Embed Size (px)
Citation preview

Proc. Natl. Acad. Sci. USAVol. 93, pp. 9858-9863, September 1996Neurobiology
Olfactory marker protein (OMP) gene deletion causes alteredphysiological activity of olfactory sensory neurons
(olfactory bulb/tyrosine hydroxylase/cholecystokinin/electroolfactogram)
0. 1. BUIAKOVA*t, H. BAKERt, J. W. Scorr§, A. FARBMANI, R. KREAMII, M. GRILLO*, L. FRANZENt,M. RICHMANI, L. M. DAVIS§, S. ABBONDANZO*, C. L. STEWART*, AND F. L. MARGOLIS*"***Roche Institute of Molecular Biology, Roche Research Center, Nutley, NJ 07110; tBurke Rehabilitation Center, Cornell University Medical College, WhitePlains, NY 10605; §Department of Anatomy and Cell Biology, Emory University, Atlanta, GA 30322; IDepartment of Neurobiology and Physiology,Northwestem University, Evanston, IL 60208; and IlTufts New England Medical Center, Boston, MA 02111
Communicated by Sidney Udenfriend, Roche Institute of Molecular Biology, Nutley, NJ, June 11, 1996 (received for review January 10, 1996)
ABSTRACT Olfactory marker protein (OMP) is anabundant, phylogenetically conserved, cytoplasmic protein ofunknown function expressed almost exclusively in matureolfactory sensory neurons. To address its function, we gener-ated OMP-deficient mice by gene targeting in embryonic stemcells. We report that these OMP-null mice are compromisedin their ability to respond to odor stimuli, providing insight toOMP function. The maximal electroolfactogram response ofthe olfactory neuroepithelium to several odorants was 20-40%smaller in the mutants compared with controls. In addition,the onset and recovery kinetics following isoamyl acetatestimulation are prolonged in the null mice. Furthermore, theability of the mutants to respond to the second odor pulse ofa pair is impaired, over a range of concentrations, comparedwith controls. These results imply that neural activity directedtoward the olfactory bulb is also reduced. The bulbar pheno-type observed in the OMP-null mouse is consistent with thishypothesis. Bulbar activity of tyrosine hydroxylase, the ratelimiting enzyme of catecholamine biosynthesis, and content ofthe neuropeptide cholecystokinin are reduced by 65% and50%, respectively. This similarity to postsynaptic changes ingene expression induced by peripheral olfactory deafferenta-tion or naris blockade confirms that functional neural activityis reduced in both the olfactory neuroepithelium and theolfactory nerve projection to the bulb in the OMP-null mouse.These observations provide strong support for the conclusionthat OMP is a novel modulatory component of the odordetection/signal transduction cascade.
Odor detection occurs in olfactory cilia where signal trans-duction is believed to be initiated by a stimulus moleculeinteracting with a seven-transmembrane domain G protein-coupled receptor. Subsequent activation of G protein-mediated second messenger systems leads to cation channelopening and action potential generation (1). Many proteins ofthe odor detection/signal tranduction cascade are either abun-dantly or selectively expressed in olfactory sensory neurons(1-4). Although these proteins are structurally homologous toproteins involved in signal transduction in other cell types,attempts to demonstrate functional activity of olfactory re-ceptor molecules in heterologous systems have been largelyunsuccessful (see, however, ref. 5). The basis for this lack ofsuccess is unknown but may relate to the difficulty in choosingan optimal stimulus/receptor pair. This is unlikely, as individ-ual olfactory sensory cells, expressing one or few receptortypes (1, 6, 7), respond to a wide range of stimuli (1, 8, 9). Otherpossibilities include the absence in heterologous systems ofessential olfactory neuron-specific components directly orindirectly involved in odor detection/signal transduction pro-
cesses. These latter components might include proteins selec-tively expressed in mature olfactory sensory neurons and theircilia. The olfactory marker protein (OMP) is a possiblecandidate because this 19-kDa phylogenetically conserved,cytoplasmic protein is expressed almost exclusively by matureolfactory neurons in essentially all vertebrate species (10, 11).OMP is present throughout the cytoplasm of olfactory sensoryneurons and is abundant in the olfactory ciliary lumen (10, 12).Here we report that olfactory sensory neurons of OMP-nullmice, from which the OMP gene has been deleted by homol-ogous recombination in embryonic stem (ES) cells, havereduced electrophysiological activity. Furthermore, this reduc-tion in neural activity of olfactory neurons influences down-stream gene expression in the olfactory bulbs.
