (1982) Features of a Clostridium, strain CV-AAI, an obligatory anaerobic bacterium producing acetic acid from methanol.
115

Effect van cobalt
Bij anaërobe behandeling van methanolhoudend afvalwater hangt het CZV-verwijderingsrendement en de stabiliteit van de reactor af van de route waarlangs methanol wordt afgebroken. Methaan is het gewenste eind-produkt, verantwoordelijk voor een effectieve CZV-verwijdering, terwijl productie van vluchtige vetzuren (VFA) weinig CZV-verwijdering geeft. Daarom is het noodzakelijk dat acetogenese wordt voorkomen, of dat eventueel gevormde VFA worden omgezet in methaan. De directe omzetting van methanol in methaan geniet echter de voorkeur boven indirecte methaanvorming via VFA, omdat de methylotrofe methanogenen een 4 tot 6 keer grotere specifieke methanogene activiteit hebben dan acetoclastische methanogenen (zie Tabel 3, Hoofdstuk 1). Bovendien kan VFA-accumulatie in slecht gebufferde systemen leiden tot instabiliteit van de reactor.
Dit proefschrift beschrijft onderzoek naar de milieu-factoren die van belang zijn voor overheersing van methylotrofe methanogenen over acetogen in een natuurlijke meng-cultuur, tijdens anaërobe afvalwaterbehandeling in bioreactoren. Zowel technische als microbiologische aspecten zijn onderzocht. Bovendien wordt de afbraakroute van methanol naar methaan opgehelderd.
Vijf belangrijke factoren bepalen het uiteindelijke lot van methanol in anaërobe bioreactoren: de concentratie cobalt in het medium, de methanol-concentratie in de reactor, de pH in de reactor, de concentratie bicarbonaat in het medium en hoge concentraties ongedissocieerde vluchtige vetzuren.
6.2 Effect van cobalt
In Hoofdstuk 2 is het onderzoek naar het effect van sporenelementen op de methanogenese van methanol en acetaat gepresenteerd. Cobalt bleek het enige geteste sporenelement te zijn dat methanogenese van methanol aanmerkelijk versnelde. Wanneer acetaat als substraat werd gebruikt, werd geen significante invloed van enig sporenelement waargenomen. In continue experimenten werd in de cobalt-deficiënte reactor minder acetaat gevormd dan in de cobalt-gevoede reactor. Deze resultaten suggereren dat methanogenen betere cobalt-gebruikers zijn dan acetogenen en dat de cobalt-concentratie mogelijk kan worden gebruikt om acetaatvorming uit methanol te voorkomen. Als hypothese wordt gesteld dat de cobalt-concentratie kan worden gebruikt als parameter om de afbraakroute van methanol, tijdens anaërobe behandeling van methanolhoudend afvalwater, te regelen.
In Hoofdstuk 3 wordt het effect van cobalt op iedere metabole populatie, betrokken bij de anaërobe omzetting van methanol, geëvalueerd. Voor dit doel werd gedurende één jaar slib gekweekt op methanol en mineraal medium zonder cobalt. Activiteits-testen met
118

Discussie en conclusies
specifieke remmers gaven aan dat methaan direct werd gevormd uit methanol, en niet via tussentijdse vorming van acetaat of H2/C02. De toevoeging van cobalt stimuleerde slechts die metabole populaties, die direct methanol gebruikten. De bacteriën die H2/C02 of acetaat gebruiken, werden nauwelijks beïnvloed. Daaropvolgend werd de invloed van toenemende cobalt-concentraties op de groei-snelheid en de specifieke activiteit van methylotrofe methan-ogenen en acetogenen bepaald. Bij lage cobalt-concentraties hadden beide metabole populaties vergelijkbare groei-snelheden en specifieke activiteiten. Bij optimale cobalt-concentraties echter, hadden de acetogenen iets grotere groei-snelheden en specifieke activiteiten. Beide metabole populaties hadden vergelijkbare cobalt-optima voor groei en activiteit, met waarden rond 0,05 mg • l"1.
De hogere cobalt behoefte van de methylotrofe bacteriën, vergeleken met hydroge-notrofe bacteriën acetoclastische methanogenen, heeft verscheidene technologische en microbiologische implicaties. Vanuit technologisch standpunt is cobalt een belangrijke nutriënt die in beschouwing moet worden genomen wanneer afvalwater met methylotrofe substraten moet worden behandeld. Vanuit microbiologisch standpunt is deze hoge cobalt behoefte het gevolg van de produktie van corrinoïden, waarin cobalt het centale ion is.10 Uit de literatuur blijkt dat methylotrofe methanogenen en acetogenen een veel hoger gehalte aan corrinoïde (vitamine B12) hebben dan de dezelfde of andere micro-organismen die andere substraten consumeren (Tabel 5, Hoofdstuk 1). Dit wordt waarschijnlijk veroorzaakt door de aanwezigheid van een uniek corrinoïde-bevattend methyltransferase dat betrokken is bij de initiële stap van methanol-omzetting in zowel methanogenen als acetogenen.6'7 Het metabolisme van methanol induceert de produktie van deze corrinoïde-bevattende enzymen, zoals ondermeer is aangetoond in de acetogene bacterie Sporomusa ovata, groeiend op methanol.11'12 In de meng-cultuur opgehoopt in het onderzoek van dit proefschrift, leidde toevoeging van cobalt tot een veel hogere groei-snelheid en specifieke activiteit van de methylotrofe micro-organismen (Hoofdstuk 2 en 3). Waarschijnlijk vergroot cobalt deze snelheden door de produktie van corrinoïden te stimuleren. Onlangs is aangetoond dat toevoeging van vitamine B12 de biotransformatie van tetrachloormethaan onder anaërobe omstandigheden enorm versnelde.4 Daarom kan toevoeging van cobalt mogelijk in de praktijk worden gebruikt om de biologische afbraak van zowel methanol als halomethanen in anaërobe reactoren te versnellen.
6.3 Effect van de concentratie methanol
Omdat de concentratie cobalt niet alléén verantwoordelijk kan zijn voor de dominantie van methanogenen in onze mengcultuur, zijn ook andere factoren onderzocht. Een andere belangrijke factor, die mogelijk de competitie tussen methanogenen en acetogenen kan
119

