Embed Size (px)
Citation preview

of July 28, 2018.This information is current as
and Deramification of Activated MicrogliaA Dual Role of Lipocalin 2 in the Apoptosis
Kim and Kyoungho SukWon-Ha Lee, Kiyoshi Mori, Sang-Hyun Kim, In Kyeom Shinrye Lee, Jayoung Lee, Sangseop Kim, Jae-Yong Park,
http://www.jimmunol.org/content/179/5/3231doi: 10.4049/jimmunol.179.5.3231
2007; 179:3231-3241; ;J Immunol
Referenceshttp://www.jimmunol.org/content/179/5/3231.full#ref-list-1
, 16 of which you can access for free at: cites 64 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists All rights reserved.Copyright © 2007 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on July 28, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on July 28, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

A Dual Role of Lipocalin 2 in the Apoptosis and Deramificationof Activated Microglia1
Shinrye Lee,* Jayoung Lee,* Sangseop Kim,* Jae-Yong Park,† Won-Ha Lee,‡ Kiyoshi Mori,§
Sang-Hyun Kim,* In Kyeom Kim,* and Kyoungho Suk2*
Activated microglia are thought to undergo apoptosis as a self-regulatory mechanism. To better understand molecularmechanisms of the microglial apoptosis, apoptosis-resistant variants of microglial cells were selected and characterized. Theexpression of lipocalin 2 (lcn2) was significantly down-regulated in the microglial cells that were resistant to NO-inducedapoptosis. lcn2 expression was increased by inflammatory stimuli in microglia. The stable expression of lcn2 as well as theaddition of rLCN2 protein augmented the sensitivity of microglia to the NO-induced apoptosis, while knockdown of lcn2expression using short hairpin RNA attenuated the cell death. Microglial cells with increased lcn2 expression were moresensitive to other cytotoxic agents as well. Thus, inflammatory activation of microglia may lead to up-regulation of lcn2expression, which sensitizes microglia to the self-regulatory apoptosis. Additionally, the stable expression of lcn2 in BV-2microglia cells induced a morphological change of the cells into the round shape with a loss of processes. Treatment ofprimary microglia cultures with the rLCN2 protein also induced the deramification of microglia. The deramification ofmicroglia was closely related with the apoptosis-prone phenotype, because other deramification-inducing agents such ascAMP-elevating agent forskolin, ATP, and calcium ionophore also rendered microglia more sensitive to cell death. Takentogether, our results suggest that activated microglia may secrete LCN2 protein, which act in an autocrine manner tosensitize microglia to the self-regulatory apoptosis and to endow microglia with an amoeboid form, a canonical morphologyof activated microglia in vivo. The Journal of Immunology, 2007, 179: 3231–3241.
M icroglia are the CNS-resident immune cells (1–3).These phagocytic cells function as the first line of de-fense in the CNS. When stimulated, ramified resting
microglia transform into the amoeboid form (4) and they activelyparticipate in the CNS immune and inflammatory responses (5, 6).Although the morphological transformation from the ramifiedmicroglia into the amoeboid microglia is observed under a widevariety of in vitro as well as in vivo conditions (7–9), the pre-cise mechanism of the changes in the microglial morphologyremains largely unknown. Activated microglia migrate to areaof injured nervous tissue and they engulf and destroy microbesand cellular debris (10). The role of microglia as inflammatorycells is not always beneficial. Uncontrolled microglial activa-tion and the subsequent excessive neuroinflammation are nowbelieved to contribute to a variety of the CNS pathologies in-cluding neurodegenerative diseases (11–14). Thus, inflamma-
tory activation of microglia has to be strictly regulated, and theapoptotic elimination of activated microglia is thought to be onemechanism of the microglial self-regulation (15–19). Evidencehas been provided that inflammatory cells in the CNS may un-dergo apoptosis upon activation in a manner similar to activa-tion-induced cell death of lymphocytes (16, 20, 21). However,little is known about the molecular mechanisms underlying thetermination of neuroinflammation and the autoregulatory apo-ptosis of microglia.
Lipocalin 2 (LCN2)3 is a member of lipocalin family, whichbinds or transports lipid and other hydrophobic molecules (22,23). LCN2 is also known as 24p3 (24), 24-kDa superinducibleprotein (SIP24) (25), and neutrophil gelatinase-associated li-pocalin (NGAL; a human homolog of lcn2) (26, 27). LCN2 hasdiverse functions. In vitro studies have shown that LCN2 isimportant for both cellular apoptosis and survival (28 –32). Italso plays a central role in the induction of cellular differenti-ation in the kidney during embryogenesis (33) and protects thekidney from ischemic injury (34, 35). In various forms of gas-trointestinal injury, LCN2 facilitates mucosal regeneration bypromoting cell migration (36). In vivo studies based on lcn2-deficient mice, however, argued against the role of LCN2 in therenal protection (37). Moreover, lcn2-deficient mice exhibitedan increased susceptibility to bacterial infections because of thefailure of iron sequestration, indicating a critical role of lcn2 inprotection against bacterial infection (38). Previous works havealso suggested that lcn2 protects neutrophil gelatinase fromdegradation (so, the name NGAL) (39), and it may function as
*Department of Pharmacology, Kyungpook National University School of Medicine,Daegu, Korea; †Department of Physiology, Institute of Health Science, GyeongsangNational University, Jinju, Korea; ‡Department of Genetic Engineering, School ofLife Sciences and Biotechnology, Kyungpook National University, Daegu, Korea;and §Department of Medicine and Clinical Science, Kyoto University GraduateSchool of Medicine, Kyoto, Japan
Received for publication March 6, 2007. Accepted for publication June 22, 2007.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was supported by the Neurobiology Research Program from the KoreaMinistry of Science and Technology and by Grant R01-2006-000-10314-0 from theBasic Research Program of the Korea Science and Engineering Foundation. S.L., J.L.,and S.K. were supported by the Brain Korea 21 Project in 2006. J.-Y.P. and K.S. werethe recipients of the Korea Research Foundation Grant funded by the Korean Gov-ernment (MOEHRD, Basic Research Promotion Fund) (KRF-2006-005-J04202;KRF-2006-311-E00045).2 Address correspondence and reprint requests to Dr. Kyoungho Suk, Department ofPharmacology, School of Medicine, Kyungpook National University, 101 Dong-In,Joong-gu, Daegu, 700-422, Korea. E-mail address: [email protected]
3 Abbreviations used in this paper: LCN2, lipocalin 2; SNAP, S-nitroso-N-acetylpeni-cillamine; SNP, sodium nitroprusside; DFO, deferoxamine mesylate; shRNA, shorthairpin RNA.
Copyright © 2007 by The American Association of Immunologists, Inc. 0022-1767/07/$2.00
The Journal of Immunology
www.jimmunol.org
by guest on July 28, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

an acute phase protein (40). Recently, two cellular receptors forlcn2 have been identified. Megalin, a member of the low-den-sity lipoprotein receptor family, has been shown to bind humanlcn2 and to mediate its cellular uptake (41). Brain type organiccation transporter is another cell surface receptor for mouselcn2, which has been shown to selectively mediate apoptosis(29). Despite the receptor identification, the precise role of lcn2in cell survival and death has yet to be determined.
In the present study, we used a transcriptomic approach com-bined with the phenotypic selection of apoptosis-resistant mi-croglial cells to better understand the mechanisms of microglialapoptosis. One of the genes that was significantly down-regu-lated in the apoptosis-resistant microglial cells was lcn2. Sub-sequent studies including the forced expression and knockdownof gene expression revealed that lcn2 is critical for apoptosissensitization as well as for the amoeboid transformation of ac-tivated microglia.
Materials and MethodsReagents and cells
LPS from Escherichia coli 0111:B4 prepared by phenolic extraction andgel filtration chromatography, S-nitroso-N-acetylpenicillamine (SNAP),sodium nitroprusside (SNP), etoposide, cisplatin, hydrogen peroxide,PMA, deferoxamine mesylate (DFO), thrombin, ATP, A23187, forskolin,and dexamethasone were obtained from Sigma-Aldrich. Staurosporine andganglioside mixtures were purchased from Calbiochem. Recombinantmouse IFN-� and anti-mouse LCN2 polyclonal Ab were purchasedfrom R&D Systems. Iron-saturated enterochelin (0.7 kDa) was pur-chased from EMC Microcollections. All other chemicals were obtainedfrom Sigma-Aldrich, unless stated otherwise. BV-2 mouse microglialcell line (42) which exhibits phenotypic and functional properties com-parable to those of primary microglial cells (43) were grown in DMEMcontaining 5% FBS, 2 mM glutamine, and penicillin-streptomycin (In-vitrogen Life Technologies). Mouse primary microglial cultures wereprepared by mild trypsinization as previously described with minormodifications (44). In brief, forebrains of newborn ICR mice werechopped and dissociated by mechanical disruption using a nylon mesh.The cells were seeded into poly-L-lysine-coated flasks. After in vitroculture for 10 –14 days, microglial cells were isolated from mixed glialcultures by mild trypsinization. Mixed glial cultures were incubatedwith a trypsin solution (0.25% trypsin, 1 mM EDTA in HBSS) diluted1/4 in PBS containing 1 mM CaCl2 for 30 – 60 min. This resulted in thedetachment of an upper layer of astrocytes in one piece, whereas mi-croglia remained attached to the bottom of the culture flask. The de-tached layer of astrocytes was aspirated and the remaining microgliawere used for experiments. The purity of microglial cultures was �95%as determined by isolectin B4 staining. Animals used in the currentresearch have been acquired and cared for in accordance with the guide-lines published in the National Institutes of Health Guide for the Careand Use of Laboratory Animals.