MATERIALS AND METHODSTargeting Vector. The mouse OMP targeting construct
contained 9 kb of 5' and 1.5 kb of 3' homology, along withPGK-neomycin resistance (PGK-neo) and herpes simplexvirus-thymidine kinase (HSV-TK) cassettes in pBluescriptKS(+) (Stratagene) as illustrated (see Fig. 1).
Generation of Chimeric Mice. ES cell line W9.5 was grownon a feeder layer of mitomycin C-inactivated primary mouseembryonic fibroblasts in Dulbecco's modified Eagle's mediumsupplemented with 20% (vol/vol) fetal bovine serum and 0.1mM 2-mercaptoethanol. The cells were electroporated with 25,ug of linearized vector and clones selected using positive(PGK-neo) and negative (HSV-TK) selection with G418 and1-(2'-deoxy-2'-fluoro-1-beta-D-arabinofuranosyl)-5-iodou-racil (FIAU), respectively, as described (13). EcoRI diagnosticrestriction enzyme digests ofDNA from surviving clones werescreened for homologous recombination by Southern analysisusing the 3' single copy probe (see Fig. la). The overalltargeting frequency was 10% of clones surviving double se-lection. Following verification of homologous recombination,cells from two clones (see Fig. lb) were used to generatechimeric mice (13). Germ-line transmission of the targetedallele was monitored by Southern analysis as above and byPCR. Tail tip DNA preparations and PCR were performed asdescribed (14). The wild-type OMP allele was detected usinga 5' primer homologous to DNA sequence in the coding regionof the OMP gene (5'-AAGCTGCAGTTCGATCACTGGA-3') and a 3' primer (5'-TGTTCCTGTCCAGTCTCAGTCT-
Abbreviations: OMP, olfactory marker protein; CCK, cholecystokinin;ES, embryonic stem cell; EOG, electroolfactogram.tPresent address: Departments of Psychiatry and Genetics, ColumbiaUniversity, New York Psychiatric Institute Unit 58, 722 West 168Street, New York, NY 10032.
* *To whom reprint requests should be sent at the present address:Department of Anatomy, University of Maryland School of Med-icine, 685 West Baltimore Street, Baltimore, MD 21201. e-mail:[email protected].
9858
The publication costs of this article were defrayed in part by page chargepayment. This article must therefore be hereby marked "advertisement" inaccordance with 18 U.S.C. §1734 solely to indicate this fact.

Proc. Natl. Acad. Sci. USA 93 (1996) 9859
3') homologous to the 3' untranslated portion of the OMPgene transcript that lies within the 3' homology region of thetargeting construct. The targeted OMP allele was identified byusing a 5' primer homologous to the promoter region of theneo gene (5'-TACCGGTGGATGTGGAATGTGT-3') andthe same 3' primer as for the wild-type OMP allele above. A700-bp amplicon was generated in the presence of the wild-type allele and a 390-bp amplicon in the presence of thetargeted allele.Western Blot Analysis. Mice were sacrificed by CO2 and
exsanguination, and olfactory mucosa and bulbs were dis-sected, frozen, weighed, and homogenized in SDS buffer (1%SDS/10 mM Tris HCl, pH 7.4/100 AM NaF/10 mM sodiumpyrophosphate/10 ,g of aprotinin per ml/5 mg of leupeptinper ml/0.2 mM phenylmethylsulfonyl fluoride). Aliquots wereelectrophoresed in 15% polyacrylamide gels, transferred to0.45 ,um nitrocellulose, blocked with 5% nonfat milk and 0.5%BSA in PBST (137 mM NaCl/2.5 mM KCl/10 mM Na2HPO4/1.75 mM KH2PO4, pH 7.2/0.25% Tween 20), and sequentiallyincubated with 1/250 rabbit-anti-OMP, 1/1000 goat anti-rabbit-IgG phosphatase conjugated; the color was developedwith 100 mg of nitroblue tetrazolium per ml and 50 mg/ml ofX-phosphate (Boehringer Mannheim).Immunocytochemistry. Olfactory epithelia from mice at 2
days and 2 months postnatal were fixed in Zamboni's fixative,cryoprotected in 30% sucrose and frozen in OCT (Tissue Tech,Miles). The tissue blocks were sectioned at 8 to 10 ,um at-20°C. The sections were washed in PBS containing 0.2%glycine, incubated with goat polyclonal anti-OMP antibodies(1:1000) or rabbit polyclonal anti-carnosine antibodies (1:500)overnight at room temperature, and washed in PBS. Anti-OMP-treated sections were incubated with rhodamine-conjugated donkey anti-goat IgG antibodies (1:80) (Chemi-con); anti-camosine treated sections were incubated withfluorescein isothiocyanate-conjugated swine anti-rabbit IgGantibodies (1:40) (Dako). In both cases incubations wereperformed for 1 hr at room temperature in the dark withsubsequent washes in PBS.
