Embed Size (px)
Citation preview

THE JOURNAL OF BIOLOGICAL CHEMISTRY 8 1987 by The American Society of Biological Chemists, Inc.
Vol. 262, No. 7, Issue of March 5, pp. 3376-3381, 1987 Printed in U.S.A.
Association of Heparan Sulfate Proteoglycan and Laminin with the Cytoskeleton in Rat Liver*
(Received for publication, September 8, 1986)
David J. Carey$, Colleen M. Rafferty, and Margaret M. Schramm From the Department of Physiology, The Milton S. Hershey Medical Center, Hershey, Pennsylvania 17033
Rats were injected with 35S04 and after 2 h their livers were removed and used to prepare a detergent- insoluble cytoskeleton fraction. Spectrin, cytokera- tins, and actin were major protein components of the isolated cytoskeletons. The cytoskeleton fraction ac- counted for approximately 14% of the total trichloro- acetic acid-insoluble 35S04 radioactivity incorporated into the liver. The cytoskeleton-associated radioactiv- ity was present in a single species of macromolecule. This molecule was not present to a significant extent in the detergent-soluble fraction containing the cell supernatant and dissolved membrane proteins. Further characterization revealed the cytoskeleton-associated molecule was a heparan sulfate proteoglycan: it was eluted from a Sepharose CL-4B column under denatur- ing conditions at K,, = 0.4; following mild alkaline hydrolysis the radioactivity was eluted at a Kay = 0.7; when this material was subjected to nitrous acid hy- drolysis all of the radioactivity was eluted near the column included volume. The isolated cytoskeletons contained attached nuclei. Pure nuclei isolated without associated cytoskeletal elements contained less than 1% of the total liver trichloroacetic acid-insoluble “5s04 radioactivity and no detectable heparan sulfate proteoglycan. These results suggested that other ma- trix proteins might be associated with the liver cyto- skeleton. When the subcellular distribution of laminin was monitored by immunostaining proteins trans- ferred to nitrocellulose, laminin was detected exclu- sively in the cytoskeleton fraction. These results pro- vide evidence for an association between extracellular connective tissue proteins and intracellular structural proteins.
Heparan sulfate proteoglycans are ubiquitous components of extracellular matrices and cell surfaces (1-5). Although the precise functions of these molecules are not known, they have been suggested to play a role in extracellular matrix assembly (6-8), cell-matrix adhesion (9, lo), receptor modulation (ll), and assembly of supramolecular membrane complexes (12).
An important principle that has emerged concerning the function of extracellular matrix and cell surface molecules is that they interact, through the cell membrane, with the cell’s cytoskeleton (13-19). Such an interaction provides a plausible explanation for the diverse effects of extracellular matrix contact on cell behavior, which includes effects on cell shape (19, 20), motility (21-23), and differentiation (14, 20-24).
* This work was supported by Research Grant NS-21925 from the National Institutes of Health. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “aduertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
$ To whom correspondence should be addressed.
Unfortunately, the details of the association between extra- cellular matrix proteins and the cytoskeleton are not known.
Recently, we have presented evidence that a cell surface heparan sulfate proteoglycan produced by cultured rat Schwann cells is bound to the Schwann cell cytoskeleton (25). Thus, this molecule could serve as a receptor for basement membrane proteins secreted by Schwann cells and provide a molecular link between the extracellular matrix and the Schwann cell cytoskeleton. Schwann cell terminal differentia- tion, which includes dramatic changes in Schwann cell mor- phology, is dependent upon contact with the Schwann cell basement membrane (20).
The purpose of these studies was to investigate in more detail the association of heparan sulfate proteoglycan with the cytoskeleton in a tissue that was amenable to biochemical analysis and that would permit us to study the nature of the heparan sulfate proteoglycan-cytoskeleton interaction at the molecular level, including the eventual isolation and purifi- cation of the proteoglycan. The data presented here show that purified rat liver cytoskeleton fractions contain a unique heparan sulfate proteoglycan that is not present in the cell supernatant or nuclear fractions. The data also indicate that the basement membrane glycoprotein laminin is tightly but noncovalently associated with the liver cytoskeleton.
MATERIALS AND METHODS
Radiolabeling-Adult female Sprague-Dawley rats were injected intraperitoneally with 35S04 (1 mCi in 1 ml of 0.05 M sodium phos- phate, pH 7.5, 0.15 M NaCl; purchased from ICN Radiochemicals, Irvine, Irvine, CA as carrier-free sulfuric acid). Two hours after injection the rats were anesthetized with ether and the livers were removed and placed into ice-cold 0.05 M sodium phosphate, pH 7.5, 0.15 M NaC1. The excised livers were processed immediately as described below. All subsequent procedures were conducted at 4 “C. Livers from these rats contained 4-8 X lo6 cpm of trichloroacetic acid-insoluble radioactivity per liver.
Isolation of Liver Cytoskeletons-A purified cytoskeleton fraction from rat liver was isolated by a modification of a previously published method (25). The excised livers were homogenized in 2 volumes (ml/ g, wet weight) of cytoskeleton buffer (0.15 M NaCl, 10 mM Tris-HC1, pH 7.2, 5 mM MgC12, 2 mM EGTA,’ 0.25 mM dithiothreitol, 1 mM phenylmethylsulfonyl fluoride, 1% Triton X-100) in a hand-held dounce homogenizer (three passes each with a loose and a tight pestle). Anti-foam A (Sigma) was included in the homogenization buffer at a concentration of 0.01% to reduce foaming. The homoge- nates were filtered through four layers of gauze and then centrifuged in a Sorvall SS-34 rotor at 11,000 rpm for 2 min. The resulting supernatants (equivalent to the cell supernatant fraction) were re- moved and the pellets were resuspended in a volume of fresh cyto- skeleton buffer equal to that of the original homogenate. The resus- pended pellets were centrifuged as above. The supernatants (wash fraction) were removed and the pellets (cytoskeletons) were resus- pended in a volume of cytoskeleton buffer equivalent to 0.6 ml/g of tissue using a dounce homogenizer.
The abbreviations used are: EGTA, [ethylenebis(oxyethylene- nitri1o)tetraacetic acid; SDS, sodium dodecyl sulfate.