Microarray analysis
For the gene expression profiling studies, DNA microarray analysis wasconducted by GenoCheck using a Platinum Biochip mouse 7.4 k cDNAchip containing 7636 cDNA spots. Data analysis was done usingGenePix pro 4.1 software. Log-transformed spot intensity was plotted(M vs A scatter plot), normalized, and further analyzed as previouslydescribed (45). A complete listing of the genes on the microarray anddetailed experimental protocols is available at the GenoCheck web site(www.genocheck.com).
Assessment of cytotoxicity by MTT assay or TUNEL staining
For MTT assay, cells (5 � 104 cells in 200 �l/well) were seeded in 96-wellplates and treated with various stimuli for the indicated time periods. Aftertreatment, the medium was removed and MTT (0.5 mg/ml) was added,followed by incubation at 37°C for 2 h in CO2 incubator. After insolublecrystals were completely dissolved in DMSO, absorbance at 570 nm wasmeasured using a microplate reader (Anthos Labtec Instruments). Apopto-sis of microglia was scored by TUNEL assay using a commercially avail-able kit according to the manufacturer’s protocol (In Situ Cell Death De-tection kit, POD; Roche Applied Science). The percentage of apoptotic
cells was quantitated by counting TUNEL-positive cells in 10 random mi-croscope fields.
Evaluation of mitochondrial membrane potential
Mitochondrial potential was measured using a lipophilic cationic probe5,5�,6,6�-tetrachloro-1,1�,3,3�-tetraethylbenzimidazolcarbocyanine io-dide (JC-1; Molecular Probes). After incubating the cells with 10 �g/mlJC-1 at 37°C for 10 min, emission at 525 and 590 nm was measuredusing a fluorescent microplate reader. The ratio of A590 (red) to A525
(green) was calculated as an indicator of mitochondrial potential. Adecrease in membrane potential leads to a decrease in the ratio of red:green fluorescence.
Morphological analyses of microglial deramification andapoptosis
Morphological analysis of microglia was performed using phase contrastand fluorescence microscope. Isolectin B4 staining was done as previouslydescribed (46). Microscopic images were processed using MetaMorph Im-aging System (Molecular Devices). Deramification of microglia was quan-titated as previously described with a slight modification (47). Ramifiedcells were defined as having at least two processes, among which oneprocess was longer than one cell body diameter. Nonramified cells werethose that did not fulfill these criteria. The percentage of ramified cells wasdetermined from a minimum of five randomly chosen fields containing atleast 250 cells. Morphological changes in the nuclear chromatin of cellsundergoing apoptosis were detected by staining with 2.5 �g/ml Hoechst33342 fluorochrome (Molecular Probes), followed by examination on afluorescence microscope (Olympus BX50).
Flow cytometric analysis of apoptosis
Microglia were detached with trypsin-EDTA and washed twice with coldPBS. The cells were then resuspended in 250 �l of binding buffer (10 mMHEPES, 140 mM NaCl, and 2.5 mM CaCl2 (pH 7.4)) and incubated with3 �l of FITC-conjugated annexin V (Molecular Probes) according to themanufacturer’s specifications. Afterward, cells were gently vortexed andincubated for 15 min at room temperature in the dark. Propidium iodide (20�g/ml) was then added and flow cytometry was performed within 1 h usingFACSAria (BD Biosciences).
RT-PCR
Total RNA was extracted from BV-2 cells or primary microglial cells byTRIzol reagent (Invitrogen Life Technologies) according to the manufac-turer’s protocol. Reverse transcription was conducted using Superscript(Invitrogen Life Technologies) and oligo(dT) primer. PCR amplificationusing specific primer sets was conducted at 55°C annealing temperature for30–40 cycles. Nucleotide sequences of the primers were based on pub-lished cDNA sequences (Table I).
Western blot analysis
Cells were lysed in triple-detergent lysis buffer (50 mM Tris-HCl (pH 8.0),150 mM NaCl, 0.02% sodium azide, 0.1% SDS, 1% Nonidet P-40, 0.5%sodium deoxycholate, and 1 mM PMSF). Protein concentration in celllysates was determined using the Bio-Rad protein assay kit. An equalamount of protein for each sample was separated by 12% SDS-PAGE andtransferred to Hybond ECL nitrocellulose membranes (Amersham Bio-sciences). The membranes were blocked with 5% skim milk and sequen-tially incubated with primary Abs (goat polyclonal anti-LCN2 Ab, R&DSystems; rabbit polyclonal anti-BAD Ab, Cell Signaling Technology; rab-bit polyclonal anti-BAX Ab, Cell Signaling Technology; rat monoclonalanti-BIM Ab, Calbiochem; rabbit polyclonal anti-STAT1 Ab, Cell Signal-ing Technology; rabbit polyclonal anti-IRF-1 Ab, Santa Cruz Biotechnol-ogy; monoclonal anti-�-tubulin clone B-5-1-2 mouse ascites fluid; Sigma-Aldrich) and HRP-conjugated secondary Abs (anti-rabbit or -mouse IgG;Amersham Biosciences), followed by ECL detection (Amersham Bio-sciences). For the detection of secreted LCN2 protein, Western blot anal-ysis of culture medium was performed. In brief, microglial cells grown in100-mm culture dishes were washed five times with PBS. Cells were thencovered with a minimal volume of culture medium, and then treated withstimulating agents at 37°C. Conditioned medium were collected and cen-trifuged successively at 2,000 � g (5 min), 15,600 � g (10 min), to removenonadherent cells and debris. Samples were then precipitated with a tri-chloroacetic acid and acetone mixture (10% trichloroacetic acid and 10mM DTT in acetone) at �20°C overnight. Precipitated proteins were sub-jected to the SDS-PAGE and ECL detection.
3232 Lcn2 IN MICROGLIAL APOPTOSIS AND DERAMIFICATION
by guest on July 28, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
Stable transfection of lcn2 cDNA or short hairpin RNA (shRNA)
BV-2 cells in 6-well plates were transfected with 2 �g of mouse lcn2 orlcn2-specific shRNA constructs using Lipofectamine reagent (InvitrogenLife Technologies). Mammalian expression construct of mouse lcn2 clonedinto pcDNA3 was a gift from Dr. J. P. Kehrer (University of Texas, Austin,TX) (32), and lcn2-specific shRNA construct was a gift from Dr. L. G.Cantley (Yale University, New Haven, CT) (48). The shRNA correspond-ing to the nucleotides 192–213 of lcn2 cDNA followed by a 9-base loopand the inverted repeat was cloned into pSuppressor Retro vector (Im-genex). An empty pcDNA3 vector and a vector encoding the scrambledRNA were used as a control for the stable expression of lcn2 and lcn2-specific shRNA, respectively. Stable transfectants were selected in thepresence of G418 (400 �g/ml) at 2 days after the transfection. Up- ordown-regulation of LCN2 protein in the stable transfectants was confirmedby Western blot analysis.
rLCN2 protein
Recombinant mouse LCN2 protein has been prepared as previously de-scribed (33). In brief, rLCN2 protein was expressed as a GST fusion pro-tein in the BL21 strain of E. coli, which does not synthesize siderophore.The protein was purified using glutathione-Sepharose 4B beads (Amer-sham Biosciences), followed by elution with thrombin or glutathione. Foriron and enterochelin loading, a 5-fold molar excess of ion-saturated en-terochelin (EMC Microcollections) was mixed with purified rLCN2protein.
Statistical analysis
All data were presented as means � SD from three or more independentexperiments. Statistical comparison between different treatments was done
by either the Student t test or one-way ANOVA with Dunnett’s multiplecomparison tests using the GraphPad Prism program (GraphPad Software).Differences with a p value �0.05 were considered statistically significant.