Histology. Olfactory epithelia from 3-month-old mice werefixed in Bouin's fixative and decalcified with RDO (ApexEngineering, Plainfield, IL). The samples were dehydrated,embedded in paraffin, and sectioned at 6-8 Am. Sections weredeparaffined, rehydrated, and stained with hematoxylin/eosin.To count dendritic knobs, olfactory tissue from 5-week-oldmice (two wild type, three heterozygotes, and three OMP-nullmice) was fixed in 0.1 M sodium phosphate (pH 7.2) containing1.6% formaldehyde and 2% glutaraldehyde at 4°C. The tissuewas subsequently processed and embedded in an epon-aralditeplastic mixture. Olfactory knobs were counted either from sixnonadjacent sections in each of two areas per mouse for a totalolfactory epithelial length of --20 microns (electron micro-scope) or in three to four nonadjacent sections from each offour areas per mouse along 200-600 Am of olfactory epitheliallength (light microscope). In total, --400-500 knobs werecounted per mouse in light and electron microscopy, repre-senting 1.5-2.0 mm of epithelial length.
Biochemistry. Mice, at 1 and 2 months postnatal, weresacrificed by CO2 inhalation followed by exsanguination; theolfactory bulbs were dissected individually, frozen on dry ice,coded, and weighed. Single olfactory bulbs from each pair werehomogenized in 5 mM potassium phosphate buffer (pH 7.0)containing 0.2% Triton X-100; tyrosine hydroxylase activityand protein content were determined as described (15). In theassay '4C-dopa generated from 14C-tyrosine substrate is sep-arated on alumina, quantified by scintillation spectrometry,and reported as nmol dopa/bulb/15 min. For quantification oflevels of cholecystokinin (CCK)-like immunoreactivity (CCK-LI) by RIA, the second member of each pair of frozenolfactory bulbs, stored at -80°C, was extracted using proce-dures previously described in detail (16). Briefly, bulbs were
each homogenized in 2 M acetic acid followed by the additionof 20 volumes of cold 50% acetonitrile/0.1% trifluoroaceticacid. Following centrifugation (10,000 x g for 30 minutes), thepeptide-containing supernatants were subsequently delipi-dated and concentrated. The acidified upper aqueous phasewas dried and reconstituted in RIA buffer. Overall, this simpleextraction procedure yields recoveries of exogenously addedCCK >90%.The RIA procedure for quantification of CCK-LI uses a
highly selective antiserum generated against synthetic sulfatedCCK-8 coupled to keyhole limpet hemocyanin using previ-ously described conjugation procedures (16). The antibodyrecognizes sulfated CCK-8 and sulfated gastrin 17 on anequimolar basis, with nonsulfated CCK-8 and gastrin 17displaying <10% cross-reactivity. Other methionine-con-taining neuropeptides such as substance P, methionine en-kephalin, and f3-endorphin display <0.1% cross-reactivity inthe RIA. The RIA tracer is a radioiodinated CCK-BoltonHunter reagent conjugate, prepared by previously describedprocedures and purified by HPLC. In the RIA, partiallypurified CCK antiserum is used at a final dilution of 1:20,000at which 35% of the added radiolabeled peptide is specificallybound. Initial incubation with the antibody is for 24 hr,followed by addition of 20,000 cpm of 125I-labeled CCK tracerand an additional 24-hr incubation. Finally, precipitation ofantibody-bound radioactive tracer is achieved by addition of 1ml of cold absolute ethanol. After centrifugation, supernatantsare aspirated and the pellets counted in a y counter at >70%efficiency. Nonspecific binding of radiolabel is typically 2%and is subtracted from each tube. For unknown samples,peptide concentrations are quantified by interpolation from alogit-log plot of the standard dose-response displacementcurve which utilizes synthetic sulfated CCK-8 as the RIAstandard. In these analyses, the lowest detectable dose is -1fmol peptide/tube and 50% displacement is achieved at '12fmol peptide/tube.