3376
Cytoskeleton-associated Proteoglycan and Laminin 3377 Additional purification of the cytoskeleton fraction was achieved
by centrifugation in sucrose density step gradients. The resuspended cytoskeleton fraction was layered over a step gradient formed by 8 ml each of 40% and 20% (w/w) sucrose in cytoskeleton buffer. The tubes were centrifuged for 10 min at 12,000 X gav. The contents were divided into 4 fractions equivalent to the sample solution plus the 20% sucrose layer (fraction 1, containing soluble proteins), the 20/ 40% sucrose interphase (fraction 2, containing the purified cytoskel- etons), the 40% sucrose layer (fraction 3), and the pellet (fraction 4, containing small amounts of cytoskeletal proteins and nuclei).
Isolation of Rat Liuer Nuclei-Nuclei were purified from rat liver by the method of Blobel and Potter (26). Briefly, excised livers were homogenized in ice-cold 0.25 M sucrose, 50 mM Tris-HCI, pH 7.5, 25 mM KCI, 5 mM MgClz, in a motor driven glass Teflon tissue grinder. The homogenate was filtered through four layers of gauze, diluted with 2 volumes of 2.3 M sucrose, 50 mM Tris-HCI, pH 7.5, 25 mM KCI, 5 mM MgC12, and then layered over a solution of 2.3 M sucrose, in the same buffer. After centrifugation for 30 min at 124,000 X g,,, the supernatant was removed. The pellet, which contained the puri- fied nuclei, was resuspended in a small volume of homogenization buffer.
Electrophoresis and Zmmunoblotting-Aliquots of various cellular fractions were subjected to SDS-slab gel electrophoresis using the buffer system of Laemmli (27). The gels were either stained with Coomassie Brilliant Blue R, dried, and subjected to autoradiography or transferred to nitrocellulose for staining with antisera. In some cases gels that had been stained with Coomassie Blue were scanned with a Bio-Rad Model 1650 scanning densitometer to quantitate the amounts of various protein bands.
Gels to be immunostained were soaked for 30 min in transfer buffer (12.5 mM Tris base, 96 mM glycine) and the proteins were electro- phoretically transferred to nitrocellulose (Bio-Rad) at a constant voltage of 70 V for 40 min. Following transfer the nitrocellulose sheets were soaked for 1 h in blocking solution (50 mM Tris-HCI, pH 7.4, 100 mM NaCI, 5% Carnation instant non-fat dry milk (28)). The sheets were then incubated overnight a t 4 "C in the first antibody diluted in blocking solution. After rinsing, the sheets were soaked for 1-2 h in a solution containing affinity purified goat anti-rabbit IgG- conjugated to horseradish peroxidase (Bio-Rad, diluted 1:2000 in blocking solution). After rinsing, the bound antibodies were visualized by their peroxidase reaction products with 4-chloro-1-naphthol as substrate.
Proteoglycan Analysis-35S04-labeled proteoglycans were analyzed as described previously (25, 29). Aliquots of cell supernatant or purified cytoskeleton fractions were diluted into guanidine buffer (4 M guanidine HCI, 50 mM sodium acetate, pH 5.0, 0.1% Triton X- loo), applied to a 1 X 100-cm column of Sepharose CL-4B, and eluted with guanidine buffer at a flow rate of 6 ml/h. Aliquots of column fractions were analyzed by liquid scintillation counting to localize radiolabeled peaks. In all the experiments reported here, the recovery of radioactive material from the columns was 85% or greater. Peak tubes were pooled and the radiolabeled macromolecules were precip- itated at -20 "C by addition of 4 volumes of 1.3% potassium acetate in 95% ethanol. Glycosaminoglycan chains were released from pro- teoglycans by hydrolysis in 0.05 M NaOH, 1 M NaBH, at 45 "C for 16 h. Heparan sulfate chains were hydrolyzed by nitrous acid (1.9 N acetic acid, 0.25 M sodium nitrite) for 2 h at room temperature.
Antisera-Antiserum to laminin was produced in a rabbit immu- nized with purified mouse laminin (generously provided by Dr. Hynda Kleinman, National Institutes of Health, Bethesda, MD). Multiple subcutaneous injections of 100 gg of laminin in complete (first injec- tion) or incomplete (subsequent injections) Freund's adjuvant were given at 2-week intervals. Antibody titer and specificity were tested by staining of proteins transferred to nitrocellulose as described above.
Antibodies to mouse brain spectrin, mouse red blood cell protein 4.1, and mouse red blood cell protein 2.1 (ankryn) (30) were gener- ously provided by Dr. Steven Goodman, Hershey Medical Center.