ResultsEstablishment and characterization of apoptosis-resistantvariant of microglial cells
We have previously shown that inflammatory activation of cul-tured microglia induces a self-regulatory apoptosis. To investigate
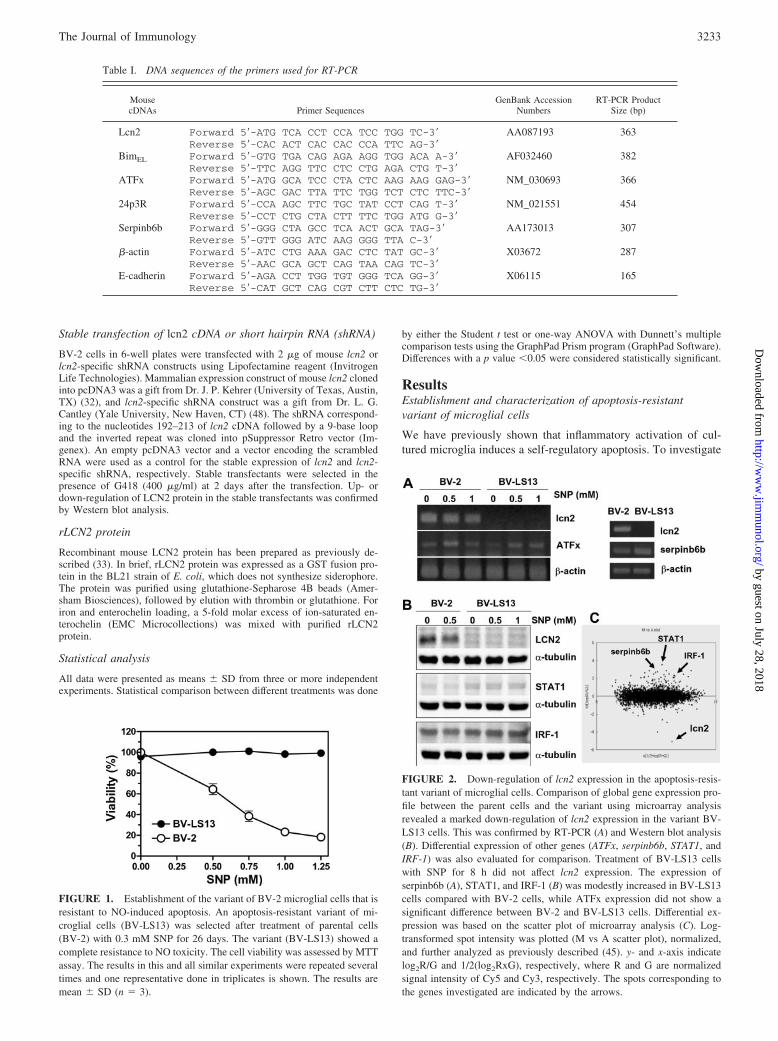
FIGURE 1. Establishment of the variant of BV-2 microglial cells that isresistant to NO-induced apoptosis. An apoptosis-resistant variant of mi-croglial cells (BV-LS13) was selected after treatment of parental cells(BV-2) with 0.3 mM SNP for 26 days. The variant (BV-LS13) showed acomplete resistance to NO toxicity. The cell viability was assessed by MTTassay. The results in this and all similar experiments were repeated severaltimes and one representative done in triplicates is shown. The results aremean � SD (n � 3).
FIGURE 2. Down-regulation of lcn2 expression in the apoptosis-resis-tant variant of microglial cells. Comparison of global gene expression pro-file between the parent cells and the variant using microarray analysisrevealed a marked down-regulation of lcn2 expression in the variant BV-LS13 cells. This was confirmed by RT-PCR (A) and Western blot analysis(B). Differential expression of other genes (ATFx, serpinb6b, STAT1, andIRF-1) was also evaluated for comparison. Treatment of BV-LS13 cellswith SNP for 8 h did not affect lcn2 expression. The expression ofserpinb6b (A), STAT1, and IRF-1 (B) was modestly increased in BV-LS13cells compared with BV-2 cells, while ATFx expression did not show asignificant difference between BV-2 and BV-LS13 cells. Differential ex-pression was based on the scatter plot of microarray analysis (C). Log-transformed spot intensity was plotted (M vs A scatter plot), normalized,and further analyzed as previously described (45). y- and x-axis indicatelog2R/G and 1/2(log2RxG), respectively, where R and G are normalizedsignal intensity of Cy5 and Cy3, respectively. The spots corresponding tothe genes investigated are indicated by the arrows.
Table I. DNA sequences of the primers used for RT-PCR
MousecDNAs Primer Sequences
GenBank AccessionNumbers
RT-PCR ProductSize (bp)
Lcn2 Forward 5�-ATG TCA CCT CCA TCC TGG TC-3� AA087193 363Reverse 5�-CAC ACT CAC CAC CCA TTC AG-3�
BimEL Forward 5�-GTG TGA CAG AGA AGG TGG ACA A-3� AF032460 382Reverse 5�-TTC AGG TTC CTC CTG AGA CTG T-3�
ATFx Forward 5�-ATG GCA TCC CTA CTC AAG AAG GAG-3� NM_030693 366Reverse 5�-AGC GAC TTA TTC TGG TCT CTC TTC-3�
24p3R Forward 5�-CCA AGC TTC TGC TAT CCT CAG T-3� NM_021551 454Reverse 5�-CCT CTG CTA CTT TTC TGG ATG G-3�
Serpinb6b Forward 5�-GGG CTA GCC TCA ACT GCA TAG-3� AA173013 307Reverse 5�-GTT GGG ATC AAG GGG TTA C-3�
�-actin Forward 5�-ATC CTG AAA GAC CTC TAT GC-3� X03672 287Reverse 5�-AAC GCA GCT CAG TAA CAG TC-3�
E-cadherin Forward 5�-AGA CCT TGG TGT GGG TCA GG-3� X06115 165Reverse 5�-CAT GCT CAG CGT CTT CTC TG-3�
3233The Journal of Immunology
by guest on July 28, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

the molecular mechanism(s) of the self-regulatory apoptosis ofactivated microglia, we selected and characterized apoptosis-resis-tant variants of BV-2 mouse microglial cells. Because NO is the
major cytotoxic mediator in the apoptosis of activated microglia,variant clones of BV-2 microglial cells that were resistant to theNO toxicity were selected. After long-term culture of microglial
FIGURE 3. Apoptosis-enhancing effect of lcn2 in mi-croglia. BV-2 microglial cells with increased lcn2 expres-sion were selected by stable transfection of lcn2 cDNA.The increased lcn2 expression in the stable transfectantsof lcn2 (S1 and S3) compared with the cells transfectedwith an empty vector (E1) was confirmed by RT-PCR andWestern blot analysis (A). BV-2 microglial cells with in-creased lcn2 expression showed an enhanced sensitivity tothe NO donor (SNP or SNAP)-induced apoptosis as de-termined by MTT assay (B) and annexin V-binding assay(data not shown). The lcn2 transfectants (S1 and S3) orthe empty vector transfectant (E1) were exposed to theindicated concentrations of SNP or SNAP for 24 h, andthen cell viability or apoptosis was evaluated by MTTassay (B) or flow cytometric analysis (data not shown).Values represent mean � SD. �, Statistically significantdifferences from the empty vector transfectant (E1) (B)(p � 0.05). Results are the representative of three inde-pendent experiments.
FIGURE 4. Decreased apoptotic sensitivity in thelcn2-knockdown cells. Stable knockdown of lcn2 ex-pression by transfection of BV-2 cells with lcn2-spe-cific shRNA construct decreased the apoptotic sensi-tivity compared with the scrambled shRNAtransfectant. Knockdown of lcn2 expression was con-firmed by Western blot analysis of the transfectants(A). The viability of transfectants was measured byMTT assay after treatment with the indicated concen-trations of SNP or SNAP for 24 h (B). The lcn2 knock-down effect was rescued by rLCN2 protein treatment(C). The rLCN2 protein reversed the decrease in theapoptotic sensitivity. Values are mean � SD (n �3). �, Statistically significant differences from thescrambled shRNA transfectant (p � 0.05).
3234 Lcn2 IN MICROGLIAL APOPTOSIS AND DERAMIFICATION
by guest on July 28, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

cells in the presence of NO donor SNP, a variant clone of micro-glial cells that were resistant to the NO cytotoxicity was estab-lished and named BV-LS13. The NO-induced cell death in theparental BV-2 cells was due to apoptosis as determined by nuclearmorphology, TUNEL staining, and induction of mitochondrial per-meability transition (data not shown). The BV-LS13 was com-pletely resistant to the NO-induced apoptosis, as opposed to pa-rental cells (Fig. 1). We next compared the global gene expressionprofile of the parental cells and the variant cells by cDNA mi-croarray analysis (Fig. 2C). One of the genes that showed a sig-nificant change in the mRNA expression levels in the BV-LS13cells was lcn2: �5-fold decrease in the lcn2 mRNA expression hasbeen detected in the BV-LS13 cells by the microarray analysis.The decrease in the lcn2 expression in the BV-LS13 cells wasconfirmed by RT-PCR and Western blot analysis (Fig. 2, A and B).The expression of ATFx has been also investigated, because it hasbeen previously reported that lcn2 and ATFx are oppositely reg-ulated in the process of hemopoietic cellular apoptosis (30, 49). Asignificant change in ATFx expression between the parental andvariant cells was not detected. The expression of serpinb6b,STAT1, and IRF-1 was also assessed for comparison, because thedifferences in the expression of these genes were shown in themicroarray analysis. In the current study, we have focused on lcn2.