Effect van concentratie methanol
beïnvloeden, is de beschikbare substraatconcentratie in de bioreactor. Voor beide bovengenoemde metabole populaties is daarom de substraat-affiniteit bepaald (Hoofdstuk 3). De substraat-affiniteit en de groeisnelheid zijn tesamen de belangrijkste factoren voor de evaluatie van substraatcompetitie. Tabel 1 geeft een opsomming van de kinetische parameters van de methanogenen en acetogenen van onze, op methanol gekweekte, mengcultuur. Deze resultaten zijn ook weergegeven in Figuur 6, Hoofdstuk 3. Het blijkt dat de methanogenen een 60-maal zo grote affiniteit voor methanol hebben dan de acetogenen. Zolang de methanol concentratie in de reactor beneden 80 mM blijft, hebben de methanogene bacteriën een hogere groeisnelheid dan de acetogene bacteriën. Hierdoor zullen de methanogenen de acetogenen overgroeien. Dit is bevestigd in continue experimenten, waar de VF A produktie direct viel te relateren aan die perioden waarin overbelasting van methanol ( > 1000 mg CZV • l"1) optrad (Hoofdstuk 5). Bij lage cobalt concentraties (< 0,0001 mg Co • l"1) hebben beide metabole populaties echter vergelijkbare groeisnelheden, waardoor methanogenen gemakkelijk de acetogenen kunnen overgroeien in een veel breder gebied van methanol-concentraties. Cobalt-defficiëntie leidt daarom voornamelijk tot vergroting van het competieve voordeel van methanogenen over acetogene bacteriën.
Tabel 1 - Groeisnelheid en Ks van methanogenen en acetogenen, groeiend in een meng cultuur op methanol met specifieke remmers.
Metabole populatie / i , œ Ks
(mM) + Co
0,3 0,3
(d-1) -Co
1,2 1,0
acetogenen 0,3 1,2 16 methanogenen 0,3 1,0 0,25
6.4 Effect van externe toevoeging van bicarbonaat, pH en ongedis-sociëerde vluchtige vetzuren
Door verschillende niveau's van alkaliteit toe te passen is het effect van de reactor pH onderzocht. In Hoofdstuk 4 is de anaërobe behandeling van methanol-houdend afvalwater onderzocht, zonder toevoeging van alkaliteit. Ondanks de lage pH-waarden van het alkaliteit-vrije medium, kan methanol succesvol worden omgezet in methaan. Een verbazingwekkend hoge specifieke methanogene activiteit van 3,57 g CZV • g"1 VSS "' werd bereikt. Dit resulteerde in een omzettingssnelheid van 13 g CZV • l"1 • d"1 naar methaan, gedurende continue bedrijfsvoering bij pH 4,2. Echter, significante methanogenese trad alleen op wanneer NaCl werd toegevoegd. Natrium was dus een limiterend nutriënt. Een
120

Discussie en conclusies
Methanosarcina-achtige bacterie was de belangrijkste methanogeen, verantwoordelijk voor de omzetting van methanol naar methaan. De methanogene activiteit in de reactor was echter zeer gevoelig voor tijdelijke onderbrekingen van de voeding. Bovendien leidde substraat-uitputting, in batch-experimenten bij lage pH, tot inactivatie van de methanogenen. Deze resultaten suggereren dat de afsterving van de acidofiele methanogenen hoog is, wat betekent dat onderbrekingen in de voeding kunnen leiden tot ontregeling van het zuiveringssysteem. Omdat natrium en substraat nodig zijn, vereisen methanogenen groeiend bij lage pH, blijkbaar veel energie voor onderhoud. Dit lijkt logisch omdat veel energie nodig is om protonen uit het celcytoplasma te pompen voor regulatie van de intracellulaire pH. Toekomstig onderzoek moet uitwijzen of een pH-verhoging tijdens onderbreking van de voeding, de afsterving van acidofiele methylotrofe methanogenen kan voorkomen.
Bicarbonaat speelt een belangrijke rol in de anaërobe omzetting van methanol. Als zwak zuur zorgt het niet alleen voor buffering van het systeem, ook is het nodig als co-substraat in de acetogene afbraak van methanol. Bicarbonaat wordt gevormd bij de omzetting van methanol in methaan. Van het endogeen geproduceerd bicarbonaat, kan volgens de reactie stoichiometric tot één-derde van het methanol worden geconsumeerd door acetogene bacteriën. Voor de volledige omzetting van methanol in acetaat, is toevoeging van extra bicarbonaat noodzakelijk. Voor de volledige omzetting van methanol in acetaat is de totale hoeveelheid benodigd bicarbonaat (endogeen plus extern) 0,64 g HC03" • g"1 CZV.
De geringe endogene bicarbonaat-produktie is niet voldoende voor significante acetogenese. Zonder toevoeging van bicarbonaat zijn de methanogene bacteriën daarom de voornaamste metabole populatie. Als extra bicarbonaat wordt toegevoegd, worden gunstiger omstandigheden voor de acetogenen geschapen. Wanneer onvoldoende buffercapaciteit aanwezig is, verlaagd de eventuele produktie van C02 en VFA de pH. Een lage pH is zelf niet giftig voor methylotrofe methanogenen (Hoofdstuk 4). De fractie ongedissociëerde VFA neemt echter toe bij dalende pH. Deze ongedissociëerde VFA zijn wel giftig voor methanogene bacteriën. Als de concentratie ongedissociërde VFA gedurende lange tijd hoog blijft, spoelen de methanogenen uit en gaan de acetogene bacteriën overheersen. Is wel voldoende buffercapaciteit aanwezig, dan zal de eventuele VFA-produktie bij tijdelijke overbelasting niet de pH doen dalen. Hierdoor zal de fractie ongedissociëerde VFA te klein zijn om de methanogenese wezenlijk te verstoren.
6.5 Conclusies en Aanbevelingen
Door de ongewenste ophoping van VFA, werd tot nu toe de anaërobe behandeling van methanol-houdend afvalwater als lastig ervaren. Vier factoren zijn van belang bij de competi-
121