RESULTS
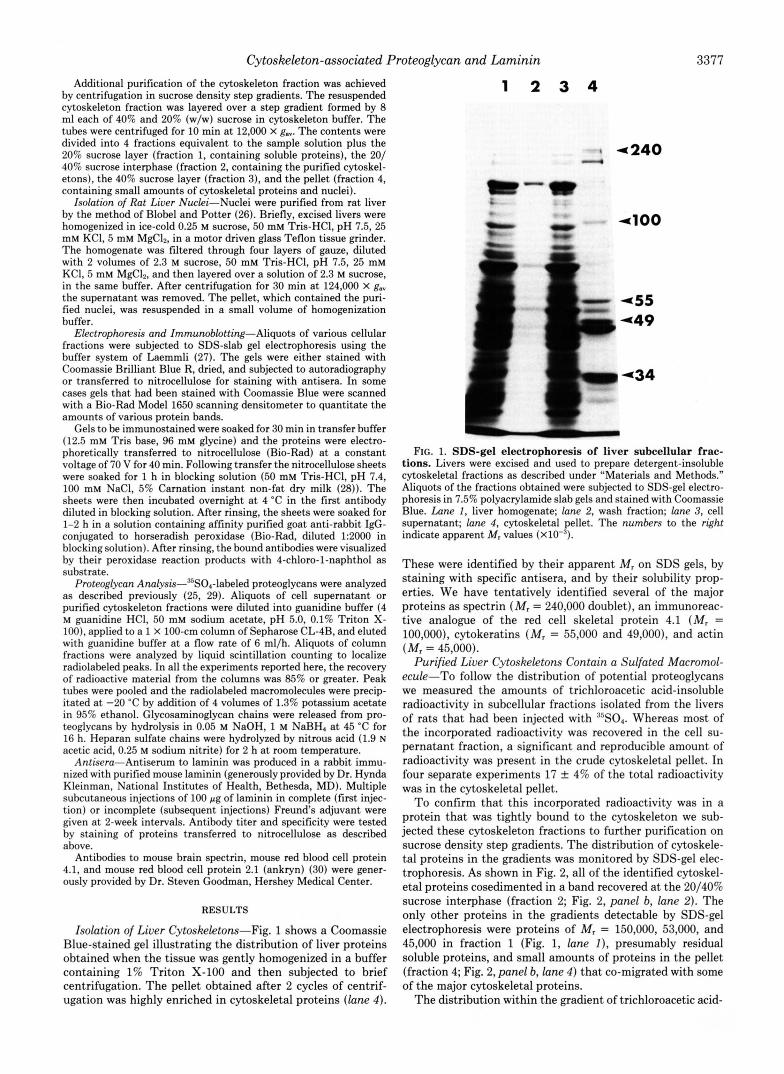
Isolation of Liver Cytoskeletom-Fig. 1 shows a Coomassie Blue-stained gel illustrating the distribution of liver proteins obtained when the tissue was gently homogenized in a buffer containing 1% Triton X-100 and then subjected to brief centrifugation. The pellet obtained after 2 cycles of centrif- ugation was highly enriched in cytoskeletal proteins (lane 4 ) .
1 2 3 4
a100
455 tao -7.
434
FIG. 1. SDS-gel electrophoresis of liver subcellular frac- tions. Livers were excised and used to prepare detergent-insoluble cytoskeletal fractions as described under "Materials and Methods." Aliquots of the fractions obtained were subjected to SDS-gel electro- phoresis in 7.5% polyacrylamide slab gels and stained with Coomassie Blue. Lane I , liver homogenate; lane 2, wash fraction; lane 3, cell supernatant; lane 4, cytoskeletal pellet. The numbers to the right indicate apparent M, values ( x ~ O - ' ) .
These were identified by their apparent M, on SDS gels, by staining with specific antisera, and by their solubility prop- erties. We have tentatively identified several of the major proteins as spectrin (M, = 240,000 doublet), an immunoreac- tive analogue of the red cell skeletal protein 4.1 (M, = 100,000), cytokeratins (MI = 55,000 and 49,000), and actin (M, = 45,000).
Purified Liver Cytoskeletons Contain a Sulfated Macromol- ecule-To follow the distribution of potential proteoglycans we measured the amounts of trichloroacetic acid-insoluble radioactivity in subcellular fractions isolated from the livers of rats that had been injected with "SO,. Whereas most of the incorporated radioactivity was recovered in the cell su- pernatant fraction, a significant and reproducible amount of radioactivity was present in the crude cytoskeletal pellet. In four separate experiments 17 & 4% of the total radioactivity was in the cytoskeletal pellet.
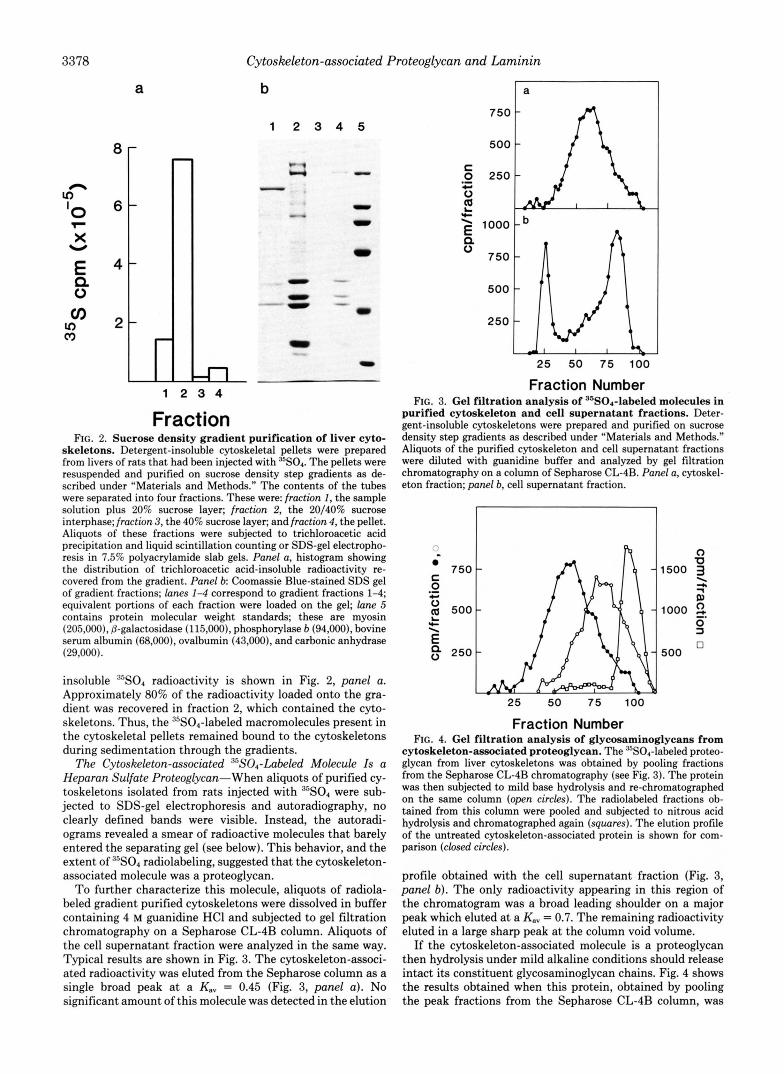
To confirm that this incorporated radioactivity was in a protein that was tightly bound to the cytoskeleton we sub- jected these cytoskeleton fractions to further purification on sucrose density step gradients. The distribution of cytoskele- tal proteins in the gradients was monitored by SDS-gel elec- trophoresis. As shown in Fig. 2, all of the identified cytoskel- eta1 proteins cosedimented in a band recovered at the 20/40% sucrose interphase (fraction 2; Fig. 2, panel b, lane 2) . The only other proteins in the gradients detectable by SDS-gel electrophoresis were proteins of M, = 150,000, 53,000, and 45,000 in fraction 1 (Fig. 1, lane I ) , presumably residual soluble proteins, and small amounts of proteins in the pellet (fraction 4; Fig. 2, panel b, lane 4 ) that co-migrated with some of the major cytoskeletal proteins.