Proapoptotic activity of lcn2 in microglia
As the expression of lcn2 was down-regulated in the apoptosis-resistant microglia and lcn2 has been previously involved in thecell death and survival, we hypothesized that the apoptosis-resis-tant phenotype of the BV-LS13 cells may be due to the markeddown-regulation of proapoptotic lcn2 expression. To test this hy-
pothesis, we used three different approaches: 1) stable overexpres-sion of lcn2; 2) knockdown of lcn2 expression using shRNA; and3) treatment with rLCN2 protein. First, BV-2 cells overexpressinglcn2 were obtained by stable transfection. An increase in the lcn2mRNA as well as protein expression in the stable transfectants (S1and S3) was confirmed by RT-PCR and Western blot analysis (Fig.3A). Stable overexpression of lcn2 enhanced the sensitivity of mi-croglial cells to NO donors such as SNP and SNAP (Fig. 3B). ANO donor SNP releases iron along with NO (50). A small amountof iron may affect microglial viability. To exclude this possibility,the effect of SNP was confirmed by another NO donor SNAP,which does not release iron. Both SNP and SNAP gave rise to asimilar result. The apoptosis-enhancing effects of lcn2 in the stabletransfectants were assessed by MTT assay (Fig. 3B) as well asannexin V staining followed by flow cytometric analysis (data notshown). Transfection of lcn2 also increased the sensitivity of mi-croglia to other cytotoxic agents such as etoposide, H2O2, andstaurosporine (data not shown), indicating that lcn2 indeed hasproapoptotic activity. The lcn2 transfectants S1 and S3, however,showed a slight difference in their sensitivity to cytotoxic drugs,which cannot be explained currently. Second, stable knockdown oflcn2 expression by the transfection of BV-2 cells with the lcn2-specific shRNA construct decreased the NO sensitivity (Fig. 4B).Knockdown of lcn2 expression in the stable transfectants was con-firmed by Western blot analysis (Fig. 4A). To make sure that theeffect of lcn2 knockdown observed is not due to an off-target effect,a shRNA rescue experiment was performed (Fig. 4C). rLCN2 pro-tein (Fig. 5C) rescued the lcn2 knockdown effects (Fig. 4C). Ad-ditionally, the recombinant mouse LCN2 protein sensitized BV-2microglial cells to NO donors, whereas LCN2 protein alone did not
FIGURE 5. rLCN2 protein sensi-tized microglia to apoptosis. Additionof rLCN2 protein increased the sen-sitivity of microglia to the NO toxic-ity. The apoptosis-enhancing effect ofLCN2 protein was observed in bothBV-2 cells (A and D) and primarymicroglia cultures (B). GST-fusedLCN2 protein was expressed in BL21cells (C, left), which was then cleavedby thrombin to release the pure LCN2protein (C, right). The GST proteinwas run on the same gel for compar-ison. In contrast to apo-LCN2 protein,LCN2 protein complexed with ironand siderophore did not exert the ap-optosis-enhancing effect (D). Valuesare mean � SD (n � 3). Asterisksindicate statistically significant differ-ences from the control treated withthe same NO donor concentrations inthe absence of LCN2 (�, p � 0.05;��, p � 0.01) (A, B, and D).
3235The Journal of Immunology
by guest on July 28, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

affect microglial cell viability (Fig. 5A). A similar result was ob-tained in primary microglia cultures (Fig. 5B). Because it has beenpreviously shown that the proapoptotic effect of LCN2 is associ-ated with iron metabolism (29), the effect of iron chelator DFO onthe microglial cell viability has been investigated. It has been re-ported that LCN2 containing the iron complex of the bacterialsiderophore donates iron to cells via the lcn2 receptor (lcn2/24p3R) (29). Internalization of LCN2 and its receptor leads to theuptake of iron from the siderophore-iron complex. Donation ofiron to the cell leads to a decrease in TfR1 expression and anincrease in ferritin levels. In addition, donation of iron to thecell prevents apoptosis by decreasing the expression of the pro-apoptotic protein Bim. In contrast, LCN2 without an iron com-plex binds to lcn2/24p3R and is internalized into the cell, andthen a putative intracellular mammalian siderophore iron com-plex becomes bound to LCN2, which is subsequently releasedfrom the cell by exocytosis. Depletion of iron from the cellresults in the up-regulation of the proapoptotic molecule, Bim,which leads to apoptosis. DFO alone was modestly toxic tomicroglia cells (20 �M, 98.2 � 1.2% viability; 100 �M, 86.3 �1.3% viability), and did not significantly affect the NO donor(SNP or SNAP)-induced microglial cell death (data not shown).Moreover, the LCN2-induced apoptosis sensitization was abol-ished by the concurrent addition of siderophore-iron complex(Fig. 5D). Concentrations of LCN2, DFO, and siderophore-ironcomplex used in the present study are comparable to those inthe previous report (29). These results indicate that the pro-apoptotic activity of LCN2 in microglia may be related with theiron transport and metabolism: apo-LCN2 may be proapoptoticby depleting intracellular iron, while holo-LCN2 containing theiron complex of siderophore may not be.
Expression and regulation of lcn2, lcn2 receptor (lcn2/24p3R),and Bcl-2 family proteins in microglia
Previously, lcn2 has been proposed as an acute phase protein (40),and the expression of lcn2 has been shown to be modulated byinflammatory stimuli in macrophages (40, 51, 52). Thus, wesought to determine how the expression of lcn2 is regulated byinflammatory or toxic stimuli in microglia. The expression of lcn2was strongly enhanced by LPS, serum withdrawal, PMA, IFN-�,and calcium ionophore A23187 (Fig. 6A). The secretion of LCN2was also increased by LPS, serum withdrawal, and PMA as de-termined by Western blot analysis of conditioned medium of mi-croglia cultures (Fig. 6B). The results indicate that microglialexpression and secretion of lcn2 may be increased underinflammatory condition in the CNS, as is in macrophages in theperiphery. Recently, the presence of lcn2 receptor (lcn2/24p3R)that mediates the lcn2-induced apoptosis has been reported (29).The expression of this lcn2 receptor was detected in microglia(Fig. 6C). Bcl-2 family proteins have been previously implicatedin the effects of lcn2 on the cell death and survival (28, 29, 31). Inparticular, up-regulation of proapoptotic Bim expression was es-sential for the apoptosis-inducing effects of lcn2 (29). However,none of the proapoptotic Bcl-2 family proteins tested was signif-icantly influenced by the lcn2 overexpression (E1 vs S1 or S3)with or without SNP treatment (Fig. 6D). Most importantly, theexpression of Bim was not changed by lcn2 transfection at mRNA(BimEL mRNA expression assessed by RT-PCR; data not shown)or protein levels (Fig. 6D). These results suggest that the regula-tion of proapoptotic Bcl-2 family proteins may not be directlyinvolved in the apoptosis-sensitizing effects of lcn2. Nevertheless,it is yet possible that the proapoptotic effect of lcn2 on microgliamay be mediated through the newly identified lcn2 receptor(lcn2/24p3R).
FIGURE 6. Expression of lcn2, lcn2 receptor (lcn2/24p3R), andBcl-2 family proteins and their regulation by inflammatory signals andother stimulants in microglia. The expression of lcn2 was increased byLPS (100 ng/ml), serum withdrawal for 8 h (SW), PMA (100 �g/ml),IFN-� (50 U/ml), or calcium ionophore A23187 (5 �M). Treatmentwith SNP (0.5 mM), gangliosides mixture (G-mixture; 50 �g/ml),thrombin (10 �g/ml), ATP (3 mM), or forskolin (10 �M), however, didnot significantly affect the lcn2 expression levels. BV-2 microglial cellswere treated with the indicated stimuli for 24 h (A, upper), 8 h (A,lower), or were incubated under serum-free condition for 8 h (SW), andthen LCN2 protein was detected by Western blot analysis (A). SecretedLCN2 protein was detected by Western blot analysis of culture mediaafter the similar treatment for 8 h (B). RT-PCR analysis revealed thatBV-2 and BV-LS13 cells as well as primary microglia cultures expresslcn2 receptor (24p3R), which has been recently shown to mediate theapoptotic effect of lcn2 (C). The lcn2 receptor (24p3R) expression wasnot detected in the reaction without reverse transcriptase (data notshown). The expression of proapoptotic Bcl-2 family proteins such asBIM, BAX, and BAD was not significantly influenced by the lcn2 up-regulation or SNP treatment (D). lcn2 receptor expression was detectedby RT-PCR analysis (C), and Bcl-2 family proteins were detected byWestern blot analysis after treatment of BV-2 cells, empty vector (E1),or lcn2 (S1 and S3) transfectants with 0.5 mM SNP for 8 h (D). EitherPonceau S staining or �-tubulin detection was done to confirm the equalloading of the samples.