Electrophysiology. Male and female mice (20-35 g, 10-14weeks old) were anesthetized with nembutal (70 mg/kg) orsacrificed by CO2 inhalation. A cannula was placed in thetrachea to allow stimuli to be aspirated through the nose at 50ml/min. The nonluminal side (lamina propria) of the olfactoryepithelium was exposed by drilling through the bone with adental burr. Two exposures were made in each animal, one inthe dorsal recess of the epithelium and one in the lateral recessbetween the base of the turbinate bones. Recordings weremade with micropipettes (resistance 5-10 mfl) filled withRinger's solution and driven through the epithelium until alarge negative response was evoked by odor stimulation. Cleanair flowed into a chamber in front of the nose at 500 ml/min.Air saturated with odors was injected into the chamber by acomputer controlled syringe pump. The rate of flow producedby the syringe pump determines the odor concentration (ex-pressed as percent of vapor saturation). All odors were rou-tinely tested at 2, 7, and 18% of saturation. Each odorpresentation was accomplished by infusion of compound intothe stimulus chamber 10 sec before the beginning of anartificial sniff produced by opening a valve in the vacuum lineconnected to the tracheal cannula. This preparation is similarto that previously described (17). One-minute intervalselapsed between odor presentations. Isoamyl acetate wasalways the first odor tested, and these data were used forcomparisons of kinetics. The same results were observed witheither anesthetized or C02-killed mice, but only data from thelatter were used for the analysis of responses to single pulses(see Fig. 3 a and c). These data were evaluated by analysis ofvariance across OMP status, electrode position, and stimulusconcentration. Recordings of responses to dual odor pulseswere made primarily from the dorsal recess. Results from someanesthetized mice were included in this analysis. Analysis of
Neurobiology: Buiakova et aL

9860 Neurobiology: Buiakova et al.
variance showed that the difference between anesthetized andkilled mice was not a significant contribution to the variance.
RESULTS AND DISCUSSIONTo address the function of OMP, we used gene targeting inmouse ES cells to generate a null allele by replacing theintronless coding region of the OMP gene (11) with a selectioncassette containing the neomycin phosphotransferase (neo)gene (Fig. 1 a and b). The absence of OMP expression in micehomozygous for the targeted allele was confirmed immuno-chemically (Figs. lc and 2 c and e). Central nervous systemareas of ectopic OMP expression (18) were also devoid ofOMP (not shown). Quantification of OMP levels by ELISArevealed that heterozygous mutant mice had 40-50% of the
aMx x
NEO TK pBluescript
v - aD
wr
MUT
b
c
OMP
M2 F: M2 - 1; PROBE
NEO
$ + +
+ +
19 lkb
(1)
+ +
+iI9 kD) - i
(1)
19 kb$-X
9.5kb
(2)
+ +
19)kDa -
FIG. 1. Gene targeting at the mouse OMP locus. (a) Schematicrepresentation of the OMP targeting vector (Top), the OMP locus(Middle), and the expected recombination events (Bottom). Successfulgene replacement by the targeting vector results in the deletion of theentire coding region of the OMP gene and insertion of a PGK-neoselection cassette in the opposite orientation. The location of the DNAprobe used to identify putative targeted ES cell clones and theirprogeny is indicated. (b) Southern blot analysis of gene targeting at theOMP locus and transmission of the targeted allele. (1) A 19-kb EcoRIfragment, indicative of a wild-type allele, is detected in the parental EScell line (+/ +). An additional fragment of 9.5 kb corresponding to thetargeted allele, is detected in DNA from two clones (-/+). Each ofthese clones was used to generate an independent OMP-null line. Mostof the data presented here derive from analysis of one of these twolines. (2) The three lanes represent DNA samples from tail tips of3-week-old heterozygous (-/+), homozygous mutant (-/-), andwild-type (+/+) progeny derived from intercrosses between miceheterozygous for the targeted allele. (c) Western immunoblot analysisof extracts from olfactory epithelium (1) and olfactory bulbs (2) ofmice that are genotypically wild-type (+/+), or homozygous (-/-)for the targeted OMP locus. Goat anti-OMP antiserum identifies the19-kDa OMP in wild type but not in homozygous OMP-null mice. Inwild-type olfactory bulbs OMP is present only in axon terminals ofolfactory sensory neurons but not in any bulbar cells.
OMP level of wild-type littermates indicating that each alleleis autonomous. Immunoblots probed with antiserum directedagainst Golf, a component of the olfactory transduction cas-cade, indicated that there was no difference in Golf expressionbetween the wild-type and OMP-null mice. This observationsuggests that OMP gene deletion was not accompanied byglobal alterations in gene expression.