The distribution within the gradient of trichloroacetic acid-
Cytoskeleton-associated Proteoglycan and Laminin
a
8 -
6 -
4 -
2- n b
1 2 3 4 5
1 Y W
LlLbl - 1 2 3 4
Fraction FIG. 2. Sucrose density gradient purification of liver cyto-
skeletons. Detergent-insoluble cytoskeletal pellets were prepared from livers of rats that had been injected with 35s04. The pellets were resuspended and purified on sucrose density step gradients as de- scribed under "Materials and Methods." The contents of the tubes were separated into four fractions. These were: fraction 1, the sample solution plus 20% sucrose layer; fraction 2, the 20/40% sucrose interphase; fraction 3, the 40% sucrose layer; and fraction 4, the pellet. Aliquots of these fractions were subjected to trichloroacetic acid precipitation and liquid scintillation counting or SDS-gel electropho- resis in 7.5% polyacrylamide slab gels. Panel a, histogram showing the distribution of trichloroacetic acid-insoluble radioactivity re- covered from the gradient. Panel b Coomassie Blue-stained SDS gel of gradient fractions; lanes 1-4 correspond to gradient fractions 1-4; equivalent portions of each fraction were loaded on the gel; lane 5 contains protein molecular weight standards; these are myosin (205,000), 6-galactosidase (115,000), phosphorylase b (94,000). bovine serum albumin (68,000), ovalbumin (43,000), and carbonic anhydrase (29,000).
insoluble 35s04 radioactivity is shown in Fig. 2, panel a. Approximately 80% of the radioactivity loaded onto the gra- dient was recovered in fraction 2, which contained the cyto- skeletons. Thus, the 35S04-labeled macromolecules present in the cytoskeletal pellets remained bound to the cytoskeletons during sedimentation through the gradients.
The Cytoskeleton-associated ""S4-Labeled Molecule Is a Heparan Sulfate Proteoglycan-When aliquots of purified cy- toskeletons isolated from rats injected with 35s04 were sub- jected to SDS-gel electrophoresis and autoradiography, no clearly defined bands were visible. Instead, the autoradi- ograms revealed a smear of radioactive molecules that barely entered the separating gel (see below). This behavior, and the extent of 3sS04 radiolabeling, suggested that the cytoskeleton- associated molecule was a proteoglycan.
To further characterize this molecule, aliquots of radiola- beled gradient purified cytoskeletons were dissolved in buffer containing 4 M guanidine HCl and subjected to gel filtration chromatography on a Sepharose CL-4B column. Aliquots of the cell supernatant fraction were analyzed in the same way. Typical results are shown in Fig. 3. The cytoskeleton-associ- ated radioactivity was eluted from the Sepharose column as a single broad peak at a K., = 0.45 (Fig. 3, panel a). No significant amount of this molecule was detected in the elution
l a
25 50 75 100
Fraction Number hc. 3. Gel filtration analysis of S5S04-labeled molecules in
purified cytoskeleton and cell supernatant fractions. Deter- gent-insoluble cytoskeletons were prepared and purified on sucrose density step gradients as described under "Materials and Methods." Aliquots of the purified cytoskeleton and cell supernatant fractions were diluted with guanidine buffer and analyzed by gel filtration chromatography on a column of Sepharose CL-4B. Panel a, cytoskel- eton fraction; panel b, cell supernatant fraction.
3 .. e C 0
750
.- w
500 .c \
E 2 250
n 0
1500 3 2 z
1000 g 0 3
500 0
25 50 75 100
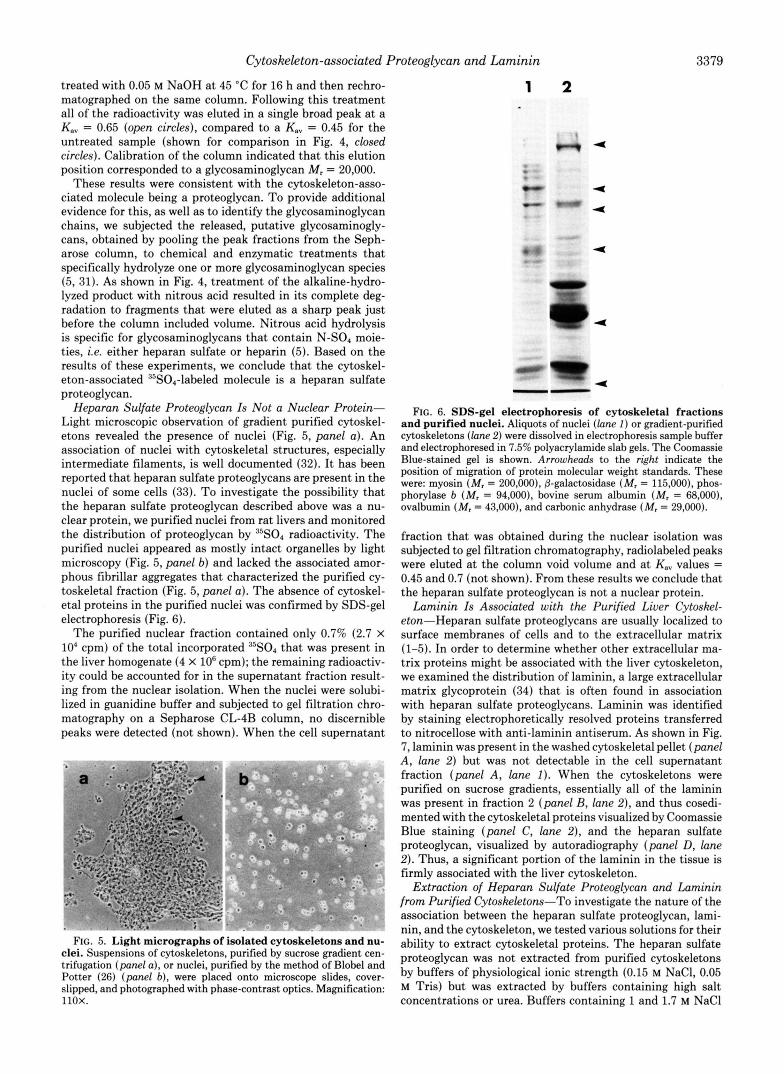
Fraction Number FIG. 4. Gel filtration analysis of glycosaminoglycans from
cytoskeleton-associated proteoglycan. The 35S04-labeled proteo- glycan from liver cytoskeletons was obtained by pooling fractions from the Sepharose CL-4B chromatography (see Fig. 3). The protein was then subjected to mild base hydrolysis and re-chromatographed on the same column (open circles). The radiolabeled fractions ob- tained from this column were pooled and subjected to nitrous acid hydrolysis and chromatographed again (squares). The elution profile of the untreated cytoskeleton-associated protein is shown for com- parison (closed circles).