3236 Lcn2 IN MICROGLIAL APOPTOSIS AND DERAMIFICATION
by guest on July 28, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

Deramification of microglia by lcn2
Stable expression of lcn2 in BV-2 microglia cells not only in-creased their sensitivity to apoptotic signals, but also induced amorphological change that reflected amoeboid transformation ofmicroglia upon activation (Fig. 7). A similar morphologicalchange was also observed in primary microglia cultures treatedwith rLCN2 protein (Figs. 8 and 9). LPS has been previouslyshown to induce microglial apoptosis through TLR4 (53) and tocause microglial deramification (4, 47, 54). Activation of microgliawith LPS treatment similarly induced deramification under the cur-rent condition (Fig. 8).
Relationship between microglial deramification andapoptosis-prone phenotype
We next examined whether the LCN2-induced microglial derami-fication is related with the apoptosis-sensitive phenotype. We haveused other signaling mediators or reagents that are previously
known to induce microglial deramification in vitro, such as ATP,forskolin, and calcium ionophore A23187. As expected, all thesestimuli induced the deramification of primary microglia cultures(Fig. 9). These stimuli also increased the NO-induced cell death ofmicroglia to varying degree (Fig. 10). However, only calciumionophore A23187 enhanced lcn2 expression (Fig. 6A), suggestingthat an increase in the intracellular calcium may lead to the mi-croglial deramification possibly through lcn2 up-regulation. Theeffect of calcium ionophore A23187 was also assessed after the
FIGURE 7. Induction of morphological change by the forced expres-sion of lcn2. Stable expression of lcn2 (S1 and S3) induced the deramifi-cation of BV-2 microglial cells. Parental BV-2 cells and the empty vector-transfectant (E1) are shown for comparison (original magnification, �200).Scale bar, 25 �m.
FIGURE 8. The effect of purified rLCN2 protein onmicroglial morphology. Addition of rLCN2 protein (10�g/ml) induced the morphological change in primarymicroglia cultures in a manner similar to the lcn2-ex-pressing BV-2 cells (B). Treatment with LPS (100 ng/ml) also induced microglial deramification (C). Primarymicroglia cultures were either left untreated (A) ortreated for 24 h with LCN2 protein or LPS, and thencellular morphology was examined under phase contrastmicroscope. The percentage of deramification was thenquantitated (D). Several randomly chosen fields wereexamined to obtain the percentage of ramified cells,which were defined as the cells harboring the processesthat are longer than the diameter of the cell body. Re-sults are one representative of more than three indepen-dent experiments. GST (10 �g/ml) was used as a controlprotein (photo not shown). Original magnification,�100 (A–C). Scale bar, 50 �m.
FIGURE 9. Comparison of LCN2 protein with other stimuli with re-spect to the effects on microglial morphology. After treatment of primarymicroglia cultures for 24 h with culture media, rLCN2 protein (10 �g/ml),ATP (3 mM), forskolin (10 �M), or A23187 (5 �M), cellular morphologywas examined by isolectin B4 staining. Afterward, the percentage of de-ramification was quantitated as in Fig. 8 (A). Representative cellular mor-phologies after the isolectin B4 staining of untreated or LCN2-treated mi-croglia are shown (original magnification, �100) (B). Scale bar, 50 �m.
3237The Journal of Immunology
by guest on July 28, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
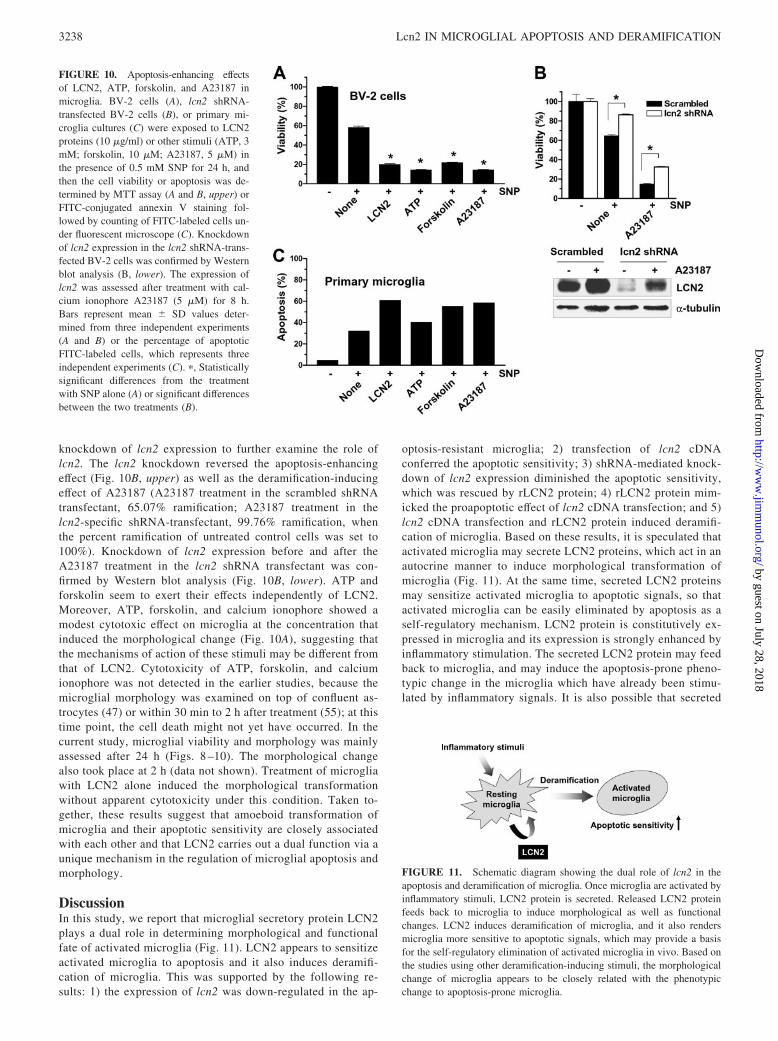
knockdown of lcn2 expression to further examine the role oflcn2. The lcn2 knockdown reversed the apoptosis-enhancingeffect (Fig. 10B, upper) as well as the deramification-inducingeffect of A23187 (A23187 treatment in the scrambled shRNAtransfectant, 65.07% ramification; A23187 treatment in thelcn2-specific shRNA-transfectant, 99.76% ramification, whenthe percent ramification of untreated control cells was set to100%). Knockdown of lcn2 expression before and after theA23187 treatment in the lcn2 shRNA transfectant was con-firmed by Western blot analysis (Fig. 10B, lower). ATP andforskolin seem to exert their effects independently of LCN2.Moreover, ATP, forskolin, and calcium ionophore showed amodest cytotoxic effect on microglia at the concentration thatinduced the morphological change (Fig. 10A), suggesting thatthe mechanisms of action of these stimuli may be different fromthat of LCN2. Cytotoxicity of ATP, forskolin, and calciumionophore was not detected in the earlier studies, because themicroglial morphology was examined on top of confluent as-trocytes (47) or within 30 min to 2 h after treatment (55); at thistime point, the cell death might not yet have occurred. In thecurrent study, microglial viability and morphology was mainlyassessed after 24 h (Figs. 8 –10). The morphological changealso took place at 2 h (data not shown). Treatment of microgliawith LCN2 alone induced the morphological transformationwithout apparent cytotoxicity under this condition. Taken to-gether, these results suggest that amoeboid transformation ofmicroglia and their apoptotic sensitivity are closely associatedwith each other and that LCN2 carries out a dual function via aunique mechanism in the regulation of microglial apoptosis andmorphology.