Before analysis of the OMP-null mice for potential func-tional deficits, it was essential to evaluate the morphologicalorganization of the olfactory neuroepithelium in the OMP-nullmice to determine if there were any gross anatomical abnor-malities that could compromise interpretation of the func-tional studies. To characterize the morphological organizationof the olfactory epithelium in the OMP-null mice, histological,immunocytochemical, and electron microscopy analyses wereperformed. Hematoxylin/eosin-stained histological sectionsof olfactory mucosa from mutant and wild-type siblings wereindistinguishable (Fig. 2 a and b). The ratio of immature tomature sensory neurons seemed to be unaffected. Immuno-staining with anticarnosine antibodies (Fig. 2 d and f), thatstain mature olfactory sensory neurons (19), or with anti-GAP43 antibodies (not shown), that identify immature neu-rons (20), was similar in OMP-null and wild-type mice despitethe absence of any OMP in the OMP-null mice (Fig. 2).Quantification of the olfactory epithelial dendritic knob den-sity (by electron microscopy and light microscopy of plasticsections) was the same for OMP-null and wild-type controls(31 ± 5.5/200 microns and 33 ± 3.6/200 microns, respective-ly). A dendritic knob decorated by long cilia at the surface ofthe epithelium and the formation of synaptic contact withtarget neurons in the bulb are key morphological character-istics of a mature olfactory sensory neuron (1). Our electronmicroscopy, histological, and immunocytochemical observa-tions demonstrate that the olfactory neuroepithelium of theOMP-null mice appears morphologically normal.We recorded electroolfactograms (EOGs) from the olfac-
tory epithelium of wild-type and OMP-null mice to evaluatetheir responses to odors (Fig. 3a). For the initial tests withisoamyl acetate, the response magnitudes (Fig. 3b) were 25%lower in the OMP-null mice. Responses to all other odorchemicals tested (benzaldehyde, menthone, benzene, li-monene, and cyclooctane) were also 20-40% smaller in theOMP-null mice (data not shown). These decreases werestatistically significant (P < 0.02) for all except limonene andcyclooctane. We observed large effects of the mutation onresponse kinetics. The slopes of the initial and decay phases ofthe response to isoamyl acetate (Fig. 3b) were 53% and 43%smaller in the mutant mice, indicating that both responsegeneration and recovery processes are compromised in theolfactory epithelium of OMP-nulls (Fig. 3 a and b). The initialresponse slopes were significantly slower (P < 0.05) for allodors tested. To further test the impairment in responserecovery, we analyzed the responses to two successive pulses ofisoamyl acetate separated by a 6-sec interval. Recovery of theresponse was substantially slower for the OMP-null mice at fivedifferent concentrations (Fig. 3 c and d). This diminishedrecovery probably leads to a sustained reduction in responseto repeated sniffs that is far greater than the 25% smallerresponse that we observed with single pulse stimulation. In theOMP-null mice, the reductions in magnitude of the initialEOG response, the alterations in the kinetics, and the relativemagnitude decrement in response to successive stimulus pulsesall indicate that the olfactory neuroepithelium is severelycompromised in its ability to respond to, and recover from,odor stimulation.
Is this deficit in the ability of olfactory sensory neurons torespond to odor stimulation also reflected in a decreasedability to communicate with their neuronal targets in theolfactory bulb? Mature olfactory sensory neurons synapse onintrinsic bulbar neurons whose morphological and biochemical
Proc. Natl. Acad. Sci. USA 93 (1996)
rz1.
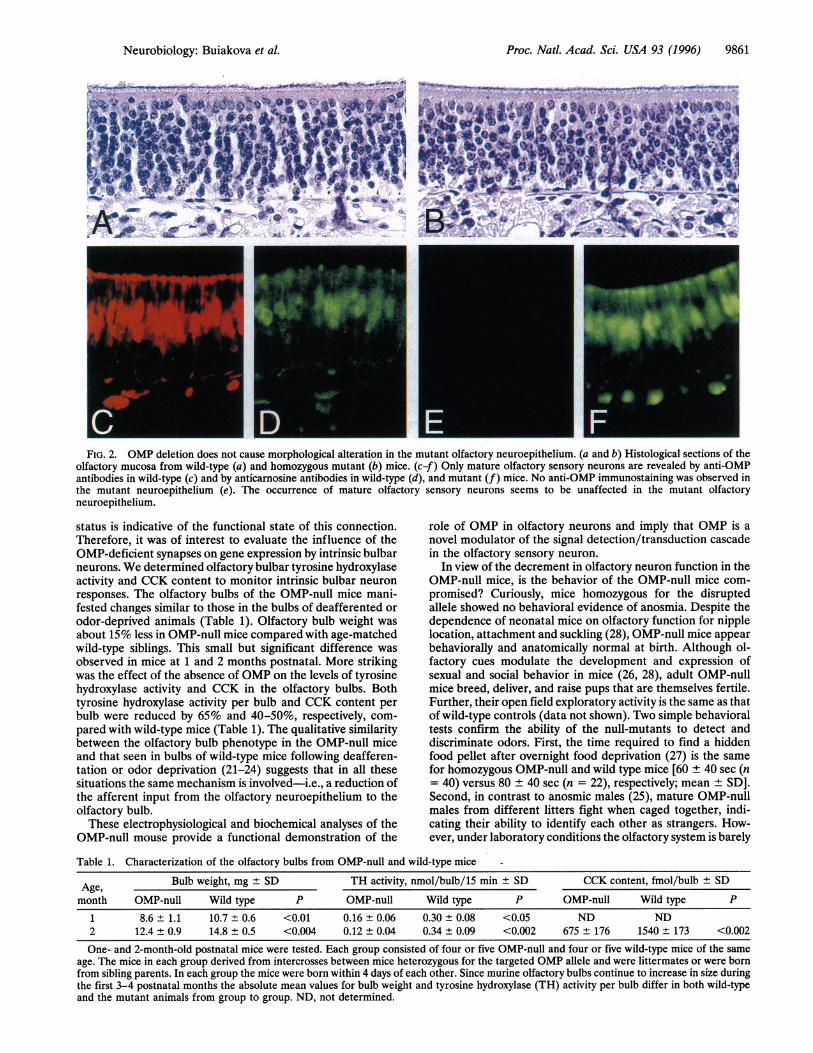
Proc. Natl. Acad. Sci. USA 93 (1996) 9861
FIG. 2. OMP deletion does not cause morphological alteration in the mutant olfactory neuroepithelium. (a and b) Histological sections of theolfactory mucosa from wild-type (a) and homozygous mutant (b) mice. (c-f) Only mature olfactory sensory neurons are revealed by anti-OMPantibodies in wild-type (c) and by anticarnosine antibodies in wild-type (d), and mutant (f) mice. No anti-OMP immunostaining was observed inthe mutant neuroepithelium (e). The occurrence of mature olfactory sensory neurons seems to be unaffected in the mutant olfactoryneuroepithelium.