profile obtained with the cell supernatant fraction (Fig. 3, panel b). The only radioactivity appearing in this region of the chromatogram was a broad leading shoulder on a major peak which eluted a t a K., = 0.7. The remaining radioactivity eluted in a large sharp peak at the column void volume.
If the cytoskeleton-associated molecule is a proteoglycan then hydrolysis under mild alkaline conditions should release intact its constituent glycosaminoglycan chains. Fig. 4 shows the results obtained when this protein, obtained by pooling the peak fractions from the Sepharose CL-4B column, was
Cytoskeleton-associated Proteoglycan and Laminin
treated with 0.05 M NaOH at 45 "C for 16 h and then rechro- matographed on the same column. Following this treatment all of the radioactivity was eluted in a single broad peak a t a K., = 0.65 (open circles), compared to a K,, = 0.45 for the untreated sample (shown for comparison in Fig. 4, closed circles). Calibration of the column indicated that this elution position corresponded to a glycosaminoglycan M , = 20,000.
These results were consistent with the cytoskeleton-asso- ciated molecule being a proteoglycan. To provide additional evidence for this, as well as to identify the glycosaminoglycan chains, we subjected the released, putative glycosaminogly- cans, obtained by pooling the peak fractions from the Seph- arose column, to chemical and enzymatic treatments that specifically hydrolyze one or more glycosaminoglycan species (5, 31). As shown in Fig. 4, treatment of the alkaline-hydro- lyzed product with nitrous acid resulted in its complete deg- radation to fragments that were eluted as a sharp peak just before the column included volume. Nitrous acid hydrolysis is specific for glycosaminoglycans that contain N-SO, moie- ties, i.e. either heparan sulfate or heparin (5). Based on the results of these experiments, we conclude that the cytoskel- eton-associated 35S04-labeled molecule is a heparan sulfate proteoglycan.
Heparan Sulfate Proteoglycan Is Not a Nuclear Protein- Light microscopic observation of gradient purified cytoskel- etons revealed the presence of nuclei (Fig. 5, panel a ) . An association of nuclei with cytoskeletal structures, especially intermediate filaments, is well documented (32). It has been reported that heparan sulfate proteoglycans are present in the nuclei of some cells (33). To investigate the possibility that the heparan sulfate proteoglycan described above was a nu- clear protein, we purified nuclei from rat livers and monitored the distribution of proteoglycan by 35s04 radioactivity. The purified nuclei appeared as mostly intact organelles by light microscopy (Fig. 5, panel b) and lacked the associated amor- phous fibrillar aggregates that characterized the purified cy- toskeletal fraction (Fig. 5, panel a) . The absence of cytoskel- eta1 proteins in the purified nuclei was confirmed by SDS-gel electrophoresis (Fig. 6).
The purified nuclear fraction contained only 0.7% (2.7 X lo4 cpm) of the total incorporated 35s04 that was present in the liver homogenate (4 x lo6 cpm); the remaining radioactiv- ity could be accounted for in the supernatant fraction result- ing from the nuclear isolation. When the nuclei were solubi- lized in guanidine buffer and subjected to gel filtration chro- matography on a Sepharose CL-4B column, no discernible peaks were detected (not shown). When the cell supernatant
FIG. 5. Light micrographs of isolated cytoskeletons and nu- clei. Suspensions of cytoskeletons, purified by sucrose gradient cen- trifugation (panel a), or nuclei, purified by the method of Blobel and Potter (26) (panel b), were placed onto microscope slides, cover- slipped, and photographed with phase-contrast optics. Magnification: 11ox.
FIG.
1 2
4
4
4
4
4
-= "
4
3379
._actions It-uurified r
6. SDS-gel electrophoresis of cytoskelets and Durified nuclei. Aliauots of nuclei (lane I ) or mad
11 fr: iel
cytosieletons (lane 2) were dissolved in electrophorek sampie buffer and electrophoresed in 7.5% polyacrylamide slab gels. The Coomassie Blue-stained gel is shown. Arrowheads to the right indicate the position of migration of protein molecular weight standards. These were: myosin (M, = 200,000), j3-galactosidase (M, = 115,000), phos- phorylase b (M, = 94,000), bovine serum albumin (M, = 68,000), ovalbumin (M, = 43,000), and carbonic anhydrase (M, = 29,000).
fraction that was obtained during the nuclear isolation was subjected to gel filtration chromatography, radiolabeled peaks were eluted at the column void volume and at K,, values = 0.45 and 0.7 (not shown). From these results we conclude that the heparan sulfate proteoglycan is not a nuclear protein.