Conclusies en Aanbevelingen
tie tussen acetogenen en methanogenen: de methanol-concentratie in de reactor, de cobalt-concentratie, de aanwezigheid van exogeen bicarbonaat en hoge concentraties ongedissociëerde VFA.
Figuur 1 geeft een simpel conceptueel model, gebaseerd op de resultaten van dit proefschrift. Methanogenen zullen overheersen als óf de methanol concentratie in de reactor, óf het bicarbonaat-gehalte, óf de cobalt-concentratie laag is. Bovendien zal methanol direct door methylotrofe methanogenen worden omgezet in methaan, zonder tussentijdse vorming van VFA. Aan de andere kant kan significante acetogenese allen worden verwacht wanneer én de concentratie methanol in de reactor is hoog, én exogeen bicarbonaat aanwezig is, én cobalt is aanwezig, én methanogenen worden geremd door bijv. ongedissociëerde VFA. Aan alle vier voorwaarden moet worden voldaan.
Afhankelijk van het gewenste eind-produkt, moeten verschillende maatreglen worden getroffen om methanogenen dan wel acetogenen te bevoordelen. Hieronder worden een aantal richtlijnen gegeven voor methaan-produktie door directe methanogenese. Gedurende de start-periode van de reactor, moet de cobalt-concentratie laag worden gehouden. Bovendien moet door onderbelasting de methanol-concentratie in de reactor laag worden gehouden. Wanneer eenmaal de methanogenese is gaan overheersen, kan de belasting worden verhoogd. Eveneens kan de methanogense worden gestimuleerd door toevoeging van cobalt. Om vorming van ongedissociëeerde VFA te voorkomen, moet ook een hoge alkaliteit worden toegepast. Voorzichtigheid is geboden wanneer NaOH wordt gebruikt om de pH te verhogen. Als er geen bufferend zwak zuur aanwezig is kan toevoeging van NaOH de gevoelige pH-balans verstoren. Toevoeging van NaHC03 geniet de voorkeur omdat zowel alkaliteit als buffercapaciteit wordt toegevoegd. Wassen van het geproduceerde biogas met NaOH, kan een goedkope bron van bicarbonaat zijn. Een alternatieve aanpak om methaan-produktie te verkrijgenis door geen alkaliteit to te voegen en de acidofiele methanogenen te bevorderen. Deze laatste mogelijkheid moet echter nog verder worden onderzocht voordat deze in de praktijk kan worden toegepast.
Voor de overheersing van de acetogene bacteriën moet: de optimale cobalt-concentratie worden toegepast, de methanol-concentratie hoog worden gehouden door overbelasting van de reactor en een overmaat aan exogeen bicarbonaat worden toegevoegd. Bovendien moeten de methanogenen worden geremd door hoge concentraties ongedissociërde VFA. Toevoeging van matige hoeveelheden bicarbonaat (ongeveer 10-20 meq • l"1) creëerde deze omstandigheden. Waar ondissociërde ophoopten. Omdat dichloormethaan en tri-chloormethaan specifieke methanogene remmers zijn, zullen acetogenen waarschijnlijk overheersen bij anaërobe reiniging van plaatsen verontreinigd met halomethanen. Slib-pasteurisatie kan ook van belang zijn, omdat veel methylotrofe acetogenen sporevormers zijn.
122
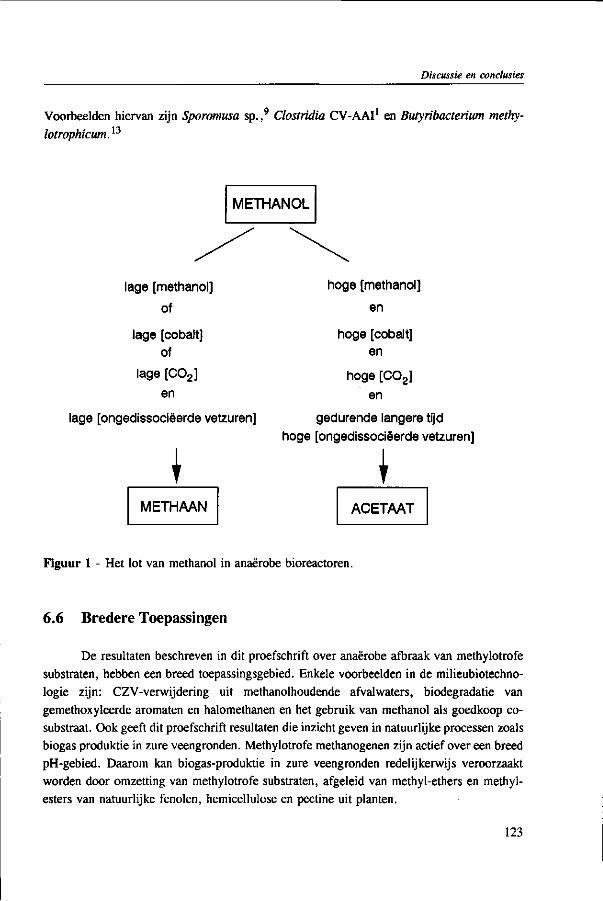
Discussie en conclusies
Voorbeelden hiervan zijn Sporomusa sp.,9 Clostridia CV-AAI1 en Butyribacterium methy-
lotrophicum.13
METHANOL
lage [methanol]
of
lage [cobalt] of
lage [C02]
en
edissociëerde vetzuren]
hoge [methanol]
en
hoge [cobalt] en
hoge [C02] en
gedurende langere tijd hoge [ongedissociëerde vetzuren]
{ { METHAAN ACETAAT
Figuur 1 - Het lot van methanol in anaërobe bioreactoren.
6.6 Bredere Toepassingen
De resultaten beschreven in dit proefschrift over anaërobe afbraak van methylotrofe substraten, hebben een breed toepassingsgebied. Enkele voorbeelden in de milieubiotechnologie zijn: CZV-verwijdering uit methanolhoudende afvalwaters, biodegradatie van gemethoxyleerde aromaten en halomethanen en het gebruik van methanol als goedkoop co-substraat. Ook geeft dit proefschrift resultaten die inzicht geven in natuurlijke processen zoals biogas produktie in zure veengronden. Methylotrofe methanogenen zijn actief over een breed pH-gebied. Daarom kan biogas-produktie in zure veengronden redelijkerwijs veroorzaakt worden door omzetting van methylotrofe substraten, afgeleid van methyl-ethers en methyl-esters van natuurlijke fenolen, hemicellulose en pectine uit planten.
123