status is indicative of the functional state of this connection.Therefore, it was of interest to evaluate the influence of theOMP-deficient synapses on gene expression by intrinsic bulbarneurons. We determined olfactory bulbar tyrosine hydroxylaseactivity and CCK content to monitor intrinsic bulbar neuron
responses. The olfactory bulbs of the OMP-null mice mani-fested changes similar to those in the bulbs of deafferented or
odor-deprived animals (Table 1). Olfactory bulb weight wasabout 15% less in OMP-null mice compared with age-matchedwild-type siblings. This small but significant difference wasobserved in mice at 1 and 2 months postnatal. More strikingwas the effect of the absence of OMP on the levels of tyrosinehydroxylase activity and CCK in the olfactory bulbs. Bothtyrosine hydroxylase activity per bulb and CCK content perbulb were reduced by 65% and 40-50%, respectively, com-
pared with wild-type mice (Table 1). The qualitative similaritybetween the olfactory bulb phenotype in the OMP-null miceand that seen in bulbs of wild-type mice following deafferen-tation or odor deprivation (21-24) suggests that in all thesesituations the same mechanism is involved-i.e., a reduction ofthe afferent input from the olfactory neuroepithelium to theolfactory bulb.These electrophysiological and biochemical analyses of the
OMP-null mouse provide a functional demonstration of the
role of OMP in olfactory neurons and imply that OMP is a
novel modulator of the signal detection/transduction cascadein the olfactory sensory neuron.
In view of the decrement in olfactory neuron function in theOMP-null mice, is the behavior of the OMP-null mice com-
promised? Curiously, mice homozygous for the disruptedallele showed no behavioral evidence of anosmia. Despite thedependence of neonatal mice on olfactory function for nipplelocation, attachment and suckling (28), OMP-null mice appearbehaviorally and anatomically normal at birth. Although ol-factory cues modulate the development and expression ofsexual and social behavior in mice (26, 28), adult OMP-nullmice breed, deliver, and raise pups that are themselves fertile.Further, their open field exploratory activity is the same as thatof wild-type controls (data not shown). Two simple behavioraltests confirm the ability of the null-mutants to detect anddiscriminate odors. First, the time required to find a hiddenfood pellet after overnight food deprivation (27) is the samefor homozygous OMP-null and wild type mice [60 ± 40 sec (n= 40) versus 80 ± 40 sec (n = 22), respectively; mean ± SD].Second, in contrast to anosmic males (25), mature OMP-nullmales from different litters fight when caged together, indi-cating their ability to identify each other as strangers. How-ever, under laboratory conditions the olfactory system is barely
Table 1. Characterization of the olfactory bulbs from OMP-null and wild-type mice
Age, Bulb weight, mg ± SD TH activity, nmol/bulb/15 min SD CCK content, fmol/bulb ± SDmonth OMP-null Wild type P OMP-null Wild type P OMP-null Wild type P
1 8.6 ± 1.1 10.7 ± 0.6 <0.01 0.16 ± 0.06 0.30 ± 0.08 <0.05 ND ND2 12.4 ± 0.9 14.8 ± 0.5 <0.004 0.12 ± 0.04 0.34 ± 0.09 <0.002 675 ± 176 1540 ± 173 <0.002
One- and 2-month-old postnatal mice were tested. Each group consisted of four or five OMP-null and four or five wild-type mice of the sameage. The mice in each group derived from intercrosses between mice heterozygous for the targeted OMP allele and were littermates or were bornfrom sibling parents. In each group the mice were born within 4 days of each other. Since murine olfactory bulbs continue to increase in size duringthe first 3-4 postnatal months the absolute mean values for bulb weight and tyrosine hydroxylase (TH) activity per bulb differ in both wild-typeand the mutant animals from group to group. ND, not determined.