DiscussionIn this study, we report that microglial secretory protein LCN2plays a dual role in determining morphological and functionalfate of activated microglia (Fig. 11). LCN2 appears to sensitizeactivated microglia to apoptosis and it also induces deramifi-cation of microglia. This was supported by the following re-sults: 1) the expression of lcn2 was down-regulated in the ap-
optosis-resistant microglia; 2) transfection of lcn2 cDNAconferred the apoptotic sensitivity; 3) shRNA-mediated knock-down of lcn2 expression diminished the apoptotic sensitivity,which was rescued by rLCN2 protein; 4) rLCN2 protein mim-icked the proapoptotic effect of lcn2 cDNA transfection; and 5)lcn2 cDNA transfection and rLCN2 protein induced deramifi-cation of microglia. Based on these results, it is speculated thatactivated microglia may secrete LCN2 proteins, which act in anautocrine manner to induce morphological transformation ofmicroglia (Fig. 11). At the same time, secreted LCN2 proteinsmay sensitize activated microglia to apoptotic signals, so thatactivated microglia can be easily eliminated by apoptosis as aself-regulatory mechanism. LCN2 protein is constitutively ex-pressed in microglia and its expression is strongly enhanced byinflammatory stimulation. The secreted LCN2 protein may feedback to microglia, and may induce the apoptosis-prone pheno-typic change in the microglia which have already been stimu-lated by inflammatory signals. It is also possible that secreted
FIGURE 10. Apoptosis-enhancing effectsof LCN2, ATP, forskolin, and A23187 inmicroglia. BV-2 cells (A), lcn2 shRNA-transfected BV-2 cells (B), or primary mi-croglia cultures (C) were exposed to LCN2proteins (10 �g/ml) or other stimuli (ATP, 3mM; forskolin, 10 �M; A23187, 5 �M) inthe presence of 0.5 mM SNP for 24 h, andthen the cell viability or apoptosis was de-termined by MTT assay (A and B, upper) orFITC-conjugated annexin V staining fol-lowed by counting of FITC-labeled cells un-der fluorescent microscope (C). Knockdownof lcn2 expression in the lcn2 shRNA-trans-fected BV-2 cells was confirmed by Westernblot analysis (B, lower). The expression oflcn2 was assessed after treatment with cal-cium ionophore A23187 (5 �M) for 8 h.Bars represent mean � SD values deter-mined from three independent experiments(A and B) or the percentage of apoptoticFITC-labeled cells, which represents threeindependent experiments (C). �, Statisticallysignificant differences from the treatmentwith SNP alone (A) or significant differencesbetween the two treatments (B).
FIGURE 11. Schematic diagram showing the dual role of lcn2 in theapoptosis and deramification of microglia. Once microglia are activated byinflammatory stimuli, LCN2 protein is secreted. Released LCN2 proteinfeeds back to microglia to induce morphological as well as functionalchanges. LCN2 induces deramification of microglia, and it also rendersmicroglia more sensitive to apoptotic signals, which may provide a basisfor the self-regulatory elimination of activated microglia in vivo. Based onthe studies using other deramification-inducing stimuli, the morphologicalchange of microglia appears to be closely related with the phenotypicchange to apoptosis-prone microglia.
3238 Lcn2 IN MICROGLIAL APOPTOSIS AND DERAMIFICATION
by guest on July 28, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

LCN2 protein may influence neighboring cells (astrocytes, neu-rons, and endothelial cells) in the brain. In the inflammatorysites in the CNS, the number of microglia may increase byproliferation or inward migration. Thus, the secretion of LCN2may be a part of an autoregulatory mechanism by which acti-vated microglia in the inflammatory sites could be safely re-moved (Fig. 11). This is in agreement with a previous report,where the role of secreted LCN2/24p3 in the termination of theimmune response has been proposed (30). Previously, it hasbeen reported that the production of RBC is also under thecontrol of LCN2 in a similar manner (56). LCN2 acted as anautocrine factor that negatively regulates RBC production byinducing apoptosis and inhibiting differentiation.
In the healthy brain, microglia have a typical ramified mor-phology consisting of a small cell body and long processes withsecondary branching. Changes in the microglial morphology areobserved in a wide range of the CNS pathologies such as braininjury, ischemia, infection, autoimmunity, and neurodegenera-tive diseases (7–9). Under these pathological conditions, ram-ified microglia transform into amoeboid microglia, with theirprocesses retracted and their cell body enlarged. Such a mor-phological transformation is associated with microglial activa-tion and is induced in vitro by a variety of stimuli includingLPS, IFN-�, laminin, ATP, etc. (4, 47, 54, 55, 57, 58). Never-theless, the mechanism underlying the morphological transfor-mation from ramified microglia into amoeboid microglia re-mains unknown. In the present study, we have unexpectedlyfound that LCN2, one of the secreted proteins of activated mi-croglia, plays a central role in the morphological changes. Upontransfection of BV-2 microglial cells with lcn2 cDNA, such amorphological transformation occurred. This was confirmed bythe treatment of primary microglia cultures with rLCN2 protein.Our results indicate for the first time that a single secretoryprotein of microglia actually mediates their own deramification.A similar type of morphological change has been also observedafter the transient expression of lcn2 in NIH3T3 fibroblast cells(J. Lee and K. Suk, unpublished results), indicating that theeffect of lcn2 on the cellular morphology may not be limited tomicroglia.
Having shown that lcn2 plays a dual role in microglial apo-ptosis and deramification, our next question was how these twoeffects of lcn2 are related with each other. To answer this ques-tion, we have used other stimuli that are known to induce themorphological change of microglia (47, 55, 57). Calcium iono-phore, ATP, and forskolin all induced the deramification of mi-croglia, and concurrently enhanced the sensitivity of microgliato the NO-induced apoptosis. These results indicate that micro-glial deramification is closely associated with the apoptotic sen-sitivity. The morphological change may be sufficient to increasethe apoptotic sensitivity. This speculation, however, needs to befurther tested using other conditions where microglial derami-fication takes place. Calcium ionophore exerted a unique effecton microglia in that it induced the microglial deramificationwith a concurrent induction of LCN2 expression, which furthersupported that lcn2 is responsible for microglial deramification.ATP and forskolin, however, did not enhance LCN2 expression,and yet induced the deramification and apoptotic sensitivity,suggesting that 1) ATP and forskolin may induce microglialderamification independently of LCN2 and 2) the microglialmorphology is the major factor in determining cell survival anddeath.
The results reported in this study were obtained using micro-glial cells cultured in vitro, which may not be the same as brainmicroglia in vivo. Functional significance of lcn2 in the micro-
glial apoptosis and morphological change in vivo has yet to bedetermined. The disruption of the blood-brain barrier followingCNS injury in vivo can lead to the recruitment of blood-bornemonocytes that are difficult to distinguish from the brain-resi-dent microglia (54, 59). Thus, the results on microglial mor-phology either in vitro or in vivo alone have to be interpretedwith caution. Recently, genome-wide expression profiling hasshown that LCN2 is up-regulated in brain tissue after focalcerebral ischemia (60, 61). Whether microglia are the cellularsource of LCN2 in brain, and whether the microglia-derivedLCN2 controls their own apoptosis and morphology in vivo,needs further investigation.
Proapoptotic activity of LCN2 is controversial. LCN2-in-duced apoptosis seems to occur in a cell type-specific fashion.A previous study even reported the role of LCN2 as a survivalfactor (32). Differences between mouse and human LCN2 interms of proapoptotic activity have been also reported (62).Devireddy et al. (30) has shown that most leukocytes are sen-sitive to LCN2-mediated apoptosis, while nonleukocytic celllines are not. Among many primary cells of leukocytic origintested, only macrophages as an exception were resistant to theLCN2-mediated apoptosis (30). As microglia are the CNSequivalent of macrophages, they were also resistant to theLCN2-mediated apoptosis as demonstrated in the current study.LCN2 alone did not induce apoptosis of microglia. LCN2merely increased the sensitivity of microglia to other apopto-genic stimuli. Thus, as least in macrophages and microglia, itcan be concluded that LCN2 is proapoptotic. It should be, how-ever, noted that molecular mechanisms underlying the apopto-sis-inducing effect and the apoptosis-sensitizing effect of LCN2may be different. Under the condition where LCN2 inducedapoptosis, it increased the expression of proapoptotic memberof Bcl-2 family proteins such as Bim through the recently iden-tified cell surface receptor for LCN2 (lcn2/24p3R). On the con-trary, such a change in the expression of Bcl-2 family proteinswas not observed in microglia, where LCN2 merely increasedthe apoptotic sensitivity. Also, phosphorylation of BAD atserine 136 was not detected (data not shown). Although theLCN2 receptor (lcn2/24p3R) is also expressed on microglia, itis possible that unidentified LCN2 receptors may be present onmicroglia whose proapoptotic pathways are not directly asso-ciated with the modification of Bcl-2 family protein expression.It should be also noted that lcn2-induced apoptosis occurred inthe cells that overexpressed lcn2/24p3R (29). The level of lcn2/24p3R may influence the fate of microglia; it may determinewhether LCN2 actually causes apoptosis or merely facilitatesthe apoptotic processes.