Referenties
6.7 Referenties.
1. Adamse, A.,D., en C. T. M. Velzeboer (1982) Features of a Clostridium, strain CV-AA1, an obligatory anaerobic bacterium producing acetic acid from methanol. Antonie van Leeuwenhoek 48:305-313.
2. Cord-Ruwisch, R., en B. Ollivier (1986) Interspecific hydrogen transfer during methanol degradation by Sporomusa acidovorans and hydrogenophilic anaerobes. Arch. Microbiol. 144:163-165.
3. DiStefano, T. D., J. M. Gossett, en S. H. Zinder (1992) Hydrogen as an electron donor for dechlorination of tetrechloroethene by an anaerobic mixed culture. Appl. Environ. Microbiol. 58:3622-3629.
4. Freedman, D. L., en S. Hashsham (1993) Enhanced biotransformation of carbon tetrachloride under methanogenic conditions. Abstract in the proceedings of The second International Symposium - In situ and on-site bioreclamation - . San Diego, California, April 5-8, 1993.
5. Heijthuijsen, J. H. F. G., en T. A. Hansen. (1986) Interspecies hydrogen transfer in co-cultures of methanol-utilizing acidogens and sulfate-reducing or methanogenic bacteria. FEMS Microbiol. Lett. 38:57-64.
6. van der Meijden, P., C. van der Drift, en G. D. Vogels (1984) Methanol conversion in Eubacterium limosum. Arch. Microbiol. 138:360-364.
7. van der Meijden, P., H. J. Heythuysen, H. Sliepenbeek, F. P. Houwen, C. van der Drift, en G. D. Vogels (1984) Activation and inactivation of methanol: 2-mercaptoethanesulfonic acid methyltransferase from Methanosarcina barkeri. J. Bacteriol. 153:6-11.
8. Minami, K., K. Okamura, S. Ogawa, en T. Naritomi (1991) Continuous anaerobic treatment of wastewater from a kraft pulp mill. J. Ferment. Bioeng. 71:270-274.
9. Moller, B., R. Oßmer, B. H. Howard, G. Gottschalk, en H. Hippe (1984) Sporomusa, a new genus of gram-negative anaerobic bacteria including Sporomusa sphaeroides spec. nov. and Sporomusa ovata spec. nov. Arch. Microbiol. 139:388-396.
10. Stupperich, E., H. Eisinger, en S. Schurr (1990) Corrinoids in anaerobic bacteria. FEMS Microbiol. Rev. 87:355-360.
11. Stupperich, E., P. Aulkemeyer, en C. Eckerskorn (1992) Purification and characterization of a methanol-induced cobamide-containing protein from Sporomusa ovata. Arch. Microbiol. 158:370-373.
12. Stupperich, E., en R. Konle (1993) Corrinoid-dependent methyl transfer reactions are involved in methanol and 3,4-dimethyoxybenzoate metabolism by Sporomusa ovata. Appl. Environ. Microbiol. 59:3110-3116.
13. Zeikus, J. G., L. H. Lynd, T. E. Thompson, J. A. Krzycki, P. J. Weimer, en P. W. Hegge (1980) Isolation and characterization of a new, methylotrophic, acidogenic anaerobe, the Marburg strain. Curr. Microbiol. 3:381-386.
124

Discussäo e Conclusöes
6.1 Introduçâo
O metanol se encontra presente tanto em âguas residuârias industrials diversas, resultando em importante poluente orgânico, bem como no meio ambiente na forma de composto de origem natural. Nas âguas residuârias geradas na indüstria papeleira que utiliza o processo de condensaçâo "kraft", o metanol é o principal poluente orgânico, podendo constituir em até 87% da demanda quimica de oxigênio total (DQO).8 As âguas residuârias geradas no processo de condensaçâo nào contém nutrientes inorgânicos nem alcalinidade. Todavia, os nutrientes inorgânicos sâo essenciais para o tratamento biológico e a alcalinidade é necessâria para se manter urn valor de pH estâvel nos reatores.
Em condiçôes anaeróbias o metanol é uma fonte em potencial para uma cadeia alimentär complexa, a quai se compöe de uma variedade de muitos grupos trófïcos. Um destes grupos sâo as bactérias metanogênicas metilotróficas que podem converter o metanol diretamente para metano. Outro grupo sâo as bactérias acetogênicas metilotróficas que produzem acetato e butirato, sendo esta conversào, porém, limitada pela disponibilidade de carbono inorgânico. Embora a conversào de metanol para H2/C02 seja em gérai termodinamicamente desfavorâvel, aquelas bactérias acetogênicas também sâo capazes de produzir H2/C02 a partir do metanol, através de sintrofismo corn as bactérias consumidoras de hidrogênio, cujos exemplos sâo as bactérias redutoras de sulfato2'5 e as bactérias redutoras de tetra-cloro etano.3 Desta forma, o metanol também pode ser uma fonte alimentär para os grupos das bactérias hidrogenotróficas e metanogênicas acetoclâsticas, através da produçâo de acetato ou de H2/C02 produzidos pelas bactérias acetogênicas.
125

O efeito do cobalto
A eficiência de remoçâo da DQO e a estabilidade de um reator anaeróbio, durante o tratamento de âguas residuârias metanólicas, sâo dependentes do caminho em que o metanol seja convertido. O alvo final é o metano, cuja produçâo é responsâvel pela remoçâo efetiva da DQO, ao passo que corn a produçâo de âcidos graxos volâteis (AGV) apenas uma pequena remoçâo da DQO é obtida. Assim, é necessârio que a acetogênese seja evitada ou que os AGV eventualmente acumulados também sejam convertidos para metano. Porém, é prefen'vel a conversâo direta de metanol para metano do que a formaçâo intermediâria de AGV, uma vez que as bactérias metanogênicas metilotróficas têm uma atividade metanogênica especifica de 4 a 6 vezes maior do que a das bactérias metanogênicas acetoclâsticas (ver Tabela 3, Capitulo 1). Além do mais, a acumulaçâo de AGV pode levar à instabilidade do reator num sistema sem um tamponamento adequado.
Nesta tese sâo pesquisados os fatores ambientais importantes na predominância das bactérias metanogênicas metilotróficas em relaçâo as acetogênicas, numa cultura mista natural, durante o tratamento anaeróbio de âguas residuârias em bioreatores. Os aspectos tecnológicos e microbiológicos sâo pesquisados e, adicionalmente, se apresenta o caminho em que metanol é convertido para metano.
Os cinco importantes fatores encontrados que contribuem para o destino final do metanol em reatores anaeróbios sâo: a presença de cobalto no meio, a concentraçâo de metanol no reator, o pH no reator, o ni'vel de bicarbonato e as altas concentraçôes de âcidos graxos volâteis nâo-dissociados.
6.2 O efeito do cobalto
No Capitulo 2 se estudou o efeito dos micronutrientes sobre a metanogênese a partir do metanol e do acetato. O cobalto foi o dnico elemento traço dentre os testados que aumentou significativamente a metanogênese do metanol. Ao contrario, nâo houve qualquer influencia marcante de nenhum dos micronutrientes quando se utilizou o acetato como substrato. Num experimento em sistema continuo formou-se menos acetato no reator corn ausência de cobalto do que no reator com a sua presença. Estes resultados sugerem que as bactérias metanogênicas sâo mais aptas do que as acetogênicas para a utilizaçâo do cobalto, e que as concentraçôes de cobalto podem ser usadas para evitar a formaçâo de acetato a partir do metanol. A hipótese formulada foi que a concentraçâo de cobalto poderia ser usada como um paramètre de controle do fluxo de substrato, durante o tratamento anaeróbio de âguas residuârias metanólicas.
No Capitulo 3 se avaliou separadamente o efeito do cobalto sobre cada grupo trófico
126