Neurobiology: Buiakova et al.
I

9862 Neurobiology: Buiakova et al.
a OMP nuU W.d ty
I mV
1 second markem
b
cOMP nuN;~
"W-Wid tp
I secondmam
d1.5
Wild type OMP nullmeen menn
.4
Peek mV 9.51 7.13 p<0.001 0.9
NormalIed 0.75 0.35 p<0.001 0.inWal slope
NomZedskpe
-0.07 -0.04 p<0.0010.3
0
. Wild eo OMP MAlII
o.001 0.01 0.1 1DiUkion fom satured odor
FIG. 3. EOG responses evoked in OMP-null and wild-type mice by isoamyl acetate. (a) Responses at the dorsomedial recording site to a singlepulse of isoamyl-acetate at 7% of saturation at room temperature (onset and duration indicated by the solid bar). The thicker trace is from anOMP-null and the thin trace is from a wild-type mouse. Positive is upward. Note that both the initial response and return to baseline are morerapid for the wild-type mice. (b) Summary of peak voltages and time courses of responses for all recording sites to single pulses of isoamyl acetate.The normalized slope was calculated by dividing the slope of the steepest portion of the response by the peak voltage. This normalization correctedthe initial and decay slopes for response size. This method of comparison of response kinetics was more accurate than measurement of time topeak because of difficulty in measuring the slow response peaks (see a). (c) Responses of an OMP-null mouse (thicker trace) and a wild-type mouse(thin trace) to two successive pulses of isoamyl acetate separated by a 6-sec interval. Stimulus parameters were as in a. (d) Means and standarderrors for the second response (as in c) as a fraction of the first response. Closed circles, wild-type mice (n = 10), and open circles, OMP-null mice(n = 4), recorded at the dorsomedial position. There was no significant difference in these curves for the lateral recording position (n = 3). Theoverall difference attributable to OMP status was significant at P < 0.01 by analysis of variance.
challenged. For example, rodents with >90% of their olfactoryepithelium or olfactory bulbs destroyed can still detect anddiscriminate odors (25-27). The EOG data suggest an expla-nation for these observations. First, they demonstrate that,although the olfactory sensory neurons of OMP-null mice arefunctionally compromised, they are still responsive. Second,the EOG response decrements that we have observed are torepetitive stimulation with a single pure odor. In contrast, thebehaviors we monitored are in response to complex odormixtures. Alternatively, the OMP-null mice may have adoptedsubtle, novel behavioral strategies (e.g., altered sniff fre-quency) to compensate for their profound deficit to repetitivestimulation. Detailed behavioral evaluation of these mice willbe critical to our understanding in the future.
In conclusion, our biochemical and physiological analyses ofthe OMP-null mouse provide insight into the enigmatic role ofthis protein in the function of the olfactory neuron. TheOMP-null mice exhibit a defect in the EOG characterized bya reduction in the initial stimulus responsivity of the olfactoryepithelium to odors, altered kinetics of response generationand recovery, and a reduced ability to respond to the secondstimulus of a pair. These observations imply that overall thereis a reduction of neural activity in the olfactory projection tothe bulb. Consistent with this prediction, the profile of reducedgene expression in intrinsic olfactory bulb neurons is similar tothat seen in animals after either deafferentation or narisclosure, and reflects the reduced neural activity of the olfac-tory sensory neurons. This bulbar phenotype independentlysupports the conclusion that the OMP-null phenotype ischaracterized by a dramatic reduction in neural activity in theolfactory nerve to bulb projection. Together these indepen-dent observations are consistent with the compelling conclu-sion that OMP is a novel modulator of the signal detection/transduction cascade in the olfactory sensory neuron.
This work was supported in part by grants from the NationalInstitutes on Aging (H.B.), the National Institute on Deafness andOther Communication Disorders (A.F. and J.W.S.), and the NationalInstitiute on Drug Abuse (R.K.).