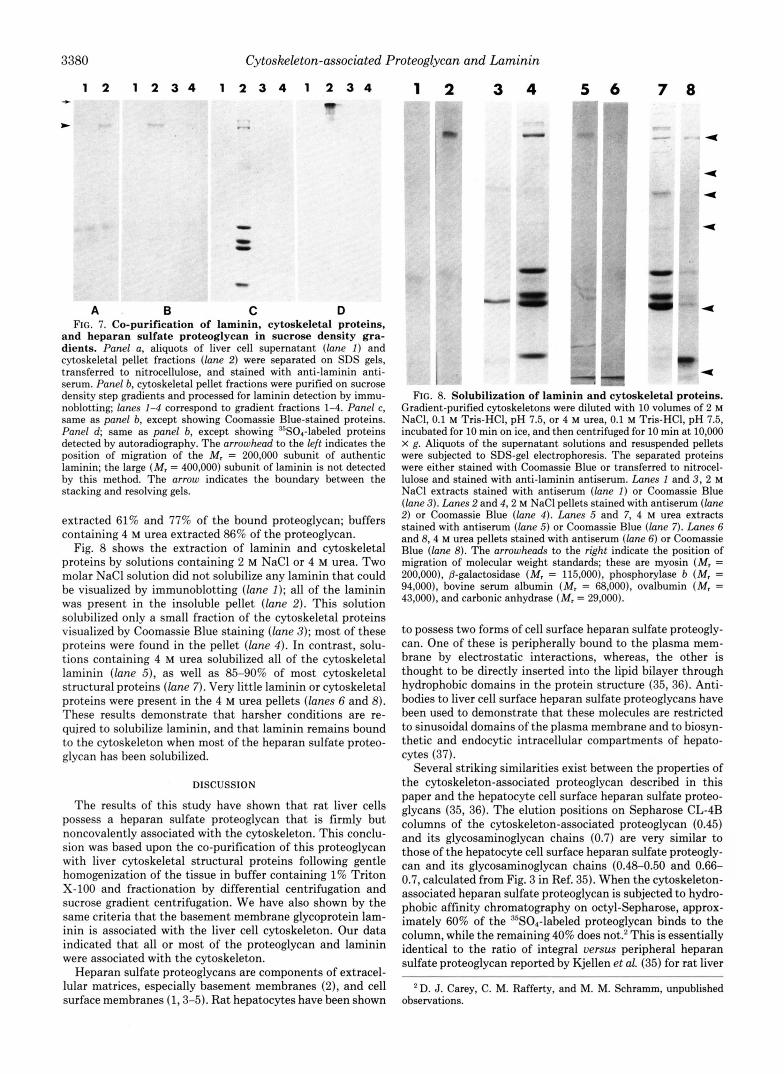
Laminin Is Associated with the Purified Liver Cytoskel- eton-Heparan sulfate proteoglycans are usually localized to surface membranes of cells and to the extracellular matrix (1-5). In order to determine whether other extracellular ma- trix proteins might be associated with the liver cytoskeleton, we examined the distribution of laminin, a large extracellular matrix glycoprotein (34) that is often found in association with heparan sulfate proteoglycans. Laminin was identified by staining electrophoretically resolved proteins transferred to nitrocellose with anti-laminin antiserum. As shown in Fig. 7, laminin was present in the washed cytoskeletal pellet (panel A , lane 2) but was not detectable in the cell supernatant fraction (panel A , lane I). When the cytoskeletons were purified on sucrose gradients, essentially all of the laminin was present in fraction 2 (panel B, lane 2), and thus cosedi- mented with the cytoskeletal proteins visualized by Coomassie Blue staining (panel C, lane 2), and the heparan sulfate proteoglycan, visualized by autoradiography (panel D, lane 2). Thus, a significant portion of the laminin in the tissue is firmly associated with the liver cytoskeleton.
Extraction of Heparan Sulfate Proteoglycan and Laminin from Purified Cytoskeletons-To investigate the nature of the association between the heparan sulfate proteoglycan, lami- nin, and the cytoskeleton, we tested various solutions for their ability to extract cytoskeletal proteins. The heparan sulfate proteoglycan was not extracted from purified cytoskeletons by buffers of physiological ionic strength (0.15 M NaCl, 0.05 M Tris) but was extracted by buffers containing high salt concentrations or urea. Buffers containing 1 and 1.7 M NaCl
3380 Cytoskeleton-associted Proteoglycan and Laminin
1 2 1 2 3 4 1 2 3 4 1 2 3 4 *
A B C D FIG. 7. Co-purification of laminin, cytoskeletal proteins,
and heparan sulfate proteoglycan in sucrose density gra- dients. Panel a, aliquots of liver cell supernatant ( l a n e I) and cytoskeletal pellet fractions (lane 2) were separated on SDS gels, transferred to nitrocellulose, and stained with anti-laminin anti- serum. Panel b, cytoskeletal pellet fractions were purified on sucrose density step gradients and processed for laminin detection by immu- noblotting; lanes 1-4 correspond to gradient fractions 1-4. Panel c, same as panel b, except showing Coomassie Blue-stained proteins. Panel d; same as panel b, except showing DsS04-labeled proteins detected by autoradiography. The arrowhead to the left indicates the position of migration of the M, = 200,000 subunit of authentic laminin; the large (M, = 400,000) subunit of laminin is not detected by this method. The arrow indicates the boundary between the stacking and resolving gels.
extracted 61% and 77% of the bound proteoglycan; buffers containing 4 M urea extracted 86% of the proteoglycan.
Fig. 8 shows the extraction of laminin and cytoskeletal proteins by solutions containing 2 M NaCl or 4 M urea. Two molar NaCl solution did not solubilize any laminin that could be visualized by immunoblotting (lane I); all of the laminin was present in the insoluble pellet (lane 2). This solution solubilized only a small fraction of the cytoskeletal proteins visualized by Coomassie Blue staining (lane 3); most of these proteins were found in the pellet (lane 4) . In contrast, solu- tions containing 4 M urea solubilized all of the cytoskeletal laminin (lane 5 ) , as well as 85-90% of most cytoskeletal structural proteins (lane 7). Very little laminin or cytoskeletal proteins were present in the 4 M urea pellets (lanes 6 and 8). These results demonstrate that harsher conditions are re- quired to solubilize laminin, and that laminin remains bound to the cytoskeleton when most of the heparan sulfate proteo- glycan has been solubilized.
DISCUSSION
The results of this study have shown that rat liver cells possess a heparan sulfate proteoglycan that is firmly but noncovalently associated with the cytoskeleton. This conclu- sion was based upon the co-purification of this proteoglycan with liver cytoskeletal structural proteins following gentle homogenization of the tissue in buffer containing 1% Triton X-100 and fractionation by differential centrifugation and sucrose gradient centrifugation. We have also shown by the same criteria that the basement membrane glycoprotein lam- inin is associated with the liver cell cytoskeleton. Our data indicated that all or most of the proteoglycan and laminin were associated with the cytoskeleton.