Discussäo e conclusöes
que estivesse potencialmente envolvido na conversäo anaeróbia do metanol. Para este objetivo se cultivou lodo com metanol durante um pen'odo de um ano, num meio mineral sem a presença de cobalto. Os resultados de testes de atividade usando inibidores especificos indicam que o metano foi sendo formado diretamente a partir do metanol, e nâo através da formaçâo intermediâria de acetato ou H2/C02. A adiçâo de cobalto estimulou somente aqueles grupos trófïcos que utilizam o metanol diretamente, ao passo que os outros grupos tróficos que utilizam H2/C02 ou acetato como produtos intermediaries, foram afetados muito pouco. Consequentemente, determinou-se a influência de concentraçôes crescentes de cobalto sobre a taxa de crescimento e a atividade especifica, para as metanogênicas metilotróficas e as acetogênicas. No caso de baixas concentraçôes de cobalto, as taxas de crescimento e atividades especi'ficas foram similares para ambos os grupos tróficos, ao passo que no caso de concentraçôes ótimas de cobalto, a taxa de crescimento e a atividade especi'fica das acetogênicas foram levemente maiores. As concentraçôes ótimas de cobalto para ambos os grupos tróficos foram similares, aproximadamente 0,05 mg • l"1.
Esta exigência de mais cobalto por parte dos microrganismos metilotróficos, comparada com a dos hidrogenotróficos e metanogênicos acetoclâsticos, têm diversas implicaçôes tecnológicas e microbiológicas. Do ponto de vista tecnológico, o cobalto é um nutriente importante a ser considerado no tratamento anaeróbio de âguas residuârias contendo substrates para os metilotróficos. Do ponto de vista microbiológico, presume-se que esta maior exigência de cobalto seja dévida à produçâo de corrinoides, nos quais o cobalto é o ion central.10 As metanogênicas metilotróficas e as acetogênicas sâo citadas na literatura, como possuidoras de uma quantidade muito maior de corrinoides (por exemplo, vitamina B12) do que nos mesmos ou em outros microrganismos que consumam outros tipos de substrates (Tabela 5, Capitulo 1). É provâvel que isto seja devido à participaçâo exclusiva de metil transferases que contém corrinoides na fase inicial de conversäo do metanol, tante nas metanogênicas como nas acetogênicas6'7 O metabolismo do metanol induz a produçâo dessas enzimas que contém os corrinoides, como é o caso na bacteria acetogênica Sporomusa ovata, cultivada em metanol.11'12 Os resultados da adiçâo de cobalto na cultura mista utilizada neste estudo foram taxas de crescimento e atividades especi'ficas muito maiores para os microrganismos metilotróficos (Capitulos 2 e 3). Presume-se que o cobalto seja a causa do aumento daquelas taxas dévida à produçâo de corrinoides. Em relate recente, publicou-se que, em condiçôes anaeróbias, a adiçâo de vitamina B12 aumenta significativamente a biotransformaçâo de tetra-cloro metano.4 Por conseguinte, a adiçâo de cobalto pode, em potencial, ser utilizada na pratica para acelerar a biodegradaçào, tante de metanol como de halometanos, em bioreatores anaeróbios.
127
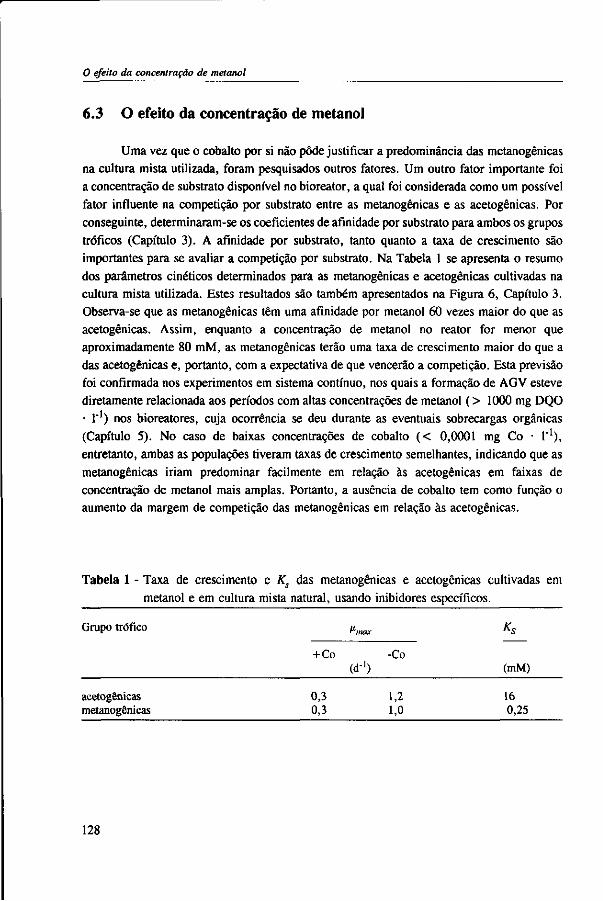
O efeito da concentraçào de metanol
6.3 O efeito da concentraçào de metanol
Uma vez que o cobalto por si näo pôde justificar a predominância das metanogênicas
na cultura mista utilizada, foram pesquisados outros fatores. Um outro fator importante foi
a concentraçào de substrato disponivel no bioreator, a quai foi considerada como um possivel
fator influente na competiçâo por substrato entre as metanogênicas e as acetogênicas. Por
conseguinte, determinaram-se os coeficientes de afinidade por substrato para ambos os grupos
tróficos (Capftulo 3). A afinidade por substrato, tanto quanto a taxa de crescimento sâo
importantes para se avaliar a competiçâo por substrato. Na Tabela 1 se apresenta o resumo
dos paramétras cinéticos determinados para as metanogênicas e acetogênicas cultivadas na
cultura mista utilizada. Estes resultados sâo também apresentados na Figura 6, Capftulo 3.
Observa-se que as metanogênicas têm uma afinidade por metanol 60 vezes maior do que as
acetogênicas. Assim, enquanto a concentraçào de metanol no reator for menor que
aproximadamente 80 mM, as metanogênicas terâo uma taxa de crescimento maior do que a
das acetogênicas e, portanto, com a expectativa de que vencerâo a competiçâo. Esta previsâo
foi confïrmada nos experimentos em sistema continuo, nos quais a formaçâo de AGV esteve
diretamente relacionada aos periodos corn al tas concentraçôes de metanol ( > 1000 mg DQO
• l"1) nos bioreatores, cuja ocorrência se deu durante as eventuais sobrecargas orgânicas
(Capftulo 5). No caso de baixas concentraçôes de cobalto ( < 0,0001 mg Co • l"1),
entretanto, ambas as populaçôes tiveram taxas de crescimento semelhantes, indicando que as
metanogênicas iriam predominar facilmente em relaçâo as acetogênicas em faixas de
concentraçào de metanol mais amplas. Portanto, a ausência de cobalto tem como funçào o
aumento da margem de competiçâo das metanogênicas em relaçâo as acetogênicas.
Tabela 1 - Taxa de crescimento e Ks das metanogênicas e acetogênicas cultivadas em
metanol e em cultura mista natural, usando inibidores especi'ficos.
Grupo trófico nlnax K, s
+ Co -Co (d1) (mM)
acetogênicas 0,3 1,2 16 metanogênicas 0,3 1,0 0,25
128