1. Shepherd, G. M. (1994) Neuron 13, 771-790.2. Buck, L. & Axel, R. (1991) Cell 65, 175-187.3. Reed, R. R. (1992) Neuron 8, 205-209.4. Liman, E. R. & Buck, L. B. (1994) Neuron 13, 611-621.5. Raming, K., Krieger, J., Strotmann, J., Boekhoff, I., Kubick, S.,
Baumstrack, C. & Breer, H. (1993) Nature (London) 361, 353-356.
6. Chess, A., Simon, I., Cedar, H. & Axel, R. (1994) Cell 78,823-834.
7. Ressler, K. J., Sullivan, S. L. & Buck, L. B. (1994) Curr. Opin.Neurobiol. 4, 588-596.
8. Ivanova, T. T. & Caprio, J. (1993)J. Gen. Physiol. 102,1085-1105.9. Sicard, G. & Holley, A. (1984) Brain Res. 292, 283-296.
10. Margolis, F. L. (1980) in Proteins of the Nervous System, eds.Bradshaw, R. A. & Schneider, D. M. (Raven, New York), pp.59-84.
11. Buiakova, 0. I., Rama Krishna, N. S., Getchell, T. V. & Margolis,F. L. (1994) Genomics 20, 452-462.
12. Menco, B. P. (1989) Cell Tissue Res. 256, 275-281.13. Lau, M. M., Stewart, C. E., Liu, Z., Bhatt, H., Rotwein, P. &
Stewart, C. L. (1994) Genes Dev. 8, 2953-2963.14. Kudrycki, K.,Stein-Izsak, C., Behn, C., Grillo, M., Akeson, R. &
Margolis, F. L. (1993) Mol. Cell. Biol. 13, 3002-3014.15. Joh, T. H., Geghman, C. & Reis, D. J. (1973) Proc. Natl. Acad.
Sci. USA 70, 2667-2771.16. Shimonaka, H., Marchand, J. E., Connelly, C. S. & Kream, R.
(1992) J. Neurochem. 59, 81-92.17. Ezeh, P. I., Davis, L. M. & Scott, J. W. (1995) J. Neurophysiol. 73,
2207-2220.18. Baker, H., Grillo, M. & Margolis, F. L. (1989) J. Comp. Neurol.
285, 246-261.19. Biffo, S., Grillo, M. & Margolis, F. L. (1990) Neuroscience 35,
637-651.
Proc. Natl. Acad. Sci. USA 93 (1996)
122

Neurobiology: Buiakova et al.
20. Verhaagen, J., Oestreicher, A. B., Gispen, W. H. & Margolis,F. L. (1989) J. Neurosci. 9, 683-691.
21. Baker, H., Morel, K., Stone, D. M. & Maruniak, J. A. (1993)Brain Res. 614, 109-116.
22. Baker, H., Towle, A. C. & Margolis, F. L. (1988) Brain Res. 450,69-80.
23. Ehrlich, M. E., Grillo, M., Joh, T. H., Margolis, F. L. & Baker, H.
Proc. Natl. Acad. Sci. USA 93 (1996) 9863
(1990) Mol. Brain Res. 7, 115-122.24. Henegar, J. R. & Maruniak, J. A. (1991) Brain Res. 568,230-234.25. Liebenauer, L. L. & Slotnick, B. M. (1996) Physiol. Behav., in press.26. Doty, R. L. (1986) Experientia 42, 257-271.27. Harding, J. W., Getchell, T. V. & Margolis, F. L. (1978) Brain
Res. 140, 271-285.28. Blass, E. M. & Teicher, M. H. (1980) Science 210, 15-22.


![OpenMP - IPSltodi.est.ips.pt/aabreu/openMP.pdf · #pragma omp sections [clause ...] newline {#pragma omp section newline structured_block #pragma omp section newline structured_block}](https://img.pdfslide.net/doc/110x75/5e752997f76299646930422c/openmp-pragma-omp-sections-clause-newline-pragma-omp-section-newline.jpg)
![Orbital Motors Type OMP, OMR and OMH · Type OMP OMP OMP OMP OMP OMP OMP Motor size 110 125 160 200 250 315 400 Max. torque N•m [lbf•in] cont. 215 [1900] 240 [2120] 300 [2660]](https://img.pdfslide.net/doc/110x75/5f5b5c3b3301e9386c6593ea/orbital-motors-type-omp-omr-and-type-omp-omp-omp-omp-omp-omp-omp-motor-size-110.jpg)

