Heparan sulfate proteoglycans are components of extracel- lular matrices, especially basement membranes (2), and cell surface membranes (1,3-5). Rat hepatocytes have been shown
1 2 3 4 5 6 7 8 9" -" am
4
4
4
4
<
FIG. 8. Solubilization of laminin and cytoskeletal proteins. Gradient-purified cytoskeletons were diluted with 10 volumes of 2 M NaCI, 0.1 M Tris-HCI, pH 7.5, or 4 M urea, 0.1 M Tris-HCI, pH 7.5, incubated for 10 min on ice, and then centrifuged for 10 min at 10,OOO X g. Aliquots of the supernatant solutions and resuspended pellets were subjected to SDS-gel electrophoresis. The separated proteins were either stained with Coomassie Blue or transferred to nitrocel- lulose and stained with anti-laminin antiserum. Lanes I and 3, 2 M NaCl extracts stained with antiserum (lone I) or Coomassie Blue (lane 3) . Lanes 2 and 4 ,2 M NaCl pellets stained with antiserum (lane 2) or Coomassie Blue (lane 4). Lanes 5 and 7, 4 M urea extracts stained with antiserum (lane 5) or Coomassie Blue (lane 7). Lanes 6 and 8 , 4 M urea pellets stained with antiserum (lane 6 ) or Coomassie Blue (lane 8). The arrowheads to the right indicate the position of migration of molecular weight standards; these are myosin (M, = 200,000), 8-galactosidase (M, = 115,000), phosphorylase b (M, = 94,000), bovine serum albumin (M, = 68,000), ovalbumin (M, = 43,000), and carbonic anhydrase (M. = 29,000).
to possess two forms of cell surface heparan sulfate proteogly- can. One of these is peripherally bound to the plasma mem- brane by electrostatic interactions, whereas, the other is thought to be directly inserted into the lipid bilayer through hydrophobic domains in the protein structure (35,36). Anti- bodies to liver cell surface heparan sulfate proteoglycans have been used to demonstrate that these molecules are restricted to sinusoidal domains of the plasma membrane and to biosyn- thetic and endocytic intracellular compartments of hepato- cytes (37).
Several striking similarities exist between the properties of the cytoskeleton-associated proteoglycan described in this paper and the hepatocyte cell surface heparan sulfate proteo- glycans (35, 36). The elution positions on Sepharose CL-4B columns of the cytoskeleton-associated proteoglycan (0.45) and its glycosaminoglycan chains (0.7) are very similar to those of the hepatocyte cell surface heparan sulfate proteogly- can and its glycosaminoglycan chains (0.48-0.50 and 0.66- 0.7, calculated from Fig. 3 in Ref. 35). When the cytoskeleton- associated heparan sulfate proteoglycan is subjected to hydro- phobic affinity chromatography on octyl-Sepharose, approx- imately 60% of the RsSOI-labeled proteoglycan binds to the column, while the remaining 40% does not.2 This is essentially identical to the ratio of integral uersus peripheral heparan sulfate proteoglycan reported by Kjellen et al. (35) for rat liver
*D. J. Carey, C. M. Rafferty, and M. M. Schramm, unpublished observations.

Cytoskeleton-associated Proteoglycan and Laminin 3381
plasma membranes. These similarities, together with the fact that no other similar sized 35S0,-labeled macromolecules were detected in our liver fractions, suggest that the cytoskeleton- associated proteoglycan described in this paper is identical to the hepatocyte cell surface heparan sulfate proteoglycan (35, 36).
The mechanism of binding of heparan sulfate proteoglycan and laminin to the liver cytoskeleton is not known. There is considerable evidence to suggest that cells interact with base- ment membranes via specific binding interactions with lami- nin (38, 39). Presumably, the binding of laminin to the he- patocyte cytoskeleton is mediated by a cell surface laminin receptor. A laminin-binding protein of M , 70,000 has been isolated from several cell types (40-42). Interestingly, the laminin-binding protein isolated from fibrosarcoma cells in culture can bind to actin filaments in vitro (43), suggesting a plausible explanation for the findings reported here. On the other hand, our results are also consistent with the possibility that laminin or the laminin receptor could be complexed to another cell surface molecule that is bound to intracellular proteins. Conceivably, this could be accomplished by binding between laminin and the heparan sulfate chains of the cell surface proteoglycan. However, our data on the differential extraction of these molecules from isolated cytoskeletons, which showed that laminin remained bound under conditions in which most of the proteoglycan was solubilized, argue against this possibility.
Evidence suggesting a functional interaction between the cytoskeleton and basement membrane components, especially laminin, have also been reported. Addition of soluble laminin to isolated corneal epithelia cause a smoothing of the basal surface and the reorganization of a network of actin microfi- laments located near the cell surface (19). Recent evidence suggests that this effect of laminin is mediated by specific cell surface receptors (44). In addition, it has been reported that cell surface laminin on cultured fibrosarcoma cells is resistant to detergent extraction following cross-linking by antibodies or excess exogenous laminin (45). This was interpreted as suggesting an association of laminin receptors with the cyto- skeleton.
There are obvious functional consequences that would re- sult from a direct association of extracellular matrix and intracellular cytoskeletal proteins. The anchoring of the main structural elements of the cytoplasm to the extracellular con- nective tissue matrix would provide structural stability and support to the cell. Furthermore, this association helps to explain some of the known effects of extracellular matrix contact on cell behavior, such as alterations in cell morphol- ogy and cell migration. The ability to isolate complexes of cytoskeletal and extracellular matrix proteins from tissue in amounts sufficient to permit biochemical characterization will be an essential step towards understanding these processes at the molecular level.
1.
2.
3.
4.
5.
REFERENCES
Oldberg, A., Kjellen, L., and Hook, M. (1979) J. Bid. Chem. 254,
Hassell, J. R., Robey, P. G., Barrach, H. J., Wilszek, J., Rennard, S. I., and Martin, G. R. (1980) Proc. Natl. Acad. Sci. U. S. A.