LCN2 has been implicated in mesenchymal-epithelial transi-tion. LCN2 is an endogenous epithelial inducer (33) and stim-ulates the epithelial phenotype of transformed cells suppressinginvasion and metastasis (63, 64). LCN2 also promotes tubulo-genesis by regulating epithelial morphogenesis (48). The ex-pression of E-cadherin was, however, not detected in primarymicroglia cultures by RT-PCR even after 40 cycles of amplifi-cation (data not shown). Also, there was no change in its ex-pression after LPS or LCN2 treatment, arguing against the roleof LCN2 in the mesenchymal-epithelial transition of microglia.It is not currently understood how the role of LCN2 as an ep-ithelial inducer is related with the proapoptotic and amoeboid-transforming effects of LCN2 found in the present study. Nev-ertheless, what these studies have in common is that LCN2 hasan important role in determining cellular morphology in addi-tion to its role in the cell survival and death.
3239The Journal of Immunology
by guest on July 28, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

In conclusion, we present evidence that a secretory proteinLCN2 is involved in the morphological transformation and apo-ptosis of activated microglia (Fig. 11). The amoeboid transforma-tion of microglia may be closely related with their vulnerability toapoptosis. Thus, activated microglia in vivo may secrete LCN2protein, which induces morphological transformation and facili-tates the self-regulatory apoptotic elimination. However, the exactmolecular mechanisms underlying the proapoptotic activity ofLCN2 and the morphological changes of microglia need to beelucidated in the future studies.
DisclosuresThe authors have no financial conflict of interest.
References1. Stoll, G., and S. Jander. 1999. The role of microglia and macrophages in the
pathophysiology of the CNS. Prog. Neurobiol. 58: 233–247.2. Kim, S. U., and J. de Vellis. 2005. Microglia in health and disease. J. Neurosci.
Res. 81: 302–313.3. Streit, W. J., R. E. Mrak, and W. S. Griffin. 2004. Microglia and neuroinflam-
mation: a pathological perspective. J. Neuroinflammation 1: 14.4. Giulian, D., J. Li, S. Bartel, J. Broker, X. Li, and J. B. Kirkpatrick. 1995. Cell
surface morphology identifies microglia as a distinct class of mononuclear phago-cyte. J. Neurosci. 15: 7712–7726.
5. Aloisi, F. 2001. Immune function of microglia. Glia 36: 165–179.6. Suk, K. 2006. Proteomics-based discovery of biomarkers and therapeutic targets
in neurodegenerative diseases: perspective of microglia and neuroinflammation.Expert Opin. Ther. Pat. 16: 237–247.
7. Perry, V. H., and S. Gordon. 1988. Macrophages and microglia in the nervoussystem. Trends Neurosci. 11: 273–277.
8. Raivich, G., M. Bohatschek, C. U. Kloss, A. Werner, L. L. Jones, andG. W. Kreutzberg. 1999. Neuroglial activation repertoire in the injured brain:graded response, molecular mechanisms and cues to physiological function.Brain Res. Brain Res. Rev. 30: 77–105.
9. Suzumura, A., T. Marunouchi, and H. Yamamoto. 1991. Morphological trans-formation of microglia in vitro. Brain Res. 545: 301–306.
10. Gehrmann, J., Y. Matsumoto, and G. W. Kreutzberg. 1995. Microglia: intrinsicimmuneffector cell of the brain. Brain Res. Brain Res. Rev. 20: 269–287.
11. Block, M. L., L. Zecca, and J. S. Hong. 2007. Microglia-mediated neurotoxicity:uncovering the molecular mechanisms. Nat. Rev. Neurosci. 8: 57–69.
12. Gonzalez-Scarano, F., and G. Baltuch. 1999. Microglia as mediators of inflam-matory and degenerative diseases. Annu. Rev. Neurosci. 22: 219–240.
13. Suk, K. 2005. Regulation of neuroinflammation by herbal medicine and its im-plications for neurodegenerative diseases: a focus on traditional medicines andflavonoids. Neurosignals 14: 23–33.
14. Mrak, R. E., and W. S. Griffin. 2005. Glia and their cytokines in progression ofneurodegeneration. Neurobiol. Aging 26: 349–354.
15. Liu, B., K. Wang, H. M. Gao, B. Mandavilli, J. Y. Wang, and J. S. Hong. 2001.Molecular consequences of activated microglia in the brain: overactivation in-duces apoptosis. J. Neurochem. 77: 182–189.
16. Lee, P., J. Lee, S. Kim, H. Yagita, M. S. Lee, S. Y. Kim, H. Kim, and K. Suk.2001. NO as an autocrine mediator in the apoptosis of activated microglial cells:correlation between activation and apoptosis of microglial cells. Brain Res. 892:380–385.
17. Kingham, P. J., M. L. Cuzner, and J. M. Pocock. 1999. Apoptotic pathwaysmobilized in microglia and neurones as a consequence of chromogranin A-in-duced microglial activation. J. Neurochem. 73: 538–547.
18. Jones, L. L., R. B. Banati, M. B. Graeber, L. Bonfanti, G. Raivich, andG. W. Kreutzberg. 1997. Population control of microglia: does apoptosis play arole? J. Neurocytol. 26: 755–770.
19. Lee, J., J. Hur, P. Lee, J. Y. Kim, N. Cho, M. S. Lee, S. Y. Kim, H. Kim, andK. Suk. 2001. Dual role of inflammatory stimuli in activation-induced cell deathof mouse microglial cells: initiation of two separate apoptotic pathways via in-duction of interferon regulatory factor-1 and caspase-11. J. Biol. Chem. 276:32956–32965.
20. Suk, K., J. Lee, J. Hur, Y. S. Kim, M. S. Lee, S. H. Cha, S. Y. Kim, and H. Kim.2001. Activation-induced cell death of rat astrocytes. Brain Res. 900: 342–347.
21. Suk, K. 2005. Role of caspases in activation-induced cell death of neuroglia.Curr. Enzyme Inhibition 1: 43–50.
22. Flower, D. R., A. C. North, and C. E. Sansom. 2000. The lipocalin protein family:structural and sequence overview. Biochim. Biophys. Acta 1482: 9–24.
23. Kjeldsen, L., J. B. Cowland, and N. Borregaard. 2000. Human neutrophil gela-tinase-associated lipocalin and homologous proteins in rat and mouse. Biochim.Biophys. Acta 1482: 272–283.
24. Flower, D. R., A. C. North, and T. K. Attwood. 1991. Mouse oncogene protein24p3 is a member of the lipocalin protein family. Biochem. Biophys. Res. Com-mun. 180: 69–74.
25. Hamilton, R. T., M. Nilsen-Hamilton, and G. Adams. 1985. Superinduction bycycloheximide of mitogen-induced secreted proteins produced by BALB/c 3T3cells. J. Cell. Physiol. 123: 201–208.
26. Borregaard, N., and J. B. Cowland. 2006. Neutrophil gelatinase-associated li-pocalin, a siderophore-binding eukaryotic protein. Biometals 19: 211–215.
27. Kjeldsen, L., A. H. Johnsen, H. Sengelov, and N. Borregaard. 1993. Isolation andprimary structure of NGAL, a novel protein associated with human neutrophilgelatinase. J. Biol. Chem. 268: 10425–10432.
28. Yousefi, S., and H. U. Simon. 2002. Granulocyte apoptosis: death by a secretedlipocalin? Cell Death Differ. 9: 595–597.
29. Devireddy, L. R., C. Gazin, X. Zhu, and M. R. Green. 2005. A cell-surfacereceptor for lipocalin 24p3 selectively mediates apoptosis and iron uptake. Cell123: 1293–1305.
30. Devireddy, L. R., J. G. Teodoro, F. A. Richard, and M. R. Green. 2001. Inductionof apoptosis by a secreted lipocalin that is transcriptionally regulated by IL-3deprivation. Science 293: 829–834.
31. Tong, Z., X. Wu, and J. P. Kehrer. 2003. Increased expression of the lipocalin24p3 as an apoptotic mechanism for MK886. Biochem. J. 372: 203–210.
32. Tong, Z., X. Wu, D. Ovcharenko, J. Zhu, C. S. Chen, and J. P. Kehrer. 2005.Neutrophil gelatinase-associated lipocalin as a survival factor. Biochem. J. 391:441–448.
33. Yang, J., D. Goetz, J. Y. Li, W. Wang, K. Mori, D. Setlik, T. Du,H. Erdjument-Bromage, P. Tempst, R. Strong, and J. Barasch. 2002. An irondelivery pathway mediated by a lipocalin. Mol. Cell. 10: 1045–1056.
34. Mishra, J., K. Mori, Q. Ma, C. Kelly, J. Yang, M. Mitsnefes, J. Barasch, andP. Devarajan. 2004. Amelioration of ischemic acute renal injury by neutrophilgelatinase-associated lipocalin. J. Am. Soc. Nephrol. 15: 3073–3082.
35. Mori, K., H. T. Lee, D. Rapoport, I. R. Drexler, K. Foster, J. Yang,K. M. Schmidt-Ott, X. Chen, J. Y. Li, S. Weiss, et al. 2005. Endocytic deliveryof lipocalin-siderophore-iron complex rescues the kidney from ischemia-reper-fusion injury. J. Clin. Invest. 115: 610–621.