Discussào e conclusöes
6.4 O efeito da adiçâo externa de bicarbonato, do pH e dos âcidos graxos volâteis nâo-dissociados
O efeito do pH no reator foi avaliado através do uso de diversas concentraçôes de alcalinidade. No Capftulo 4 estudou-se o tratamento anaeróbio de metanol sem a adiçâo de qualquer alcalinidade. A conversào de metanol para metano pode ser obtido com sucesso mesmo em valores baixos de pH, os quais sâo estabelecidos no meio sem nenhuma alcalinidade. A obtençâo de uma impressionante alta atividade metanogênica especifica, de 3,57 g DQO • g"1 VSS • d"1, foi evidente e possibilitou a conversào de 13 g DQO • l"1 • d"1
para metano durante a operaçâo continua em pH 4,2. Entretanto, a significativa metanogênese ocorrida somente após a adiçâo de NaCl, indica que o sódio foi um nutriente necessârio. Urn microrganismo semelhante à Methanosarcina foi a principal metanogênica responsâvel pela conversào de metanol para metano. Entretanto, a atividade metanogênica do reator esteve muito sensîvel as interrupçôes acidentais na alimentaçâo. Em adiçâo, a conversào de substrata nos ensaios em batelada resultou na inativaçào das metanogênicas em pH baixo. Estes resultados sugerem que a taxa de decaimento é aparentemente alta para as metanogênicas acidofflicas envolvidas, o que significa que as interrupçôes na alimentaçâo podem levar o sistema à instabilidade. Uma vez que o sódio e o substrata sâo necessârios, as metanogênicas que se desenvolvem em baixo pH requerem, provavelmente, um alto ni'vel de energia para a sua sobrevivência, necessârio para forçar os protons para o exterior do citoplasma celular para que o pH intracelular seja regulado. As pesquisas futuras deverào investigar se a elevaçâo do pH durante as interrupçôes na alimentaçâo podem evitar a morte das metanogênicas metilotróficas acidofflicas.
A funçâo do bicarbonato é importante na conversào anaeróbia do metanol, nâo somente por agir como um âcido fraco no sistema de tamponamento do pH, mas também porque ele é um co-substrato necessârio na conversào acetogênica do metanol. Quando o metanol é convertido para metano produz-se bicarbonato. De acordo corn a estequeometria, até um terço do metanol pode, em potencial, ser consumido pelas acetogênicas, devido ao bicarbonato endógeno gerado na metanogênese. Para se obter uma conversào compléta de metanol para acetato, bicarbonato exógeno tem ser adicionado. A quantidade total de bicarbonato (endôgena mais exógena), para se obter a conversào compléta de metanol para acetato, é de 0,64 g HC03" • g"1 DQO.
Sem a adiçâo exógena de bicarbonato as metanogênicas serâo os principals grupos tróficos, uma vez que os parcos recursos endógenos de bicarbonato gerados na metanogênese, sâo insuficientes para se ter uma acetogênese significativa. A adiçâo exógena de bicarbonato cria condiçôes favorâveis para o desenvol vi men to das acetogênicas. Caso haja insuficiente capacidade de tamponamento, a resultante produçào de C02 e AGV abaixa o pH. O baixo
129