Rapraeger, A. C., and Bernfield, M. (1983) J. Bid. Chem. 258,
Hook, M., Kjellen, L., Johansson, S., and Robinson, J. (1984)
Gallagher, J. T., Lyon, M., and Steward, W. P. (1986) Biochem.
8505-8510
77,4494-4498
3632-3636
Annu. Reu. Biochem. 53, 847-869
J. 236, 313-325
6.
7.
8.
9.
10.
11. 12.
13. 14.
15. 16.
17.
18.
19. 20.
21.
22.
23.
24.
25.
26. 27. 28.
29.
30.
31.
32.
33.
34.
35.
36.
37.
38.
39.
40.
41.
42.
43.
44. 45.
Kleinman, H. K., McGarvey, M. L., Hassell, J . R., Star, V. L., Cannon, F. B., Laurie, G. W., and Martin, G. R. (1986) Bio- chemistry 25, 312-318
Laurie, G. W., Bing, J . T., Kleinman, H. K., Hassell, J. R., Aumailley, M., Martin, G. R., and Feldmann, R. J . (1986) J.
Yurchenco, P. D., Tsilbary, E. C., Charonis, A. S., and Furthmayr,
Laterra, J., Silbert, J . E., and Culp, L. A. (1983) J. Cell Biol. 96,
Cole, G. J., Schubert, D., and Glaser, L. (1985) J . Cell Biol. 100,
Cole, G. J., and Glaser, L. (1986) J. Cell Bid. 102, 403-412 Anderson, M. J., Klier, F. G., and Tanguay, K. E. (1984) J . Cell
Singer, I., and Paradisco, P. R. (1981) Cell 24, 481-492 Bissell, M. J., Hall, H. G., and Parry, G. (1982) J. Theor. Bid.
Chen, W. T., and Singer, S. J. (1982) J. Cell Biol. 95, 205-222 Woods, A,, Hook, M., Kjellan, L., Smith, C. G., and Rees, D. A.
Chen, W. T., Hasegawa, E., Hasegawa, T., Weinstock, C., and
Drenckhahn, D., and Wagner, J . (1986) J. Cell B i d . 102, 1738-
Sugrue, S. P., and Hay, E. D. (1981) J. Cell B i d . 91, 45-54 Carey, D. J., Todd, M. S., and Rafferty, C. M. (1986) J . Cell Biol.
102, 2254-2263 Baron-van Evercooren, A,, Kleinman, H. K., Seppa, H. E. J.,
Rentier, B., and Dubois-Dalcq, M. (1982) J. Cell Biol. 93, 211- 216
Lander, A. D., Fujii, D. K., and Reichardt, L. F. (1985) Proc. Natl. Acad. Sci. U. S. A. 82, 2183-2187
Duband, J. L., Rocher, S., Chen, W. T., Yamada, K. M., and Thiery, J. P. (1986) J. Cell Bid. 102, 160-178
Wicha, M. S., Lowrie, G., Kohn, E., Bagavandoss, P., and Mahn T. (1982) Proc. Natl. Acad. Sci. U. S. A. 79, 3213-3217
Carey, D. J., and Todd, M. S. (1986) J . Biol. Chem. 261, 7518- 7525
Blobel, G., and Potter, V. R. (1966) Science 154, 1662-1665 Laemmli, U. K. (1970) Nature 227, 680-685 Johnson, D. A., Gautsch, J. W., Sportsman, J. R., and Elder, J.
Mehta, H., Orphe, C., Todd, M. S., Cornbrooks, C. J., and Carey,
Goodman, S. R., and Zagon, I. S. (1986) Am. J . Physiol. 250,
Toole, B. P. (1976) in Neuronal Recognition (Barondes, S., ed)
Fey. E. G.. Krochmalnic. G.. and Penman. S. (1986) J. Cell Biol.
Mol. B i d . 189, 205-216
H. (1986) J. Histochem. Cytochem. 34, 93-102
112-123
1192-1199
Bid. 99, 1769-1784
99,31-68
(1984) J. Cell Bid. 99, 1743-1753
Yamada, K. M. (1985) J. Cell Biol. 100, 1103-1114
1747
H. (1984) Gene Anal. Technol. 1,3-8
D. J. (1985) J. Cell Bid. 101, 660-666
C347-C360
pp. 275-329, Plenum Press, New York
i02,1654-1665 . , , . ,
Fedarko. N. S.. and Conrad. H. E. (1986) J. Cell B id . 102. 587- 599
Timpl, R., Rohde, H., Robey, P. G., Rennard, R. I., Foidart, J. M., and Martin, G. R. (1979) J. Bid. Chem. 254, 9933-9937
Kjellen, L., Oldberg, A., and Hook, M. (1980) J. Bid. Chem. 255,
Kjellen, L., Pettersson, I., and Hook, M. (1981) Proc. Natl. Acad.
Stow, J. L., Kjellen, L., Unger, E., Hook, M., and Farquhar, M.
Tarranova, V. P., Rohrbach, D. H., and Martin, G. X. (1980) Cell
Johansson, S., Kjellen, L., Hook, M., and Timpl, R. (1981) J. Cell
Lesot, H., Kuhl, U., and von der Mark, K. (1983) E M B O J. 2,
Malinoff, H. L., and Wicha, M. S. (1983) J. Cell B i d . 96, 1475-
Rao, N. C., Barsky, S. H., Terranova, V. P., and Liotta, L. A.
Brown, S. S., Malinoff, H. L., and Wicha, M. S. (1983) Proc.
Sugrue, S. P., and Hay, E. D. (1986) J. Cell Biol. 102, 1907-1916 Cody, R. L., and Wicha, M. S. (1986) Exp. Cell Res. 165, 107-
. ,
10407-10413
Sci. U. S. A. 78,5371-5375
G. (1985) J. Cell Biol. 100, 975-980
22,719-726
Bid. 90, 260-264
861-865
1479
(1983) Biochem. Biophys. Res. Commun. 11 1, 804-808
Natl. Acad. Sci. U. S. A. 80, 5927-5930
116





