36. Playford, R. J., A. Belo, R. Poulsom, A. J. Fitzgerald, K. Harris, I. Pawluczyk,J. Ryon, T. Darby, M. Nilsen-Hamilton, S. Ghosh, and T. Marchbank. 2006.Effects of mouse and human lipocalin homologues 24p3/lcn2 and neutrophil ge-latinase-associated lipocalin on gastrointestinal mucosal integrity and repair.Gastroenterology 131: 809–817.
37. Berger, T., A. Togawa, G. S. Duncan, A. J. Elia, A. You-Ten, A. Wakeham,H. E. Fong, C. C. Cheung, and T. W. Mak. 2006. Lipocalin 2-deficient miceexhibit increased sensitivity to Escherichia coli infection but not to ischemia-reperfusion injury. Proc. Natl. Acad. Sci. USA 103: 1834–1839.
38. Flo, T. H., K. D. Smith, S. Sato, D. J. Rodriguez, M. A. Holmes, R. K. Strong,S. Akira, and A. Aderem. 2004. Lipocalin 2 mediates an innate immune responseto bacterial infection by sequestrating iron. Nature 432: 917–921.
39. Yan, L., N. Borregaard, L. Kjeldsen, and M. A. Moses. 2001. The high molecularweight urinary matrix metalloproteinase (MMP) activity is a complex of gelati-nase B/MMP-9 and neutrophil gelatinase-associated lipocalin (NGAL): modula-tion of MMP-9 activity by NGAL. J. Biol. Chem. 276: 37258–37265.
40. Liu, Q., and M. Nilsen-Hamilton. 1995. Identification of a new acute phase pro-tein. J. Biol. Chem. 270: 22565–22570.
41. Hvidberg, V., C. Jacobsen, R. K. Strong, J. B. Cowland, S. K. Moestrup, andN. Borregaard. 2005. The endocytic receptor megalin binds the iron transportingneutrophil-gelatinase-associated lipocalin with high affinity and mediates its cel-lular uptake. FEBS Lett. 579: 773–777.
42. Blasi, E., R. Barluzzi, V. Bocchini, R. Mazzolla, and F. Bistoni. 1990. Immor-talization of murine microglial cells by a v-raf/v-myc carrying retrovirus. J Neu-roimmunol. 27: 229–237.
43. Bocchini, V., R. Mazzolla, R. Barluzzi, E. Blasi, P. Sick, and H. Kettenmann.1992. An immortalized cell line expresses properties of activated microglial cells.J. Neurosci. Res. 31: 616–621.
44. Saura, J., J. M. Tusell, and J. Serratosa. 2003. High-yield isolation of murinemicroglia by mild trypsinization. Glia 44: 183–189.
45. Jung, J. W., J. S. Park, J. W. Hwang, K. S. Kang, Y. S. Lee, B. S. Song, G. J. Lee,C. D. Yeo, J. S. Kang, W. S. Lee, et al. 2004. Gene expression analysis ofperoxisome proliferators- and phenytoin-induced hepatotoxicity using cDNA mi-croarray. J. Vet. Med. Sci. 66: 1329–1333.
46. Lee, H., Y. O. Kim, S. Y. Kim, H. Kim, H. S. Noh, S. S. Kang, G. J. Cho,W. S. Choi, and K. Suk. 2003. Flavonoid wogonin from medicinal herb is neu-roprotective by inhibiting inflammatory activation of microglia. FASEB J. 17:1943–1944.
47. Kalla, R., M. Bohatschek, C. U. Kloss, J. Krol, X. Von Maltzan, and G. Raivich.2003. Loss of microglial ramification in microglia-astrocyte cocultures: involve-ment of adenylate cyclase, calcium, phosphatase, and Gi-protein systems. Glia41: 50–63.
48. Gwira, J. A., F. Wei, S. Ishibe, J. M. Ueland, J. Barasch, and L. G. Cantley. 2005.Expression of neutrophil gelatinase-associated lipocalin regulates epithelial mor-phogenesis in vitro. J. Biol. Chem. 280: 7875–7882.
49. Persengiev, S. P., L. R. Devireddy, and M. R. Green. 2002. Inhibition of apo-ptosis by ATFx: a novel role for a member of the ATF/CREB family of mam-malian bZIP transcription factors. Genes Dev. 16: 1806–1814.
50. Kim, H. J., I. Tsoy, M. K. Park, Y. S. Lee, J. H. Lee, H. G. Seo, and K. C. Chang.2006. Iron released by sodium nitroprusside contributes to heme oxygenase-1induction via the cAMP-protein kinase A-mitogen-activated protein kinase path-way in RAW 264.7 cells. Mol. Pharmacol. 69: 1633–1640.
51. Meheus, L. A., L. M. Fransen, J. G. Raymackers, H. A. Blockx, J. J.Van Beeumen, S. M. Van Bun, and A. Van de Voorde. 1993. Identification bymicrosequencing of lipopolysaccharide-induced proteins secreted by mouse mac-rophages. J. Immunol. 151: 1535–1547.
52. Cowland, J. B., O. E. Sorensen, M. Sehested, and N. Borregaard. 2003. Neutro-phil gelatinase-associated lipocalin is up-regulated in human epithelial cells byIL-1�, but not by TNF-�. J. Immunol. 171: 6630–6639.
3240 Lcn2 IN MICROGLIAL APOPTOSIS AND DERAMIFICATION
by guest on July 28, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

53. Jung, D. Y., H. Lee, B. Y. Jung, J. Ock, M. S. Lee, W. H. Lee, and K. Suk. 2005.TLR4, but not TLR2, signals autoregulatory apoptosis of cultured microglia: acritical role of IFN-� as a decision maker. J. Immunol. 174: 6467–6476.
54. Kloss, C. U., M. Bohatschek, G. W. Kreutzberg, and G. Raivich. 2001. Effect oflipopolysaccharide on the morphology and integrin immunoreactivity of ramifiedmicroglia in the mouse brain and in cell culture. Exp. Neurol. 168: 32–46.
55. Xiang, Z., M. Chen, J. Ping, P. Dunn, J. Lv, B. Jiao, and G. Burnstock. 2006.Microglial morphology and its transformation after challenge by extracellularATP in vitro. J. Neurosci. Res. 83: 91–101.
56. Miharada, K., T. Hiroyama, K. Sudo, T. Nagasawa, and Y. Nakamura. 2005.Lipocalin 2 functions as a negative regulator of red blood cell production in anautocrine fashion. FASEB J. 19: 1881–1883.
57. Bohatschek, M., C. U. Kloss, R. Kalla, and G. Raivich. 2001. In vitro model ofmicroglial deramification: ramified microglia transform into amoeboid phago-cytes following addition of brain cell membranes to microglia-astrocyte cocul-tures. J. Neurosci. Res. 64: 508–522.
58. Chamak, B., and M. Mallat. 1991. Fibronectin and laminin regulate the in vitrodifferentiation of microglial cells. Neuroscience 45: 513–527.
59. Andersson, P. B., V. H. Perry, and S. Gordon. 1992. The acute inflammatoryresponse to lipopolysaccharide in CNS parenchyma differs from that in otherbody tissues. Neuroscience 48: 169–186.
60. MacManus, J. P., T. Graber, C. Luebbert, E. Preston, I. Rasquinha, B. Smith, andJ. Webster. 2004. Translation-state analysis of gene expression in mouse brainafter focal ischemia. J. Cereb. Blood Flow Metab. 24: 657–667.
61. Trendelenburg, G., K. Prass, J. Priller, K. Kapinya, A. Polley, C. Muselmann,K. Ruscher, U. Kannbley, A. O. Schmitt, S. Castell, et al. 2002. Serial analysisof gene expression identifies metallothionein-II as major neuroprotective gene inmouse focal cerebral ischemia. J. Neurosci. 22: 5879–5888.
62. Klausen, P., C. U. Niemann, J. B. Cowland, K. Krabbe, and N. Borregaard. 2005.On mouse and man: neutrophil gelatinase associated lipocalin is not involved inapoptosis or acute response. Eur. J. Haematol. 75: 332–340.
63. Hanai, J., T. Mammoto, P. Seth, K. Mori, S. A. Karumanchi, J. Barasch, andV. P. Sukhatme. 2005. Lipocalin 2 diminishes invasiveness and metastasis ofRas-transformed cells. J. Biol. Chem. 280: 13641–13647.
64. Lee, H. J., E. K. Lee, K. J. Lee, S. W. Hong, Y. Yoon, and J. S. Kim. 2006.Ectopic expression of neutrophil gelatinase-associated lipocalin suppresses theinvasion and liver metastasis of colon cancer cells. Int. J. Cancer 118:2490–2497.
3241The Journal of Immunology
by guest on July 28, 2018http://w
ww
.jimm
unol.org/D
ownloaded from