Conclusöes e recomendaçôes
pH por si proprio näo é tóxico para as metanogênicas metilotrófïcas (Capftulo 4). Entretanto, a fraçâo de AGV nâo-dissociado aumenta quando o pH abaixa, dévida à produçâo de AGV pelas acetogênicas. Os AGV nâo-dissociados exercem toxicidade para as metanogênicas. Quando uma alta concentraçâo de AGV nâo-dissociado se mantém por longos periodos, as metanogênicas sâo eliminadas pouco a pouco e as acetogênicas passam a predominar nos bioreatores. Caso haja sufïciente capacidade de tamponamento, a resultante produçâo de AGV durante as sobrecargas eventuais nâo ira abaixar o pH e, em consequência, a fraçâo de AGV nâo-dissociado sera tâo pequena que nâo ira afetar as metanogênicas.
6.5 Conclusöes e Recomendaçôes
O tratamento anaeróbio de âguas residuârias metanólicas tem sido considerado até o presente como sendo problemâtico, devido à acumulaçâo indesejada de AGV. Quatro fatores sâo importantes para a predominância das acetogênicas ou das metanogênicas: a concentraçâo de metanol no reator, a concentraçâo de cobalto, a presença de carbono inorgânico exógeno e a alta concentraçâo de AGV nâo-dissociado.
A Figura 1 mostra um modelo conceptual simples baseado nos resultados obtidos nesta tese. As metanogênicas predominarâo caso a concentraçâo de metanol, carbono inorgânico ou cobalto seja baixa. Ademais, o metanol é convertido diretamente para metano pelas metanogênicas metilotrófïcas e nâo através da formaçâo intermediâria de AGV. Por outro lado, só se pode esperar a predominância de significativa acetogênese, caso a concentraçâo de metanol no reator seja alta, carbono inorgânico exógeno seja adicionado, cobalto esteja disponivel e as metanogênicas sejam inibidas, por exemplo, por AGV nâo-dissociado. Todas estas quatro condiçôes têm que ser satisfeitas.
Dependendo do alvo como produto final, as medidas serâo distintas para se obter a predominância de metanogênicas ou de acetogênicas. No caso de produçâo de metano, se apresentam a seguir algumas medidas para que prevaleçam as metanogênicas. Durante o perîodo da partida do reator devem ser utilizadas baixas concentraçôes de cobalto. Em adiçâo, a concentraçâo de metanol no reator deve ser mantida baixa, aplicando baixa carga orgânica volumétrica. Quando a metanogênese começar a predominar, pode-se aplicar cargas orgânicas maiores, estimulando as metanogênicas pelas adiçâo suplementar de cobalto. Para se evitar a acumulaçâo de AGV nâo-dissociado deve-se aplicar também maiores concentraçôes de alcalinidade. Deve-se tornar um cuidado especial ao se usar NaOH para aumentar o pH. Na ausência de um âcido fraco, o quai é necessârio para se criar uma capacidade de tamponamento, a adiçâo de NaOH pode romper o delicado equilfbrio do pH e, acidentalmente, provocar a sua elevaçâo para valores demasiadamente alcalinos. É
130
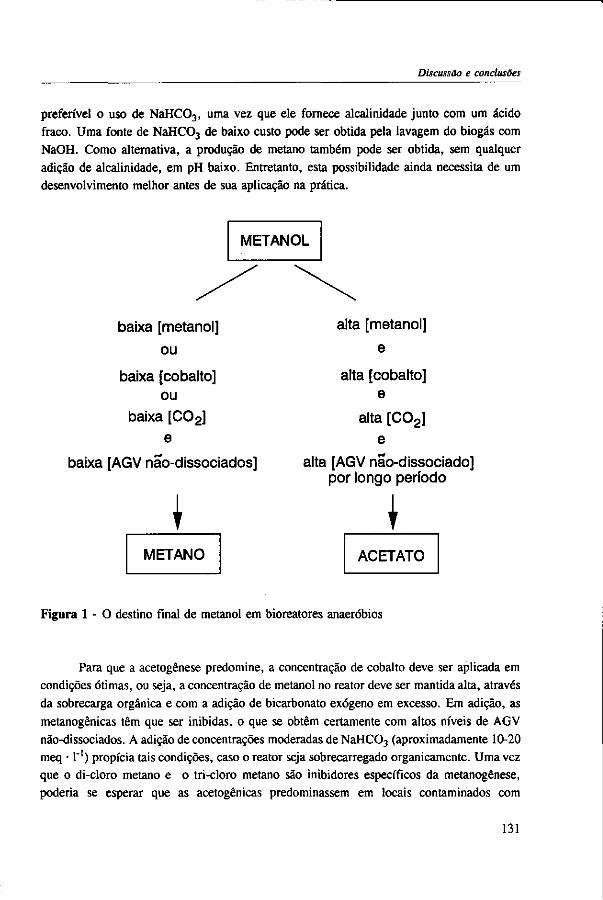
Discussäo e conclusôes
preferîvel o uso de NaHC03, uma vez que ele fornece alcalinidade junto com um âcido fraco. Uma fonte de NaHC03 de baixo custo pode ser obtida pela lavagem do biogäs com NaOH. Como alternativa, a produçâo de metano também pode ser obtida, sem qualquer adiçâo de alcalinidade, em pH baixo. Entretanto, esta possibilidade ainda nécessita de um desenvolvimento melhor antes de sua aplicaçâo na prâtica.
METANOL
baixa [metanol] ou
baixa [cobalto] ou
baixa [C02] e
baixa [AGV nao-dissociados]
I
alta [metanol]
e
alta [cobalto] e
alta [C02] e
alta [AGV nao-dissociado] por longo perîodo
t METANO ACETATO
Figura 1 - 0 destino final de metanol em bioreatores anaeróbios
Para que a acetogênese prédomine, a concentraçào de cobalto deve ser aplicada em condiçôes ótimas, ou seja, a concentraçào de metanol no reator deve ser mantida alta, através da sobrecarga orgânica e com a adiçâo de bicarbonato exógeno em excesso. Em adiçâo, as metanogênicas têm que ser inibidas, o que se obtêm certamente corn altos nfveis de AGV nao-dissociados. A adiçâo de concentraçôes moderadas de NaHC03 (aproximadamente 10-20 meq • l"1) propfcia tais condiçôes, caso o reator seja sobrecarregado organicamente. Uma vez que o di-cloro metano e o tri-cloro metano sâo inibidores especfficos da metanogênese, poderia se esperar que as acetogênicas predominassem em locais contaminados com
131

Outras aplicaçôes
halometanos, em caso de bioremediaçâo anaeróbia. A pasteurizaçâo de lodo também merece consideraçào, uma vez que muitas acetogênicas metilotróficas sâo formadoras de esporos, como sâo os casos de Sporomusa sp.,9 Clostridium CV-AAI1 and Butyribacterium methylotrophicum.13
6.6 Outras Aplicaçôes
Os resultados obtidos nesta tese com a degradaçâo anaeróbia de substrates metilotróficos permitem outras aplicaçôes mais amplas. Alguns exemplos de tais aplicaçôes no campo da biotecnologia ambiental, além da remocäo de DQO das âguas residuânas metilotróficas, inclüem: a biodegradaçâo de compostes aromâticos metoxilados e o uso de metanol como um co-substrato barato. Além do mais, os resultados desta tese fornecem fortes indfcios dos processos que ocorrem na natureza, tais como a formaçâo de biogâs em solos âcidos de turfa. As metanogênicas metilotróficas sâo ativas numa faixa ampla de pH. Logo, a produçâo de biogâs em turfas âcidas pode ser muite bem devido à conversâo de substrates metilotróficos derivados de metil-éteres ou ésteres em fenóis naturais de plantas, hemi-celulose e pectina.
6.7 Literatura citada
1. Adamse, A.,D., e C. T. M. Velzeboer (1982) Features of a Clostridium, strain CV-AA1, an obligatory anaerobic bacterium producing acetic acid from methanol. Antonie van Leeuwenhoek 48:305-313.
2. Cord-Ruwisch, R., e B. Ollivier (1986) Interspecific hydrogen transfer during methanol degradation by Sporomusa acidovorans and hydrogenophilic anaerobes. Arch. Microbiol. 144:163-165.
3. DiStefano, T. D., J. M. Gossett, e S. H. Zinder (1992) Hydrogen as an electron donor for dechlorination of tetrechloroethene by an anaerobic mixed culture. Appl. Environ. Microbiol. 58:3622-3629.
4. Freedman, D. L. e S. Hashsham (1993) Enhanced biotransformation of carbon tetrachloride under methanogenic conditions. Resumo nas atas do Segundo Simposio Internacional - In situ and on-site bioreclamation. San Diego, California, 5-8 de abril, 1993.
5. Heijthuijsen, J. H. F. G., e T. A. Hansen. (1986) Interspecies hydrogen transfer in co-cultures of methanol-utilizing acidogens and sulfate-reducing or methanogenic bacteria. FEMS Microbiol. Lett. 38:57-64.
6. van der Meijden, P., C. van der Drift, e G. D. Vogels (1984) Methanol conversion
132

Discussào e conclusöes
in Eubacterium limosum. Arch. Microbiol. 138:360-364. 7. van der Meijden, P., H. J. Heythuysen, H. Sliepenbeek, F. P. Houwen, C. van der
Drift, e G. D. Vogels (1984) Activation and inactivation of methanol: 2-mercaptoethanesulfonic acid methyltransferase from Methanosarcina barken. J. Bacteriol. 153:6-11.
8. Minami, K., K. Okamura, S. Ogawa, e T. Naritomi (1991) Continuous anaerobic treatment of wastewater from a kraft pulp mill. J. Ferment. Bioeng. 71:270-274.
9. Moller, B., R. Oßmer, B. H. Howard, G. Gottschalk, e H. Hippe (1984) Sporomusa, a new genus of gram-negative anaerobic bacteria including Sporomusa sphaeroides spec. nov. and Sporomusa ovata spec. nov. Arch. Microbiol. 139:388-396.
10. Stupperich, E., H. Eisinger, e S. Schurr (1990) Corrinoids in anaerobic bacteria. FEMS Microbiol. Rev. 87:355-360.
11. Stupperich, E., P. Aulkemeyer, e C. Eckerskorn (1992) Purification and characterization of a methanol-induced cobamide-containing protein from Sporomusa ovata. Arch. Microbiol. 158:370-373.
12. Stupperich, E., e R. Konle (1993) Corrinoid-dependent methyl transfer reactions are involved in methanol and 3,4-dimethyoxybenzoate metabolism by Sporomusa ovata. Appl. Environ. Microbiol. 59:3110-3116.
13. Zeikus, J. G., L. H. Lynd, T. E. Thompson, J. A. Krzycki, P. J. Weimer, e P. W. Hegge (1980) Isolation and characterization of a new, methylotrophic, acidogenic anaerobe, the Marburg strain. Curr. Microbiol. 3:381-386.
133

Acknowledgements
This thesis would not have been accomplished without the work and help of many people, whom I would like to thank here.
First I would like to express my sincere gratitude to Gatze Lettinga, my "promotor", for giving me the opportunity to work at the department, introducing me in the "very interesting subject" of anaerobic degradation of methanol, for his guidance as well as for his warmth and understanding throughout these past five years. I also would like to express my deep thanks to Jim Field, my "co-promotor", for his extensive help and delightful guidance in every step of this work; without his support this dissertation would never have been completed.
Part of the research in this dissertation was the result of the efforts of several students, colleagues, researchers and visiting researchers of the Department of Environmental Technology and Department of Microbiology. I am grateful to Fons Stams for his thorough and careful review of the manuscript, for his valuable comments and pleasant cooperation. Anita van Langerak deserves special thanks for her hard experimental work, for her stimulating interest, and enjoyable companionship. It was also a great pleasure to work together with Adrie Veeken, Alla Nozhevnikova, André Visser, Bram Damman, Pavel Jenicek, Richard Tichy, Ronaldo Petruy, and Sonja Viester. Cooperation with them enabled me to perform studies beyond the scope of my skills and possibilities.
A dedicated analytical team at the Department of Environmental Technology has supported me. I am deeply indebted to them all: Use Bennehey, Johannes van der Laan, and Martin de Wit for making all chromatographic determinations possible; Paul de Jong, Ron Sleyster for the chemical analyses; and Jo Ackerman-Jacobs, Sjoerd Hobma and Vinnie de Wilde for their willingness to help when required. Appreciation is likewise extended to Heleen Vos and Liesbeth Kesaulya-Monster for their timely help at our secretary's office. Acknowledgement should be made to the staff of the technical services at Biotechnion, stockroom, photograph services, drawing room, maintenance and the shop.
135

I thank Renze van Houten for helping with the Dutch translation of the discussion and conclusions, and also for helpful discussion and valuable bibliographic references, that is also extended to Adrianus van Haandel, José Luis Sanz Martin, Jukka Rintala, Jules van Lier Martin Brockmann, and Miriam van Eekert. Many thanks are also to Albert Janssen, André van der Last, Arne Alphenaar, Brian Donlon, Ching-Shyung Hwu, Elias Razo Flores, Erna Noordkamp, Gao Yan, Greg Malina, Grietje Zeeman, Joost van Buuren, Jos Frijns, Julia Rousskova, Look Hulshoff Pol, Marjo Lexmond, Paula Catunda, Salih Rebac, Sjon Kortekaas, Viet Nguyen Trung, and all other members of the department who created a very pleasant working atmosphere.
My special thanks are to Reyes Sierra, who helped me in so many different ways, for her continuing support and friendship. Many other people also contributed to my enjoyable stay in The Netherlands, namely Dora Lettinga, Helen and Tony Haigh, Jane Hulshoff Pol, Katja Grolle, Lucilla Bassetti, Marinka van Ommen, Marta Perez Soba, Ricardo Bernardes, Ritva Kumpulainen, Teresa Cesario, Wim van der Maas, and Yuzo Fujikura. I also express my appreciation to my family and friends in Brazil, specially Joséte Florencio, Silvia Passos, and Silvia Romitelli for their constant support.
I thank the Department of Civil Engineering from the Federal University of Pernambuco at Recife for giving the opportunity to come to The Netherlands. The financial support of the Brazilian Ministry of Education (CAPES - Coordenaçâo de Apoio ao Ensino Superior) is also greatfully acknowledged.
I would like to thank my former professors in Brazil, Samuel Murgel Branco, Eugenio Foresti, Ivanildo Hespanhol, and José Roberto Campos for their early guidance, encouragement and career inspiration.
Last but not least, my most special thanks to Mario Kato for his thoughtfulness, constant encouragement and unerring support that were of fundamental importance for the conclusion of this work.
136

Curriculum vitae
The author of this dissertation, born in Caruaru - Pernambuco, Brazil - obtained her degree in Civil Engineering at Federal University of Pernambuco (UFPE). From University of Sâo Paulo at Sao Carlos, she was granted her master of science degree with magna cum lauda from the Department of Hydraulics and Sanitary Engineering. The topic of her master thesis was on bulking sludge occuring in aerobic wastewater treatment. Between 1982 and 1988, she has been a faculty member of the Department of Civil Engineering, UFPE. Since 1989 until present, the author has been a colleague of the Department of Environmetal Technology at the Agricultural University in Wageningen (The Netherlands). In June, 1994, the author will return to the Department of Civil Engineering, UFPE, in Recife, to conduct research on the topic of anaerobic wastewater treatment.
137





























